
Submitted:
14 January 2023
Posted:
16 January 2023
You are already at the latest version
Abstract
Arboviruses such as yellow fever virus (YFV), dengue virus (DENV), and chikungunya virus (CHIKV) presenting wide global dissemination and the pathogenic profile developed in infected individuals, which develop from nonspecific clinical conditions to severe forms, characterized by the promotion of significant lesions in different organs of the harborer, culminating in multiple organ dysfunction. To characterize, quantify, and compare the patterns of histopathological alterations in human liver samples from patients with yellow fever (YF), dengue fever (DF), and chikungunya fever (CF). Analytical cross-sectional study by histopathological analysis with 70 samples of liver patients, collected from 2000 to 2017, with confirmed laboratory diagnosis who died due to infection and complications by the YF, DF, and CF. Of the histopathological findings in human liver samples there was a significant difference between the control and infection groups, with a predominance of alterations in the midzonal area of the three cases analyzed, among the arboviruses studied, the hepatic involvement in cases of YF showed greater intensity of histopathological changes. Among the alterations evaluated, cell swelling, microvesicular steatosis and apoptosis were classified as degree of tissue damage from severe to very severe. The pathological abnormalities associated with infection by YFV, DENV, and CHIKV showed predominance of changes in the midzonal area. We also noted that in among of the arboviruses studied, liver involvement in cases of YFV infection was more intense.
Keywords:
Arboviruses
; Histopathology
; Cell death
; Necrosis
; Steatosis
; Liver
1. Introduction
Arboviruses (arthropod-borne viruses) develop part of their replication cycle in insects and can be transmitted to humans and other animals by the bite of hematophagous arthropods. These viruses have been identified in almost all continents, and tend to have a geographic distribution as part of a special ecological subsystem represented by viruses, vectors, amplifying hosts, and reservoirs [1,2,3].
In recent years, the importance of increased climate and environmental changes due to anthropogenic factors such as deforestation favor the transposition of the barrier between species [4]. In addition, the increase in activities, such as trade and tourism, helps spread these viruses from endemic regions to different parts of the world [5].
Favor the increase in arbovirus infections. Many countries have experienced the periodic emergence of arboviruses [6]. The establishment of arboviruses in certain places is based on the complex interaction of biotic and abiotic factors, which favor the occurrence of epidemics and epizootics in different continents [7].
Arboviruses such as yellow fever virus (YFV), dengue virus (DENV), and chikungunya virus (CHIKV), belonging to the Flavivirus genus and Flaviviridae family (YFV and DENV) and Alphavirus genus and Togaviridae family (CHIKV), are related to seasonal circulation in tropical and subtropical areas [8,9,10]. According to the World Health Organization, 47 countries are endemic or have endemic regions of yellow fever (YF), of which approximately 72% are located in Africa and 28% in Central and South America [11].
In South America, countries with a concentration of confirmatory diagnoses for the disease are Peru, Bolivia, and Brazil [12]. DENV, on the other hand, has a more extensive area of circulation, with cases of dengue fever (DF) being common in 100 countries worldwide [13,14]. In the Americas, approximately 500 million people are at risk of infection. Brazil and Colombia are the South American countries that led the ranking of cases of the disease [15].
Unlike YFV and DENV, which are more widely circulated viruses, CHIKV has recently demonstrated an impact on a global scale. Autochthonous transmission has been reported in 40 countries worldwide. However, 115 countries have reported cases of chikungunya fever (CF), with approximately 41% of the reported cases being in the Americas, and when observing South America, as well as in DF, chikungunya cases are concentrated in Brazil and Colombia [15].
These arboviruses develop transmission cycles in wild and urban environments and are transmitted by the bites of mosquitoes belonging to the Culicidae family [16,17]. Currently, in Brazil and South America, DF and CF cases are predominantly urban, while in YF, the prevalence is in the sylvatic cycle of the disease, determined by epizootics in different regions of the country, mainly in the southeast, northeast, and north [18]. Brazil is considered the main source of the circulation and dispersion of these arboviruses in other Latin American countries [19,20,21,22].
YFV, DENV, and CHIKV are responsible for causing different clinical manifestations in infected individuals, several factors influence the aggravation of the disease. They are associated with both the characteristics of the host and the characteristics of the viruses, including the patient's age, viral load, pattern of immune response, secondary infection, and interaction of viral proteins and host cell receptors. This host-virus relationship, plays an important role in the clinical outcome of the disease, as it determines not only the viral tissue tropism but also a multitude of intracellular mechanisms that can alter the overall outcome of the infection [23,24,25].
Approximately 15% to 60% of individuals infected with YFV develop symptoms of the disease, with evolution to death in 20% to 50% of cases [26,27]. In DF cases, lethality is also associated with the infecting serotype, which has four serotypes (DENV-1 to DENV-4), which influence the patient's clinical outcome [28]. In CF cases, most infected individuals develop symptoms. Severe cases tend to have atypical characteristics of the disease (hepatitis, myocarditis, and neuropathies) [29,30].
These arboviruses have been observed to replicate and cause significant damage to various organs of the host, such as the heart, lung, thymus, spleen, kidney, and liver; therefore, they are considered to be multi organic pathogens [30,31,32,33,34]. The liver is considered a target organ in YFV and DENV infections and is characterized by CHIKV infection [35,36]. The pathogenesis of liver injury involves several mechanisms, including direct viral cytopathic effects and immune-mediated injury [37].
The diagnosis of patients infected with these viruses can be performed using several techniques, including serology, viral isolation, immunohistochemistry (IHC), molecular biology, and histopathology [38,39,40,41,42,43,44,45]. Previous studies in animal models and histopathological results in humans have allowed the characterization of clinical and pathological changes that develop during viral pathogenesis [46].
The hepatic histopathological patterns observed in the pathogenic profiles of these arboviruses show different degrees of degenerative lesions, with the presence of inflammatory infiltrate, macro-and microvesicular steatosis, necrosis, alterations in hepatic sinusoidal endothelial cells (HSEC), and Kupffer cells [12,47,48,49,50,51,52,53,54,55,56].
Taking into account the viral pathogenesis of YFV, DENV, and CHIKV that are associated with hepatic impairment, this study aimed to characterize, quantify, and compare the patterns of histopathological alterations in human liver samples from patients with YF, DF, and CF, contributing to the improvement of the criteria for histopathological diagnosis of these diseases through systematic and comparative characterization, which can also help expand clinical-epidemiological criteria and expand knowledge regarding the harmful processes developed in human liver tissue in YFV, DENV, and CHIKV infections.
2. Materials and Methods
This was an analytical cross-sectional study with retrospective samples of patients with confirmed laboratory diagnoses who died due to infection and complications by YFV, DENV, and CHIKV. Patients' ages were 15 and 63 years (mean age = 37 years) and 60% were male. The samples came from the states of Tocantins, Goiás, Distrito Federal and Paraíba, where most cases of infections came from the state of Goiás, between 2000 and 2016.
Samples collected from patients with a confirmed diagnosis of YFV, DENV, and CHIKV infections by clinical/epidemiological, histopathological, IHC, and molecular criteria were included.
Samples that did not have confirmatory histopathological, IHC, or molecular reports were excluded. Samples that did not have enough material to make slides, that is, the amount of material was not enough for making slides, and its use could compromise the storage of a part for the return to the laboratory block file. In addition, the samples that were identified were impaired.
Seventy liver tissue samples from individuals with YF, DF, and CF were included in the study (36 samples of YF, 17 of DF, and 17 of CF) from the biobank of the Pathology Section of Institute Evandro Chagas (SAPAT/IEC), located in the Municipality of Ananindeua, arising from the epidemiological surveillance of the period from 2000 to 2017. These cases were diagnosed based on clinical/epidemiological criteria, histopathological examination, detection of viral antigens by IHC, or viral genome by polymerase chain reaction with reverse transcription-polymerase chain reaction in blood or frozen liver samples for diagnosis (RT-PCR).
Liver samples obtained during routine autopsies were included as negative controls. Twenty liver samples from patients with a negative molecular diagnosis for the main hepatotropic viruses, with no morphological changes, and from the Death Verification Service of the Metropolitan Region of the city of Belém, State do Pará, Brazil (Supplementary Figure).
Samples that did not have confirmatory histopathological, IHC, or molecular reports were excluded. Samples that did not have enough material to make slides, that is, the amount of material was not enough for making slides, and its use could compromise the storage of a part for the return to the laboratory block file. In addition, the samples that were identified were impaired.
All procedures performed during this study were approved by the Research Ethics Committee of the Institute Evandro Chagas, following the recommendations by the resolution of National Health Council No. 466/12. It was approved with perish number 2.364.226. Free and informed consent terms were waived, as the samples collected were obtained post-mortem, and all data will be handled and analyzed anonymously, without the nominal identification of individuals.
Histological Analysis
Liver tissue samples were fixed for 24 h in 10% buffered formalin solution and kept in ethyl alcohol. Tissue fragments were passed through several increasing solutions of alcohol (70–100%). Two passages were carried out in xylene and two in paraffin at 60°C. After cooling, the tissue blocks were sectioned using a rotating microtome to obtain 5µm-thick sections. Tissue sections were stained with hematoxylin and eosin (H&E).
To evaluate the histopathological patterns, the stained samples were visualized using microscope at a total magnification of 400x in the area or zone of the hepatic acinus of Rappaport (Z1 = periportal, Z2 = midzonal, and Z3 = centrilobular) and the portal tract (PT), measured in terms of damage according to a semiquantitative scale from 0 to 4 (0: absent, 1: mild+, 2: moderate ++, 3: intense +++, 4: very intense ++++) to classify the degree of tissue damage observed, as described by Quaresma et al. [47].
Morphological alterations in the hepatic acinus (Z1, Z2, and Z3), PT, and inflammatory infiltrate included cell swelling, macrovesicular steatosis, microvesicular steatosis, lytic necrosis, coagulative necrosis, apoptosis (Councilman-Rocha Lima body), hyperplasia of Kupffer cells, Kupffer cell hypertrophy, sinusoidal endothelial alteration (SEA), sinusoidal congestion, sinusoidal dilatation, edema, congestion, portal vein alteration, portal artery alteration, biliary canaliculus alteration, and lymphocytes, neutrophils, plasma cells, and macrophages.
Immunohistochemistry
Tissue immunostaining with hyperimmune mouse ascitic fluids to YFV, DENV and CHIKV produced in young mice were titrated to choose the best dilution for immunohistochemical assay [58]. Initially, tissue samples were deparaffinized in xylene and hydrated in a series of decreasing concentrations of ethyl alcohol (90%, 80%, and 70%). Subsequently, antigenic recovery was performed in protease solution (Sigma-Aldrich, Poole, UK) at 37 °C. Non-specific proteins were blocked by a solution of 5% skim milk; 0,01 M phosphate buffered saline (PBS) and normal horse serum (Vector Laboratories, Burlingame, USA). The anti-YFV, DENV and CHIKV antibodies diluted 1:100 in PBS pH 7.2 were then added on the slides and after they were incubated at room temperature for 1 h. After three washes with PBS, incubation with biotinylated anti-mouse IgG reagent secondary antibody (Vector Laboratories, Burlingame, USA) diluted 1: 5 was performed for 1 h, followed by three washes in PBS. After this step, the sections were incubated with streptavidin-alkaline phosphatase complex (Invitrogen Corporation, Camarillo, CA, USA) 1:20 dilution for 1 h at room temperature; finally, the slides were washed three times with 0,1 M TRIS solution. For reveal the reaction, HistoMarkRed phosphatase kit (KPL, Gaithersburg, USA) was used on the slides for 30 min at room temperature. Then, the sections were counterstained with Harris haematoxylin and the slides were assembled with entellan.
Molecular Biology
All samples used in the present study were submitted to refrigeration, followed by freezing at -20oC, -70oC, or immersed in liquid nitrogen. Biological samples of human blood and tissue were subjected to diagnostic tests for YFV, DENV, and CHIKV by real-time PCR according to the protocol described by Mori et al. [42].
Statistical Analysis
Data were analyzed using descriptive and inferential statistics, according to the nature of the variables, summarized by measures of central tendency and dispersion, in addition to the confidence interval. This is expressed in the form of graphics. To compare the different groups in terms of morphological changes, inflammatory infiltrate, and SEA in each acinar zone, in addition to structural changes and inflammatory infiltrate observed in the PT, Kruskal–Wallis analysis of variance was used, followed by Dunn's post-test.
In comparisons of the same group in relation to a given variable but in different zones, the comparison was made using Friedman's analysis of variance with Dunn's post-test. The results obtained during the research were stored in Microsoft Excel 2019 spreadsheets, and graphs were constructed using GraphPad Prism 9.2 program (GraphPad Software, San Diego, CA, USA). Differences were considered to be statistically significant at P ≤ .05.
Results
Qualitative Histology
Qualitative analysis of the histopathological aspects showed that cell swelling, steatosis, and apoptosis were the most frequent hepatic alterations observed in the YF, DF, and CF samples in all areas of the hepatic acinus. However, there was a prevalence in the midzonal area. Histopathological changes and the presence of immunostaining of viral antigens were absent in normal controls samples (Supplemental Figure).
Councilman's acidophilic bodies, representing hepatocytes in an apoptotic process, demonstrated by a homogeneous, intensely acidophilic mass, were frequently observed in the three zones of the YF and DF samples and were absent in CF cases. However, the process showed a greater comparative intensity in the midzonal area of the YF cases (Figure 1A–F).
Small focal points of lytic and coagulative necrosis were also observed in the hepatocytes of the cases of YF, DF, and CF, which were generally mild in the three analyzed zones; however, when lytic necrosis was observed in Z2 of patients with YF, DF, and CF, a predominance of this alteration was observed in the area. These focal points of necrosis were visualized as areas of parenchyma showing a complete absence of cells and were generally replaced by an amorphous and slightly eosinophilic mass containing cellular debris (Figure 2A–F).
Other alterations observed in the acinar zones (Z1, Z2, and Z3) were hyperplasia and hypertrophy of Kupffer cells, which were mild to moderate, with emphasis on Z2 in cases of YF, DF, and CF, presenting a higher frequency of alterations. In addition, the fatty changes were characterized by macro-and microvesicular steatosis in the cytoplasm of hepatocytes, showing a characteristic aspect of these types of steatosis with large and small droplets in the cytoplasm of hepatocytes, respectively. These changes were accompanied by mild to moderate acinar, and portal inflammatory infiltrates consisting of lymphocytes, plasma cells, neutrophils, and macrophages, with a higher frequency of lymphoplasmacytic infiltrates, especially around foci of necrosis. The distribution of inflammation in the lobule maintained a pattern similar to that of other acinar changes, that is, with a clear preference for the midzonal area (Figure 3A–F).
Among the endothelial alterations observed in the samples, there was a difference in histopathological findings between the arboviruses studied; in the SEA cases, YF and CF showed a higher frequency of this alteration in relation to cases. As for sinusoidal congestion, cases of YF and DF stood out and were more intense than cases of CF. In sinusoidal dilatation, the highlight was for CF cases, with no change in YF and DF cases. There was a predominance of endothelial alterations in Z2.
In PT, the main alterations analyzed were moderate, with edema equivalent to the three arboviruses studied, and in portal vein alteration, DF cases were more intense. Congestion and alteration of the biliary canaliculus were mild and observed only in cases of YF and DF and absent in cases of CF. However, the alteration of the portal artery was mild for the three arboviruses studied.
Semiquantitative Histology
Histopathological Changes in Hepatic Acinus
The semiquantitative data on the histopathological changes in the hepatic acinus showed a significant difference between the control and analyzed variables in all zones of the hepatic acinus. With the presence of intense cellular swelling in the three hepatic zones of CF cases, no significant difference was observed between the zones of the samples in these cases. In the case of YF, only Z1 showed a high swelling intensity, while the other two zones were of moderate intensity. In DF cases, swelling was moderate in the three analyzed zones. In the individual analysis, the DF and CF cases presented a homogeneous frequency and intensity between the hepatic acinus zones for this alteration. In the CF cases, the intensity was high, and in the DF, it was moderate; in the YF, there was an intensity variation between the analyzed areas, with emphasis on Z1 (Figure 4 and Figure 5).
As for the data referring to the fatty alterations characterized by macrovesicular and microvesicular steatosis. In macrovesicular steatosis, the results were equivalent between the arboviruses and the studied zones; that is, no significant difference between the cases of YF, DF, and CF when compared to each other in a given zone, but there was a significant difference between the control and the cases (Z1 and Z2: ctrl × yfv, P < .001 / ctrl × denv, P < .001 / ctrl × chikv, P < .001; Z3: ctrl × yfv, P = .02 / ctrl × denv, P < .001 / ctrl × chikv, P < .001; ). In Z1 and Z3, the intensity of the alteration was mild, whereas in Z2 of all arboviruses, an increase in the quantification of the alteration was observed, which was moderate in all cases studied. Thus, the macrovesicular steatosis identified in the midzonal area of YF, DF, and CF cases seemed to show higher values than other areas of the hepatic acinus (Figure 4A–C).
In microvesicular steatosis, there was a clear difference between the arboviruses analyzed, except between YF and DF (Z1, Z2 and Z3: yfv x chikv, P < .001 / denv x chikv, P < .001; Z1 and Z3, yvf × denv, P > .99; Z2: yfv x denv, P = .30); in Z1 and Z3, the YF and DF cases presented moderate intensity, and in Z2, the change was intense for both infections. An increase in the intensity of microvesicular steatosis was observed in the midzonal area in the YF and DF cases (Figure 4D–F).
Regarding the necrotic processes evaluated, regarding lytic necrosis, liver samples from patients with YF, DF, and CF showed no significant difference when compared to each other (Z1 and Z3: yfv × denv, P > .99 / yfv × chikv, P > .99 / denv × chikv, P > .99; Z2: yfv × denv, P > .99 / yfv × chikv, P = .10 / denv × chikv, P = .25); however, there was a significant difference between the control and infection groups (All Zones: ctrl × yfv, P < .001 / ctrl × denv, P < .001 / ctrl × chikv, P < .001). Although the values referring to lytic necrosis were equivalent between arboviruses, in Z1 and Z3 the necrosis was mild; however, in Z2, lytic necrosis was moderate (Figure 5). Regarding coagulative necrosis, all areas of the three infections evaluated corresponded to a mild degree of this alteration (Figure 6), with differents results between patients with YF, DF and CF in Z2 (yfv × denv, P > .99 / yfv × chikv, P = .02 / denv × chikv, P = .02) and Z3 (yfv × denv, P > .99 / yfv × chikv, P = .03 / denv × chikv, P = .16), but without significant differences in Z1 (yfv × denv, P > .99 / yfv × chikv, P = .90 / denv × chikv, P > .99) (Figure 6).
Regarding apoptosis, CF cases expressed the absence of this alteration in all analyzed zones (ctrl × chikv, P > .99). In DF cases, there was a significant difference between the control and infection group (Z1: ctrl × denv, P = .02; Z2: ctrl × denv, P < .01; Z3: ctrl × denv, P < .001), with moderate apoptosis in all zones. The cases of YF were related to the highest values of apoptotic processes of the arboviruses studied, emphasizing the midzonal area, presenting a very intense lesion (All Zones: ctrl x yfv, P < .001). The periportal and centrilobular areas of YF samples showed intense and moderate apoptosis, respectively (Figure 6).
Other changes analyzed included Kupffer cell hyperplasia and hypertrophy. In hyperplasia, DF cases significantly differed between the control and infection group in all areas evaluated (Z1: ctrl × denv, P < .01; Z2: ctrl × denv, P < .001; Z3: ctrl × denv, P = .04). However, the cases of YF and CF showed no change in Z3 (ctrl x yfv, P = .17 / ctrl x chikv: P > .99 / yfv x chikv: P > .99). Z1 and Z2 of the cases referring to the three arboviruses had equivalent lesion intensities, with mild and moderate hyperplasia, respectively.
In the case of Kupffer cell hypertrophy, all arboviruses showed a significant difference between the control and infection groups in the different hepatic zones: for Z1 and Z2, the values corresponded to the arboviruses with mild and moderate hypertrophy, respectively (Z1: ctrl × yfv, P < .001 / ctrl × denv, P = .02 / ctrl × chikv, P < .001; Z2: ctrl × yfv, P < .001 / ctrl × denv, P < .01 / ctrl × chikv, P < .001). In Z3, however, there was a difference between arboviruses: CF cases showed moderate cellular hypertrophy, while YF and FD had mild intensity (yfv × chikv, P = .02 / denv × chikv, P < .01 / ctrl x yfv, P < .001 / ctrl x denv, P = .01 / ctrl x chikv, P < .01). Unlike hyperplasia, in hypertrophy, CF cases showed a higher frequency of moderate intensity than YF and DF. Regarding the evaluated zones, the midzonal area showed a greater commitment. However, in CF cases, the midzonal and centrilobular areas of the hepatic acinus were equally affected (Figure 7).
Regarding the semiquantification of endothelial changes in the hepatic acinus of YF, DF, and CF cases, three variables, including SEA, showed a significant difference between the control and infection groups. In all three areas evaluated, cases of YF and CF expressed a moderate profile of SEA (Z1 and Z2: yfv × chikv, P > .99 / Z3: yfv × chikv, P = .27), whereas in cases of DF, only Z2 showed moderate injury (yfv × denv: P = .77 / denv × chikv: P > .99), and in Z1 and Z3, the SEA was light (Z1: yfv × denv, P = .26 / denv × chikv, P = .045; Z3: yfv × denv, P = .35 / denv × chikv, P < .01). Among the arboviruses studied, SEA was predominant in cases of YF and CF, with homogeneous intensity between the different areas analyzed. In all infections, Z2 was the most affected (Figure 8).
The second variable analyzed was sinusoidal congestion. This change was absent in CF cases in all analyzed zones (Z1: ctrl × chikv, P = .30; Z2: ctrl × chikv, P = .77; Z3: ctrl × chikv, P = .43). Significant values refer to cases of YF and DF (Z1: ctrl × yfv and ctrl × denv, P < .01; Z2: ctrl × yfv, P < .01 / ctrl × denv, P = .03; Z3: ctrl × yfv, P = .02 / ctrl × denv, P = .12 ), with corresponding results in Z1 and Z3, with the presence of mild congestion (Z1 and Z3: yfv × denv, P > .99). However, sinusoidal congestion in Z2 of the YF cases was moderate, whereas in DF, the same area analyzed presented mild congestion (yfv × denv: P > .99). Sinusoidal congestion was prevalent in the YF and DF cases, with greater intensity in the Z2 of YF cases.
The last alteration associated with the hepatic endothelium is sinusoidal dilatation. Alteration was absent in the cases of YF and DF in all analyzed zones (All zones: ctrl × yfv, ctrl × denv and yfv × denv, P > .99). However, for CF cases, dilation was moderate and homogeneous in all three studied areas (All zones: ctrl × chikv, yfv × chikv and denv × chikv, P < .001). We observed a clear predominance of sinusoidal dilatation in CF cases compared to YF and DF, without a specific zone of involvement.
Among the changes observed in PT, edema was moderate in cases of YF, DF, and CF (ctrl × yfv: P < .001 / ctrl × denv: P < .01 / ctrl × chikv: P < .001), and there was no significant difference between arboviruses (yfv × denv: P = .84 / yfv × chikv: P > .99 / denv × chikv: P > .99) (Figure 6A). In the case of portal congestion, the cases of YF and DF were mild (yfv × chikv: P < .001 / denv × chikv: P < .01 / yfv × denv: P > .99), and in CF cases, changes were absent (ctrl × chikv: P > .99) (Figure 8). Regarding the alteration of the portal vein, DF cases presented higher values than YF and CF, although it was not statistically significant (yfv × denv: P > .99) / denv × chikv: P = .21), being moderate in DF cases and light in YF and CF (Figure 8).
There was no significant difference in portal artery alterations between the arboviruses studied (yfv × denv: P > .99 / yfv × chikv: P > .99 / denv × chikv: P = .38) presenting a mild degree, only between the control and samples. However, when bile canaliculus alteration was evaluated, there was a predominance in cases of YF and DF (ctrl × yfv: P = .02 / ctrl × denv: P < .01 / yfv × denv: P = .12) with a mild degree, while there was absence in CF cases (ctrl × chikv: P > .99 / yfv × chikv: P = .36) / denv × chikv: P = .01) (Figure 8). Of the five alterations evaluated in the PT of fatal cases of YF, DF, and CF, homogeneous values were observed among the arboviruses related to the variables edema and alteration of the portal artery, with no alteration observed only in cases of CF associated with the variable congestion and alteration of the bile canaliculus. DF cases showed higher values in hepatic portal vein alteration when compared to YF and CF cases. In a general evaluation of the alterations in PT, the cases of YF and DF presented a higher frequency of alterations in relation to FC.
The acinar inflammatory infiltrate was mild to moderate among the arboviruses studied, with a significant difference between the control and infection groups. Of the cell types of inflammatory cells, lymphocytes in Z1 and Z3 of the cases of YF, DF, and CF were identified, which showed mild intensity, with no significant difference between the inflammatory processes of the infections evaluated in these areas of the hepatic acinus (Z1 and Z3: yfv × denv, P > .99 / yfv × chikv, P > .99 / denv × chikv, P > .99), the Z2 of the YF and DF cases had higher values when compared to the Z2 of the CF cases, with moderate infiltration and light, respectively (yfv × denv: P > .99 / yfv × chikv: P < .001 / denv × chikv: P < .01).
The plasma cell infiltrates identified in the hepatic acinus of fatal cases of YF, DF, and CF were equivalent to the lymphocytic infiltrates in Z1 and Z3 of the cases studied, with mild intensity, with no significant difference between arboviruses (Z1 and Z3: yfv × denv, P > .99 / yfv × chikv, P > .99 / denv × chikv, P > .99). However, unlike the lymphocytic infiltrate, the evaluation of the plasma cell infiltrate in Z2 of arboviruses showed an increase only in cases of YF, with a moderate degree of intensity (yfv × denv: P = .52)/ yfv × chikv: P = .01 / denv × chikv: P = .25).
Neutrophil acinar inflammatory infiltrate was not identified in Z1 and Z3 of CF cases (Z1: ctrl × chikv, P > .99 / yfv × chikv, P = .19 / denv × chikv, P = .10; Z3: ctrl × chikv, P = .32 / yfv × chikv, P > .99 / denv × chikv, P > .99). In cases of YF, in Z1 and Z2 zones they have mild intensity. Neutrophil infiltration was absent in Z3 of the YF cases. In DF cases, the infiltrate was homogeneous between the analyzed zones, all of which had a mild intensity of neutrophils. A preferential occurrence of acinar inflammatory infiltrate in the middle zone of the hepatic acinus was observed. Among the arboviruses studied, cases of YFV and DENV infection showed greater intensity of inflammatory cells in the liver tissue when compared to CHIKV infections.
As for the portal inflammatory infiltrate, the lymphocytic inflammatory infiltrate was moderate in cases of YF and DF and mild in CF cases (yfv × denv: P = .11 / yfv × chikv: P < .01 / denv × chikv: P > .99). Similar to the acinar infiltrate values, the plasma cell infiltrates in the YF cases were comparatively higher than those in the DF and CF cases, corresponding to moderate and mild degrees, respectively, although it was not statistically significant (yfv × denv: P = .86 / yfv × chikv: P > .99 / denv × chikv: P > .99). In PT, no neutrophil inflammatory infiltrate was identified in CF cases (ctrl × chikv: P > .99 / yfv × chikv: P < .01 / denv × chikv: P < .01), only mild infiltrate was identified in cases of YF and DF (yfv × denv: P > .99). The last inflammatory cell specimens identified in the PT were macrophages, and we observed that there was no significant difference between the YF, DF, and CF samples, which showed a light macrophage infiltrate (yfv × denv: P > .99 / yfv × chikv: P > .99 / denv × chikv: P > .99).
According to the portal inflammatory infiltrate data, a more intense occurrence of inflammatory cells was observed in the cases of YF and DF compared to the cases of CF. The same pattern was identified in the acinar inflammatory infiltrate where YFV and DENV infections showed greater intensity of inflammatory cells in the liver tissue. Another aspect found was that the cell specimens with the highest concentration were lymphocytes and plasma cells.
4. Discussion
Arboviruses, such as YF, DF, and CF, are considered diseases that pose risks to global health, as they behave epidemiologically in the form of epidemics and epizootics, with outbreaks of a seasonal pattern of greater or lesser impact on public health. In the tropical regions of Africa and South America, biotic and abiotic factors favor the establishment and circulation of these arboviruses, such as rainfall, climate, temperature, presence of dispersing vectors, and amplification of hosts and reservoirs [8,59].
Brazil is considered the main source of the circulation and dispersion of these arboviruses in other Latin American countries [19]. In South America, Brazil, Bolivia, and Peru are the countries that lead the ranking of confirmed cases of YFV, DENV, and CHIKV infections. In Brazil, cases of DF and CF are predominantly urban, while in YF, the prevalence is in the sylvatic cycle of the disease, determined by epizootics in different regions of the country, mainly in the southeast, northeast, and north [18]. In northern Brazil, the Amazon region predominates these arboviruses and is classified as an endemic area for these diseases [20,21,22].
Such diseases have several clinical outcomes, framed as asymptomatic, mild, and fatal cases; the latter is characterized by multiple organ dysfunction, hemorrhage, severe jaundice, hepatitis, myocarditis, and neuropathies [26,27,60]. These clinical manifestations depend on several factors, such as viral load, tissue tropism, genetic factors, and immune evasion mechanisms, referring to both the virus and host. In addition, the interaction of viral proteins and host cell receptors may be relevant, as their interaction dictates not only viral tissue tropism but also a multitude of intracellular mechanisms that can alter the overall outcome of the infection [24,25].
Although several studies have addressed the clinical manifestations and hepatic histological changes of YF and DF, little is known about the hepatic involvement in CHIKV infection or hepatic histopathological differences in fatal cases of YF, DF, and CF. It is known that these three arboviruses are correlated with the development of liver tissue damage mechanisms, and worsening of the infection is associated with a greater probability of a lethal outcome. What are the histopathological similarities and differences in liver involvement among these three arboviruses?
Previous studies have reported several causes of the pathogenesis of these viruses, which are inherent to the host and virus. The viral proteins nsP2 and nsP3 of the alphavirus CHIKV and NS1, NS4A/B, and NS5, which belong to the flaviviruses YFV and DENV, play roles in the development of viral cytopathic effects. In addition to being associated with persistence in host cells, it promotes the attenuation of antiviral responses by inhibiting key factors of the immune response, resulting in viral dissemination in the host organism [25,61,62,63,64,65,66].
Considering the multi organic character of these arboviruses, related to infection and proliferation in different organs of the host [52,67]. the hepatic histopathological findings referring to YF, DF, and CF associate YFV infection, DENV, and CHIKV to different degrees of degenerative lesions, with the presence of inflammatory infiltrate, macro and microvesicular steatosis, necrosis, and changes in HSEC and Kupffer cells [30,31,47,48,51,52,68].
In our study on the semiquantitative histopathology of 70 liver samples from patients with YF, DF, and fatal CF, the quantification of histopathological changes showed abnormalities associated with cell swelling, macro and microvesicular steatosis, lytic and coagulative necrosis, apoptosis, hyperplasia, hypertrophy of Kupffer cells, endothelial changes, and inflammatory infiltrate. Cellular swelling in DF and CF cases showed homogeneous intensity, with no differences between the analyzed acinus zones; however, for CF cases, the swelling was intense, and in cases of moderate DF, in YF, an intensity variation was observed between the zones, in which in Z1 the swelling was intense, and moderate in Z2 and Z3.
Although there are no data on swelling in the liver tissue in CF cases, several studies have shown that YFV and DENV infections are associated with the development of hepatocyte swelling [69,70,71,72,73,74]. According to a study by Quaresma et al., the hepatocytes in Z1 and Z3 showed a greater intensity of this alteration. Cell swelling is attributed to changes in the mechanisms controlling water and ion concentrations due to direct viral action on the cell membrane or the immune response generated by the host [75,76].
As for macro-and microvesicular steatosis, the results showed that there was no significant difference between arboviruses with uniform intensity between them in macrovesicular steatosis. However, a moderate difference in intensity was observed between the acinar zones, and the cases of YF, DF, and CF showed a moderate increase in macrovesicular steatosis in Z2. In microvesicular steatosis, this alteration was not identified in CF cases in any of the three areas evaluated, whereas the intensity observed in YF and DF cases was equivalent, with moderate microvesicular steatosis in Z1 and Z3 and intense in Z2. Similar to macrovesicular steatosis, a preferential concentration of small lipid vesicles was observed in the cytoplasm of hepatocytes located in the midzonal area. A predominance of microvesicular steatosis was also observed in cases of YF and DF, unlike the results for macrovesicular steatosis, in which there was no difference between the arboviruses studied (Figure 9).
In YF, macro-and microvesicular steatosis is a change commonly observed in infected patients, present in the three zones of the hepatic acinus, but it is predominant in Z2 [57,77,78]. DF is associated with fatty liver degeneration [79,80]. Recently, Win et al. reported findings in human liver samples from patients who died of dengue hemorrhagic fever (DHF), which showed extensive cell damage and microvesicular steatosis [37]. Most studies relate YFV and DENV infections to the detection of microgoticular steatosis, the presence of which is related to changes in lipid metabolism, one of the mechanisms involved in hepatomegaly, a clinical manifestation associated with hepatitis observed in YF and DF [31,57,70].
Although data on hepatic fatty degeneration in CF are scarce, flavivirus and alphavirus infections have lipid metabolic implications, influencing the immune response against infection and the maintenance of viral pathogenic mechanisms [62]. In addition, studies have reported that the synthesis of lipid mediators during viral infection is related to the signaling, control, and maintenance of the immune response and viral pathogenesis [82,83,84,85,86].
During infection, the virus can control the lipid metabolism of the host cell in order to meet the demands required in the viral replication process [87,88]. Since the mobilization and recruitment of lipid droplets is an essential mechanism in the bioenergetic demand for viral proliferation, which occurs through the formation of replication complexes in regions of lipid concentration [89,90,91,92]. Thus, lipid metabolism is directly linked to the development of YFV, DENV, and CHIKV pathogenesis [93,94,95].
According to a study by Quaresma et al., the intensity of steatotic hepatocytes coincides with the intensity of apoptotic processes. Two studies on chronic hepatitis correlated steatosis with unfavorable prognosis [96,97]. Pre-existing non-alcoholic steatohepatitis is also an aggravating clinical factor that may potentiate liver failure during the disease [98].
Regarding the necrotic processes analyzed in liver samples from patients infected with YFV, DENV, and CHIKV, the degree of injury ranged from mild to moderate. There was no significant difference between arboviruses in lytic and coagulative necrosis, with equivalent intensity of injury. However, in lytic necrosis, there was a preferential concentration of lesions in the midzonal area of the YF, DF, and CF samples, with a moderate degree of intensity. In evaluating coagulative necrosis, all cases in the three zones of the hepatic acinus corresponded to a mild degree of this alteration, with no significant difference between the arboviruses or zones (Figure 10).
According to the histopathological findings of other studies, severe forms of DF are determined by the presence of hepatocellular necrosis, usually observed in Z1 and Z2 of the hepatic acinus [36,99]. In addition, several studies have associated the frequency of distinct histopathological findings in YF and DF with the Z2 of the hepatic acinus, but this pattern has also been observed in infections by other arboviruses [51,57,76,100,101,102].
In our findings, Z2 was the area with the highest concentration of damage, with the degree of injury varying from moderate to very intense, particularly in the case of YF, which presented the highest levels of harmful processes in the hepatic parenchyma. According to Pan et al., the development of injurious mechanisms in Z2 of the hepatic acinus is characteristic of the pathogenesis of flavivirus infections.
This pattern of injury in the midzonal area is attributed to the intrinsic characteristics of hepatocytes in this area, which have a lower presence of oxygen and nutrients when compared to Z1 [70,99,104]. In a study by Olímpio et al., lytic and coagulative necrosis were recurrent findings in the three hepatic zones of patients with YF, but the highest frequency was in Z2. The triggering of these cell destruction processes may be related to the activation of a varied set of defense cells, generating a strong immune response characterized by a cytokine storm that causes tissue damage [47].
Hepatic histopathological findings from human cases of co-infection with DENV-CHIKV showed that patients had coagulative necrosis, especially around the hepatic vein [30,33,68]. In animal models of CHIKV infection, infected mice presented on the third day after infection, foci of hepatocytic necrosis dispersed in the liver parenchyma without a specific zone [105,106,107,108].
Regarding apoptosis, we observed significant differences between arboviruses; CF cases showed the absence of this alteration in all analyzed areas. In DF cases, the results were homogeneous between the zones; that is, apoptotic hepatocytes showed a homogeneous distribution in the liver parenchyma, with moderate intensity in the evaluated areas. The cases of YF were related to the highest values of apoptotic processes of the arboviruses studied, emphasizing the midzonal area, presenting a very intense lesion. The periportal and centrilobular areas of YF samples showed intense and moderate apoptosis, respectively. From these data, it was possible to observe a clear predominance of the alteration in the cases of YF, especially in Z2 (Figure 6).
Apoptosis is a prominent histopathological feature of YFV infection, seen in hepatocytes, Kupffer cells, and endothelial cells. In YF, liver injury mechanisms are complex and involve the cytopathic effects inherent to viral replication and the development of the host's immune response to neutralize viral replication by activating, producing, and releasing cytokines, chemokines, and reactive oxygen species (ROS) and immune cells (Figure 11) [48,57,70,74,76,109,110].
The intense apoptotic event in hepatocytes in YF is also associated with the development of coagulopathy, a clinical manifestation of severe YF. Since hepatocytes are the main producers of most circulating clotting factors, the loss of these factors via hepatocyte destruction or some other mechanism can also result in an imbalance in the coagulation cascade, promoting hemorrhagic manifestation, a classic clinical feature in the fatal outcome of the disease [111]. The presence of apoptosis, lytic necrosis, swelling, and steatosis of hepatocytes is related to liver failure, a clinical manifestation observed in severe YF [112].
In the liver, two pathways of apoptosis were observed: extrinsic and intrinsic, which are dependent and independent of death receptors. Death receptors are the main mediators of apoptosis in the liver [113]. Enveloped viruses such as flavivirus and alphavirus are present in the outer layer of the host cell membrane derived from the endoplasmic reticulum (ER) and are mainly rich in phosphatidylserine (PS) and phosphatidylethanolamine (PE) (Figure 11) [114,115,116,117,118].
PS and PE are relevant components for interaction with the host's entry receptors and are important for the processes of tropism, pathogenicity, and viral infectivity. PS is mainly found in the outer membrane of apoptotic cells and is used as a “signal” to tissue macrophages, this mechanism triggered by PS by viruses as binding molecules trigger processes of “apoptotic mimicry” [119].
Regarding the hyperplasia and hypertrophy of Kupffer cells related to YF, DF, and CF cases. Data regarding hyperplasia showed a clear difference between arboviruses in terms of alteration. DF cases had a higher frequency of hyperplastic Kupffer cells, and an increase in the intensity of hyperplasia in the midzonal area of YF, DF, and CF cases was also identified. As for hypertrophy, Z1 and Z2 in the cases of YF, DF, and CF showed corresponding values with mild and moderate hypertrophy, respectively. In Z3, there was a difference between arboviruses; CF cases showed moderate cellular hypertrophy, while YF and DF cases showed mild hypertrophy. Unlike hyperplasia, in hypertrophy, CF cases showed a higher frequency of moderate intensity than YF and DF. Regarding the evaluated zones, the midzonal area was more affected, but in the case of CF, the midzonal and centrilobular areas were equally affected.
Kupffer cell hyperplasia and hypertrophy in YF have been widely documented in the scientific literature by several authors [7,100,120]. Hyperplasia and Kupffer cell destruction have been identified in most fatal DF cases [37,80,121,122]. Liver injury observed in hemorrhagic fevers, such as DF and YF, revealed that hepatocytes and Kupffer cells might be the target cells for viral replication [69,123]. Viral antigens were detected in Kupffer cells and hepatocytes from patients with DHF near damaged areas, suggesting an association between viral replication and liver damage [124]. Viral antigens were also detected in Kupffer cells in liver samples from fatal CF cases [34].
Three axes referring to hepatic endothelial alterations in the cases of YF, DF, and CF were evaluated. In the comparative analysis, SEA was predominant in cases of YF and CF, with moderate intensity in the three different zones of the hepatic acinus. In relation to DF, the midzonal area of the cases was more affected, with a moderate degree of alteration, while in Z1 and FD Z3, the change was slight. In sinusoidal congestion, CF cases did not show any change in any of the analyzed areas; therefore, the prevalence of congestion was in YF and DF cases, emphasizing the Z2 of YF cases. In contrast to congestion, in sinusoidal dilatation, we observed that the cases of YF and DF did not present any alteration in any of the analyzed zones; however, in the cases of CF, sinusoidal dilatation was moderate in all the zones, that is, there was no specific zone of involvement.
Viral antigens have been detected in HSEC and Kupffer cells in arbovirus infections [125,126,127]. In a study by Zellweger et al., a DENV-infected mouse model showed an intense HSEC infection. Local HSEC injury may result in changes in portal microcirculation, resulting in secondary ischemic injury and portal hypertension that may be more pronounced in some patients [37]. Reversal of portal flow has also been demonstrated in flavivirus infection, which, together with circulatory dysfunction of the splanchnic vascular bed, may explain several features of liver dysfunction [129].
Sinusoidal congestion has been characterized in YF infections [77]. Sinusoidal capillaries show intense circulation of macrophages/lymphocytes, and immune processes generate hepatocyte injury and necrosis of sinusoid capillaries due to the release of cytokines and activation of the complement system, as well as endothelial dysfunction [130]. Two other studies related this change to fatal DF cases, pointing to hemorrhagic congestion in the hepatic sinusoids [121,122]. Most patients have moderate-to-severe hepatic sinusoidal congestion [36,121,131].
In a study by Sharp et al., liver samples from fatal CF cases showed the presence of the viral antigen in vessels, Kupffer cells, HSEC, and portal connective tissue; leukocyte concentration was also observed within the vascular channels of the liver. The most frequently identified histopathological features were chronic portal hepatitis (mild), steatosis (mild), rare hepatocyte necrosis/apoptosis, and sinusoidal congestion with leukocytosis. A study by Agarwal et al. involving hepatic histopathological analysis of mice infected with CHIKV demonstrated the presence of dilatation of the hepatic sinusoids of these animals as a result of viral infection.
Of the alterations identified in the study referring to PT, the main alterations analyzed were moderate, with edema being equivalent to the three arboviruses studied (Figure 11), and in the alteration of the portal vein, the cases of DF were more intense. Congestion and alteration of the biliary canaliculus were mild and observed only in cases of YF and DF, and absent in cases of CF. However, the alteration of the portal artery was mild for the three arboviruses studied. In the study by Duarte-neto et al., microscopic liver analysis of YFV-infected individuals demonstrated the presence of edema in the PT and the space of Disse. None of the cases had bile duct injury.
Although the results obtained in the research related to histopathological alterations in DF, in general, were classified as mild to moderate, an explanation for these results would be the infectious serotype of the analyzed samples. Given that the study did not aim at the molecular characterization of the serotypes responsible for the infection, it is unknown whether the serotype of the samples examined was related to a different manifestation of DF, since the different clinical manifestations of the disease are also correlated with the DENV serotype. Patients infected with DENV-2 have a higher frequency of severe forms of the disease than those infected with other serotypes [132,133]. These patients manifest persistent vomiting, epigastric pain, plasma leakage, and shock [134].
According to Halsey et al., patients infected with DENV-3 showed a prevalence of musculoskeletal and gastrointestinal manifestations, whereas DENV-4 was associated with respiratory and cutaneous manifestations. Among the serotypes, DENV-2 was the most frequently isolated in cases of DHF and dengue shock syndrome, followed by DENV-3, DENV-1, and DENV-4 [28,136]. Another possibility is the individual aspects of each case, considering that the intrinsic characteristics of the host also influence the outcome of the disease.
As for the acinar and portal inflammatory infiltrates, our results were characterized by mild to moderate intensity, with the main cell types identified as lymphocytes and plasma cells. In YF and DF cases, the concentration of acinar inflammatory infiltrate was more evident in Z2 than in other zones of the hepatic acinus. Given the intense apoptotic process observed in the YF samples, the values referring to inflammation were disproportionate to the intensity of the liver injury.
Our data corroborate the previous findings of YF, in which the involvement of the liver parenchyma is disproportionate to the inflammatory findings. Furthermore, as previously mentioned, the pathogenic profile of YFV is correlated with the promotion of extensive apoptosis, a process linked to the non-induction of significant inflammatory response, thus explaining the scarcity of inflammatory infiltrates [47,48,57,137].
In cases of DF, as the processes of necrosis and apoptosis were generally associated with mild to moderate intensity, the limitation of inflammatory cells in the samples is predictable. Several studies have identified that the inflammatory infiltrates in YFV and DENV infection cases consist mainly of lymphocytes [26,69,76,138]. YF cases are characterized by the predominance of mild-to-moderate lymphocytes, both in the acinar area and in the PT [77,100,101,120].
Data from the two human studies collectively propose an immunometabolic basis for symptomatic YFV infection. The anabolic demands of infection can exacerbate cellular stress and trigger ROS responses, cell death, and inflammation [139,140]. According to Chan et al., immunometabolic responses support symptomatic YF and possibly other flavivirus infections.
Although CF is characterized by extensive inflammatory responses related to musculoskeletal injuries, hepatic inflammatory involvement was not intense in our study. Studies analyzing liver tissue from human cases and animal models infected with CHIKV have shown foci of mild inflammatory infiltrates composed of mixed inflammatory cells [68,108].
Infection promoted by flaviviruses and alphaviruses is based on the manipulation of the host's metabolism through the control of the survival of infected cells, thus affecting cell death pathways. Cell death pathways can be divided into two opposing processes: accidental cell death (ACD) and regulated cell death (RCD). While ACD is a consequence of severe and rapid injury (osmotic forces, pH variations, and lytic viral replication), RCD is based on regulated molecular machinery, implying modulation by pharmacological, genetic, or infectious components [142,143].
The different cell death processes have different morphological characteristics, with three main types: apoptosis, autophagy-dependent cell death, and necrosis. Apoptosis is characterized by chromatin condensation, nuclear fragmentation, cytoplasmic shrinkage, and the formation of “apoptotic bodies”, which are subsequently destroyed by regional phagocytes [143]. Necrosis is mainly characterized by swelling, rupture of the plasma membrane, and efflux of cytoplasmic content into the extracellular environment without evident phagocytosis or lysosomal degradation by neighboring cells [142].
Autophagy-dependent cell death is defined by the distinctive features of extensive vacuolization of the cytoplasm, leading to phagocytosis and degradation by lysosomes. The triggering of this process usually takes longer, allowing a longer period of viral proliferation before cell destruction. However, in a broader context, autophagy is associated with inhibition rather than induction of cell death and is considered a mechanism for maintaining cell homeostasis after stress signals (hypoxia, ROS, PRR activation, and others) [144,145].
The activation of other described mechanisms of non-apoptotic RCD is directly linked to viral load during infection. Necroptosis, which has morphological characteristics similar to necrosis, is an independent process of caspases [146]. Pyroptosis is a highly inflammatory form of lytic RCD that is dependent on the formation of pores in the plasma membrane by members of the gasdermin (GSDM), protein family which generally correlates with the activation of inflammatory caspases (CASP1, 4, or 5) [142,147]. These processes have been described in infections by intracellular pathogens, and are related to the induction of an antimicrobial response and involvement in viral pathogenesis [148,149].
Viral cytopathic mechanisms develop through the interaction between viral molecular components and cellular machinery. Flaviviruses and alphaviruses induce, activate, and inhibit several antiviral response mechanisms, including the development of different cell death processes, influencing the different morphological characteristics presented by cases of YFV, DENV, and CHIKV infection.
Finally, our results indicated differences in the patterns of histopathological alterations in human liver samples from fatal infections by YFV, DENV, and CHIKV, related to hepatic acinus, portal tract, and inflammatory infiltrate, with intensity variation among the studied arboviruses and predominance of lesions in certain areas of the liver parenchyma, resulting in tissue damage and liver dysfunction (Figure 12).
5. Conclusions
The cases of YFV, DENV, and CHIKV infections promoted different intensities of hepatic lesions in the analyzed hepatic acinus zones (Z1, Z2, and Z3). There was a predominance of alterations in the hepatic acinus among the analyzed areas, referring to the midzonal area (Z2) in YF, DF, and CF. Among the arboviruses studied, hepatic involvement in cases of YFV infection was more intense, characterizing the classic hepatic profile of the disease.
In the semiquantitative evaluation, high-intensity cell swelling, microvesicular steatosis, and apoptosis were observed, classified as intense to very intense. Regarding other histopathological changes evaluated in the hepatic acinus, macrogoticular steatosis, lytic/coagulative necrosis, and Kupffer cell hyperplasia/hypertrophy ranged from mild to moderate.
From the endothelial alterations in the hepatic acinus of the cases of YF, DF, and CF, there was a variation from mild to moderate, with the highlights of the cases of YF and CF presenting a higher frequency of moderate intensity. In PT, there was also a variation in intensity from mild to moderate, emphasizing edema and alteration of the portal vein, with all arboviruses presenting a moderate degree of edema. DF cases had relatively high values in portal vein alteration compared to YF and CF.
Infection by YFV, DENV, and CHIKV was accompanied by a mild-to-moderate inflammatory infiltrate consisting predominantly of lymphocytes and plasma cells, especially in cases of YF and DF, with a degree of intensity disproportionate to the involvement of the hepatic parenchyma. These data substantially contribute to understanding the pathogenesis of these arboviruses and the process of hepatotropic injury developed during infections, contributing to the improvement of diagnostic criteria for diseases and expanding our knowledge about the pathogenesis of YFV, DENV, and CHIKV in human liver samples.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Normal negative controls of liver samples with molecular and immunohistochemical tests for the main hepatotropic viruses, such as hepatitis A, B, C and D viruses and with preserved histological morphology, Figure S1: Negative controls.
Author Contributions
Designed the study, Y.P.R., L.F.M.F., V.C.S., J.R.d.S., C.P., E.C.S.F., A.C.R.C., J.O.C., L.C.M., J.A.L.N., F.S.S.V., L.C.S., M.P.F., H.T.F., M.V.B.F. L.D.C., M.I.S.D., T.L.S.A., I.P.F., J.A.S.Q. and P.F.d.C.V.; performed lab tests, Y.P.R., L.F.M.F., V.C.S., J.R.d.S., C.P., E.C.S.F., A.C.R.C., J.O.C., L.C.M., J.A.L.N., F.S.S.V., L.C.S., M.P.F., H.T.F., M.V.B.F. L.D.C., M.I.S.D., T.L.S.A. and I.P.F.; furnished reagents, A.J.M.F., J.A.S.Q., and P.F.d.C.V.; drafted the manuscript, M.I.S.D., T.L.S.A., P.F.d.C.V and J.A.S.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Institute of Science and Technology for Emerging and Reemerging Viruses – INCT-VER/CNPq–406360/2022-7. National Council for Scientific and Technological Development (CNPq)/Brazil - 308600/2022-3 (JASQ).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of Instituto Evandro Chagas (protocol code number 2.364.226 in 11 June 2020).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The database used and/or analysed during the current study is not publicly accessible but can be available, upon reasonable request, from the corresponding authors.
Acknowledgments
The authors are grateful to PAPQ program (UFPA).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Young PR. Arboviruses: A Family on the Move. Adv Exp Med Biol. 2018, 1062, 1-10. [CrossRef]
- Halstead SB. Travelling arboviruses: A historical perspective. Travel Med Infect Dis. 2019, 31, 101471. [CrossRef]
- Huang Y-J, Higgs S, Vanlandingham DL. Arbovirus-Mosquito Vector-Host Interactions and the Impact on Transmission and Disease Pathogenesis of Arboviruses. Front Microbiol. 2019, 10, 22. [CrossRef]
- Braack L, Gouveia de Almeida AP, Cornel AJ, Swanepoel R, de Jager C. Mosquito-borne arboviruses of African origin: review of key viruses and vectors. Parasit Vectors. 2018, 11, 29. [CrossRef]
- Findlater A, Bogoch II. Human Mobility and the Global Spread of Infectious Diseases: A Focus on Air Travel. Trends Parasitol. 2018, 34, 9, 772-783. [CrossRef]
- Ushijima Y, Abe H, Ondo GN, et al. Surveillance of the major pathogenic arboviruses of public health concern in Gabon, Central Africa: increased risk of West Nile virus and dengue virus infections. BMC Infect Dis. 2021, 21, 1, 265. [CrossRef]
- Girard M, Nelson CB, Picot V, Gubler DJ. Arboviruses: A global public health threat. Vaccine. 2020, 38, 24, 3989-3994. [CrossRef]
- Jing QL, Cheng Q, Marshall JM, Hu WB, Yang ZC, Lu JH. Imported cases and minimum temperature drive dengue transmission in Guangzhou, China: evidence from ARIMAX model. Epidemiol Infect. 2018, 146, 10, 1226-1235. [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV) Virus Taxonomy Profile: Flaviviridae, Journal of General Virology. 2019. https://pubmed.ncbi.nlm.nih.gov/28218572/.
- International Committee on Taxonomy of Viruses (ICTV) Virus Taxonomy Profile: Togaviridae, Journal of General Virology. 2018. https://pubmed.ncbi.nlm.nih.gov/29745869/.
- Pan American Health Organization/World Health Organization, “Epidemiological Update Yellow Fever 6 March 2019” (PAHO/WHO, 2019). https://www3.paho.org/hq/index.php?option=com_topics&view=rdmore&cid=2194&Itemid=40%20784&lang=pt.
- Monath TP, Vasconcelos PFC. Yellow fever. J Clin Virol. 2015, 64, 160-173. [CrossRef]
- Costa RL, Voloch CM, Schrago CG. Comparative evolutionary epidemiology of dengue virus serotypes. Infect Genet Evol. 2012, 12, 2, 309-314. [CrossRef]
- Centers for Disease Control and Prevention (CDC). National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), Division of Vector-Borne Diseases (DVBD). 2019. https://www.cdc.gov/dengue/index.html.
- Romero D, Olivero J, Real R, Guerrero JC. Applying fuzzy logic to assess the biogeographical risk of dengue in South America. Parasit Vectors. 2019, 12, 1, 428. [CrossRef]
- Hamlet A, Jean K, Perea W, et al. The seasonal influence of climate and environment on yellow fever transmission across Africa. PLoS Negl Trop Dis. 2018, 12, 3. [CrossRef]
- Souza-Neto JA, Powell JR, Bonizzoni M. Aedes aegypti vector competence studies: A review. Infect Genet Evol. 2019, 67, 191-209. [CrossRef]
- Goes de Jesus J, Gräf T, Giovanetti M, et al. Yellow fever transmission in non-human primates, Bahia, Northeastern Brazil. PLoS Negl Trop Dis. 2020, 14, 8. [CrossRef]
- Auguste AJ, Lemey P, Bergren NA, et al. Enzootic Transmission of Yellow Fever Virus, Venezuela. Emerg Infect Dis. 2015, 21, 1, 99-102. [CrossRef]
- Ministry of Health of Brazil. Health Surveillance Department. Epidemiological surveillance guide. (6th ed. Brasilia: Ministry of Health). Series A. Standards and Technical Manuals. 2005. https://bvsms.saude.gov.br/bvs/publicacoes/health_brazil_2015_2016.pdf.
- Ministry of Health of Brazil. Dengue Epidemiological Aspects, Diagnosis and Treatment. (Brasilia: Ministry of Health). 2019. https://bvsms.saude.gov.br/bvs/publicacoes/dengue_aspecto_epidemiologicos_diagnostico_tratamento.pdf.
- Ministry of Health of Brazil. Monitoring of cases of urban arboviruses caused by viruses transmitted by Aedes (dengue, chikungunya and zika), epidemiological weeks 1 to 53, 2020. (Brasilia: Ministry of Health). 2021. https://www.gov.br/saude/pt-br/centrais-de-conteudo/publicacoes/boletins/boletins-epidemiologicos/edicoes/2020/boletim_epidemiologico_svs_48.pdf.
- Lovera D, Martínez-Cuellar C, Galeano F, Amarilla S, Vazquez C, Arbo A. Clinical manifestations of primary and secondary dengue in Paraguay and its relation to virus serotype. J Infect Dev Ctries. 2019, 13, 12, 1127-1134. [CrossRef]
- Oliveira LG, Peron JPS. Viral receptors for flaviviruses: Not only gatekeepers. J Leukoc Biol. 2019, 106, 3, 695-701. [CrossRef]
- Kril V, Aïqui-Reboul-Paviet O, Briant L, Amara A. New Insights into. Chikungunya Virus Infection and Pathogenesis. Annu Rev Virol. 2021, 8, 1, 327-347. [CrossRef]
- Quaresma JAS, Pagliari C, Medeiros DBA, Duarte MIS, Vasconcelos PFC. Immunity and immune response, pathology and pathologic changes: progress and challenges in the immunopathology of yellow fever. Rev Med Virol. 2013, 23, 5, 305-318. [CrossRef]
- Chen Z, Liu L, Lv Y, et al. A fatal yellow fever virus infection in China: description and lessons. Emerg Microbes Infect. 2016, 5, 7. [CrossRef]
- Yung C-F, Lee K-S, Thein T-L, et al. Dengue serotype-specific differences in clinical manifestation, laboratory parameters and risk of severe disease in adults, Singapore. Am J Trop Med Hyg. 2015, 92, 5, 999-1005. [CrossRef]
- Honório NA, Câmara DCP, Calvet GA, Brasil P. Chikungunya: an arbovirus infection in the process of establishment and expansion in Brazil. Rep Public Health. 2015, 31, 1-3. [CrossRef]
- Gasque P, Bandjee MCJ, Reyes MM, Viasus D. Chikungunya Pathogenesis: From the Clinics to the Bench. J Infect Dis. 2016, 214(suppl 5), S446-S448. [CrossRef]
- Póvoa TF, Alves AMB, Oliveira CAB, Nuovo GJ, Chagas VLA, Paes MV. The Pathology of Severe Dengue in Multiple Organs of Human Fatal Cases: Histopathology, Ultrastructure and Virus Replication. PLoS One. 2014, 9, 4, e83386. [CrossRef]
- Kularatne SAM, Rajapakse MM, Ralapanawa U, Waduge R, Pathirage LPMMK, Rajapakse RPVJ. Heart and liver are infected in fatal cases of dengue: three PCR based case studies. BMC Infect Dis. 2018, 18, 681. [CrossRef]
- Mercado-Reyes M, Acosta-Reyes J, Navarro-Lechuga E, et al. Dengue, chikungunya and zika virus coinfection: results of the national surveillance during the zika epidemic in Colombia. Epidemiol Infect. 2019, 147, e77, 1-7. [CrossRef]
- Sharp TM, Keating MK, Shieh W-J, et al. Clinical Characteristics, Histopathology, and Tissue Immunolocalization of Chikungunya Virus Antigen in Fatal Cases. Clin Infect Dis. 2021, 73, 2, e345-e354. [CrossRef]
- Ustafa M, Illzam EM, Jeffree MS, et al. Dengue Fever: Clinical Spectrum, And Management. IOSR-JDMS. 2017, 16, 53-59. [CrossRef]
- Chia PY, Thein T-L, Ong SWX, Lye DC, Leo YS. Severe dengue and liver involvement: an overview and review of the literature. Expert Rev Anti Infect Ther. 2020, 18, 3, 181-189. [CrossRef]
- Dissanayake HA, Seneviratne SL. Liver involvement in dengue viral infections. Rev Med Virol. 2018, 28, 2, e1971. [CrossRef]
- Valentine MJ, Murdock CC, Kelly PJ. Sylvatic cycles of arboviruses in non-human primates. Parasit Vectors. 2019, 12, 1, 463. [CrossRef]
- Kerkhof K, Falconi-Agapito F, Van Esbroeck M, Talledo M, Ariën KK. Reliable Serological Diagnostic Tests for Arboviruses: Feasible or Utopia? Trends Microbiol. 2020, 28, 4, 276-292. [CrossRef]
- Tadeu de Araújo LJ, Gonzalez LL, Buss LF, et al. Surveillance of hemorrhagic fever and/or neuroinvasive disease: challenges of diagnosis. J Public Health. 2021, 55, 41. [CrossRef]
- Fischer C, Jo WK, Haage V, Moreira-Soto A, Filho EFO, Drexler JF. Challenges towards serologic diagnostics of emerging arboviroses. Clin Microbiol Infect. 2021, 27, 9, 1221-1229. [CrossRef]
- Mori A, Pomari E, Deiana M, et al. Molecular techniques for the genomic viral RNA detection of West Nile, Dengue, Zika and Chikungunya arboviruses: a narrative review. Expert Rev Mol Diagn. 2021, 21, 6, 591-612. [CrossRef]
- Murillo DFB, Piche-Ovares M, Gamboa-Solano JC, et al. Serological Positivity against Selected Flaviviruses and Alphaviruses in Free-Ranging Bats and Birds from Costa Rica Evidence Exposure to Arboviruses Seldom Reported Locally in Humans. Viruses. 2022, 14, 1, 93. [CrossRef]
- Campos S, Figueredo Thiel S, Bellassai J, Rodríguez I. Immunohistochemistry in the diagnosis of yellow fever. Experience in the Department of Pathology, Health Sciences Research Institute and the Department of Pathology, Faculty of Medical Sciences, National University of Asuncion. Mem Inst Investig Cienc Salud. 2009, 7, 33-39.
- Melo-Lima BL, Espósito DLA, Lopes da Fonseca BA, Figueiredo LTM, Moreau P, Donadi EA. The Attenuated Live Yellow Fever Virus 17D Infects the Thymus and Induces Thymic Transcriptional Modifications of Immunomodulatory Genes in C57BL/6 and BALB/C Mice. Autoimmune Dis. 2015, 1-12, . [CrossRef]
- Tesh RB, Guzman H, da Rosa AP, et al. Experimental yellow fever virus infection in the Golden Hamster (Mesocricetus auratus). I. Virologic, biochemical, and immunologic studies. J Infect Dis. 2001, 183, 10, 1431-1436. [CrossRef]
- Quaresma JAS, Barros VLRS, Pagliari C, et al. Revisiting the liver in human yellow fever: virus-induced apoptosis in hepatocytes associated with TGF-beta, TNF-alpha and NK cells activity. Virology. 2006, 345, 1, 22-30. [CrossRef]
- Quaresma JAS, Barros VLRS, Fernandes ER, et al. Immunohistochemical examination of the role of Fas ligand and lymphocytes in the pathogenesis of human liver yellow fever. Virus Res. 2006, 116, 1-2, 91-97. [CrossRef]
- Idirisinghe KAP. Histopathological study of Dengue Haemorrhagic Fever. J Diagn Pathol. 2014, 8, 50–58. [CrossRef]
- Rathi KR, Arora MM, Sahai K, et al. Autopsy findings in fatal dengue haemorrhagic fever - 06 Cases. Med J Armed Forces India. 2013, 69, 3, 254–259. [CrossRef]
- Leal SG, Romano APM, Monteiro RV, Barros de Melo C, Vasconcelos PFC, Botelho de Castro M. Frequency of histopathological changes in Howler monkeys (Alouatta sp.) naturally infected with yellow fever virus in Brazil. J Braz Soc Trop Med. 2016, 49, 1, 29-33. [CrossRef]
- Begum F, Das S, Mukherjee D, Mal S, Ray U. Insight into the Tropism of Dengue Virus in Humans. Viruses. 2019, 11, 12, 1136. [CrossRef]
- Devarbhavi H, Ganga D, Menon M, Kothari K, Singh R. Dengue hepatitis with acute liver failure: Clinical, biochemical, histopathological characteristics and predictors of outcome. J Gastroent Hepat. 2020, 35, 7, 1223-1228. [CrossRef]
- Dhanoa A, Hassan SS, Ngim CF, et al. Impact of dengue virus (DENV) co-infection on clinical manifestations, disease severity and laboratory parameters. BMC Infect Dis. 2016, 16, 1, 406. [CrossRef]
- Londono-Renteria B, Marinez-Angarita JC, Troupin A, Colpitts TM. Role of Mast Cells in Dengue Virus Pathogenesis. DNA Cell Biol. 2017, 36, 6, 423-427. [CrossRef]
- Gaythorpe KA, Hamlet A, Jean K, et al. The global burden of yellow fever. Elife. 2021, 10:e64670. [CrossRef]
- Quaresma JA, Barros VL, Fernandes ER, et al. Reconsideration of histopathology and ultrastructural aspects of the human liver in yellow fever. Acta Trop. 2005, 94, 116-127. [CrossRef]
- Azevedo RSS, Araujo MT, Oliveira CS, et al. Zika Virus Epidemic in Brazil. II. Post-Mortem Analyses of Neonates with Microcephaly, Stillbirths, and Miscarriage. J Clin Med. 2018, 7, 496. [CrossRef]
- Goodman AG, Rasmussen AL. Editorial: Host-Pathogen Interactions During Arboviral Infections. Front Cell Infect Microbiol. 2019, 9, 77. [CrossRef]
- Her Z, Malleret B, Chan M, et al. Active infection of human blood monocytes by Chikungunya virus triggers an innate immune response. J Immunol. 2010, 184, 10, 5903-13. [CrossRef]
- Akhrymuk I, Kulemzin SV, Frolova EI. Evasion of the Innate Immune Response: the Old World Alphavirus nsP2 Protein Induces Rapid Degradation of Rpb1, a Catalytic Subunit of RNA Polymerase II. J Virol. 2012, 86, 13, 7180-7191. [CrossRef]
- Randall G. Lipid Droplet Metabolism during Dengue Virus Infection. Trends Microbiol. 2018, 26, 8, 640-642. [CrossRef]
- Zhang J, Lan Y, Li MY, et al. Flaviviruses Exploit the Lipid Droplet Protein AUP1 to Trigger Lipophagy and Drive Virus Production. Cell Host Microbe. 2018, 23, 819-831.e5. [CrossRef]
- Acharya B, Gyeltshen S, Chaijaroenkul W, Na-Bangchang K. Significance of Autophagy in Dengue Virus Infection: A Brief Review. Am J Trop Med Hyg. 2019, 100, 4, 783-790. [CrossRef]
- Akhrymuk I, Lukash T, Frolov I, Frolova EI. Novel Mutations in nsP2 Abolish Chikungunya Virus-Induced Transcriptional Shutoff and Make the Virus Less Cytopathic without Affecting Its Replication Rates. J Virol. 2019, 93, 4, e02062-18. [CrossRef]
- Bovay A, Marraco SAF, Speiser DE. Yellow fever virus vaccination: an emblematic model to elucidate robust human immune responses. Hum Vaccin Immunother. 2021, 17, 8, 2471-2481. [CrossRef]
- Schilte C, Couderc T, Chretien F, et al. Type I IFN controls chikungunya virus via its action on nonhematopoietic cells. J Exp Med. 2010, 207, 2, 429–442. [CrossRef]
- Mercado M, Acosta-Reyes J, Parra E, et al. Clinical and histopathological features of fatal cases with dengue and chikungunya virus co-infection in Colombia, 2014 to 2015. Euro Surveill. 2016, 21, 22. [CrossRef]
- Lin C-F, Wan S-W, Chen M-C, et al. Liver injury caused by antibodies against dengue virus nonstructural protein 1 in a murine model. Lab Invest. 2008, 88, 10, 1079-1089. [CrossRef]
- Paes MV, Lenzi HL, Nogueira ACM, et al. Hepatic damage associated with dengue-2 virus replication in liver cells of BALB/c mice. Lab Invest. 2009, 89, 10, 1140-1151. [CrossRef]
- Wang J, Chen Y, Gao N, et al. Inhibitory effect of glutathione on oxidative liver injury induced by dengue virus serotype 2 infections in mice. PLoS One. 2013, 8, 1, e55407. [CrossRef]
- Engelmann F, Josset L, Girke T, et al. Pathophysiologic and Transcriptomic Analyses of Viscerotropic Yellow Fever in a Rhesus Macaque Model. PLoS Negl Trop Dis. 2014, 8, 11, e3295. [CrossRef]
- Drumond BP, Fagundes LGS, Rocha RP, et al. Phylogenetic analysis of Dengue virus 1 isolated from South Minas Gerais, Brazil. Braz J Microbiol. 2016, 47, 1, 251-258. [CrossRef]
- Sakinah S, Priya SP, Kumari S, et al. Impact of dengue virus (serotype DENV-2) infection on liver of BALB/c mice: A histopathological analysis. Tissue Cell. 2017, 49, 1, 86-94. [CrossRef]
- Nogueira RMR, Schatzmayr HG, Bispo de Filippis AM, et al. Dengue Virus Type 3, Brazil, 2002. Emerg Infect Dis. 2005, 11, 9, 1376–1381. [CrossRef]
- Dias Jr LB, Alves VAF, Kanamura C, Oikawa RTC, Wakamatsu A. Fulminant hepatic failure in northern Brazil: morphological, immunohistochemical and pathogenic aspects of Lábrea hepatitis and yellow fever. Trans R Soc Trop Med Hyg. 2007, 101, 8, 831-839. [CrossRef]
- Pierrotti LC, Duarte-Neto AN, Song ATW, Ventura CG, David-Neto E, Azevedo LS. Fatal Yellow Fever in a Kidney Transplant Patient. Clin Infect Dis. 2020, 70, 1, 144-148. [CrossRef]
- Olímpio FA, Falcão LFM, Carvalho MLG, et al. Endothelium Activation during Severe Yellow Fever Triggers an Intense Cytokine-Mediated Inflammatory Response in the Liver Parenchyma. Pathogens. 2022, 11, 1, 101. [CrossRef]
- Pagliari C, Quaresma JAS, Fernandes ER, et al. Immunopathogenesis of dengue hemorrhagic fever: contribution to the study of human liver lesions. J Med Virol. 2014, 86, 7, 1193-1197. [CrossRef]
- Rivera JA, Rengifo AC, Parra ÉA, Castellanos JE, Caldas ML. Illustrated histopathological features of fatal dengue cases in Colombia. Biomedica. 2020, 40, 3, 438-447. [CrossRef]
- Win MM, Charngkaew K, Punyadee N, et al. Ultrastructural Features of Human Liver Specimens from Patients Who Died of Dengue Hemorrhagic Fever. Trop Med Infect Dis. 2019, 4, 2, 63. [CrossRef]
- Cui L, Lee YH, Kumar Y, et al. Serum Metabolome and Lipidome Changes in Adult Patients with Primary Dengue Infection. PLoS Negl Trop Dis. 2013, 7, 8, e2373. [CrossRef]
- Murakami M. Bioactive Lipid Mediators: Current Reviews and Protocols. Tokyo: Springer Japan. 2015.
- Melo CFOR, Noin de Oliveira D, Lima EO, et al. A Lipidomics Approach in the Characterization of Zika-Infected Mosquito Cells: Potential Targets for Breaking the Transmission Cycle. PLoS One. 2016, 11, 10, e0164377. [CrossRef]
- Gong W, Jia J, Zhang B, et al. Serum Metabolomic Profiling of Piglets Infected with Virulent Classical Swine Fever Virus. Front Microbiol. 2017, 8, 731. [CrossRef]
- Melo CFOR, Delafiori J, Dabaja MZ, et al. The role of lipids in the inception, maintenance and complications of dengue virus infection. Sci Rep. 2018, 8, 1, 11826. [CrossRef]
- Singh R, Kaushik S, Wang Y, et al. Autophagy regulates lipid metabolism. Nature. 2009, 458 (7242), 1131-1135. [CrossRef]
- Heaton NS, Randall G. Dengue virus-induced autophagy regulates lipid metabolism. Cell Host Microbe. 2010, 8, 5, 422-432. [CrossRef]
- Sagan SM, Rouleau Y, Leggiadro C, et al. The influence of cholesterol and lipid metabolism on host cell structure and hepatitis C virus replication. Biochem Cell Biol. 2006, 4, 1, 67-79. [CrossRef]
- Albulescu L, Wubbolts R, Van Kuppeveld FJM, Strating JRPM. Cholesterol shuttling is important for RNA replication of coxsackievirus B3 and encephalomyocarditis virus. Cell Microbiol. 2015, 17, 8, 1144-1156. [CrossRef]
- Zhang J, Zhang Z, Chukkapalli V, et al. Positive-strand RNA viruses stimulate host phosphatidylcholine synthesis at viral replication sites. Proc Natl Acad Sci U S A. 2016, 113, 8, E1064- E1073. [CrossRef]
- Stoeck IK, Lee J-Y, Tabata K, et al. Hepatitis C Virus Replication Depends on Endosomal Cholesterol Homeostasis. J Virol. 2017, 92, 1, e01196-17. [CrossRef]
- Hong C, Tontonoz P. Liver X receptors in lipid metabolism: opportunities for drug Discovery. Nat Rev Drug Discov. 2014, 13, 6, 433-444. [CrossRef]
- Howe V, Sharpe LJ, Alexopoulos SJ, et al. Cholesterol homeostasis: How do cells sense sterol excess? Chem Phys Lipids. 2016, 199, 170-178. [CrossRef]
- Hwang J, Wang Y, Fikrig E. Inhibition of Chikungunya Virus Replication in Primary Human Fibroblasts by Liver X Receptor Agonist. Antimicrob Agents Chemother. 2019, 63, 9, 01220-19. [CrossRef]
- Asselah T, Rubbia-Brandt L, Marcellin P, Negro F. Steatosis in chronic hepatitis C: why does it really matter? Gut. 2006, 55, 1, 123-130. [CrossRef]
- Petta S, Amato M, Cabibi D, et al. Visceral adiposity index is associated with histological findings and high viral load in patients with chronic hepatitis C due to genotype 1. Hepatology. 2010, 52, 5, 1543-1552. [CrossRef]
- Boeckmans J, Rombaut M, Demuyser T, et al. Infections at the nexus of metabolic-associated fatty liver disease. Arch Toxicol. 2021, 95, 7, 2235-2253. [CrossRef]
- França RFO, Zucoloto S, Lopes da Fonseca BA. A BALB/c mouse model shows that liver involvement in dengue disease is immune-mediated. Exp Mol Pathol. 2010, 89, 3, 321-326. [CrossRef]
- Mascheretti M, Tengan CH, Sato HK, et al. Yellow fever: reemerging in the state of Sao Paulo, Brazil, 2009. Public Health Journal. 2013, 47, 5. [CrossRef]
- Guarner J, Hale GL. Four human diseases with significant public health impact caused by mosquito-borne flaviviruses: West Nile, Zika, dengue and yellow fever. Semin Diagn Pathol. 2019, 36, 3, 170-176. [CrossRef]
- Casadio L, Nastri AC, Malta FM, et al. Late-Onset Relapsing Hepatitis Associated with Yellow Fever. N Engl J Med. 2020, 382, 21, 2059-2061. [CrossRef]
- Pan Y, Cheng A, Wang M, Yin Z, Jia R. The Dual Regulation of Apoptosis by Flavivirus. Front Microbiol. 2021, 12, 654494. [CrossRef]
- Paes MV, Pinhão AT, Barreto DF, et al. Liver injury and viremia in mice infected with dengue-2 virus. Virology. 2005, 338, 2, 236-246. [CrossRef]
- Wang E, Volkova E, Adams AP, et al. Chimeric Alphavirus Vaccine Candidates for Chikungunya. Vaccine. 2008, 26, 39, 5030-5039. [CrossRef]
- Ziegler SA, Lu L, Travassos da Rosa APA, Xiao S-Y, Tesh RB. An animal model for studying the pathogenesis of chikungunya virus infection. Am J Trop Med Hyg. 2008, 79, 1, 133-139.
- Mallilankaraman K, Shedlock DJ, Bao H, et al. A DNA vaccine against chikungunya virus is protective in mice and induces neutralizing antibodies in mice and nonhuman primates. PLoS Negl Trop Dis. 2011, 5, 1, e928. [CrossRef]
- Agarwal A, Joshi G, Nagar DP, et al. Mosquito saliva induced cutaneous events augment Chikungunya virus replication and disease progression. Infect Genet Evol. 2016, 40, 126-135. [CrossRef]
- Sitia G, Iannacone M, Aiolfi R, et al. Kupffer Cells Hasten Resolution of Liver Immunopathology in Mouse Models of Viral Hepatitis. PLoS Pathog. 2011, 7, 6, e1002061. [CrossRef]
- Leowattana W, Leowattana T. Dengue hemorrhagic fever and the liver. World J Hepatol. 2021, 13, 1968–1976. [CrossRef]
- Bailey AL, Kang L-I, Zanella LGFAB, et al. Consumptive coagulopathy of severe yellow fever occurs independently of hepatocellular tropism and massive hepatic injury. Proc Natl Acad Sci U S A. 2020, 117, 51, 32648-32656. [CrossRef]
- Tuboi SH, Costa ZGA, Vasconcelos PFC, Hatch D. Clinical and epidemiological characteristics of yellow fever in Brazil: analysis of reported cases 1998-2002. Trans R Soc Trop Med Hyg. 2007, 101, 2, 169-175. [CrossRef]
- Holz LE, Bowen DG, Bertolino P. Mechanisms of T cell death in the liver: to Bim or not to Bim? Dig Dis. 2010, 28, 1, 14-24. [CrossRef]
- Meertens L, Carnec X, Lecoin MP, et al. The TIM and TAM Families of Phosphatidylserine Receptors Mediate Dengue Virus Entry. Cell Host Microbe. 2012, 12, 4, 544-557. [CrossRef]
- Moller-Tank S, Kondratowicz AS, Davey RA, Rennert PD, Maury W. Role of the phosphatidylserine receptor TIM-1 in enveloped-virus entry. J Virol. 2013, 87, 15, 8327-8341. [CrossRef]
- Jemielity S, Wang JJ, Chan YK, et al. TIM-family proteins promote infection of multiple enveloped viruses through virion-associated phosphatidylserine. PLoS Pathog. 2013, 9, 3, e1003232. [CrossRef]
- Perera-Lecoin M, Meertens L, Carnec X, Amara A. Flavivirus Entry Receptors: An Update. Viruses. 2014, 6, 1, 69-88. [CrossRef]
- Morizono K, Chen ISY. Role of Phosphatidylserine Receptors in Enveloped Virus Infection. J Virol. 2014, 88, 8, 4275-4290. [CrossRef]
- Amara A, Mercer J. Viral apoptotic mimicry. Nat Rev Microbiol. 2015, 13, 8, 461-469. [CrossRef]
- Duarte-Neto AN, Cunha MDP, Marcilio I, et al. Yellow fever and orthotopic liver transplantation: new insights from the autopsy room for an old but re-emerging disease. Histopathology. 2019, 75, 5, 638-648. [CrossRef]
- Smith DR, Khakpoor A. Involvement of the liver in dengue infections. Dengue Bulletin. 2009, 33, 75-86.
- Aye KS, Charngkaew K, Win N, et al. Pathologic highlights of dengue hemorrhagic fever in 13 autopsy cases from Myanmar. Hum Pathol. 2014, 45, 6, 1221-1233. [CrossRef]
- Huerre MR, Lan NT, Marianneau P, et al. Liver histopathology and biological correlates in five cases of fatal dengue fever in Vietnamese children. Virchows Arch. 2001, 438, 2, 107-115. [CrossRef]
- Limonta D, Capó V, Torres G, Pérez AB, Guzmán MG. Apoptosis in tissues from fatal dengue shock syndrome. J Clin Virol. 2007, 40, 1, 50-54. [CrossRef]
- Jessie K, Fong MY, Devi S, Lam SK, Wong KT. Localization of dengue virus in naturally infected human tissues, by immunohistochemistry and in situ hybridization. J Infect Dis. 2004, 189, 8, 1411-1418. [CrossRef]
- Modhiran N, Watterson D, Muller DA, et al. Dengue virus NS1 protein activates cells via Toll-like receptor 4 and disrupts endothelial cell monolayer integrity. Sci Transl Med. 2015, 7, 304, 304ra142. [CrossRef]
- Pang X, Zhang R, Cheng G. Progress towards understanding the pathogenesis of dengue hemorrhagic fever. Virol Sin. 2017, 32, 1, 16-22. [CrossRef]
- Zellweger RM, Prestwood TR, Shresta S. Enhanced infection of liver sinusoidal endothelial cells in a mouse model of antibody-induced severe dengue disease. Cell Host Microbe. 2010, 7, 2, 128-139. [CrossRef]
- Khongphatthanayothin A, Lertsapcharoen P, Supachokchaiwattana P, et al. Hepatosplanchnic circulatory dysfunction in acute hepatic infection: the case of dengue hemorrhagic fever. Shock. 2005, 24, 5, 407-411. [CrossRef]
- Basílio-de-Oliveira CA, Aguiar GR, Baldanza MS, Barth OM, Eyer-Silva WA, Paes MV. Pathologic study of a fatal case of dengue-3 virus infection in Rio de Janeiro, Brazil. Braz J Infect Dis. 2005, 9, 4, 341-347. [CrossRef]
- Cunha MP, Duarte-Neto AN, Pour SZ, et al. Systemic dengue infection associated with a new dengue virus type 2 introduction in Brazil - a case report. BMC Infect Dis. 2021, 21, 1, 311. [CrossRef]
- Kumaria R. Correlation of disease spectrum among four Dengue serotypes: a five years hospital based study from India. Braz J Infect Dis. 2010, 14, 2, 141-146. [CrossRef]
- Chen Y-C, Lu J-W, Yeh C-T, Lin T-Y, Liu F-C, Ho Y-J. Micafungin Inhibits Dengue Virus Infection through the Disruption of Virus Binding, Entry, and Stability. Pharmaceuticals (Basel). 2021, 14, 4, 338. [CrossRef]
- Suppiah J, Ching S-M, Amin-Nordin S, et al. Clinical manifestations of dengue in relation to dengue serotype and genotype in Malaysia: A retrospective observational study. PLoS Negl Trop Dis. 2018, 12, 9, e0006817. [CrossRef]
- Halsey ES, Marks MA, Gotuzzo E, et al. Correlation of serotype-specific dengue virus infection with clinical manifestations. PLoS Negl Trop Dis. 2012, 6, 5, e1638. [CrossRef]
- Baltina LA, Tasi Y-T, Huang S-H, et al. Glycyrrhizic acid derivatives as Dengue virus inhibitors. Bioorg Med Chem Lett. 2019, 29, 20, 126645. [CrossRef]
- Monath TP. Yellow fever: an update. Lancet Infect Dis. 2001, 1, 1, 11-20. [CrossRef]
- Xavier EA, et al. Immunocompetent mice model for dengue virus infection. Scientific World Journal. 2012, 525947. [CrossRef]
- Fox CJ, Hammerman PS, Thompson CB. Fuel feeds function: energy metabolism and the T-cell response. Nat Rev Immunol. 2005, 5, 11, 844-852. [CrossRef]
- Wang A, Luan HH, Medzhitov R. An evolutionary perspective on immunometabolism. Science. 2019, 363(6423), eaar3932. [CrossRef]
- Chan KR, Gan ES, Chan CYY, et al. Metabolic perturbations and cellular stress underpin susceptibility to symptomatic live-attenuated yellow fever infection. Nat Med. 2019, 25, 8, 1218-1224. [CrossRef]
- Galluzzi L, Vitale I, Aaronson SA, et al. Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 3, 486-541. [CrossRef]
- Cappuccio L, Maisse C. Infection of Mammals and Mosquitoes by Alphaviruses: Involvement of Cell Death. Cells. 2020, 9, 12, 2612. [CrossRef]
- Boya P, González-Polo R-A, Casares N, et al. Inhibition of Macroautophagy Triggers Apoptosis. Mol Cell Biol. 2005, 25, 3, 1025-1040. [CrossRef]
- Roy SG, Sadigh B, Datan E, Lockshin RA, Zakeri Z. Regulation of cell survival and death during Flavivirus infections. World J Biol Chem. 2014, 5, 2, 93-105. [CrossRef]
- Choi ME, Price DR, Ryter SW, Choi AMK. Necroptosis: a crucial pathogenic mediator of human disease. JCI Insight. 2019, 4, 15, e128834. [CrossRef]
- Man SM, Karki R, Kanneganti T-D. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunol Rev. 2017, 277, 1, 61-75. [CrossRef]
- Vanden Berghe T, Vanlangenakker N, Parthoens E, et al. Necroptosis, necrosis and secondary necrosis converge on similar cellular disintegration features. Cell Death Differ. 2010, 17, 6, 922-930. [CrossRef]
- Kepp O, Galluzzi L, Zitvogel L, Kroemer G. Pyroptosis - a cell death modality of its kind? Eur J Immunol. 2010, 40, 3, 627-630. [CrossRef]
Figure 1.
Histopathology of hepatic infection by YFV. Where it is observed in the presence of multiple apoptotic bodies (black arrows) and mononuclear acinar inflammatory infiltrate (circle) (A). In (B), apoptotic bodies (councilman bodies) and hemorrhage are observed (black arrows). In (C) and (D), multiple apoptotic and councilman bodies and macro and microvesicular steatosis are observed (black arrows). In (E), portal tracts with inflammatory infiltrate of mononuclear cells (circle) and the presence of antigen for YFV in liver tissue marked by IHC in (F) are observed (black arrows). (Hematoxylin and Eosin, A and E: 200x; B, C, D and F: 400x).
Figure 1.
Histopathology of hepatic infection by YFV. Where it is observed in the presence of multiple apoptotic bodies (black arrows) and mononuclear acinar inflammatory infiltrate (circle) (A). In (B), apoptotic bodies (councilman bodies) and hemorrhage are observed (black arrows). In (C) and (D), multiple apoptotic and councilman bodies and macro and microvesicular steatosis are observed (black arrows). In (E), portal tracts with inflammatory infiltrate of mononuclear cells (circle) and the presence of antigen for YFV in liver tissue marked by IHC in (F) are observed (black arrows). (Hematoxylin and Eosin, A and E: 200x; B, C, D and F: 400x).
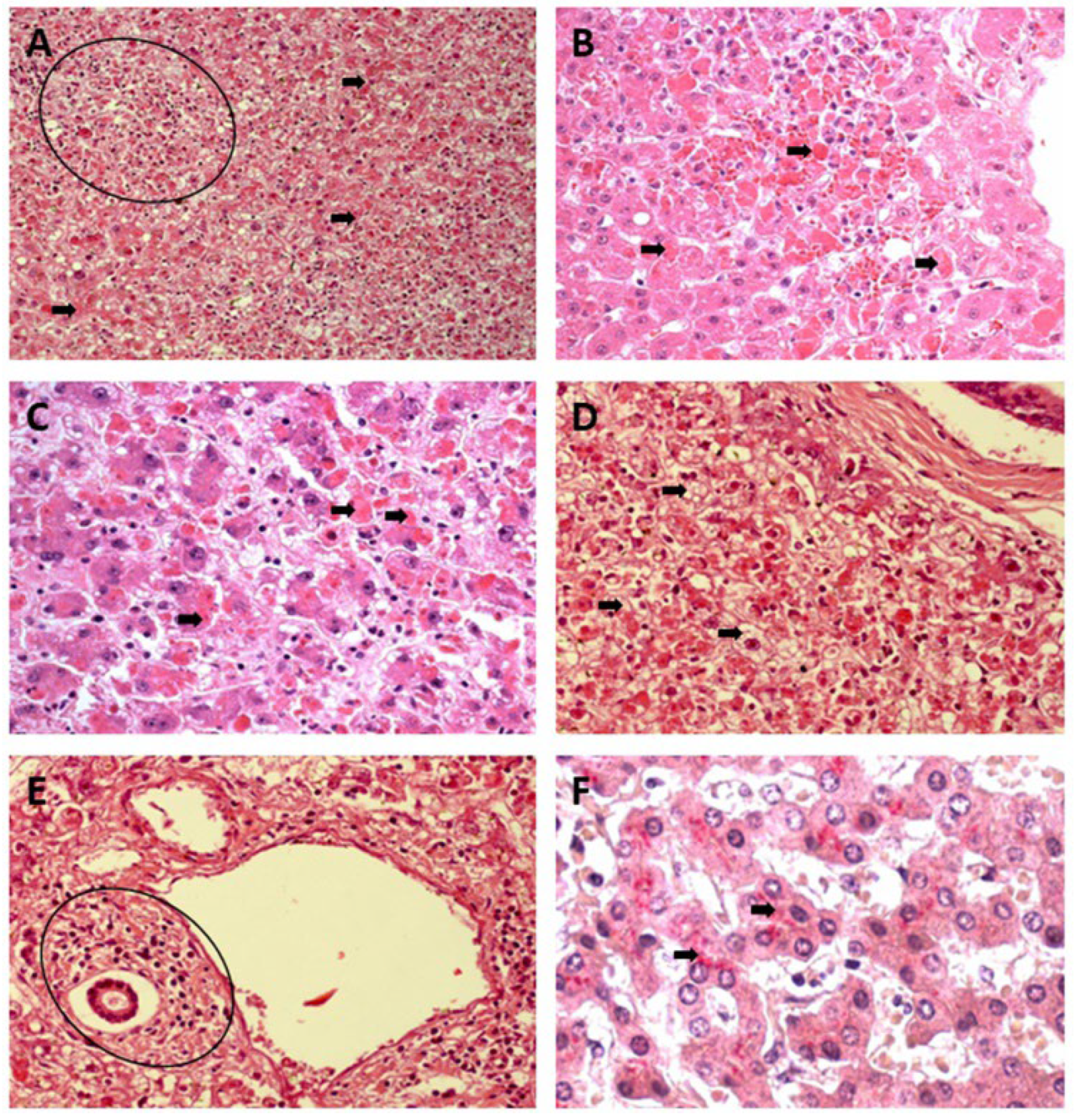
Figure 2.
Histopathology of hepatic infection by DENV. Where it is observed in the presence of mild cell swelling (black arrows) (A) associated with a mononuclear inflammatory infiltrate (circle) in (B) and the presence of macrovesicular steatosis (black arrows) and apoptosis patterns (open arrows) in (C) and (D). Changes in endothelial cells associated with the inflammatory response of mononuclear cells (black arrows) in (D). Hepatocytic ballooning and lytic necrosis (black arrows) of hepatocytes in (E) and immunostaining by IHC by alkaline phosphatase (black arrows) in (F) in liver tissue. (Hematoxylin and Eosin, A and B: 200x; C – F: 400x).
Figure 2.
Histopathology of hepatic infection by DENV. Where it is observed in the presence of mild cell swelling (black arrows) (A) associated with a mononuclear inflammatory infiltrate (circle) in (B) and the presence of macrovesicular steatosis (black arrows) and apoptosis patterns (open arrows) in (C) and (D). Changes in endothelial cells associated with the inflammatory response of mononuclear cells (black arrows) in (D). Hepatocytic ballooning and lytic necrosis (black arrows) of hepatocytes in (E) and immunostaining by IHC by alkaline phosphatase (black arrows) in (F) in liver tissue. (Hematoxylin and Eosin, A and B: 200x; C – F: 400x).
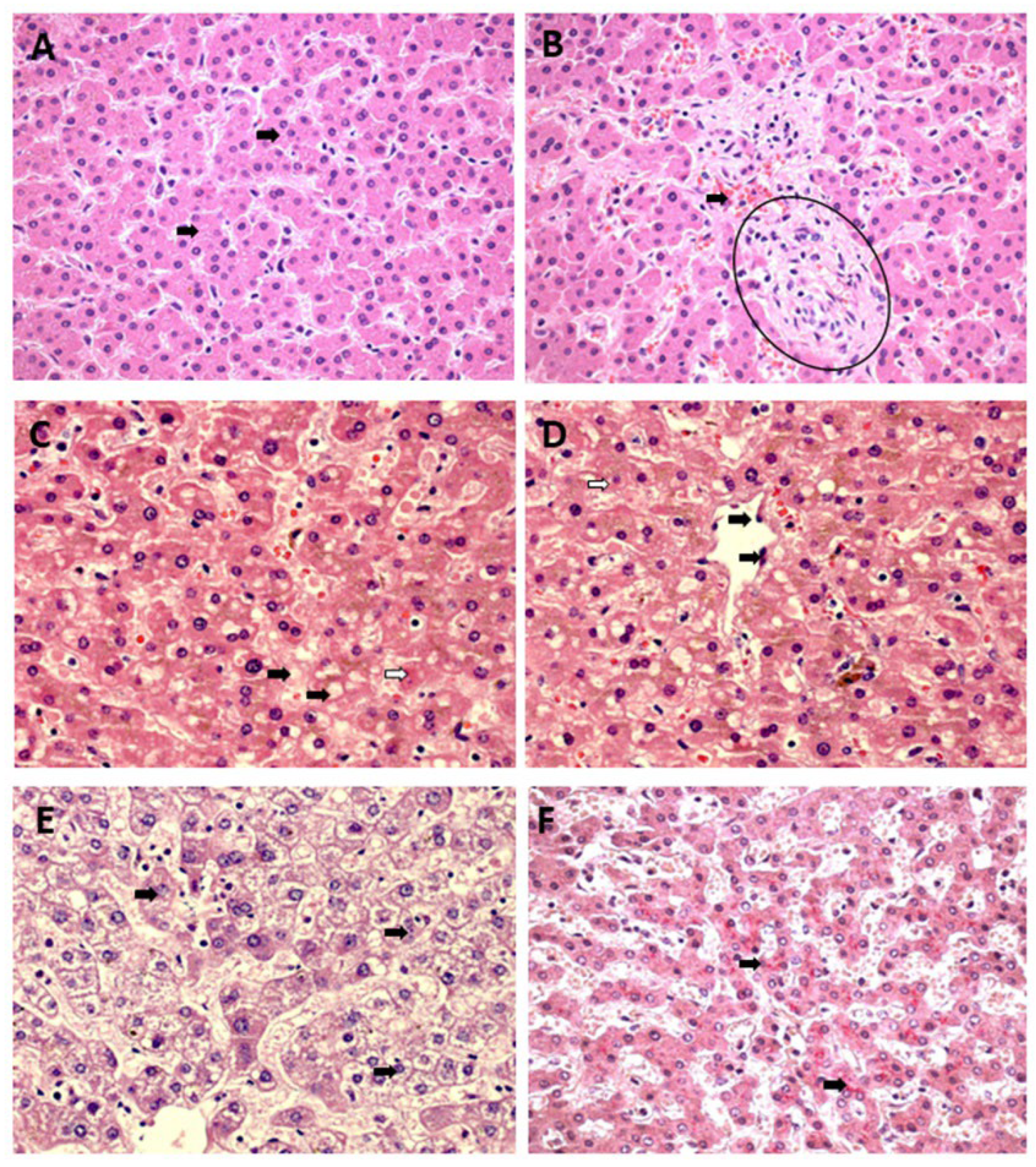
Figure 3.
Histopathology of hepatic infection by CHIKV. Where the presence of macrovesicular steatosis (black arrows) (A) in the midst of sinusoidal congestion (open arrows) and cell swelling (open arrows) is observed in (B) and the presence of macro (black arrows) and microdroplet (open arrows) steatosis patterns of steatosis in (C). Figures of cells with morphology in apoptosis (black arrows) and necrosis (open arrows) in (D) associated with the response mononuclear inflammatory disease (black arrows and circle) in (E) and the presence of immunostaining for the virus in tissue by IHC by alkaline phosphatase (black arrows) in (F). (Hematoxylin and Eosin, A: 200x; B – F: 400x).
Figure 3.
Histopathology of hepatic infection by CHIKV. Where the presence of macrovesicular steatosis (black arrows) (A) in the midst of sinusoidal congestion (open arrows) and cell swelling (open arrows) is observed in (B) and the presence of macro (black arrows) and microdroplet (open arrows) steatosis patterns of steatosis in (C). Figures of cells with morphology in apoptosis (black arrows) and necrosis (open arrows) in (D) associated with the response mononuclear inflammatory disease (black arrows and circle) in (E) and the presence of immunostaining for the virus in tissue by IHC by alkaline phosphatase (black arrows) in (F). (Hematoxylin and Eosin, A: 200x; B – F: 400x).

Figure 4.
Semiquantification of histopathological changes in hepatic acini (Z1, Z2, and Z3) of patients infected with YFV, DENV, and CHIKV. Macrovesicular Steatosis (A–C) and Microvesicular Steatosis (D–F).
Figure 4.
Semiquantification of histopathological changes in hepatic acini (Z1, Z2, and Z3) of patients infected with YFV, DENV, and CHIKV. Macrovesicular Steatosis (A–C) and Microvesicular Steatosis (D–F).
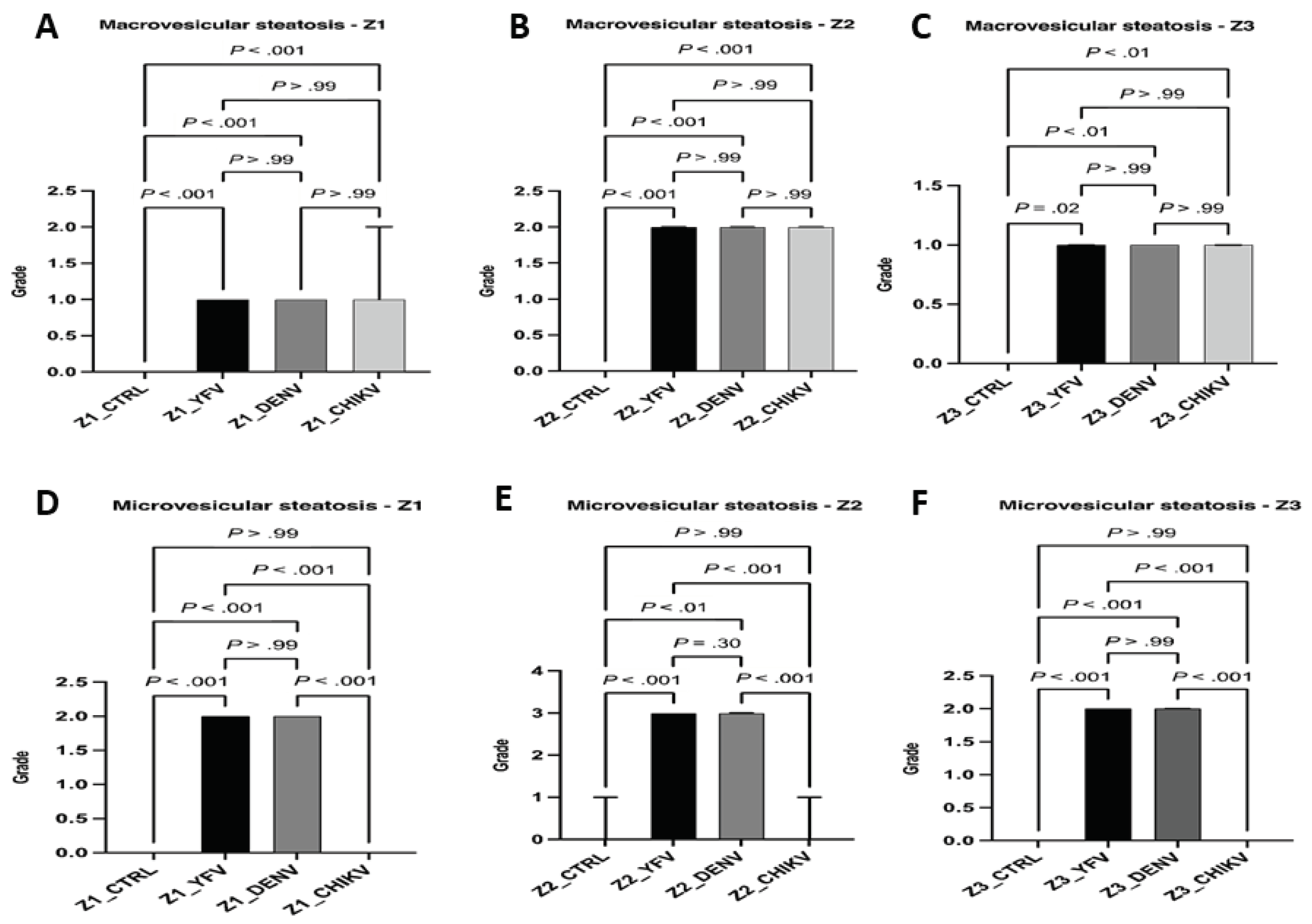
Figure 5.
Semiquantification of histopathological changes in hepatic acini (Z1, Z2, and Z3) of patients infected with YFV, DENV, and CHIKV. Cell Swelling (A–C) and Lytic Necrosis (D–E).
Figure 5.
Semiquantification of histopathological changes in hepatic acini (Z1, Z2, and Z3) of patients infected with YFV, DENV, and CHIKV. Cell Swelling (A–C) and Lytic Necrosis (D–E).
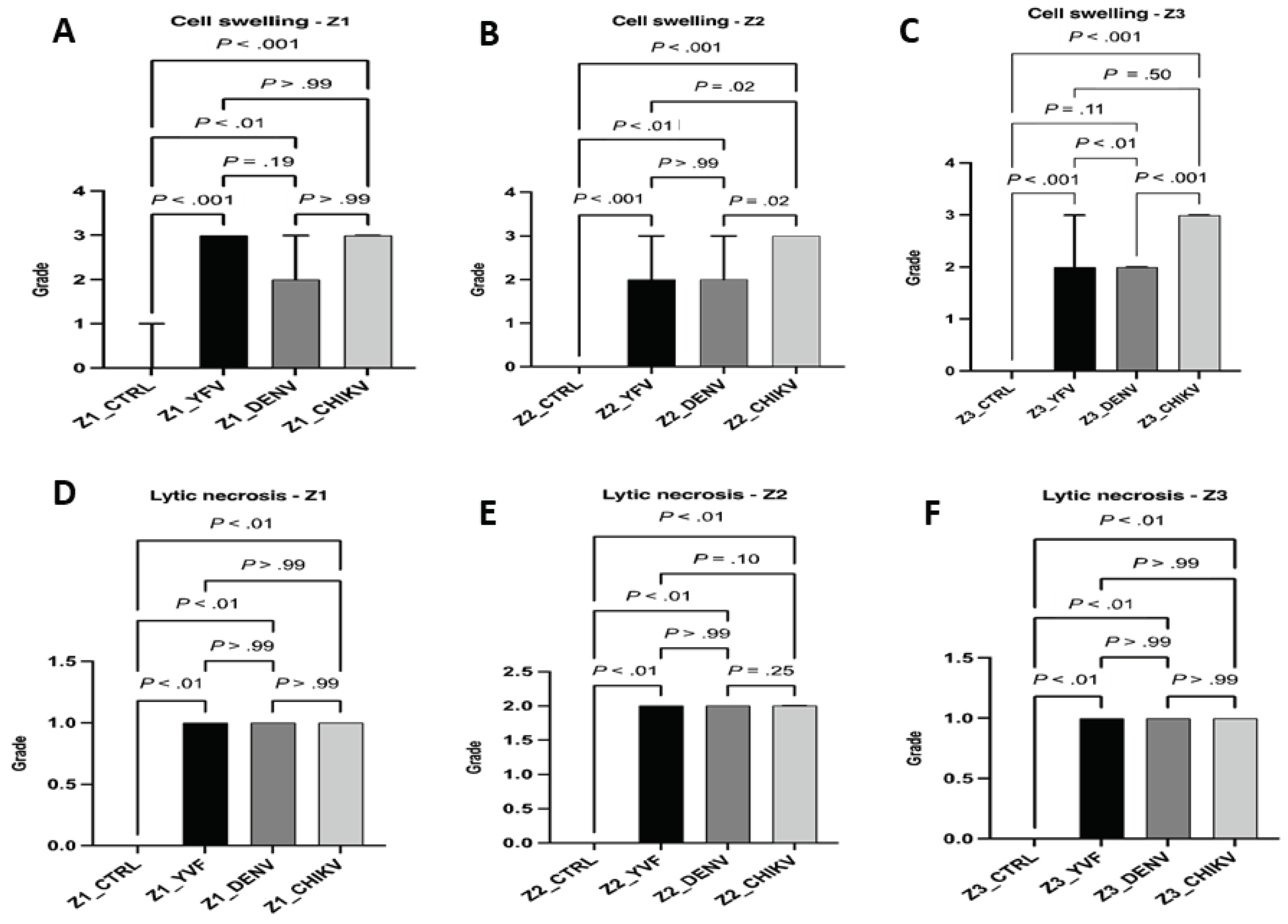
Figure 6.
Semiquantification of histopathological changes in hepatic acini (Z1, Z2, and Z3) of patients infect.
Figure 6.
Semiquantification of histopathological changes in hepatic acini (Z1, Z2, and Z3) of patients infect.
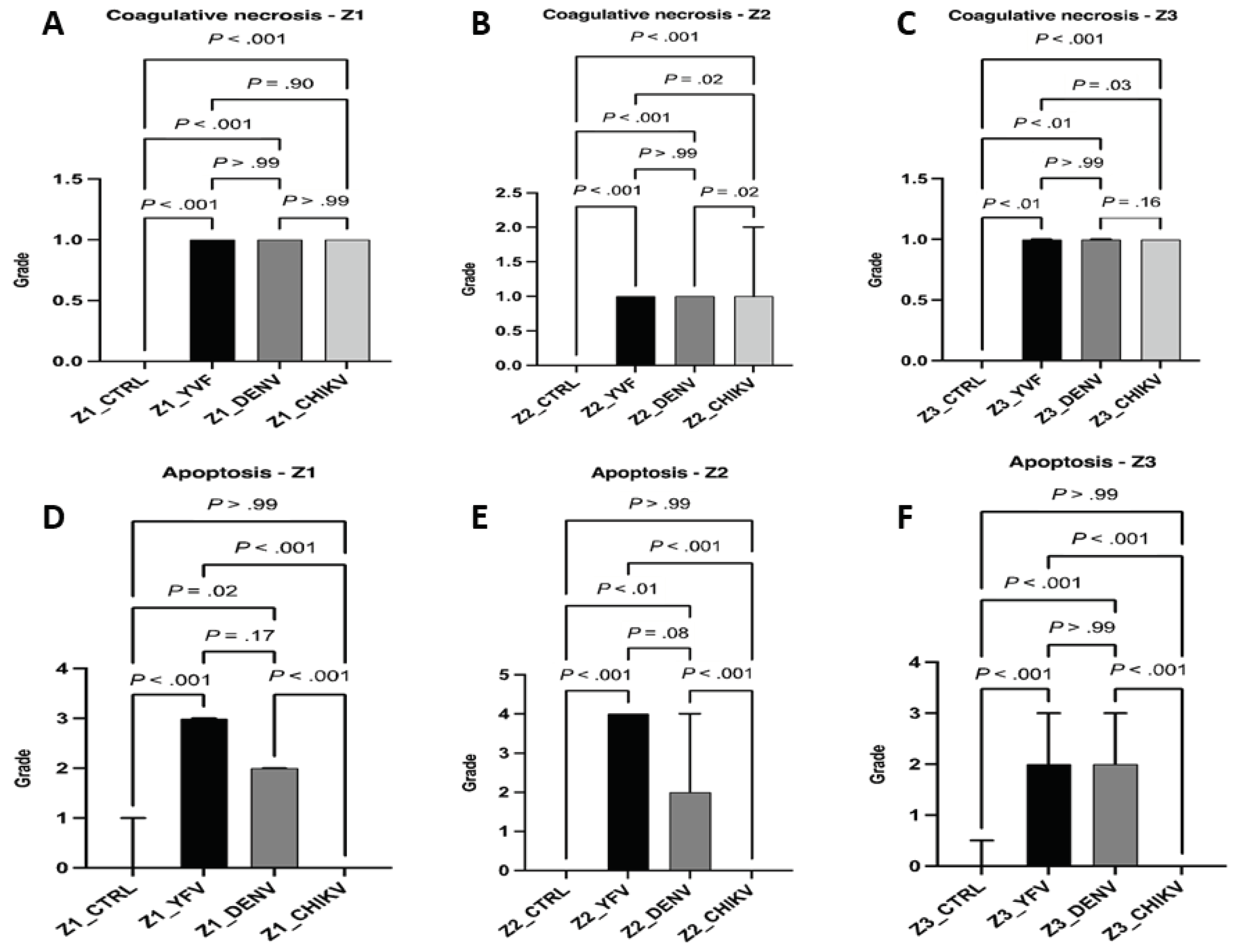
Figure 7.
Semiquantification of histopathological changes in hepatic acini (Z1, Z2, and Z3) of patients infected with YFV, DENV, and CHIKV. Kupffer Cell Hyperplasia (A–C) and Kupffer Cell Hypertrophy (D–F).
Figure 7.
Semiquantification of histopathological changes in hepatic acini (Z1, Z2, and Z3) of patients infected with YFV, DENV, and CHIKV. Kupffer Cell Hyperplasia (A–C) and Kupffer Cell Hypertrophy (D–F).
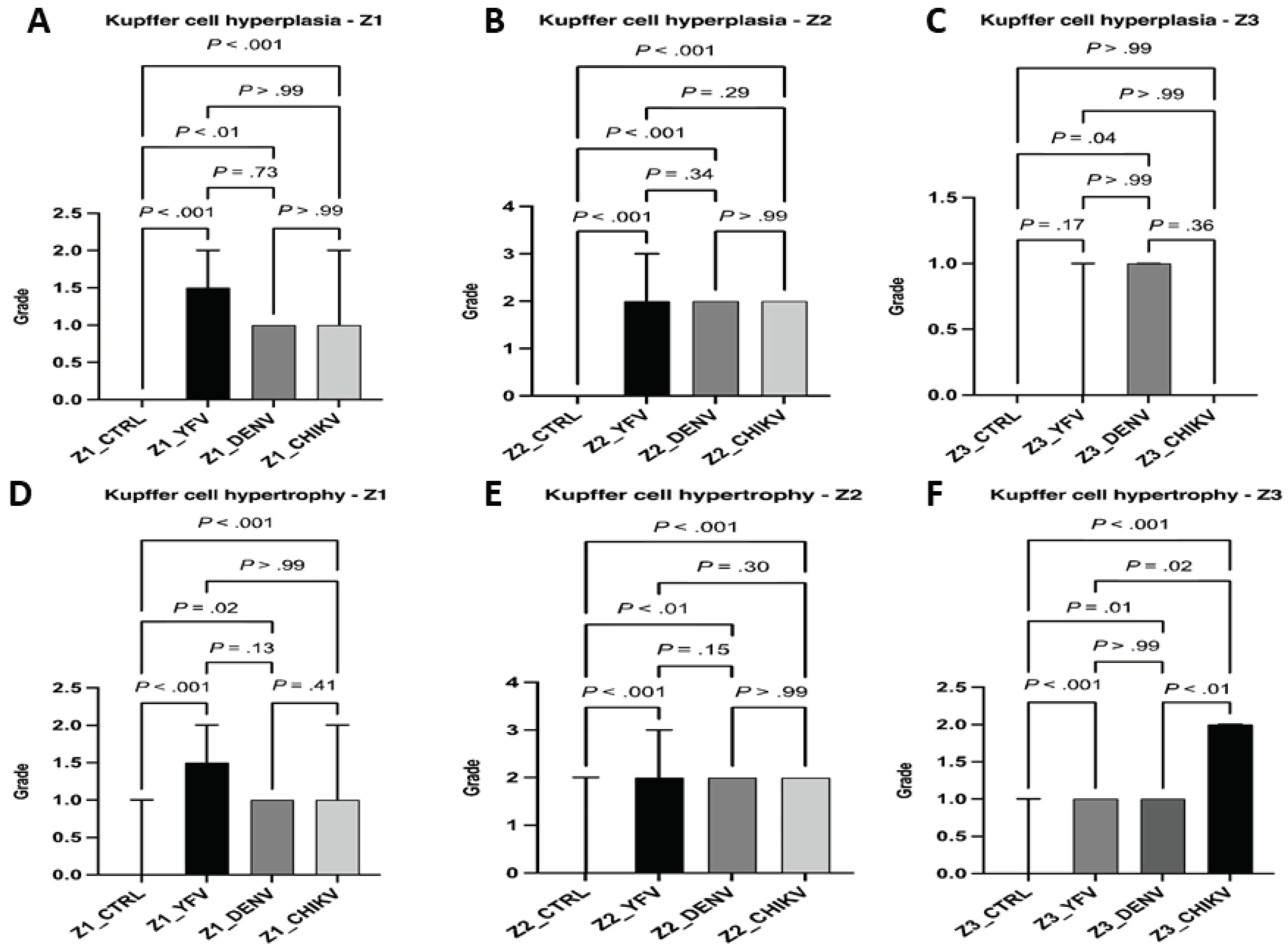
Figure 8.
Semiquantification of histopathological changes in the portal tract of patients infected with YFV, DENV, and CHIKV. Edema (A), Congestion (B), Portal Vein Alteration (C), Portal Artery Alteration (D), Bili Canaliculus Alteration (E).
Figure 8.
Semiquantification of histopathological changes in the portal tract of patients infected with YFV, DENV, and CHIKV. Edema (A), Congestion (B), Portal Vein Alteration (C), Portal Artery Alteration (D), Bili Canaliculus Alteration (E).
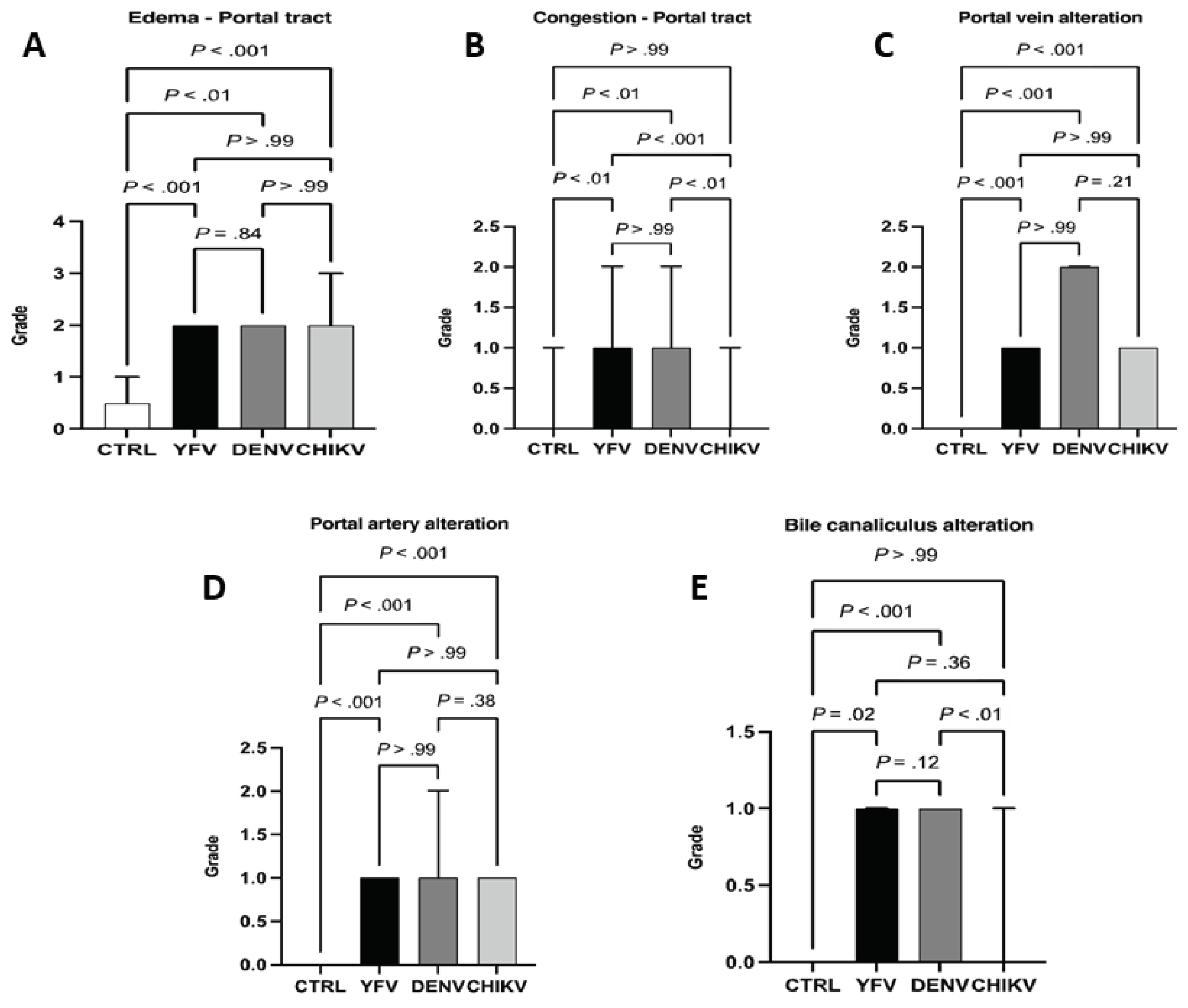
Figure 9.
Steatosis in liver tissue of YFV, DENV, and CHIKV-infected patients. Hepatic steatosis is defined by an increase in lipids in hepatocytes, microscopically, lipid droplets are observed in the liver parenchyma. The figure shows the mechanisms of macrovesicular and microvesicular steatosis.
Figure 9.
Steatosis in liver tissue of YFV, DENV, and CHIKV-infected patients. Hepatic steatosis is defined by an increase in lipids in hepatocytes, microscopically, lipid droplets are observed in the liver parenchyma. The figure shows the mechanisms of macrovesicular and microvesicular steatosis.
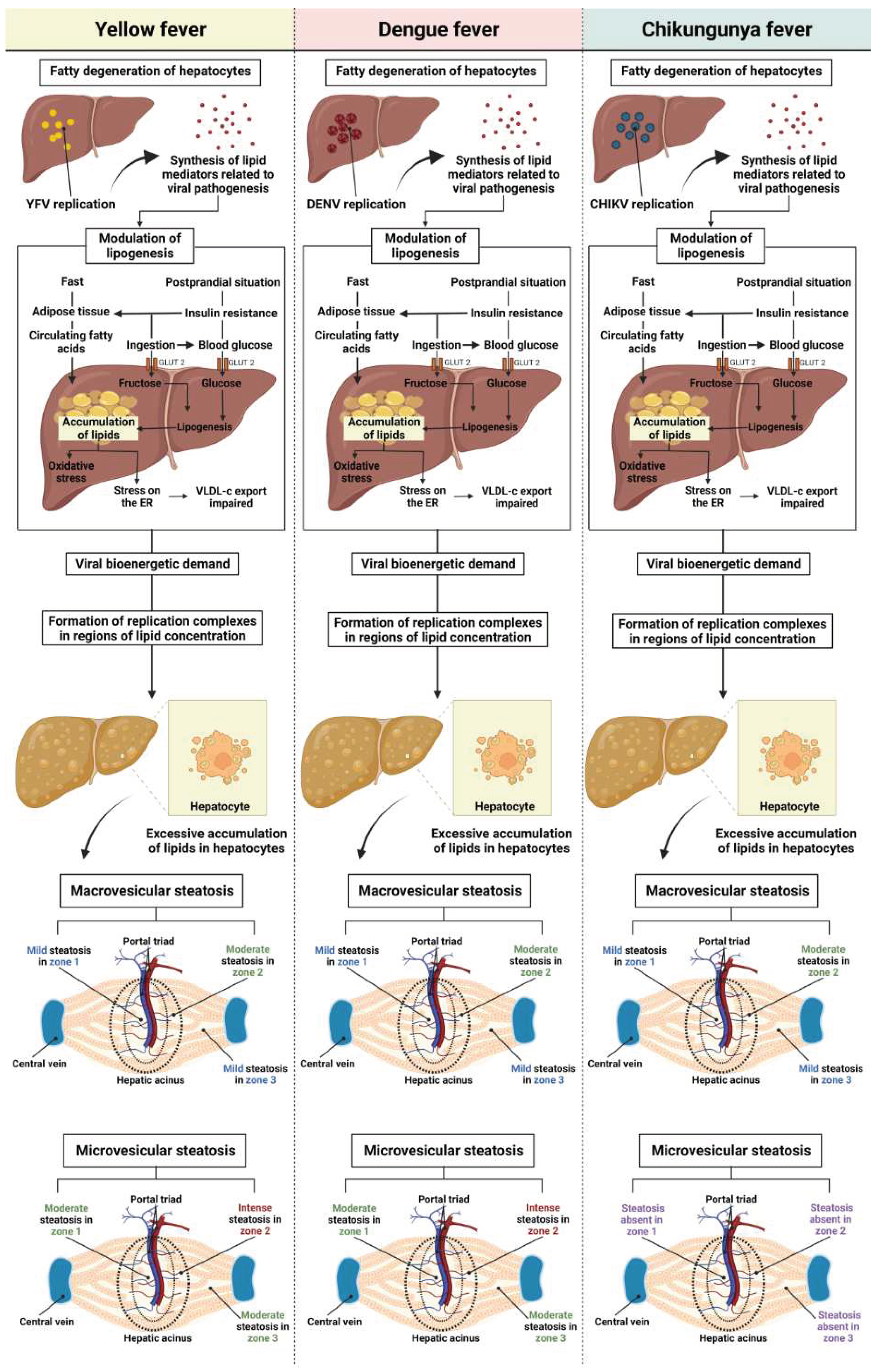
Figure 10.
Necrosis in liver tissue of YFV, DENV, and CHIKV-infected patients. During the viral infection, infected hepatocytes undergo necrotic processes, due to direct viral action on the infected cell and/or immunomodulation, activating cytotoxic cells, such as NK and TC8+.
Figure 10.
Necrosis in liver tissue of YFV, DENV, and CHIKV-infected patients. During the viral infection, infected hepatocytes undergo necrotic processes, due to direct viral action on the infected cell and/or immunomodulation, activating cytotoxic cells, such as NK and TC8+.
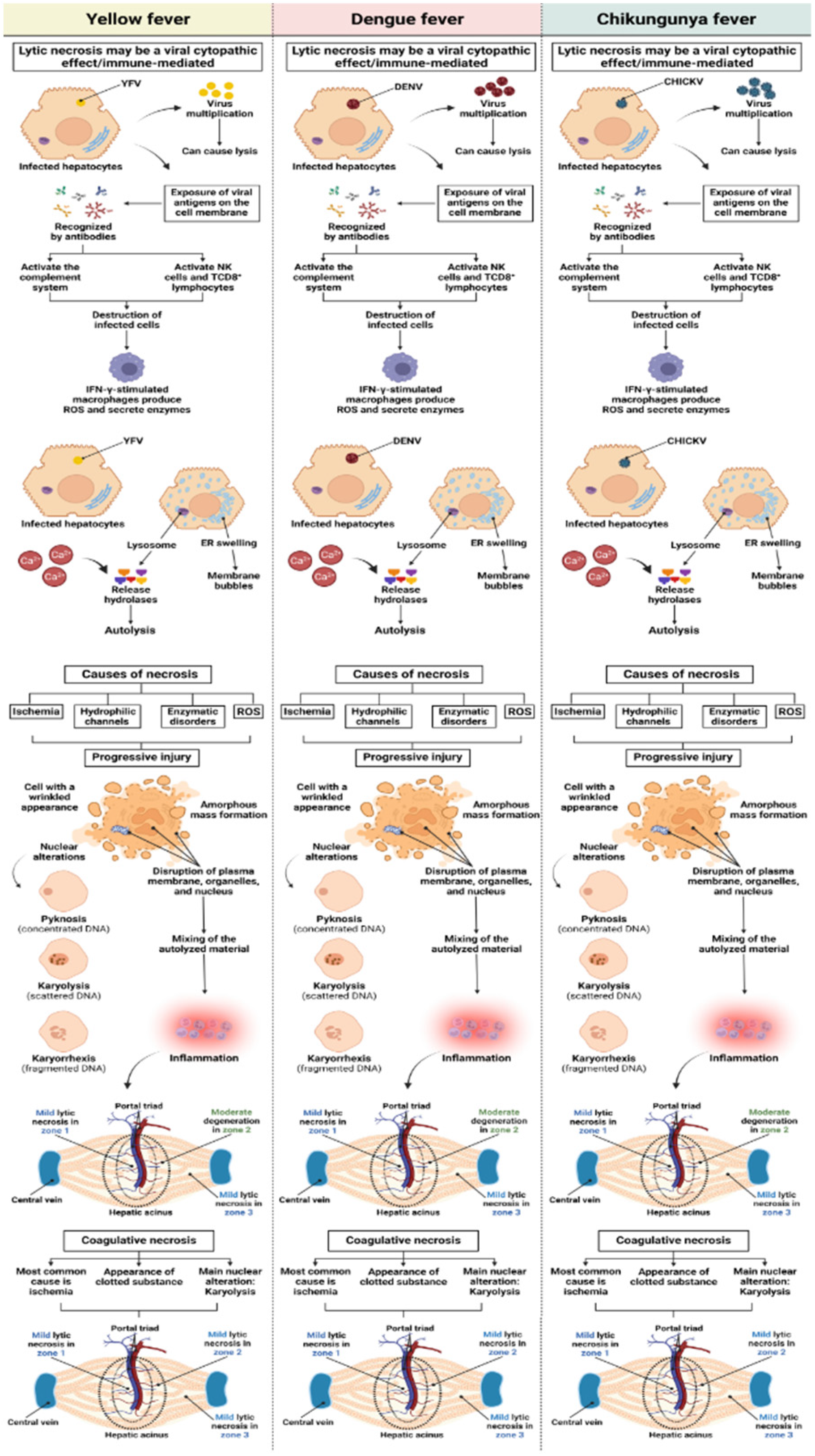
Figure 11.
Apoptosis in liver tissue of YFV, DENV, and CHIKV-infected patients. In viral infection the development of apoptotic process is observed. The mechanisms of apoptosis induction involve intrinsic signals coming from mitochondrial alteration or by the activation of cell death receptors on the cell surface.
Figure 11.
Apoptosis in liver tissue of YFV, DENV, and CHIKV-infected patients. In viral infection the development of apoptotic process is observed. The mechanisms of apoptosis induction involve intrinsic signals coming from mitochondrial alteration or by the activation of cell death receptors on the cell surface.
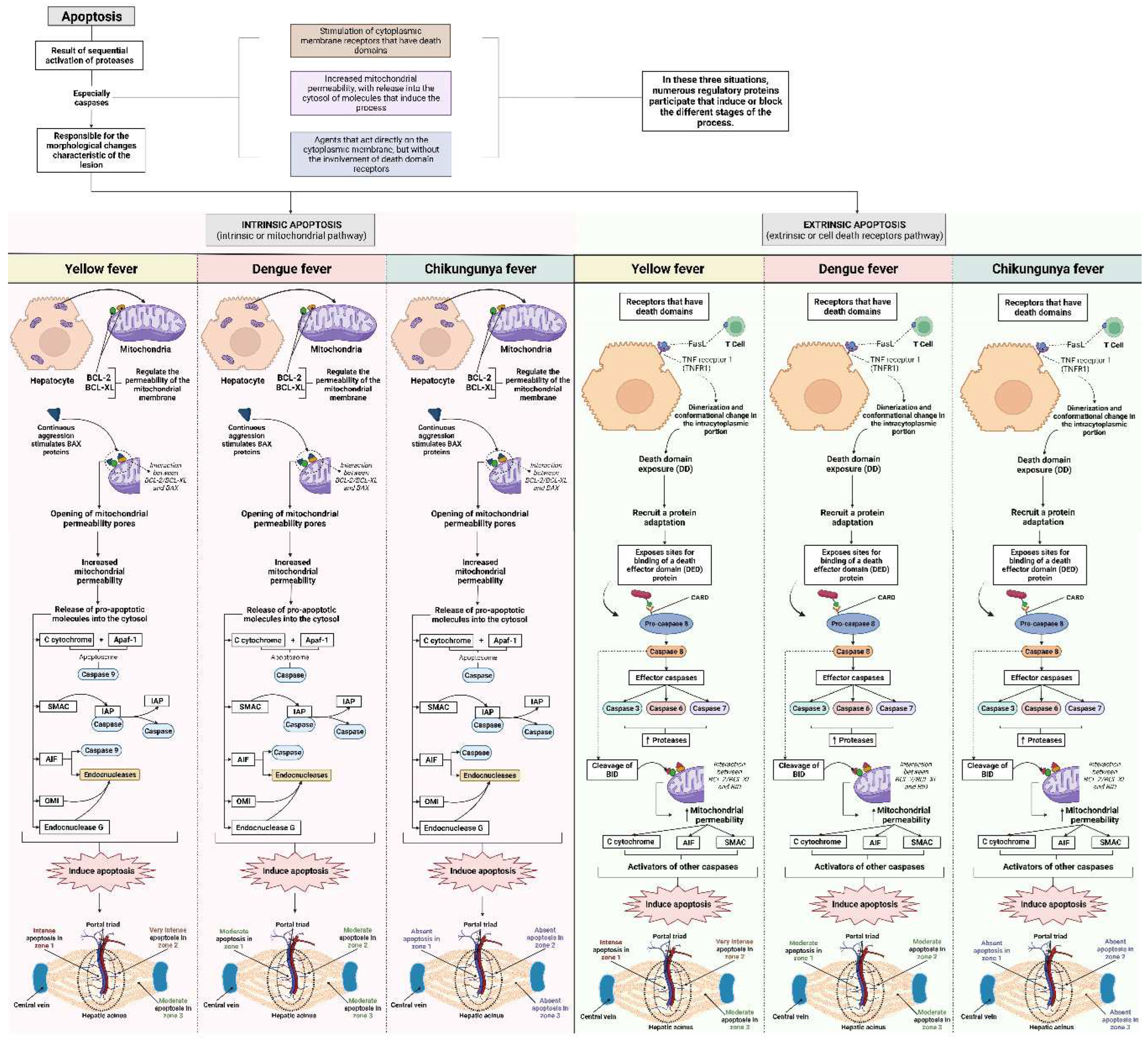
Figure 12.
Vascular alteration in liver tissue of YFV, DENV, and CHIKV-infected patients. Changes in vascular microcirculation include, arteriolar vasodilation, smaller venules dilate, while larger venules undergo minor constriction increasing pressure in microcirculation, increasing vascular permeability, plasma outflow into the interstitium.
Figure 12.
Vascular alteration in liver tissue of YFV, DENV, and CHIKV-infected patients. Changes in vascular microcirculation include, arteriolar vasodilation, smaller venules dilate, while larger venules undergo minor constriction increasing pressure in microcirculation, increasing vascular permeability, plasma outflow into the interstitium.
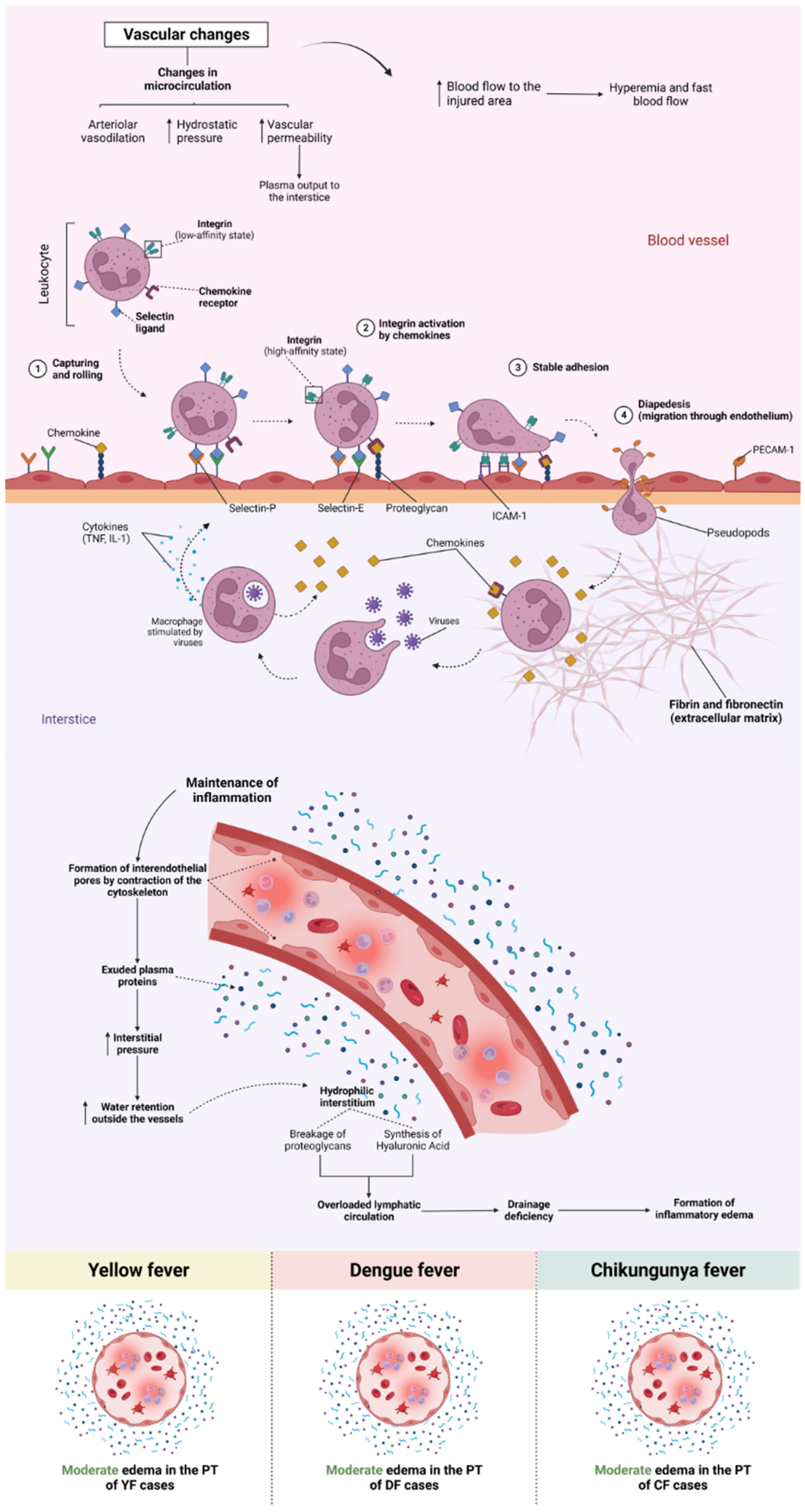
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



