
Submitted:
19 January 2023
Posted:
20 January 2023
You are already at the latest version
Abstract
Cryptococcus neoformans is a human multidrug-resistant yeast with high mortality rates in immunocompromised patients. Recently, the synthetic peptide Mo-CBP3-PepII emerged as a potent anticryptococcal molecule with an MIC50 at low concentration. Here, the mechanisms of action of Mo-CBP3-PepII were deeply analyzed to provide new information about how it led C. neoformans cells to death. Light and fluorescence microscopies, analysis of enzymatic activities, and proteomic analysis were employed to understand the effect of Mo-CBP3-PepII on C. neoformans cells. Light and fluorescence microscopies revealed Mo-CBP3-PepII induced the accumulation of anion superoxide and hydrogen peroxide in C. neoformans cells. In addition to a reduction in the activity of superoxide dismutase (SOD), ascorbate peroxidase (APX), and catalase (CAT) in the cells treated with Mo-CBP3-PepII. In the presence of Ascorbic acid (AsA), no ROS were detected and Mo-CBP3-PepII lost the inhibitory activity against C. neoformans. Yet, Mo-CBP3-PepII inhibited the activity of lactate dehydrogenase (LDH), ergosterol biosynthesis, and induced the decoupling of cytochrome c from the mitochondrial membrane. Proteomic analysis revealed a reduction in the abundance of proteins related to energetic metabolism, DNA and RNA metabolism, pathogenicity, protein metabolism, cytoskeleton, and cell wall organization and division. Our findings indicated that Mo-CBP3-PepII might have multiple mechanisms of action against C. neoformans cells, mitigating the development of resistance and thus being a potent molecule to be employed in the production of new drugs against C. neoformans infections.
Keywords:
alternative drugs
; cryptococcosis
; oxidative stress
; synthetic peptides
1. Introduction
Two decades have passed since the last new antifungal drug was released; however, the incidence of invasive infections by multidrug-resistant Fungi has increased [1]. Three main antifungal classes are available to treat fungal infections: azoles, polyenes, and echinocandins. Nonetheless, their use includes limitations, such as the spectrum of activity, resistance, toxicity, drug-drug interactions, and poor bioavailability [2].
In a list of priority regarding fungal pathogens recently published by the World Health Organization (WHO) [3], Cryptococcus neoformans appears in the top three due to some reasons: (1) low susceptibility to current antifungal agents; (2) poorly understood resistance to drugs; and (3) high mortality rate ranging from 41% to 61% [3]. C. neoformans causes cryptococcosis, which mainly occurs in the lungs and leads to pneumonia but also affects the brain, causing meningitis, mostly in patients with a compromised immune system [3,4]. HIV+ patients, individuals with cancer, or taking any drug that weaken the immune system are predisposed to acquire cerebral cryptococcosis, which forms lesions known as cryptococcomas [3]. C. neoformans is naturally resistant to azole antifungals and the latest antifungal drug, Caspofungin, causing global health concerns [5,6]. Therefore, the search for new alternatives to treat cryptococcosis becomes emerging. In this context, antimicrobial peptides (AMPs) appear as a promise due to their mechanisms of action on the membrane [7].
Natural AMPs have limitations in their use and are generally cytotoxic. Moreover, the rational design of synthetic antimicrobial peptides (SAMPs) is an alternative to this problem. This approach is cost-effective for active short peptides [7,8,9]. Recently, our research group assessed the anticryptococcal potential of Mo-CBP3-PepII [10], a synthetic peptide bioinspired in the sequence of Mo-CBP3, a protein purified from Moringa oleifera seeds [11]. This study demonstrated that Mo-CBP3-PepII has an MIC50 of 25 µg mL-1 and induced membrane pore formation, DNA degradation, and apoptosis in cryptococcal cells. Additionally, it was reported that Mo-CBP3-PepII interacts with ergosterol in the membrane of C. neoformans favoring pore formation [10].
Here, new insights were provided about the mechanisms of action employed by Mo-CBP3-PepII against C. neoformans. Our analysis revealed that Mo-CBP3-PepII stimulates oxidative stress, inhibits ergosterol biosynthesis, and induces the decoupling of cytochrome c from mitochondria. In addition, Mo-CBP3-PepII promotes changes in the proteomic profile of C. neoformans.
2. Results and Discussion
2.1. Mo-CBP3-PepII Induced ROS Overaccumulation in C. neoformans Cells
All experiments in this study were carried out using the MIC50 concentration of 25 µg mL-1 of Mo-CBP3-PepII against C. neoformans as defined in our previous published study by Aguiar et al. [10]. The Mo-CBP3-PepII-treated C. neoformans cells showed an accumulation of reactive oxygen species (ROS) anion superoxide (O2•–) and hydrogen peroxide (H2O2) (Figure 1). The light microscopy analysis revealed the accumulation of O2•– in Mo-CBP3-PepII-treated C. neoformans cells (Figure 1 - blue or cyan dots – black arrows). As expected, the same was not observed in control cells (DMSO-NaCl) (Figure 1). The detection of H2O2 was carried out by fluorescence microscopy using 2′,7′ dichlorofluorescein diacetate (DCFH-DA, Sigma, St. Louis, MI, 364 USA) assay. The green fluorescence on cells indicates the accumulation of H2O2 after treatment with Mo-CBP3-PepII. In contrast, DMSO-NaCl not showed the same (Figure 1 - H2O2 panel).
ROS are inherent molecules of life as a byproduct of aerobic cell metabolism, and it is impossible to leave without them. Besides, ROS plays several essential roles in cell life, growth, development, defense, and signaling [12]. For pathogens such as Fungi, ROS play critical roles in biofilm biogenesis, infection, virulence, and developmental process [13]. Even though these are good effects of ROS, the induction of ROS overproduction is a mechanism of action employed by antimicrobial peptides to kill human pathogenic Fungi [7,14,15,16,17]. What is essential to discuss is that ROS are good at low and controlled levels; if an external insult disturbs the ROS homeostasis, they could be lethal for cells. That is what antimicrobial peptides do.
Here, Mo-CBP3-PepII induced an uncontrolled accumulation of •O2− and H2O2 (Figure 1). At higher levels, •O2− and H2O2 could cause severe damage to cells by interacting and degrading critical cellular components such as DNA, proteins, lipids, inducing membrane pore formation, apoptosis and death [16,17,18,19,20]. For example, higher levels of •O2− as demonstrated in Figure 1, led to DNA oxidation mediated by iron [20]. High levels of •O2− increase free iron levels by releasing it from heme-oxidized proteins and enzyme clusters. The free iron interacts with DNA molecules oxidizing them and inducing fragmentation [20]. This result agrees with Aguiar et al. [10], that reported Mo-CBP3-PepII induced DNA fragmentation in C. neoformans cells.
In the case of H2O2, higher levels of H2O2 drive the lipid peroxidation in the membrane and, thus, pore formation [18]. Recently, it was reported that two synthetic peptides, RcAlb-PepII and PepGAT, lost the ability to induce pore formation in the C. neoformans membrane in the presence of an antioxidant agent where no H2O2 accumulation. This result suggests both peptides induce pore formation by ROS-dependent pathway [21]. Besides damaging the membrane, higher levels of H2O2 could also result in a higher level of oxidized protein and loss of activity. Proteomic analysis of Klebsiella pneumoniae cells in the presence of a synthetic peptide Mo-CBP3-PepI, which induced higher levels of H2O2, revealed an increase in the accumulation of proteins involved in the repair of oxidized proteins [22]. This data suggests that K. pneumoniae cells are trying to fix the damaged proteins by H2O2.
2.2. Mo-CBP3-PepII Affects the Activity of Redox System Enzymes
Aguiar et al. [10] revealed Mo-CBP3-PepII forms a 6-kDa pore in the membrane of C. neoformans. So, it was reasoned that Mo-CBP3-pepII could move by this pore and interfere with the activity of some cytoplasmic enzymes involved in ROS metabolism, leading to ROS accumulation, as shown in Figure 1. As discussed above, ROS are essential to life but only if controlled to safer levels [12]. The safer levels of ROS were maintained by two mechanisms: enzymatic and non-enzymatic [23]. The enzymatic is mediated by scavenger enzymes that either convert highly unstable ROS molecules to more stable or completely oxidize them to H2O and O2 [23]. Here, the activity of three enzymes, SOD, CAT, and APX, was analyzed (Figure 2).
The first enzyme analyzed was SOD because it is involved in the conversion of •O2− to H2O2 [24]. In control C. neoformans cells, the SOD activity was 5.67 AU mgP-1. In contrast, no activity was detected in Mo-CBP3-PepII-treated cells (Figure 2A). In addition, two other enzymes were analyzed, CAT and APX, both involved in the conversion of H2O2 into H2O in O2 [25,26]. Regarding the CAT activity, control cells of C. neoformans presented an activity of 0.52 AU mgP-1 and Mo-CBP3-PepII-treated cells showed a value of 0.121 AU mgP-1. A reduction of 4.0-fold in CAT activity (Figure 2B). The APX activity from C. neoformans cells was also affected by Mo-CBP3-PepII (Figure 3C). Mo-CBP3-PepII (0.77 AU mgP-1) induced a reduction of 4.5-fold in APX activity compared to control cells (3.40 AU mgP-1) (Figure 2C).
As demonstrated in Figure 2, Mo-CBP3-PepII induced alteration in the activity of enzymes involved in the ROS scavenger process. By an unclear mechanism, Mo-CBP3-PepII insults the balance of ROS in C. neoformans cells. First, the inhibition of SOD activity (Figure 2A) agrees with the accumulation of •O2− (Figure 1). Second, the reduction in the activity of CAT and APX let high the levels of H2O2 (Figure 2B and C). The number of studies showing such alteration in redox enzymes is scarce. Two synthetic peptides, RcAlb-PepII and RcAlb-PepII, could also completely inhibit the SOD activity and reduce CAT and APX activities in C. neoformans cells, leading to ROS accumulation [21]. In another study, not with peptides but with an antifungal protein, Mo-CBP2, purified from Moringa oleifera seeds, presented perturbation in the activity of those enzymes in Candida albicans [27].
This scenario strongly corroborates the previous results of Mo-CBP3-PepII [10]. Recently, Aguiar et al. [10] Mo-CBP3-PepII induced DNA fragmentation and apoptosis in C. neoformans cells. Altogether, previous results and those presented here showed a high accumulation of ROS in C. neoformans cells that could drive DNA fragmentation and apoptosis.
2.3. Ascorbic Acid (AsA) Affects the Anticryptococcal Potential of Mo-CBP3-PepI
Figure 1 and Figure 2 revealed Mo-CBP3-PepII induced the overaccumulation of ROS by negatively modulating the activity of scavenger enzymes. These results correspond to previous damage caused by Mo-CBP3-PepII in C. neoformans cells [10]. Based on that, one question emerged: is the activity of Mo-CBP3-PepII ROS-dependent? To answer this question, Mo-CBP3-PepII was assayed against C. neoformans in the presence of 10 mM of AsA. At the concentration of 25 µg mL-1, Mo-CBP3-PepII reaches the MIC50 without AsA (Figure 3 – graphic). In the presence of 10 mM of AsA, the anticryptococcal activity of Mo-CBP3-PepII dropped to 10%, a reduction of 40% in activity (Figure 3 – graphic).
Aguiar et al. [10] reported that Mo-CBP3-PepII could induce pore formation in membranes of C. neoformans cells, as revealed by the propidium iodide (PI) uptake assay. In the presence of AsA, fluorescence microscopy revealed no H2O2 accumulation and pore formation (Figure 3), indicating that Mo-CBP3-PepII lost the ability to induce pore formation. This result suggests the ability of Mo-CBP3-PepII to induce pore formation is H2O2-dependent.
As happened to ROS, pore formation is also a common mechanism of action played by peptides because it could result from ROS accumulation and lipid peroxidation or the direct action of peptides in the membrane [18,28,29]. These data suggest the activity and mechanism of action of Mo-CBP3-PepII are dependent on ROS accumulation. This result is different from others presented by peptides RcAlb-PepIII and PepKAA [21], where the ability of pore formation is independent of ROS.
It seems that Mo-CBP3-PepII first interacts with C. neoformans cells interfering with the ROS scavenger enzyme activity leading to the accumulation of ROS and, thus, several other damages to C. neoformans, driving them to death.
2.4. Mo-CBP3-PepII Interferes in the Biosynthesis of Ergosterol
Ergosterol is a fungal membrane component, and its primary function is acting as a stabilizer [30]. The biosynthesis of ergosterol occurs in the cytoplasm of the cell and is transported to the membrane [30]. Aguiar et al. [10] demonstrated that Mo-CBP3-PepII was able to bind to ergosterol. Therefore, we experimented with evaluating the ability of Mo-CBP3-PepII to inhibit the ergosterol synthesis (Figure 4A). As expected, in control with DMSO-NaCl, there was no inhibition of ergosterol synthesis. The positive control for inhibition, itraconazole (ITR), was able to inhibit 47.65% of ergosterol biosynthesis (Figure 4A). Meanwhile, Mo-CBP3-PepII inhibited the ergosterol biosynthesis in 72% (Figure 4A), a value 1.5 times higher than ITR.
By inhibiting the biosynthesis of ergosterol, Mo-CBP3-PepII destabilizes the membrane facilitating the pore formation on the membrane leading to loss of cytoplasmic content and cell death. Mo-CBP3-PepII targets ergosterol in two ways: interacting with it in the membrane [10] or inhibiting its synthesis in C. neoformans cells (Figure 4A).
2.5. Energetic Metabolism is Affected in C. neoformans Treated with Mo-CBP3-PepII
Still looking for possible cytoplasmic targets of Mo-CBP3-PepII, the lactate dehydrogenase (LDH) activity was assayed in C. neoformans cells after contact. The activity of LDH was wholly inhibited in cells treated with Mo-CBP3-PepII (MIC50) (Figure 4B). As expected, control cells did not suffer any alteration in LDH activity (Figure 4B).
LHD is an essential enzyme in carbohydrate metabolism because it is involved in the regeneration of NAD+ from NADH produced in the glycolysis pathway. This reaction maintains a high ratio of NAD+/NADH in the cytoplasm, favoring glycolysis [31]. By inhibiting the LDH activity, Mo-CBP3-PepII interferes in the regeneration of NAD+, which is essential for glycolysis still works. The inhibition of LDH can jeopardize all energetic metabolism in C. neoformans cells.
Another parameter of energetic metabolism evaluated was the decoupling of Cyt c from the mitochondrial membrane (Figure 4C). In cells treated with Mo-CBP3-PepII (MIC50), there was a significant increase of 51% of Cyt c content in the cytoplasm. The positive control H2O2 promotes 100% of Cyt c release from mitochondrial to cytoplasm (Figure 4C). DMSO did not promote any release of Cyt c. At that time, Mo-CBP3-PepII is interfering with energy production in mitochondria. The release of Cyt c from the mitochondrial membrane impairs the complete function of the electron transport chain (ETC) in the mitochondrion leading to a depletion in ATP levels [32]. These analyses suggest Mo-CBP3-PepII has targets within C. neoformans cells and might reach the cytoplasm and other organelles.
One fact is, Mo-CBP3-PepII affects the energy production of C. neoformans cells in both stages in the cytoplasm and mitochondria. Energy is critical for any organism to cope with stresses of any kind, for example, to produce defense proteins [33]. Here, we showed that Mo-CBP3-PepII imposes different types of stress on C. neoformans cells and shutting down the energy production mitigates the chances of C. neoformans fighting back.
The role in ETC is the canonical function of Cyt c. However, its role goes beyond that [32]. Cyt c is also crucial for the health of mitochondria. The decoupling of Cyt c from mitochondrial membranes signals mitochondrion misfunction and thus starts cell apoptosis [32]. Therefore, by inducing the release of o Cyt c from the mitochondrial membrane, Mo-CBP3-PepII induces C. neoformans cells to enter apoptosis. Aguiar et al. [10] revealed that Mo-CBP3-PepII induced apoptosis in C. neoformans cells corroborating the results reported here.
2.6. Mo-CBP3-PepII Promotes Several Changes in the Cell Structure of C. neoformans
Atomic force microscopy (AFM) analysis revealed the effects that Mo-CBP3-PepII induces in the structure of C. neoformans cells. The control showed a cryptococcal cell in its morphology with no damage (Figure 5A,B), healthy and continuous cytoplasmatic membrane, homogenous cytoplasm, and no damage to the capsule. By contrast, C. neoformans treated with Mo-CBP3-PepII showed several damages in its structure, such as damage to the membrane, cell wall thickness, damage to fungal capsules, wrinkled cytoplasm, and roles across the whole cells (Figure 5C,D – brown dots). Also, it is possible to see that Mo-CBP3-PepII-treated cells have a diameter and height and hence higher volume than control cells indicating treated cells are bigger than control cells (Figure 5 C,D). This result suggests that Mo-CBP3-PepII induced pore formation in the C. neoformans membrane by changing its permeability. In that case, cells cannot control the osmotic potential. Additionally, 3D images (Figure 5D) showed that C. neoformans cells treated with Mo-CBP3-PepII are flattened, presenting structures like depression in the middle of the cell.
Recently, the morphology of C. neoformans treated with synthetic peptides was revealed by scanning electron microscopy [10]. In this study, authors revealed several damages to the cell structure; in some cases, those are similar to data presented in this study, but AFM analysis brought more details on the effect of Mo-CBP3-PepII on C. neoformans cells. Similar AFM data on C. neoformans cells were reported by Ishida et al. [34] using silver nanoparticles. In the study, the authors reported that C. neoformans cells presented damage in the cell wall, cell capsule, and membrane.
2.7. Proteomic Profile of C. neoformans Treated with Mo-CBP3-PepII
2.7.1. Overview
Proteomic analysis is an important technique to see the whole picture of a cell by evaluating the protein profile after treatment with peptides [35,36,37,38,39]. Proteomic analysis has already been employed to study the response of resistant pathogens such as Clostridioides difficile [37]. Regarding C. neoformans, proteomic analysis was employed to understand the changes in protein profile during the transition from planktonic to biofilm lifestyle [40]. As far as we know, no study has employed proteomics to analyze the C. neoformans response to antimicrobial peptides, reinforcing the pioneering of our work.
As shown above, Mo-CBP3-PepII presented different mechanisms against C. neoformans cells. However, all of those are despite only the cell structure itself. After that, one new question arises: is Mo-CBP3-PepII able to change the protein profile of C. neoformans? A proteomic analysis was performed to obtain the answer (Figure 6, Table 1, and Supplementary Tables 1 and 2).
Five hundred seventy-three proteins were identified (Figure 6A, Table 1, and supplementary Tables 1 and 2). Of these, 265 were exclusively identified in control cells (Supplementary Table 1), 266 were exclusively detected in Mo-CBP3-PepII-treated cells (Supplementary Table 2), and 42 were detected in both groups (Figure 6A and Table 1).
The proteins identified in both groups were called overlapping proteins. For these proteins, fold-change was calculated based on the intensity of peptides for proteins in Mo-CBP3-PepII-treated/control cells [41]. Proteins with a fold-change value ≥ 1.5 (p < 0.05, Tukey’s test) [41] were called up-accumulated (increase in abundance). Proteins with a fold-change value of ≤ 0.5 (p < 0.05, Tukey’s test) [41] were called down-accumulated (decrease in abundance). Proteins with a fold-change value ranging from 0.5 to 1.5 (p < 0.05, Tukey’s test) were considered they did not change in abundance [41] (Table 1). Among the overlapping proteins, 10 were up-accumulated, and 05 were down-accumulated in Mo-CBP3-PepII-treated cells compared to control cells (Figure 6A and Table 1). Twenty-seven overlapping proteins did not change their accumulation in response to Mo-CBP3-PepII treatment in C. neoformans cells (Figure 6A and Table 1).
The overlapping proteins were classified based on gene ontology for molecular function and biological process (Figure 6B,C). Regarding the molecular function, proteins were classified into 9 groups DNA and RNA Binding (14.28%), Hydrolase (7.14%), Oxidoreductase (9.52%), Ligase (2.38%), Multifunctional enzyme (2.38%), Protein Binding (9.52%), Transport or Structural Activity (11.90%), Transferase (26.19%), and Unknown (16.66%) (Figure 6B, Table 1).
The biological process also revealed 9 groups of proteins Amino acid metabolism (9.52%), Cell Function and Structure (21.42%), Energetic metabolism (9.52%), Lipid metabolism (4.76%), Nucleic acid metabolism (23.80%), Protein folding (9.52%), Regulation Factor or Signaling (4.76%), Transport (4.76%), and Unknown (11.90%) (Figure 6C, Table 1).
2.7.2. DNA and RNA Binding Proteins
In this group, two isoforms, DNA topoisomerase I and DNA topoisomerase 2, were exclusively detected in the C. neoformans cells treated with Mo-CBP3-PepII (Supplementary table 2). DNA topoisomerases are proteins that solve topological problems in DNA molecules, such as supercoiling and catenation [42,43]. Topoisomerases are a well-known target for antifungal drugs such as echinocandins [42]. Despite their function in regulating DNA torsion, topoisomerases are also involved in the repair of damaged DNA. The exclusive detection of two isoforms of topoisomerases in treated cells indicates Mo-CBP3-PepII [44]. Additionally, Pommier et al. [44] reported topoisomerase I is induced during cellular stress to prevent DNA hence cell death mediated by apoptosis [44]. The increase of topoisomerase 1 suggests the Mo-CBP3-PepII is inducing damage to DNA. It is trustworthy to notice that Aguiar et al. [10] revealed that Mo-CBP3-PepII induced DNA fragmentation and apoptosis; maybe these processes are related to apoptosis in C. neoformans cells induced by Mo-CBP3-PepII.
Another protein only identified in Mo-CBP3-PepII-treated C. neoformans cells was the Mitochondrial escape protein 2 (MET2) (Supplementary table 2). MET2 protein controls the escape of mitochondrial DNA during stress to prevent degradation [45,46]. Once mitochondria suffer any external insult that could lead to DNA damage, the MET2 proteins are involved in the protection, repair, or even DNA escape to prevent worse damage [45,46]. Here, the increase in the abundance of MET2 protein only in treated cells suggests that Mo-CBP3-PepII attacks mitochondrial, which follows the results above-mentioned that Mo-CBP3-PepII induces the decoupling of Cyt c from the mitochondrial membrane (Figure 4C).
Yet, another protein that deserved attention, the clustered mitochondria protein (CMP), was only detected in the control cells and not in treated cells (Supplementary tables 1 and 2). The CMP in Fungi and many other eukaryotic organisms is essential for mitochondrial health and functioning [47,48,49]. The absence of CMP leads to misfunction of mitochondria interfering with the normal function of the cell. This result indicates that Mo-CBP3-PepII caused several damages to the mitochondrion reflectin to the whole C. neoformans cell.
2.7.3. Ligase- and Amino Acid Metabolism-Related Proteins
In the ligase group, one protein was found in both treated and non-treated C. neoformans cells, the carbamoyl-phosphate synthase (Table 1). The fold-change value, 0.07, indicates that carbamoyl-phosphate synthase (CPS) decreased in abundance after treatment with Mo-CBP3-PepII (Table 1). The CPS protein is involved in the biosynthesis of arginine [50,51]. Arginine is an essential amino acid involved in protein synthesis and in many other physiological and biochemical processes [51]. Liu et al. [51] reported that a CPS-mutant of pathogenic fungi Magnaporthe oryzae cannot produce arginine and affect the pathogenicity and development processes. Here, reducing CPS protein in C. neoformans cells could lead to arginine depletion, affecting vital cellular processes and, ultimately, death. Another protein, the Arginine biosynthesis bifunctional protein, in which two isoforms were exclusively from control cells, reinforces the hypothesis that Mo-CBP3-PepII interferes in arginine biosynthesis in C. neoformans cells (Supplementary tables 1 and 2).
Proteomic analysis revealed many proteins involved in amino acid metabolism were only detected in control cells (supplementary table 1), such as Alanine-tRNA ligase, histidine biosynthesis trifunctional protein, C-1-tetrahydrofolate synthase, glycine dehydrogenase. After treatment with the peptide, the decrease in abundance in all those proteins indicates a shutdown in protein synthesis [52]. The inability of the cell to produce proteins after the treatment with a drug causes severe forms of stress and inhibits cells from responding to stresses leading to death [52]. An antifungal drug called sordarin inhibits protein synthesis in pathogenic yeasts [53]. However, many yeasts, including C. neoformans, are intrinsically resistant to sordarin, making this drug useless for treatment. Nowadays, there are no drugs available that affect the protein synthesis of Fungi, which makes more attractive the effect of Mo-CBP3-PepII in inhibiting the protein synthesis of C. neoformans.
2.7.4. Oxidoreductase-Related Proteins
In this group, one protein, D-2-hydroxyglutarate-pyruvate transhydrogenase, presented a fold-change value indicating its up-accumulation of it after exposure of C. neoformans cells to Mo-CBP3-PepII (Table 1). The D-2-hydroxyglutarate-pyruvate transhydrogenase is mainly involved in the metabolism of lactate and oxidation of NADH to produce NAD+ in the absence of LDH activity [54,55]. In Saccharomyces cerevisiae, the D-2-hydroxyglutarate-pyruvate transhydrogenase, also known as minor D-LDH, catalyzes the conversion of D-2-hydroxyglutarate into α-ketoglutarate using FAD+ as a cofactor, and pyruvate as the donor of electrons producing lactate. The α-ketoglutarate in the cytosol is rapidly converted into 2-hydroxyglutarate, which is degraded in the cytosol by D-2-hydroxyglutarate-pyruvate transhydrogenase consuming NADH restoring NAD+ [54,55].
As shown before, the activity of LDH is inhibited in cells after treatment with Mo-CBP3-PepII interfering in NAD+ restoring and impairing the glycolysis to keep stand. The up-accumulation of D-2-hydroxyglutarate-pyruvate transhydrogenase suggests the C. neoformans cells are trying to find a way to continue with the glycolysis pathway by employing an alternative way to maintain acceptable levels of NAD+ essential do glycolysis pathway.
2.7.5. Protein Binding-Related Proteins
By analyzing proteins in this group, an unexpected result was seen. Many proteins components of the 26S proteasome, such as 26S proteasome regulatory subunit RPN1, 26S proteasome regulatory subunit RPN5, U3 small nucleolar RNA-associated, ERAD-associated E3 ubiquitin-protein, Uba3-binding protein but2, Ubiquinone biosynthesis protein, were only detected in control cells being absent in treated cells (Supplementary tables 1 and 2). In eukaryotic cells, the proteasome is a multicomplex enzymatic system that plays a role in protein turnover and many other cell processes, such as development, growth, division, cell-cycle progression, and defense [56,57,58]. The misfunction of proteasome could trap cells in a cell-cycle arrest and, consequently, apoptosis [57].
Here, the proteomic analysis revealed many proteasome subunits present in control cells, which is essential to its function, disappeared after the treatment with Mo-CBP3-PepII (Supplementary Table 2). This result suggests that C. neoformans do not have a functional proteasome and could start apoptosis. This idea agrees with the results of Aguiar et al. [10], where Mo-CBP3-PepII induces apoptosis in C. neoformans cells.
Another protein unique from the control cell was ASI1 (Supplementary Table 1). The ASI1 is a nuclear inner membrane-attached protein involved in correctly maintaining gene expression regulation [59,60,61]. The protein ASI1 is involved in the functional folding of a group of transcription factors known as Stp proteins. In the absence of ASI1, unprocessed forms of Stp proteins were produced, leading to cell failure in control gene expression, involved in ribosomal RNA (rRNA) production [61]. So, somehow Mo-CBP3-PepII induces alteration in the expression of rRNA and consequently reduces ribosome in C. neoformans treated cells. This result agrees with the shutting down in protein synthesis discussed above.
2.7.6. Transferase-Related Proteins
Eleven proteins in this group presented overlapping in the treated and control cells (Table 1). Of these, 6 proteins were up-accumulated, and 5 did not change. First is the atypical kinase COQ8, with a fold-chance value of 1.81 (Table 1). The atypical kinase COQ8 is a mitochondrial protein involved in coenzyme Q (CoQ) biosynthesis [62,63]. CoQ is a molecule involved in at least two critical processes in eukaryotic cells: (1) acting as electron transport in ETC and (2) working as an antioxidant [62,63]. Here, we reasoned the increase in atypical kinase COQ8 to increase the CoQ levels is a response of C. neoformans cells to two stresses imposed by Mo-CBP3-PepII. First, high production of CoQ could be involved in a repair process of ETC, which is affected by the decoupling of Cyt c from the mitochondrial membrane (Figure 4C). As discussed above, the release of Cyt c from mitochondrial jeopardizes the ETC and depletes the ATP synthesis. Second, higher CoQ levels might be associated with its antioxidant activity as a defense against ROS overaccumulation induced by Mo-CBP3-PepII (Figure 1).
The threonylcarbamoyl-AMP synthase presented a fold-change of 8.81 (Table 1), one of the higher evaluated in C. neoformans cells treated with Mo-CBP3-PepII. The threonylcarbamoyl-AMP synthase is an important enzyme involved in the production of threonylcarbamoyl-AMP, a central metabolite essential for biosynthesis L-threonine and a universal tRNA nucleoside N6-threonylcarbamoyl adenosine involved in the maturation of tRNA [64,65]. The increase of threonylcarbamoyl-AMP synthase might be an attempt of C. neoformans cells to overcome the reduction in protein synthesis, as suggested above.
The enzyme (2E.6E)-farnesyl diphosphate synthase (FPPS) presented a fold-change of 11.12 in treated cells compared to control cells (Table 1). The FPPS protein is a crucial enzyme involved in a central biochemical pathway for eukaryotic cells, the isoprenoid biosynthesis pathway [66,67,68,69]. The isoprenoid biosynthesis produces sesquiterpenes that supply the production of many essential metabolites such as ubiquinone, dolichols, and sterols; in our case, for fungi, the participation in sterols biosynthesis leads to ergosterol biosynthesis [66,67,68,69]. Here, it was reported that Mo-CBP3-PepII inhibits 60% of the biosynthesis of ergosterol, which in turn compromises the health of cellular membranes [67,68,69]. Altogether, this high fold-change value of FPPS led us to hypothesize that C. neoformans is trying to compensate for the inhibition of ergosterol biosynthesis to alleviate the damage to the membrane keeping it healthy and functional.
Another protein that was up-accumulated in the treated cells compared to the control was the spindle assembly checkpoint kinase (SAC). The SAC is a signal protein that indicates the mistaken attachment of the mitotic spindle to the kinetochores of chromosomes [70,71,72]. The SAC protein is vital during the checkpoint in the cell cycle. SAC protein is essential to check the correct position of chromosomes during the transition of metaphase-to-anaphase. If something goes wrong in this process, SAC protein accumulates and negatively regulates the CDC20 inhibiting cell division to prevent duplicated chromosome separation. The cell cycle is stopped until chromosomes are correctly aligned to the spindle [70,71,72]. The higher accumulation of SAC proteins in C. neoformans cells treated with Mo-CBP3-PepII indicates an inhibition of the cellular cycle, decreasing the rate of cell division and, thus, in the case of C. neoformans, infection and pathogenicity [70,71,72].
2.7.7. Transport or Structural Activity-Related Proteins
Most of the overlapping proteins in this group decrease in accumulation in treated/control cells (Table 1), including Low-affinity methionine permease. Methionine is an essential proteinogenic amino acid [73,74]. As an essential amino acid, cells cannot synthesize methionine and must obtain it from the environment. To do so, cells use transports attached to the membrane to collect amino acids [73,74]. The low-affinity methionine permease transporter is used by C. neoformans cells to collect methionine from the environment. The Mo-CBP3-PepII-treated C. neoformans cells presented a decrease in abundance of this transporter, leading to a decrease of methionine concentration within the cell, interfering in protein synthesis [73,74].
Another transporter, histidine permease, also decreased in abundance in C. neoformans cells after exposure to Mo-CBP3-PepII (Supplementary Table 1). As happens to methionine, histidine is a proteinogenic essential amino acid that has to be collected by cells from the environment [75,76,77,78]. However, otherwise, than methionine permease, histidine permease functions go beyond histidine transport. In Fungi, histidine permeases are essential to keep fungi safe, virulent, vigorous, and normal morphogenesis and development [77,78]. The absence of this protein in C. neoformans cells treated with Mo-CBP3-PepII suggests that all processes developed by it are compromised, making it hard for the fungus to overcome stresses imposed by the peptide.
Another protein that decreased in abundance in C. neoformans cells exposure to Mo-CBP3-PepII was the oligomycin resistance ATP-dependent permease YOR1 (Table 1). Oligomycin is a molecule produced by bacteria from the Streptomyces genus and is used as an antibiotic. Oligomycin binds to ATP synthase inhibiting ATP synthesis [79,80,81]. Over the years, pathogenic yeasts have developed resistance to oligomycin by producing oligomycin resistance ATP-dependent permease YOR1 [80]. Thus, the reduction in the abundance of oligomycin resistance ATP-dependent permease YOR1 suggests that C. neoformans cells became susceptible to oligomycin after treatment with Mo-CBP3-PepII.
A similar result was recently published by Branco et al. [22], revealing that the treatment of Klebsiella pneumoniae cells with the Mo-CBP3-PepI synthetic peptide also induces the reduction in three multidrug resistance proteins such as multidrug resistance protein MdtN, UPF0194 membrane protein YbhG, and multidrug resistance protein EmrK. This result suggests Mo-CBP3-PepI increases the susceptibility of K. pneumoniae to drugs. The same result was found in C. neoformans after treatment with Mo-CBP3-PepII.
The BNI4 protein with the highest fold-change of 66.18 shows its significant accumulation in C. neoformans cells after treatment with Mo-CBP3-PepII (Table 1). The BNI4 protein is involved in the recruitment of chitin synthase to produce the chitin to be incorporated in the cell wall of the new buddy [82,83]. Yeasts divide by cytokinesis, a process where one cell divides itself in two [84]. During this process, the cell wall needs to be produced for two cells; for this process, chitin synthase has to be recruited by the BNI4 protein [82,83,84].
We have one hypothesis for this higher level of BNI4 protein. Mo-CB3-PepII is a synthetic peptide designed from the sequence of a chitin-binding protein from Moringa oleifera seeds, Mo-CBP3 [11]. Lima et al. [85] proved that Mo-CBP3-PepII is a chitin-binding peptide and causes damage to the cell wall of Candida albicans by interacting with chitin. Scanning electron microscopy reported by Aguiar et al. [10] and AFM analysis done here (Figure 5) strongly suggest that Mo-CBP3-PepII also in C. neoformans cell wall. As such, we hypothesize that Mo-CBP3-PepII might interact with chitin and thus interfere with producing cell walls for new buds. To cope with this insult, C. neoformans cells raised the levels of BNI4 as much as possible to recruit chitin synthase to produce cell wall for new buds.
In contrast to BNI4, coronin is the protein with the lowest fold-change value of 0.04 (Table 1) and thus decreases in abundance in C. neoformans cells treated with Mo-CBP3-PepII. Coronin is a protein critical for cell structure because it interacts with actin filaments and microtubules, promoting cellular processes such as remodeling the cell cytoskeleton, cell motility, endocytosis, and phagocytosis [86,87,88]. In yeasts, Cai et al. [88] reported that coronin protects actin filaments from depolymerization keeping the cytoskeleton functional. In contrast, in coronin-mutant yeasts, the function of actin filaments and cytoskeleton are compromised [88]. Those coronin-mutant yeasts lost the ability to remodel the cytoskeleton as well as have other cytoskeleton-dependent functions compromised. Here, the dramatic reduction in the levels of coronin in peptide-treated C. neoformans cells suggests that Mo-CBP3-PepII is affecting the well-function of the cytoskeleton and inhibiting critical processes to cell life.
2.7.8. Energetic Metabolism-Related Proteins
The most exciting proteins were identified exclusively in C. neoformans cells treated com Mo-CBP3-PepII (Supplementary Table 2). The first protein found was Cytochrome c mitochondrial import. The Cytochrome c mitochondrial import is a protein that imports the cytoplasmic cytochrome c to the mitochondrial membrane [89,90,91,92]. The exclusive identification of this protein in C. neoformans cells treated with Mo-CBP3-PepII is very exciting. As pointed out above, Mo-CBP3-PepII induced the decoupling of Cyt c from the mitochondrial membrane (Figure 4C). Therefore, the exclusive identification of Cytochrome c mitochondrial import in C. neoformans treated cells indicates strongly suggests the cell is trying to respond to the stresses caused by peptide either by producing new Cyt c molecules or even recoupling the molecules released by the peptide.
Another protein exclusively identified in treated cells was alcohol dehydrogenase 4 (supplementary table 2). Alcohol dehydrogenase functions in the alcoholic fermentation of yeasts [93,94]. In alcohol fermentation, the pyruvate is produced in glycolysis. After glycolysis, pyruvate is driven to a two-reaction reaction, where it is converted into ethanol in a step catalyzed by alcohol dehydrogenase [93,94]. The point in the alcoholic formation is that, in the reduction of pyruvate to ethanol, the NADH cytosolic produced in the glycolysis is converted into NAD+, raising the ratio NAD+/NADH and thus favoring the glycolysis [93,94].
The point is, in the C. neoformans treated cells, a reduction in the activity of LDH induced by Mo-CBP3-PepII (Figure 4A). LDH recovered the NAD+ by lactic fermentation, keeping high the NAD+/NADH ratio to favor glycolysis [31]. So, it is feasible to suggest that the high level of Alcohol dehydrogenase is an attempt of C. neoformans cells to compensate for the inhibition of LDH by Mo-CBP3-PepII and stand high the NAD+/NADH ratio to favor glycolysis.
2.7.9. Pathogenicity-Related Proteins
In this group, one protein exclusively detected in control cells and absent in the treated deserves attention, the Subtilisin-like protease 6 (Supplementary table 1 and table 2). The subtilisin-like proteases are enzymes in fungi autophagy and fungal virulence, enhancing invasion and colonization [95,96]. The absence of Subtilisin-like protease 6 after treatment with Mo-CBP3-PepII (supplementary table 2) indicates the C. neoformans cells stay with a limited capacity to cause infection.
3. Conclusion
The results presented here showed that Mo-CBP3-PepII displays its anticryptococcal activity by multiple mechanisms of action affecting several cellular processes essential for cell life, such as development, pathogenesis, cell division, and metabolism. Acting on cells in multiple ways, Mo-CBP3-PepII difficult the development of resistance by C. neoformans. Additionally, our results suggest that Mo-CBP3-PepII could act as an adjuvant for drugs that are becoming useless. Additionally, recent studies showed that Mo-CBP3-PepII is a safe molecule presenting no toxicity to human erythrocytes, MRC-5, HaCAT, L292 human cell lines, and zebrafish embryos [11,97]. Therefore, it is feasible to suggest that Mo-CBP3-PepII is a potential molecule to be employed alone or in combination with other drugs to develop new treatments against C. neoformans.
4. Materials and Methods
4.1. Fungal Strains, Chemicals, and Synthetic Peptides
Cryptococcus neoformans (ATCC 32045) was obtained from the Department of Biochemistry and Molecular Biology at the Federal University of Ceará (UFC), Fortaleza, Brazil. All the chemicals used in the experiments were obtained from Sigma Aldrich (São Paulo, SP, Brazil). The Mo-CBP3-PepII (NIQPPCRCC) synthetic peptide was chemically synthesized by the company ChemPeptide (Shanghai, China).
4.2. Antifungal Assay
The antifungal assay was performed according to Aguiar et al. [10]. The cryptococcal cells were cultivated in yeast potato dextrose (YPD) agar for 10 days. Then cells were collected and transferred to a liquid YPD medium. The concentration of Mo-CBP3-PepII was the MIC50 (25 µg mL−1) defined by Aguiar et al. [10]. This MIC50 concentration was used in all experiments performed in this study. Therefore, 25 µL of C. neoformans cells (106 cells mL−1) and 25 µL of Mo-CBP3-PepII (25 µg mL−1) were incubated for 24 h at 30 °C before each assay. DMSO-NaCl was the control used.
4.3. Detection of Mo-CBP3-PepII-Induced Overproduction of ROS
Fluorescence microscopy assay was performed to evaluate the Mo-CBP3-PepII-induced H2O2 overproduction, as described previously by Dias et al. [98]. After the antifungal assay (Section 4.2), the samples were washed with 0.15 M NaCl and centrifugated (5000 × g 5 min at 4 °C) three times and then were incubated with 9 µL of 2′,7′ dichlorofluorescein diacetate (DCFH-DA, Sigma, St. Louis, MI, 364 USA) for 30 min in the dark at 22 ± 2 °C. Next, the samples were washed and centrifuged as mentioned, transferred to slides, and observed with a fluorescence microscope (Olympus System BX 41, Tokyo, Japan) with an excitation wavelength of 535 nm and 370 emission wavelength of 617 nm.
Additionally, the qualitative of anion superoxide was performed according to Choi et al. [99]. The antifungal assay was done as above. Then C. neoformans cells were washed, recovered in 0.15 M of NaCl, and incubated with 100 µM nitroblue tetrazolium chloride (NBT) (Sigma Aldrich, USA) for 2 hours at 22 ± 2 °C. Then, cells were washed to remove the excess NBT and visualized in a light microscope (Olympus System BX 41, Tokyo, Japan).
The experiments to evaluate the anticryptococcal activity in the presence of the antioxidant agent ascorbic were done following the methodology described by Neto et al. [27]. The experiments to perform the H2O2 overaccumulation and pore formation in ascorbic acid at 10 mM were performed as described by Aguiar et al. [21].
4.4. Protein Extraction from C. neoformans Cells
The extraction of proteins from C. neoformans cells was done following Branco et al. [22]. After the antifungal assay, the samples were washed three times to remove the media with 50 mM Na+-acetate pH 5.2 and centrifugated at 12,000 × g for 15 min at 4 °C. Then, samples were resuspended in 200 µL of extraction buffer and frozen at -20 °C for 24 h. After that, the samples were submitted to sonication for 30 min to break the cell wall and plasmatic membrane, centrifuged again, and the supernatant was collected. The Bradford assay [100] was performed to determine the protein concentration using bovine serum albumin (BSA). The extracted proteins were used for enzymatic assay and proteomic analysis.
4.5. Activity Redox System Enzymes
4.5.1. Ascorbate Peroxidase (APX) Activity
To evaluate the APX activity, the methodology was performed according to Souza et al. [101]. In tubes, 100 μL of the treated or control samples were mixed with 800 μL of 50 mM K+-phosphate buffer, pH 6.0, containing 0.5 mM of L-ascorbic acid and incubated at 30 °C for 10 minutes. Then, 100 μL of 2 mM H2O2 was added to start the reaction. The reaction was tracked every 10 seconds until 1 min in the spectrophotometer (Biochron, Libra 394 S12) at the wavelength 290 nm. APX activity unity (AU) was expressed as the reduction in the absorbance by 0.01 at 290 nm, indicating the use of ascorbate to remove H2O2 by milligram of the protein (UA mgP-1).
4.5.2. Catalase (CAT) Activity
Following Souza et al. [101], 200 μL of proteins extracted from Mo-CBP3-PepII-treated and non-treated C. neoformans cells were incubated with 700 μL 50 mM K+-phosphate buffer pH 7.0 at 30 °C for 10 min. Next, 100 μL of H2O2 was added to start the reaction. The reaction medium was transferred to a quartz cuvette (1 cm-1), and the absorbance was measured in a spectrophotometer (Biochron, Libra 394 S12). The decrease in absorbance at 240 nm was observed at intervals of 10 seconds to 1 minute. CAT activity unity (AU) was expressed as the reduction in the absorbance by 0.01 at 240 nm, indicating the use of ascorbate to remove H2O2 by milligram of the protein (UA mgP-1).
4.5.3. Superoxide Dismutase (SOD) Activity
SOD activity was performed following the methodology described by Souza et al. [101]. In a flat-bottom 96 well-plate was mixed 10 μL of 1 M K+-phosphate buffer pH 7.8, 20 μL 1 mM 2,2’,2’’,2’’’- ethylenediaminetetraacetic acid (EDTA), 10 μL of 0.25% Triton X-100, 20 μL of 130 mM L-Methionine, 100 μL of protein extract from C. neoformans cells in the presence and absence of Mo-CBP3-PepII (MIC50), 20 μL 100 mM of riboflavin, 20 μL of 700 μM of BNT. The mixture was homogenized and kept in the dark for 5 min. After that, the plate was exposed to a 32-W fluorescent light and the absorbances were measured in 1 min intervals up to 5 min at 630 nm in a microplate reader (Epoch, Biotek, Santa Clara, CA, USA). Blanks consisted of all reagents used without protein extracts, it was replaced with deionized water. The SOD was expressed in activity units per mg of protein (AU mgP-1). One unit of SOD activity (1 AU) corresponded to the amount of the sample needed to inhibit the photoreduction of NBT by 50%.
4.6. Ergosterol Inhibition Synthesis
The inhibition of ergosterol biosynthesis was evaluated according to Neto et al. [27]. The C. neoformans cells were cultivated in the presence of Mo-CBP3-PepII (MIC50), 5% DMSO, and Itraconazole (ITR) (1000 µg mL-1) for 24 h at 30 °C. Then, the cells were centrifugated at 3000 × g for 5 min. Subsequently, the pellet was dried and weighed. Two mL of 25% alcoholic sodium hydroxide solution (m/v) was added to each pellet and strongly vortexed for 1 min. To extract the sterol, 4 mL of sterile 75% n-hexane was added and vortexed for 3 min. Then, 400 μL of pure ethanol was added to 200 μL sterol extract. Next, the samples were transferred to a quartz cuvette (1 cm) and the absorbance was measured in a spectrophotometer (Biochron, Libra 394 S12) at 230 nm and 282 nm. Ergosterol content was calculated based on three equations:
Eq. 1: % ergosterol + 24(28) [DHE=(Abs282/290) x F] / pellet weight
Eq. 2: % 23(28) DHE =[(Abs230/518) x F] / pellet weight
Eq. 3: % ergosterol = % ergosterol + 24(28) DHE - % 24(28) DHE
24 (28) DHE refers to dehydroergosterol, where the absorbance is similar to ergosterol at 282 nm. F, in both equations, represents the factor of dilution of ethanol.
4.7. Lactate Dehydrogenase (LDH) Activity
The LDH Liquiform™ kit (Labtest Diagnóstica, BR) was measured according to the manufacturer’s instructions.
4.8. Cytochrome C Release
The assay was performed according to Neto et al. [27]. The cells were incubated with Mo-CBP3-PepII (MIC50), H2O2 (10 mM), and DMSO-NaCl for 24 h at 30 °C. Then, 100 µL of buffer 50 mM Tris-HCl pH 7.5 containing 2 mM EDTA, 1 mM phenylmethylsulphonyl fluoride, and 6% glucose were added to the cell suspension and homogenized. Subsequently, the samples were centrifuged at 2000 x g at 4 °C for 10 min and the supernatant was collected and added into microtubes. The pellet was washed with the buffer 50 mM Tris pH 7.5 containing 2 mM EDTA and centrifugated (6000 x g for 30 sec). The supernatant was discarded, and the mitochondria were suspended in 100 µL of the same buffer. Next, the cytosolic and mitochondrial suspensions were treated with 30 mM of ascorbic acid for 5 min at 30 °C. Then, the optical density of the obtained solution was measured using a microtiter plate reader (Epoch, Biotek, Santa Clara, CA, USA) at 550 nm.
4.9. Atomic Force Microscopy (AFM)
After the antifungal assay, the cells were prepared for AFM analysis following the methodology described by Souza et al. [102]. The samples were washed three times with sterile water and centrifugated at 5000 x g for 5 min at 4 °C. Then, 10 μL of the cells were transferred to a freshly washed glass surface previously treated with (v/v) poly-L-lysine and incubated at room temperature (22 ± 2°C) for 30 min. All samples were analyzed on Multimode 8 (Bruker, Santa Barbara, CA, USA). The probes used were SNL (Bruker) with 0.06 N/m nominal spring constant, a resonance frequency of 320 kHz, and a nominal tip radius of 8 nm.
4.10. Gel-Free Proteomic Analysis
For proteomic analysis, proteins were extracted as described in Section 4.4. Subsequently, proteins from the control and treated cells were treated with 10 mM DTT and incubated for 1 h at 37 °C in the dark to reduce the proteins. Next, 15 mM iodoacetamide was added and incubated for 30 min in a dark room to alkylate the reduced proteins. Then, the proteins were digested using trypsin gold (Promega, Madison, WI, USA) to a final concentration of 1:20 (w/w) as described by manufacturers for 16 h at 37 °C. After that, the samples were dried in a speed vacuum (Eppendorf, Hamburg, Germany) for 3 h and analyzed by an ESI-QUAD-TOF mass spectrometer.
4.11. Protein Identification
The protein identification was performed following Branco et al. [22]. Tandem mass spectra were exported as .pkl files and loaded in the MASCOT MS/MS ions search from MATRIX SCIENCE (https://www.matrixscience.com/cgi/search_form.pl?FORMVER=2&SEARCH=MIS, accessed on 10 November 2022) against UP2311_S_cerevisiae (protein database), UP219602_F_oxysporum (protein database), and SwissProt databases (protein database). The search was done by following parameters: fixed modifications to Carbamidomethyl (C), variable modifications to Oxidation (O); the peptide charge was set to 2+, 3+, and 4+; and the instrument was set to ESI-QUAD-TOF. The proteins identified were searched for in UNIPROT and separated into 3 sets: 1) unique from control for those only identified in control samples, 2) unique from the cells treated with Mo-CBP3-PepII for those only identified in treated samples, and 3) Mo-CBP3-PepII x control overlapping proteins.
The proteins with a fold-change value ≥ 1.5 (p < 0.05, Tukey’s test) were up-accumulated (increased the abundance), and proteins with a fold-change value ≤ 0.5 (p < 0.05, Tukey’s test) were down-accumulated (decreased the abundance) and considered for comparisons. Proteins with a fold-change value between 0.5 to 1.5 were considered that did not change. For each protein, its corresponding FASTA file was downloaded. The blast2go program (https://www.blast2go.com/, accessed on 25 November 2022) was used to categorize the proteins detected by Gene Ontology (GO) annotation according to Molecular function, Biological Activity, and subcellular location.
4.12. Statistical Analysis
All experiments were performed in triplicates. The statistical analyses were performed using GraphPad Prism (version 5.01) for Microsoft Windows. All data obtained in the assays were compared using the one-way analysis of variance (ANOVA), followed by the Tukey test (p < 0.05).
Author Contributions
All authors made substantial contributions. The conception and design of the study and acquisition of data, analysis, and interpretation were performed by T.K.B.A., N.A.S.N., F.I.R.G., C.D.T.F., F.P.M., R.F.C., C.S.N., L.M.R.A., R.S.-O., J.T.A.O., and P.F.N.S. Microscopic analyses were carried out by T.K.B.A., N.A.S.N., L.M.R.A., R.S.-O., and P.F.N.S. Writing and revision of the article were carried out by T.K.B.A. and P.F.N.S. Lastly, P.F.N.S. performed the final approval and submission. All authors have read and agreed to the published version of the manuscript.
Funding
Special thanks to CAPES for providing the postdoctoral grant to Pedro F. N. Souza (grant number 88887.318820/2019-00).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting this study’s findings are available upon request from the corresponding author.
Acknowledgments
We are grateful to the staff of the central analytical facilities of UFC, Brazil.
Conflicts of Interest
The authors report no conflicts of interest.
References
- Seiler, G.T.; Ostrosky-Zeichner, L. Investigational Agents for the Treatment of Resistant Yeasts and Molds. Curr. Fungal Infect. Rep. 2021, 15, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.G.; Paterson, D.L. How Urgent Is the Need for New Antifungals? Expert Opin. Pharmacother. 2021, 22, 1857–1870. [Google Scholar] [CrossRef] [PubMed]
- Organization, W.H. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action 2022, 1–48.
- Woo, Y.H.; Martinez, L.R. Cryptococcus Neoformans –Astrocyte Interactions: Effect on Fungal Blood Brain Barrier Disruption, Brain Invasion, and Meningitis Progression. Crit. Rev. Microbiol. 2021, 47, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.; Casadevall, A. Cryptococcal Therapies and Drug Targets: The Old, the New and the Promising. Cell. Microbiol. 2016, 18, 792–799. [Google Scholar] [CrossRef] [PubMed]
- Maligie, M.A.; Selitrennikoff, C.P. Cryptococcus Neoformans Resistance to Echinocandins: (1,3)β-Glucan Synthase Activity Is Sensitive to Echinocandins. Antimicrob. Agents Chemother. 2005, 49, 2851–2856. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic Antimicrobial Peptides: From Choice of the Best Sequences to Action Mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Erak, M.; Bellmann-Sickert, K.; Els-Heindl, S.; Beck-Sickinger, A.G. Peptide Chemistry Toolbox – Transforming Natural Peptides into Peptide Therapeutics. Bioorg. Med. Chem. 2018, 26, 2759–2765. [Google Scholar] [CrossRef] [PubMed]
- Kundu, R. Cationic Amphiphilic Peptides: Synthetic Antimicrobial Agents Inspired by Nature. ChemMedChem 2020, 15, 1887–1896. [Google Scholar] [CrossRef]
- Aguiar, T.K.B.; Neto, N.A.S.; Freitas, C.D.T.; Silva, A.F.B.; Bezerra, L.P.; Malveira, E.A.; Branco, L.A.C.; Mesquita, F.P.; Goldman, G.H.; Alencar, L.M.R.; et al. Antifungal Potential of Synthetic Peptides against Cryptococcus Neoformans: Mechanism of Action Studies Reveal Synthetic Peptides Induce Membrane–Pore Formation, DNA Degradation, and Apoptosis. Pharmaceutics 2022, 14, 1678. [Google Scholar] [CrossRef]
- Oliveira, J.T.A.; Souza, P.F.N.; Vasconcelos, I.M.; Dias, L.P.; Martins, T.F.; Van Tilburg, M.F.; Guedes, M.I.F.; Sousa, D.O.B. Mo-CBP3-PepI, Mo-CBP3-PepII, and Mo-CBP3-PepIII Are Synthetic Antimicrobial Peptides Active against Human Pathogens by Stimulating ROS Generation and Increasing Plasma Membrane Permeability. Biochimie 2019, 157, 10–21. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, Y.; Li, B.; Chen, T.; Tian, S. Reactive Oxygen Species: A Generalist in Regulating Development and Pathogenicity of Phytopathogenic Fungi. Comput. Struct. Biotechnol. J. 2020, 18, 3344. [Google Scholar] [CrossRef]
- Cho, J.; Hwang, I. sok; Choi, H.; Hwang, J.H.; Hwang, J.S.; Lee, D.G. The Novel Biological Action of Antimicrobial Peptides via Apoptosis Induction. J. Microbiol. Biotechnol. 2012, 22, 1457–1466. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento Dias, J.; de Souza Silva, C.; de Araújo, A.R.; Souza, J.M.T.; de Holanda Veloso Júnior, P.H.; Cabral, W.F.; da Glória da Silva, M.; Eaton, P.; de Souza de Almeida Leite, J.R.; Nicola, A.M.; et al. Mechanisms of Action of Antimicrobial Peptides ToAP2 and NDBP-5.7 against Candida Albicans Planktonic and Biofilm Cells. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Cho, J.; Lee, D.G. Oxidative Stress by Antimicrobial Peptide Pleurocidin Triggers Apoptosis in Candida Albicans. Biochimie 2011, 93, 1873–1879. [Google Scholar] [CrossRef] [PubMed]
- Maurya, I.K.; Pathak, S.; Sharma, M.; Sanwal, H.; Chaudhary, P.; Tupe, S.; Deshpande, M.; Chauhan, V.S.; Prasad, R. Antifungal Activity of Novel Synthetic Peptides by Accumulation of Reactive Oxygen Species (ROS) and Disruption of Cell Wall against Candida Albicans. Peptides 2011, 32, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Uhl, L.; Gerstel, A.; Chabalier, M.; Dukan, S. Hydrogen Peroxide Induced Cell Death: One or Two Modes of Action? Heliyon 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Wang, R.; Ding, N.; Liu, X.; Zheng, N.; Fu, B.; Sun, L.; Gao, P. Reactive Oxygen Species-Mediated Caspase-3 Pathway Involved in Cell Apoptosis of Karenia Mikimotoi Induced by Linoleic Acid. Algal Res. 2018, 36, 48–56. [Google Scholar] [CrossRef]
- Keyer, K.; Imlay, J.A. Superoxide Accelerates DNA Damage by Elevating Free-Iron Levels. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 13635. [Google Scholar] [CrossRef]
- Aguiar, T.K.B.; Neto, N.A.S.; Silva, R.R.S.; Freitas, C.D.T.; Mesquita, F.P.; Alencar, L.M.R.; Santos-oliveira, R.; Goldman, G.H.; Souza, P.F.N. Behind the Curtain : In Silico and In Vitro Experiments Brought to Light New Insights into the Anticryptococcal Action of Synthetic Peptides. 2023, 1–18.
- Branco, L.A.C.; Souza, P.F.N.; Neto, N.A.S.; Aguiar, T.K.B.; Silva, A.F.B.; Carneiro, R.F.; Nagano, C.S.; Mesquita, F.P.; Lima, L.B.; Freitas, C.D.T. New Insights into the Mechanism of Antibacterial Action of Synthetic Peptide Mo-CBP3-PepI against Klebsiella Pneumoniae. Antibiotics 2022, 11, 1753. [Google Scholar] [CrossRef]
- Belozerskaya, T.A.; Gessler, N.N. Reactive Oxygen Species and the Strategy of Antioxidant Defense in Fungi: A Review. Appl. Biochem. Microbiol. 2007, 43, 506–515. [Google Scholar] [CrossRef]
- Asakura, H.; Kitahora, T. Antioxidants and Polyphenols in Inflammatory Bowel Disease: Ulcerative Colitis and Crohn Disease. Polyphenols Prev. Treat. Hum. Dis. 2018, 279–292. [Google Scholar] [CrossRef]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.-P.; Fleury, M.J.J. Microbial Antioxidant Defense Enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and Function of Ascorbate Peroxidase Isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef]
- da Silva Neto, J.X.; da Costa, H.P.S.; Vasconcelos, I.M.; Pereira, M.L.; Oliveira, J.T.A.; Lopes, T.D.P.; Dias, L.P.; Araújo, N.M.S.; Moura, L.F.W.G.; Van Tilburg, M.F.; et al. Role of Membrane Sterol and Redox System in the Anti-Candida Activity Reported for Mo-CBP2, a Protein from Moringa Oleifera Seeds. Int. J. Biol. Macromol. 2020, 143, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Cirac, A.D.; Moiset, G.; Mika, J.T.; Koçer, A.; Salvador, P.; Poolman, B.; Marrink, S.J.; Sengupta, D. The Molecular Basis for Antimicrobial Activity of Pore-Forming Cyclic Peptides. Biophys. J. 2011, 100, 2422. [Google Scholar] [CrossRef] [PubMed]
- Lipkin, R.; Lazaridis, T. Computational Studies of Peptide-Induced Membrane Pore Formation. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Lemetais, G.; Ferreira, T.; Cayot, P.; Gervais, P.; Beney, L. Ergosterol Biosynthesis: A Fungal Pathway for Life on Land? Evolution 2012, 66, 2961–2968. [Google Scholar] [CrossRef]
- Farhana, A.; Lappin, S.L. Biochemistry, Lactate Dehydrogenase. StatPearls 2022. [Google Scholar]
- Ow, Y.L.P.; Green, D.R.; Hao, Z.; Mak, T.W. Cytochrome c: Functions beyond Respiration. Nat. Rev. Mol. Cell Biol. 2008, 9, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Swan, C.L.; Sistonen, L. Cellular Stress Response Cross Talk Maintains Protein and Energy Homeostasis. EMBO J. 2015, 34, 267–269. [Google Scholar] [CrossRef]
- Ishida, K.; Cipriano, T.F.; Rocha, G.M.; Weissmüller, G.; Gomes, F.; Miranda, K.; Rozental, S. Silver Nanoparticle Production by the Fungus Fusarium Oxysporum: Nanoparticle Characterisation and Analysis of Antifungal Activity against Pathogenic Yeasts. Mem. Inst. Oswaldo Cruz 2014, 109, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, M.; Chiriac, A.I.; Otto, A.; Zweytick, D.; May, C.; Schumacher, C.; Gust, R.; Albada, H.B.; Penkova, M.; Krämer, U.; et al. Small Cationic Antimicrobial Peptides Delocalize Peripheral Membrane Proteins. Proc. Natl. Acad. Sci. U. S. A. 2014, 111, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Senges, C.H.R.; Stepanek, J.J.; Wenzel, M.; Raatschen, N.; Ay, Ü.; Märtens, Y.; Prochnow, P.; Hernández, M.V.; Yayci, A.; Schubert, B.; et al. Comparison of Proteomic Responses as Global Approach to Antibiotic Mechanism of Action Elucidation. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef]
- Maaß, S.; Bartel, J.; Mücke, P.A.; Schlüter, R.; Sura, T.; Zaschke-Kriesche, J.; Smits, S.H.J.; Becher, D. Proteomic Adaptation of Clostridioides Difficile to Treatment with the Antimicrobial Peptide Nisin. Cells 2021, 10, 1–21. [Google Scholar] [CrossRef]
- Tsakou, F.; Jersie-Christensen, R.; Jenssen, H.; Mojsoska, B. The Role of Proteomics in Bacterial Response to Antibiotics. Pharmaceuticals 2020, 13, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Moyer, T.B.; Purvis, A.L.; Wommack, A.J.; Hicks, L.M. Proteomic Response of Escherichia Coli to a Membrane Lytic and Iron Chelating Truncated Amaranthus Tricolor Defensin. BMC Microbiol. 2021, 21, 1–12. [Google Scholar] [CrossRef]
- Santi, L.; Beys-Da-Silva, W.O.; Berger, M.; Calzolari, D.; Guimarães, J.A.; Moresco, J.J.; Yates, J.R. Proteomic Profile of Cryptococcus Neoformans Biofilm Reveals Changes in Metabolic Processes. J. Proteome Res. 2014, 13, 1545–1559. [Google Scholar] [CrossRef] [PubMed]
- Noronha Souza, P.F.; Abreu Oliveira, J.T.; Vasconcelos, I.M.; Magalhães, V.G.; Albuquerque Silva, F.D.; Guedes Silva, R.G.; Oliveira, K.S.; Franco, O.L.; Gomes Silveira, J.A.; Leite Carvalho, F.E. H2O2Accumulation, Host Cell Death and Differential Levels of Proteins Related to Photosynthesis, Redox Homeostasis, and Required for Viral Replication Explain the Resistance of EMS-Mutagenized Cowpea to Cowpea Severe Mosaic Virus. J. Plant Physiol. 2020, 245, 153110. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.L.; Baranowski, J.; Fostel, J.; Montgomery, D.A.; Lartey, P.A. DNA Topoisomerases from Pathogenic Fungi: Targets for the Discovery of Antifungal Drugs. Antimicrob. Agents Chemother. 1992, 36, 2778–2784. [Google Scholar] [CrossRef]
- Costa-Silva, H.M.; Resende, B.C.; Umaki, A.C.S.; Prado, W.; da Silva, M.S.; Virgílio, S.; Macedo, A.M.; Pena, S.D.J.; Tahara, E.B.; Tosi, L.R.O.; et al. DNA Topoisomerase 3α Is Involved in Homologous Recombination Repair and Replication Stress Response in Trypanosoma Cruzi. Front. cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y.; Redon, C.; Rao, V.A.; Seiler, J.A.; Sordet, O.; Takemura, H.; Antony, S.; Meng, L.H.; Liao, Z.Y.; Kohlhagen, G.; et al. Repair of and Checkpoint Response to Topoisomerase I-Mediated DNA Damage. Mutat. Res. - Fundam. Mol. Mech. Mutagen. 2003, 532, 173–203. [Google Scholar] [CrossRef]
- Saita, S.; Shirane, M.; Nakayama, K.I. Selective Escape of Proteins from the Mitochondria during Mitophagy. Nat. Commun. 2013 41 2013, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Treviño, P.; Velásquez, M.; García, N. Mechanisms of Mitochondrial DNA Escape and Its Relationship with Different Metabolic Diseases. Biochim. Biophys. Acta - Mol. Basis Dis. 2020, 1866, 165761. [Google Scholar] [CrossRef]
- Huang, P.; Yu, T.; Yoon, Y. Mitochondrial Clustering Induced by Overexpression of the Mitochondrial Fusion Protein Mfn2 Causes Mitochondrial Dysfunction and Cell Death. Eur. J. Cell Biol. 2007, 86, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Fields, S.D.; Arana, Q.; Heuser, J.; Clarke, M. Mitochondrial Membrane Dynamics Are Altered in CluA- Mutants of Dictyostelium. J. Muscle Res. Cell Motil. 2002, 23, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Hulen, D.; Liu, T.; Clarke, M. The CluA- Mutant of Dictyostelium Identifies a Novel Class of Proteins Required for Dispersion of Mitochondria. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 7308–7313. [Google Scholar] [CrossRef] [PubMed]
- Nicoloff, H.; Hubert, J.C.; Bringel, F. In Lactobacillus Plantarum, Carbamoyl Phosphate Is Synthesized by Two Carbamoyl-Phosphate Synthetases (CPS): Carbon Dioxide Differentiates the Arginine-Repressed from the Pyrimidine-Regulated CPS. J. Bacteriol. 2000, 182, 3416–3422. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cai, Y.; Zhang, X.; Zhang, H.; Zheng, X.; Zhang, Z. Carbamoyl Phosphate Synthetase Subunit MoCpa2 Affects Development and Pathogenicity by Modulating Arginine Biosynthesis in Magnaporthe Oryzae. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- W, P.; CG, P.; G, M. Shut-down of Translation, a Global Neuronal Stress Response: Mechanisms and Pathological Relevance. Curr. Pharm. Des. 2007, 13, 1887–1902. [Google Scholar] [CrossRef]
- Domínguez, J.M.; Kelly, V.A.; Kinsman, O.S.; Marriott, M.S.; Gómez De Las Heras, F.; Martín, J.J. Sordarins: A New Class of Antifungals with Selective Inhibition of the Protein Synthesis Elongation Cycle in Yeasts. Antimicrob. Agents Chemother. 1998, 42, 2274–2278. [Google Scholar] [CrossRef] [PubMed]
- Signalling between Mitochondria and the Nucleus Regulates the Expression of a New D-lactate Dehydrogenase Activity in Yeast - Chelstowska - 1999 - Yeast - Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/pdf/10.1002/%28SICI%291097-0061%2819990930%2915%3A13%3C1377%3A%3AAID-YEA473%3E3.0.CO%3B2-0 (accessed on 17 January 2023).
- Becker-Kettern, J.; Paczia, N.; Conrotte, J.F.; Kay, D.P.; Guignard, C.; Jung, P.P.; Linster, C.L. Saccharomyces Cerevisiae Forms D-2-Hydroxyglutarate and Couples Its Degradation to d-Lactate Formation via a Cytosolic Transhydrogenase. J. Biol. Chem. 2016, 291, 6036–6058. [Google Scholar] [CrossRef] [PubMed]
- DH, L.; AL, G. Proteasome Inhibitors: Valuable New Tools for Cell Biologists. Trends Cell Biol. 1998, 8, 397–403. [Google Scholar] [CrossRef]
- Adams, J. The Proteasome: Structure, Function, and Role in the Cell. Cancer Treat. Rev. 2003, 29, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K. The Proteasome: Overview of Structure and Functions. Proc. Jpn. Acad. Ser. B. Phys. Biol. Sci. 2009, 85, 12. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.G.; Wang, X.; Zhang, L.; Xiong, X.; Zhang, Z.; Tang, K.; Pan, L.; Hsu, C.C.; Xu, H.; Tao, W.A.; et al. A Protein Complex Regulates RNA Processing of Intronic Heterochromatin-Containing Genes in Arabidopsis. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, E7377–E7384. [Google Scholar] [CrossRef]
- Calvanese, E.; Gu, Y. Towards Understanding Inner Nuclear Membrane Protein Degradation in Plants. J. Exp. Bot. 2022, 73, 2266–2274. [Google Scholar] [CrossRef]
- Zargari, A.; Boban, M.; Heessen, S.; Andréasson, C.; Thyberg, J.; Ljungdahl, P.O. Inner Nuclear Membrane Proteins Asi1, Asi2, and Asi3 Function in Concert to Maintain the Latent Properties of Transcription Factors Stp1 and Stp2. J. Biol. Chem. 2007, 282, 594–605. [Google Scholar] [CrossRef]
- Awad, A.M.; Nag, A.; Pham, N.V.B.; Bradley, M.C.; Jabassini, N.; Nathaniel, J.; Clarke, C.F. Intragenic Suppressor Mutations of the COQ8 Protein Kinase Homolog Restore Coenzyme Q Biosynthesis and Function in Saccharomyces Cerevisiae. PLoS One 2020, 15. [Google Scholar] [CrossRef]
- Clarke, C.F. Insights into an Ancient Atypical Kinase Essential for Biosynthesis of Coenzyme Q. Cell Chem. Biol. 2018, 25, 123–125. [Google Scholar] [CrossRef]
- Harris, K.A.; Bobay, B.G.; Sarachan, K.L.; Sims, A.F.; Bilbille, Y.; Deutsch, C.; Iwata-Reuyl, D.; Agris, P.F. NMR-Based Structural Analysis of Threonylcarbamoyl-AMP Synthase and Its Substrate Interactions. J. Biol. Chem. 2015, 290, 20032–20043. [Google Scholar] [CrossRef] [PubMed]
- Swinehart, W.; Deutsch, C.; Sarachan, K.L.; Luthra, A.; Bacusmo, J.M.; de Crécy-Lagard, V.; Swairjo, M.A.; Agris, P.F.; Iwata-Reuyl, D. Specificity in the Biosynthesis of the Universal TRNA Nucleoside N6-Threonylcarbamoyl Adenosine (T6A)—TsaD Is the Gatekeeper. RNA 2020, 26, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Kim, D.K.; Bae, S.H.; Gwak, H.R.; Jeon, J.H.; Kim, J.K.; Lee, B. Il; You, H.J.; Shin, D.H.; Kim, Y.H.; et al. Farnesyl Diphosphate Synthase Is Important for the Maintenance of Glioblastoma Stemness. Exp. Mol. Med. 2018, 50. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, S.M.S.; Kellogg, B.A.; Poulter, C.D. Farnesyl Diphosphate Synthase. Altering the Catalytic Site to Select for Geranyl Diphosphate Activity. Biochemistry 2000, 39, 15316–15321. [Google Scholar] [CrossRef]
- Thulasiram, H. V.; Poulter, C.D. Farnesyl Diphosphate Synthase: The Art of Compromise between Substrate Selectivity and Stereoselectivity. J. Am. Chem. Soc. 2006, 128, 15819–15823. [Google Scholar] [CrossRef] [PubMed]
- Dhar, M.K.; Koul, A.; Kaul, S. Farnesyl Pyrophosphate Synthase: A Key Enzyme in Isoprenoid Biosynthetic Pathway and Potential Molecular Target for Drug Development. N. Biotechnol. 2013, 30, 114–123. [Google Scholar] [CrossRef]
- Szymiczek, A.; Carbone, M.; Pastorino, S.; Napolitano, A.; Tanji, M.; Minaai, M.; Pagano, I.; Mason, J.M.; Pass, H.I.; Bray, M.R.; et al. Inhibition of the Spindle Assembly Checkpoint Kinase Mps-1 as a Novel Therapeutic Strategy in Malignant Mesothelioma. Oncogene 2017, 36, 6501–6507. [Google Scholar] [CrossRef]
- Maia, A.R.R.; De Man, J.; Boon, U.; Janssen, A.; Song, J.Y.; Omerzu, M.; Sterrenburg, J.G.; Prinsen, M.B.W.; Willemsen-Seegers, N.; De Roos, J.A.D.M.; et al. Inhibition of the Spindle Assembly Checkpoint Kinase TTK Enhances the Efficacy of Docetaxel in a Triple-Negative Breast Cancer Model. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2015, 26, 2180–2192. [Google Scholar] [CrossRef]
- Marston, A.L.; Wassmann, K. Multiple Duties for Spindle Assembly Checkpoint Kinases in Meiosis. Front. Cell Dev. Biol. 2017, 5, 109. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, K.T.; Chiba, Y.; Jigami, Y. Mutation of High-Affinity Methionine Permease Contributes to Selenomethionyl Protein Production in Saccharomyces Cerevisiae. Appl. Environ. Microbiol. 2010, 76, 6351–6359. [Google Scholar] [CrossRef]
- Chaumontet, C.; Azzout-Marniche, D.; Blais, A.; Piedcoq, J.; Tomé, D.; Gaudichon, C.; Even, P.C. Low-Protein and Methionine, High-Starch Diets Increase Energy Intake and Expenditure, Increase FGF21, Decrease IGF-1, and Have Little Effect on Adiposity in Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 316, R486–R501. [Google Scholar] [CrossRef]
- Li, D.; Agrellos, O.A.; Calderone, R. Histidine Kinases Keep Fungi Safe and Vigorous. Curr. Opin. Microbiol. 2010, 13, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Defosse, T.A.; Sharma, A.; Mondal, A.K.; de Bernonville, T.D.; Latgé, J.P.; Calderone, R.; Giglioli-Guivarc’h, N.; Courdavault, V.; Clastre, M.; Papon, N. Hybrid Histidine Kinases in Pathogenic Fungi. Mol. Microbiol. 2015, 95, 914–924. [Google Scholar] [CrossRef]
- Nostadt, R.; Hilbert, M.; Nizam, S.; Rovenich, H.; Wawra, S.; Martin, J.; Küpper, H.; Mijovilovich, A.; Ursinus, A.; Langen, G.; et al. A Secreted Fungal Histidine- and Alanine-Rich Protein Regulates Metal Ion Homeostasis and Oxidative Stress. New Phytol. 2020, 227, 1174–1188. [Google Scholar] [CrossRef] [PubMed]
- Doige, C.A.; Ames, G.F.L. ATP-Dependent Transport Systems in Bacteria and Humans: Relevance to Cystic Fibrosis and Multidrug Resistance. Annu. Rev. Microbiol. 1993, 47, 291–319. [Google Scholar] [CrossRef] [PubMed]
- Hundal, T.; Norling, B.; Ernster, L. The Oligomycin Sensitivity Conferring Protein (OSCP) of Beef Heart Mitochondria: Studies of Its Binding to F1 and Its Function. J. Bioenerg. Biomembr. 1984, 16, 535–550. [Google Scholar] [CrossRef]
- Parker, J.H.; Trimble, I.R.; Mattoon, J.R. Oligomycin Resistance in Normal and Mutant Yeast. Biochem. Biophys. Res. Commun. 1968, 33, 590–595. [Google Scholar] [CrossRef]
- Devenish, R.J.; Prescott, M.; Boyle, G.M.; Nagley, P. The Oligomycin Axis of Mitochondrial ATP Synthase: OSCP and the Proton Channel. J. Bioenerg. Biomembr. 2000, 32, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Kozubowski, L.; Panek, H.; Rosenthal, A.; Bloecher, A.; DeMarini, D.J.; Tatchell, K. A Bni4-Glc7 Phosphatase Complex That Recruits Chitin Synthase to the Site of Bud Emergence. Mol. Biol. Cell 2003, 14, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Larson, J.R.; Kozubowski, L.; Tatchell, K. Changes in Bni4 Localization Induced by Cell Stress in Saccharomyces Cerevisiae. J. Cell Sci. 2010, 123, 1050–1059. [Google Scholar] [CrossRef]
- Marchant, R.; Smith, D.G. Bud Formation in Saccharomyces Cerevisiae and a Comparison with the Mechanism of Cell Division in Other Yeasts. J. Gen. Microbiol. 1968, 53, 163–169. [Google Scholar] [CrossRef]
- Lima, P.G.; Souza, P.F.N.; Freitas, C.D.T.; Oliveira, J.T.A.; Dias, L.P.; Neto, J.X.S.; Vasconcelos, I.M.; Lopes, J.L.S.; Sousa, D.O.B. Anticandidal Activity of Synthetic Peptides: Mechanism of Action Revealed by Scanning Electron and Fluorescence Microscopies and Synergism Effect with Nystatin. J. Pept. Sci. 2020, 1–13. [Google Scholar] [CrossRef]
- Gandhi, M.; Goode, B.L. Coronin: The Double-Edged Sword of Actin Dynamics. Subcell. Biochem. 2008, 48, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Makhov, A.M.; Bear, J.E. F-Actin Binding Is Essential for Coronin 1B Function in Vivo. J. Cell Sci. 2007, 120, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Marshall, T.W.; Uetrecht, A.C.; Schafer, D.A.; Bear, J.E. Coronin 1B Coordinates Arp2/3 Complex and Cofilin Activities at the Leading Edge. Cell 2007, 128, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, D.W.; Neupert, W. Import of Cytochrome c into Mitochondria: Reduction of Heme, Mediated by NADH and Flavin Nucleotides, Is Obligatory for Its Covalent Linkage to Apocytochrome C. Proc. Natl. Acad. Sci. U. S. A. 1989, 86, 4340–4344. [Google Scholar] [CrossRef] [PubMed]
- Dumont, M.E.; Cardillo, T.S.; Hayes, M.K.; Sherman, F. Role of Cytochrome c Heme Lyase in Mitochondrial Import and Accumulation of Cytochrome c in Saccharomyces Cerevisiae. Mol. Cell. Biol. 1991, 11, 5487–5496. [Google Scholar] [CrossRef]
- Dumont, M.E. Mitochondrial Import of Cytochrome C. Adv. Mol. Cell Biol. 1996, 17, 103–126. [Google Scholar] [CrossRef]
- NICHOLSON, D.W.; KÖHLER, H.; NEUPERT, W. Import of Cytochrome c into Mitochondria. Cytochrome c Heme Lyase. Eur. J. Biochem. 1987, 164, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Plapp, B. V.; Green, D.W.; Sun, H.W.; Park, D.H.; Kim, K. Substrate Specificity of Alcohol Dehydrogenases. Adv. Exp. Med. Biol. 1993, 328, 391–400. [Google Scholar] [CrossRef]
- Reid, M.F.; Fewson, C.A. Molecular Characterization of Microbial Alcohol Dehydrogenases. Crit. Rev. Microbiol. 1994, 20, 13–56. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Chung, K.R.; Liu, X.; Li, H. Aaprb1, a Subtilsin-like Protease, Required for Autophagy and Virulence of the Tangerine Pathotype of Alternaria Alternata. Microbiol. Res. 2020, 240, 126537. [Google Scholar] [CrossRef] [PubMed]
- Bryant, M.K.; Schardl, C.L.; Hesse, U.; Scott, B. Evolution of a Subtilisin-like Protease Gene Family in the Grass Endophytic Fungus Epichlo Festucae. BMC Evol. Biol. 2009, 9, 1–13. [Google Scholar] [CrossRef]
- Souza, P.F.N.; vanTilburg, M.F.; Mesquita, F.P.; Amaral, J.L.; Lima, L.B.; Montenegro, R.C.; Lopes, F.E.S.; Martins, R.X.; Vieira, L.; Farias, D.F.; et al. Neutralizing Effect of Synthetic Peptides toward SARS-CoV-2. ACS Omega 2022, 7, 16222–16234. [Google Scholar] [CrossRef]
- Dias, L.P.; Souza, P.F.N.; Oliveira, J.T.A.; Vasconcelos, I.M.; Araújo, N.M.S.; Tilburg, M.F. V; Guedes, M.I.F.; Carneiro, R.F.; Lopes, J.L.S.; Sousa, D.O.B. RcAlb-PepII, a Synthetic Small Peptide Bioinspired in the 2S Albumin from the Seed Cake of Ricinus Communis, Is a Potent Antimicrobial Agent against Klebsiella Pneumoniae and Candida Parapsilosis. Biochim. Biophys. Acta - Biomembr. 2020, 1862, 183092. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, J.W.; Cha, Y.; Kim, C. A Quantitative Nitroblue Tetrazolium Assay for Determining Intracellular Superoxide Anion Production in Phagocytic Cells. J. Immunoass. Immunochem. 2006, 27, 31–44. [Google Scholar] [CrossRef]
- MM, B. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Silva, F.D.A.; Carvalho, F.E.L.; Silveira, J.A.G.; Vasconcelos, I.M.; Oliveira, J.T.A. Photosynthetic and Biochemical Mechanisms of an EMS-Mutagenized Cowpea Associated with Its Resistance to Cowpea Severe Mosaic Virus. Plant Cell Rep. 2017, 36, 219–234. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Vasconcelos, I.M.; Silva, F.D.A.; Moreno, F.B.; Monteiro-Moreira, A.C.O.; Alencar, L.M.R.; Abreu, A.S.G.; Sousa, J.S.; Oliveira, J.T.A. A 2S Albumin from the Seed Cake of Ricinus Communis Inhibits Trypsin and Has Strong Antibacterial Activity against Human Pathogenic Bacteria. J. Nat. Prod. 2016, 79. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Detection of anion superoxide and hydrogen peroxide in C. neoformans cells. Light and fluorescence microscopies analysis to detect, respectively, the conversion of NBT into formazan (blue or cyan dots – black arrows) and the accumulation of hydrogen peroxide in C. neoformans cells treated with Mo-CBP3-PepII at MIC50 concentration. White bars indicate 100 µm.
Figure 1.
Detection of anion superoxide and hydrogen peroxide in C. neoformans cells. Light and fluorescence microscopies analysis to detect, respectively, the conversion of NBT into formazan (blue or cyan dots – black arrows) and the accumulation of hydrogen peroxide in C. neoformans cells treated with Mo-CBP3-PepII at MIC50 concentration. White bars indicate 100 µm.
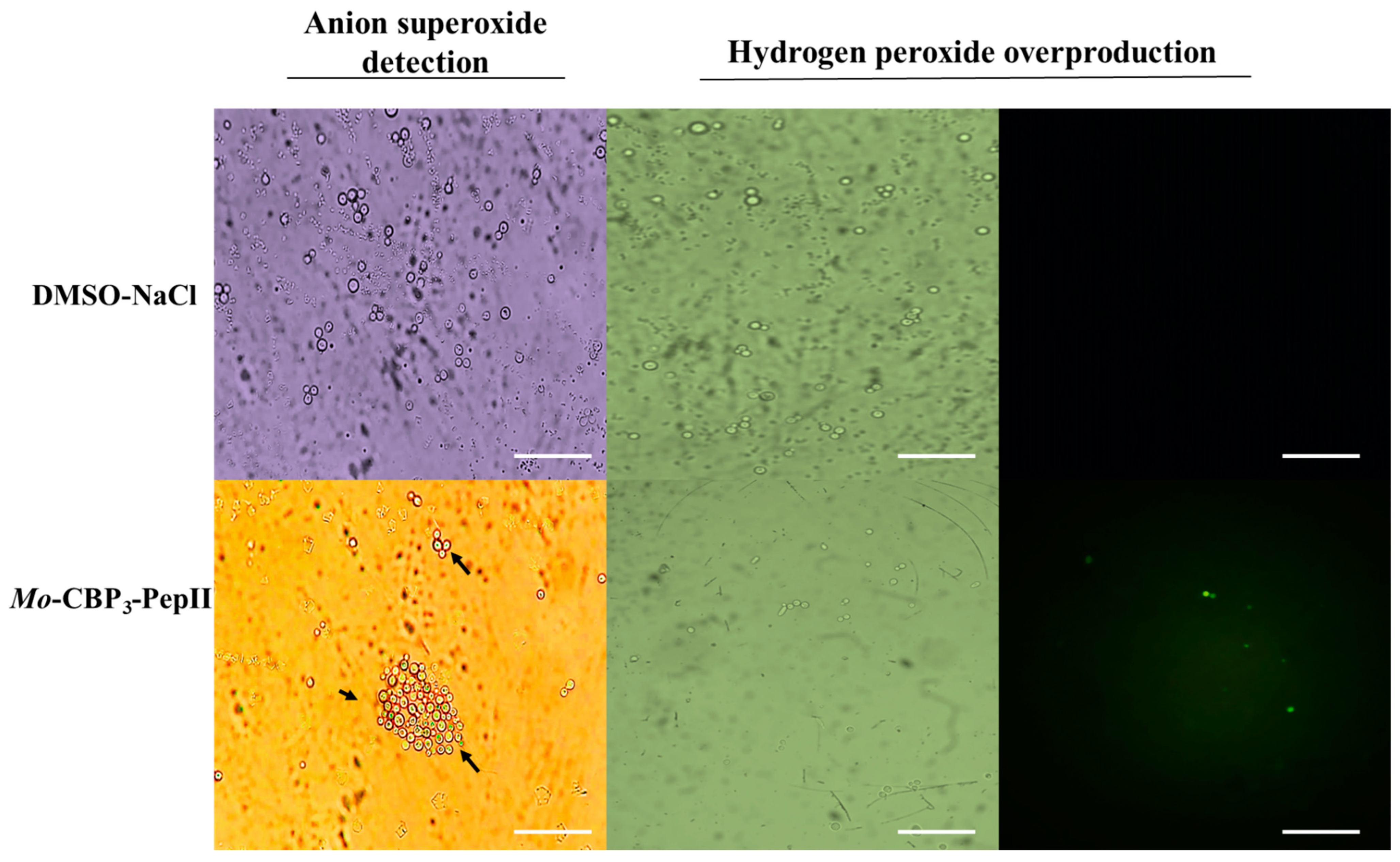
Figure 2.
The activity of enzymes involved in ROS in C. neoformans cells after treatment with Mo-CBP3-PepII. The activity of enzymes was tested in C. neoformans cells treated (Mo-CBP3-PepII) and non-treated (DMSO) with the synthetic peptide. The different lowercase letters indicate statistical significance at p > 0.05.
Figure 2.
The activity of enzymes involved in ROS in C. neoformans cells after treatment with Mo-CBP3-PepII. The activity of enzymes was tested in C. neoformans cells treated (Mo-CBP3-PepII) and non-treated (DMSO) with the synthetic peptide. The different lowercase letters indicate statistical significance at p > 0.05.
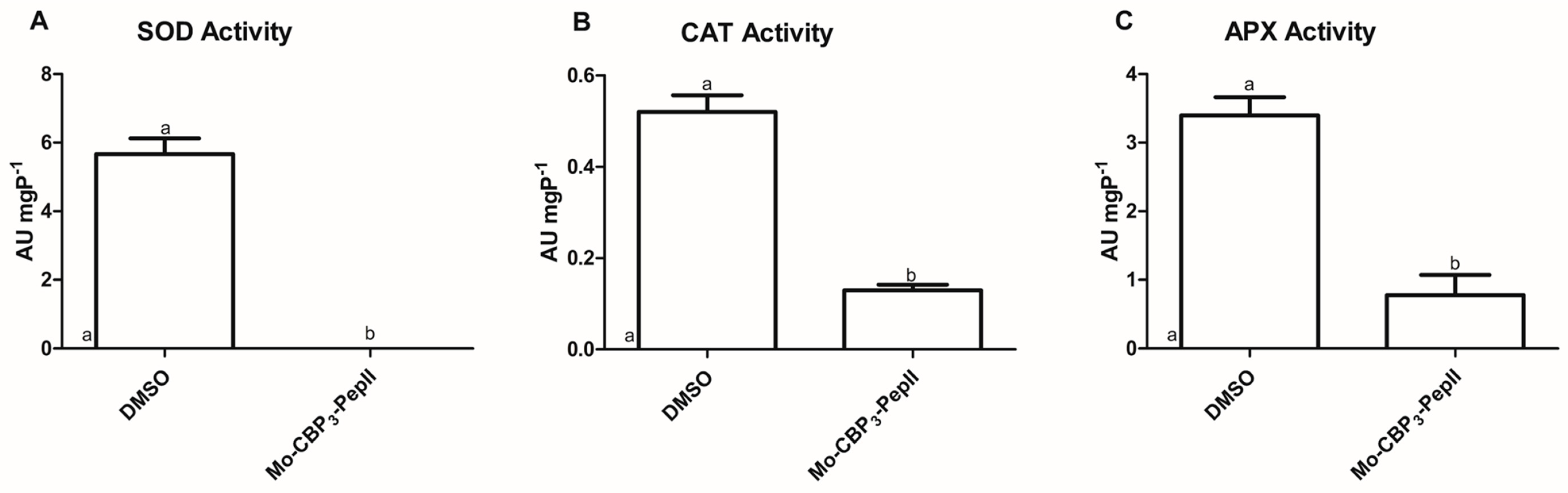
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



