Submitted:
26 January 2023
Posted:
27 January 2023
You are already at the latest version
Abstract
The article presents the results of a morpho-physiological assessment of rice samples by the strength of growth, as well as an assessment of the degree of development of the conducting system of the flag leaves of rice plants. The resistance of varieties to flooding is very relevant in the fight against weeds due to the deep layer of water that weeds cannot overcome. Currently, there are no zoned varieties in Russia that would meet these requirements. Therefore, the problem of creating such rice varieties is urgent, as it will reduce production costs, reduce grain losses during harvesting, improve the quality of the products obtained, and also reduce the pesticide load on the ecosystem. As a result of research, a number of studied samples revealed the potential to quickly lengthen the first leaves, overcome a large layer of water and accumulate vegetative mass. As a result of the evaluation of the conducting system of the flag leaf of rice plants, it was found that in plants with the flood resistance gene Sub1A, which stops growth under water, the conductive beams were smaller in comparison with other fast-growing samples with the AG, Sk genes. Consequently, fast-growing samples have bigger sizes of the conducting system as a resistance mechanism.
Keywords:
rice
; samples
; resistance to flooding
; conductive system
; power of growth
1. Introduction
Rice resistance to flooding is of great importance in the world rice growing, since the peculiarities of the natural conditions of its cultivation in the tropics and subtropics are associated with complete flooding of rice crops for a short period of several days to two weeks, due to heavy precipitation.
Flooding is one of the main environmental stress factors affecting many artificial and natural ecosystems around the world. An increase in the frequency and duration of heavy rains due to climate change negatively affects the growth and development of plants, which ultimately leads to plant death if it persists for several days. As a result, crops of unstable varieties are dying, a loss of yield occurs [1].
Most agricultural crops, including rice, being a semi-aquatic plant, suffer greatly from lack of oxygen during flooding, which leads to annual crop loss. At the same time, the main molecular features that determine the resistance of rice to anoxia differ depending on the stage of development [2].
During vegetative growth, rice uses two different strategies to overcome long-term flooding resistance to flooding. The genetic variability of the plant response to flooding includes various schemes: 1) rest, which allows withstanding a long time under water, 2) a strategy of rapid stem elongation with changes in plant structure and metabolism [3].
The first one is the transition of plants to hibernation under water. The dormant state is observed in rice varieties with the Sub1A-1 locus, which limit the mobilization of starch and, thus, produce less ethanol and other fermentation products, and also affect the processes associated with aging [4]. The reaction to oxygen deficiency in rice coleoptiles at the gene level includes the expression of genes encoding proteins involved in fermentation, cell wall expansion, carbohydrate and lipid metabolism, as well as the induction of expression of ethylene response factors [5].
The study of the mechanism of flood survival in rice has provided important information about the development, physiology, and molecular strategies of survival during immersion in water. Significant progress in the breeding of flood-resistant rice varieties has been achieved over the past decade after the successful identification and mapping of the quantitative trait loci (QTL) of flood resistance, designated as Submergence 1 (Sub1) from the old FR13A variety. Using return scratching with markers, Sub1 was included in many elite varieties in a short time and with high accuracy compared to traditional methods of selection [6].
The high energy and speed of the initial growth of plants is a second strategy for the survival of plants in conditions of prolonged flooding. It consists in the rapid growth and elongation of the coleoptile during development under anaerobic conditions, called the “tube effect”, which is controlled by genes of tolerance to immersion (Snorkel 1, 2).
Most rice varieties are unable to germinate and reach the surface of the water when fully submerged. Therefore, resistance to anaerobic conditions during germination is an important feature for rice cultivation by direct sowing in irrigated ecosystems [7]. Therefore, it is necessary to study the polymorphism of various forms of rice in terms of resistance to prolonged deep water levels.
To overcome prolonged flooding, rice uses two strategies. The first strategy is the transition of plants to hibernation under water. High energy and the rate of initial plant growth is the second strategy for plant survival in conditions of prolonged flooding. Therefore, it is necessary to study the polymorphism of various forms of rice in terms of resistance to prolonged deep water levels.
Early germination energy is an important goal of rice breeding, especially in direct sowing. Chinese scientists have identified 3 quantitative trait loci (QTL) influencing this feature. This study provides a theoretical basis and genetic resources for the selection of rice for direct sowing [8].
In India, lines with strong early growth energy and rapid uniform germination combined with good yields were selected for the purpose of genetic improvement of rice varieties. This is done to achieve uniformity of the plant population in the field due to effective and rapid germination of seeds from the depth of the soil and early emergence of seedlings, which helps to reduce the number of weeds [9].
The internodes of deep-water rice lengthen in response to an increase in the water level so that its leaves remain above the water surface and avoid oxygen depletion. This elongation is stimulated by ethylene-regulated genes, Snorkel1 and Snorkel2. On the contrary, when a flash flood occurs at the germination stage, the submerged rice carrying the Sub1A gene remains stunted and lies in the water for several weeks to avoid energy consumption associated with the elongation of the plant, and then resumes its growth using its stored energy after the water recedes. However, deep-water and flood-resistant rice has the opposite reaction to flooding; namely, to rise above the surface by lengthening the stem or to remain in a stunted state underwater until the flood recedes. The genes Snorkel 1 (SK1) and Snorkel 2 (SK2), identified on chromosome 12, determine the sensitivity to ethylene and regulate the elongation of the internodes of deep-sea rice in response to immersion. Normal cultivated rice has no SK genes [10].
The resistance of varieties to flooding is very relevant in Russia when fighting weeds due to the deep layer of water that weeds cannot overcome and die [11]. This leads to such positive effects as the uselessness of herbicides application, reducing the cost of producing a unit of production, improving the quality of the products that can be used in the production of products for children and dietary nutrition, and the absence of environmental damage since there is no need to treat crops with herbicides [12].
Tolerance to flooding is a genetic property that manifests itself under stress due to the involvement of many uncontrolled environmental factors. In the presence of a large number of genetic resources of rice, promising genotypes with desirable traits can be selected from them [13].
For large-scale screening of rice germ plasm with resistance to stress caused by immersion, Japanese scientists have developed two new methods of analysis, which were based mainly on the strength of seedlings, estimated by the ability of rapid elongation of shoots under immersion conditions. Their results show that the growth force of seedlings serves as a mechanism for rice resistance to flooding. The relationship between the germination energy, based on the rapid elongation of the shoot, and resistance to immersion in water, due to recovery from stress caused by immersion, is shown [14].
The main element of the conducting system are conducting beams, the development of which affects the adaptation to growing conditions and plant productivity. The parameters of conducting beams and their number are largely influenced by the specific and varietal characteristics of rice.
Conducting beams form the basis of leaf venation, provide the mesophyll with salt solutions and water, and also carry out the outflow of plastic substances obtained as a result of assimilation. Venation is especially pronounced on the underside of the leaf. The veins have approximately the same size, they are connected either in the upper part of the leaf or on both sides of the plate.
The number and size of conducting beams affects the formation of plant productivity and its resistance to the negative influence of the environment. The conductive system of leaves supplies plants with water and distributes assimilates, which are stable organic compounds that are end products in the process of photosynthetic fixation and reduction of carbon dioxide in plants [15].
Currently, there are no zoned varieties in Russia that would meet these requirements. Therefore, the problem of creating such rice varieties is urgent, as it will reduce production costs, reduce grain losses during harvesting, improve the quality of the products obtained, and also reduce the pesticide load on the ecosystem.
Thus, the research of rice resistance to flooding and the development of methodology in this area is an important fundamental problem for this crop.
2. Materials and Methods
The object of the study was 180 rice samples obtained as a result of hybridization of Russian varieties with Asian donors of genes for resistance to prolonged flooding with water in the Laboratory of Rice Breeding and Seed Production of the Agrarian Scientific Center “Donskoy”. The locus of resistance to flooding Sub 1A was transferred from donors Br-11, CR-1009, Inbara-3, TDK-1, Khan Dan. The donors of the Snorkel 1, 2 genes were the varieties Khao Hlan On, Kharsu 80A, Mazhan Red. Local varieties Kuboyar and Kontakt were used as controls.
Laboratory studies were carried out using several physiological methods developed by us.
1. Assessment of the strength of growth in aerobic conditions. The strength of seed growth of 180 samples was evaluated by germinating seeds under controlled aerobic conditions in Petri dishes on wet filter paper, at a temperature of 28 °C in the dark, in a double repetition of 5 seeds each. After 10 days, germination was determined and the length of sprouts and roots was measured. This experience allows revealing the potential of varieties to quickly accumulate vegetative mass.
2. Evaluation of rice on the ability to anaerobic germination. The seeds were placed in a glass tube (2.5 cm in diameter and 15 cm in height), filled with distilled water 10 cm deep and incubated at a temperature of 28 ° C without changing the water. On the 5th, 7th, 9th, 12th, and 14th days, the length of the sprout was measured to identify the growth dynamics.
3. Assessment of growth in glass cylinders. Cylinders 42 cm high were filled with field soil by 5 cm, filled with tap water and 10 seeds were lowered to the surface twice (Figure 1). Cultivation was carried out in the light in room conditions at a temperature of 25-30 °C. Measurements of the length of rice sprouts were carried out on the 3rd, 8th and 13th day of growth to identify its dynamics.
The evaluation of conducting beams of rice plant leaves was carried out by microscopy of a cross-section of a flag leaf. Sections of ten flag leaves of each sample were studied in threefold repetition. The “Shoptop SX40” was used.
3. Results and Discussion
To overcome the deep water layer, intensive growing samples with the maximum length of sprouts are of great interest. If plants cannot overcome a thick layer of water, they will die from lack of oxygen and nutrients [10].
The germination of seeds during aerobic germination in Petri dishes on the 10th day from laying for germination varied from 60 to 100%. Out of 180 samples, 156 germinated all the seeds, 20 - 80% and 4 - 60%. The analysis showed that the length of the sprout in the samples ranged from 2.4 to 7.4 cm. Samples with a small sprout length – 3...4 cm prevailed. The maximum values of the average sprout length of 7.4 cm were at lines 1191 (Inbara-3 × Contact) × Khao Hlan On × Kuboyar and 1544 R6 (Kharsu 80A) × Contact. Their parent forms Khao Hlan On and Kharsu 80A are very fast growing varieties.
The average length of the spine varied from 2.2 to 15.3 cm. Samples with a sign value of 4...8 cm prevailed. The greatest length of the spine was noted in the lines 1191 (Inbara-3 × Contact) × (Khao Hlan On × Kuboyar) (15.3 cm) and 2032 KD Sub-1 D-149 × Comandor (13.8 cm). Comparison of the length of the sprout and the root in rice samples showed that the correlation between them is weak (r=0.10 ± 0.05).
Flood tolerance or anaerobic tolerance is characterized by the ability of plants to recover after flooding at different stages of development. Restoration involves the regeneration of plants damaged by sudden flooding. The grounds for tolerance to flooding are associated with a high initial carbohydrate content, high levels of chlorophyll and minimal growth under immersion conditions. High energy and speed of initial plant growth is one of the main strategies for plant survival in conditions of prolonged flooding. Therefore, an experiment was conducted to study the ability of rice to overcome a large layer of water.
In the process of studying promising breeding samples, a morpho-physiological assessment of this material for resistance to flooding was carried out.
One of the strategies for the survival of rice plants during prolonged flooding is to stop growth under water, fall into a kind of hibernation, and when the water layer decreases, they continue their growth and development [16].
Therefore, an experiment was conducted on germination of seeds in test tubes from under a 10 cm water layer (Figure 2a). As a result of the research, it was found that with anaerobic germination after 5 days, most of the studied varieties and lines (71%) had an average sprout length from 0.1 to 2.0 cm, only 6% had a maximum average sprout length from 3.0 to 4.0 cm (Figure 2b).
The largest sprout length (3.0...4.0 cm) was in the lines 1665 (Atlant × Neutron), 1176 (Inbara-3 (Sub-1) × Innovator × Contact) and 1337 ((BR-11 × Innovator) × Kuban). Most of the lines had no roots. Only three lines had a sprouted root, its length was 2...3 mm.
With anoxia, the rice seedling lengthens the internode and firstly forms a sprout longer than with aerobic germination, which causes the insignificant development or absence of a root in most rice plants. During the study, the seedlings of 19 rice lines in test tubes grew very slowly, from 0.1 to 0.5 cm, which suggested the presence of the Sub 1A gene in them. According to the data of changes in the value of leaf growth in the samples, it can be assumed that the Sub1A gene is present or absent.
On the 7th day of growth in test tubes, the length of the sprout in the samples varied from 0.9 to 4.8 cm, on average – 2.7 cm (Figure 3a).
The smallest increase or resting state in the studied samples indicates the possibility of the presence of the Sub1A gene in them. The smallest increase in the length of the sprout (0.2...0.5 cm) was observed in samples 1177 [(Inbara-3 × Innovator) × Contact], 3137 (Inbara-3 × Stepnyak), 2407 (Bacchus × Inbara-3), 1337 [BR-11 × Innovator) × Kuban 3], 1176 [(Inbara-3 × Innovator) × Contact], 766 (Inbara-3 × Contact). Most of them literally have the Inbara-3 variety, which carries the Sub1A locus in its genotype.
On the 9th day, the length of the sprout varied from 1.8 to 7.0 cm, on average 3.8 cm (Fig. 3b). The smallest sprout length (2.1...2.2 cm) was in samples 766 (Inbara-3 × Contact), 3137 (Inbara-3 × Steppe), 2407 (Bacchus × Inbara-3).
Another 25 samples had a small sprout length of up to 3 cm . Many forms were light green, they were not resistant to this kind of stress and did not survive. Some of these plants were removed from the framework of the experiment, so that they would not die, the water level was lowered to 1 cm. This had a positive impact on their further growth and development. Aerobic processes were activated in their cells, the synthesis of auxins and gibberellins was normalized, as a result of which the plants continued their normal growth.
On the 12th day of the experiment, the length of the seedling in the samples varied from 3.4 to 11.5 cm, on average 6.3 cm (Figure 4a).
Samples with a sprout length of 3...7 cm prevailed, there were 64% of them. About 6% of the samples grew rapidly and overcame the layer of water in the test tube, going out into the air. These are such lines as 2035 (KD Sub-1 D-149 × Commander) and 350 (Inbara 3 × Innovator) × Contact. Their length was 11.4 cm. However, weakly growing forms are also of interest. In particular, sample 766 (Inbara-3 × Contact) grew only to 2.2 cm, adding 1 mm in 3 days. This indicates the possible presence of the Sub1A gene in him.
After 14 days, the average length of the seedlings was 8.6 cm, varying from 3.0 to 16.5 cm (Figure 4b). The tallest samples were 1191 [(Inbara-3 × Contact) × (Khao Hlan On × Kuboyar)] – 16.5 cm, and 1542 (Kharsu 80A × Contact) – 16.3 cm, and low-growing again No. 766 – 3 cm. Apparently, samples 1191 and 1542 were given the energetic growth gene (Snorkel) from the Khao Hlan On variety, and sample 766 received the Sub1A gene from the Inbara-3 variety.
The results of studying plants in cylinders showed significant differences between the growth rates under water (Figure 5).
After 3 days from sowing, the length of the sprouts varied from 0.1 to 1.7 cm. The maximum sprout length during this period (1.5-1.7 cm) was observed in samples whose parent was mainly the Kharsu 80A variety: 1134 (Kharsu 80A × Contact), 1537 (Kharsu 80A × Contact), 1533 (Kharsu 80A × Contact), 1032 (Kuboyar × Kharsu 80A), 1063 (Kuboyar × Mazhan Red), minimum (0.1-0.2 cm) – in samples: 995 (Contact × Khao Hlan On), 996 (Contact × Khao Hlan On) (Table 1).
On the 8th day, the differences in plant length of the samples became more significant: from 2.0 to 12.0 cm (on average 6.42 cm). The largest values of length (11.5-12.0 cm) during this period were shown by samples from another hybrid combination: Contact × Khao Hlan On. These are samples numbered 985, 998 and 1007. The minimum length (2 cm) at this stage was for samples 1001 (Contact × Khao Lan On) and 1036 (Kuboyar × Kharsu 80A).
On the 13th day, the average length of the stems ranged from 8.4 to 47.0 cm (on average 21.1 cm). The largest length of seedlings (more than 30 cm) was in the samples from the hybrid combination Contact × Khao Hlan On: 1006 (47.0 cm), 998 (45.3 cm), 997 (40.3 cm), 991 (39.7 cm), 985 (39.7 cm), etc., the smallest, in the samples of combinations Kuboyar × Mazhan Red: 1065 (8.4 cm); Kuboyar × Kharsu 80A: 1033 (8.8 cm), 1038 (8.6 cm), and 1044 (8.6 cm).
Vigorously growing plants rose above the water layer and the upper edge of the cylinder and began to supply oxygen through the aerenchyma to the root system. The undersized plants did not grow up to the middle of the cylinder.
The dynamics of the growth of rice plants with maximum and minimum values on the 13th day in cylinders under water is shown in Figure 6.
The graph shows that on the third day the differences in length between these samples were imperceptible (0.30-0.75 cm), on the 8th day they became more noticeable (2.58-12.00 cm), and on the 13th day, they became significant (8.55-47.00 cm). At the same time, samples with minimum values of 1033, 1038, and 1044 had almost identical curve configurations, and with maximum values they differed slightly. The curves of samples 998 and 1006 had similar configurations, and the 997 line was located slightly lower. The average values of this trait for the entire group of studied samples on the third day were greater than those of these six samples (0.86 cm), on the eighth day they were in the middle (6.42 cm), and on the thirteenth day they were closer to smaller values (21.12 cm).
This indicates a rightward asymmetry in the distribution of plant length during this period; shorter plants prevailed.
This is clearly demonstrated by the histogram of the distribution of the studied rice samples along the length of plants with leaves on the 13th day after sowing (Figure 7).
The histogram has 3 peaks, one in the 10-15 cm class, the second one in 35-40 cm, the third one in 45-50 cm. Most of the samples (75%) had a small plant height: up to 30 cm. A fourth part of the samples exceeded this value. Two samples (998 and 1006), noted earlier, stood out in particular.
The energy index of the initial growth of rice plants in laboratory conditions, combined with an assessment of the conducting system of flag leaves, allowed identifying differences in the test samples in response to this abiotic stress, which made it possible to identify samples resistant to flooding.
The main structural and functional component of the leaves are conductive beams of rice plant leaves, which provide trophic, transpiration, regulatory and mechanical functions. The shell of the conductive beam of rice performs the function of assimilation of nutrients that are necessary both for central metabolism and for reaction to biotic and abiotic stresses.
A small number of conducting beams and the small size of their area delay the movement and accumulation of assimilates in plant leaves, the functions of respiration, transpiration, photosynthesis decrease and growth slows down. In conditions of prolonged flooding due to the smaller diameter of the conductive beams and their total area, the plant resists the influence of this stress. This is primarily due to the ability of the conductive system of stable samples to be rebuilt under the influence of external factors without disturbing the functions of plant growth, respiration, photosynthesis and transpiration.
Leaves contain several types of cells underlying their functional specialization. Knowledge about the structure of these cells, in the presence of certain genes in them, is fragmentary. To understand the structure of the conducting system of the leaves of rice plants grown in flood conditions and having the resistance gene Sub1, microscopy of the cross section of the flag leaf was used (Figure 8 and Figure 9).
The resistance of rice samples to prolonged flooding made it necessary to find out which physiological systems affect the ability of plants to adapt and withstand stress conditions. Studies have been conducted to assess the degree of development of the conducting system of the flag leaf of rice plants grown in flood conditions.
It was found that of the seven studied samples, two with the Sub1A resistance to prolonged flooding gene: 4526 INBARA-3 × Innovator (Sub-1A, AG-1) and 4617 INBARA-3 × Contact (Sub-1) differ in the smallest beam diameters, the area of one beam and the total area of conducting beams: 153.2 and 165.6 microns, 18424 and 21527 microns, 0.92 and 1.46 mm2, respectively (Table 2).
Samples with various combinations of vigorous growth genes Sk, AG (4641, 4642, 4732, 4758, 4773) had higher values of the diameter and area of the beams: from 25717 to 31904 mcm2.
Thus, as a result of the conducted research, it was found that adaptive varieties are anatomically equipped to absorb less water in flood conditions and maintain the normal functioning of all plant systems.
4. Conclusion
1. Thus, as a result of morpho-physiological assessment of rice samples, more vigorously growing rice samples with the greatest potential for growth and development were identified. Rapid plant growth is an adaptive reaction of rice to oxygen production, which is necessary for survival during prolonged immersion in water. A number of the studied samples revealed the potential to grow rapidly, overcome a large layer of water and accumulate vegetative mass.
2. Evaluation of 180 rice samples by the intensity of growth of seedlings under aerobic conditions on the 10th day showed a significant variation in the length of the sprout: from 2.4 to 7.4 cm and the root: from 2.2 to 15.3 cm. The maximum values of the average length of the sprout and root were at line 1191 [(Inbara-3 × Contact) x (Khao Hlan On × Kuboyar)].
3. As a result of anaerobic germination of rice seeds in test tubes, it was established that after 5 days, most of the studied varieties and lines (71%) had an average sprout length from 0.1 to 2.0 cm, only 6% had a maximum average sprout length from 3.0 to 4.0 cm. Samples with minimal stem growth (0.2...0.5 cm) were isolated under anaerobic conditions: 1177, 3137, 2407, 1337, 1176, 766, which had the Inbara-3 variety in their pedigree. The largest sprout length (3.0...4.0 cm) was in the lines 1665 (Atlant × Neutron), 1176 ((Inbara-3 × Innovator) × Contact) and 1337 ((BR-11 × Nova-tor) × Kuban). 14 days after the seeds were filled with water, the average length of the seedlings was 8.6 cm, varying from 3.0 to 16.5 cm. The tallest samples were 1191 [(Inbara-3 × Contact) x (Khao Hlan On × Kuboyar)] – 16.5 cm, and 1542 (Kharsu 80A × Contact) – 16.3 cm, and stunted No. 766 – 3.0 cm.
4. The results of germination in cylinders with a height of 42 cm showed significant differences between plants in terms of growth rate under water. After 3 days from sowing the seeds, the length of the shoots varied from 0 to 8 mm, on the 8th day, from 2.0 to 11.8 cm, on the 13th day, from 18.0 to 47.0 cm. The greatest length of seedlings was in samples 1006 (47.0 cm) and 998 (45.3 cm), which can be grown in the field without herbicides using the technology of obtaining seedlings from under the water layer or used as donors of high growth energy.
5. As a result of the evaluation of the conducting system of the flag leaf of rice plants, it was found that in plants with the flood resistance gene Sub1A, which stops growth under water, the conducting bundles were smaller in comparison with other fast-growing samples with the AG, Sk genes. Consequently, fast-growing samples have large sizes of the conducting system as a resistance mechanism.
Author Contributions
Golubova V.A. – performing laboratory experiments, data analysis, writing the text of the article; Kostylev P. I. – general scientific guidance, setting goals and objectives, analysis of published data, formation of research methodology and concept of the article, data analysis, preparation of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by grant No. 22-26-00246 of the Russian Science Foundation (RSF).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kawano, N.; Ito, O.; Sakagami, J.-I. Morphological and physiological responses of rice seedlings to complete submergence (flash flooding). Annals of Botany. 2009, 103, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu Rev Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nature Reviews Genetics. 2015, 1–15. [Google Scholar] [CrossRef]
- Fukao, T.; Yeung, E.; Bailey-Serres, J. The submergence tolerance regulator Sub1A mediates crosstalk between submergence and drought tolerance in rice. The Plant Cell. 2011, 23, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Narsai, R.; Edwards, J.M.; Roberts, T.H.; Whelan, J.; Joss, G.H.; Atwell, B.J. Mechanisms of growth and patterns of gene expression in oxygen-deprived rice coleoptiles. Plant J. 2015, 82, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Oladosu, Y.; Rafii, M.Y.; Arolu, F.; Chukwu, S.C.; Muhammad, I.; Kareem, I.; Salisu, M.A.; Arolu, I.W. Submergence Tolerance in Rice: Review of Mechanism, Breeding and Future. Sustainability. 2020, 12, 1632. [Google Scholar] [CrossRef]
- Septiningsih, E.M.; Ignacio, J.C.I.; Sendon, P.M.D.; Sanchez, D.L.; Ismail, A.M.; Mackill, D.J. QTL mapping and confirmation for tolerance of anaerobic conditions during germination derived from the rice landrace Ma-Zhan Red. Theor. Appl. Genet. 2013, 126, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Guo, Z.; Luo, L.; Gao, Q.; Xiao, W.; Wang, J.; Wang, H.; Chen, Z.; Guo, T. Identification of QTL and candidate genes involved in early seedling growth in rice via high-density genetic mapping and RNA-seq. The Crop Journal. 2021, 9, 360–371. [Google Scholar] [CrossRef]
- Singh, U.M.; Yadav, S.; Dixit, S.; Ramayya, P.J.; Devi, M.N.; Raman, K.A.; Kumar, A. QTL Hotspots for Early Vigor and Related Traits under Dry Direct-Seeded System in Rice (Oryza sativa L.). Frontiers in Plant Science. 2017. V. 8. Art. 286. P.1-14. [CrossRef]
- Nagai, K.; Kurokawa, Y.; Mori, Y.; Minami, A.; Reuscher, S.; Wu, J.; Matsumoto, T.; Ashikari, M. Snorkel genes relating to flood tolerance were pseudogenized in normal cultivated rice. Plants (Basel). 2022. V. 11(3). P. 376. [CrossRef]
- Panda, D.; Barik, J. Flooding Tolerance in Rice: Focus on Mechanisms and Approaches. Rice Science. 2021. V. 28. Is. 1, P. 43-57. [CrossRef]
- Barik, S.R.; Pandit, E.; Pradhan, S.K.; Mohanty, S.P.; Mohapatara, T. Genetic mapping of morpho-physiological traits involved during reproductive stage drought tolerance in rice. PLoS ONE. 2019. No. 14 (12). Article number e0214979. [CrossRef]
- Bin Rahman, A.N.M.R.; Zhang, J.H. Flood and drought tolerance in rice: Opposite but may coexist. Food Energy Security. 2016. No. 5 (2). P. 76-88. [CrossRef]
- Manangkil, O.E.; Vu, H.T.T.; Yoshida, S.; Mori, N.; Nakamura, C. A simple, rapid and reliable bioassay for evaluating seedling vigor under submergence in indica and japonica rice (Oryza sativa L.). Euphytica. 2008, 163, 267–274. [Google Scholar] [CrossRef]
- Sage, T.L.; Sage, R.F. The Functional Anatomy of Rice Leaves: Implications for Refixation of Photorespiratory CO2 and Efforts to Engineer C4 Photosynthesis into Rice. Plant and Cell Physiology 2009, 50, 756–772. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Yeung, E.; Bailey-Serres, J. The submergence tolerance regulator Sub1A mediates crosstalk between submergence and drought tolerance in rice. The Plant Cell. 2011, 23, 412–427. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Rice seed germination and plant growth in cylinders under water.

Figure 2.
Anaerobic germination of rice in test tubes and distribution of samples along the length of the sprout (cm) on the 5th day.
Figure 2.
Anaerobic germination of rice in test tubes and distribution of samples along the length of the sprout (cm) on the 5th day.
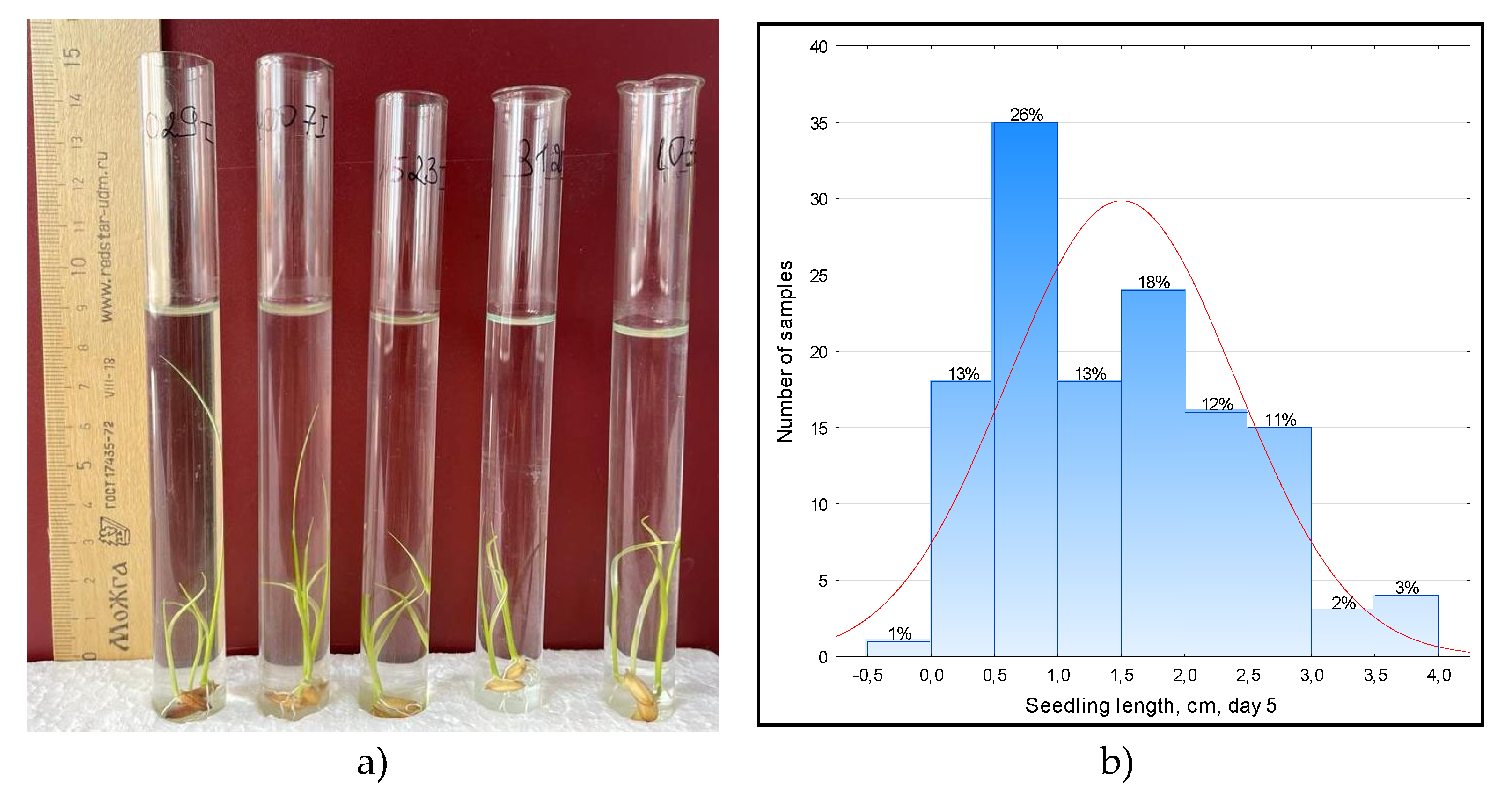
Figure 3.
Distribution of rice varieties and lines along the average length of the seedling during anaerobic germination: a) on the 7th day; b) on the 9th day.
Figure 3.
Distribution of rice varieties and lines along the average length of the seedling during anaerobic germination: a) on the 7th day; b) on the 9th day.
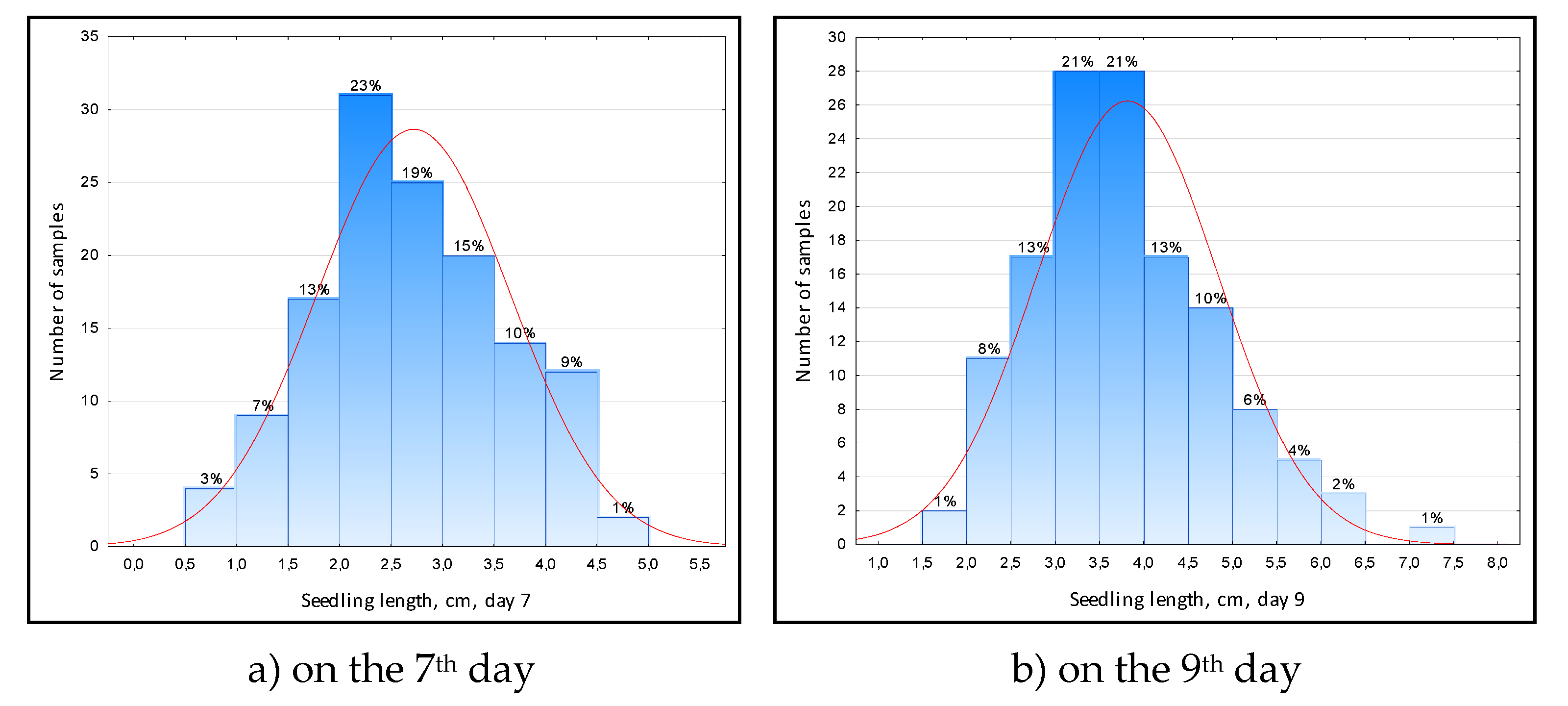
Figure 4.
Distribution of rice varieties and lines along the average length of the seedling during anaerobic germination: a) on the 12th day; b) on the 14th day.
Figure 4.
Distribution of rice varieties and lines along the average length of the seedling during anaerobic germination: a) on the 12th day; b) on the 14th day.
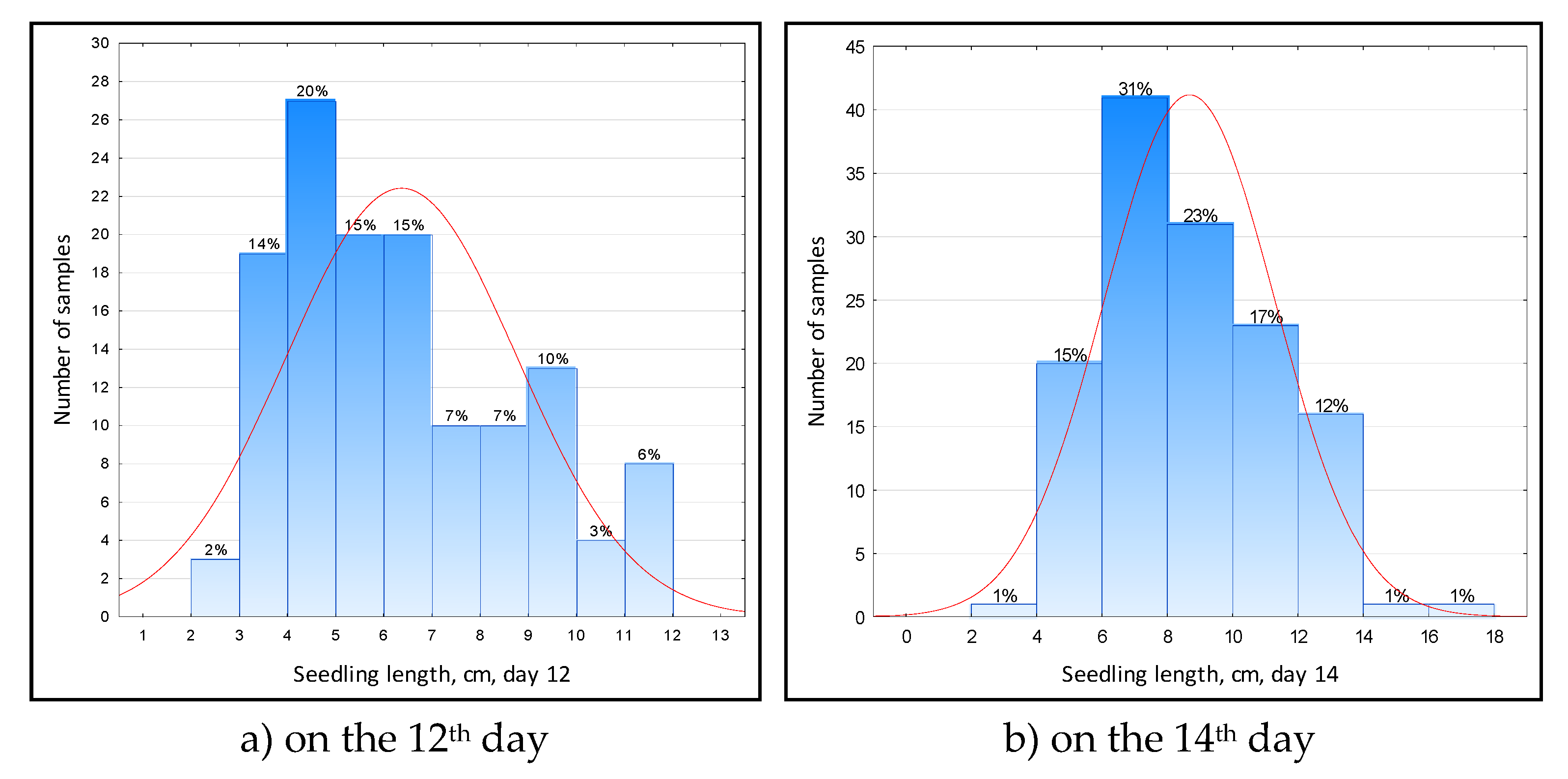
Figure 5.
Growth of rice plants in cylinders.

Figure 6.
Dynamics of rice plant growth in cylinders under water (maximum and minimum values on the 13th day).
Figure 6.
Dynamics of rice plant growth in cylinders under water (maximum and minimum values on the 13th day).
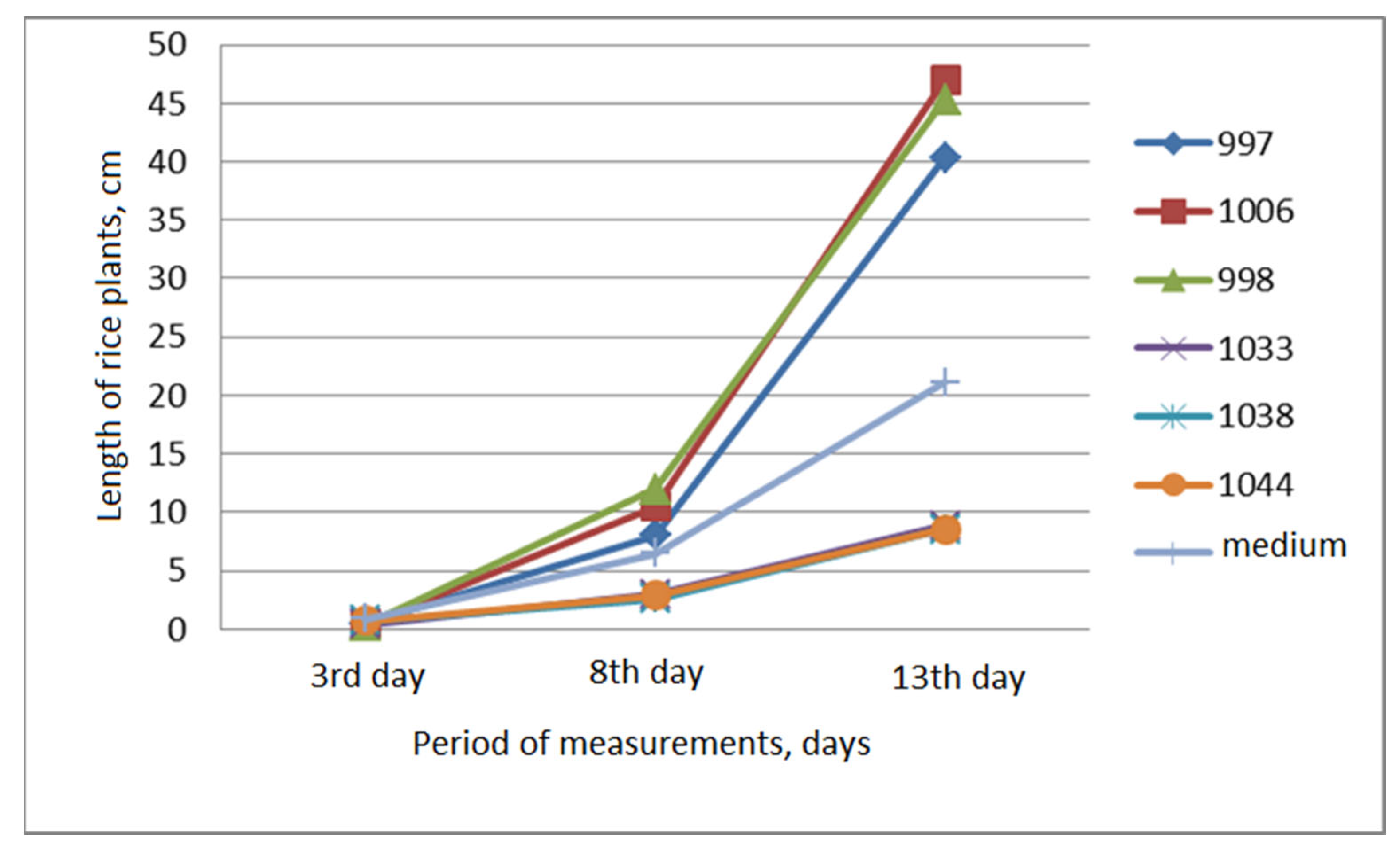
Figure 7.
Distribution of rice samples along the length of plants with leaves on the 13th day after sowing.
Figure 7.
Distribution of rice samples along the length of plants with leaves on the 13th day after sowing.
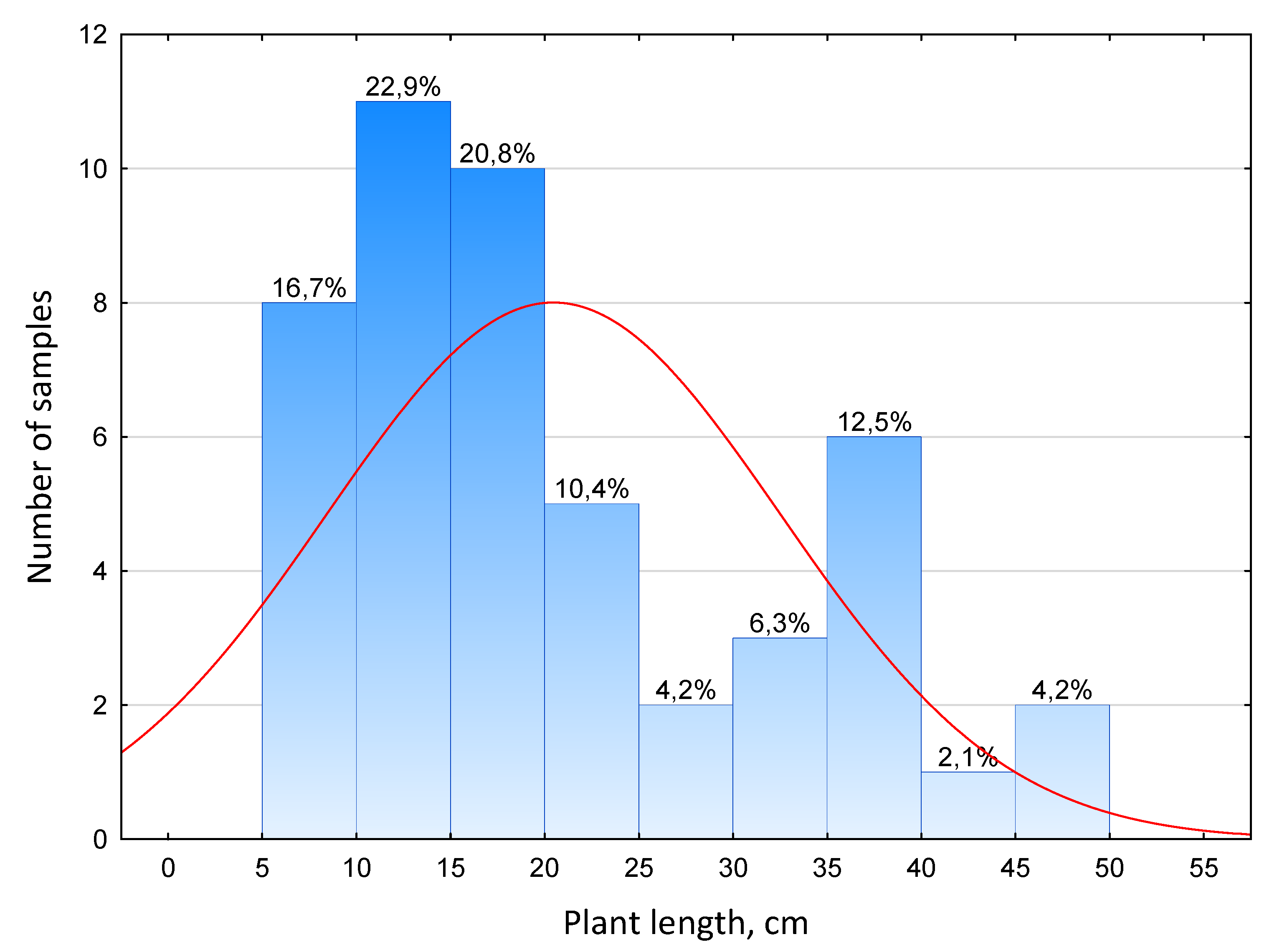
Figure 8.
Cross section of rice sample leaf 4526 Inbar-3 × Innovator (Sub1A, AG-1).
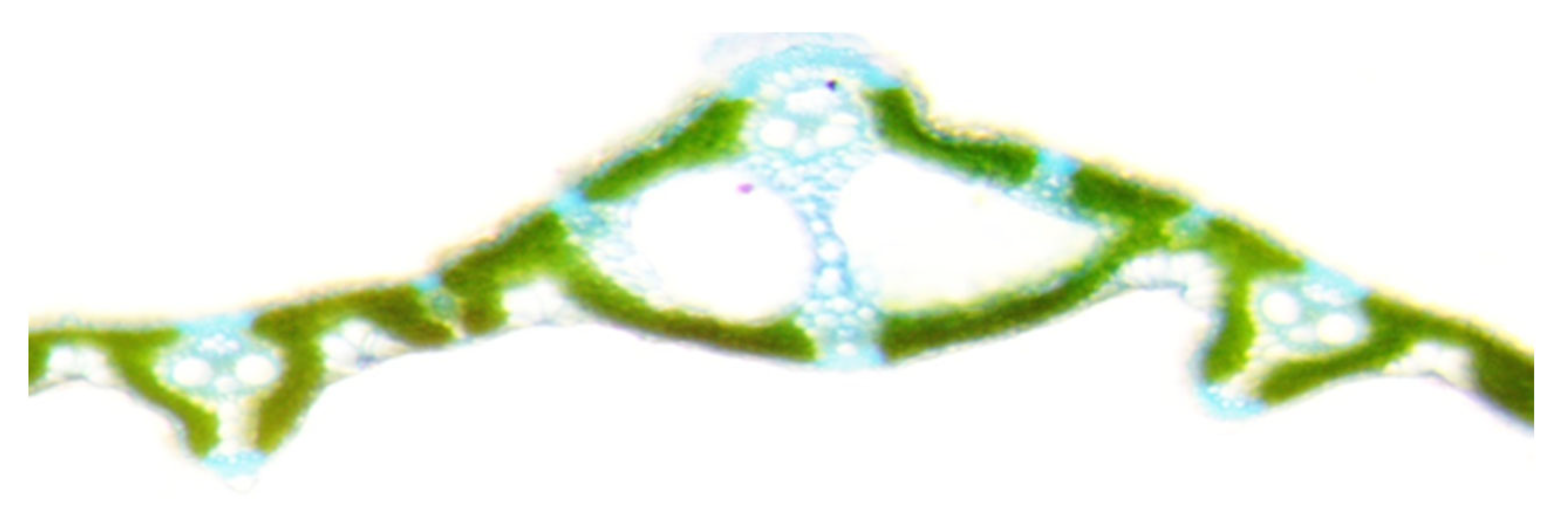
Figure 9.
Cross section of a rice sample leaf 4758 Contact × Khao Hlan On (Sk-2).
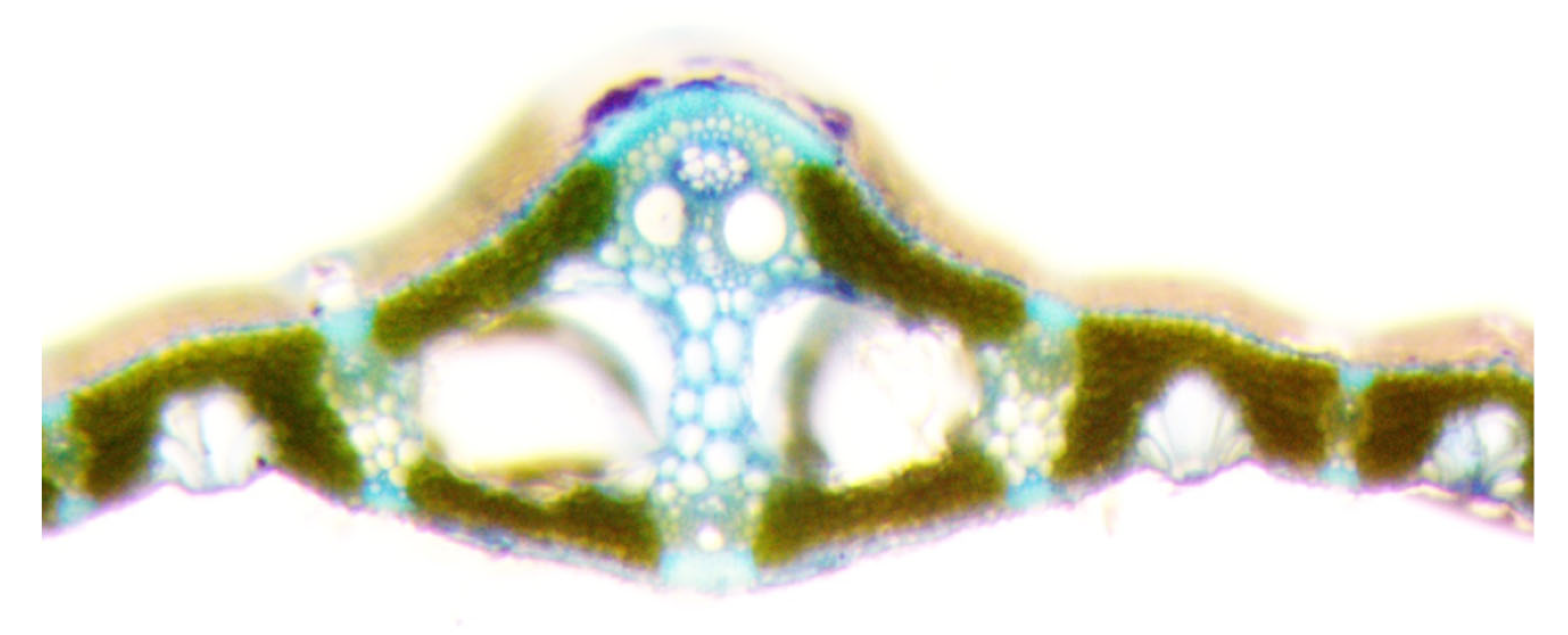

Table 1.
Length of rice plants (cm) when germinating in cylinders (2022).
| № of sample | Name | Length of plants by days of measurements, cm | ||
|---|---|---|---|---|
| 3rd day | 8th day | 13th day | ||
| 983 | Contact × Khao Hlan On | 0.30 | 8.30 | 36.00 |
| 985 | Contact × Khao Hlan On | 0.50 | 11.50 | 39.67 |
| 986 | Contact × Khao Hlan On | 0.50 | 8.00 | 31.67 |
| 987 | Contact × Khao Hlan On | 0.50 | 6.50 | 31.00 |
| 988 | Contact × Khao Hlan On | 0.80 | 9.30 | 39.17 |
| 991 | Contact × Khao Hlan On | 0.50 | 10.00 | 39.67 |
| 992 | Contact × Khao Hlan On | 1.00 | 6.00 | 28.67 |
| 994 | Contact × Khao Hlan On | 0.50 | 6.50 | 31.00 |
| 995 | Contact × Khao Hlan On | 0.10 | 6.00 | 38.33 |
| 996 | Contact × Khao Hlan On | 0.20 | 4.30 | 25.00 |
| 997 | Contact × Khao Hlan On | 0.50 | 8.00 | 40.33 |
| 998 | Contact × Khao Hlan On | 0.30 | 12.00 | 45.33 |
| 1001 | Contact × Khao Hlan On | 0.30 | 2.00 | 18.00 |
| 1004 | Contact × Khao Hlan On | 0.50 | 6.30 | 26.33 |
| 1006 | Contact × Khao Hlan On | 0.40 | 10.50 | 47.00 |
| 1007 | Contact × Khao Hlan On | 0.50 | 11.80 | 35.33 |
| 1008 | Contact × Khao Hlan On | 0.75 | 4.51 | 13.94 |
| 1029 | Kuboyar × Kharsu 80A | 0.50 | 2.75 | 11.11 |
| 1032 | Kuboyar × Kharsu 80A | 1.50 | 3.75 | 9.19 |
| 1033 | Kuboyar × Kharsu 80A | 0.40 | 3.00 | 8.85 |
| 1035 | Kuboyar × Kharsu 80A | 0.75 | 3.40 | 12.28 |
| 1036 (1) | Kuboyar × Kharsu 80A | 0.60 | 4.50 | 11.72 |
| 1036 (2) | Kuboyar × Kharsu 80A | 0.30 | 2.00 | 9.95 |
| 1037 (1) | Kuboyar × Kharsu 80A | 0.75 | 3.25 | 11.08 |
| 1037 (2) | Kuboyar × Kharsu 80A | 0.50 | 3.13 | 9.39 |
| 1038 | Kuboyar × Kharsu 80A | 0.75 | 2.58 | 8.56 |
| 1041 | Kuboyar × Kharsu 80A | 1.17 | 4.75 | 24.94 |
| 1044 | Kuboyar × Kharsu 80A | 0.75 | 2.90 | 8.55 |
| 1047 | Kuboyar × Kharsu 80A | 1.17 | 4.00 | 11.03 |
| 1058 | Kuboyar × Mazhan Red | 1.00 | 3.88 | 9.25 |
| 1063 | Kuboyar × Mazhan Red | 1.50 | 5.33 | 15.85 |
| 1065 | Kuboyar × Mazhan Red | 1.25 | 3.17 | 8.40 |
| 1133 | Kharsu 80A × Contact | 1.39 | 9.38 | 18.50 |
| 1134 | Kharsu 80A × Contact | 1.70 | 9.00 | 25.00 |
| 1417 | Khao Hlan On × Contact | 1.27 | 7.25 | 18.17 |
| 1515 | Khao Hlan On × Kuboyar | 1.23 | 8.17 | 16.43 |
| 1516 | Khao Hlan On × Kuboyar | 1.34 | 6.75 | 20.00 |
| 1518 | Khao Hlan On × Kuboyar | 1.13 | 6.25 | 16.42 |
| 1522 | Khao Hlan On × Kuboyar | 1.23 | 10.00 | 14.57 |
| 1523 | Khao Hlan On × Kuboyar | 1.41 | 9.88 | 18.29 |
| 1525 | Kharsu 80A × Contact | 1.14 | 6.75 | 16.07 |
| 1526 | Kharsu 80A × Contact | 1.16 | 7.00 | 16.14 |
| 1528 | Kharsu 80A × Contact | 1.05 | 6.25 | 14.71 |
| 1529 | Kharsu 80A × Contact | 1.07 | 7.00 | 14.48 |
| 1532 | Kharsu 80A × Contact | 0.93 | 6.00 | 12.60 |
| 1533 | Kharsu 80A × Contact | 1.54 | 7.50 | 23.25 |
| 1535 | Kharsu 80A × Contact | 0.92 | 6.25 | 12.20 |
| 1537 | Kharsu 80A × Contact | 1.57 | 11.00 | 20.33 |
| Medium | 0.86 | 6.42 | 21.12 | |
| Minimum | 0.10 | 2.00 | 8.40 | |
| Maximum | 1.70 | 12.00 | 47.00 | |
| Standard deviation | 0.43 | 2.79 | 11.23 | |
Table 2.
The average values of the parameters of the beams of the conductive system of the leaf of rice plants.
Table 2.
The average values of the parameters of the beams of the conductive system of the leaf of rice plants.
| № | Samples | Number of beams, pcs | Beam diameter, mcm | Area of one beam, mcm2 | Total beam area, mm2 | Leaf thickness, mcm |
|---|---|---|---|---|---|---|
| 1 | 4526 | 50 | 153.2 | 18424 | 0.92 | 700.9 |
| 2 | 4617 | 68 | 165.6 | 21527 | 1.46 | 684.8 |
| 3 | 4641 | 65 | 181.7 | 25917 | 1.69 | 651.1 |
| 4 | 4642 | 72 | 181.4 | 25831 | 1.86 | 879.8 |
| 5 | 4732 | 59 | 181.0 | 25717 | 1.52 | 826.9 |
| 6 | 4758 | 65 | 201.6 | 31904 | 2.07 | 893.6 |
| 7 | 4773 | 60 | 187.4 | 27568 | 1.65 | 709.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



