
Submitted:
30 January 2023
Posted:
02 February 2023
You are already at the latest version
Abstract
Information about molecular mechanism of pesticide resistance in the rust-red flour beetle, a major pest destroying grains and flour across Nigeria is grossly lacking, hindering evidence-based con-trol. Here, we identified to species level three populations of the red-flour beetle from Kano, Ni-geria, as Tribolium castaneum, and investigated the mechanism driving their insecticide resistance. IRAC susceptibility bioassays established high cypermethrin resistance, with LC50s of 4.35-5.46mg/ml in the three populations, NNFM, R/Zaki and Yankaba. High DDT and malathion resistance was observed in NNFM, with LC50s of 15.32- and 3.71mg/ml, respectively. High sus-ceptibility was observed towards dichlorvos in all three populations with LC50s of 0.17-0.35mg/ml, respectively. Synergist bioassay with piperonylbutoxide significantly restored cypermethrin sus-ceptibility, with mortality increasing almost threefold, from 24.8% obtained from 1.25mg/ml of cypermethrin, to 63.3% in synergised group (p=0.013), suggesting a preeminent role of P450s. The two major knockdown resistance (kdr) mutations, T929I and L1014F in the IIS4 and IIS6 fragments of voltage-gated sodium channel were not detected in both cypermethrin-alive and cyperme-thrin-dead beetles, suggesting lesser role of target-site insensitivity. These findings highlight the need to explore alternative control tools for this pest and/or explore incorporation of synergists, like piperonylbutoxide as additional chemistries into pesticides formulations to improve their ef-ficacy.

Keywords:
red flour beetle
; Tribolium castaneum
; pesticides
; insecticides
; resistance
; metabolic
1. Introduction
In Nigeria, wheat is the third most consumed grain after corn and rice [1], with a rise in consumption from 300,000 tonnes in 1970 to about 6 million tonnes as of 2021 [2]. Nigeria imports 98 % of its wheat needs; in 2020 alone $ 2.15 billion was spent in wheat importation, making it the third most imported product in Nigeria, and the country the fourth largest importer of wheat in the world (https://oec.world/en/profile/bilateral-product/wheat/reporter/nga). In Nigeria wheat is milled into flour for conventional bakery products such as bread, cakes, biscuits, as well as making pasta, with the top companies — the Flour Mills of Nigeria Plc, Olam, Dangote, Charghoury and Honeywell milling 32 %, 24 %, 19 %, 11 % and 10 % of the flour market share, respectively, while the rest of other companies mill only 4 % [1].
One of the major challenges to storage of milled flour is infestation by pests, such as the rust-red flour beetle, which in Nigeria is known to destroy/contaminate all the important cereals and its flour, e.g. maize (Zea mays), sorghum (Sorghum bicolor), wheat (Triticum aestivam), rice (Oryza sativa) and millet (Pennisetum glaucum) [3,4]. Indeed, this insect is a serious, cosmopolitan pest of stored grains and grain products in tropical and subtropical regions of the world [5], shown to destroy not only maize, wheat, and other granaries, but also rice bran, oilseed, oil cake, nuts, dried fruit, spices, chocolate, and even bones of animals [6].
Infestation by the rust-red flour beetle results in loss of quality and quantity of food products [4,7,8] as well as bad smells and bad tastes imparted to the food materials from their excrement [8] containing pungent benzoquinones [9]. Several studies in Nigeria have described damage to staple meals and flour from infestation by the red flour beetle. For example, a study in north-eastern Nigeria had described index of susceptibility of flour to infestation by this beetle of 19.65–20.76 % in unpolished flour and 18.89–19.76 % in polished flour [3], while another study from north-central Nigeria had described the potential of this pest to destroy millet, wheat, cassava, yam, etc [10,11]. Another recently published study had documented lower mean weight loses of 8.75 % for cassava and millet on which the red flour beetle was grown compared with higher weight loses seen in maize (25 %) [12].
The control of this pest rely heavily on the use of chemical insecticides/pesticides [13] and several classes of insecticides are used, including pyrethroids, organophosphates, organochlorides, carbamates and abamectin [14,15]. However, continuous application of the pesticides in the field and in storage facilities has accelerated the development of resistance in this agricultural pest to diverse insecticides from several classes [16], threatening the success of pesticide-based control measures [14,17]. For example, multiple resistance towards three different classes of insecticides (deltamethrin, abamectin and chlorpyrifos) have been reported in the Iranian population of the red flour beetle [13]. In contrast, susceptibility to pyrethroids (cypermethrin) was observed in the Egyptian population, which was resistant to pirimiphos-methyl [18]. One of the first studies on resistance in this beetle and its potential molecular mechanism, conducted over six decades ago included a population from Nigeria, shown to be resistant to malathion [17]. Unfortunately, since then little effort has been invested to explore the resistance status of this pest in Nigeria, and its molecular mechanism, despite the anecdotal reports of resistance to pesticides by farmers and flour mills. Most studies conducted in Nigeria evaluated the influence of cereal flours on the growth of this pest, or the efficacy of traditional plants and/or their essential oils, e.g. [19,20]. However, a recently published study on a population from Ondo, south-western Nigeria [21] has evaluated the impact of different rearing substrates on the insecticide resistance response, showing that composite (wheat + cassava) flour increase cypermethrin resistance.
Investigating the pesticide brands/formulations used for protecting stored grains and flours against the red flour beetle and the resistance status of this pest towards the active ingredients will provide information to enhance rationale choice of the pesticides based on evidence, promoting food security in Nigeria, and elsewhere. In this study three populations of the red flour beetle from Kano, northern Nigeria were investigated for insecticide/pesticide resistance using three different insecticides (cypermethrin, dichlorvos, DDT and malathion) which constitute the bulk of the active ingredients utilised in the brand of pesticides in northern Nigeria to treat stored grains and flours. The beetles were identified morphologically and to species level using molecular assays. The molecular mechanism of pyrethroid resistance was explored using cypermethrin, synergized with piperonyl butoxide, to detect the contribution of P450 monooxygenases. Amplification of fragments of the voltage-gated sodium channel encompassing the 929 and 1014 codons detected none of the two mutations, 929I and 1014F associated with pyrethroid/DDT knockdown resistance suggesting a limited role of target-site insensitivity in the nerve agent pesticides.
2. Materials and Methods
2.1. Collection and Rearing of Beetles
The rust-red flour beetles were collected from three different locations within Kano Metropolis (11°58′17″N, 8°35′9″E): (i) Northern Nigeria Flour Mill company (NNFM), (ii) Yankaba market and (iii) Rijyar Zaki (R/Zaki) flour milling/storage facility. Parent stock of the beetles were recovered by sieving from infested flours and reared in an insectarium in Bayero University Kano, under insectary condition advised by the United States Department of Agriculture [22]. The insects were introduced to non-infested dried-wheat flour, mixed with brewer’s yeast at a ratio of 95:5 (w/w) and maintained at 26 °C. Emerged adults were periodically sub-cultured onto fresh flour/yeast mix to prevent overcrowding and starvation.
2.2. Morphological Identification of the Beetles
Morphological identifications of the eggs, larvae, pupae and adult forms of the beetles, as well as the sex of the adults and pupae were performed microscopically, using a combination of the protocol provided by the Department of Entomology and Nematology, University of Florida, United States [7] and the work of Dia and colleagues [23]. Adult males were distinguished by swelling of glands on their prothoracic legs. The sexes were confirmed by observing the genital papillae on the ventral side of the distal abdominal segment under a binocular microscope. The female pupae possess well-developed, protruding genital papillae while the males possess reduced genital papillae compared to females [24].
2.3. Insecticides Susceptibility Filter Paper Bioassays
The choice of insecticides to be tested was made based on the three most popular formulations/brands (Figure S1) used in storage facilities and warehouses in Kano, to control stored products insect pests. These brands are: (i) DAKSH, made up of 100 % w/v dichlorvos; (ii) Cypertrap, made up of 10 % cypermethrin; and (iii) MULTIPHOS, made up 560 g/kg aluminium phosphide plus inert material at 400 g/kg. However, phosphine was not tested due to the unavailability to acquire it. Instead, in addition to dichlorvos and cypermethrin, DDT and malathion were also tested using the NNFM population. This is because previous studies have described populations of the red-flour beetles resistant to these two insecticides.
Analytical grade dichlorvos (98 %), cypermethrin (100 %), DDT (98 %) and malathion (98 %) were purchased from SIGMA Aldrich (Merck KGaA, Darmstadt, Germany). The bioassay was conducted using a modified protocol (Method No. 006) of Insecticide Resistance Action Committee [25]. A total of 9 different concentrations of cypermethrin, dichlorvos DDT, and malathion (ranging from 0.0625 to 10 mg/ml) were made from the stock solution of 200 mg/ml in Dow Corning (Silicone) oil and acetone for cypermethrin and DDT, or Olive oil and acetone for dichlorvos and malathion. Whatman filter papers were cut into 4×4 cm2 and impregnated with 1 ml each of the insecticide solutions above. The filter papers were dried in the dark at room temperature and kept in fridge (4 oC) prior to use. The beetles were held without food for 1 h at 26 oC and 70 % relative humidity prior to tests.
For each concentration four replicates of 20 beetles aged 2 to 4 d old were transferred into 4×4 cm2 plastic tray containing the 4×4 cm2 insecticide impregnated filter paper. The beetles were exposed for 5 h after which they were transferred into another 4×4 cm2 plastic tray containing the 4×4 cm2 non-impregnated paper and knockdown plus delayed mortalities recorded at 24 h. Control groups were exposed to papers impregnated with acetone and carrier oil only.
2.4. Investigation of the Role of Metabolic Resistance Using Synergist Bioassay
To investigate the potential role of metabolic mechanism in resistance, synergist bioassays were conducted using adult beetles with piperonyl butoxide (PBO: an inhibitor of cytochrome P450 monooxygenase), adopting a previous protocol we have utilized for tomato leaf miner, Tuta absoluta [26]. In total, four replicates each of 20 beetles pre-exposed to 4 % PBO-impregnated papers were transferred to filter papers impregnated with 1.5 mg/ml cypermethrin (a concentration that produced ~20 % mortality) and maintained for 5 h, before being transferred into non-impregnated papers. Mortalities were recorded at 24 h. Two control groups were used (i) comprise of four replicates of 20 beetles exposed to PBO-impregnated papers only, and (ii) comprised of four replicates of 20 beetles exposed to acetone plus Dow Corning oil impregnated papers.
2.5. Investigation of the Role of Target-Site Pyrethroid/DDT Insensitivity Resistance Mutations
To investigate the potential role of target site insensitivity resistance mechanism known to reduce the efficacy of pyrethroids and DDT, presence of the knockdown resistance (kdr) mutations in the voltage-gated sodium channel (VGSC) was investigated. The cDNA sequence of T. castaneum para sodium channel (XM962927,2) described in previous publications [27,28] was retrieved from GenBank and used to create the primers below, flanking the IIS4 to IIS6 transmembrane domain of subunit II of the VGSC, which encompass the 929th and 1014th codons, respectively. Nine beetles each, randomly selected from alive and dead following exposure to cypermethrin were used for detection of the above mutations. Genomic DNA was extracted from the beetles using the ethanol precipitation of Livak [29]. The PCR mix comprise 2 µL of gDNA, 2.5 µL of 10x TaqA Buffer, ~0.4 µM (0.85 µL) each of forward (Tcast_IIS4-IIS6_vgscF2: 5’-CCGACTGTTGAGGGTTTTTA-3’) and reverse (Tcast_IIS4-IIS6 _vgscR2: 5’-GCTATTTTATTAGTGTCGTT-3’) primers, 1.25 mM (1.5 µL) of MgCl2, 0.25 mM (0.2 µL) of dNTP mixes and 0.2 µL of KapaTaq DNA polymerase (KAPA Biosystems, Wilmington, MA, USA), in ddH2O. Amplification was carried out using the following conditions: initial denaturation of 3 min at 94 °C, followed by 35 cycles each of 30 s at 94 °C (denaturation), 60 s at 51 °C (primer annealing) and 30 s at 72 °C (extension). This was followed with 5 min final extension at 72 °C. The PCR amplicons were separated on agarose gel stained with pEqGREEN and visualized using Ingenius 3 gel doc (Syngene, Cambridge, UK). Amplicons were purified using Qiaquick purification kit (Qiagen, Hilden, Germany) and sequenced on both strands using the above primers, by GeneWiz (https://www.genewiz.com/en-GB/).
2.6. Molecular Identification of the Beetles
A total of 32 randomly selected F1 parent beetles, 16 cypermethrin-alive and 16 dead were used for DNA extraction. Genomic DNA was extracted [29] and fragments of the mitochondrial DNA cytochrome oxidase subunit I (COI) were amplified to identify the beetles to the species level using polymerase chain reaction [30]. The PCR reaction was carried out using KAPA Taq DNA polymerase kit (KAPA Biosystems, MA, USA) in a total volume of 25 µL. The reaction mix comprise 2 µL of gDNA, 2.5 µL of 10x TaqA Buffer, ~0.4 µM (0.85 µL) each of forward and reverse primers, Tca33F26: 5’-GAATAGTAGGCACTTCATTAAGACTC-3’ and Tca346R24: 5’-CCATGTGCAATGTTTGATGA GAGG-3’, 1.25 mM (1.5 µL) of MgCl2, 0.25 mM (0.2 µL) of dNTP mixes and 0.2 µL of Taq DNA polymerase, in ddH2O. Amplification was carried out using the following conditions: initial denaturation of 3 min at 94 °C, followed by 35 cycles each of 30 s at 94 °C (denaturation), 60 s at 51 °C (primer annealing) and 30 s at 72 °C (extension). This was followed with 10 min final extension at 72 °C. PCR amplicons were separated using agarose gel electrophoresis and visualized to confirm presence of 337 bp fragments diagnostic of T. castaneum.
2.7. Data Analysis
Insecticide susceptibility bioassays data were corrected for mortality in the control using the Abbott’s formula [31]. Linear-probit analysis of larval bioassay result for LC50 was performed using PASW statistics 18 software (http://www.spss.com.hk/statistics/). Insecticide susceptibility bioassay results were analysed using Microsoft excel, which was also used to plot bar charts. Chi square test of significance was performed to assess significance from synergist bioassay using an online tool (https://www.socscistatistics.com/tests/chisquare/default2.aspx).
3. Results
3.1. Morphological and Molecular Identification of Beetles to Species Level
The beetles were morphologically identified as T. castaneum (Herbst 1797) based on morphological keys [7,23]. Bodies of adult beetles were flat, reddish-brown, and measuring about 3-4 mm long (Figure S2). Thorax is slightly darker than the elytra and densely punctuated. The last three antennal segments were larger, almost square. The elytra have a conspicuous field of trichoid sensilla, with numerous gland openings when viewed from dorsal side. Primers targeting the mitochondrial cytochrome oxidase I (mtCOI) generated a band of 337 bp [30] for 31 samples which amplified successfully (Figure S3), confirming that the beetles were T. castaneum.
3.2. Insecticides Resistance Profile of the T. castaneum Populations
Filter paper bioassays were conducted to assess the resistance profile of the T. castaneum beetles to four insecticides from three different classes. While samples from NNFM were tested with cypermethrin, dichlorvos, DDT and malathion, those from Yankaba and R/Zaki were tested with dichlorvos and cypermethrin only. Resistance was observed toward cypermethrin, for all concentrations, with no mortalities at 0.025 mg/ml and 0.25 mg/ml for NNFM and Yankaba populations (Figure 1a).
Only around 50–55 % of the flies were killed at the median concentration of 2.5 mg/ml and average mortalities of 76.7 %, 73.3 % and 63 % were obtained from 5 mg/ml exposure for NNFM, R/Zaki and Yankaba beetles, respectively, which did not increase for the first two populations. The cypermethrin LC50s (lethal concentrations that killed 50 % of the beetles) for NNFM, R/Zaki and Yankaba beetles were 4.35, 4.45 and 5.46 mg/ml, respectively (Table 1).
In the contrary dichlorvos inflicted the highest mortality of all insecticides tested with even low concentrations of 0.25 mg/ml killing 58 %, 60 % and 40 % of the NNFM, R/Zaki and Yankaba populations, while the median concentration of 2.5 % killed between 80-90 % of the beetles (Figure 1b). LC50s of 0.28, 0.17 and 0.35 mg/ml were calculated, respectively for NNFM, R/Zaki and Yankaba (Table 1).
Resistance was observed towards malathion and DDT, with no mortalities at 0.0625- and 0.25 mg/ml, and mortalities of 26.7 and 10.8 % respectively, at the median concentration of 2.5 mg/ml, which increased to only 58.3 % and 26.7 % at the 5 mg/ml (Figure 1c). The LC50s for malathion and DDT were 3.71 and 15.32 mg/ml, respectively.
3.3. Assessment of the Role of Metabolic Resistance in Cypermethrin Resistance
Synergist bioassay was conducted with PBO to investigate the role of CYP450 monooxygenases on cypermethrin resistance. Pre-exposure to 4 % PBO for 1 h followed by 1.25 mg/mL cypermethrin significantly restored susceptibility, with mortalities increasing approximately threefold, from 24.8 % with 1.25 mg/ml cypermethrin alone, to 63.3 % in synergised group (p = 0.013) (Figure 1d).
3.4. Assessment of the Role of VGSC Target-Site Insensitivity Kdr Mutations in Cypermethrin Resistance
A 550 bp fragment of the T. castaneum para sodium channel gene encompassing domains IIS4-IIS6 of the voltage-gated sodium channel was successfully amplified from gDNA of 9 each of cypermethrin-alive and -dead beetles, and sequenced (Figure 2a) to investigate presence of T929I and L1014F kdr mutations. For both alive and dead beetles, the above mutations were absent, with the codons ACG (Figure 2b) and CTT (Figure 2c) coding for threonine and leucine, respectively present in both the two contrasting phenotypes.
4. Discussion
Reducing post-harvest and storage losses of food stocks is of crucial importance to promoting food availability and security in sub-Saharan Africa, where a significant food deficit is experienced in most countries [32], partly due to biotic agents like agricultural insect pests which contaminate and destroy food items. However, one of the major challenges facing control of the pests is resistance in most the insect species. In this study we established resistance to three chemicals which are used as pesticides for control of the rust-red flour beetle affecting grains and stored flour in northern Nigeria and explored the possible molecular mechanism of pyrethroids (one of the major ingredients used in pesticide formulations) resistance.
Morphological and molecular identification of the beetles from this study confirmed that they are T. castaneum (Herbst, 1797). Several studies have utilized a combination of morphological and molecular approaches to differentiate the species of beetles infesting stored food produce [5,23,30,33] and its ability to destroy grains, cereals and flour, in Nigeria, Africa and globally [7,18,34,35]. Indeed, many studies have identified red flour beetle as an important pest that destroyed flour in Nigeria. For example, Ehisianya and colleagues [12] have demonstrated the role of this beetle in invasion and destruction of different types of flours including maize, wheat, millet, sorghum and cassava flour from Umudike, Southern-eastern Nigeria. Another study on the major flour pests from south-western Nigeria have revealed this beetle as the major species alongside Tenebrioides mauritanicus and Dinoderus porcellus [36]. A study from north-eastern Nigeria [4] has also described this pest to be predominant among flour pests in Maiduguri.
The development and spread of resistance in primary and secondary agricultural pests is of interest to farmers and warehousing businesses [37]. And without understanding the resistance profile of the major pests and its mechanism it will not be possible to control the pest through resistance management. In this study, high cypermethrin resistance was observed in all the three populations of the T. castaneum studied. A recent paper on a population from Ondo, south-western Nigeria [21] has evaluated the impact of different rearing substrates on the insecticide resistance response of red flour beetles, showing that composite (wheat + cassava) flour increase cypermethrin resistance. Several studies have reported similar or contrasting resistance cypermethrin phenotype. For example, in Egypt, a recent study [38] has revealed cypermethrin exhibited low toxicity to a population of T. castaneum when tested using thin film technique, with LC50 value 2.20 mg/ml obtained 24 h after treatment, a value which is on average half the LC50S from our study. On the contrary, Naeem and colleagues [39] had described a population of T. castaneum highly susceptible to cypermethrin, with mortalities of > 94 % when exposed to 5 mg/mL of cypermethrin for 24 h. The LC50 obtained by Naeem and colleagues (0.09%, ~0.9 mg/mL) was four to five times lower than LC50s obtained in all 3 populations from Kano. Another study conducted by Khalequzzaman and Khanom [40] had reported very high toxicity of cypermethrin to the adult flour beetles from Bangladesh, which is contrary to our findings. High resistance to type II pyrethroids cypermethrin had also been previously reported from Italian population of T. castaneum [35].
Highest susceptibility was obtained from dichlorvos exposure, even at low concentrations, which is similar to the findings of Andric et al. [41] who reported high mortalities (> 95 %) and another study which reported absolute mortality upon exposure of Indian populations to this organophosphate [42]. The finding of high malathion resistance in our study was not surprising.
Malathion resistance had been described several decades ago in Nigerian population of the red flour beetle, and its potential molecular mechanism explored [17,43]. Several other studies have described malathion resistance in T. castaneum, for example, studies from Australia [44,45].
Highest resistance was observed with DDT, which was not surprising, as one of the pioneering studies on resistance in the red flour beetle was a selection study in which a colony with a slight tolerance at 1.8 ppm was selected for several generations until stocks with resistance factor of x166 were generated [46].
PBO is increasingly being used as a synergist to investigate the role of P450 monooxygenases in metabolic resistance [47], for example in several studies targeting insect pests of public health and agricultural importance, using mostly pyrethroids [26,48,49,50]. The contribution of monooxygenases in T. castaneum resistance has been demonstrated in insects by [51] and other researchers. For example, using a combination of functional genomics and reverse genetics approaches, Zhu and colleagues [50] have shown that overexpression of a P450 CYP6BQ9 was associated with deltamethrin resistance in QTC279 strain of T. castaneum compared to the susceptible strain. In a similar finding, an ATP-binding cassette transporter, TcABCG–H has been found overexpressed in response to insecticide induction [52]. The synergist assay conducted in our study demonstrated that P450 monooxygenases-mediated metabolic mechanism is associated with cypermethrin resistance.
Study of mutation in knockdown resistance gene (kdr) has led to the identification several other mutations that alone or in combination with other mechanisms were known to confer resistance to pyrethroids in arthropod species [55]. And previous studies have associated polymorphisms in the T. castaneum voltage gated sodium channel (VGSC) with insecticide resistance [27,52,53,54]. The absence of the T929I and 1014F kdr mutations in the VGSC from the T. castaneum in this study suggest the preeminent role of metabolic mechanism in pyrethroid resistance. The T929I mutation has been shown to be associated with DDT resistance in the maize weevil, Sitophilus zeamais, in a previous study [28]. Resistance in pyrethroid-resistant rice weevils was associated with the combination of the L1014F kdr mutation, and increases in walking and detoxification activities, while the T929I mutation combined with increases in walking activity were the primary pyrethroid resistance mechanisms in maize weevils [27].
5. Conclusions
In conclusion, the rust-red fluor beetle, T. castaneum from Kano, northern Nigeria is multiple resistant to several pesticides in use for its control (including pyrethroids, DDT and organophosphates), threatening the quality and quantity of stored grains, cereals and fluor. The resistance is probably driven by metabolic mechanism, particularly the cytochrome P450 monooxygenases. The absence of the two major kdr mutations suggest the lesser role of target-site mutations in pyrethroids and/or DDT resistance. These findings highlight the challenges associated with the control of this secondary pest of stored products and underscore the need to explore alternative approaches for its control, as well as using synergists against the major metabolic genes driving the resistance, such as the CYP450s.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Three of the most widely used pesticide formulations for control of beetles of stored flour in northern Nigeria., Figure S2: Agarose gel of amplified fragments of mitochondrial cytochrome oxidase I for identification of the red-rust flower beetles from Kano, Nigeria, to species level. First and last lanes (L) of each panel is a hyper ladder IV DNA ladder [Bioline 100–1013 bp (40–200 ng/band)]. 31 samples produced 337 bp band sizes characteristic of T. castaneum., Figure S3: Morphology of the posterior and anterior body of red flour beetles from Kano. Pictures were captured using a stereomicroscope at ×2 magnification., Files S1: Genotyping of kdr mutations in cypermethrin-alive and -dead T. castaneum. Sequences of fragment of the voltage-gated sodium channel encompassing the IIS4-IIS6 transmembrane domain, harboring the T929I and L1014F mutations.
Author Contributions
Conceptualization, SSI.; methodology, MMM, MAM and MA.; software, SSI and MMM.; validation, SSI.; formal analysis, MMM and SSI.; investigation, MMM, MAM, MA and SSI.; resources, SSI.; data curation, SSI and MMM.; writing—original draft preparation, MMM.; writing—review and editing, SSI.; visualization, SSI.; supervision, SSI.; project administration, SSI.; funding acquisition, SSI. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated in this research is contained within the article or Supplementary Material File S1 provided.
Conflicts of Interest
The authors declare no conflict of interest.
References
- C. Lyddon, “Focus on Nigeria,” WORLD-GRAIN.com SOSLAND PUBLISHING COMPANY, 2022, 4, 88–100, Accessed: Jan. 22, 2022.
- J. Reidy, “Nigeria wheat initiative reaches first-year goal.,” SOSLAND PUBLISHING COMPANY 2022, www.world-grain.com. Available online: https://www.world-grain.com/articles/17449-nigeria-wheat-initiative-reaches-first-year-goal# (accessed on 9 December 2022).
- J. M. Turaki, B. M. Sastawa, B. G. J. Kabir, and N. E. S. Lale, “Susceptibility of flours derived from various cereal grains to infestation by the rust-red flour beetle (Tribolium castaneum Herbst) (Coleoptera: Tenebrionidae) in different seasons,” J. Plant Prot. Res. 2007, 47, 279–288. Available online: http://www.plantprotection.pl/Susceptibility-of-flours-derived-from-various-cereal-grains-to-infestation-by-the.91119.0.2.html.
- D. M. Mailafiya, Y. T. Maina, Y. M. Degri, and U. U. Gadzama, “Traders’ perception of food grain storage and pest management in Dalwa market, Borno State, Nigeria.” J. Agric Crop Res. 2014, 2, 62-70, ISSN: 2384-731X.
- R. Jagadeesan, P. J. Collins, G. J. Daglish, P. R. Ebert, and D. I. Schlipalius, “Phosphine resistance in the rust red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae): Inheritance, gene interactions and fitness costs,” PLoS ONE 2012, 7, 2012. [CrossRef]
- R. Ahmad, S. Hassan, S. Ahmad, S. Nighat, Y. K. Devi, K. Javeed, S. Usmani, M. J. Ansari, S. Erturk, M. Alkan and B. Hussain “Stored Grain Pests and Current Advances for Their Management,” in Postharvest Technology - Recent Advances, New Perspectives and Applications. IntechOpen, 2022. [CrossRef]
- 7. R. Baldwin and T. R. Fasulo, “Confused Flour Beetle, Tribolium confusum Jacquelin du Val and Red Flour Beetle, Tribolium castaneum (Herbst) (Insecta: Coleoptera: Tenebrionidae),” Edis, 1–5, 2020. [CrossRef]
- 8. E. M. Pires, E. Q. Souza, R. M. Nogueira, M. A. Soares, T. K. R. Dias, and M. A. Oliveira, “Damage caused by Tribolium castaneum (Coleoptera: Tenebrionidae) in Stored Brazil nut,” Sci. Electron. Arch. 2017, 10, 1–5. [CrossRef]
- 9. J. F. Campbell and C. Runnion, “Patch exploitation by female red flour beetles, Tribolium castaneum,” J. Insect Sci. 2003, 3, 20. https://pubmed.ncbi.nlm.nih.gov/15841236/. [CrossRef] [PubMed]
- M. I. Nafiu, N. Lawal, A. M. Aminu, A. S. Muohammed, R. I. Abdullahi, and T. A. Bello, “Productivity of Tribolium Species (Coleoptera; Tenebrionidae) within Flours Derived from Edible Tubers and Effects on Proximate Composition.” Int. J. Sci. Glob Sustain. 2020, 6, 12. https://fugus-ijsgs.com.ng/index.php/ijsgs/article/view/68.
- F. A. Ajayi and S. A. Rahman, “Susceptibility of some staple processed meals to red flour beetle, Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae),” Pak. J. Biol. Sci. 2006, 9, 1744–1748, 2006. [CrossRef]
- C. N. Ehisianya, A. G. Stephen, and B. N. Onunka, “Development of Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) and Damage to Selected Flours in Storage,” Nig. Agric. J. 2022, 53, 193–198. https://www.ajol.info/index.php/naj/article/view/232673.
- S. Vojoudi, M. Saber, V. Mahdavi, H. Golshan, and Z. Abedi, “Efficacy of some Insecticides Against Red Flour Beetle, Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) Adults Exposed on Glass, Ceramic Tile, Plastic and Paper Disc Surfaces,” J. Lif Sci. 2012, 6, 405–410.
- H. Pieterse, G. G. M. Schulten, and W. Kuyken, “A study on insecticide resistance in Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae) in Malawi (Central Africa),” J. Stored Prod. Res. 1972, 8, 183–191. [CrossRef]
- Rauf and R. M. Wilkins, “Malathion-resistant Tribolium castaneum has enhanced response to oxidative stress, immunity, and fitness, Pest. Biochem. Physiol. 2022, 184. ISSN 0048-3575. [CrossRef]
- U. Shamjana and T. Grace, "Review of Insecticide Resistance and Its Underlying Mechanisms in Tribolium castaneum,” Insecticides - Impact and Benefits of Its Use for Humanity," Review of Insecticide Resistance and Its Underlying Mechanisms in Tribolium castaneum, 2022, IntechOpen. [CrossRef]
- E. Dyte and D. G. Blackman, “The spread of insecticide resistance in Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae),” J. Stored Prod. Res. 1970, 6, 255–261, 1970. [CrossRef]
- M. A. Attia, T. F. Wahba, N. Shaarawy, F. I. Moustafa, R. N. C. Guedes, and Y. Dewer, “Stored grain pest prevalence and insecticide resistance in Egyptian populations of the red flour beetle Tribolium castaneum (Herbst) and the rice weevil Sitophilus oryzae (L.),” J. Stored Prod. Res. 2020, 87, 101611. [CrossRef]
- F. K. Ayo, A. S. Hezekiah, and A. A. Temitope, “Comparative Pesticidal Activities of Essential Oils Extracted from Indigenous Plants Against Tribolium castaneum Herbst (Coleoptera: Tenebrionidae),” Asian J. Sci. Res. 2019, 12, 502–507. [CrossRef]
- A. Ukeh and S. B. A. Umoetoka, “Repellent effects of five monoterpenoid odours against Tribolium castaneum (Herbst) and Rhyzopertha dominica (F.) in Calabar, Nigeria,” Crop Prot. 2011, 30, 1351–1355. [CrossRef]
- A. Gbaye, E. A. Oyeniyi, E. I. Ogunleye, and O. V. Aloba, “The impact of cassava and wheat flour substrates on the biological parameters and insecticide response in Tribolium castaneum (Herbst),” Biocatal. Agric. Biotechnol. 2021, 38. [CrossRef]
- R. W. Beeman, S. Haas, and K. Friesen, “An Introduction to the Care and Handling of Tribolium castaneum,” USDA Tribolium stock maintenance, 2022. https://www.ars.usda.gov/plains-area/mhk/cgahr/spieru/docs/tribolium-stock-maintenance/.
- A. K. M. Dia, A. G. R. J. Sarr, A. Kafom, D. Ngom, T. Diome, C. Thiaw, S. Ndiaye and M. Sembene., “Morphological Identification of Trophic Tribolium Castaneum Populations Herbst (Coleoptera, Tenebrionidae) in West Africa.,” Int. J. Adv. Res. 2018, 6, 203–216. [CrossRef]
- R. Parthasarathy, A. Tan, H. Bai, and S. R. Palli, “Transcription factor broad suppresses precocious development of adult structures during larval–pupal metamorphosis in the red flour beetle, Tribolium castaneum R., 2008, 125, 299–313. [CrossRef]
- IRAC, “IRAC Test Methods Series. Test no. 006.,” Insecticides Resistance Action Committee Susceptibility., 2009, Version 3. https://irac-online.org/content/uploads/2009/09/Method_006_v3_june09.pdf (accessed on 2 February 2022).
- Bala, M. M. Mukhtar, H. K. Saka, N. Abdullahi, and S. S. Ibrahim, “Determination of insecticide susceptibility of field populations of tomato leaf miner (Tuta absoluta) in northern Nigeria,” Agriculture 2019, 9, 7. [CrossRef]
- Haddi, W. R. Valbon, L. O. Viteri Jumbo, L. O. de Oliveira, R. N. C. Guedes, and E. E. Oliveira, “Diversity and convergence of mechanisms involved in pyrethroid resistance in the stored grain weevils, Sitophilus spp.,” Sci. Rep. 2018, 8, 1–16. [CrossRef]
- R. A. Araújo, M. S. Williamson, C. Bass, L. M. Field, and I. R. Duce, “Pyrethroid resistance in Sitophilus zeamais is associated with a mutation (T929I) in the voltage-gated sodium channel,” Insect Mol. Biol. 2011, 20, 437–445. [CrossRef]
- J. Livak, “Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis,” Genetics, 1984, 107, 611–634. [CrossRef]
- T. Zhang, Y. Wang, W. Guo, D. Luo, Y. Wu, Z. Kučerová, V. Stejskal, G. Opit, Y. Cao, F. Li and Z. Li “DNA barcoding, species-specific PCR and real-Time PCR techniques for the identification of six Tribolium pests of stored products,” Sci. Rep., 2016, 6, 1–11. [CrossRef]
- W. S. Abbott, “A method of computing the effectiveness of an insecticide,” J. Econ. Entomol., 1925, 18, 265–267. [CrossRef]
- R. J. Hodges, J. C. Buzby, and B. Bennett, “Postharvest losses and waste in developed and less developed countries: Opportunities to improve resource use,” J. Agric. Sci., 2011, 149, 37–45. [CrossRef]
- Q. Ming, A. Wang, and C. Cheng, “Molecular identification of Tribolium castaneum and T. confusum (Coleoptera: Tenebrionidae) using PCR-RFLP analysis,” J. Genet., 2014, 93, e17–e21. https://pubmed.ncbi.nlm.nih.gov/24823304/.
- Y. Kayode, C. O. Adedire, and R. O. Akinkurolere, “Influence of four cereal flours on the growth of Tribolium castaneum Herbst (Coleoptera: Tenebrionidae),” Ife J. Sci. 2014, 16, 505–516.
- Rossi, S. Cosimi, and A. Loni, “Insecticide resistance in Italian populations of Tribolium flour beetles,” Bull. Insectolog. 2010, 63, 251–258, ISSN 1721-8861.
- Abo and E. Ja, “An evaluation of infestation of insect pests of flours in Benin City, Edo State, Nigeria,” J. Appl. Sci. Environ. Manag. 2014, 18, 487–494. Available online: https://www.ajol.info/index.php/jasem/article/view/109917.
- J. A. McFarlane, “Guidelines for pest management to reduce stored food losses caused by insects and mites.,” Greenwich Academic Literature Archive (GALA)., 1989, no. 22, p. 62. ODNRI Bulletin No. 22. Available online: https://gala.gre.ac.uk/id/eprint/10727/1/Doc-0115.pdf (accessed on 12 December 2022).
- S. Abouelkassem, A. A. Salem, and A. R. B. Arab, “Toxicity and development of resistance in Tribolium castaneum and Sitophilus oryzae oryzae to certain selected insecticides,” Egypt. J. Plant Prot. Res. Inst. 2018, 1, 188-198. http://www.ejppri.eg.net/pdf/v1n2/11.pdf.
- Naeem, S. I. Anjum, M. Ismail, J. Khan, S. Bibi., “Laboratory assessment of different botanical extracts and cypermethrin against insect pests,” J. Entomol. Zool. Stud. 2015, 3, 84–88. https://www.entomoljournal.com/archives/2015/vol3issue5/PartB/3-5-36.pdf.
- Khalequzzaman and M. Khanom, “Effects of cypermethrin alone and in combination with leaf and seed extracts of neem against adult Tribolium castaneum (Herbst),” Univ. J. Zool. Rajshahi Univ. 2006, 25, 45–49. [CrossRef]
- Andrić, P. Kljajić, I. Perić, and M. Pražić Golić, “Susceptibility of red flour beetle Tribolium castaneum (Herbst) populations from Serbia to contact insecticides,” 10th Int. Work. Conf. Stored Prod. Prot. 2010, 425, 869–873. [CrossRef]
- S. D. Reddy and C. Srivastava, “Persistent Toxicity of Malathion and Dichlorvos on Jute Surface Against Tribolium castaneum (Herbst),” Ann. Plant Prot. Sci. 2004, 12, 41–44. Available online: https://www.indianjournals.com/ijor.aspx?target=ijor:apps&volume=12&issue=1&article=011.
- E. A. Parkin, E. I. C. Scott, and R. Forster, "Increased resistance of stored-product insects to insecticides. The resistance of field strains of beetles," Pest Infest. Res. 1961, 34-35, 1962.
- B. R. Champ and M. J. Campbell-Brown, “Insecticide resistance in Australian Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae) - II. Malathion resistance in eastern Australia,” J. Stored Prod. Res. 1970, 6, 111–131. [CrossRef]
- G. GREENIN, “Malathion Resistance in the Red Flour Beetle,” Aust. J. Entomol. 1970, 9, 160–162. [CrossRef]
- C. E. Dyte and D. G. Blackman, “Selection of a DDT-resistant strain of Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae),” J. Stored Prod. Res. 1967, 2, 211–228. [CrossRef]
- R. Feyereisen, “Insect P450 inhibitors and insecticides: Challenges and opportunities,” Pest Manag. Sci. 2015, 71, 793–800. [CrossRef]
- S. S. Ibrahim et al., “Exploring the mechanisms of multiple insecticide resistance in a highly plasmodium-infected malaria vector Anopheles funestus sensu stricto from Sahel of northern Nigeria,” Genes 2020, 11, 454. [CrossRef]
- M. Mukhtar and S. S. Ibrahim, “Temporal Evaluation of Insecticide Resistance in Populations of the Major Arboviral Vector Aedes Aegypti from Northern Nigeria,” Insects 2022, 13, 187. [CrossRef]
- F. Zhu, R. Parthasarathy, H. Bai, K. Woithe, M. Kaussmann, R. Nauen, D. A. Harrison, and S. R. Palli., “A brain-specific cytochrome P450 responsible for the majority of deltamethrin resistance in the QTC279 strain of Tribolium castaneum,” Proc. Natl. Acad. Sci. USA 2010, 107, 8557–8562. [CrossRef]
- C. T. Zimmer and R. Nauen, “Cytochrome P450 mediated pyrethroid resistance in European populations of Meligethes aeneus (Coleoptera: Nitidulidae),” Pestic. Biochem. Physiol. 2011, 100, 264–272. [CrossRef]
- J. Rösner, J. Tietmeyer, and H. Merzendorfer, “Functional analysis of ABCG and ABCH transporters from the red flour beetle, Tribolium castaneum,” Pest Manag. Sci. 2021, 77, 2955–2963. [CrossRef]
- M. Abd El Halim, B. M. H. Alshukri, M. S. Ahmad, E. Y. T. Nakasu, M. H. Awwad, E. M. Salama, A. M. R. Gatehouse and M. G. Edwards “RNAi-mediated knockdown of the voltage gated sodium ion channel TcNav causes mortality in Tribolium castaneum,” Sci. Rep. 2016, 6, 29301. [CrossRef]
- Rösner, B. Wellmeyer, and H. Merzendorfer, “Tribolium castaneum: A Model for Investigating the Mode of Action of Insecticides and Mechanisms of Resistance,” Curr. Pharm. Des. 2020, 26, no. 29, 3554–3568. [CrossRef]
- Dong, Du Y, Rinkevich F, Nomura Y, Xu P, Wang L, Silver K, Zhorov B.S., "Molecular biology of insect sodium channels and pyrethroid resistance." Insect Biochem Mol Biol. 2014, 50, 1-17. [CrossRef]
Figure 1.
Insecticide susceptibility bioassays with various concentrations of insecticides. Each bar represents percentage mortality from four individual replicates for each concentration of (a) cypermethrin, (b) dichlorvos, (c) malathion and DDT for the NNFM population only, and (d) PBO synergist bioassay with cypermethrin. Error bars indicate standard deviation of means. ** p = 0.01 for comparison between synergized and un-synergized bioassays.
Figure 1.
Insecticide susceptibility bioassays with various concentrations of insecticides. Each bar represents percentage mortality from four individual replicates for each concentration of (a) cypermethrin, (b) dichlorvos, (c) malathion and DDT for the NNFM population only, and (d) PBO synergist bioassay with cypermethrin. Error bars indicate standard deviation of means. ** p = 0.01 for comparison between synergized and un-synergized bioassays.
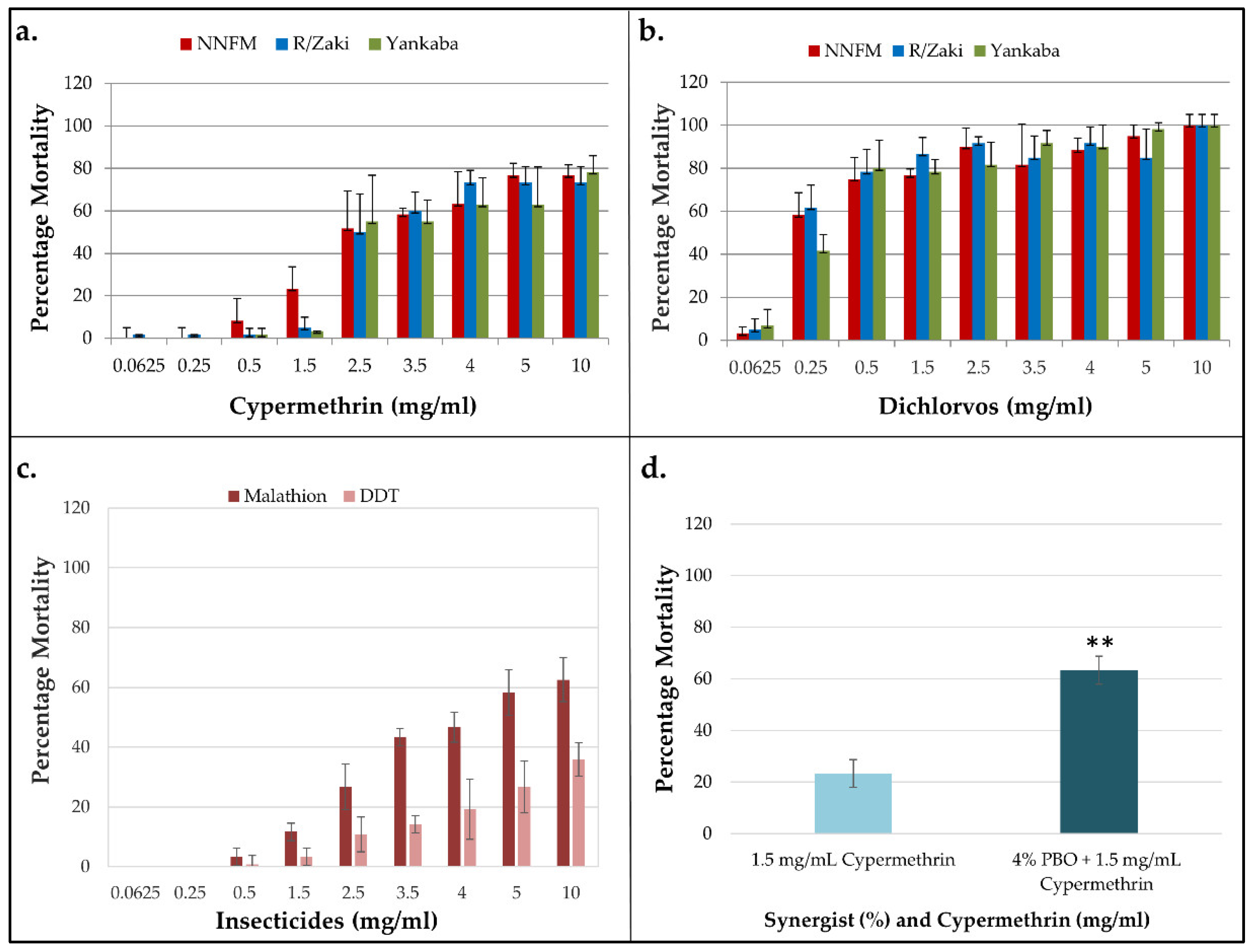
Figure 2.
Analysis of fragments of voltage-gated sodium channel gene. a. Amplified segments of the T. castaneum VGSC. L is hyperladder IV DNA ladder [Bioline 100–1013 bp (40–200 ng/band)]; b. Analysis of the polymorphism of fragments of the VGSC gene spanning the T929I and L1014F kdr mutations codons.
Figure 2.
Analysis of fragments of voltage-gated sodium channel gene. a. Amplified segments of the T. castaneum VGSC. L is hyperladder IV DNA ladder [Bioline 100–1013 bp (40–200 ng/band)]; b. Analysis of the polymorphism of fragments of the VGSC gene spanning the T929I and L1014F kdr mutations codons.
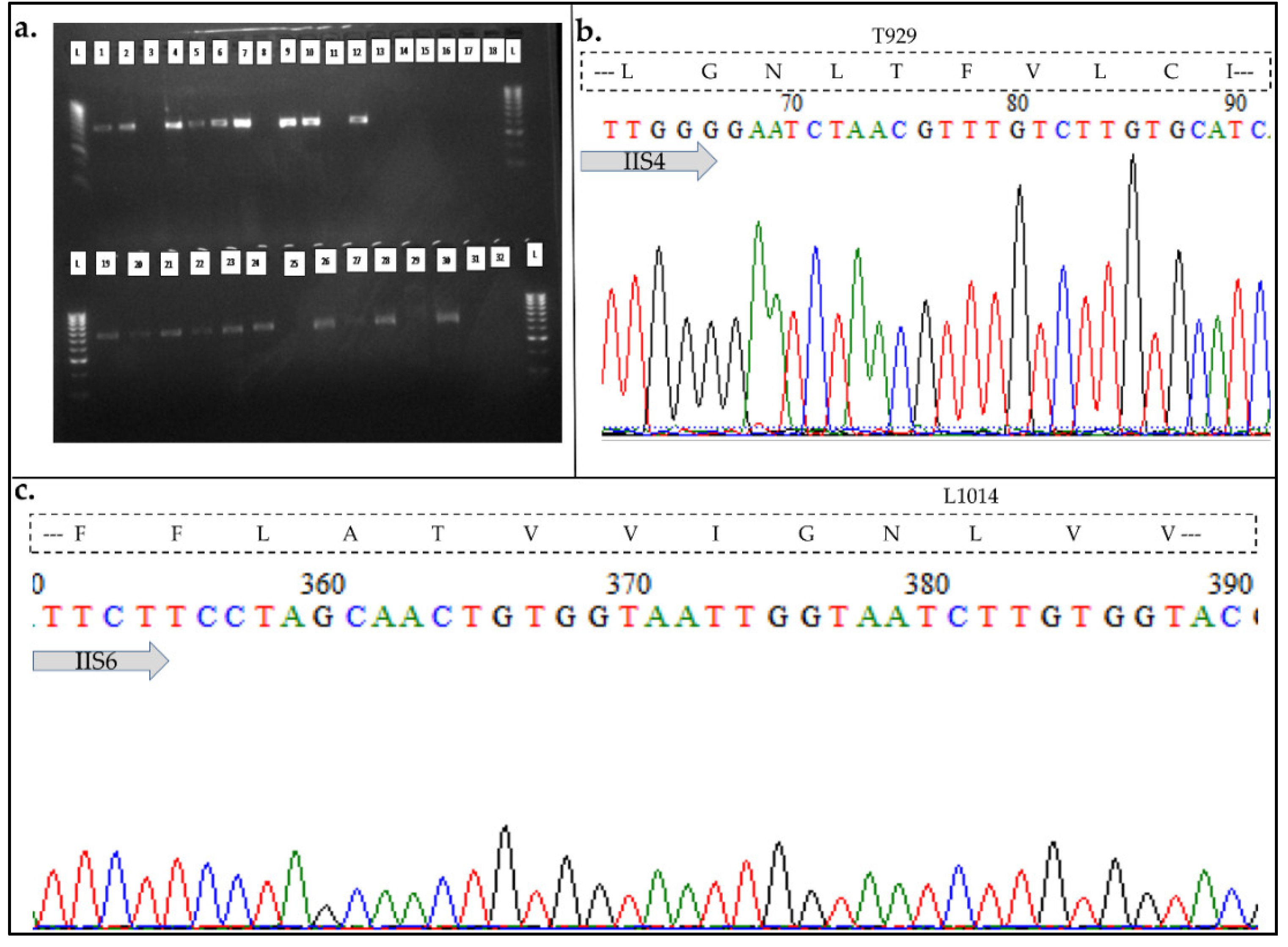

Table 1.
Results of probit analysis for insecticide susceptibility bioassays with various T. castaneum populations.
Table 1.
Results of probit analysis for insecticide susceptibility bioassays with various T. castaneum populations.
| Population | Insecticide | n | LC50 (mg/ml) (95% CI) |
|---|---|---|---|
| NNFM | Cypermethrin | 720 | 4.35 (3.68-5.01) |
| DDT | 720 | 15.32 (5.09-20.33) | |
| Dichlorvos | 720 | 0.28 (0.07-0.63) | |
| Malathion | 720 | 3.71 (1.72-5.92) | |
| R/Zaki | Cypermethrin | 720 | 4.45 (3.83-5.08) |
| Dichlorvos | 720 | 0.17 (0.03-0.46) | |
| Yankaba | Cypermethrin | 720 | 5.46 (4.64-6.27) |
| Dichlorvos | 720 | 0.35 (0.06-0.63) |
n = number of adult T. castaneum used per experiment; LC50 = concentration that killed 50% of the experimental insects; CI = 95 % confidence interval.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



