
Submitted:
09 February 2023
Posted:
13 February 2023
You are already at the latest version
Abstract
Viruses with encephalitogenic potential can cause neurological conditions of clinical and epidemiological importance, such as Saint Louis encephalitis virus, Venezuelan equine encephalitis virus, Eastern equine encephalitis virus, Western equine encephalitis virus, dengue virus, Zika virus, Chikungunya virus, Mayaro virus and West Nile virus. The objective of the present study was to determine the number of arboviruses with neuroinvasive potential isolated in Brazil that corresponds to the collection of viral samples belonging to the Section of Arbovirology and Hemorrhagic Fevers (SAARB/IEC) of the Laboratory Network of National Reference for Arbovirus Diagnosis from 1954 to 2022. A total of 1,090 arbovirus samples with encephalitogenic potential were isolated from mice. From a total of 7,054 samples intended for cell culture. The isolates from mosquitoes totaled 659 viruses. The emergence of new arboviruses may be responsible for diseases still unknown to humans, making the Amazon region a hotspot for infectious diseases due to its fauna and flora species characteristics. The detection of circulating arboviruses with the potential to cause neuroinvasive diseases is constant, which justifies the continuation of active epidemiological surveillance work that offers adequate support to the public health system regarding the virological diagnosis of circulating arboviruses in Brazil.
Keywords:
arbovirus
; viral isolation
; cell culture
; neurotropic
; public health
1. Introduction
The term arbovirus is derived from the union of the first syllables of the English expression arthropod-borne (transmitted by arthropods), followed by the suffix virus. According to the concept established by the World Health Organization (WHO) [1], “Arboviruses are viruses maintained in nature through biological transmission between susceptible vertebrate hosts and hematophagous arthropods, or between arthropod hosts by the transovarian and/or venereal route. They are able to reproduce in vertebrate hosts, multiply in arthropod tissues and be passed on to new susceptible vertebrates by arthropod bites after an extrinsic incubation period”.
Arbovirus infections can result in a wide spectrum of clinical syndromes, from mild febrile illness to hemorrhagic fevers and neuroinvasive forms. However, most human arbovirus infections are asymptomatic or oligosymptomatic [2].
The Centers for Disease Control and Prevention (CDC) defines meningitis (aseptic), encephalitis and acute flaccid paralysis as main neurological manifestations of arbovirus infection, although many cases are not recognized as manifestations caused by these viruses, which contributes so that the diagnosis remains a challenge to be faced [3].
Population growth, disorderly urbanization and increased human traffic and international trade have contributed to the emergence and spatial spread of arboviruses in recent decades [4,5]. Every day, other forms of arbovirus transmission, such as blood transfusion, organ transplantation, sexual or perinatal transmission and laboratory exposure, gain importance [6].
Currently, approximately seven viral families (Peribunyaviridae, Togaviridae, Flaviviridae, Sedoreoviridae, Rhabdoviridae, Asfaviridae and Phenuiviridae) harbor arboviruses capable of infecting humans and animals and have become important in public health [7,8,9,10], with emphasis on dengue virus (DENV), Chikungunya virus (CHIKV), Zika virus (ZIKV), West Nile virus (WNV) and Rocio virus (ROCV) as neuroinvasive viruses of greatest epidemiological interest in Brazil [2].
The Alphavirus genus belongs to the Togaviridae family, which is composed of 32 viruses, including the Mayaro virus (MAYV), Eastern equine encephalitis virus (EEEV), Chikungunya virus (CHIKV) and Western equine encephalitis virus (WEEV), which are important in public health. Additionally, included are the Aura virus (AURAV), Una virus (UNAV), Mucambo virus (MUCV), Triniti virus (TNTV) and Pixuna virus (PIXV) [11]. Alphaviruses are approximately 70 nm in diameter, with an icosahedral capsid enveloped with positive-sense, single-stranded RNA genomes ranging from 10 to 12 kb [12]. These viruses are transmitted by mosquitoes, although other blood-sucking insects, including ticks, lice and bed bugs, have been implicated in transmission [13,14]. Vertebrate hosts include humans, nonhuman primates, equids, birds, amphibians, reptiles, rodents and pigs (Figure 1) [15].
The Flavivirus genus belongs to the Flaviviridae family and is composed of 53 viral representatives, including Japanese encephalitis virus (JEV), DENV, ZIKV, and WNV, of public health importance. The following viruses are also included: Cacipacoré virus (CACV), Ilhéus virus (ILHV), Bussuquara virus (BUSV) and Iguape virus (IGUV) [11]. Flaviviruses have positive-sense RNA of approximately 9.0-13 kb with an icosahedral symmetrical structure with an approximate diameter of 40 to 60 nm [16].
Flaviviruses are transmitted by arthropod vectors through transovarian or vertical transmission and by cycles between vectors and vertebrate hosts. Ticks also contribute to the maintenance of these viruses via vertical and transstadial routes. Rodents and bats can also be part of the flavivirus transmission cycle, with no known arthropod vectors. Humans are not definitive hosts but accidental hosts when they enter the habitat of these mosquitoes (Figure 1). In certain cases, flaviviruses can be transmitted to humans by blood products, organ transplantation, unpasteurized milk, or aerosols. It is worth mentioning that individuals who move to areas endemic with these vectors are at greater risk of acquiring the disease [16].
2. Alphavirus Genus
The Togaviridae family includes Alphavirus genus comprising 32 viral species [17], with single-stranded RNA genome, spherical and enveloped. The envelope contains spikes of glycoproteins (E1 and E2). The viral particle measures 70 nm in diameter and infect vertebrates and invertebrates. The Alphaviruses include mosquito transmitted diseases such as Eastern, Western, or Venezuelan equine encephalitis, which are characterized by fever, malaise, headache and encephalitis, and diseases caused by the Chikungunya, Ross River, Mayaro, and Sindbis viruses, which produce arthralgias. The syndrome produced by viruses such as Chikungunya result in intense joint pain, high fevers, and a rash, and although infection is self-limited, symptoms may persist for years. In addition to mosquito transmission, the virus can be transmitted vertically as well as with infected blood transfusions [12,15].
2.1. Mayaro Virus (MAYV)
MAYV originates from the Americas and was first isolated in 1954 in Trinidad and Tobago, islands in the Caribbean region, and was also isolated in primates and migratory birds in the United States. In Brazil, the first identification took place in 1955 during an outbreak of an acute febrile illness with headache that affected rural workers on a farm located on the left banks of the Guamá River in Belém. After three decades, new outbreaks were confirmed in localities in Pará, such as Benevides (1991), Belterra (1978), Santa Bárbara and Conceição do Araguaia (1981) [18,19,20,21,22].
In a recent epidemiological investigation, MAYV was detected in human populations residing in the municipalities of Parauapebas and Canaã dos Carajás in the Pará state, evidencing the circulation of arboviruses in this area, which reinforces the importance of epidemiological surveillance in areas that are under pressure from environmental changes [23].
The transmission of MAYV occurs through the bite of a female mosquito belonging to the genus Haemagogus, particularly the species Haemagogus janthinomys [24], which has diurnal and wild habits and lives in treetops, facilitating its contact with the host and consequently its transmission. The global expansion of the two main urban vectors (Aedes aegypti and Aedes albopictus) and the increasing number of international travelers certainly contribute to the increased risk of MAYV transmission [25]
In the case of viral infection, variations in the habitats and behaviors of viruses have been noted in recent decades, such as the spread of MAYV by infected birds or humans, resulting in a transmission cycle that may involve humans as a reservoir [26].
Considering the lack of licensed vaccines and proven therapeutics, prevention focuses mainly on domestic mosquito control to contain the spread of MAYV [27].
2.2. Eastern Equine Encephalitis Virus (EEEV)
The encephalitis manifests itself with severe neurological conditions in humans and horses and has high lethality [28,29]. In Brazil, the isolations of animal cases had been registered at the present decade [30]. EEEV was isolated from horses in the states of Bahia, Pernambuco, Pará, São Paulo, Minas Gerais, Rio de Janeiro, Mato Grosso, and Paraná [31,32,33,34,35,36,37,38,39,40]. Currently, equine viral encephalomyelitis is sporadic in most Brazilian states [41].
Epidemiological studies carried out in the Brazilian Amazon were able to positively isolate EEEV [42]. Previously, in humans, positivity was restricted to seroepidemiological surveys, with no neurological clinical manifestations associated with the virus. Therefore, a fatal human infection was reported, and its occurrence was attributed to climate changes and human encroachment on previously wild areas [43].
The EEEV transmission cycle occurs between birds and mosquitoes, and the genera Aedes, Culex, and Mansonia have already been implicated in the transmission of EEEV among humans and horses, as they feed on birds and mammals [44]. Another form of transmission reported for this virus is by aerosol, which is of great concern due to the threat of its use as a possible bioterrorism agent [45].
2.3. Western Equine Encephalitis Virus (WEEV)
Western equine encephalitis is a serious neurological disease capable of affecting humans and horses and is widespread in Canada, Latin America and the USA. The first isolation of this virus occurred in 1938 [46] in the USA. In Brazil, WEEV has already been isolated in forests in Rio de Janeiro and Pará [47,48]. Known vectors belong to the genera Aedes and Culex, and viral isolation has also been obtained in chickens, pheasants, rodents, rabbits, ungulates, turtles and snakes and blood from wild birds [49].
The WEEV complex contains the following viruses: WEEV, AURAV, UNAV, TNTV, PIXV, and MUCV. Some viruses originally isolated in the Amazon region have not yet been fully characterized [11] to justify their inclusion in a taxon. AURAV and UNAV were isolated only from mosquitoes, mainly Aedes serratus and Psorophora ferox, respectively. MUCV was isolated from all four types of sources (humans, wild animals, sentinels and arthropods). PIXV was isolated twice from mosquitoes (Anopheles nimbus and Trichoprosopon digitatum) and once from a wild rodent (Proechimys guyannensis). AURAV, found in South America, has not been linked to any disease in humans or animals.
TNTV was isolated from rodents (Dasyprocta aguti and Orizomys sp.) and mosquitoes (Sabethes spp.) [44] in the Brazilian Amazon.
2.4. Chikungunya Virus (CHIKV)
Chikungunya fever received this name because of the language spoken in East Africa, where the first patients were treated; the term means ‘those who bend over’, which corresponds to the position adopted by the patient due to the severe pain caused by CHIKV. There are also reports of sudden-onset fever greater than 38.5 °C and severe arthralgia or arthritis with acute onset, which may also include headache, muscle pain and skin rash [50].
The first record of the disease in Brazilian territory was described in 2010 in the state of Rio de Janeiro as a case imported from Indonesia. However, from 2014 onward, there were reports of autochthonous infections caused by CHIKV in Amapá and Bahia, with a consequent increase in the number of cases [51].
The disease is transmitted by Ae. aegypti and Ae. albopictus mosquitoes, which are widespread in all Brazilian states and are widely dispersed in urban areas facilitating the transmission and spread and increase of CHIKV in Brazil [52]. As a result, it is necessary to maintain care regarding the epidemiological surveillance of CHIKV and invest in studies related to the development of vaccines and educational campaigns to combat vectors.
2.5. Pixuna Virus (PIXV), Mucambo Virus (MUCV), Una Virus (UNAV), Aura Virus (AURAV) and Triniti Virus (TNTV)
PIXV was isolated from arthropods in 1961 [34] in the state of Pará and from some human cases, one of which was related to a fatal laboratory infection [42], after which its isolation was not frequently reported; very little about this virus has been studied, with detection achieved only through serological records among horses in the state of Paraná [37] and among inhabitants of the Amazon region [53].
MUCV and many isolates from the Amazon are pathogenic to humans, and in this region, MUCV has been isolated from humans, sentinel animals, wild animals and mosquitoes. The Una, Aura and Triniti viruses were isolated from mosquitoes, and the TNTV was also isolated from a Oryzomys sp. rodent and maintains the ability to cause unknown human infection until the present [11].
3. Flavivirus Genus
The Flaviviridae family consists of four genera (Flavivirus, Pestivirus, Pegivirus and Hepacivirus) comprising 89 viral species [17], of which more than 50 species are transmitted by arthropods, infecting mosquitoes or ticks [54]. Mammals and birds can also be their hosts, which can present hemorrhagic fever, neurological disease or even be asymptomatic. Yellow fever virus (YFV), DENV, JEV, WNV and tick-borne encephalitis viruses are pathogenic to humans. Other members cause economically important diseases in domestic or wild animals. Other viruses in this group include SLEV, Rocio virus (ROCV), CACV, ILHV, BUSV and IGUV [7,55].
The viral particle measures 40 to 60 nm in diameter and has a protein capsid with icosahedral symmetry, a single-stranded RNA genome, and positive polarity, with approximately 11 kb in length [16]. Blood-sucking mosquitoes such as Ae. aegypti and Ae. albopictus are implicated as transmitters of flaviviruses. These are able to mutate and/or adapt to new zoonotic cycles, re-emerging in places that have suffered environmental impacts as a result of human action in nature [56].
3.1. Dengue Virus (DENV)
DENV is represented by four distinct serotypes: DENV-1 to DENV-4, which are maintained in cycles involving small primates and Aedes mosquitoes. In urban centers, this transmission occurs between mosquitoes and humans, with Ae. aegypti as the main vector, and Ae. albopictus and Ae. polynesiensis as secondary vectors. Ae. aegypti originated from the African continent and later spread to Asia and America by sea, arriving in Brazil in the mid-18th century by slave ships. The reports mentioned in the scientific literature mention the 1920s (20th century) as the beginning of epidemics that occurred in Brazilian territory, with the first reports occurring in the state of São Paulo in 1916 and Rio de Janeiro (Niterói) in 1923 [7].
In 1955, campaigns were carried out in an attempt to eradicate the Ae. aegypti vector in Brazil without success, causing viral circulation again in 1963 with the reemergence of DENV-2 and DENV-3 in several countries. At the end of the 1960s, the vector was already circulating in cities such as Belém, Salvador and Rio de Janeiro [57].
In the 1980s, there was an expansion of dengue throughout the Brazilian territory, with the gradual emergence of the four serotypes, which resulted in the emergence of arbovirus epidemics that are still observed in the country [58]. Regarding the epidemiological profile of dengue in Brazil between 2014 and 2019, dengue cases were predominant in the Southeast and Midwest regions, with a predominance of serotype 2 (DENV-2) [59].
Clinically, dengue manifests as an acute infectious disease characterized by fever, myalgia, headache, joint pain, retro-orbital pain, rash, thrombocytopenia, lymphadenopathy and leukopenia. Fever is sudden and high, persisting for 2 to 7 days. Rash and pruritus are evident in the convalescent phase of the disease [60].
The most severe clinical phase is called dengue hemorrhagic fever (DHF), which is characterized by increased vascular permeability and results in hypovolemic shock, which can lead to tissue damage and multiple organ failure [60].
In addition to the bite of Ae. aegypti mosquitoes, there are also reports of vertical transmission and blood transfusion transmission. Studies, especially in endemic areas such as the Amazon region, should be initiated in blood banks to determine the risk of infection for DENV and other flaviviruses for blood product recipients, especially in epidemic periods [61,62].
3.2. Saint Louis Encephalitis Virus (SLEV)
SLEV is transmitted by mosquitoes of the Culex genus, and birds are considered amplifying hosts found in urban and rural environments. On the other hand, men and domestic animals are hosts that show symptoms but do not develop significant viremia [63].
Reservoirs of this virus mainly include birds, primates, marsupials and other wild animals. The first record of the isolation of this virus was in the USA from an insectivorous bat, and in Brazil [64], isolation was carried out in 1960 from Sabethes belisarioi mosquitoes collected on the Belém-Brasília highway [65].
In Brazil, only three cases of SLEV in humans were reported in the 1970s, two in the northern region and a third identified in the municipality of São Pedro/SP in 2004 [65]. More recently, the detection of SLEV was described in humans in the states of São Paulo and Mato Grosso during outbreaks of DENV and ZIKV [66].
Most reported cases of SLEV go unnoticed because they are asymptomatic, subclinical, or have symptoms similar to those of a common cold. The main signs and symptoms observed include fever, headache, myalgia, nausea/vomiting and drowsiness [67].
3.3. West Nile Virus (WNV)
WNV was identified in the West Nile district of Uganda [69]. Over the years, it has also been observed in Africa, Europe, North America, the Middle East and Asia [70,71]. In Brazil, in 2009, antibodies against WNV were detected in horses in the state of Mato Grosso; in subsequent years, serological evidence continued to be detected in poultry, horses and humans [72,73,74]. Subsequently, another study carried out in the same region detected the presence of neutralizing antibodies not only against WNV but also against other flaviviruses in horses, sheep and alligators, showing wide circulation of these viruses in the region.
However, it was only in 2018 that the virus was identified and isolated from the central nervous system (CNS) of horses with neurological disease in the state of Espírito Santo [75,76]. Additionally, the identification of positive cases among humans and horses in the states of Piauí, São Paulo and Minas Gerais indicated the need to expand the study and epidemiological surveillance of WNV in the country [77,78,79]. Equine surveillance by veterinarians is essential to provide constant information about the circulation of WNV in the national territory [79].
Symptoms vary in each host, ranging from the asymptomatic form, common in horses and humans, to neurological manifestations in birds such as disorientation, ataxia, tremors and head tilt, followed by death. Approximately 20% of humans who develop symptoms have nonspecific clinical signs such as fever, body pain and myalgia, and a smaller number of those affected may have the neuroinvasive form, characterized by meningitis, encephalitis and paralysis progressing to death [80,81].
WNV transmission is maintained in an enzootic cycle, that is, between arthropods and wild birds, but it can also infect horses and mammals, including humans, the latter being considered accidental and final hosts because they develop a low viral load, which does not allow them to be involved in the transmission cycle. Other forms of transmission can be transovarian, which favors the permanence of the virus in mosquitoes, provided that there are favorable conditions for its maintenance and consequent transfer to progeny [80,82]; organ transplantation; placental route; and blood transfusion [83,84,85,86].
In 2002, the first case of suspected WNV infection after an obstetric procedure in a patient who received blood and blood products was reported [87]. The worldwide prevalence of arboviruses has grown dramatically in recent decades, and in areas where these diseases are endemic, transmission by transfusion has rarely been investigated. Concerns about the transmission of arboviruses by blood transfusion increased after the documentation of the transmission of WNV by transfusion in the USA. In Brazil, studies indicated that asymptomatic individuals can be blood donors and serve as a source of virus dissemination in the community [61].
3.4. Cacipacoré Virus (CACV), Ilhéus Virus (ILHV) and Bussuquara Virus (BUSV)
CACV was first isolated from birds (Formicarius analis) in 1977 in an Amazonian area in the state of Pará. The presence of antibodies against this arbovirus was detected in free-living monkeys captured in the state of Mato Grosso do Sul, with two animals showing antibodies against flaviviruses and one showing a monotypic reaction against CACV. More recent studies reported the isolation of CACV from mosquitoes collected in the Amazon region and from human blood in the state of Rondônia [88,89].
ILHV was isolated from a pool of mosquitoes of the genera Aedes and Psorophora during an epidemiological investigation of yellow fever in the city of Ilhéus, state of Bahia, in 1947 [90]. Birds are the main vertebrate hosts, and humans are considered accidental and terminal hosts, as they develop low viremia. ILHV was detected in invertebrates (Aedes genus mosquitoes) and vertebrates in the Pantanal region and in the state of Pará, São Paulo, respectively [7,91,92].
BUSV was originally isolated from primate blood near the city of Belém in 1959 [93]. The transmission cycle occurs between mosquitoes of the Culex genus and rodents, although birds can act as reservoirs. In 1971, it was reported the first case of a human febrile illness by BUSV, detected in Panamá [94]. In 2015, sera from nonhuman primates from Goiânia, in the Brazilian Midwest, were positive for BUSV [95].
The Evandro Chagas Institute (IEC) was created in 1936, with the name of North Institute for Experimental Pathology Institute, by the state government of Pará, and in 1942, it was federalized, becoming a scientific research body of the Special Service Foundation for Public Health. In 1954, with the support of the Rockefeller Foundation, studies on arboviruses in the Amazon region were hosted at the IEC, which brought innovations in field and laboratory procedures that have contributed in a decisive and invaluable way to public health and the scientific community during these five decades.
The objective of this work was to present the encephalitogenic viruses that have been registered in the Department of Arbovirology and Hemorrhagic Fevers (SAARB) of the IEC over the past six decades.
4. Viral Isolation
4.1. Isolation in Mice
For viral isolation, samples such as blood, serum, suspensions of human or animal viscera, and arthropods were used. The aforementioned samples (viscera and arthropods) were ground in 1000 μL of diluent buffer (5% fetal bovine serum (FBS)), penicillin (100 U/ml), streptomycin (100 μg/ml) and gentamicin (0.05 mg/ml) in TissueLyser equipment (Qiagen, Valencia, CA) using 5-mm diameter “beads” (stainless steel microspheres) at 26,000 movements per minute for 4 minutes, followed by freezing at -70 °C and later centrifugation at 10,000 rpm for 10 minutes at 4 °C. Then, each sample was centrifuged at 15,652 g for 10 minutes at 4 °C. After centrifugation, the samples remained at that temperature until inoculation into a newborn Swiss mouse of 2 to 3 days of life. The animals were inoculated intracerebrally with a volume of 0.02 mL of the suspensions with the aid of a 1-mL syringe and a 25x6 disposable needle. This inoculation step was also valid for the other biological samples. The cages that contained the animals that were to be used were identified with batch numbers and internal records of the sample, material type and inoculation date. They contained a lactating female and 6 newborn Swiss albino mice (two days old) [96].
After inoculation, the animals were observed daily for up to 21 days, and records were made on a card (inoculation card).
Mice that showed signs of disease were collected and stored in a freezer at -70 °C until laboratory tests were carried out, while those that did not show signs were disposed of following institutional protocols.
4.2. Cell Culture
The Laboratory of Viral Isolation/SAARB/IEC have its own Vero (renal cells of Chlorocebus aethiops) and C6/36 (Ae. albopictus) cells which were constantly used during these decades.
Vero cells were maintained in Medium 199 supplemented with glutamine (2 mM), sodium bicarbonate (3 mM), penicillin/streptomycin (104 IU/mL), fungizone (2.5 mg/mL) and 10% FBS for cell growth, with the amount of FBS being reduced to 5% for cell maintenance. The cells were trypsinized every 7 days, and the medium was replaced every 2 days and kept at 37 °C under a humid atmosphere of 95% H2O and 5% CO2. For the C6/36 cells, L-15 medium was used, trypsinization was performed manually, and the cells remained in a room with a temperature of approximately 25 °C [97].
Briefly, 100 µL of each diluted viral sample were inoculated into monolayers of the aforementioned cells propagated in 16x125-mm tubes and then incubated at 37 °C and 28 °C for a period of one hour (adsorption), and then 1.5 ml of overlay medium (maintenance medium 199 and L-15) were added to the cells. Cell cultures were observed daily to monitor possible cytopathogenic effects (CPEs) with the aid of an Axiovert S100 microscope (Zeiss). Negative controls were made from the inoculation of uninfected cell fluids. The observation of infected cultures was performed with the aid of an Axiovert S100 microscope (Zeiss).
4.3. Origin of Viral Isolates
The samples registered by the SAARB/IEC from 1954 to 2012 correspond to 665 flaviviruses and 739 alphaviruses from the state of Pará (Figure 2, Figure 3 and Figures S1 and S2) from forested areas in the region, such as the Utinga Forest and Uriboca, around highways such as Belém-Brasília, Transamazônica, and Santarém-Cuiabá, a region where projects are developed by the IEC, such as the Jari project and the localities of the Guamá ecological research area (APEG), Bosque Rodrigues Alves, the airport road, Guamá colony, farms (Matinha, Oriboca-Pirelli, Piramucal), the Northern Agronomic Institute (IAN), Northern Agricultural Research and Experimentation Institute (IPEAN), Pará [Abaetetuba, Afuá, Água Preta, Alenquer, Altamira, Barcarena, Belém, Benevides, Bragança, Breves, Cachoeira Porteira (Cacipacoré, Oriximiná), Castanhal (Alto do Capim), Catu, Faro, Itaituba, Marabá, Marajó (Ponta de Pedras), Medicilândia, Monte Alegre, Mucambo, Tucuruí, Ourém, Pacajá, Parauapebas, Manatee, Portel, Santa Bárbara, Bragança, Santarém] and other states, Amapá (Serra do Navio, ICOMI), Mato Grosso (Aripuanã), Acre (Boca do Acre), Goiás (Uruaçu), Boa Vista (Roraima), Brasilia, Fortaleza (Ceará), Goiânia (Goiás), Tocantins, Macapá (Amapá), Bom Jardim (Maranhão), Bahia Institute of Biology (IBIO/UFBA), Manaus (Amazonas), Rio Grande do Sul, Mato Grosso, Mato Grosso do Sul, Bahia, Paraná, Minas Gerais, Curitiba and the city of Iquitos (Peru).
5. Alphavirus and Flavivirus Collection
5.1. Isolated Samples from Mice
Between 1938 and 2012, 1,090 samples of arboviruses with encephalitogenic potential were isolated from newborn Swiss mice, of which 370 were flaviviruses [14 DENV, 202 SLEV, 109 BUSV, 44 ILHV, and 1 CACV] and 720 were togaviruses [39 VEEV, 211 EEEV, 13 WEEV, 155 MAYV, 4 PIXV and 236 MUCV]. In addition, a total of 7 togaviruses and 4 flaviviruses were partially identified. In both viral families studied, isolation was achieved in animals, arthropods and humans, with declining success. Approximately 293 viruses are in the families Togaviridae (9), Rhabdoviridae (3), Sedoreoviridae (4), Picornaviridae (21), Flaviviridae (4), Peribuniviridae (225) and Arenaviridae (27) with incomplete characterization; eight are partially identified and therefore have unknown pathogenesis: Iriri virus, Uruará virus, Galibi virus, Codajás virus, Cajazeira virus, Marajó virus, Cantá virus, Tracambé virus, and Naranjal virus (NJLV).
Regarding the 659 samples isolated from Sabethes sp., T. digitatum, Aedes sp., Mansonia, Psorophora, Ae. scapularis, Ae. serratus, Ae. fulvus, Ae. oligopistus, Ae. sexlineatus, Ae. septemstriatus, Ae. argyrothorax, Ae. aegpypti, Anopheles nuneztovari, Anopheles species, Anopheles triannulatus, Anopheles nimbus and Culex melanoconion mosquitoes, it was possible to detect BUSV, MUCV, ILHV, AURAV, DENV-2, EEEV, MAYV, PIXV, SLEV, TNTV, Trocara virus, UNAV, and WEEV.
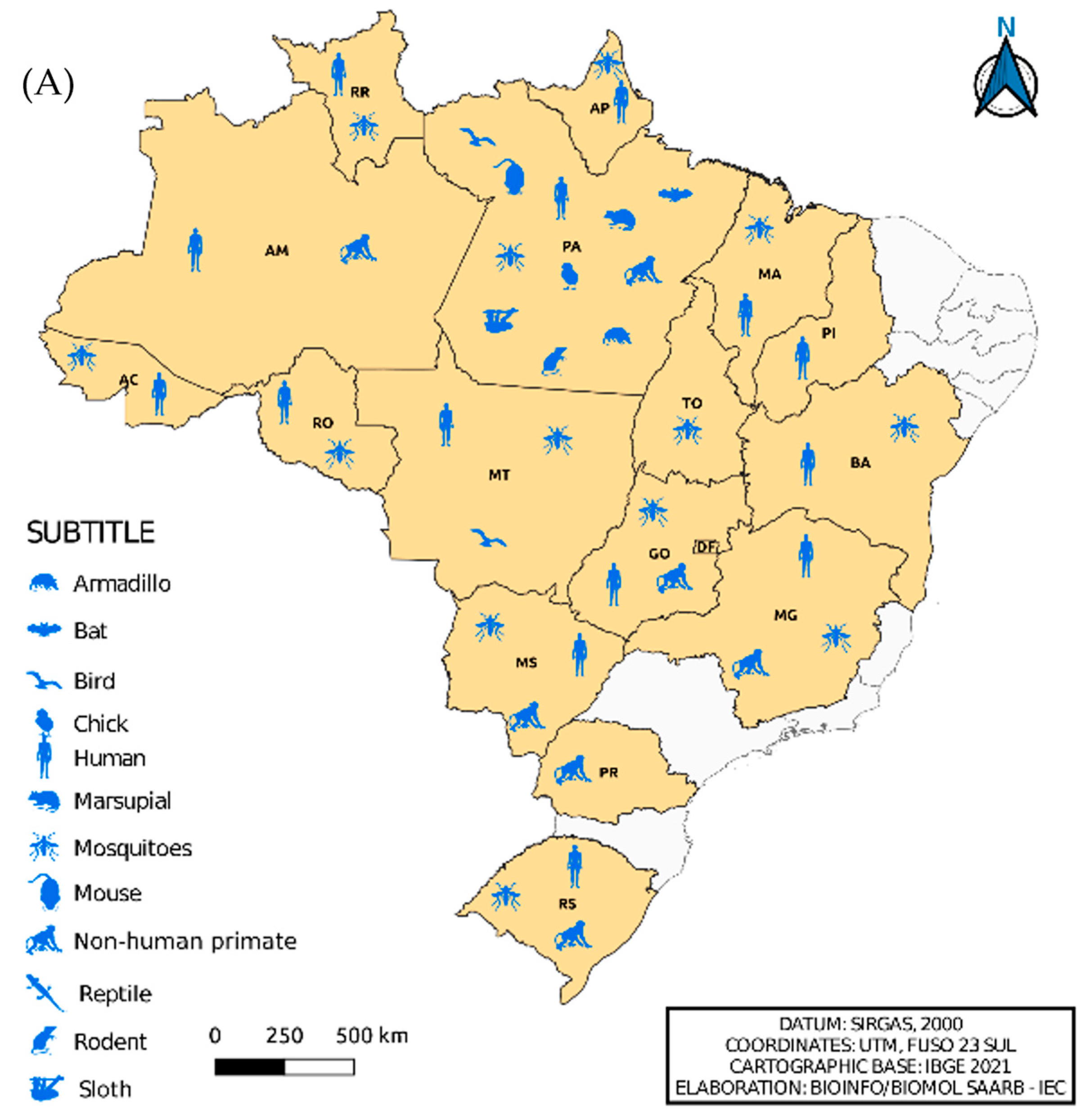
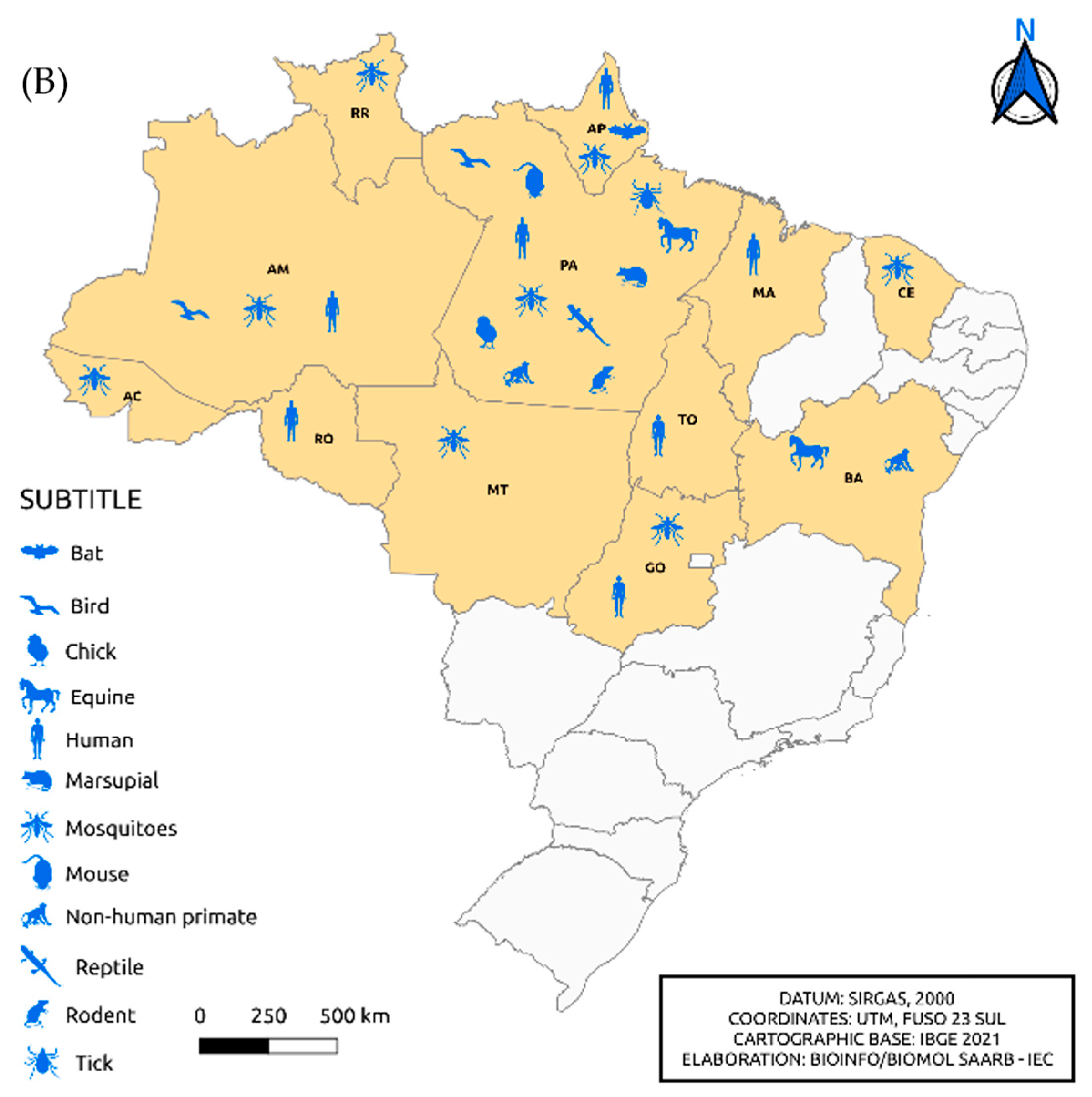
Figure 4 shows the isolation of viruses belonging to the Flavivirus and Alphavirus genera according to the distribution by federative unit and organism in which the isolation occurred, respectively. In the north region, specifically in the state of Pará, the isolation of flaviviruses in armadillos and sloths and alphaviruses in ticks and horses was observed, differing from other Brazilian states. We also noted viral isolation in horses from the state of Bahia. When comparing Figure 4A,B, it was remarkable to note the absence of alphavirus isolation in the states of Minas Gerais, Mato Grosso, Paraná, Rio Grande do Sul and Piauí.
5.2. Isolated Samples in Cell Culture
From a total of 7,054 samples intended for cell culture, it was possible to isolate 442 arboviruses with neuroinvasive potential from 2015 to 2022, with 191 DENV-1, 93 DENV-2, 82 DENV-4, 64 CHIKV and 14 ZIKV. The isolates from mosquitoes totaled 323 viruses, including ILHV (26), UNAV (35), TNTV (16), SLEV (60) and EEEV (239), in the period from 1955 to 2022.
6. Legacy and Future Concerns
The growing deforestation in the 16th century was associated with the challenge of eradicating arbovirus-transmitting vectors and the uncertain diagnosis of these viruses, whether due to antigenic similarity, causing cross-reactions (in arbovirus-endemic areas) in laboratory tests or similarity of symptoms among patients, constituting old and recurring problems in public health [8,98]. Concomitantly, disorderly urbanization, with the accumulation of garbage in urban areas, favors the proliferation of arbovirus vectors and contributes to the emergence and/or re-emergence of diseases in Brazilian territory, especially arboviruses caused by flaviviruses and alphaviruses [99].
Many of the arboviruses isolated by the IEC in epidemiological research on the circulation of arboviruses, especially in the Amazon region, led to a vast biological collection, with approximately 10,000 arboviruses, including those unknown to science, many of which still have incomplete taxonomic characterization [100].
Over all these years, the information generated by the SAARB/IEC has revealed ecological, environmental and epidemiological information about arboviruses. Studies of this nature become relevant, even more so in the Amazon region, as it has favorable conditions for the maintenance cycle of these agents, demonstrating an ideal environment for the emergence or re-emergence of arboviruses [96].
Clinical manifestations may be absent at the time of infection, which causes the free movement of individuals, resulting in viral spread, even to nonendemic regions, a situation that imposes the need for constant surveillance of arboviruses [101,102]. This incessant work in public health helps to promote the detection and knowledge of the circulation of viruses, thus enabling the identification of the potential agents responsible for outbreaks and epidemics; the prevention of arboviruses and vector control are the most effective measures in the current scenario where there is no vaccine or specific treatment available for most arboviruses [2].
In view of the epidemiological scenario demonstrating the simultaneous circulation of arboviruses with encephalitogenic potential that are of public health importance, in 2017, the Ministry of Health established changes in the Sentinel Surveillance Manual for Arbovirus Neuroinvasive Diseases, with the purpose of expanding the surveillance of neuroinvasive diseases caused by arboviruses [2].
In view of what was observed in this study, it was possible to conclude that the isolation of ZIKV coincided with the first reports published by the Pan American Health Organization (PAHO), leaving physical and emotional sequelae in the affected population, especially among women of childbearing age due to congenital infections that resulted in fetal malformations and deaths [103,104].
In view of the above, the need for constant epidemiological surveillance has become fundamentally important to investigate the circulation of arboviruses, especially in the Amazon region, which has peculiar characteristics, many of which are still unknown to man, remaining a true hotspot for known viruses and for those completely new to science.
With over 80 years of experience, the IEC has collaborated with the Ministry of Health in reporting outbreaks and epidemics, whether by reporting cases, implementing training programs, or developing human resources that will contribute to the reduction in arbovirus cases [102].
We reiterate the concern regarding the need for urgent actions to combat forest degradation that influences the spread of diseases transmitted by arthropods and the adaptation to the domestic environment and the possibility of emerging or re-emerging zoonoses [105].
Regarding the data presented in this review, it is of great importance to understand that health education constitutes a mechanism that should be seen as a need of the population to contribute to behaviors associated with prevention.
Currently, for most arboviruses, there is no vaccine available for the diseases discussed in this article, and the cycle of sylvatic transmission of viruses cannot be prevented. Thus, preventive measures are necessary, such as avoiding contact with the areas of occurrence and minimizing exposure to vector bites, either by means of individual protection (use of repellents and full-length clothes) or collective protection (use of curtains and mosquito nets), especially in rural and wild areas, in addition to avoiding exposure in affected areas; this is performed with the aim of reducing contact between man and vector. Therefore, it is recommended to avoid exposures in forested areas and riverbanks, especially at times when the vector is most active [2].
The data presented in the present study reinforce the constant need for epidemiological surveillance, which provides support to prevent the spread of arboviruses with neuroinvasive potential.
There are numerous arboviruses present in different environments worldwide, especially in the Amazon, due to the favorable conditions found there; the emergence of more virulent forms of these viruses are constantly emerging and invading new habitats. Therefore, knowing the epidemiology of these arboviruses is of fundamental importance to predict future mechanisms of emergence and/or re-emergence of the same and thus implement preventive measures as well as control and prevent the spread of arboviruses in urban areas. In addition to encephalitogenic viruses, there are many arboviruses with little information available in the literature [1,2,3]. Therefore, it is only a matter of time before we see the emergence of these viruses with a more virulent phenotype that can spread to a susceptible population that has no immunological memory for these viruses. The ZIKV pandemic is a recent classic example that showed us the power of arbovirus dispersion and, most importantly, its association with congenital malformations associated with ZIKV infection [103,104]. Unfortunately, the prevention of infection caused by the majority of arboviruses depends almost exclusively on efforts to control the populations of potential vectors, whether through the use of repellents or protective clothing and mainly by raising the awareness of the population to reduce outbreaks at breeding sites. Despite great efforts to develop vaccines against arboviral infections, especially those that cause severe encephalitis, only a few have been successful [106].
The search for vaccines is a necessity, and several live attenuated vaccines based on modern molecular biology techniques are in full pharmaceutical development. Some, such as the ChimeriVax candidates developed against WNV, Japanese encephalitis and dengue, have already shown good results in terms of safety and immunogenicity [106]. Our collection shows the great viral diversity that has already been isolated in our country, which can contribute to future possibilities for the development of vaccines as well as for the development of promising antiviral drugs aimed at isolated and known viruses, which represents another aspect that needs progress in research, since effective and specific antiviral treatments aimed at arboviruses of public health interest are still lacking.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Distribution of flaviviruses isolated by the IEC in the Pará state and other federal units in Brazil; Figure S2: Distribution of alphaviruses isolated by the IEC in the Pará state and other federal units in Brazil.
Author Contributions
Conceptualization: Pinto, E.V. Methodology: Barros, L.J.L.; Freitas, M.N.O.; Santos, M.M.; Gonçalves, E.J.; Pantoja, J. Software: Silva, F.S. Validation: Silva, F.S. Formal Analysis: Hernández, L.H.A. Investigation: Lima, C.S.; Lima, M.F.; Costa, L.R.O.; Silva, I.F.; Neto, J.P.N; Cunha, T.C.A.S; Nascimento, B.L.S. Resources: Evandro Chagas Institute Writing - Original Draft Preparation: Wanzeller, A.L.M.; Pinto, E.V. Writing – Review & Editing: Pinto, E.V Visualization: Azevedo, R.S.S.; Carvalho, V.L.; Chiang, J.O Supervision: Pinto, E.V. Project Administration: Wanzeller, A.L.M Data Curation: Travassos da Rosa, E.S.; Rodrigues, S.G.; Vasconcelos, H.B.; Vasconcelos, P.F.C.; Cruz, A.C.R. Funding Acquisition: Evandro Chagas Institute.
Funding
Evandro Chagas Institute
Acknowledgments
Thank you to the technicians and researchers who collaborated during all these decades in isolating arboviruses at SAARB/IEC: MONTEIRO, H.O. (in memoriam); CASTRO, F.S.; TRAVASSOS DA ROSA, A.P.A. (in memoriam); CARVALHO, C.L.; BUNA S.B.; PINHEIRO, F.P.; and BENSABATH, G. (in memoriam).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- WHO (World Health Organization). 1985. Arthropod-borne and rodent-borne viral diseases. Technical Report Series, n. 719. Geneva: World Health Organization.
- Brazil. Ministry of Health. Manual de Vigilância Sentinela de Doenças Neuroinvasivas por Arbovírus; Ministry of Health: Brasília, Brazil, 2017. [Google Scholar]
- CDC (Centers for Disease Control and Prevention). Arboviral diseases, neuroinvasive and non-neuroinvasive. Available online: https://ndc.services.cdc.gov/case-definitions/arboviral-diseases-neuroinvasive-and-non-neuroinvasive-2015/ (accessed on 17 March 2022).
- Wilder-Smith, A.; Gubler, D.J.; Weaver, S.C.; Monath, T.P.; Heymann, D.L.; Scott, T.W. Epidemic arboviral diseases: priorities for research and public health. Lancet Infect Dis. 2017, 17, e101–e106. [Google Scholar] [CrossRef]
- WHO (World Health Organization). 2009. Dengue: guidelines for diagnosis, treatment, prevention and control. Geneva: World Health Organization.
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. New Eng J Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Lopes, N.; Nozawa, C.; Linhares, R.E.C. Características gerais e epidemiologia dos arbovírus emergentes no Brasil. Rev Pan-Amaz Saude 2014, 5, 55–64. [Google Scholar] [CrossRef]
- Donalisio, M.R.; Freitas, R.R.; Von Zuben, A.P.B. Arboviruses emerging in Brazil: challenges for clinic and implications for public health. Rev. Saude Publica 2017, 51, 1–6. [Google Scholar] [CrossRef]
- Pimentel, V.; Afonso, R.; Nunes, M.; Vieira, M.L.; Bravo-Barriga, D.; Frontera, E.; Martinez, M.; Pereira, A.; Maia, C.; Paiva-Cardoso, M.N.; et al. Geographic dispersal and genetic diversity of tick-borne phleboviruses (Phenuiviridae, Phlebovirus) as revealed by the analysis of L segment sequences. Ticks Tick Borne Dis. 2019, 10, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis, N.; Tesh, R.B.; Popov, V.L.; Widen, S.G.; Wood, T.G.; Forrester, N.L.; Gonzalez, J.P.; Saluzzo, J.F.; Alkhovsky, S.; Lam, S.K.; et al. Exploiting the Legacy of the Arbovirus Hunters. Viruses 2019, 11, 471. [Google Scholar] [CrossRef] [PubMed]
- Travassos da Rosa, J.F.S.; Travassos da Rosa, A.P.A.; Vasconcelos, P.F.C.; Pinheiro, F.P.; Rodrigues, S.G.; Travassos da Rosa, E.S.; Dias, L.B.; Cruz, A.C.R. Arboviruses isolated in the Evandro Chagas Institute, including some described for the first time in the Brazilian Amazon region, their know hosts, and their pathology for man. In An Overview of Arbovirology in Brazil and Neighbouring Countries; Travassos da Rosa, A.P., Vasconcelos, P.F.C., Travassos da Rosa, J.F.S., Eds.; Instituto Evandro Chagas: Belém, Brazil, 1998; pp. 19–31. [Google Scholar]
- Chen, R.; Mukhopadhyay, S.; Merits, A.; Bolling, B.; Nasar, F.; Coffey, L.L.; Powers, A.; Weaver, S.C.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Togaviridae. J Gen Virol. 2018, 99, 761–762. [Google Scholar] [CrossRef] [PubMed]
- Lwande, O.W.; Lutomiah, J.; Obanda, V.; Gakuya, F.; Mutisya, J.; Mulwa, F.; Michuki, G.; Chepkorir, E.; Fischer, A.; Venter, M.; et al. Isolation of tick and mosquito-borne arboviruses from ticks sampled from livestock and wild animal hosts in Ijara District, Kenya. Vector Borne Zoonotic Dis. 2013, 13, 637–642. [Google Scholar] [CrossRef]
- Hayes, R.O.; Francy, D.B.; Lazuick, J.S.; Smith, G.C.; Gibbs, E.P.J. Role of the Cliff Swallow Bug (Oeciacus vicarius) in the Natural Cycle of a Western Equine Encephalitis-Related Alphavirus. J Med Entomol. 1977, 14, 257–262. [Google Scholar] [CrossRef]
- Kuhn, R.J. Togaviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, USA, 2013; pp. 629–650. [Google Scholar]
- Simmond, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J Gen Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef]
- ICTV (International Committee on Taxonomy of Viruses). Currently ICTV Taxonomy Release. Available on: https://talk.ictvonline.org/ taxonomy/ (accessed on 04 April 2022).
- Pinheiro, F.P.; Freitas, R.B.; Travassos da Rosa, J.F.; Gabbay, Y.B.; Mello, W.A.; LeDuc, J.W. An outbreak of Mayaro virus disease in Belterra, Brazil. I. Clinical and virological findings. Am J Trop Med Hyg. 1981, 30, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, P.F.C.; Travassos da Rosa, A.P.A.; Rodrigues, S.G.; Rosa, E.S.T.; Dégallier, N.; Rosa, J.F.S.T.; et al. Inadequate management of natural ecosystem in the Brazilian Amazon region results in the emergence and reemergence of arboviruses. Cad. Saude Publica 2001, 17, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, T.L.M.; Santos, C.L.S.; Suzuki, A.; Petrella, S.M.C.; Bisordi, I.; Nagamori, A.H.; Marti, A.T.; Santos, R.N.; Fialho, D.M.; Lavigne, S.; et al. Mayaro virus: imported cases of human infection in São Paulo State, Brazil. Rev Int Med Trop. 2007, 4, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, R.S.; Silva, E.V.; Carvalho, V.L.; Rodrigues, S.G.; Nunes-Neto, J.P.; Monteiro, H.; Peixoto, V.S.; Chiang, J.O.; Nunes, M.R.; Vasconcelos, P.F. Mayaro Fever Virus, Brazilian Amazon. Emerg Infect Dis. 2009, 15, 1830–1832. [Google Scholar] [CrossRef] [PubMed]
- Zuchi, N.; Heinen, L.B.; Santos, M.A.; Pereira, F.C.; Slhessarenko, R.D. Molecular detection of Mayaro virus during a dengue outbreak in the state of Mato Grosso, Central-West Brazil. Mem Inst Oswaldo Cruz 2014, 109, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.P.S.; Chiang, J.O.; Ferreira, M.S.; Henriques, D.F.; Oliveira, C.F.; Carvalho, V.L.; Silva, E.V.P.; Azevedo, R.S.S.; Martins, L.C. Research on the circulation of arboviruses in human populations living in the Municipalities of Parauapebas and Canaã de Carajás, located in the southeastern mesoregion of the state of Pará. Research, Society and Development 2022, 11, e6211326043. [Google Scholar] [CrossRef]
- Pereira, N.T.; Rocha, M.N.; Sucupira, P.H.F.; Carvalho, F.D.; Moreira, L.A. Wolbachia significantly impacts the vector competence of Aedes aegypti for Mayaro virus. Sci Rep 2018, 10, 6889. [Google Scholar] [CrossRef] [PubMed]
- Theilacker, C.; Held, J.; Allering, L.; Emmerich, P.; Schmidt-Chanasit, J.; Kern, W.V.; Panning, M. Prolonged polyarthralgia in a German traveller with Mayaro virus infection without inflammatory correlates. BMC Infect Dis. 2013, 13, 369. [Google Scholar] [CrossRef]
- Neufeld, P.M. About Mayaro Fever: an emergin arbovirus. Rev Bras Anal Clin. 2017, 49, 118–119. [Google Scholar] [CrossRef]
- Diagne, C.T.; Bengue, M.; Choumet, V.; Hamel, R.; Pompon, J.; Missé, D. Mayaro Virus Pathogenesis and Transmission Mechanisms. Pathogens 2020, 9, 738. [Google Scholar] [CrossRef]
- Barba, M.; Fairbanks, E.L.; Daly, J.M. Equine viral encephalitis: prevalence, impact, and management strategies. Vet Med (Auckl). 2019, 10, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Beer, J. Doenças infecciosas em animais domésticos. Roca: São Paulo, Brazil, 1999; ISBN 978-8572412483.
- Gil, L.H.V.G.; Magalhaes, T.; Santos, B.S.A.S.; Oliveira, L.V.; Oliveira-Filho, E.F.; Cunha, J.L.R.; Fraiha, A.L.S.; Rocha, B.M.M.; Longo, B.C.; Ecco, R.; et al. Active Circulation of Madariaga Virus, a Member of the Eastern Equine Encephalitis Virus Complex, in Northeast Brazil. Pathogens 2021, 10, 983. [Google Scholar] [CrossRef] [PubMed]
- Alice, F.J. Encefalomielite equina na Bahia, estudo de três amostras isoladas. Rev Bras Biol. 1951, 11, 125–144. [Google Scholar]
- Cunha, R. Estudos sobre uma amostra de vírus da encefalomielite equina isolada de material proveniente de Recife. Bol Soc Bras Med Vet. 1954, 14, 201–215. [Google Scholar]
- Causey, O.R.; Shope, R.E.; Laemmert, H.W. Report of an epizootic of encephalomyelitis virus in Pará, Brazil. Rev Serv Esp Saude Publica. 1962, 12, 47–50. [Google Scholar]
- Correa, W.M.; Correa, C.N.M. Encefalomielite equina. In Enfermidades infecciosas dos mamíferos domésticos, 2nd ed. MEDSI: Rio de Janeiro, Brazil, 1992; pp. 635–642.
- Iversson, L.B.; Silva, R.A.M.S.; Travassos da Rosa, A.P.A.; Barros, V.L.R.S. Circulation of Eastern Equine Encephalitis, Western equine encephalitis, Ilhéus, Maguari and Tacaiuma viruses in equines of the Brazilian Pantanal, South America. Rev Inst Med Trop. 1993, 35, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.R.; Sugay, W. Ocorrência da encefalomielite equina em Itaporanga, Estado de São Paulo. Arq Inst Biol. 1962, 29, 63–68. [Google Scholar]
- Campos, K.F.; de Oliveira, C.H.S.; Reis, A.B.; Vamasaki, E.M.; Brito, M.F.; Andrade, S.T.J.; Duarte, M.D.; Barbosa, J.D. Surto de encefalomielite equina Leste na Ilha de Marajó, Pará. Pesq Vet Bras. 2013, 33, 443–448. [Google Scholar] [CrossRef]
- Casseb, A.R.; Casseb, L.M.N.; Silva, S.P.; Vasconcelos, P.F.C. Abovírus: Importante zoonose na Amazônia Brasileira. Vet e Zootec. 2013, 20, 391–403. [Google Scholar]
- Fernández, Z.; Richartz, R.; Travassos da Rosa, A.P.A.; Soccol, V.T. Identification of the encephalitis equine virus, Brazil. Rev Saude Pública. 2000, 34, 232–235. [Google Scholar] [CrossRef]
- Wigg, M.D. Isolamento de uma amostra de vírus WEE em Haemagogous janthinomys. Master’s dissertation, Rio de Janeiro Federal University, Rio de Janeiro, Brazil, 1977.
- Heinemann, M.B.; Souza, M.C.C.; Cortez, A.; Ferreira, F.; Homem, V.S.F.; Ferreira-Neto, J.S.; Soares, R.M.; Cunha, E.M.S.; Richtzenhain, L.J. Soroprevalência da encefalomielite equina do leste e do oeste no Município de Uruará, PA, Brasil. Brazilian Journal of Veterinary Research and Animal Science 2006, 43, 137–139. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C.; Travassos da Rosa, J.F.S.; Travassos da Rosa, A.P.A.; Dégallier, N.; Pinheiro, F.D.P. Epidemiologia das encefalites por arbovírus na Amazônia brasileira. Rev Inst Med Trop S Paulo. 1991, 33, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Dexter, E.P.; Dexter, D.D.; Lindsay, C.W.; Ross, R.R.; Lutwick, L. Case of fatal eastern equine encephalitis. ID Cases 2021, 26, e01288. [Google Scholar] [CrossRef]
- Hervé, J.P.; Dégallier, N.; Travassos da Rosa, A.P.A.; Pinheiro, F.P.; Sá Filho, G.C. Arboviroses: aspectos ecológicos. In Instituto Evandro Chagas: 50 Anos de Contribuição às Ciências Biológicas e à Medicina Tropical. Fundação SESP: Belém Brazil, 1986; pp. 409–437.
- Rust, R.S. Human arboviral encephalitis. Semin Pediatr Neurol. 2012, 19, 130–151. [Google Scholar] [CrossRef] [PubMed]
- Acha, P.N.; Szyfres, B. Zoonosis y enfermedades transmisibles comunes al hombre y a los animales. Volumen 1: Bacteriosis y Micosis, 3rd ed. Pan American Health Organization: Washington, USA, 2003.
- Bruno-Lobo, G.; Bruno-Lobo, M.; Travassos, J.; Pinheiro, F.; Pazin, I.P. Estudos sobre arbovírus. III. Isolamento de vírus sorologicamente relacionado ao sub-grupo Western - Sindbis de um caso de encefalomielite equina no Rio de Janeiro. An Microbiol. 1961, 9, 183–195. [Google Scholar]
- Jonkers, A.H.; Downs, W.G.; Spence, L.; Aitken, T.H.G. Arthropod borne encephalitis viruses in Northern South America. II. A serological survey of northeastern Venezuela. Am J Trop Med Hyg. 1965, 14, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Kenney, J.L.; Deardoff, E.; Wang, E.; Weaver, S.C. Western Equine Encephalitis submergence: lack of evidence for a decline in virus virulence. Virology 2008, 380, 170–172. [Google Scholar] [CrossRef] [PubMed]
- BVS (Biblioteca Virtual em Saúde). Febre Chikungunya. Available online: https://bvsms.saude.gov.br/febre-de-chikungunya/ (accessed on 17 May 2022).
- Montalbano, C.A.; Bezerra, W.S.P.; Ribeiro, K.M.; Rosa, S.B.A. Doenças infecciosas de relevância no Brasil. Atena Editora: Ponta Grossa, Brazil, 2021, 340 pp. ISBN 978-655-983-610-9.
- Brazilian Ministry of Health. Boletim epidemiológico v. 54, n. 01, 2023. Available online: https://www.gov.br/saude/pt-br/centrais-de-conteudo/publicacoes/boletins/epidemiologicos/edicoes/2023/boletim-epidemiologico-volume-54-no-01/ (accessed on 07 February 2022).
- Cruz, A.C.R.; Vasconcelos, P.F.C. Arbovírus no Brasil. Biológico, 2008, 70, 45–46. [Google Scholar]
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A structural perspective of the Flavivirus life cycle. Nat Rev Microbiol. 2005, 3, 13–22. [Google Scholar] [CrossRef]
- Coimbra, T.L.; Nassar, E.S.; Nagamori, A.H.; Ferreira, I.B.; Pereira, L.E.; Rocco, I.M.; Ueda-Ito, M.; Romano, N.S. Iguape: a newly recognized flavivirus from São Paulo State, Brazil. Intervirology. 1993, 36, 144–152. [Google Scholar] [CrossRef]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antiviral Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed]
- Catão, R.C. Dengue no Brasil: abordagem geográfica na escala nacional. Cultura acadêmica: São Paulo, Brazil, 2012, 176 pp. ISBN 978-857-983-332-8.
- Bezerra, J.M.T.; Sousa, S.C.; Tauil, P.L.; Carneiro, M.; Barbosa, D.S. Entry of dengue virus serotypes and their geographic distribution in Brazilian federative units: a systematic review. Rev Bras Epidemiol. 2021, 24, E210020. [Google Scholar] [CrossRef] [PubMed]
- Oneda, R.M.; Basso, S.R.; Frasson, L.R.; Mottecy, N.M.; Saraiva, L.; Bassani, C. Epidemiological profile of dengue in Brazil between the years 2014 and 2019. Rev Assoc Med Bras. 2021, 67, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.K.; Bhattacharjee, S. Dengue virus: epidemiology, biology, and disease aetiology. Can J Microbiol. 2021, 67, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.L.; Amarilla, A.A.; Poloni, T.R.; Covas, D.T.; Aquino, V.H.; Figueiredo, L.T.M. Detection of dengue virus in sera of Brazilian blood donors. Transfusion 2012, 52, 1667–1671. [Google Scholar] [CrossRef] [PubMed]
- Slavov, S.N.; Santos, E.V.; Hespanhol, M.R.; Marta, R.; Rodrigues, E.S.; Haddad, R.; Ubiali, E.M.A.; Covas, D.T.; Kashima, S. Dengue RNA detection and seroprevalence in blood donors during an outbreak in the São Paulo State, Brazil, 2016. J Med Virol. 2021, 93, 3344–3349. [Google Scholar] [CrossRef] [PubMed]
- Kubizeski, J.R. Arboviroses emergentes no município de Sinop-MT: pesquisa de vetores. Bachelor’s dissertation, Mato Grosso Federal University, Sinop, Brazil, 2017.
- Lopes, S.F. Identificação de Flavivirus em aves silvestres da Amazônia Central. Master’s dissertation, Amazonas Federal University, Manaus, Brazil, 2011.
- Rocco, I.M.; Santos, C.L.; Bisordi, I.; Petrella, S.M.; Pereira, L.E.; Souza, R.P.; Suzuki, A. St. Louis encephalitis vírus: first isolation from a human in São Paulo state, Brasil. Rev Inst Med Trop S Paulo. 2005, 47, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Moraes, M.M.; Kubiszeski, J.R.; Vieira, C.J.S.; Gusmão, A.F.; Pratis, T.S.; Colombo, T.E.; Thies, S.F.; Araujo, F.C.; Zanelli, C.F.; Milhim, B.H.G.A.; et al. Detection of Saint Louis encephalitis virus in two Brazilian states. J Med Virol. 2021, 94, 776–781. [Google Scholar] [CrossRef]
- Lavezzo, L.C. Estudo de arboviruses em dodores de sangue na região Amazônica e em uma cidade do interior de São Paulo. Master’s dissertation, São Paulo State University, São Paulo, Brazil, 2010.
- Barbosa, B.C.M. Arbovírus emergentes no Brasil e seu risco de transmissão por transfusão sanguínea: uma revisão da literatura. Master’s dissertation, Minas Gerais Federal University, Belo Horizonte, Brazil, 2017.
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A Neurotropic Virus Isolated from the Blood of a Native of Uganda. Am J Trop Med Hyg. 1940, 20, 471–472. [Google Scholar] [CrossRef]
- Murgue, B.; Zeller, H.; Deubel, V. The ecology and epidemiology of West Nile virus in Africa, Europe and Asia. Curr Top Microbiol Immunol. 2002, 267, 195–221. [Google Scholar] [CrossRef]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile. Lancet Infect Dis. 2002, 2, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Melandri, V.; Guimarães, A.E.; Komar, N.; Nogueira, M.L.; Mondini, A.; Fernandez-Sesma, A.; Alencar, J.; Bosch, I. Serological detection of West Nile virus in horses and chicken from Pantanal, Brazil. Mem Inst Oswaldo Cruz. 2012, 107, 1073–1075. [Google Scholar] [CrossRef]
- Ometto, T.; Durigon, E.L.; de Araujo, J.; Aprelon, R.; de Aguiar, D.M.; Cavalcante, G.T.; Melo, R.M.; Levi, J.E.; de Azevedo Júnior, S.M.; Petry, M.V.; et al. West Nile virus surveillance, Brazil, 2008-2010. Trans R Soc Trop Med Hyg. 2013, 107, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Pauvolid-Corrêa, A.; Campos, Z.; Juliano, R.; Velez, J.; Nogueira, R.M.R.; Komar, N. Serological evidence of widespread circulation of West Nile virus and other flaviviruses in equines of the Pantanal, Brazil. PLoS Neg Trop Dis. 2014, 8, e2706. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.C.; Silva, E.V.P.; Casseb, L.M.N.; Silva, S.P.; Cruz, A.C.R.; Pantoja, J.A.S.; Medeiros, D.B.A.; Filho, A.J.M.; Cruz, E.R.M.; Araújo, M.T.F.; et al. First isolation of West Nile virus in Brazil. Mem Inst Oswaldo Cruz. 2019, 114, e180332. [Google Scholar] [CrossRef]
- Silva, A.S.G.; Rehfeld, I.S.; Santos, B.S.A.S.; Franklin, L.F.S.; Teixeira, R.B.C.; Lobato, Z.I.P.; Guedes, M.I.M.C.; Costa, E.A. Febre do Nilo Ocidental no Brasil: o novo desafio aos medicos-veterinários. Rev Educ Cont Med Vet Zootec. 2021, 19, e38082. [Google Scholar] [CrossRef]
- Costa, E.A.; Bayeux, J.J.M.; Silva, A.S.G.; de Queiroz, G.A.; Santos, B.S.A.S.; Rocha, M.N.; Rehfeld, I.S.; Franklin, L.F.S.; Valle, L.B. Epidemiological surveillance of West Nile virus in the world and Brazil: relevance of equine surveillance in the context of “One Health”. Brazilian Journal of Veterinary Research and Animal Science 2019, 56, e164335. [Google Scholar] [CrossRef]
- Meireles, A.R.; Fernandes, L.F.; Fernande, P.M.G.; da Cruz, G.H.S. Primeiro diagnóstico de febre do Nilo Ocidental em humano em Minas Gerais: relato de caso. Braz J Infect Dis. 2022, 26, 102294. [Google Scholar] [CrossRef]
- Vieira, M.A.C.S.; Romano, A.P.M.; Borba, A.S.; Silva, E.V.P.; Chiang, J.O.; Eulálio, K.D.; Azevedo, R.S.S.; Rodrigues, S.G.; Almeida-Neto, W.S.; Vasconcelos, P.F.C. West Nile virus encephalitis: The first human case recorded in Brazil. Am J Trop Med Hyg. 2015, 93, 377–379. [Google Scholar] [CrossRef]
- Colpitts, T.M.; Conway, M.J.; Montgomery, R.R.; Fikrig, E. West Nile Virus: biology, transmission, and human infection. Clin Microbiol Rev. 2012, 25, 635–648. [Google Scholar] [CrossRef]
- Davis, L.E.; Debiasi, R.; Goade, D.E.; Haaland, K.T.; Harrington, J.A.; Harnar, J.B.; Pergam, S.A.; King, M.K.; Demasters, B.K.; Tyler, K.L. West Nile virus neuroinvasive disease. Ann Neurol. 2006, 60, 286–300. [Google Scholar] [CrossRef]
- Chapman, G.E.; Baylis, M.; Archer, D.; Daly, J.M. The challenges posed by equine arboviruses. Equine Vet J. 2018, 50, 436–445. [Google Scholar] [CrossRef]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The Global Ecology and Epidemiology of West Nile Virus. BioMed Research International. 2015, 2015, 376230. [Google Scholar] [CrossRef] [PubMed]
- McCullough, J. Transfusion-Transmitted Diseases. In Transfus Medicine, 5th ed.; McCullough, J; Wiley-Blackwell: Hoboken, NJ, 2021, pp. 422-452.
- Pealer, L.N.; Marfin, A.A.; Petersen, L.R.; Lanciotti, R.S.; Page, P.L.; Stramer, S.L.; Stobierski, M.G.; Signs, K.; Newman, B.; Kapoor, H.; et al. Transmission of West Nile virus through blood transfusion in the United States in 2002. N Engl J Med. 2003, 349, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Rios, M.; Sylvester, D.; Chancey, C.; Hewlett, I.K.; Stramer, S.L. West Nile Virus Adheres to Human Red Blood Cells in Whole Blood. Clin Infect Dis. 2007, 45, 181–186. [Google Scholar] [CrossRef]
- Harrington, T.; Kuehnert, M.J.; Kamel, H.; Lanciotti, R.S.; Hand, S.; Currier, M.; Chamberland, M.E.; Petersen, L.R.; Marfin, A.A. West Nile virus infection transmitted by blood transfusion. Transfusion 2003, 43, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, M.L.G.; Amarilla, A.A.; Figueiredo, G.G.; Alfonso, H.L.; Lippi, V.; Maia, F.G.M.; Morais, F.A.; Costa, C.A.D.; Henriques, D.A.; Durigon, E.L.; et al. Cacipacore virus as an emergent mosquito-borne Flavivirus. Rev Soc Bras Med Trop. 2017 50, 539–542. [CrossRef]
- Batista, W.C.; Tavares, G.S.B.; Vieira, D.S.; Honda, E.R.; Pereira, S.S.; Tada, M.S. Notification of the first isolation of Cacipacore virus in a human in the State of Rondônia, Brazil. Rev Soc Bras Med Trop. 2011, 44, 528–530. [Google Scholar] [CrossRef]
- Laemmert, H.W.; Hughes, T.P. The virus of Ilhéus encephalitis: isolation, serological specificity and transmission. J Immunol. 1947, 55, 61–67. [Google Scholar] [CrossRef]
- Bernal, M.K.M.; Chiang, J.O.; Mendes, F.F.; Andrade, S.L.S.; da Silva, S.K.S.M.; Pereira, W.L.A. Study of Arboviruses in Philander opossum, Didelphis marsupialis and Nectomys rattus captured from forest fragments in the municipality of Belém, Pará, Brazil. Cienc Rural. 2021, 51, n 4, e20200515. [Google Scholar] [CrossRef]
- Pereira, L.E.; Suzuki, A.; Coimbra, T.L.M.; Souza, R.P.; Chamelet, E.L.B. Arbovírus Ilheus em aves silvestres (Sporophila caerulescens e Molothrus bonariensis). Rev. Saude Publica. 2001, 35, 119–123. [Google Scholar] [CrossRef]
- Gomes, G.; Causey, O.R. Bussuquara, a new arthropod-borne virus. Proc Soc Exp Biol Med. 1959, 101, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Srihongse, S.; Johnson, C.M. The first isolation of Bussuquara virus from man. Trans R Soc Trop Med Hyg. 1971, 65, 541–542. [Google Scholar] [CrossRef] [PubMed]
- Gibrail, M.M. Detecção de anticorpos para arbovírus em primatas não humanos no município de Goiânia, Goiás. Master’s dissertation, Goiás Federal University, Goiânia, Brazil, 2015.
- Casals, J. The arthropod-borne group of animal viruses. Trans R Soc Trop Med Hyg. 1957, 19, 219–235. [Google Scholar]
- Gubler, D.J.; Kuno, G.; Sather, G.E.; Velez, M.; Olivier, A. Mosquito cell cultures and specific monoclonal antibodies in surveillance for dengue viruses. Am J Trop Med Hyg. 1984, 33, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Meslin, F.X.; Stohr, K.; Heymann, D. Public health implications of emerging zoonoses. Rev Sci Tech. 2001, 19, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.H.N.G. Estudos epidemiológicos sobre arbovírus em populações rurais e urbanas do Estado do Amazonas. Master’s dissertation, Amazonas Federal University, Manaus, Brazil, 2009.
- IEC (Evandro Chagas Institute). O estudo da Arbovirologia em defesa da Amazônia. Available online: https://antigo.iec.gov.br/especial-85-anos-arbovirologia/ (accessed on 04 April 2022).
- Vasconcelos, H.B.; Nunes, M.R.; Casseb, L.M.; Carvalho, V.L.; Pinto da Silva, E.V.; Silva, M.; Casseb, S.M.; Vasconcelos, P.F. Molecular epidemiology of Oropouche virus, Brazil. Emerg Infect Dis. 2011, 17, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Chastel, C. [Asymptomatic infections in man: a Trojan horse for the introduction and spread of mosquito-borne arboviruses in non-endemic areas? ] Bull Soc Pathol Exot. 2011, 104, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Faria, N.R.; Azevedo, R.S.S.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika virus in the Americas: Early epidemiological and genetic findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef]
- Marbán-Castro, E.; Goncé, A.; Fumadó, V.; Romero-Acevedo, L.; Bardají, A. Zika virus infection in pregnant women and their children: a review. Eur J Obstet Gynecol Reprod Biol. 2021, 265, 162–168. [Google Scholar] [CrossRef]
- Lowe, R.; Lee, S.; Martins, L.R.; Torres, C.C.; Castro, M.C.; Pascual, M. Emerging arboviruses in the urbanized Amazon rainforest. BMJ. 2020, 371, m4385. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P.; McCarthy, K.; Bedford, P.; Johnson, C.T.; Nichols, R.; Yoksan, S.; Marchesani, R.; Knauber, M.; Wells, K.H.; Arroyo, J.; et al. Clinical proof of principle for ChimeriVax: recombinant live, attenuated vaccines against flavivirus infections. Vaccine 2002, 20, 1004–1018. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Alphaviruses and flaviviruses transmission cycles.
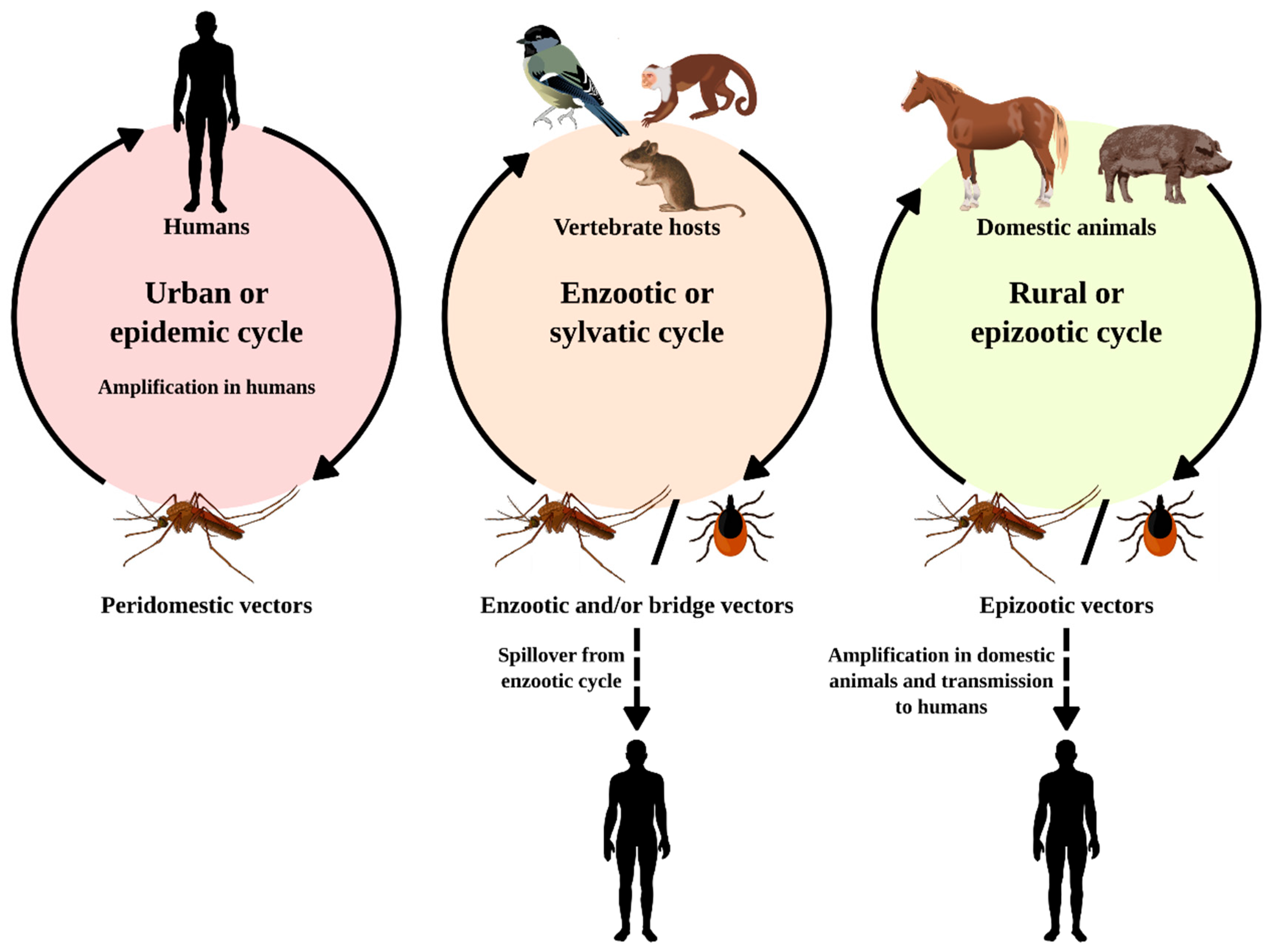
Figure 2.
Municipalities in Pará state where flaviviruses were isolated over 6 decades by the SAARB/IEC.
Figure 2.
Municipalities in Pará state where flaviviruses were isolated over 6 decades by the SAARB/IEC.
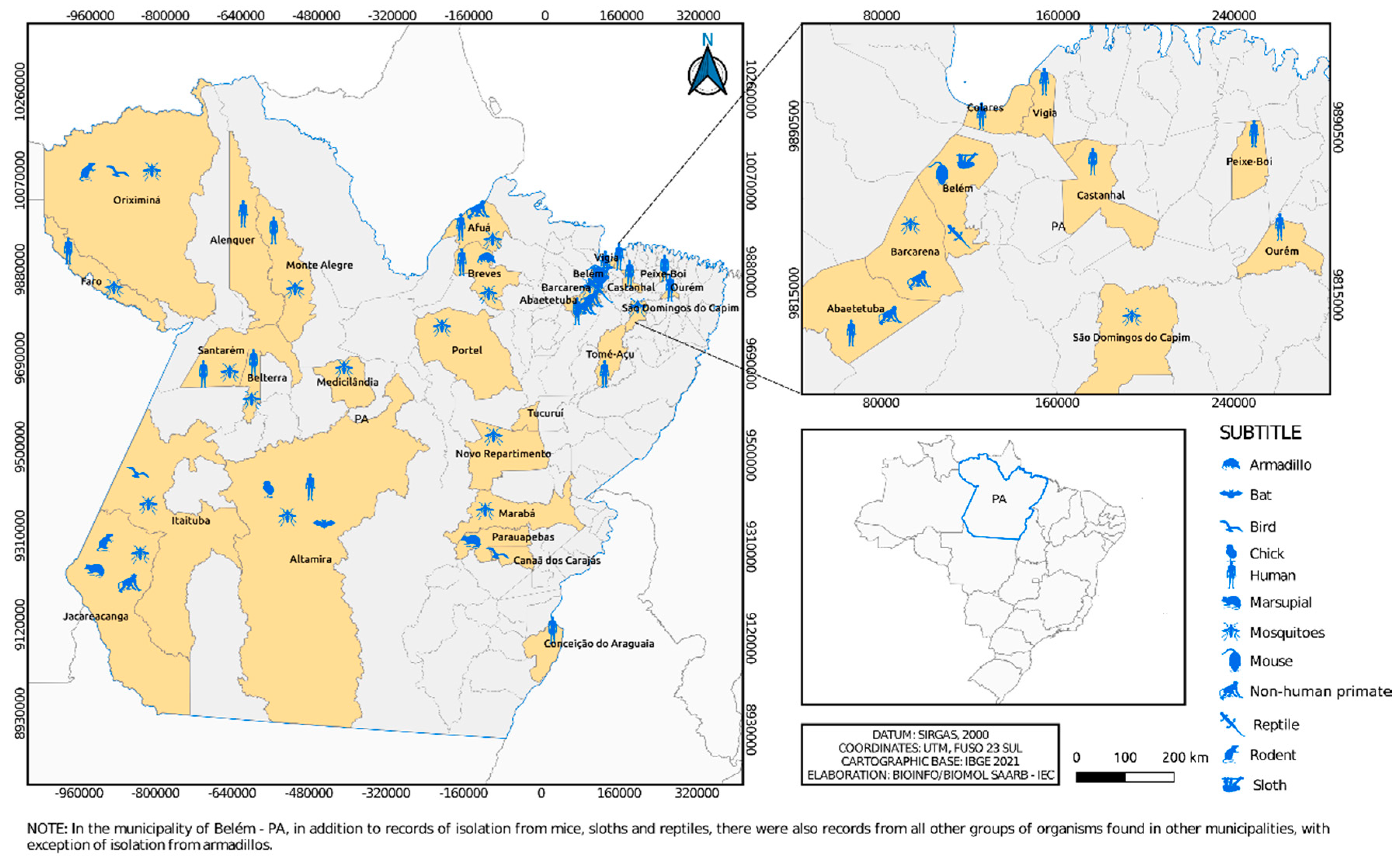
Figure 3.
Municipalities in Pará state where alphaviruses were isolated over 6 decades by the SAARB/IEC.
Figure 3.
Municipalities in Pará state where alphaviruses were isolated over 6 decades by the SAARB/IEC.
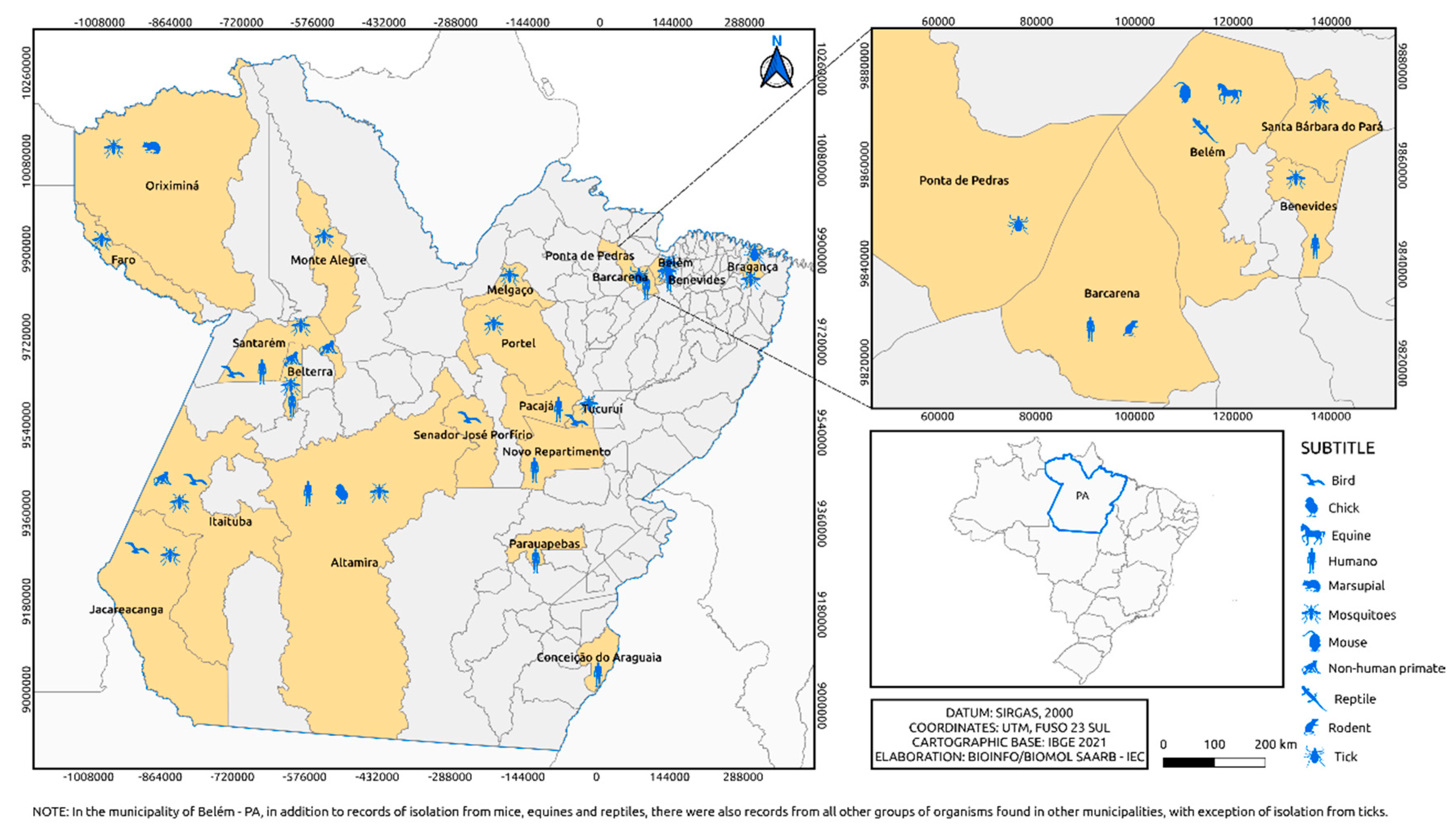
Figure 4.
Distribution of the incidence of Flavivirus (A) and Alphavirus (B) reservoirs by federative unit of Brazil.
Figure 4.
Distribution of the incidence of Flavivirus (A) and Alphavirus (B) reservoirs by federative unit of Brazil.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



