
Submitted:
14 February 2023
Posted:
15 February 2023
You are already at the latest version
Abstract
Cigar stacking fermentation is a key step in tobacco aroma enhancement and miscellaneous gas reduction, which both have a great influence on increasing cigar flavor and improving industrial availability. To analyze the effect of cigar tobacco leaf (CTLs) microbial community structure on volatiles during cigar stacking fermentation, this study used multi-omics technology to reveal the changes in microbial community structure and volatiles of different cigar varieties during stacking fermentation, in addition to exploring the interaction mechanism of microbiome and volatiles. The results showed that the dominant microbial compositions of different CTL varieties during stacking fermentation were similar, which included Staphylococcus, Corynebacterium 1, Aerococcus, and Aspergillus. These dominant microbes mainly affected the microbial community structure and characteristic microorganisms of CTLs through microbial interactions, thereby influencing the transformation of volatiles. Characteristic microorganisms of different CTLs varieties such as Trichothecium, Trichosporon, Thioalkalicoccus and Jeotgalicoccus, were found to posively correlate with characteristic volatiles like megastigmatrienone 4, pyrazine, tetramethyl-, geranyl acetone, and 2-undecanone, 6,10-dimethyl-, respectively. This in turn affected the aroma and sensory quality of the CTLs. This study provides theoretical support for the analysis of the mechanism of microorganisms on volatiles and aroma, and development of microbial agents during cigar stacking fermentation.
Keywords:
Cigar
; stacking fermentation
; microbial community
; volatiles
; interaction
; aroma
Introduction
Cigars are tobacco products rolled from cigar tobacco leaves (CTLs), and are made through a long process of preparation, fermentation, and aging, ultimately creating a unique and complex flavor [1,2]. Cigars have various aromas such as nutty, flowery, fruity, and chocolate, and the production of these complex aromas involves the transformation of many substances related to the microbial community and enzymes in CTLs [3,4]. The content of macromolecular substances and their related enzyme activity change significantly during the growth and agricultural fermentation stages of CTLs [5]. Starch, cellulose, and pectin accumulated during CTLs growth and maturation processes are then gradually degraded during the agricultural fermentation stage and subsequently converted into the aroma precursors and volatiles of CTLs [6]. At this point, the primary aroma of CTLs has been formed, although the smoke gas remains relatively rough, and miscellaneous gas, bitterness, and other bad flavors need to be further mellowed, in addition to reducing irritation through stacking fermentation to further enrich the aroma of CTLs and improve their quality [7]. Stacking fermentation is the industrial fermentation stage in the production process of cigars, and compared with the rapid degradation of macromolecular substances in the growth and agricultural fermentation process, stacking fermentation primarily transforms small molecular substances and volatiles. The content of volatiles changes with the function of microorganisms and related enzymes, although there is little variation in type, and to achieve the effect of increasing aroma, reducing miscellaneous gas, and mellowing the smoke gas [7,8,9].
In recent years, the study of aroma type, microbial community composition, and volatiles in cigar stacking fermentation has increased [7,9], yet the evolution of the microbiome structure during stacking fermentation and its mechanism for characteristic aroma formation remain poorly understood. Therefore, it is necessary to use high-throughput sequencing technology to study the changes in microbial community composition during stacking fermentation, since this is conducive to the mining of functional microorganisms and the analysis of the evolution mechanism of microbial communities, and to further improve the level to which the industrial fermentation production process can be controlled. This is very important for improving the production and process standards related to cigar fermentation [10,11,12,13]. In this study, different varieties of high-quality CTLs were used as the research objects, whilst the changes in microbial community composition and volatile changes during stacking fermentation were dynamically analyzed. The correlation network between microorganisms and volatile aroma was first established, with the changes in microbial community structure during stacking fermentation and the influence of this on volatiles and aroma subsequently analyzed. This study provides guidance for analyzing the mechanism of stacking fermentation and the excavation of functional microorganisms involved in this.
Materials and Methods
2.1. Sample collection
Eight high-quality CTL samples were collected from China Tobacco Hubei Industry Co., LTD., including different varieties of CX14, DX4, CIOLLO 98, N-Jalap HABANA, HVA, BESUKI, MATA FINA, and E-HABANO 2000. The entire stacking fermentation process consists of a total of five nodes, for every 7 d of fermentation, or temperature reached 45°C for sampling and re-stacking. Five nodes were named, as raw materials (T0, 0–7 d), pre-fermentation (T1, 7–14 d), mid-fermentation (T2, 14–21 d), post-fermentation (T3, 21–28 d), and end of fermentation (T4, 28–35 d). Further information regarding these samples is presented in Table S1.
2.2. CTLs sampling
CTL samples of 5 g were added to a sterile pre-chilled mortar, before including liquid nitrogen, and this mixture was then ground into a powder. The CTL powder was put into a 500 mL shaker flask containing 100 mL of sterile normal saline and was subsequently treated at 10°C and 220 rpm for 2–3 h. After the elution was filtered, CTL powder was removed by filtering through four layers of gauze, before the filtrate was centrifuged at 4°C and 7000 rpm for 20 min. The precipitate was then collected to represent the total microorganism content of the CTLs. These collected microbial cells were extracted using the DNeasy PowerSoil Pro Kit to show CTL microbial genomic DNA. The extracted DNA samples were then amplified through PCR using the primers ITS1-ITS4 and V4-V5. The amplification products were then electrophoresed on 1 % agarose gel, with the corresponding products being recovered for further analysis.
2.3. Amplicon sequencing
Here, the Illumina platform was used to sequence the DNA fragments of the microbial community using the two-ended sequencing method, silva_132 and unite_8, whilst the sequence analysis method used was DADA2 [14].
2.4. Profiling of volatiles by HS-SPME-GC-MS during stacking fermentation
Using a DB-5MS column with headspace solid-phase microextraction (50/30 DVB/CAR/PDMS fiber), the helium flow rate was maintained at 1.0 mL/min. The temperature procedure was as follows: At the beginning of injection, the column temperature was at 40°C (held for 2 min) and was then increased to 250°C (held for 5 min) at a rate of 10°C/min. The ion source temperature was subsequently maintained at 210°C with the transfer line temperature being maintained at 280°C. The system was operated in the EI mode with an EI voltage of 70 eV. The full scan mode used a mass scan range of 33–400 m/z alongside an acquisition rate of 10 scans per second [9]. Compared with the chemical standards of the National Institute of Standards and Technology spectral library WILEY 8.0 and NIST14, volatiles with similarity above 800 and flavor characteristics were selected for analysis, with the relative content being calculated using the area normalization method [15].
2.5. Sensory quality analysis
The sensory evaluation of CTL samples was carried out by China Tobacco Hubei Industry Co., LTD.. The indexes of the samples were scored 1–5 points, whilst the non-existent aroma and bad flavors were scored 0 points. Aromas were divided into woody, bean, flower, honey sweet, mellow sweet, burn sweet, light sweet, pollen, milk, hay, roasted, resin, fruity, herb, nutty, leather, coffee and pepper. Bad flavor was divided into green miscellaneous gas, burning, woody gas, earthy, regional gas, and protein.
2.6. Data analysis
The correlations between microorganisms and volatiles, and between volatiles aroma were calculated using the psych package for R software (version 4.2.2) [16]. Furthermore, the pheatmap package was used to create a heatmap of the volatiles, whilst Gephi and Cytoscape software were used for the network of interactions [17], the (Partial Least Squares Discriminant Analysis)PLS-DA model of SIMCA (version 14.1) was used to screen CTLs characteristic volatiles [18], and Origin and Excel software were used for data processing and analysis.
3. Results
3.1. Microbial community changes during stacking fermentation
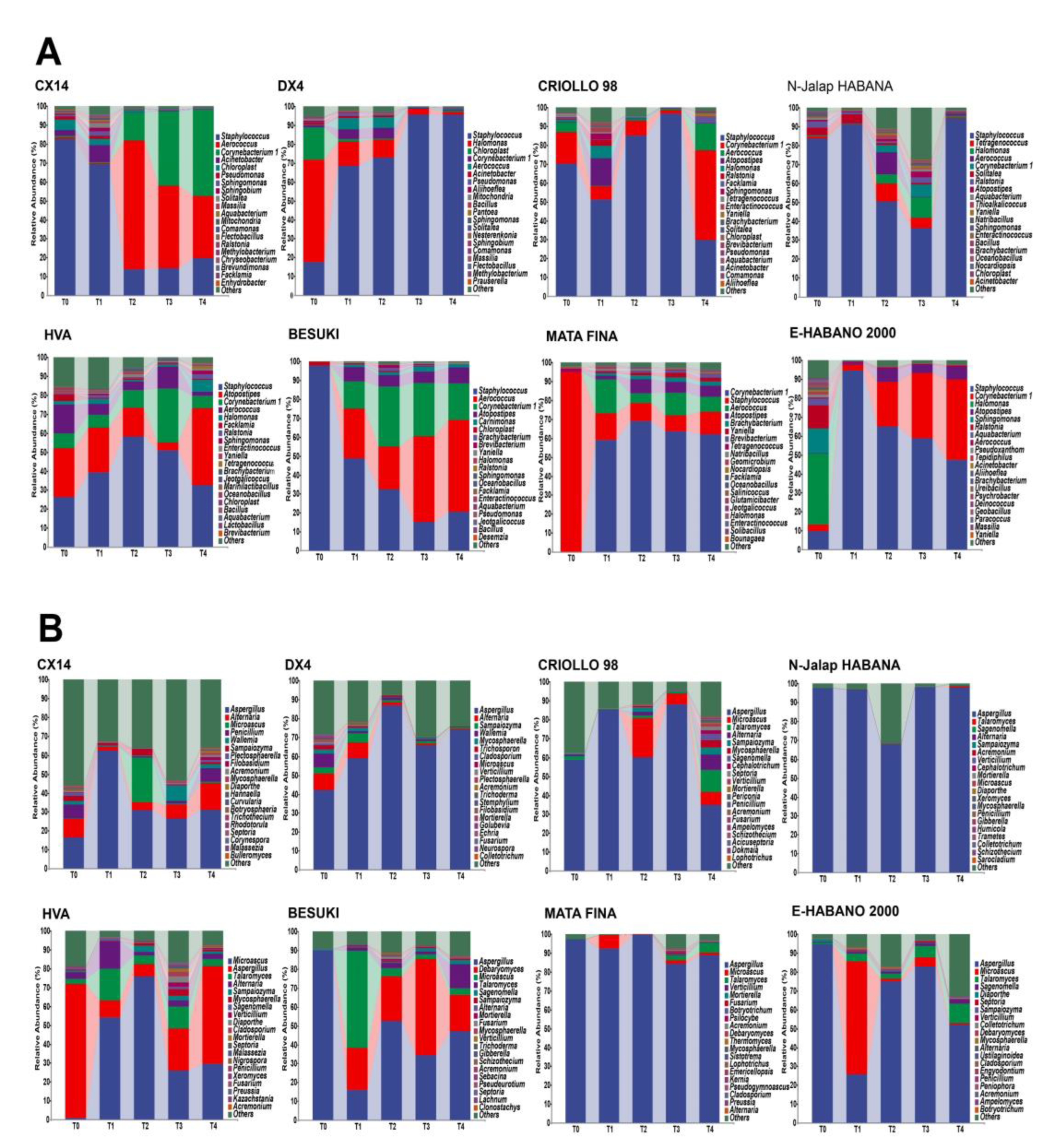
The changes in microbiota structure during cigar stacking fermentation were analyzed by 16S and ITS amplion sequencing, and the bacterial and fungal genera with relative abundances in the top 20 were selected for subsequent analysis, as shown in . The results showed that the compositions of the dominant bacterial genera in the different varieties of CTLs were similar. The three dominant bacterial genera, Staphylococcus, Corynebacterium 1 and Aerococcus, were the main components of the bacterial communities in different CTL varieties. The total relative abundance of all samples was between 67.15 % and 99.10 %; however, the relative abundance changes during stacking fermentation differed. The relative abundances of Corynebacterium 1 and Aerococcus gradually increased from the CX14, BESUKI, and MATA FINA CTLs, while the sum of the relative abundances of Corynebacterium 1 and Aerococcus at the end of fermentation were 79.59 %, 67.68 %, and 72.30 %, respectively. The CTLs from DX4 and N-Jalap HABANA had Staphylococcus as the dominant bacteria in the stacking fermentation process, with its relative abundance exceeding 90 % at the end of fermentation. Furthermore, the CTLs from CRIOLLO 98, HVA, and E-HABANO 2000 showed Staphylococcus as the main dominant bacteria, although the relative abundance of Staphylococcus gradually decreased, whilst the relative abundances of Halomonas, Atopostipes, Corynebacterium 1, and other genera showed a slight increase. During stacking fermentation, the fungal community with Aspergillus as the dominant fungal genus showed the highest proportion of relative abundance among different varieties of CTL fungi, whilst the relative abundance of Aspergillus in different CTLs varied greatly. Among them, CX14, CRIOLLO 98, HVA, BESUKI, and E-HABANO 2000 CTLs Aspergillus decreased during the fermentation process, with the community composition being richer at the end of fermentation. While Aspergillus was the most dominant fungus in the CTLs from DX4, N-Jalap HABANA, and MATA during stacking fermentation, at the end of fermentation, the relative abundances of Aspergillus were 73.98 %, 97.83 %, and 88.88 %, respectively.
Figure 1.
Microbial community changes of CTLs from different varieties. (A: Composition of bacterial community during stacking fermentation; B: Composition of fungal community during stacking fermentation)
Figure 1.
Microbial community changes of CTLs from different varieties. (A: Composition of bacterial community during stacking fermentation; B: Composition of fungal community during stacking fermentation)
3.2. Characteristic microorganisms from different varieties of CTLs
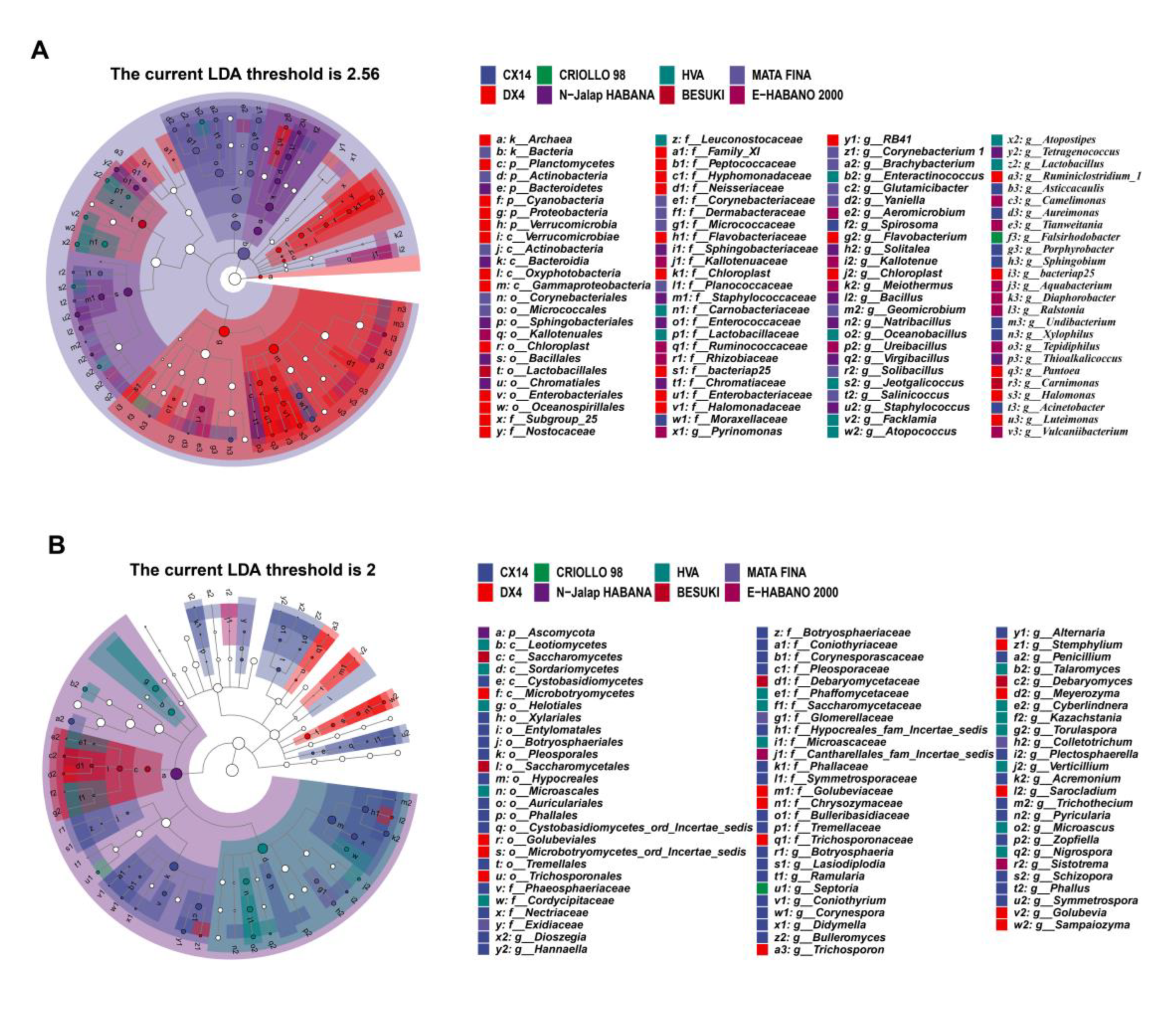
The results showed that the dominant genera and community structures of different samples affected the metabolism of microbial communities, whilst the characteristic microorganisms were significantly correlated with the characteristic volatiles. For example, Staphylococcus and Bacillus, were found to have been positively correlated with sclareolide and isophorone, respectively [8,19,20,21], therefore the characteristic microorganisms of different CTL varieties may have had an important influence on the composition of the characteristic aroma. Linear discriminant analysis effect size (LEfSe) was used to identify the characteristic microorganisms from different varieties of CTLs (LDA score > 2), and the results of this are shown in . Among these, DX4 CTLs had the largest number of characteristic bacterial genera, whilst BESUKI CTLs had the fewest characteristic bacterial genera. Additionally, CX14 CTLs showed the largest number of characteristic fungal genera, whilst N-Jalap HABANA CTLs had the fewest characteristic fungal genera.
Figure 2.
Characteristic microorganisms from different varieties of CTLs. (A: Characteristic bacterium; B: Characteristic fungus).
Figure 2.
Characteristic microorganisms from different varieties of CTLs. (A: Characteristic bacterium; B: Characteristic fungus).
3.3. Interaction relationship between microbial community
Using Spearman to analyze the interaction relationship between CTL microbial communities, ρ < 0.05, r > 0.6 were defined as significant positive correlations, whilst ρ < 0.05, r < -0.6 were defined as significant negative correlations, with the results being shown in . During stacking fermentation, among the dominant bacteria, Staphylococcus were negatively correlated with Corynebacterium 1 and Aerococcus, whilst Corynebacterium 1 and Aerococcus interacted positively. Dominant bacterial genera from different CTL varieties were significantly related to characteristic microorganisms, for example, Corynebacterium 1 was positively correlated with Facklamia, Jeotgalicoccus, Brachybacterium, Geomicrobium, Glutamicibacter, Yaniella, Salinicoccus, Solibacillus, and Natribacillus. Additionally, Aerococcus was positively correlated with Carnimonas, Facklamia, Yaniella, whilst Staphylococcus was shown to have been positively correlated with Halomonas and Solitalea. During stacking fermentation, the dominant bacterial genera were found to have affected the growth of the corresponding characteristic bacterial genera, such as Aerococcus and Corynebacterium 1, which in turn increased in the pre-fermentation period. This promoted the proliferation of Sphingobium, Aquabacterium, Halomonas, etc., and decreased when Staphylococcus increased during the middle and late fermentation stages. Characteristic bacteria such as Aquabacterium, Brachybacterium, Tetragenococcus, Ralstonia, Enteractinococcus, Jeotgalicoccus, Lactobacillus, Bacillus, and Yaniella all interacted in a positive direction, whilst characteristic fungi such as Sampaiozyma, Wallemia, Penicillium, Trichosporon, Nigrospora, Septoria, and Plectosphaerella also all interacted in a positive direction. These positive interactions between the characteristic microorganisms greatly influenced the evolution of the microbiota structure during the stacking fermentation process. Aspergillus, the dominant fungi of CTLs, was negatively correlated with characteristic fungal genera, including Alternaria, Curvularia, Fusarium, Gibberella, Mycosphaerella, Plectosphaerella, Pseudeurotium, Penicillium, Sampaiozyma, Septoria, and Trichothecium. During stacking fermentation, Aspergillus inhibits the growth of characteristic fungi, such as Alternaria, Trichosporon, and Mycosphaerella which was opposite to the change observed in Aspergillus. The abundance of Aspergillus was found to have been higher during the early stage of fermentation, whilst the characteristic fungus was then inhibited from growing and increased during the middle and late stages of fermentation with a decrease in Aspergillus. The above results showed that the abundance and interactions of dominant microbes during stacking fermentation affected the changes in characteristic microorganisms in the microbial community and subsequently influenced the microbiota structure of CTLs during the stacking fermentation process.
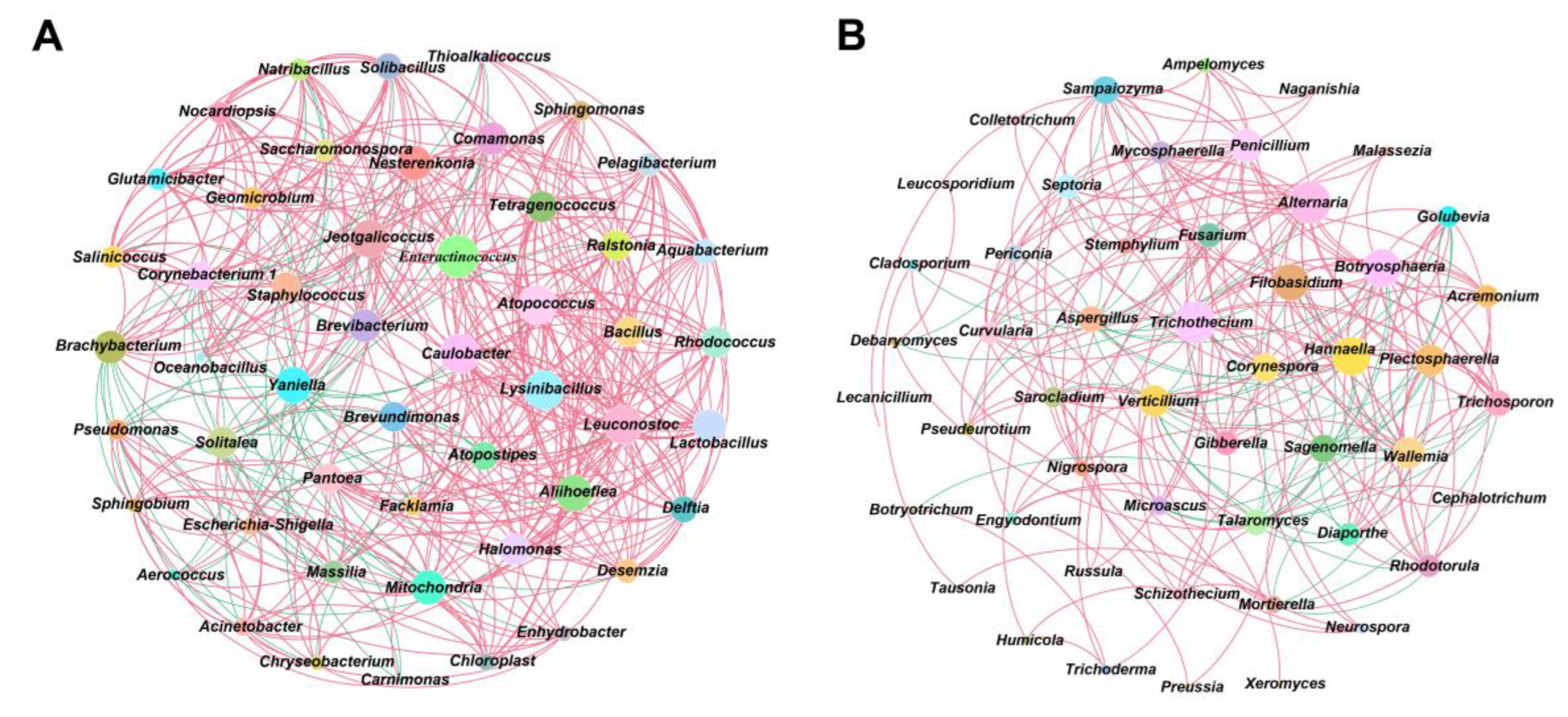
Figure 3.
Microbial community interaction networks. (A: Interaction network between CTLs bacterial communities; B: Interaction network between CTLs fungal communities. Red represents positive correlation and blue represents negative correlation)
Figure 3.
Microbial community interaction networks. (A: Interaction network between CTLs bacterial communities; B: Interaction network between CTLs fungal communities. Red represents positive correlation and blue represents negative correlation)
3.4. Changes of volatiles in the stacking fermentation
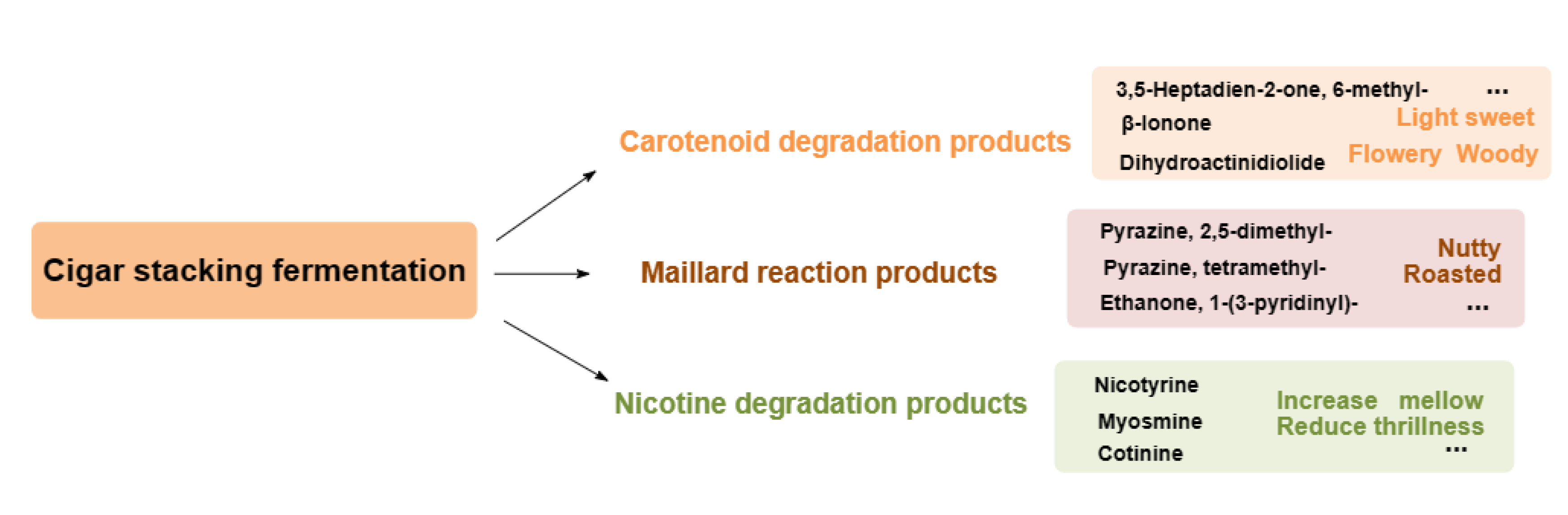
The results showed that the types of volatiles increased during stacking fermentation with the volatiles detected including 2-undecanone, 6,10-dimethyl-, geranyl acetone, β-ionone, phytone, farnesyl acetone, ethanone, 1-(3-pyridinyl)-, myosmine, and nicotyrine [7]. However, the transformation mechanism of stacking fermentation on CTL aroma and sensory quality remained unclear. The results are shown in , with the volatiles at the end of fermentation for different CTLs and the volatiles of CTLs between raw materials and the end of fermentation being found to have been quite different. During stacking fermentation, carotenoid degradation products, Maillard reaction products, and nicotinic degradation products increased, which was consistent with the past results found by Liu’s study, indicating that these three types of substances were the key volatiles for the quality improvement of CTLs [7].
Figure 4.
Cluster analysis of volatiles from different varieties of CTLs between raw materials and the end of fermentation.
Figure 4.
Cluster analysis of volatiles from different varieties of CTLs between raw materials and the end of fermentation.
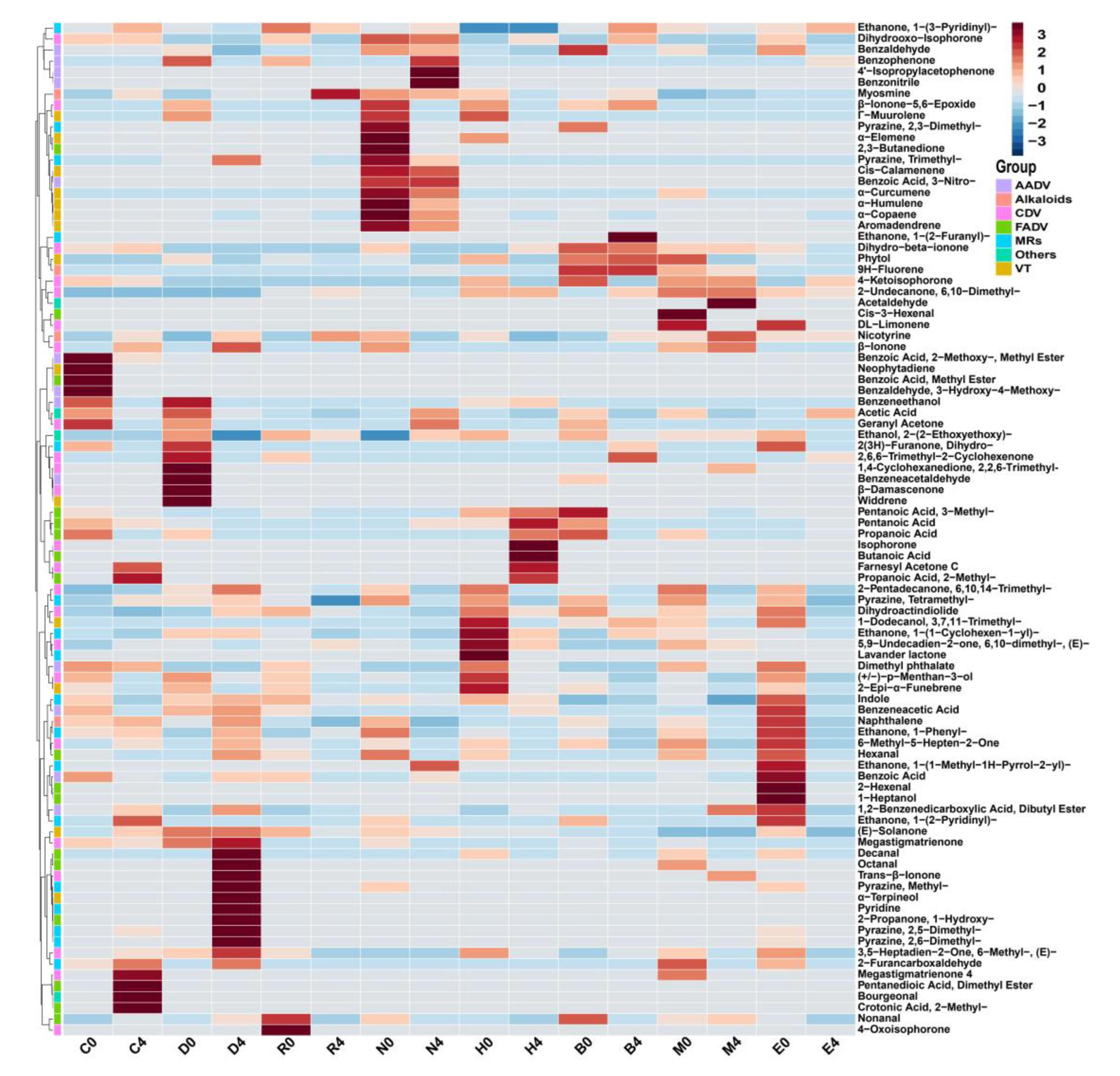
In addition to β-ionone, dihydroactinidiolide, 6-methyl-3,5-heptadien-2-one, geranyl acetone, and isophorone, the carotenoid degradation products in different varieties of CTLs also contained β-ionone-5,6-epoxide, menthol, and α-terpineol, among other aroma intermediates. Carotenoid degradation products are important components of tobacco aroma, the threshold is low, and their type and content both have an important impact on improving tobacco aroma and mellowness [22,23]. For example, β-ionone is an aroma ingredient of CTLs flowery and woody aroma; dihydroactinidiolide has a sweet and woody aroma, which could mellow the smoke gas; 6-methyl-3,5-heptadien-2-one and geranyl acetone have a rosy, leafy, and fruity aroma [23]. The content of Maillard reaction products, such as pyrazine, 2,5-dimethyl-, pyrazine, 2,6-dimethyl-, pyrazine, tetramethyl-, furfural, 2-propanone, 1-hydroxy-, ethanone, 1-(3-pyridinyl)-, and indole, all increased during stacking fermentation. Maillard reaction products are mainly generated by carbohydrates and amino acids via the production of dicarbonyl compounds and amino ketones, primarily creating nutty, roasted, chocolate, and other aromas for CTLs [24]. Nicotine, the most abundant and important alkaloid in CTLs, is irritating and bitter when its content is too high, whilst there is insufficient smoke and a bland flavor when its content is too low. CTLs can reduce irritation and increase mellowness through degrading nicotine to produce myosmine and nicotine during stacking fermentation [25]. Therefore, in stacking fermentation, the transformation of carotenoid degradation and Maillard reaction products alongside the degradation of nicotine played a role in mellowing smoke gas and enhancing the aroma of CTLs.
Figure 5.
Effects from volatiles of stacked fermented CTLs on the aroma and quality.
3.5. Characteristic volatiles from different CTL varieties
The characteristic volatiles of different CTL varieties were analyzed using partial least squares discriminant analysis (PLS-DA), and the results are shown in . The model Q2 value was 0.809, indicating a better model prediction effect. Volatiles with a VIP value greater than 1 were defined as characteristic volatiles, which contributed more to the difference between samples. The characteristic volatiles at the end of the fermentation of CTL varieties are shown in .
Figure 6.
PLS-DA Analysis of volatiles at the end of fermentation. (a: PLS-DA score plot of volatiles; b: Loading plot of volatiles; and c: VIP value of volatiles).
Figure 6.
PLS-DA Analysis of volatiles at the end of fermentation. (a: PLS-DA score plot of volatiles; b: Loading plot of volatiles; and c: VIP value of volatiles).
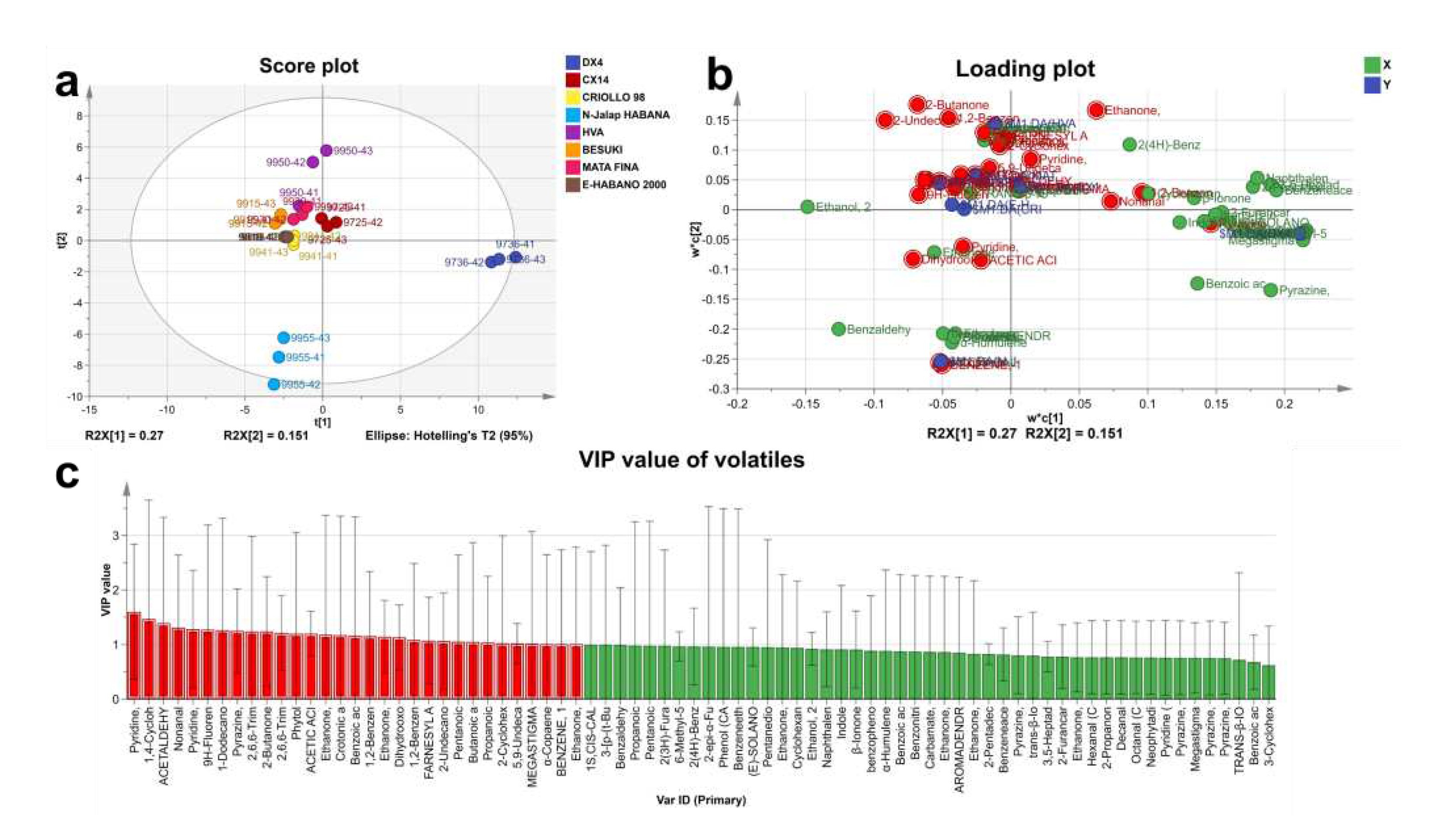

Table 1.
Characteristic aroma components of different varieties of tobacco leaves.
| Varieties | Characteristic volatiles |
| CX14 | Ethanone, 1-(2-pyridinyl)-、Tiglic acid、Benzoic acid, 2-methoxy-, methyl ester、Dimethyl phthalate、Megastigmatrienone 4、Bourgeonal |
| DX4 | Nonanal、Pyrazine, tetramethyl-、Ethanone, 1-(1-Cyclohexen-1-yl)- |
| CRIOLLO 98 | Myosmine |
| N-Jalap HABANA | Acetic Acid、Geranyl Acetone、α-Curcumene |
| HVA | Farnesyl acetone C、2-Undecanone, 6,10-dimethyl-、Pentanoic acid, 3-methyl-、Butanoic acid、Propanoic acid, 2-methyl-、Isophorone、α-Copaene |
| BESUKI | 9H-Fluorene、1-Dodecanol, 3,7,11-Trimethyl-、2,6,6-Trimethyl-2-Cyclohexenone、Dihydro-beta-ionone、Phytol、Dihydrooxo-isophorone、Ethanone, 1-(2-furanyl)- |
| MATA FINA | 1,4-Cyclohexanedione, 2,2,6-Trimethyl-、Acetaldehyde、Nicotyrine、1,2-Benzenedicarboxylic acid, dibutyl ester、4-Oxoisophorone |
3.6. Correlation analysis between microorganisms-volatiles and volatiles-aroma
The interaction between CTLs microorganisms and volatiles during stacking fermentation was analyzed by Spearman correlation analysis, with ρ < 0.05 and r greater than 0.6 being defined as significant positive correlations, and ρ < 0.05 and r less than -0.6 being defined as significant negative correlations, as shown in . The correlation between bacteria and volatiles during stacking fermentation was more complex than that between fungi and volatiles whilst also being mainly positively correlated. Characteristic volatiles of different varieties of CTLs were also positively correlated with characteristic microorganisms, such as in CX14 CTLs, Penicillium was positively correlated with dimethyl phthalate, whilst Diaporthe, Trichothecium were positively correlated with ethanone, 1-(2-pyridinyl)-, tigelic acid, megastigmatrienone 4, and bourgeonal. This provides CTLs with a fresh flowery and light sweet aroma. In DX4 CTLs, Chloroplast was positively correlated with ethanone, 1-(1-cyclohexen-1-yl)-, and Trichosporon was positively correlated with pyrazine, tetramethyl-, which could increase the roasted and burnt sweet aroma of CTLs. Among N-Jalap HABANA CTLs, Thioalkalicoccus was positively correlated with geranyl Acetone and α-curcumene, and Solitalea was positively correlated with acetic acid, which could increase the light sweet and flowery aroma of CTLs. Furthermore, Enteractinococcus, Jeotgalicoccus, and Talaromyces in the HVA CTLs were positively correlated with 2-undecanone, 6,10-dimethyl-, and Lactobacillus, whilst Atopococcus was positively correlated with butanoic acid and isophorone, which could increase the fruity and mint aroma. Carnimonas and Debaryomyces in BESUKI CTLs were also positively correlated with 1-dodecanol, 3,7,11-trimethyl- and dihydro-beta-ionone, which could increase the flowery and hay aroma. Additionally, Yaniella in MATA FINA CTLs was positively correlated with nicotine, when Salinicoccus, Nocardiopsis, Natribacillus, Solibacillus, Geomicrobium, Salinicoccus and Glutamicibacter were positively correlated with 4-oxoisophorone and 1,4-cyclohexanedione, 2,2,6-trimethyl-, which could increase the sweet, hay, nutty, and woody aroma.
Figure 7.
Interaction network between CTLs microorganisms and volatiles. (A: Interaction network between bacteria and volatiles; B: Interaction network between fungi and volatiles. Red represents positive correlation and blue represents negative correlation).
Figure 7.
Interaction network between CTLs microorganisms and volatiles. (A: Interaction network between bacteria and volatiles; B: Interaction network between fungi and volatiles. Red represents positive correlation and blue represents negative correlation).
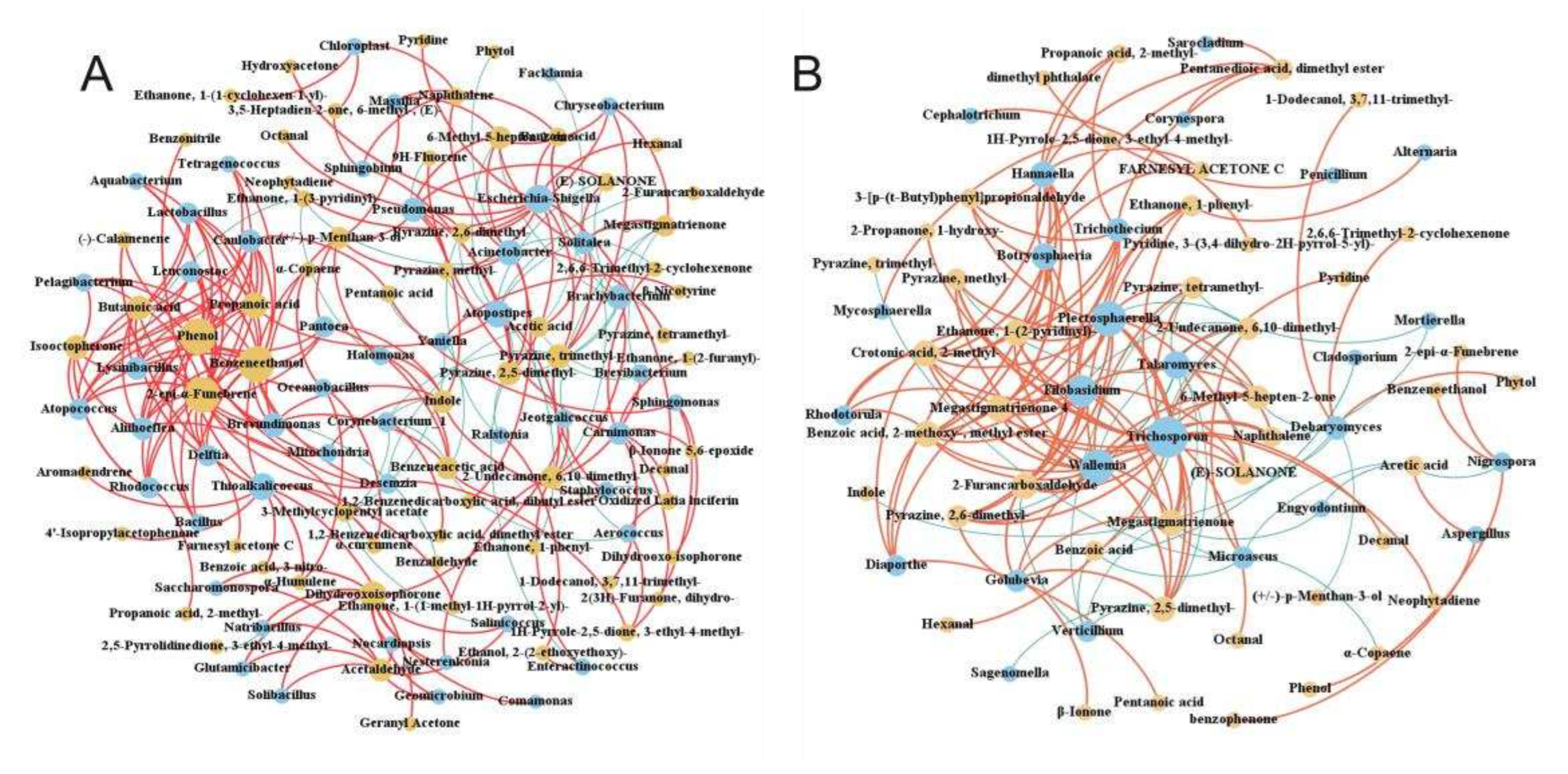
Figure 8.
Aroma composition of CTLs and interaction network between volatiles and aroma. (A: Radar chart of the aroma from different varieties of CTLs; B: Interaction network between the volatiles and aroma. Red represents positive correlation and blue represents negative correlation).
Figure 8.
Aroma composition of CTLs and interaction network between volatiles and aroma. (A: Radar chart of the aroma from different varieties of CTLs; B: Interaction network between the volatiles and aroma. Red represents positive correlation and blue represents negative correlation).
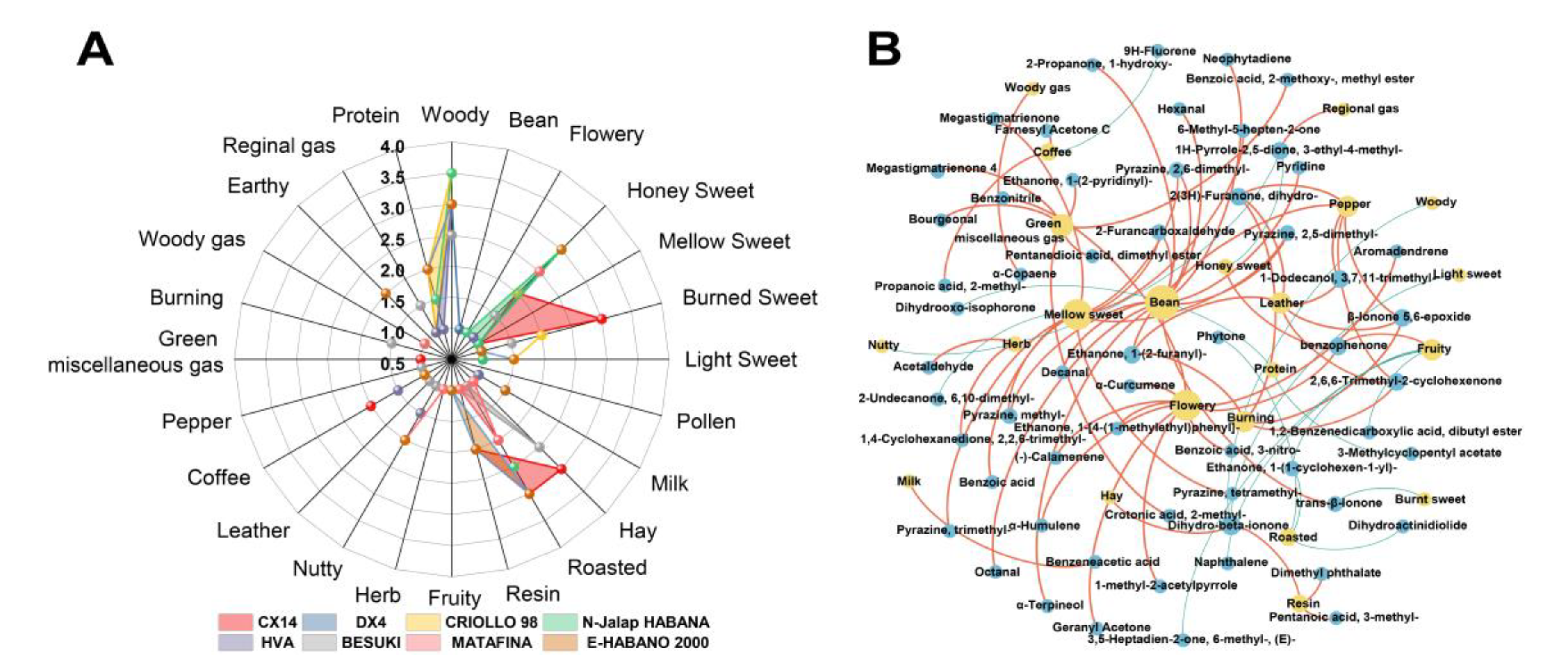
Radar charts were used to identify the aroma of different CTL varieties at the end of stacking fermentation. The results are shown in , where the woody aroma was found to be the most prominent among the different CTL varieties, with each sample being supplemented with another aroma.
Using Spearman correlation analysis, the interaction network between volatile aroma was established; ρ < 0.05 and r greater than 0.6 were defined as significant positive correlations, whilst ρ < 0.05 and r less than -0.6 were defined as significant negative correlations, with the results being shown in B. The characteristic volatiles of CTLs showed a stronger positive correlation with many aromas, such as bean aroma, which was positively related to nonanal, decanal, pyrazine, 2,5-dimethyl-, furfural, pyrazine, 2,6-dimethyl-, and α-terpineol. Meanwhile, the coffee aroma was positively correlated with farnesyl acetone C and propanoic acid, 2-methyl-, whilst the flowery aroma was positively correlated with geranyl acetone, benzophenone, benzonitrile, and α-curcumene. Hay aroma was positively correlated with dihydro-beta-ionone; Honey sweet aroma was positively correlated with benzophenone; Mellow sweet aroma was positively correlated with megastigmatrienone, pyrazine, tetramethyl-, and furfural; Pepper aroma was positively correlated with 1-dodecanol, 3,7,11-trimethyl-, 2,6,6-trimethyl-2-cyclohexenone and ethanone, 1-(2-furanyl)-. The resin aroma was positively correlated with dimethyl phthalate and dihydro-beta-ionone.
The characteristic volatiles of different CTLs varieties were positively correlated with the aroma of CTLs, such as the characteristic volatiles of DX4 CTLs, including nonanal and pyrazine, tetramethyl, which corresponded to sweet, roasted, and nutty honey aromas, respectively. In N-Jalap HABANA CTLs, geranyl acetone and α-curcumene were positively correlated with the honey sweet aroma. The characteristic volatiles farnesyl acetone C and propanoic acid, 2-methyl-, were positively correlated with coffee and roasted aroma in HVA CTLs. In BESUKI CTLs, dihydro-beta-ionone was positively correlated with hay aroma; In MATA FINA CTLs, acetaldehyde and 4-oxoisophorone were positively correlated with honey sweet and roasted aroma, respectively. In CX14 CTLs, characteristic volatiles were not strongly correlated with the aroma, but the volatiles with a pleasant aroma, such as the characteristic volatiles ethanone, 1-(2-pyridinyl)-, tigelic acid, benzoic acid, 2-methoxy-, methyl ester, and bourgeonal, corresponded to the burn sweet, roasted and hay aromas, respectively. Furthermore, the higher contents of ethanone, 1-(3-pyridinyl)-and pentanoic acid, 3-methyl- in E-HABANO 2000 CTLs were positively correlated with featured roasted and resin aromas, respectively. The above results showed that the characteristic volatiles of CTLs had an important contribution to the aroma.
4. Discussion
CTLs that go through stacking fermentation could better improve their aroma quality, aroma amount, and mellowness, whilst decreasing irritation; therefore, it is important to study the influence of the microbial community on aroma changes and the mechanism of this. In this study, through the analysis of the microbial community composition and volatiles of CTLs, the interaction network between microorganisms, volatile and aroma was established, as shown in . This which provides theoretical support for the analysis of the mechanism of microorganisms and the changes in volatiles during stacking fermentation as well as the development of cigar stacking fermentation microbial agents.
Figure 9.
Interaction network among microorganism- volatiles -aroma. (Red represents positive correlation, blue represents negative correlation).
Figure 9.
Interaction network among microorganism- volatiles -aroma. (Red represents positive correlation, blue represents negative correlation).
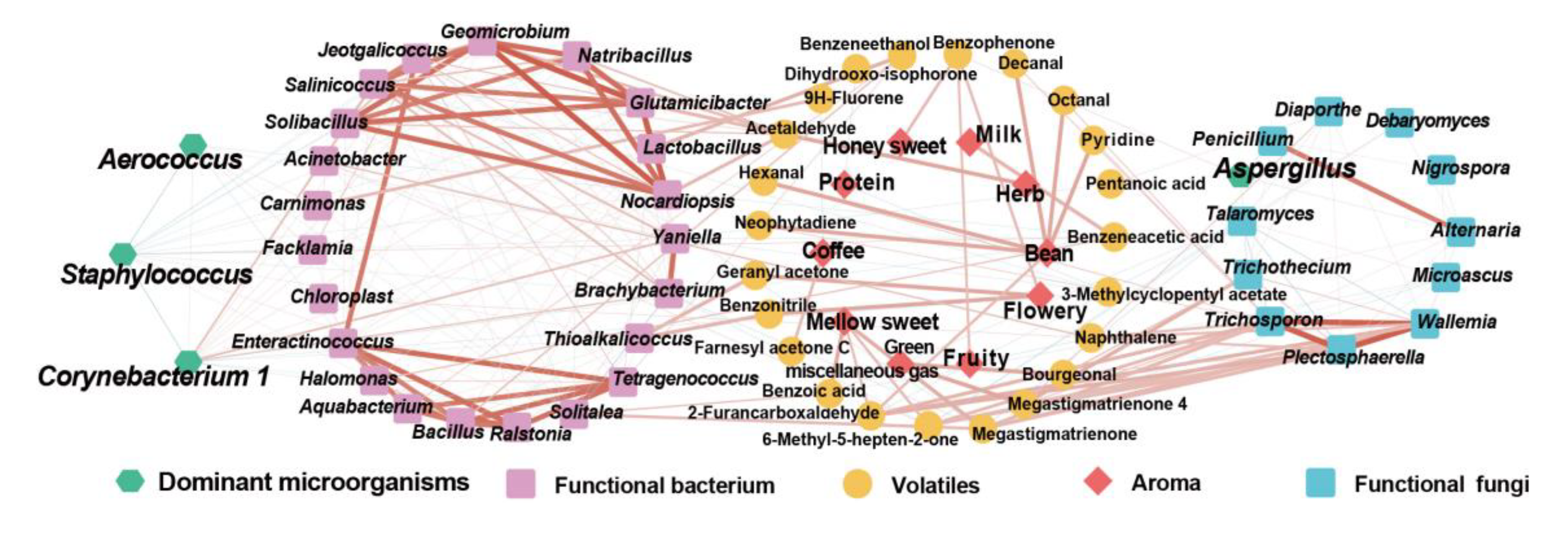
The results of amplicon sequencing showed that the microbial community structures of different CTLs varieties were quite different during stacking fermentation, whilst the dominant microbial structures remained similar. The dominant bacterial genera here were Staphylococcus, Corynebacterium 1, Aerococcus, and Aspergillus, which was similar to the results found previously by Zheng et al (2022). [9,26]. The results of microbial interaction analysis showed that during stacking fermentation, the dominant microorganisms were strongly related to the CTLs characteristic microorganisms, whilst the abundance of characteristic microorganisms was affected by the correlation and abundance of dominant microorganisms. These characteristic microorganisms also showed a positive correlation with the aroma and could be defined as functional microorganisms in the stacking fermentation process. The characteristic bacterial genera Jeotgalicoccus, Geomicrobium, Glutamicibacter, and Yaniella were positively correlated with Corynebacterium 1. Furthermore, Carnimonas and Yaniella positively correlated with Aerococcus. Among them, Carnimonas was positively correlated with Dihydro-beta-ionone; Yaniella was positively correlated with nicotine, and Geomicrobium, Salinicoccus, Glutamicibacter were positively correlated with volatiles such as 1,4-cyclohexanedione, 2,2,6-trimethyl-, and 4-oxoisophorone. Among CTLs, Jeotgalicoccus, Glutamicibacter, Yaniella, and other characteristic microorganisms have good salt tolerance, can better adapt to the alkaline environment of CTLs, and have functions including enzyme production and volatile transformation [27,28,29]. Therefore, during the stacking fermentation process, CTL-dominant microorganisms may have affected the succession changes of functional microorganisms and microbial communities through microbial interactions, thereby affecting the transformation and aroma composition of CTLs [9,30].
Stacking fermentation is an important part of mellowing smoke gas and improving tobacco permeability and aroma, with different varieties of CTLs having their own aroma and characteristic volatiles. The analysis of volatiles between raw materials and the end of fermentation showed that volatiles were mainly increased during stacking fermentation, including Maillard reaction products such as nonanal, pyrazine, tetramethyl-, furfural and ethanone, 1-(3-pyridinyl)-. This is in addition to, carotenoid degradation products such as dihydroactinidiolide, geranyl acetone, 2-pentadecanone, 6,10,14-trimethyl-, 2-undecanone, 6,10-dimethyl-, 1,4-cyclohexanedione, 2,2,6-trimethyl-, 6-methyl-3,5-heptandien-2-one, 2,6,6-trimethyl-2-cyclohexenone, as well as nicotine degradation products such as nicotyrine and myosmine. The above results showed the transformation of small molecular substances such as carotenoid degradation, Maillard reaction, and nicotine degradation during the stacking fermentation process, causing the reduction of miscellaneous gas and the improvement of quality and aroma of CTLs [22,25,31]. Carotenoid degradation products are considered to be an important source of flavor in CTLs, among which geranyl acetone and dihydroactinidiolide have a woody aroma, green flavor, and can mellow, and flue smoke gas whilst increasing the smoke concentration. 2,6,6-trimethyl-2-cyclohexenone, 2-undecanone, 6,10-dimethyl- and 1,4-cyclohexanedione, 2,2,6-trimethyl- usually have sweet and violet flavors [3]. Maillard reaction products are important aromagenic components of CTLs that burn with a sweet and roasted aroma, and studies have shown that microorganisms can synthesize aminoketones, which are the precursors of pyrazines, and further convert them into pyrazines [24,31,32]. pyrazine, tetramethyl-, pyrazine, 2,5-dimethyl-, furfural and pyrazine, 2,6-dimethyl- have a roasted aroma and burn sweet aroma, commonly used as CTLs flavor additives [3].
Through the correlation analysis of aroma components and tobacco leaf aroma characteristics, it was found that the characteristic aroma components of different varieties of tobacco leaves corresponded to the prominent aroma of tobacco leaves. For example, Diaporthe and Trichothecium in CX14 CTLs are positively correlated with ethanone, 1-(2-pyridinyl)-, tigelic acid, megastigmatrienone 4, and bourgeonal, which can enhance the flowery and light sweet aroma of CTLs. In DX4 CTLs, Chloroplast were positively correlated with ethanone, 1-(1-cyclohexen-1-yl)-, whilst Trichosporon was positively correlated with pyrazine, tetramethyl-, which was positively correlated with a roasted and burnt sweet aroma. The above results showed that the characteristic microorganisms from different varieties of CTLs during stacking fermentation might be functional microorganisms enhancing the characteristic aroma of CTLs, which had an important impact on aroma transformation and microbial community interactions. These results have theoretical significance for screening functional strains and producing cigar products with characteristic aromas.
5. Conclusions
In this study, multi-omics technology was used to analyze different CTL varieties in the stacking fermentation process. The commonality and difference changes in microbial communities and volatiles during stacking fermentation were investigated, with the correlation between microorganisms and volatiles was revealed. This provides technical support for the optimization of the stacking fermentation process whilst improving the quality of cigars. This study found that the dominant microorganisms in the process of cigar stacking fermentation were Staphylococcus, Corynebacterium 1, Aerococcus, and Aspergillus, which mainly affected the composition and community succession of stacking fermentation through microbial interactions, thereby influencing the aroma composition and aroma of CTLs. Based on the analysis of volatiles between raw materials and the end of fermentation for different varieties of CTLs, it was found that Maillard reaction products, carotenoid degradation products, and nicotine degradation products were used as the key aroma components for sensory quality improvement and aroma enhancement during the stacking fermentation process. This provides a basis for improving the quality of cigars and the controllability of cigar proportioning in industrial production. In this study, the interaction network analysis of characteristic microorganisms and volatiles was used for the screening of functional microorganisms and the development and application of microbial agents to improve the aroma quality and characteristic aromatic cigar production.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Q.W. and Z.P. designed and performed most of the study, analyzed the data, and wrote the paper. J.Z., Y.P. and L. L. participated in the tests and data analysis. J.Z., L. L., J. W. and Z.P. provided comments on the first draft of the paper. Y.P., L. L., L. L. and J. W. analyzed the sensory quality. Z.P. and J.Z. designed the study and drafted the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the China National Tobacco Technology Development Corporation Project (2021JCYL3SX2B011),.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kaur, G.; Muthumalage, T.; Rahman, I. Mechanisms of toxicity and biomarkers of flavoring and flavor enhancing chemicals in emerging tobacco and non-tobacco products. Toxicol. Lett. 2018, 288, 143–155. [Google Scholar] [CrossRef]
- Spatarella, A.; Folan, P.; Farber, H.J.; American Thoracic Society Tobacco Action, C. Cigars. Am. J. Respir. Crit. Care Med. 2018, 197, P7–P8. [Google Scholar] [CrossRef]
- Gupta, C. A Biotechnological Approach to Microbial Based Perfumes and Flavours. J. Microbiol. Exp. 2015, 2, 11–18. [Google Scholar] [CrossRef]
- Krusemann, E.J.Z.; Pennings, J.L.A.; Cremers, J.; Bakker, F.; Boesveldt, S.; Talhout, R. GC-MS analysis of e-cigarette refill solutions: A comparison of flavoring composition between flavor categories. J. Pharm. Biomed. Anal. 2020, 188, 113364. [Google Scholar] [CrossRef]
- Banozic, M.; Jokic, S.; Ackar, D.; Blazic, M.; Subaric, D. Carbohydrates-Key Players in Tobacco Aroma Formation and Quality Determination. Molecules 2020, 25. [Google Scholar] [CrossRef]
- ZHANG, L.; LUO, Z.-H.; YANG, M.-C. Diversity of Fermentation Microbes and Changes of Hydrolytic Enzyme Activities of Cigar Leaf Raw Materials. J. Agric. Sci. Technol. 2021, 23, 171–180. (In Chinese) [Google Scholar] [CrossRef]
- LIU, F.-F.; FANG, X.; LI, L.-L.; TAN, Z.-Y.; YE, M.-Q.; ZHU, T.-T.; HUANG, Y.-Y.; PAN, Y. . Changs of Volatiles Components in Cigar Wrapper During Deep Fermentation. Guangdong Agric. Sci. 2022, 49, 158–164. (In Chinese) [Google Scholar] [CrossRef]
- Liu, T.; Guo, S.; Wu, C.; Zhang, R.; Zhong, Q.; Shi, H.; Zhou, R.; Qin, Y.; Jin, Y. Phyllosphere microbial community of cigar tobacco and its corresponding metabolites. Front. Microbiol. 2022, 13, 1025881. [Google Scholar] [CrossRef]
- Zheng, T.; Zhang, Q.; Li, P.; Wu, X.; Liu, Y.; Yang, Z.; Li, D.; Zhang, J.; Du, G. Analysis of Microbial Community, Volatile Flavor Compounds, and Flavor of Cigar Tobacco Leaves From Different Regions. Front. Microbiol. 2022, 13, 907270. [Google Scholar] [CrossRef]
- ZHANG, Q.-Y.; LUO, C.; LI, D.-L.; CAI, W. . Research Progress in Curing and Fermentation Technology for Cigar Tobacco Leaf Production. Acta Tabacaria Sin. 2020, 26, 1–6. (In Chinese) [Google Scholar] [CrossRef]
- ZHOU, T.; CAI, J.-Y.; XING, L.; ZHOU, W.; JIA, Y.-H.; GAO, J. . Exploration and Refletions on the Application of the Great Wall Cigar to Dometic Cigar Tobacco. Mod. Ind. Econ. Informationization 2021, 11, 140–142. (In Chinese) [Google Scholar] [CrossRef]
- Wu, X.; Zhu, P.; Li, D.; Zheng, T.; Cai, W.; Li, J.; Zhang, B.; Zhu, B.; Zhang, J.; Du, G. Bioaugmentation of Bacillus amyloliquefaciens-Bacillus kochii co-cultivation to improve sensory quality of flue-cured tobacco. Arch. Microbiol. 2021, 203, 5723–5733. [Google Scholar] [CrossRef]
- Zheng, T.; Zhang, Q.; Wu, Q.; Li, D.; Wu, X.; Li, P.; Zhou, Q.; Cai, W.; Zhang, J.; Du, G. Effects of Inoculation With Acinetobacter on Fermentation of Cigar Tobacco Leaves. Front. Microbiol. 2022, 13, 911791. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Wu, X.; Cai, W.; Zhu, P.; Peng, Z.; Zheng, T.; Li, D.; Li, J.; Zhou, G.; Du, G.; Zhang, J. Profiling the role of microorganisms in quality improvement of the aged flue-cured tobacco. BMC Microbiol. 2022, 22, 197. [Google Scholar] [CrossRef]
- Yu, S.; Drton, M.; Promislow, D.E.L.; Shojaie, A. CorDiffViz: an R package for visualizing multi-omics differential correlation networks. BMC Bioinform. 2021, 22, 486. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Ruiz-Perez, D.; Guan, H.; Madhivanan, P.; Mathee, K.; Narasimhan, G. So you think you can PLS-DA? BMC Bioinform. 2020, 21, 2. [Google Scholar] [CrossRef]
- LIU, X.-Y.; ZHU, Q.; YANG, F.; ZHANG, J.; ZHANG, Q.-L.; LI, J.-H.; WANG, L. Muti-omics Reveal the Formation of Flavor Compounds in Sauce-flavor Daqu. Food Ferment. Ind. 2021, 47, 35–41. (In Chinese) [Google Scholar]
- ZHENG, T.-F.; ZHANG, Q.-Y.; LI, D.-L.; ZHANG, J.; DU, G.-C. . Analysis of Flavor Characteristics and Microbial Community of Cigar Tobacco Leaves from Different Regions. J. Henan Agric. Sci. 2022, 51, 45–54. (In Chinese) [Google Scholar] [CrossRef]
- YAN, T.-J.; ZHENG, L.-L.; LI, X.-N.; LIU, L.-P.; PAN, Y.; WANG, J.; LONG, F.; ZHOU, P.; SHI, X.-D. Diversity Analysis and Function Prediction of Bacterial Communities in Cigar Leaves Based on High-throughput Sequencing Technology. Acta Tabacaria Sin. 2022, 28, 8. (In Chinese) [Google Scholar] [CrossRef]
- Popova, V.; Ivanova, T.; Prokopov, T.; Nikolova, M.; Stoyanova, A.; Zheljazkov, V.D. Carotenoid-Related Volatile Compounds of Tobacco (Nicotiana tabacum L.) Essential Oils. Molecules 2019, 24, 3446. [Google Scholar] [CrossRef]
- Liang, M.H.; He, Y.J.; Liu, D.M.; Jiang, J.G. Regulation of carotenoid degradation and production of apocarotenoids in natural and engineered organisms. Crit. Rev. Biotechnol. 2021, 41, 513–534. [Google Scholar] [CrossRef]
- Mortzfeld, F.B.; Hashem, C.; Vranková, K.; Winkler, M.; Rudroff, F. Pyrazines: Synthesis and Industrial Application of these Valuable Flavor and Fragrance Compounds. Biotechnol. J. 2020, 15, 2000064. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, X.; Song, S.; Hayat, K.; Eric, K.; Majeed, H. Tobacco alkaloids reduction by casings added/enzymatic hydrolysis treatments assessed through PLSR analysis. Regul. Toxicol. Pharmacol. 2016, 75, 27–34. [Google Scholar] [CrossRef]
- ZHENG, L.-L.; ZHAO, L.; CAI, X.-H.; CHEN, Z.; CHAI, Z.-S.; SHI, X.-D. . Analysis of Bacterial and Fungal Community Diversity During Indusrial Secondary Fermentation of Cigar Core Tobacco Leaves. Acta Tabacaria Sin. 2022, 1–9. (In Chinese) [Google Scholar] [CrossRef]
- Mokashe, N.; Chaudhari, A.; Patil, U. Optimal production and characterization of alkaline protease from newly isolated halotolerant Jeotgalicoccus sp. Biocatal. Agric. Biotechnol. 2015, 4, 235–243. [Google Scholar] [CrossRef]
- Xiong, Y.-W.; Gong, Y.; Li, X.-W.; Chen, P.; Ju, X.-Y.; Zhang, C.-M.; Yuan, B.; Lv, Z.-P.; Xing, K.; Qin, S. Enhancement of growth and salt tolerance of tomato seedlings by a natural halotolerant actinobacterium Glutamicibacter halophytocola KLBMP 5180 isolated from a coastal halophyte. Plant Soil 2019, 445, 307–322. [Google Scholar] [CrossRef]
- Zhang, X.; Shan, T.; Jia, H.; Guo, C.; Wang, Z.; Yue, T.; Yuan, Y. Comparative evaluation of the effects of natural and artificial inoculation on soybean paste fermentation. Lwt 2022, 155. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, L.; Zhang, J.; Liu, J.; Zou, X. Dynamic characteristics and co-occurrence patterns of microbial community in tobacco leaves during the 24-month aging process. Ann. Microbiol. 2021, 71, 1–13. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, R.Y.; Yang, F.W.; Xie, Y.F.; Guo, Y.H.; Yao, W.R.; Zhou, W.B.A. Control strategies of pyrazines generation from Maillard reaction. Trends Food Sci. Tech. 2021, 112, 795–807. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, Y.; Tong, J.; Xu, Y. An Alkylpyrazine Synthesis Mechanism Involving l-Threonine-3-Dehydrogenase Describes the Production of 2,5-Dimethylpyrazine and 2,3,5-Trimethylpyrazine by Bacillus subtilis. Appl. Environ. Microbiol. 2019, 85, e01807–e01819. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



