
Submitted:
22 February 2023
Posted:
23 February 2023
You are already at the latest version
Abstract
Plants have acquired sets of highly regulated and complex signaling pathways to respond to unfavorable environmental conditions during evolution. Calcium signaling, as a vital mechanism, enables plants to respond to external stimuli, including abiotic and biotic stresses, and coordinate the basic processes of the growth and development. In the present study, the calcium sensor families, including CBL and CIPK, were investigated in the halophyte plant, Aeluropus littoralis, with a comprehensive analysis. Here, six AlCBL genes, and twenty AlCIPK genes were presented. The analysis of the gene structure and conserved motifs, as well as physicochemical properties, showed that these genes are highly conserved during evolution. The expression levels of AlCBL genes and AlCIPK genes were evaluated under salt stress in leaf and root tissue. Based on the real-time RT-PCR results, AlCIPK gene family had a higher variation in mRNA abundance compared to the AlCBL gene family. The AlCIPKs were found to have a higher abundance in leaves than in roots. The results suggest that the interaction pattern of AlCBL genes with AlCIPK is tissue-specific, and different interactions can be expected in leaves and roots. Based on these patterns AlCIPK3.1 - AlCBL4.1 and AlCIPK1.2 - AlCBL4.4 can interact in root tissue, while the AlCBL10 has the potential to interact with the AlCIPK5, AlCIPK26 and AlCIPK12.3 in the leaf tissue. These findings provide valuable information on the structure and function of calcium sensor families in Aeluropus littoralis a halophyte plant, for future research on the biological function of CBLs and CIPKs on salt stress resistance.
Keywords:
Calcium sensors
; CBL
; CIPK
; Salt stress
; Kinases
; Cell signaling
1. Introduction
In sessile organisms such as plants, perception and signaling of environmental stimuli is necessary for survival and growth regulation. Calcium (Ca2+) is one of the signal transduction components that acts as a second messenger in all eukaryotes [1,2,3]. Ca2+ is stored in organelles such as vacuoles, mitochondria and endoplasmic reticulum, where abiotic stresses such as salt, cold and drought cause rapid increase of Ca2+ concentration in the cytosol [3,4,5,6]. However, biotic stresses, pH dynamics, and phytohormones also can affect the Ca2+ concentration [7,8,9,10]. In addition, pollen tube development and guard cell regulation are also associated with changes in Ca2+ concentration [8]. Calcium sensors or calcium-binding proteins recognize the modification in Ca2+ concentrations in plant cell, and downstream pathways are induced by affecting the phosphorylation status of calcium sensors and activating protein kinases [11,12]. Calmodulin (CaM), calcium-dependent protein kinases (CDPKs), and calcineurin B-like (CBLs) are part of the known calcium sensors in plants [13]. CBLs are plant-specific sensors that, after sensing a specific calcium signature, can physically interact with a protein kinases, CBL-interacting protein kinases (CIPKs), to activate downstream signaling components [14,15,16]. CBL proteins share a common helix-loop-helix structural motif (the EF-hand), where acts as Ca2+ binding region [17]. Besides, it seems that the EF-hand composition could affect the affinity rate of calcium ions [17].
In the plant model system Arabidopsis diverse roles were reported for CBLs: The cbl1 mutant was very sensitive to the abiotic stresses such as drought, extreme salinity, and hyperosmotic stress. Likewise, CBL9 gene is involved in ABA signal transduction and stress-induced ABA biosynthesis pathways [18]. In addition, it was reported that CBL9 and CBL1 are participated in pollen germination and flower fertilization [19]. Furthermore, it was stated that CBL1 is involved in response to aluminum stress [20], cold stress [21,22]. Moreover, CBL7 is associated with Arabidopsis responses to alkaline stress [23]. Interestingly, it was reported that CLBs, such as CBL3 and CBL4, could modulate the potassium channel and affect the potassium homeostasis [21,24]. It has also been found that the expression patterns of the CBL genes are dependent on the tissues and developmental stages and the type of stress. For example, CBL1 expression is not affected by external application of abscisic acid (ABA), but is induced in response to environmental stresses such as salt, cold, drought, and wounding [25]. While CBL2 and CBL3 do not respond to abiotic stress stimuli, they are transcriptionally induced by light stress [26]. CIPK genes also have differential expression patterns. For example, CIPK9 transcriptional regulation is more induced in response to ABA treatment, and is mainly activated in shoot tissues [27]. In addition, CIPK genes in Medicago truncatula, including MtCIPK2, MtCIPK17, and MtCIPK18 were found to be upregulated in response to salinity, PEG and ABA treatments [28]. Recently, it has been reported that a CIPK gene from chrysanthemum, CmCIPK8, could affect the expression patterns of ion transport-related genes, and may enhance tolerance to salinity [29]. Moreover, CIPK10 in potato (StCIPK10) could increase tolerance to osmotic and drought stress by affecting the content of osmoregulation substances [30]. Also, it was reported that StCIPK10 can interact with several StCBLs, including StCBL4, StCBL8, StCBL1, StCBL6, StCBL12, and StCBL11 [30]. In Beta vulgaris it was described that, BvCIPKs, are upregulated in response to NaCl treatment [31]. In Saccharum spontaneum, CIPK genes were shown to respond to abiotic stresses as cold and water stress, and ABA treatment [32]. Overall, it seems that cell signaling networks linked with CBL-CIPK play critical roles in response to abiotic stresses [33,34,35].
Aeluropus littoralis as a halophyte model can grow under high salt concentrations [36,37]. Identifying genes related to resistance in plants such as A. littoralis, as a valuable germplasm, and determining their function can provide a better understanding of resistance mechanisms in plants [38]. According to the mentioned materials above, the genes of the CBL and CIPK family play a key role in responding to environmental stresses and regulating downstream signaling pathways, but these gene families have not been identified and investigated in A. littoralis. Here, we identified the members of CBL and CIPK families and analyzed the structure and evolution as well as their regulatory systems. In addition, the expression profile of AlCBL and AlCIPK genes were evaluated under salinity in root and leaf tissues of A. littoralis.
2. Materials and Methods
2.1. Identification of CBL and CIPK family genes in A. littoralis
In this study, the putative protein sequences of CBL and CIPK in rice were retrieved from the RGAP database (http://rice.plantbiology.msu.edu/) and for Arabidopsis thaliana from TAIR database (https://www.arabidopsis.org/). Sequences were used as query in blastp and tblastn tool, E-value< 1e-10 to identify members of the CBL and CIPK gene families from the transcriptome platform e!DAL of A. littoralis [39].The presence of PKinase- and NAF-domains in CIPK proteins, as well as EF-hand domains in CBL proteins was tested and confirmed using the CDD database [40], SMART [41], and InterPro Scan [42]. The confirmed protein sequences were renamed based on their orthologs in Arabidopsis. Further AlCBL and AlCIP proteins were analyzed by ExPASy online database ProtParam tool [43] to predict the physiochemical properties including molecular weight (MW), GRAVY, and isoelectric point (pI).
2.2. Phylogenetic analysis and classification of AlCBL and AlCIPK gene family
To investigate the evolutionary relationships in the calcium sensor gene families, the protein sequences of AlCBL and AlCIPK families along with their orthologues in Arabidopsis and rice were analyzed. First, the sequences were aligned by the ClustalW tool [44] and then a phylogenetic tree was drawn by IQ tree software [45] using the maximum likelihood (ML) method with 1000 bootstrap replications. Finally, the tree file was restored and upgraded in the iTOL database [46].
2.3. Motif analysis and gene structure of AlCBLs and AlCIPKs
2.4. Plant materials growth conditions and salt treatments
Cultivation of the A. littoralis seeds was carried out at a temperature of 25 ± 3, a photoperiod of 16 hours of light and 8 hours of darkness. Then the cloned samples were transferred to Hoagland's solution and after two months, salt stress treatment was started. In order to apply salinity stress, sodium chloride was gradually added; 100 mM salt every 3 days was added to solution until the final concentration reached 600 mM. Sampling of leaf and root tissues was done in the time series of 0 (as a control sample), 3, 12, 24 hours after the application of salt stress. The collected samples were kept in a freezer at -80 for the next steps. All experiments were performed in three biological replications.
2.5. RNA extraction and cDNA synthesis
Extraction of total RNA from leaf and root tissues was done using the Trizol kit (Threezol, Riragene). To remove genomic DNA from RNA, DNase I treatment (DNase I RNase-free, Thermo Scientific) was applied. Finally, cDNA was synthesized using a RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific) based on the company's instructions, and diluted four times.
2.6. Real-time PCR
In the present study, the levels of mRNA abundance from six AlCBL and twelve AlCIPK genes were investigated in two tissues, roots and leaves, under salinity and normal conditions. Genes were selected based on phylogenetic analysis. Primers of candidate genes were designed using AlleleID [49] (Tables S1 and S2). The Maxima SYBR Green/ROX qPCR Master Mix (Thermo Scientific) was used to evaluate the relative expression based on manufacturer's instructionswith a Bio-Rad CFX96 machine. The temperature cycle was performed in two stages according to the manufacturer's instructions: 10 minutes activation stage at 95 ºC, 40 cycles at 95 ºC for 15 seconds, and 60 ºC for 1 minute. In the current study, three reference genes, including AlUBQ, AlRPS3, and AlRPS3, for each tissue were used. The geometric mean of these genes was used to normalize the data. Finally, the relative expression levels of each target gene was calculated using the 2-△△CT method [50].
3. Results
3.1. Physicochemical properties of AlCBLs and AlCIPKs
In the present study, six AlCBL genes and twenty AlCIPK genes are identified in the genome of A. littoralis. The evaluation of the physicochemical characteristics of CBL proteins revealed variable molecular weight in the range of 18.70 (AlCBL 4.4) to 34.67 kDa (AlCBL 10), and all AlCBLs were predicted as acidophilic proteins, pI less than 5.5 (Table 1). Furthermore, all AlCBLs (except for AlCBL 10 protein) had negative GRAVY value, revealing that most AlCBLs have hydrophilic properties. In general, based on the physicochemical characteristics, AlCBL 10 protein was different from other members of the AlCBL gene family, which can be more considered in molecular functional research. According to the physicochemical characteristics, AlCIPK family members showed more diversity than AlCBLs. Molecular weight in AlCIPKs ranged of 42.04 (AlCIPK 10.6) to 58.9 kDa (AlCIPK 10.1), and pI varied from 6.21 (AlCIPK 21) to 9.28 (AlCIPK 10.2).
3.2. Phylogenetic analysis of AlCBLs
AlCBL proteins along with their orthologues in rice and Arabidopsis were subjected to a phylogenetic analysis. Results disclosed that CBL proteins could be classified in four main groups (Figure 1). None of AlCBLs could be identified in group I. AlLAC 4.1, AlLAC 4.2, AlLAC 4.3, and AlLAC 4.4 were located in group II, AlLAC 10 in group III, and AlLAC 2 in group IV. In addition, AlCBLs and rice CBLs showed more similarity to each other that Arabidopsis CBLs. Overall, our results propose that the diversity in CBL family has been occurred after the splitting of monocots and dicots.
3.3. Phylogenetic analysis of AlCIPKs
To determine the evolutionary origin of AlCIPKs, the phylogenetic tree of AlCIPKs with their orthologues in Arabidopsis (26 CIPK proteins) and rice (33 CIPK proteins) was made based on protein sequences (Figure 2). Results illustrated that CIPKs could be separated in four main groups. The highest number of CIPKs were found in group III, and the lowest number were observed in group IV. Similar to AlCBLs, AlCIPKs also showed a close relationship with rice CIPKs. In addition, it can be concluded that probably the expansion in the CIPK family has been occurred after the derivation of monocots and dicots.
3.4. Gene structure and conserved motifs of AlCBLs
AlCBLs with their orthologous in rice (OsCBLs) were analyzed based on the conserved motifs and domains, and gene structure (Figure 3). Ten conserved motifs were recognized in AlCBLs and OsCBLs that motifs 6 and 9 were not detected in CBLs from group II and motif 9 was only observed in OsCBLs from group III (Figure 3a). Calcium binding superfamily, EF-hand 7, EF-hand 5 and EF-hand 1 domains were observed in AlCBLs and OsCBLs, although they differed based on the location and number of domains (Figure 3b). In addition, two copies of EF-hand 7 and EF-hand 1 domains were found into AlCBL 10 and its orthologue, OsCBL 9, suggesting that AlCBL 10 probably has more potential to interact with downstream elements of involved pathways. Besides, AlCBLs were different based on gene structure and all AlCBLs had a high number of exons/introns (Figure 3c).
3.5. Gene structure and conserved motifs of AlCIPKs
To identify conserved motifs and determine the position of these motifs in KINAS and NAF domains, AlCIPK proteins with their orthologues in rice (OsCIPKs) were analyzed using MEME tool (Figure 4a). The results show that the spatial distribution of the motifs in the investigated proteins is strongly conserved. All ten identified motifs were observed in AlCIPK3.1, AlCIPK3.2, AlCIPK10.1, AlCIPK10.2, AlCIPK10.4, AlCIPK20, AlCIPK23 and AlCIPK26 proteins, while motif 10 was not detectable in AlCIPK1.1, AlCIPK12.1, AlCIPK12.2 and AlCIPK12.3. Motif 5 was not detected in AlCIPK4, motif 4 in AlCIPK5, motif 1 in AlCIPK10.3 and motif 2 in AlCIPK10.5. In AlCIPK1.2 protein, motifs 10 and 3, in AlCIPK11 protein, motifs 10 and 6, in AlCIPK10.6 protein, motifs 3 and 8, and in AlCIPK21 protein, motifs 10 and 4 are not present. This result indicates that the main (conserved) motifs play an important role in the function of CIPK proteins. Besides, two KINAS and NAF domains were identified in AlCIPKs and OsCIPKs (Figure 4b), that all studied proteins showed one copy of KINAS and NAF domain. Based on gene structure analysis, 60% of AlCIPK genes have one exon and no intron, about 20% of genes have 14 exons and 13 introns, and about 10% of genes have 2 exons and 1 intron, about 5% of genes have 13 exons and 12 introns, and about 5% of genes have 12 exonsand 11 are introns (Figure 4c).
3.6. Promoter analysis
In the present study, the upstream of AlCIPKs and AlCBLswas screened to identify the cis-regulatory elements related to stress, hormone and growth and development (Figure 5). Most recognized elements were related to common cis-regulatory and elements with unknown function (Figure 5a). Besides, putative cis-regulatoryelements related to transcription factors binding site, and response to phytohormones, and stresses were observed in upstream site of AlCIPKs and AlCBLs. Cis-regulatory elements involved in response to ABA hormone were frequently identified in promoter site of AlCIPKs and AlCBLs (Figure 5b). Besides, the putative cis-regulatory elements related to GA, auxin, SA and MeJA hormones were recognized in upstream site of AlCIPKs, while in AlCBLs, regulatory elements responding to GA and MeJA hormones were observed. Cis-regulatory elements involved in responsive to abiotic stresses, including low-temperature, MBS, DRE, and STRE, and biotic stresses including wound, elicitor and defense were identified in promoter region of AlCBLs and ALCIPKs (Figure 5c). In addition, the binding site of several TFs such as MYB, MYS, and WRKY were observed in upstream site of AlCIPKs and AlCBLs (Figure 5d). In general, AlCIPKs were richer than AlCBLs based on the number of stress-related cis elements.
3.7. Expression profile of AlCBL genes in response to salinity
Expression levels of AlCBL genes were investigated under salinity in root and leaf tissues. According to our results, AlCBL2 was not expressed under the tested conditions. After 3h of salinity treatment, AlCBL4.1, AlCBL4.2, and AlCBL4.4 showed an upregulation in root tissues (Figure 6). Three AlCBL genes, including AlCBL4.3, AlCBL4.4, and AlCBL10, were highly induced after 24h that all three genes were upregulated in root while they were down-regulated in leaf.
3.8. Expression profile of AlCIPK genes in response to salinity
In the root tissue, the expression levels of AlCIPK1.2 (1.96 time), AlCIPK3.1 (4.90 times), AlCIPK5 (2.32 times), AlCIPK11 (4.21 times), AlCIPK12.1 (2.62 times) and AlCIPK26 (4.63 times) were increased after three hours (hr) applying salt stress (Figure 7). In the leaf tissue at 3 hours after applying salt stress, AlCIPK11 (4.10 times), AlCIPK1.2 (2.89 times), AlCIPK4 (1.82 times), AlCIPK12.3 (1.77 times), AlCIPK5 (1.69 times), AlCIPK10.2 (-2.50 times), AlCIPK10.6 (-1.91 times) were more induced. In the root tissue, after 12 hr of salinity, AlCIPK10.2 gene (-4.46 times) just showed a sharp downregulation, while in leaf tissues, AlCIPK4 (3.43 times), AlCIPK10.2 (-6.39 times), AlCIPK11 (3.72 times), AlCIPK1.2 (2.00 times), AlCIPK3.1 (-1.82 times) and AlCIPK5 (2.13 times), AlCIPK10.6 (-1.34 times), AlCIPK12.1 (1.58 times); AlCIPK26 (2.00 times) and AlCIPK12.3 (2.61 times) showed a significant modification in their expression levels after 12 hr. Interestingly, AlCIPK10.2 was notably downregulated in both root and leaf tissues. In addition, the expression levels of AlCIPK4 (3.47 times) and AlCIPK12.3 (3.78 times) were increased in leaf tissue after 24 hours. In total, CIPKs were more expressed in leaf tissue, while AlCIPK12.3 was expressed only in leaf tissue and AlCIPK1.1 gene expression was observed only in root tissue. AlCIPK4, AlCIPK5, AlCIPK10.2, AlCIPK10.6, AlCIPK11, and AlCIPK12.3 genes were significantly expressed in leaf tissue at all times of stress.
4. Discussion
Calcium sensors such as calcineurin B-like proteins (CBL) and CBL-interacting protein kinases (CIPKs) not only participate in the processes of plant growth and development, but they are also involved in stress responses [30]. In the present study, the available genome of A. littoralis was used as resource [38] and screened for the respective gene families. Six AlCBL genes and twenty AlCIPK genes were identified. Due to the importance of calcium-dependent signaling pathways, these gene families, CBL and CIPK, have been studied in various plants. 23 CBLs and 58 CIPK genes were identified from the genome of Medicago sativa [28], 27 CIPK genes from potato [30], 9 CBLs and 30 CIPK genes from the pecan genome [51], 10 CBLs and 26 CIPKs from Arabidopsis [17], 7 CBLs and 20 CIPK genes from bread wheat [52], 7 CBLs and 23 CIPK genes from canola [53], 16 CBLs and 41 CIPK genes from quinoa [35], and 20 CIPK genes from sugar beet [31]. The different number of members of this gene family suggests that they may have been subjected to evolutionary pressures differently in each plant [54,55]. Based on the physicochemical properties, AlCBL proteins have similar properties, except for AlCBL10 protein. The proteins of the AlCIPK family show more diversity. This result support the hypothesis that AlCIPKs are highly diverse due to their involvement in different pathways [56,57]. While AlCIPKs showed a high variation in term of gene structure the AlCIPKs can be separated into two groups. This grouping is also reflected by their low intron number (less than three introns) and high intron number (more than ten introns). Moreover, this feature has also been reported in previous studies where CIPKs have been classified into two groups based on their intron structure [35,58]. It was stated that partial duplication probably has affected the intron number of gene family members [59]. Besides, it was reported that expression levels of genes can be affected by intron number and genes with low intron number could be faster induced [60]. According to phylogenetic analysis, both AlCBL family and AlCIPK family are closer related to their rice orthologs ., This finding suggest that the diversity in these gene families occurred after derivation of monocots and dicots species [61,62].
Halophyte plants have a high potential to grow in substrates of high salinity. Therefore these species are of great interest to investigating the mechanisms of tolerance to salinity. This aims at mechanisms such as absorption, transport and homeostasis of ions, osmotic regulation and salt removal from leaves [37,38]. Although the cultivation of these plants is not an easy task, while the germplasm of halophyte plants is considered as a valuable source for providing genes resistant to environmental conditions, for the implementation of plant breeding programs [63]. In the current study, the expression profiles of AlCBLs and AlCIPKs were investigated under salt stresses in roots and leaves of A. littoralis. AlCBLs and AlCIPKs showed tissue specific expression patterns. For instance, AlCIPKs mRNAs were higher abundant in leaves than in roots, while AlCBL4.3, AlCBL4.4, and AlCBL10 showed upregulation in root and downregulation in shoot. This pattern might be related to the presence of as-1 specific motifs in the promoter region of AlCBL genes. Each of AlCBL4.3, AlCBL4.1, and AlCBL2 genes had two as-1 motifs, while six as-1 motifs in AlCBL4.2, and three as-1 motifs in AlCBL4.4 and an as-1 motif 3 motifs were observed in promoter region.
The results revealed that the interaction pattern of AlCBL proteins with AlCIPK was tissue-specific and different interactions were observed in two tissues of leaves and roots. Based on the expression pattern, AlCIPK3.1-AlCBL4.1 and AlCIPK1.2-AlCBL4.4 genes can potentially interact in root tissue, while in leaf tissue AlCBL10 gene can interact with AlCIPK5, AlCIPK12.3 and AlCIPK26 genes. Positive correlation was reported between CBLs and CIPKs in response to stresses, such as salinity [64], drought [65], and disease [58]. In Arabidopsis, the interaction between CBL4 (called SOS3) and CIPK24 (called SOS2) could active the kinases and +/H+ antiporters called SOS1 and vacuolar H+-ATPase to increase stress tolerance [53,66,67]. Subsequent research in Arabidopsis showed that the AtCBL10 gene also interacts with AtCIPK24. Thus, the CBL10-CIPK24 complex interacts with vacuoles to protect the shoot from damage caused by salt stress [67]. This result suggests the fact that calcium sensors may exhibit very different functions despite high sequence similarity or close evolutionary relationship.
5. Conclusions
This review is the first comprehensive study of the family of calcium sensors with the aim of clarifying the evolution, expression patterns and possible functions of the genes of this superfamily in A. littoralis in response to abiotic stresses. These findings provide information to predict the function of calcium sensor genes in plant tolerance to salt stress. Additional studies on the expression of AlCBL and AlCIPK family genes under different abiotic stresses in future research can be useful in understanding the mechanism of gene expression adjustments related to the SOS pathway. The AlCIPK genes reported in this research, while providing preliminary information, provide a basis for identifying the functions and mechanisms of the stress response, especially the responses related to the CBL / CIPK pathway in the A. littoralis plant.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: List of primers of AlCBL genes used in qPCR analysis; Table S2: List of primers of AlCIPK genes used in qPCR analysis.
Author Contributions
Conceptualization, M.A., S.H.H., H.N.Z. and G.N.; methodology, M.A., S.H.H. and P.H.; software, M.A. and S.H.H.; validation, S.H.H., H.N.Z. and G.N.; formal analysis, S.H.H.; investigation, P.H. and M.K.; writing—original draft preparation, P.H.; writing—review and editing, P.H. and M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
Costs for open access publishing were partially funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation, grant 491250510).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.-J.; Luan, S. Regulation of calcium and magnesium homeostasis in plants: from transporters to signaling network. Curr. Opin. Plant Biol. 2017, 39, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Fakher, B.; Jakada, B.H.; Zhao, L.; Cao, S.; Cheng, Y.; Qin, Y. Genome-Wide Identification and Expression Profiling of CBL-CIPK Gene Family in Pineapple (Ananas comosus) and the Role of Ac CBL1 in Abiotic and Biotic Stress Response. Biomolecules 2019, 9, 293. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Navazio, L.; Szabo, I. The contribution of organelles to plant intracellular calcium signalling. J. Exp. Bot. 2018, 69, 4175–4193. [Google Scholar] [CrossRef]
- Kudla, J.; Becker, D.; Grill, E.; Hedrich, R.; Hippler, M.; Kummer, U.; Parniske, M.; Romeis, T.; Schumacher, K. Advances and current challenges in calcium signaling. New Phytol. 2018, 218, 414–431. [Google Scholar] [CrossRef]
- Ding, Y.; Shi, Y.; Yang, S. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef]
- Behera, S.; Xu, Z.; Luoni, L.; Bonza, M.C.; Doccula, F.G.; De Michelis, M.I.; Morris, R.J.; Schwarzländer, M.; Costa, A. Cellular Ca2+ signals generate defined pH signatures in plants. Plant Cell 2018, 30, 2704–2719. [Google Scholar] [CrossRef] [PubMed]
- Michard, E.; Simon, A.A.; Tavares, B.; Wudick, M.M.; Feijó, J.A. Signaling with ions: the keystone for apical cell growth and morphogenesis in pollen tubes. Plant Physiol. 2017, 173, 91–111. [Google Scholar] [CrossRef] [PubMed]
- De Vriese, K.; Himschoot, E.; Dünser, K.; Nguyen, L.; Drozdzecki, A.; Costa, A.; Nowack, M.K.; Kleine-Vehn, J.; Audenaert, D.; Beeckman, T. Identification of novel inhibitors of auxin-induced Ca2+ signaling via a plant-based chemical screen. Plant Physiol. 2019, 180, 480–496. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.K.; Prajapati, R.; Krishna, D.; Divakaran, K.; Pandey, Y.; Reichelt, M.; Mathew, M.K.; Boland, W.; Mithöfer, A.; Vadassery, J. The Ca2+ channel CNGC19 regulates Arabidopsis defense against Spodoptera herbivory. Plant Cell 2019, 31, 1539–1562. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Bender, K.W.; Zielinski, R.E.; Huber, S.C. Revisiting paradigms of Ca2+ signaling protein kinase regulation in plants. Biochem. J. 2018, 475, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.-P.; Thuleau, P.; Mazars, C. Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lv, Y.; Jahan, N.; Chen, G.; Ren, D.; Guo, L. Sensing of abiotic stress and ionic stress responses in plants. Int. J. Mol. Sci. 2018, 19, 3298. [Google Scholar] [CrossRef] [PubMed]
- Batistic, O.; Kudla, J. Integration and channeling of calcium signaling through the CBL calcium sensor/CIPK protein kinase network. Planta 2004, 219, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Halfter, U.; Ishitani, M.; Zhu, J.-K. The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc. Natl. Acad. Sci. 2000, 97, 3735–3740. [Google Scholar] [CrossRef] [PubMed]
- Kolukisaoglu, U.; Weinl, S.; Blazevic, D.; Batistic, O.; Kudla, J. Calcium sensors and their interacting protein kinases: genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol. 2004, 134, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.K.; Cheong, Y.H.; Kim, K.-N.; Grant, J.J.; Li, L.; Hung, W.; D’Angelo, C.; Weinl, S.; Kudla, J.; Luan, S. The calcium sensor calcineurin B-like 9 modulates abscisic acid sensitivity and biosynthesis in Arabidopsis. Plant Cell 2004, 16, 1912–1924. [Google Scholar] [CrossRef] [PubMed]
- Mähs, A.; Steinhorst, L.; Han, J.-P.; Shen, L.-K.; Wang, Y.; Kudla, J. The calcineurin B-like Ca2+ sensors CBL1 and CBL9 function in pollen germination and pollen tube growth in Arabidopsis. Mol. Plant 2013, 6, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Ligaba-Osena, A.; Fei, Z.; Liu, J.; Xu, Y.; Shaff, J.; Lee, S.; Luan, S.; Kudla, J.; Kochian, L.; Piñeros, M. Loss-of-function mutation of the calcium sensor CBL 1 increases aluminum sensitivity in Arabidopsis. New Phytol. 2017, 214, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-L.; Ren, H.-M.; Chen, L.-Q.; Wang, Y.; Wu, W.-H. A protein kinase, calcineurin B-like protein-interacting protein Kinase9, interacts with calcium sensor calcineurin B-like Protein3 and regulates potassium homeostasis under low-potassium stress in Arabidopsis. Plant Physiol. 2013, 161, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Ding, S.; Zhang, H.; Du, H.; An, L. CIPK7 is involved in cold response by interacting with CBL1 in Arabidopsis thaliana. Plant Sci. 2011, 181, 57–64. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Y.; Ma, L.; Yang, Z.; Dong, Q.; Li, Q.; Ni, X.; Kudla, J.; Song, C.P.; Guo, Y. The Ca2+ sensor SCaBP3/CBL7 fine tunes arabidopsis alkali tolerance and modulats plasma membrane H+-ATPase activity. Plant Cell 2019, 31, 1367–1384. [Google Scholar] [CrossRef] [PubMed]
- Held, K.; Pascaud, F.; Eckert, C.; Gajdanowicz, P.; Hashimoto, K.; Corratgé-Faillie, C.; Offenborn, J.N.; Lacombe, B.; Dreyer, I.; Thibaud, J.-B. Calcium-dependent modulation and plasma membrane targeting of the AKT2 potassium channel by the CBL4/CIPK6 calcium sensor/protein kinase complex. Cell Res. 2011, 21, 1116–1130. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, V.; Weinl, S.; Blazevic, D.; D’Angelo, C.; Batistic, O.; Kolukisaoglu, Ü.; Bock, R.; Schulz, B.; Harter, K.; Kudla, J. The calcium sensor CBL1 integrates plant responses to abiotic stresses. Plant J. 2003, 36, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, A.; Koizumi, N.; Sano, H. An Arabidopsis SNF1-related protein kinase, AtSR1, interacts with a calcium-binding protein, AtCBL2, of which transcripts respond to light. Plant Cell Physiol. 2001, 42, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Guo, Y.; Jagendorf, A.T.; Zhu, J.-K. Biochemical characterization of the Arabidopsis protein kinase SOS2 that functions in salt tolerance. Plant Physiol. 2002, 130, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Yang, J.; Ma, L.; Su, Q.; Pang, Y. Identification and characterization of abiotic stress responsive CBL-CIPK family genes in Medicago. Int. J. Mol. Sci. 2021, 22, 4634. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Liu, B.; Liu, H.; Sun, X.; Sun, X.; Wang, W.; Zheng, C. A new CIPK gene CmCIPK8 enhances salt tolerance in transgenic chrysanthemum. Sci. Hortic. (Amsterdam). 2023, 308, 111562. [Google Scholar] [CrossRef]
- Ma, R.; Liu, W.; Li, S.; Zhu, X.; Yang, J.; Zhang, N.; Si, H. Genome-Wide Identification, Characterization and Expression Analysis of the CIPK Gene Family in Potato (Solanum tuberosum L.) and the Role of StCIPK10 in Response to Drought and Osmotic Stress. Int. J. Mol. Sci. 2021, 22, 13535. [Google Scholar] [CrossRef]
- Wu, G.-Q.; Xie, L.-L.; Wang, J.-L.; Wang, B.-C.; Li, Z.-Q. Genome-Wide Identification of CIPK Genes in Sugar Beet (Beta vulgaris) and Their Expression Under NaCl Stress. J. Plant Growth Regul. 2022, 1–15. [Google Scholar] [CrossRef]
- Su, W.; Ren, Y.; Wang, D.; Huang, L.; Fu, X.; Ling, H.; Su, Y.; Huang, N.; Tang, H.; Xu, L. New insights into the evolution and functional divergence of the CIPK gene family in Saccharum. BMC Genomics 2020, 21, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Zhang, H.; Xie, F.; Pan, Z.-Y.; Qiu, W.-M.; Tong, Z.; Wang, Z.-Q.; He, X.-J.; Xu, Y.-H.; Sun, Z.-H. Evolution, gene expression, and protein‒protein interaction analyses identify candidate CBL-CIPK signalling networks implicated in stress responses to cold and bacterial infection in citrus. BMC Plant Biol. 2022, 22, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Mo, Z.; Yuan, G.; Xiang, H.; Visser, R.G.F.; Bai, Y.; Liu, H.; Wang, Q.; van der Linden, C.G. The CBL-CIPK network is involved in the physiological crosstalk between plant growth and stress adaptation. Plant. Cell Environ. 2022. [Google Scholar] [CrossRef] [PubMed]
- Xiaolin, Z.; Baoqiang, W.; Xian, W.; Xiaohong, W. Identification of the CIPK-CBL family gene and functional characterization of CqCIPK14 gene under drought stress in quinoa. BMC Genomics 2022, 23, 1–18. [Google Scholar] [CrossRef]
- Saad, R. Ben; Romdhan, W. Ben; Zouari, N.; Azaza, J.; Mieulet, D.; Verdeil, J.-L.; Guiderdoni, E.; Hassairi, A. Promoter of the AlSAP gene from the halophyte grass Aeluropus littoralis directs developmental-regulated, stress-inducible, and organ-specific gene expression in transgenic tobacco. Transgenic Res. 2011, 20, 1003–1018. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.H.; Nematzadeh, G.; Ahmadian, G.; Yamchi, A.; Kuhlmann, M. Identification and validation of Aeluropus littoralis reference genes for Quantitative Real-Time PCR Normalization. J. Biol. Res. 2016, 23, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hashemi-Petroudi, S.H.; Arab, M.; Dolatabadi, B.; Kuo, Y.-T.; Baez, M.A.; Himmelbach, A.; Nematzadeh, G.; Maibody, S.A.M.M.; Schmutzer, T.; Mälzer, M. Initial Description of the Genome of Aeluropus littoralis, a Halophile Grass. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Arend, D.; Lange, M.; Chen, J.; Colmsee, C.; Flemming, S.; Hecht, D.; Scholz, U. e! DAL-a framework to store, share and publish research data. BMC Bioinformatics 2014, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Copley, R.R.; Doerks, T.; Ponting, C.P.; Bork, P. SMART: a web-based tool for the study of genetically mobile domains. Nucleic Acids Res. 2000, 28, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: the proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating Maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Apte, A.; Singh, S. AlleleID. PCR Prim. Des. 2007, 329–345. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Fan, P.; Liu, H.; Tan, P.; Ma, W.; Mo, Z.; Zhao, J.; Chu, G.; Peng, F. Insight into the CBL and CIPK gene families in pecan (Carya illinoinensis): identification, evolution and expression patterns in drought response. BMC Plant Biol. 2022, 22, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, Y.; Wang, M.; Li, T.; Zhou, Y.; Wang, X.; Wei, S.; He, G.; Yang, G. Identification and comprehensive analyses of the CBL and CIPK gene families in wheat (Triticum aestivum L.). BMC Plant Biol. 2015, 15, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, B.; Liu, W.-Z.; Li, H.; Wang, L.; Wang, B.; Deng, M.; Liang, W.; Deyholos, M.K.; Jiang, Y.-Q. Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L.). BMC Plant Biol. 2014, 14, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Ahmadizadeh, M.; Rezaee, S.; Heidari, P. Genome-wide characterization and expression analysis of fatty acid desaturase gene family in Camelina sativa. Gene Reports 2020, 21, 100894. [Google Scholar] [CrossRef]
- Faraji, S.; Filiz, E.; Kazemitabar, S.K.; Vannozzi, A.; Palumbo, F.; Barcaccia, G.; Heidari, P. The AP2/ERF Gene Family in Triticum durum: Genome-Wide Identification and Expression Analysis under Drought and Salinity Stresses. Genes (Basel). 2020, 11, 1464. [Google Scholar] [CrossRef] [PubMed]
- Puresmaeli, F.; Heidari, P.; Lawson, S. Insights into the Sulfate Transporter Gene Family and Its Expression Patterns in Durum Wheat Seedlings under Salinity. Genes (Basel). 2023, 14, 333. [Google Scholar] [CrossRef] [PubMed]
- Yaghobi, M.; Heidari, P. Genome-Wide Analysis of Aquaporin Gene Family in Triticum turgidum and Its Expression Profile in Response to Salt Stress. Genes (Basel). 2023, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yu, A.; Du, Y.; Wang, G.; Li, Y.; Zhao, G.; Wang, X.; Zhang, W.; Cheng, K.; Liu, X. Foxtail millet (Setaria italica (L.) P. Beauv) CIPKs are responsive to ABA and abiotic stresses. PLoS One 2019, 14, e0225091. [Google Scholar] [CrossRef] [PubMed]
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Genome-wide analysis of NAC transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar] [CrossRef]
- Heidari, P.; Puresmaeli, F.; Mora-Poblete, F. Genome-Wide Identification and Molecular Evolution of the Magnesium Transporter (MGT) Gene Family in Citrullus lanatus and Cucumis sativus. Agronomy 2022, 12, 2253. [Google Scholar] [CrossRef]
- Heidari, P.; Abdullah; Faraji, S. ; Poczai, P. Magnesium transporter Gene Family: Genome-Wide Identification and Characterization in Theobroma cacao, Corchorus capsularis and Gossypium hirsutum of Family Malvaceae. Agronomy 2021, 11, 1651. [Google Scholar] [CrossRef]
- Faraji, S.; Ahmadizadeh, M.; Heidari, P. Genome-wide comparative analysis of Mg transporter gene family between Triticum turgidum and Camelina sativa. BioMetals 2021, 4. [Google Scholar] [CrossRef] [PubMed]
- Jaradat, A.A. Genetic resources of energy crops: biological systems to combat climate change. Aust. J. Crop Sci. 2010, 4, 309–323. [Google Scholar]
- Li, J.; Jiang, M.; Ren, L.; Liu, Y.; Chen, H. Identification and characterization of CBL and CIPK gene families in eggplant (Solanum melongena L.). Mol. Genet. Genomics 2016, 291, 1769–1781. [Google Scholar] [CrossRef] [PubMed]
- Shu, B.; Cai, D.; Zhang, F.; Zhang, D.J.; Liu, C.Y.; Wu, Q.S.; Luo, C. Identifying citrus CBL and CIPK gene families and their expressions in response to drought and arbuscular mycorrhizal fungi colonization. Biol. Plant. 2020, 64, 773–783. [Google Scholar] [CrossRef]
- Ma, X.; Li, Q.-H.; Yu, Y.-N.; Qiao, Y.-M.; Haq, S. ul; Gong, Z.-H. The CBL–CIPK pathway in plant response to stress signals. Int. J. Mol. Sci. 2020, 21, 5668. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, C.; Tang, R.-J.; Xu, H.-X.; Lan, W.-Z.; Zhao, F.; Luan, S. Calcineurin B-Like proteins CBL4 and CBL10 mediate two independent salt tolerance pathways in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 2421. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogeny tree of CBL family proteins in A. littoralis (AlCBL), O. sativa (started with LOC Os), and A. thalianas (started with AT).
Figure 1.
Phylogeny tree of CBL family proteins in A. littoralis (AlCBL), O. sativa (started with LOC Os), and A. thalianas (started with AT).
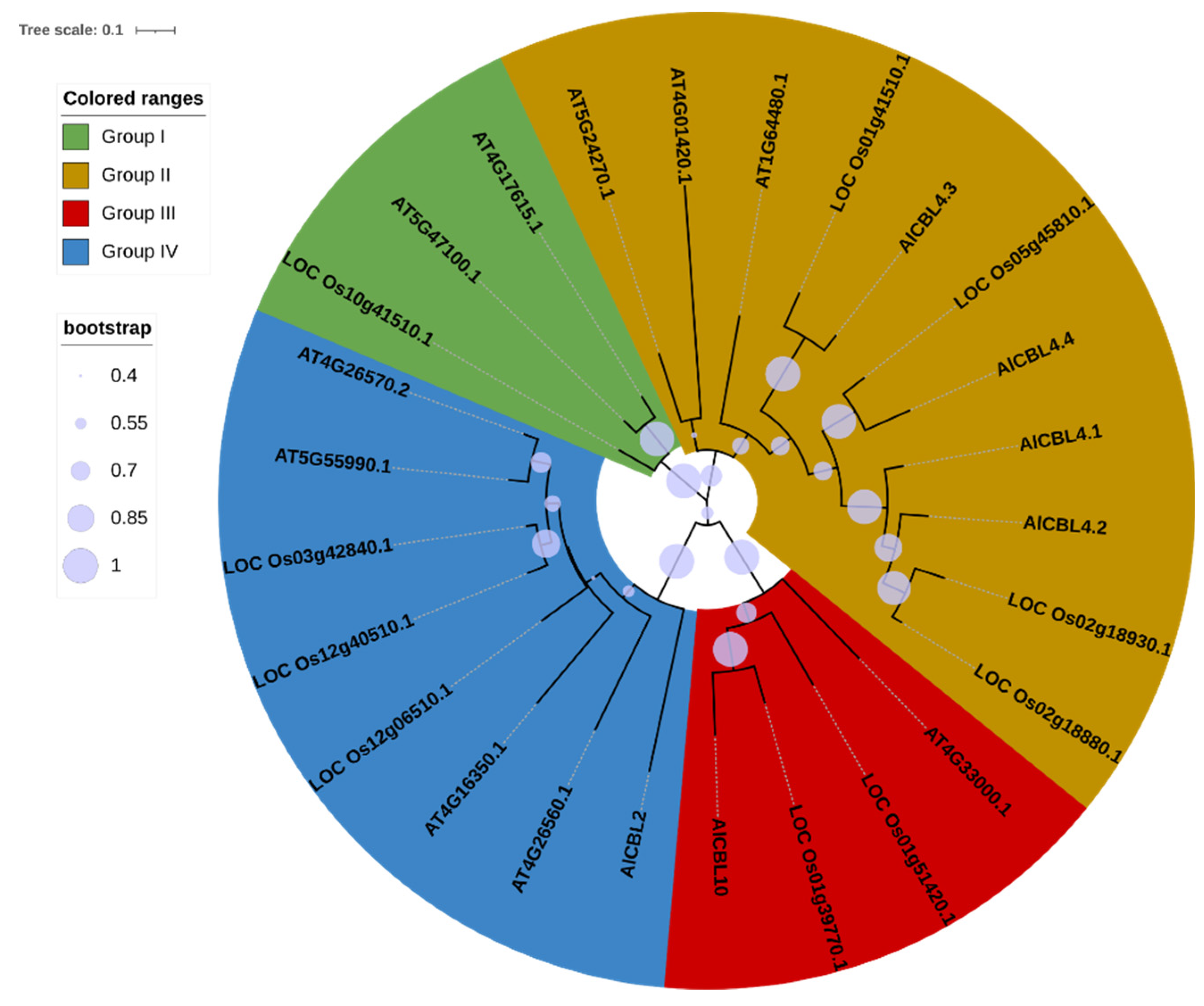
Figure 2.
Phylogeny tree of CIPK family proteins in A. littoralis (AlCBL), rice (started with LOC), and Arabidopsis (started with AT).
Figure 2.
Phylogeny tree of CIPK family proteins in A. littoralis (AlCBL), rice (started with LOC), and Arabidopsis (started with AT).
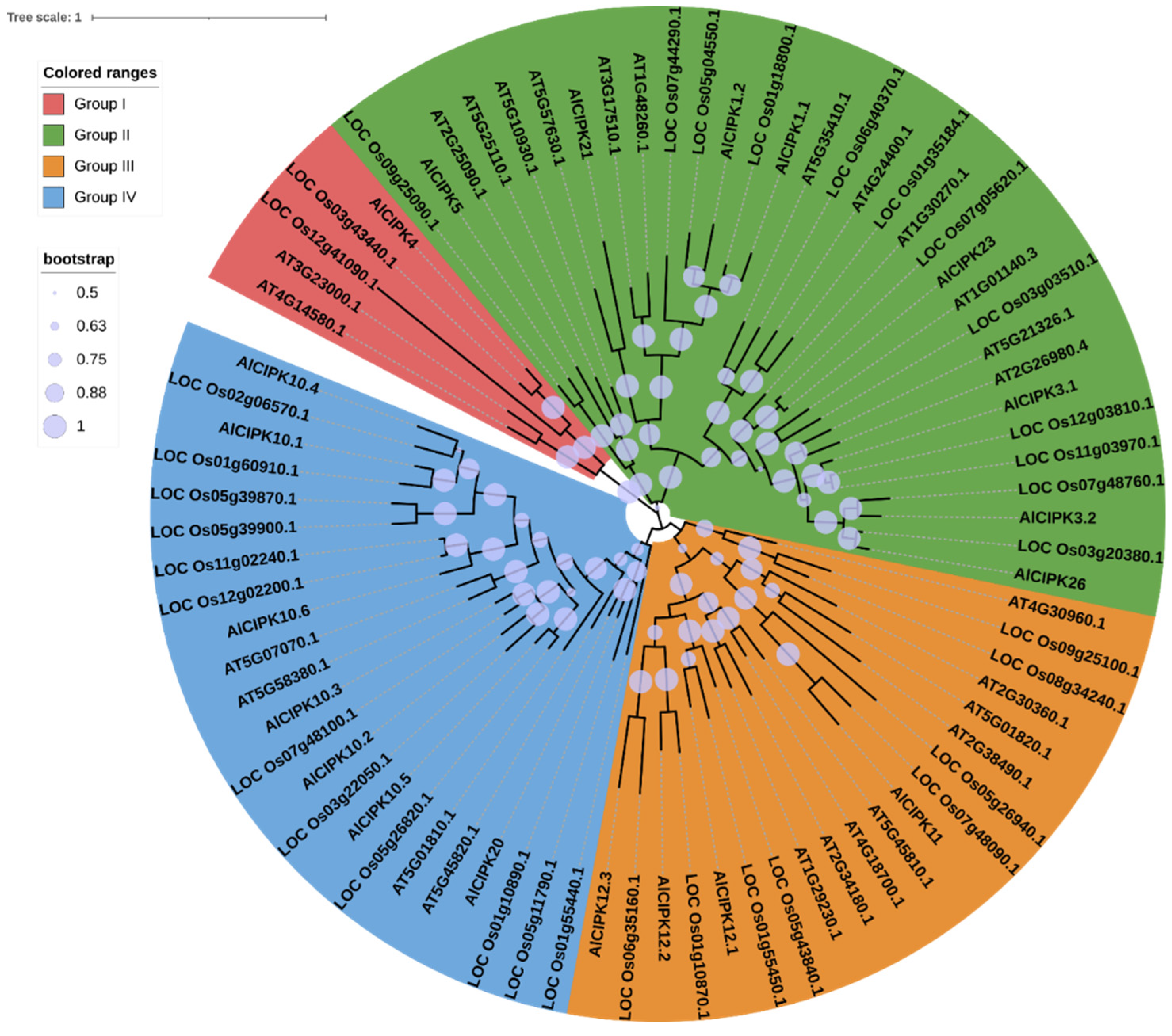
Figure 3.
Distribution of conserved motifs (a), functional domains (b), and exon/intron (c) in AlCBL and OsCBL family members.
Figure 3.
Distribution of conserved motifs (a), functional domains (b), and exon/intron (c) in AlCBL and OsCBL family members.

Figure 4.
Distribution of conserved motifs (a), functional domains (b), and exon/intron (c) in AlCIPK and OsCIPK family members.
Figure 4.
Distribution of conserved motifs (a), functional domains (b), and exon/intron (c) in AlCIPK and OsCIPK family members.
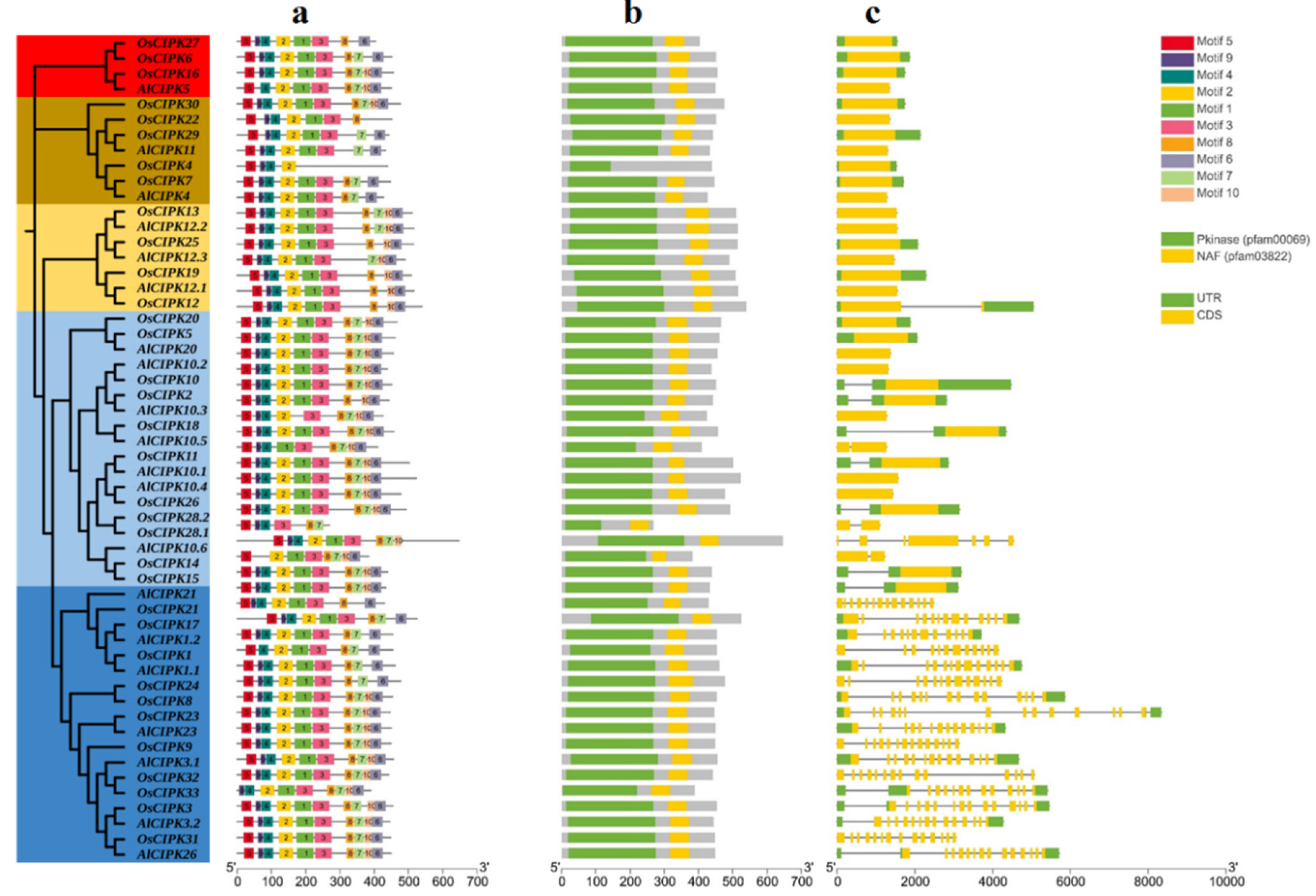
Figure 5.
Distribution of cis-regulatory elements in upstream of AlCBLs and ALCIPKs. Grouping of cis-regulatory elements based on their functions (a). Distribution of cis-regulatory elements involved in response to phytohormones (b), and stress (c), TF binding site (d).
Figure 5.
Distribution of cis-regulatory elements in upstream of AlCBLs and ALCIPKs. Grouping of cis-regulatory elements based on their functions (a). Distribution of cis-regulatory elements involved in response to phytohormones (b), and stress (c), TF binding site (d).
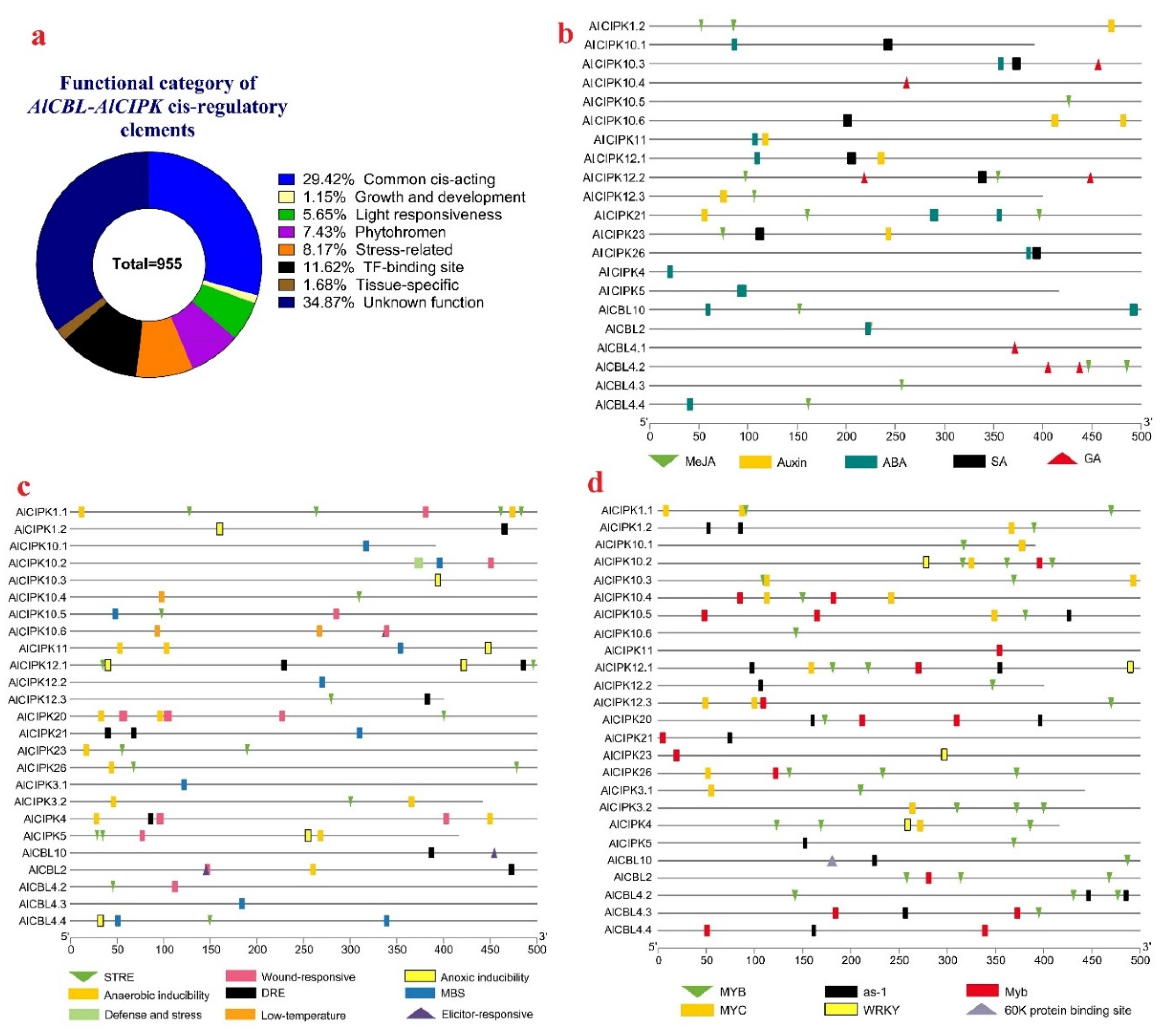
Figure 6.
Expression patterns of AlCBL genes in response to salinity in two tissues root and leaf. Expression levels are presented based on log2 fold change stress/normal condition.
Figure 6.
Expression patterns of AlCBL genes in response to salinity in two tissues root and leaf. Expression levels are presented based on log2 fold change stress/normal condition.
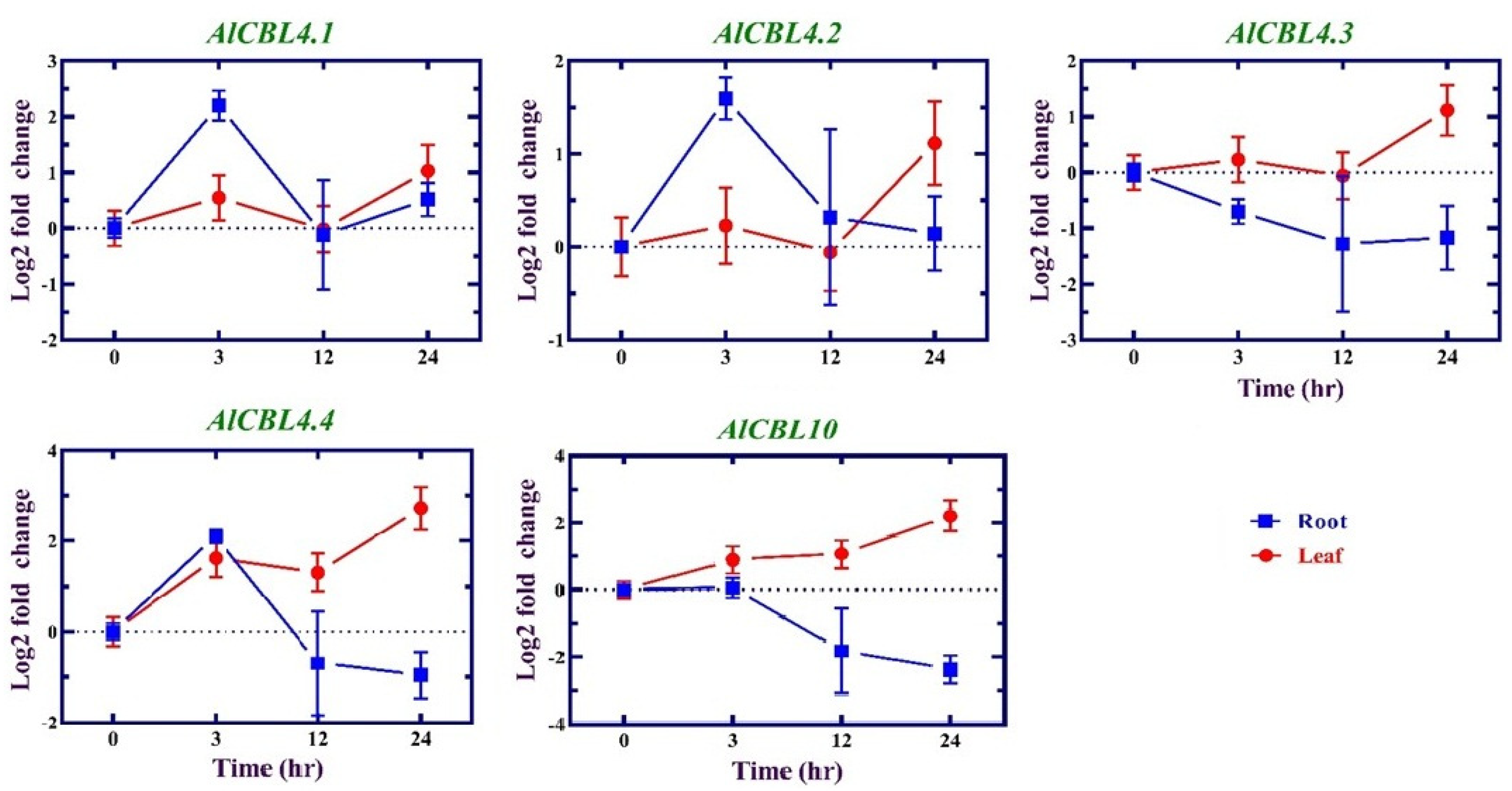
Figure 7.
Expression patterns of AlCIPK genes in response to salinity in two tissues root and leaf. Expression levels are presented based on log2 fold change stress/normal condition.
Figure 7.
Expression patterns of AlCIPK genes in response to salinity in two tissues root and leaf. Expression levels are presented based on log2 fold change stress/normal condition.
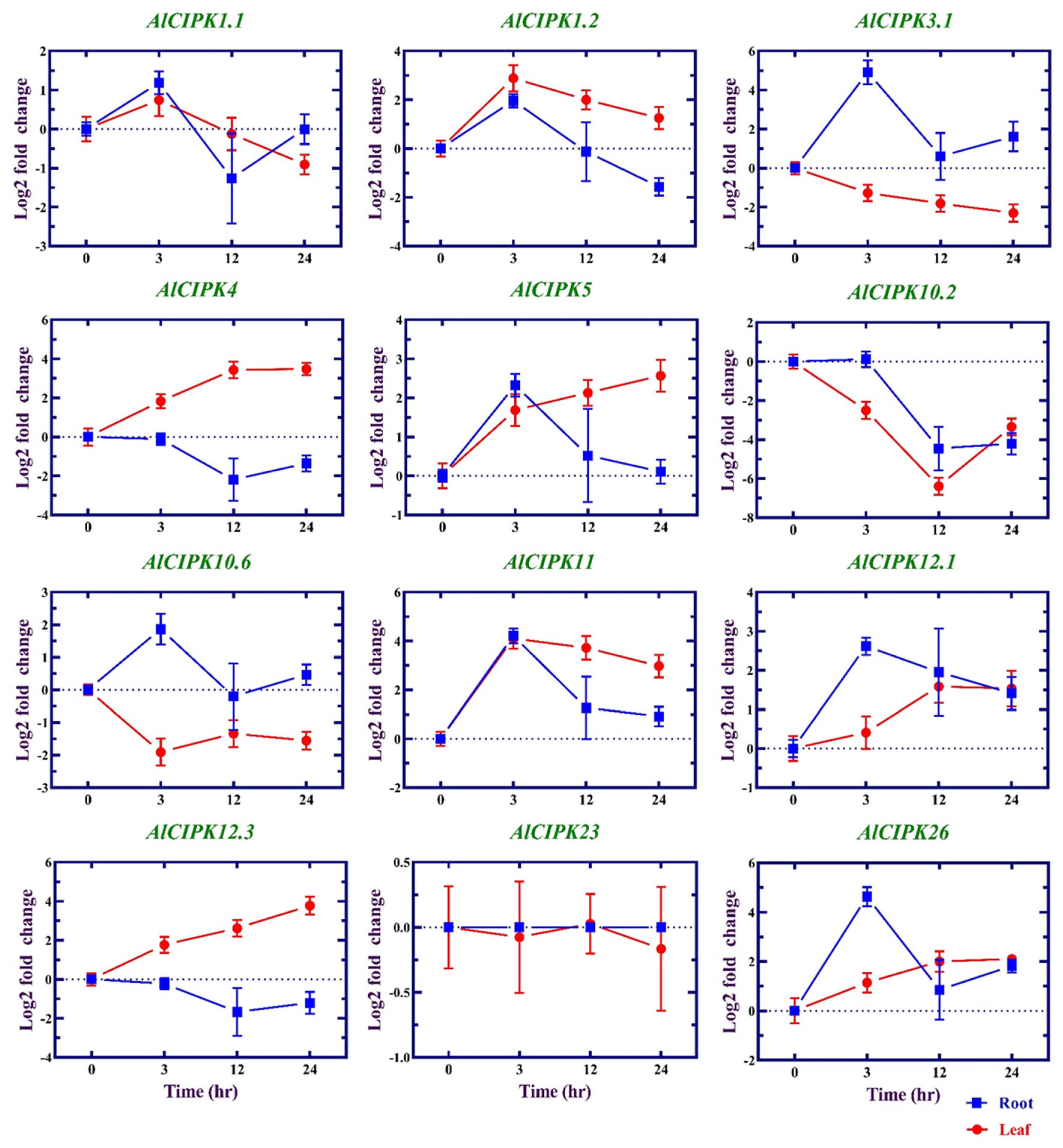

Table 1.
Physicochemical properties of identified AlCBLs and AlCIPKs encoded proteins from A. littoralis.
Table 1.
Physicochemical properties of identified AlCBLs and AlCIPKs encoded proteins from A. littoralis.
| Family | Gene ID | Gene Name | Length (aa) | Intron number | MW (kDa) | pI | GRAVY |
|---|---|---|---|---|---|---|---|
| CBL | Alg14121 | AlCBL2 | 226 | 7 | 25.87 | 4.98 | -0.219 |
| Alg15558 | AlCBL4.1 | 214 | 7 | 24.35 | 4.71 | -0.196 | |
| Alg11525 | AlCBL4.2 | 213 | 7 | 24.33 | 4.94 | -0.259 | |
| Alg8494 | AlCBL4.3 | 217 | 7 | 24.88 | 5.19 | -0.299 | |
| Alg13204 | AlCBL4. 4 | 166 | 5 | 18.70 | 4.78 | -0.341 | |
| Alg5886 | AlCBL10 | 303 | 8 | 34.67 | 5.28 | 0.133 | |
| CIPK | Alg4127 | AlCIPK1.1 | 473 | 12 | 53.48 | 6.52 | 0.372 |
| Alg7902 | AlCIPK1.2 | 454 | 11 | 50.69 | 6.62 | -0.320 | |
| Alg7566 | AlCIPK3.1 | 442 | 13 | 50.76 | 7.64 | -0.460 | |
| Alg12052 | ALCIPK3.2 | 448 | 13 | 50.63 | 8.23 | -0.407 | |
| Alg15044 | AlCIPK4 | 427 | 0 | 46.34 | 8.59 | -0.115 | |
| Alg5583 | AlCIPK5 | 450 | 0 | 48.19 | - | 0.054 | |
| Alg12300 | AlCIPK10.1 | 523 | 0 | 58.97 | 9.03 | 0.401 | |
| Alg9524 | ALCIPK10.2 | 438 | 0 | 49.72 | 9.28 | -0.260 | |
| Alg4701 | AlCIPK10.3 | 421 | 0 | 47.98 | 9.03 | 0.400 | |
| Alg3308 | AlCIPK10.4 | 478 | 0 | 54.66 | 9.13 | 0.514 | |
| Alg13906 | AlCIPK10.5 | 410 | 1 | 45.99 | 8.93 | -0.307 | |
| ALg9805 | AlCIPK10.6 | 383 | 1 | 42.04 | 8.99 | 0.480 | |
| Alg2698 | AlCIPK11 | 433 | 0 | 47.40 | 8.95 | -0.151 | |
| Alg8115 | AlCIPK12.1 | 516 | 0 | 57.36 | 8.64 | -0.341 | |
| Alg10559 | AlCIPK12.2 | 515 | 0 | 57.47 | 8.06 | -0.374 | |
| Alg11449 | AlCIPK12.3 | 490 | 0 | 54.06 | 8.84 | -0.254 | |
| Alg11347 | AlCIPK20 | 456 | 0 | 51.64 | 9.08 | -0.422 | |
| Alg8711 | AlCIPK21 | 430 | 13 | 48.54 | 6.21 | -0.303 | |
| Alg1003 | AlCIPK23 | 449 | 13 | 50.51 | 9.16 | -0.371 | |
| Alg7179 | AllCIPK26 | 448 | 13 | 50.44 | 8.41 | -0.395 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



