
Submitted:
27 February 2023
Posted:
28 February 2023
You are already at the latest version
Abstract
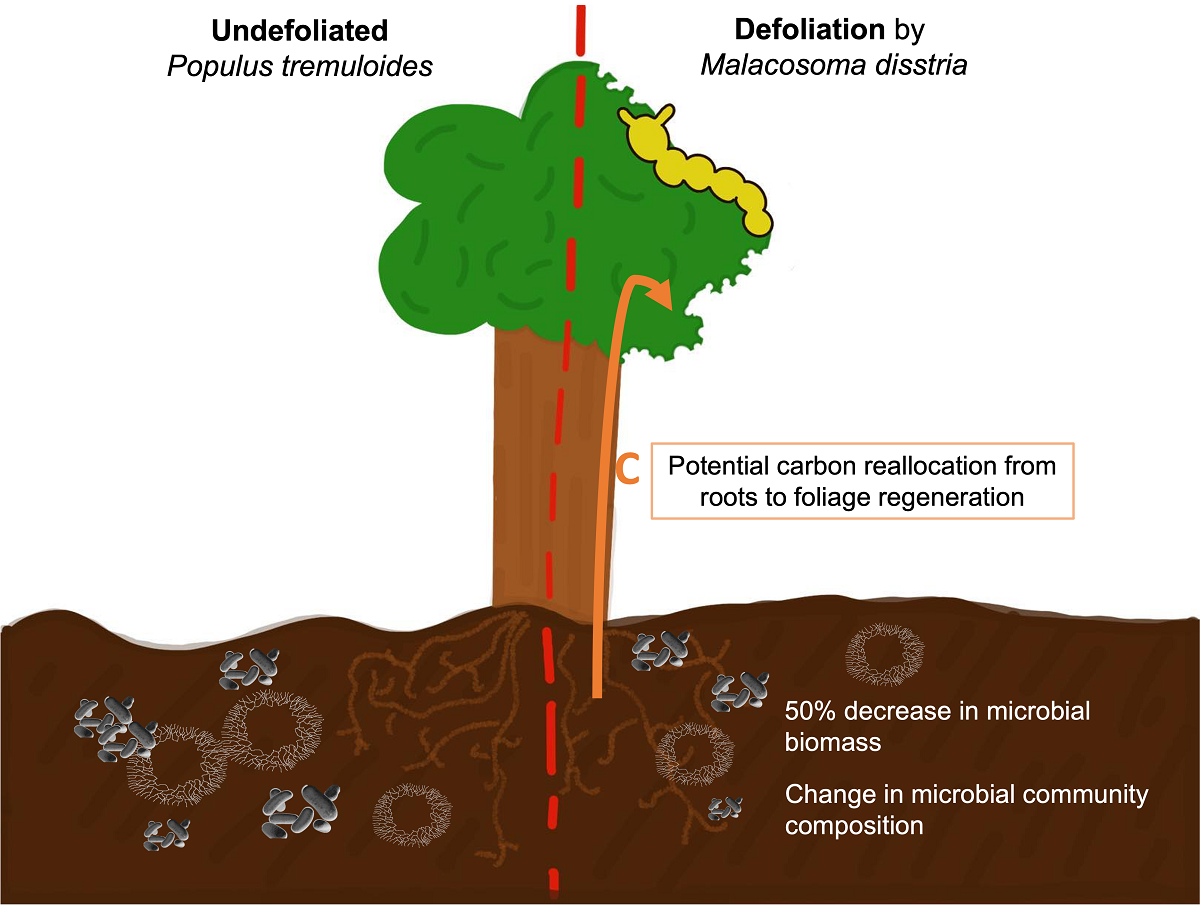
With climate change projected to increase the frequency and severity of episodic insect outbreak events, assessing potential consequences for soil microbial communities and nutrient dynamics is of importance for understanding forest resilience. The forest tent caterpillar (Malacosoma disstria) is an important defoliator of deciduous tree species in temperate and mixed forests of eastern North America with an invasion cycle every 10-12 years and outbreak events that can last 3-6 years. Following a defoliation episode on trembling aspen (Populus tremuloides) from 2015 to 2017 in Abitibi-Témiscamingue, QC, Canada, we sought to test if defoliation resulted in changes to soil bacterial and fungal communities. We hypothesized an increase in soil microbial biomass due to increased caterpillar frass inputs and potential changes in community structure following the event. Soils were sampled in July 2018, May 2019 and July 2019 from sites that had been subject to defoliation during the outbreak and from sites where no defoliation had been recorded. We assessed soil microbial biomass and fungal to total microbial activity ratio on all sampling dates, and Community Level Physiological Profiles (CLPPs) for 2018 only using a substrate induced respiration method. Contrary to our hypothesis, we observed a significant 50% decrease in microbial biomass (g biomass-C g-1 soil hour-1) in defoliated stands suggesting tree carbon normally allocated towards root exudates was reallocated towards foliage regeneration. We noted a differentiated carbon-based substrate usage following defoliation, but no change in the fungal to total microbial activity ratio. The observed changes in the two years following the defoliation event suggest that defoliation episodes aboveground could trigger changes in soil chemistry belowground with effects on soil microbial communities that may, in turn, feedback to influence forest plant dynamics.
Keywords:
Forest tent caterpillar (Malacosoma disstria)
; insect outbreak
; defoliation event
; MicroResp
; soil microbial activity
1. Introduction
Insect outbreaks are natural disturbances inherent to forest ecosystems and can alone, or coupled with droughts or fires, lead to tree mortality [1,2,3]. However, there is increasing concern that climate change will affect outbreak dynamics as higher global temperatures offer better conditions for the reproductive cycle of insects, causing an increase in frequency and intensity of invasive defoliator episodes [4,5,6]. Higher defoliating insect density may attenuate the capacity of high latitude forests to act as carbon sinks as tree growth and carbon sequestration depends in part on photosynthesis carried out by foliage [7]. Furthermore, interacting factors like drought may exacerbate tree mortality turning forests into carbon sources [8].
Consequences of altered outbreak dynamics may extend beyond host plants to soil ecosystems as vascular plants interact actively with above-ground and below-ground components of the forest ecosystem [9]. For example, insect herbivory may have both indirect and direct effects on nutrient dynamics and decomposers in soils by altering the resources flowing from above to below-ground components [9,10]. Hunter (2001) identified seven broad mechanisms by which herbivory could affect nutrient cycles. Mechanisms mediated directly by insect herbivores include increased defecation (frass) or insect cadaver inputs or changes in the nutrient content of precipitation throughfall passing through the canopy [10,11]. Indirect mechanisms mediated by host trees may include changes in leaf litter quantity and quality such as defence compounds or nutrient concentration in foliage [12], changes in the canopy structure that may modify microclimate in the understory and decreased allocation to root exudates that may alter symbiotic interactions [13]. In the medium term, changes in plant community composition may also alter nutrient cycles [10,14].
Thus, insect defoliation can reduce the amount of nutrient-rich leaves arriving to the soil while nitrogen-rich frass changes the nutritional input to the underground system, affecting carbon (C) and nitrogen (N) fluxes (eg. [15,16]). Digestion by insect herbivores reduces the C:N ratio in the frass compared to the ratio from the plant foliage [17]. This new N input is not necessarily absorbed by plant roots directly: Grüning et al. (2017) found that the new N input in the soil following a defoliation event by the nun moth (Lymantria monacha L.) on Scots pine (Pinus sylvetris L.) reduced N uptake capacity of the pine roots as the tree reallocated N from internal sources rather than increasing root N acquisition to compensate the loss of foliage. This N input could therefore become available to other organisms below-ground such as soil microbes.
Soil microbial activity, estimated typically with substrate-induced soil respiration, is an indicator of below-ground change, but studies assessing the effect of herbivorous insect outbreaks on soil microbial activity have shown mitigated results. Under artificial defoliation conditions, simulated frass addition had no effect on soil microbial activity [19]. Other laboratory studies found that litterfall and greenfall exclusion resulted in a decrease of soil microbial respiration [20], while mechanical or insect-induced defoliation increased it [15]. However, soil microbial activity following natural defoliation events generally showed a more consistent response. Multiples studies demonstrated an increase in soil microbial activity one to 16 months following an insect outbreak [11,17,21], although some found no change after 6 months [22,23] and short-term increased activity may also taper or inverse itself after three years [17].
Soil microbial community composition is increasingly studied in forest ecosystems [24], but knowledge in response to episodic insect outbreaks remains limited. Soil microbial community composition can be changed in response to recurring herbivory, as a result of changes in soil abiotic conditions such as high soil temperatures exacerbated by canopy opening during an outbreak and decreases in soil C flow from tree root exudation [25]. In response to different defoliator outbreaks, shifts of specific microbial groups have been observed. For example, Castaño et al. (2020) found a decrease in soil fungal biomass associated with a decrease in root exudates after an outbreak by the pine processionary moth. In lab cultures, Oneț et al. (2016) found an increase in fungi and a decrease in heterotrophic bacteria from soils sampled under oaks defoliated by spongy moth and attributed them to higher pH and changes in soil chemistry.
The forest tent caterpillar (Malacosoma disstria Hübner) is a major native defoliator of North American hardwood trees [28]. In the boreal zone, trembling aspen is its preferred host tree and the one on which the most important outbreaks have been recorded [29]. Forest tent caterpillar outbreaks occur in roughly 10-20 year cycles while many landscape variables like forest structure, climate and topography might affect the outbreak duration, lasting on average 4-6 years [30,31,32]. During an outbreak, the caterpillar modifies its environment by feeding on the host’s foliage efficiently, typically on the deciduous canopy [33] which can negatively affect tree growth and increase mortality [34]. We sought to better understand the medium-term effects of a recent (2015-2017) outbreak of the forest tent caterpillar in eastern Canada on soil microbial communities. Sampling beneath defoliated and undefoliated aspens, we assessed soil microbial biomass and community-level physiological profiling over two successive summers following a 3-year defoliation episode (2015-2017). Given that soil chemistry analysis at peak defoliation in 2017 showed higher amounts of N in soil under defoliated trees from our sites attributed to frass addition (Figure S1), we hypothesized an increase in soil microbial abundance and altered community composition due to additional nutritional input from the outbreak.
2. Materials and Methods
This study was conducted in the Abitibi region of western Quebec, Canada. The study area is located within the balsam fir-white birch bioclimatic domain (Saucier et al. 2011), at and around the Lake Duparquet Research and Teaching Forest (LSRTF; 48°30′ N, 79°20′ W). Forests of the LDRTF are characterized by pure and mixed stands composed of boreal coniferous and shade-intolerant deciduous species. On mesic sites, trembling aspen (Populus tremuloides Michaux), white birch (Betula papyrifera Marshall) and jack pine (Pinus banksiana Lamb.) dominate early successional stands, whereas balsam fir (Abies balsamea L.) and eastern white cedar (Thuja occidentalis L.), in association with white spruce (Picea glauca Moench) and persistent, scattered white birch, dominate late-successional stands [35]. Glaciolacustrine clays cover 55% of the LDRTF territory [36] and are the legacy of proglacial lakes Ojibway and Barlow [37]. The climate is continental and, according to the nearest weather station (Mont-Brun), for the period 1980–2010, the growing season lasted about 150 to 160 days, whereas the mean annual temperature and mean annual precipitation was 1.0 °C and 985 mm, respectively [38].
Both the defoliated and undefoliated stands were dominated by 70–90-year-old aspens (Populus tremuloides), located on moderately well-drained glacial clays developed in Luvisols and separated by approximately 10 km. In 2016 and 2017, the defoliated stand was characterized by the provincial Ministry of Forests, Wildlife and Parks through aerial surveys as severe defoliation (loss of foliage all along the crown of the majority of trees) and estimated at 70 to 90% canopy loss [39,40]. Provincial surveys record no defoliation in the region in 2018 [41]. In 2017 and 2018, caterpillar colonies were counted in both stands by the research team but were only observed in the defoliated stands in 2017.
Within each stand, eight sampling sites were selected at the base of a defoliated or undefoliated Populus tremuloides tree. Sampled trees within the stand were separated by at least 115 m from each other. In August 2018, May 2019 and July 2019, two soil cores were extracted at 0-5 cm and 5-10 cm depth within 1 m from each selected tree and subsequently pooled. Specifically, we sampled 745 ml of fresh soil that was sieved on a 2 mm test sieve (Retsch), put in sterile tubes, transported in a cooler with multiple ice packs and then frozen at -20 °C until analysis.
Using the substrate-induced soil respiration method of MicroRespTM [42] and FungiResp [43], both total microbial biomass (μg CO2-C g−1 hour−1) and the fungal ratio were tested as indicators of soil microbial activity. Soils were defrosted for twenty-four hours before being adjusted with distilled water to a water-holding capacity of 35% for all samples. After a week of incubation, bronopol then glucose was added to the samples and after six hours, the colorimetric microplate was read using a microplate spectrophotometer (Multiskan Go, Thermoscientific). A factor of 40 of glucose-induced respiration (GIR) was used as a proxy of microbial biomass [43,44]. We estimated the fungal ratio of our soils by adding a solution of bronopol at 78 μg g−1 of soil as a bacterial inhibitor (Sassi et al.,2012). A ratio of the microbial activity (μg CO2-C g−1 hour−1) induced with the inhibitor divided by the microbial activity induced with glucose allowed us to estimate the proportion of fungi compared to bacteria in our samples. Calibration curves associating percentages of CO2 to different absorbances of the detection plate were determined with the two different indicator solutions using a portable CO2 analyzer (EGM-5, PP Systems). This allowed us to convert the normalized 6-hour data to %CO2. While 8 soil samples from the 0-5 cm depth were taken from each stand type at each of the three sampling dates, 9 of the 48 samples were compromised in the lab due to the over-adjustment of water saturation capacity in very wet samples. Ultimately, this resulted in variable replication (n = 5 to 8 samples) depending on treatment and sampling date (see caption in Figure 1 for explicit details). The 5-10 cm depth was only analyzed for soils collected in August 2018.
The technique of assessing community-level physiological profiles (CLPPs) measures the ability of the soil microbiome to metabolize different carbon substrates. To determine the functional composition of the microbial communities by CLPPs, we created a catabolic fingerprint by testing four dissolved carbon sources ecologically relevant to induce differentiated mineralization between soil microbial communities [45]. Two carbohydrates (fructose and glucose), one amino acid (glycine) and one carboxylic acid (malic acid) providing different chemical types of acids and sugars were selected following Campbell et al. (2003) and Sassi et al. (2012). In the same manner as the MicroResp TM protocol, soils were defrosted for twenty-four hours before being adjusted with distilled water to a water-holding capacity of 35% for all samples. After a week of incubation, we added the four different substrate solutions (30 mg g−1). Aliquots of 25 μL per solution were inserted in each corresponding well of the MicroRespTM 96-deep-well plate according to Sassi et al. (2012). After six hours, the colorimetric microplate was read using a microplate spectrophotometer (Multiskan Go, Thermoscientific). Only 2018 samples were analyzed. To compare substrate breakdown from each treatment, relative utilization was assessed by the proportion of the total respiration induced by each substrate. To calculate the relative utilization by the microbial community for each defoliation treatment, the mean respiration per substrate per treatment was divided by the mean of total respiration across substrates per treatment.
Soil microbial biomass and the relative utilization of the four carbon-based substrates were analyzed with a mixed model analysis where defoliation and time of sampling were fixed effects and the individual tree beneath which the soil was sampled (site) was a random effect. The mixed model analysis was done with lme4 [46] and lmerTest [47] libraries. Analyses were done in the Rstudio (v.1.2.1335) (Rstudio Inc., Boston, USA) environment with R software (v.3.6.0) (R Development Core Team, Vienna, Austria).
3. Results
3.1. Soil microbial biomass
Soil microbial biomass ranged from 173 to 911 μg biomass-C g−1 soil hour−1 (Figure 1, P < 0.001), with lower biomass in the spring and higher biomass in the late summer, but no significant interaction between defoliation and sampling date was observed (Figure 1, P = 0.36). When assessing glucose-induced activity, microbial biomass in the top 5 cm of soil from previously defoliated stands was consistently about half that in control stands, across all sampling dates (Figure 1, P < 0.001). This microbial biomass decrease was also observed for all other substrates (glycin, fructose and malic acid) included to assess the catabolic fingerprint (August 2018 only) with each substrate-induced activity being two to three times the magnitude in control compared to defoliated stands (Figure 2, P < 0.001). Furthermore, there was no significant interaction between defoliation and the type of substrate used (Figure 2, P = 0.50).
In contrast, average soil microbial biomass in deeper soils (5-10 cm depth, analyzed for 2018 only) showed no effect of defoliation (P = 0.31). However, average soil microbial biomass in deeper soils was ca. 50% (65% for non-defoliated and 37% for defoliated sites) lower than that measured in the 0-5 cm layer (data not shown).
3.2. Soil microbial community
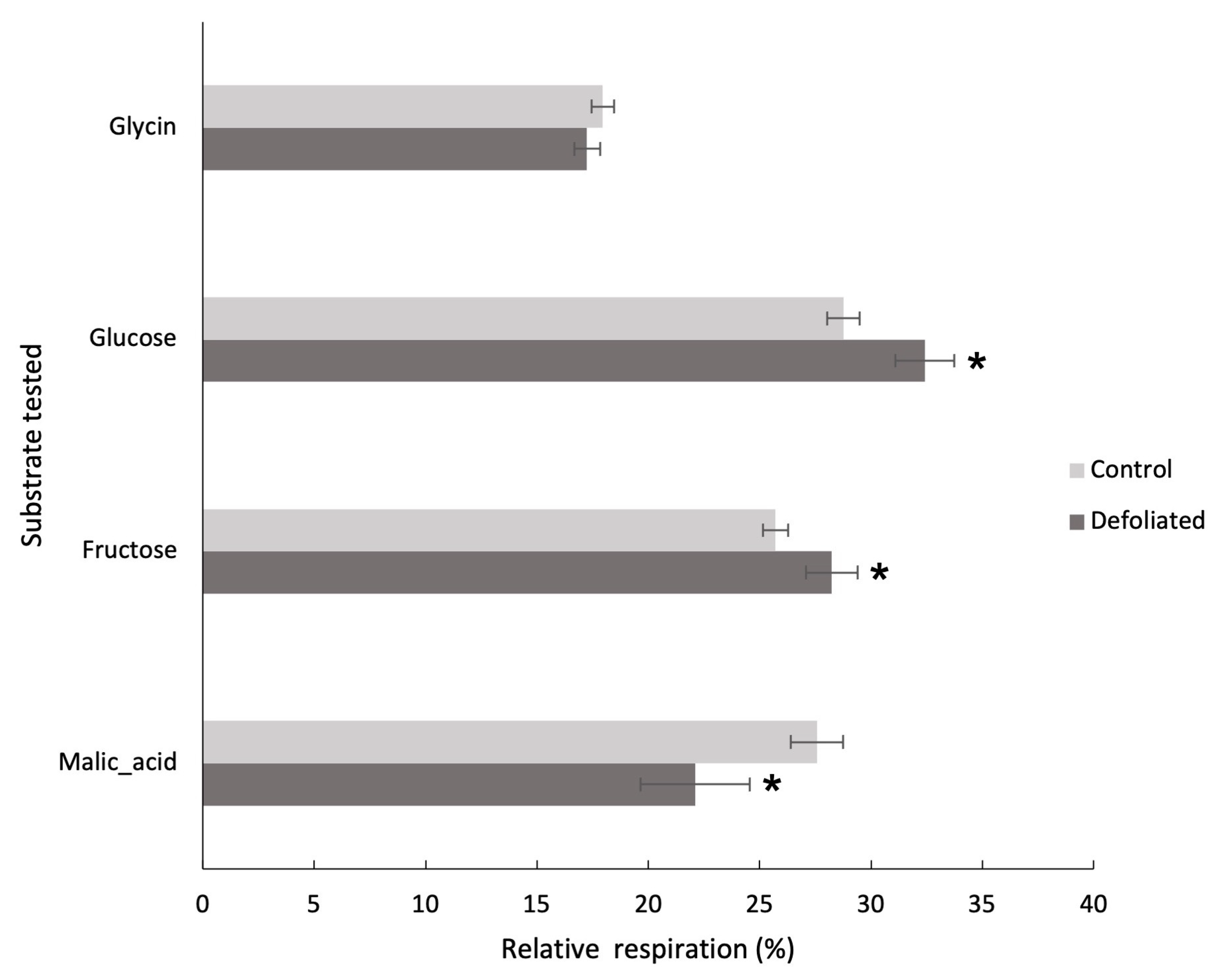
Community-level physiological profiles (CLPP) offer an indicator of microbial community functional composition. Figure 3 reshapes the data shown in Figure 2, offering a view of the proportion of respiration from each substrate relative to the total respiration induced across all substrates. Relative respiration of our microbial communities showed a differentiated usage of four carbon-based substrates between the control and the defoliated stands from soil sampled in 2018 (Figure 3). The defoliated stand microbial community used significantly less malic acid than that from soil under control trees but used more carbohydrates-based substrates (Figure 3). Relative respiration of carbohydrate substrates (glucose and fructose) was on average 12 9% higher for the defoliated treatments compared to control treatments (P < 0.05). Inversely, respiration from the carboxylic acid substrate (malic acid) was 20% lower for the defoliated treatment than the control (P < 0.05).
3.3. Fungal ratio
The average fungal to total microbial activity ratio across defoliated and control plots was 0.86 0.17, revealing a soil dominated by fungi throughout the samples. There were no significant differences between defoliated and control plots (P = 0.37).
4. Discussion
Our two-year study following a severe defoliation outbreak of the forest tent caterpillar provided us with a medium-term understanding of microbial dynamics in the trembling aspen-dominated stands of the mixed wood boreal forest. Most studies to date have been limited to short-term responses [15,19,20,22,23,33,48] or drawn general conclusions using artificial defoliation events [15,19,20,49]. While these latter artificial defoliation experiments have helped understand valuable mechanisms, such study systems are limited in their ability to reproduce the complex interactions of natural outbreaks. Our results showed that, contrary to our hypothesis that N input by frass would stimulate microbial activity [15,17,18,21], microbial biomass decreased by half in defoliated sites in the top 5 cm of soil and the response was consistent over all sampling dates (Figure 1) and tested substrates (Figure 2). As hypothesized, we observed a shift in microbial functional composition in defoliated sites based on assessments with selected substrates (Figure 3), but no change in the overall fungal activity was observed. Potential interpretations for our results include changes in the quality of aspen foliage [50], a decrease in C allocation to roots [13] and a change in vegetation regeneration benefiting balsam fir over trembling aspen [34].
Our observed decreased microbial response may be explained by changes in tree foliage quality following the major defoliation event, resulting in afterlife effects of tree litter [5,6]. A major forest tent caterpillar outbreak can lead to altered foliage quality in the second flush of leaves of host tree species with lower nitrogen and increased defence compounds. Trembling aspens produce foliar phenolic glycosides as a chemical defence in response to herbivory which is an effective deterrent to defoliators [12]. This effect can persist in time with foliage grown two years after a defoliation event having on average six times more phenolic glycosides than before defoliation (Donaldson and Lindroth 2008). Condensed tannins can also increase in remnant defoliated leaves (Donaldson and Lindroth 2008). When leaf litter falls, condensed tannins derived from plant material can form complexes with proteins [51] binding N [52,53] and chitin from fungi [54,55]. Such complexes, particularly with high molecular mass tannins, can inhibit microbial activity in soil by binding extracellular substrates [54,56,57]. Several short-term (<1 year) studies on microbial activity found an increase [11,15,17,21] or no change following insect herbivory [23,48]. However, two years after a natural defoliation event which is more similar to conditions in this study, Streminska et al. (2006) found a decrease in microbial biomass explained by lower amounts of litterfall. In her meta-analysis on N addition and microbial biomass in soil across multiple ecosystem types and biomes, [58] also found an overall decrease in microbial biomass with increasing amounts of N, particularly in studies of longer duration, indicating soil microbes are not necessarily N limited.
An alternate or complementary interpretation of our reduced microbial biomass following the outbreak event is that insect herbivory can induce an indirect response from the host tree by altering C allocation to tree roots and thus affecting microbial activity in soils. Repeated herbivory, by reducing autotrophic C supply, can lead to reduced root activity [59] and fine root biomass [60] implying that trees allocate less C below-ground than before the defoliation event. In their meta-analysis on below-ground responses to insect herbivory in ecosystems with woody plants, [13] showed that defoliators decrease carbon allocation from the host plant to its roots, provoking a decrease in root biomass and exudation. Following a disturbance, trembling aspen commonly regenerates via root suckering, but can be limited by competition from other trees [61]. We would therefore expect aspens not only to allocate C to their foliage regeneration instead of their roots but also to reduce their investment to suckering because of the competition from balsam fir that can benefit from the outbreak [34].
The forest tent caterpillar outbreak, by favouring regeneration of certain tree species such as balsam fir, could help explain the differences in CLPPs between our defoliated and undefoliated sites with soil microbial communities composition altered following a change in the availability of their preferred carbon source and soil chemistry. The caterpillar outbreak can benefit competing tree species in the forest that may ultimately alter vegetation dynamics favouring coniferous over deciduous trees as the dominant litter type changes on the forest floor [34]. The CLPPs profiles differed from soil located beneath defoliated and undefoliated control trees, indicating that a change in litter identity from aspens to firs could stimulate different microbial communities able to degrade coniferous or deciduous litter types. Light availability allowed by canopy openings following a defoliation episode can contribute to changes in the plant community composition, facilitating the growth of shade-intolerant species [10]. At the same mixed wood site in Abitibi-Témiscamingue QC, Moulinier et al. (2013) showed previous outbreaks benefited the regeneration of a conifer Abies balsamea (Balsam fir) to the detriment of the deciduous trembling aspen.
Furthermore, the CLPPs of soils located beneath defoliated trees indicated that those microbial communities used less carboxylic acid (malic acid) and more monosaccharides (fructose and glucose) than under undefoliated trees (Figure 3). Studies comparing soils from deciduous and coniferous forests also found microbes from the latter were less efficient at using carboxylic acids [63,64]. However, Chodak et al. (2016), found less efficient use of monosaccharide substrates by microbial communities from the coniferous forest. Along with a change in vegetation regeneration in the defoliated forest, a decrease in root biomass and root exudates of labile C in the topsoil can also shift the microbial communities associated with roots, as illustrated by a negative correlation between herbivory and ectomycchorizal fungi abundance [13,60]. Other studies have suggested higher soil acidity [27] or lower C: N ratio in the litterfall [17] could increase fungal biomass in topsoil under defoliated trees compared to undefoliated trees, but our fungal ratio indicator remained constant between undefoliated and defoliated sites. This possible change in forest regeneration at our site favouring balsam fir over trembling aspen may also have consequences for soil micronutrients. In 2017 at the peak of the outbreak, higher N, P, K, Ca and Mg concentrations were found in soils under defoliated compared to non-defoliated trees (Figure S1, p<0.005). For example, Ca concentrations were double at defoliated sites than at undefoliated sites which could reflect a change in nutrient absorption by dominant trees. Trembling aspen have very high Ca requirements [65] and thus gradual replacement by other species would explain the high concentrations of Ca on soil under defoliated trees.
5. Conclusions
Although the recent meta-analysis by Kristensen et al. (2020) did not report significant effects of outbreak herbivory on forest microbes, their analysis mixes outbreaks on both coniferous and deciduous tree species with deciduous stands being largely underrepresented in the 60 articles analyzed. Our study therefore suggests that microbes living in deciduous stands of mixedwood boreal forests may respond differently from those in coniferous forests, which could eventually feedback on vegetation dynamics in a warming climate where outbreaks are projected to become increasingly common. This study underlines the importance of considering soil microbes as part of a forest ecosystem perturbation’s impacts such as an insect defoliation event.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Soil chemistry analysis;
Author Contributions
Conceptualization, EDM. and TH; methodology, EDM and TH; resources, TH and ED; data curation, EDM; writing—original draft preparation, EDM and TH; writing—review and editing, ED; supervision, TH; project administration, TH; funding acquisition, TH and ED.
Funding
The QCBS provided a seed grant to ITH and ED in 2018 and a Biodiversity Science Discovery Award to EDM in 2019. SERG international provided a grant to ED. The NSERC provided a Collaborative Research and Development Grant to ED.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to AS Caron, J Jarry, S Jarry and L Rousseau for field assistance. We thank M Bouchard, F Guay, S Légaré, JP Lessard and L Nowell for their collaboration in the NSERC CRD grant. We acknowledge NSERC for financial support through a scholarship for EDM.
Conflicts of Interest
The authors declare no conflict of interest
References
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree Mortality from Drought, Insects, and Their Interactions in a Changing Climate. New Phytologist 2015, 208, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Ramsfield, T.D.; Bentz, B.J.; Faccoli, M.; Jactel, H.; Brockerhoff, E.G. Forest Health in a Changing World: Effects of Globalization and Climate Change on Forest Insect and Pathogen Impacts. Forestry 2016, 89, 245–252. [Google Scholar] [CrossRef]
- Turner, M.G. Disturbance and Landscape Dynamics in a Changing World. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain Pine Beetle and Forest Carbon Feedback to Climate Change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Mitton, J.B.; Ferrenberg, S.M. Mountain Pine Beetle Develops an Unprecedented Summer Generation in Response to Climate Warming. The American Naturalist 2012, 179, E163–E171. [Google Scholar] [CrossRef] [PubMed]
- Uelmen, J.A.; Lindroth, R.L.; Tobin, P.C.; Reich, P.B.; Schwartzberg, E.G.; Raffa, K.F. Effects of Winter Temperatures, Spring Degree-Day Accumulation, and Insect Population Source on Phenological Synchrony between Forest Tent Caterpillar and Host Trees. Forest Ecology and Management 2016, 362, 241–250. [Google Scholar] [CrossRef]
- Silfver, T.; Heiskanen, L.; Aurela, M.; Myller, K.; Karhu, K.; Meyer, N.; Tuovinen, J.-P.; Oksanen, E.; Rousi, M.; Mikola, J. Insect Herbivory Dampens Subarctic Birch Forest C Sink Response to Warming. Nature Communications 2020, 11, 2529. [Google Scholar] [CrossRef]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest Insects and Climate Change. Curr Forestry Rep 2018, 4, 35–50. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A. HERBIVORE-MEDIATED LINKAGES BETWEEN ABOVEGROUND AND BELOWGROUND COMMUNITIES. Ecology 2003, 84, 2258–2268. [Google Scholar] [CrossRef]
- Hunter, M.D. Insect Population Dynamics Meets Ecosystem Ecology: Effects of Herbivory on Soil Nutrient Dynamics: Insect Population Dynamics Meets Ecosystem Ecology. Agricultural and Forest Entomology 2001, 3, 77–84. [Google Scholar] [CrossRef]
- Grüning, M.M.; Beule, L.; Meyer, S.; Karlovsky, P.; I. -M.-Arnold, A. The Abundance of Fungi, Bacteria and Denitrification Genes during Insect Outbreaks in Scots Pine Forests. Forests 2018, 9, 497. [CrossRef]
- Donaldson, J.R.; Lindroth, R.L. GENETICS, ENVIRONMENT, AND THEIR INTERACTION DETERMINE EFFICACY OF CHEMICAL DEFENSE IN TREMBLING ASPEN. Ecology 2007, 88, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, J.Å.; Rousk, J.; Metcalfe, D.B. Below-ground Responses to Insect Herbivory in Ecosystems with Woody Plant Canopies: A Meta-analysis. J Ecol 2020, 108, 917–930. [Google Scholar] [CrossRef]
- Niklaus, P.A.; Wardle, D.A.; Tate, K.R. Effects of Plant Species Diversity and Composition on Nitrogen Cycling and the Trace Gas Balance of Soils. Plant Soil 2006, 282, 83–98. [Google Scholar] [CrossRef]
- Frost, C.J.; Hunter, M.D. Insect Canopy Herbivory and Frass Deposition Affect Soil Nutrient Dynamics and Export in Oak Mesocosms. Ecology 2004, 85, 3335–3347. [Google Scholar] [CrossRef]
- le Mellec, A.; Gerold, G.; Michalzik, B. Insect Herbivory, Organic Matter Deposition and Effects on Belowground Organic Matter Fluxes in a Central European Oak Forest. Plant Soil 2011, 342, 393–403. [Google Scholar] [CrossRef]
- Streminska Microbial Abundance and Some of Their Physiological Activities in Soil Organic Horizon of Pine Forest Affected by Insect Herbivory. Pol. J. Environ. Stud. 2006, 15, 905–914.
- Grüning, M.M.; Simon, J.; Rennenberg, H.; l-M-Arnold, A. Defoliating Insect Mass Outbreak Affects Soil N Fluxes and Tree N Nutrition in Scots Pine Forests. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Lovett, G.M.; Ruesink, A.E. Carbon and Nitrogen Mineralization from Decomposing Gypsy Moth Frass. Oecologia 1995, 104, 133–138. [Google Scholar] [CrossRef]
- Reynolds, B.C.; Hunter, M.D. Responses of Soil Respiration, Soil Nutrients, and Litter Decomposition to Inputs from Canopy Herbivores. Soil Biology and Biochemistry 2001, 33, 1641–1652. [Google Scholar] [CrossRef]
- Baranchikov, Y.N.; Perevoznikova, V.D.; Vishnyakova, Z.V. Carbon Emission by Soils in Forests Damaged by the Siberian Moth. 2002, 33, 4.
- le Mellec, A.; Habermann, M.; Michalzik, B. Canopy Herbivory Altering C to N Ratios and Soil Input Patterns of Different Organic Matter Fractions in a Scots Pine Forest. Plant Soil 2009, 325, 255–262. [Google Scholar] [CrossRef]
- Morehouse, K.; Johns, T.; Kaye, J.; Kaye, M. Carbon and Nitrogen Cycling Immediately Following Bark Beetle Outbreaks in Southwestern Ponderosa Pine Forests. Forest Ecology and Management 2008, 255, 2698–2708. [Google Scholar] [CrossRef]
- Baldrian, P. Forest Microbiome: Diversity, Complexity and Dynamics. FEMS Microbiol Rev 2017, 41, 109–130. [Google Scholar] [CrossRef] [PubMed]
- Classen, A.; Demarco, J.; Hart, S.; Whitham, T.; Cobb, N.; Koch, G. Impacts of Herbivorous Insects on Decomposer Communities during the Early Stages of Primary Succession in a Semi-Arid Woodland. Soil Biology and Biochemistry 2006, 38, 972–982. [Google Scholar] [CrossRef]
- Castaño, C.; Camarero, J.J.; Zas, R.; Sampedro, L.; Bonet, J.A.; Alday, J.G.; Oliva, J. Insect Defoliation Is Linked to a Decrease in Soil Ectomycorrhizal Biomass and Shifts in Needle Endophytic Communities. Tree Physiology 2020, 40, 1712–1725. [Google Scholar] [CrossRef] [PubMed]
- Oneț, A.; Teușdea, A.; Boja, N.; Domuța, C.; Oneț, C. Effects of Common Oak (Quercus Robur L.) Defolition on the Soil Properties of an Oak Forest in Western Plain of Romania. Ann. For. Res. 2016, 59, 1. [Google Scholar] [CrossRef]
- Mattson, W.J.; Herms, D.A.; Witter, J.A.; Allen, D.C. Woody Plant Grazing Systems: North American Outbreak Folivores and Their Host Plants. General technical report NE - U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station 1991.
- Schowalter, T.D. Biology and Management of the Forest Tent Caterpillar (Lepidoptera: Lasiocampidae). J Integr Pest Manag 2017, 8. [Google Scholar] [CrossRef]
- Cooke, B.J.; Nealis, V.G.; Régnière, J. Insect Defoliators as Periodic Disturbances in Northern Forest Ecosystems. 2009, 39.
- Cooke, B.J.; Lorenzetti, F. The Dynamics of Forest Tent Caterpillar Outbreaks in Québec, Canada. Forest Ecology and Management 2006, 226, 110–121. [Google Scholar] [CrossRef]
- Sutton, A.S.; Tardif, J.C.T.C. Dendrochronological Reconstruction of Forest Tent Caterpillar Outbreaks in Time and Space, Western Manitoba, Canada. Canadian Journal of Forest Research 2007. [Google Scholar] [CrossRef]
- Baranchikov, Y.N.; Mattson, W.J.; Hain, F.P.; Payne, T.L. Forest Insect Guilds: Patterns of Interaction with Host Trees; 1989 August 13-17; Abakan, Siberia, U.S.S.R; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, 1991; p. NE-GTR-153;
- Moulinier, J.; Lorenzetti, F.; Bergeron, Y. Effects of a Forest Tent Caterpillar Outbreak on the Dynamics of Mixedwood Boreal Forests of Eastern Canada. Écoscience 2013, 20, 182–193. [Google Scholar] [CrossRef]
- Bergeron, Y. SPECIES AND STAND DYNAMICS IN THE MIXED WOODS OF QUEBEC’S SOUTHERN BOREAL FOREST. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Spencer, K.L.; Harvey, G.L. Understanding System Disturbance and Ecosystem Services in Restored Saltmarshes: Integrating Physical and Biogeochemical Processes. Estuarine, Coastal and Shelf Science 2012, 106, 23–32. [Google Scholar] [CrossRef]
- Vincent, J.-S.; Hardy, L. L’évolution et l’extension Des Lacs Glaciaires Barlow et Ojibway En Territoire Québécois. gpq 2011, 31, 357–372. [Google Scholar] [CrossRef]
- Canada, E. and C.C. Canadian Climate Normals - Climate - Environment and Climate Change Canada. Available online: https://climate.weather.gc.ca/climate_normals/ (accessed on 16 December 2022).
- Ministère de la Faune, des Forêts et des Parc (MFFP) Aires Infestées Par La Livrées Des Forêts Au Québec En 2016; Gouvernement du Québec, Direction de la protection des forêts: Québec, 2016; p. 10;
- Ministère de la Faune, des Forêts et des Parc (MFFP) Aires Infestées Par La Livrée Des Forêts Au Québec En 2017; Gouvernement du Québec, Direction de la protection des forêts: Québec, 2017; p. 14;
- Ministère de la Faune, des Forêts et des Parc (MFFP) Aires infestées par la livrée des forêts au Québec en 2018; 2018;
- Campbell, C.D.; Chapman, S.J.; Cameron, C.M.; Davidson, M.S.; Potts, J.M. A Rapid Microtiter Plate Method To Measure Carbon Dioxide Evolved from Carbon Substrate Amendments so as To Determine the Physiological Profiles of Soil Microbial Communities by Using Whole Soil. Appl. Environ. Microbiol. 2003, 69, 3593–3599. [Google Scholar] [CrossRef] [PubMed]
- Sassi, M.B.; Dollinger, J.; Renault, P.; Tlili, A.; Bérard, A. The FungiResp Method: An Application of the MicroRespTM Method to Assess Fungi in Microbial Communities as Soil Biological Indicators. Ecological Indicators 2012, 23, 482–490. [Google Scholar] [CrossRef]
- Anderson, J.P.E.; Domsch, K.H. A Physiological Method for the Quantitative Measurement of Microbial Biomass in Soils. Soil Biology and Biochemistry 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Campbell, C.D.; Grayston, S.J.; Hirst, D.J. Use of Rhizosphere Carbon Sources in Sole Carbon Source Tests to Discriminate Soil Microbial Communities. Journal of Microbiological Methods 1997, 30, 33–41. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. Journal of Statistical Software 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. Journal of Statistical Software 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Mellec, A. le M. le; Michalzik, B.M. Impact of a Pine Lappet (Dendrolimus Pini) Mass Outbreak on C and N Fluxes to the Forest Floor and Soil Microbial Properties in a Scots Pine Forest in Germany. Canadian Journal of Forest Research 2008. [CrossRef]
- Kristensen, J.A. The Biogeochemical Consequences of Litter Transformation by Insect Herbivory in the Subarctic: A Microcosm Simulation Experiment. 2018, 14.
- Donaldson, J.R.; Lindroth, R.L. Effects of Variable Phytochemistry and Budbreak Phenology on Defoliation of Aspen during a Forest Tent Caterpillar Outbreak. Agricultural and Forest Entomology 2008, 10, 399–410. [Google Scholar] [CrossRef]
- Hagerman, A.E. Recent Advances in Polyphenol Research | Wiley Online Books; 2012;
- Coq, S.; Souquet, J.-M.; Meudec, E.; Cheynier, V.; Hättenschwiler, S. Interspecific Variation in Leaf Litter Tannins Drives Decomposition in a Tropical Rain Forest of French Guiana. Ecology 2010, 91, 2080–2091. [Google Scholar] [CrossRef]
- Joanisse, G.D.; Bradley, R.L.; Preston, C.M.; Bending, G.D. Sequestration of Soil Nitrogen as Tannin–Protein Complexes May Improve the Competitive Ability of Sheep Laurel (Kalmia Angustifolia) Relative to Black Spruce (Picea Mariana). New Phytologist 2009, 181, 187–198. [Google Scholar] [CrossRef]
- Adamczyk, B.; Sietiö, O.-M.; Biasi, C.; Heinonsalo, J. Interaction between Tannins and Fungal Necromass Stabilizes Fungal Residues in Boreal Forest Soils. New Phytologist 2019, 223, 16–21. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Sun, T.; Coq, S. The Chitin Connection of Polyphenols and Its Ecosystem Consequences. New Phytologist 2019, 223, 5–7. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Cates, R.G.; Zou, J. In¯uence of Balsam Poplar Tannin Fractions on Carbon and Nitrogen Dynamics in Alaskan Taiga ¯oodplain Soils. Soil Biology 2001, 13. [Google Scholar] [CrossRef]
- Schimel, J.P.; Cleve, K.V.; Cates, R.G.; Clausen, T.P.; Reichardt, P.B. Effects of Balsam Poplar (Populus Balsamifera) Tannins and Low Molecular Weight Phenolics on Microbial Activity in Taiga Floodplain Soil: Implications for Changes in N Cycling during Succession. Can. J. Bot. 1996, 74, 84–90. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen Additions and Microbial Biomass: A Meta-analysis of Ecosystem Studies. 2008, 10.
- Parker, T.C.; Sadowsky, J.; Dunleavy, H.; Subke, J.-A.; Frey, S.D.; Wookey, P.A. Slowed Biogeochemical Cycling in Sub-Arctic Birch Forest Linked to Reduced Mycorrhizal Growth and Community Change after a Defoliation Event. Ecosystems 2017, 20, 316–330. [Google Scholar] [CrossRef]
- Saravesi, K.; Aikio, S.; Wäli, P.R.; Ruotsalainen, A.L.; Kaukonen, M.; Huusko, K.; Suokas, M.; Brown, S.P.; Jumpponen, A.; Tuomi, J.; et al. Moth Outbreaks Alter Root-Associated Fungal Communities in Subarctic Mountain Birch Forests. Microb Ecol 2015, 69, 788–797. [Google Scholar] [CrossRef]
- Frey, B.R.; Lieffers, V.J.; Landhäusser, S.M.; Comeau, P.G.; Greenway, K.J. An Analysis of Sucker Regeneration of Trembling Aspen. Can. J. For. Res. 2003, 33, 1169–1179. [Google Scholar] [CrossRef]
- Légaré, S.; Paré, D.; Bergeron, Y. Influence of Aspen on Forest Floor Properties in Black Spruce-Dominated Stands. Plant Soil 2005, 275, 207–220. [Google Scholar] [CrossRef]
- Chodak, M.; Klimek, B.; Niklińska, M. Composition and Activity of Soil Microbial Communities in Different Types of Temperate Forests. Biol Fertil Soils 2016, 52, 1093–1104. [Google Scholar] [CrossRef]
- Gartzia-Bengoetxea, N.; Kandeler, E.; Martínez de Arano, I.; Arias-González, A. Soil Microbial Functional Activity Is Governed by a Combination of Tree Species Composition and Soil Properties in Temperate Forests. Applied Soil Ecology 2016, 100, 57–64. [Google Scholar] [CrossRef]
- Paré, D.; Bernier, P.; Lafleur, B.; Titus, B.D.; Thiffault, E.; Maynard, D.G.; Guo, X. Estimating Stand-Scale Biomass, Nutrient Contents, and Associated Uncertainties for Tree Species of Canadian Forests. Can. J. For. Res. 2013, 43, 599–608. [Google Scholar] [CrossRef]
- Carter, M.R. Soil Sampling and Methods of Analysis; CRC Press, 2007. ISBN 978-0-429-12622-2.
Figure 1.
Mean microbial biomass (μg biomass-C g−1 soil hour−1) ± standard error from glucose-induced respiration of the top 5 cm of soil sampled under undefoliated control (N = 8, 6, 6) and defoliated (N = 7, 5, 7) Populus tremuloides trees. Soils were sampled in August 2018, May 2019 and July 2019 in the Lake Duparquet Research and Teaching Forest (Abitibi, QC).
Figure 1.
Mean microbial biomass (μg biomass-C g−1 soil hour−1) ± standard error from glucose-induced respiration of the top 5 cm of soil sampled under undefoliated control (N = 8, 6, 6) and defoliated (N = 7, 5, 7) Populus tremuloides trees. Soils were sampled in August 2018, May 2019 and July 2019 in the Lake Duparquet Research and Teaching Forest (Abitibi, QC).
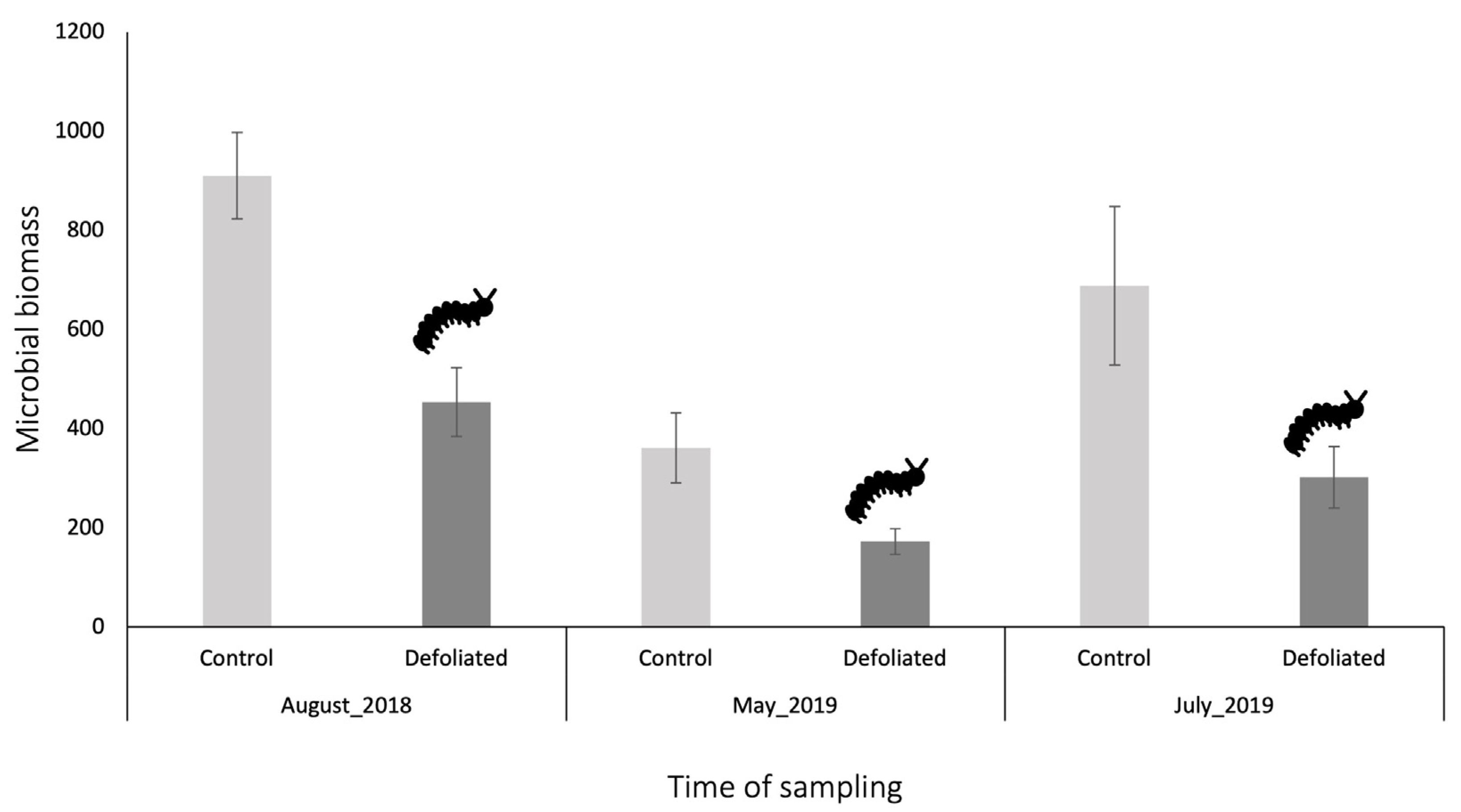
Figure 2.
Mean microbial respiration (μg biomass-C g−1 soil hour−1) ± standard error from carbon-based substrate-induced respiration of the top 5 cm of soil sampled under undefoliated control (N = 8) and defoliated (N = 7) Populus tremuloides trees. Soils were sampled in August 2018 in the Lake Duparquet Research and Teaching Forest (Abitibi, QC).
Figure 2.
Mean microbial respiration (μg biomass-C g−1 soil hour−1) ± standard error from carbon-based substrate-induced respiration of the top 5 cm of soil sampled under undefoliated control (N = 8) and defoliated (N = 7) Populus tremuloides trees. Soils were sampled in August 2018 in the Lake Duparquet Research and Teaching Forest (Abitibi, QC).
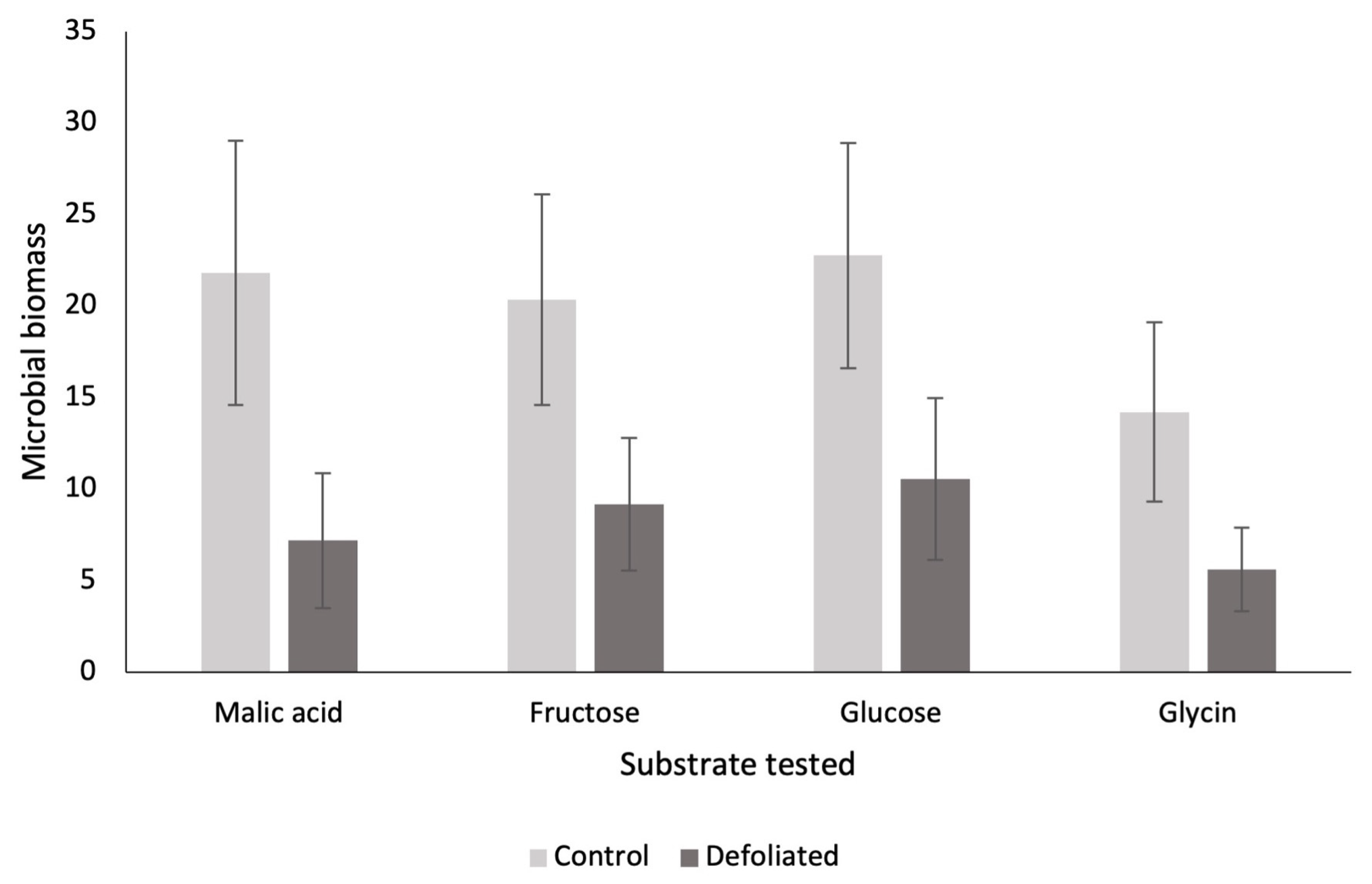
Figure 3.
Relative respiration with ± standard error from four different carbon substrate-induced respiration of the top 5 cm of soil under undefoliated control (N = 8 white) and defoliated (N = 6 black) Populus tremuloides trees. Soils were sampled in August 2018 in the Lac Duparquet Research and Training Forest (Abitibi, QC).
Figure 3.
Relative respiration with ± standard error from four different carbon substrate-induced respiration of the top 5 cm of soil under undefoliated control (N = 8 white) and defoliated (N = 6 black) Populus tremuloides trees. Soils were sampled in August 2018 in the Lac Duparquet Research and Training Forest (Abitibi, QC).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



