
Submitted:
08 March 2023
Posted:
13 March 2023
You are already at the latest version
Abstract
This study investigates the effects of the antiperspirant aluminum chlorohydrate on the development of antibiotic resistance in commensal Staphylococcus epidermidis isolates. The isolates were exposed to aluminum chlorohydrate for 30 days, respectively. The bacteria that developed resistance to oxacillin, and ciprofloxacin were isolated, and the expression levels of some antibiotic resistance genes were determined by quantitative reverse transcriptase-PCR. Before and after the exposure, the minimum inhibitory concentration (MIC) values of the bacteria were determined by the microdilution method. A time-dependent increase was observed in the number of bacteria that developed resistance and increased the MIC value. Consistent with the ciprofloxacin resistance observed after exposure, an increase in the norA, norB/C, gyrA, gyrB, parC, and parE gene expressions was observed. In addition to aluminum chlorohydrate exposure, oxacillin resistance was observed in all test bacteria in the group subcultured only in the medium, suggesting that phenotypic resistance cannot be correlated with chemical exposure in the light of these data. The increase in mecA gene expressions of selected test bacteria that acquired resistance to oxacillin after exposure compared with control groups suggests that the observed resistance may be related to aluminum chlorohydrate exposure. To our knowledge, this is the first time in the literature that the effects of aluminum chlorohydrate as an antiperspirant on the development of antibiotic resistance in Staphylococcus epidermidis have been reported.
Keywords:
aluminum chlorohydrate
; antibiotic resistance
; minimum inhibitory concentration
; quantitative reverse transcriptase-PCR
; Staphylococcus epidermidis
1. Introduction
Antibiotic resistance is one of the most serious global public health problems today. If necessary precautions are not taken, ten million people are expected to die annually from antibiotic-resistant infections by 2050. In addition to the inappropriate use of antibiotics in humans and animals for therapeutic purposes, the unnecessary and misuse of antibiotics, especially in the food and agricultural sectors for economic reasons, is one of the main reasons for the increase in antibiotic resistance. For this reason, precautions have been taken on antibiotic consumption in the fight against antibiotic resistance. Considering the increasing resistance rates, it is seen that the measures taken are not sufficient to solve the problem. New perspectives with more holistic approaches are needed to solve this problem [1,2,3,4,5,6].
Our body is rapidly colonized with microorganisms (bacteria, fungi, viruses, etc.) from the moment of birth. The community of these microorganisms is referred to as the human microbiome. The microbiota provides many benefits to the host [7,8]. Staphylococcus epidermidis, a member of the normal commensal skin flora of the human body, is primarily found in the axilla, on the head and in the nostril. S. epidermidis has a significant role in the skin ecosystem. It protects and prevents microbiota imbalance by fighting pathogens and participating in skin homeostasis through the production of beneficial bacterial metabolites. The widespread colonization of this bacterium on human skin and mucous membranes provides the opportunity for this bacterium to become infectious if suitable conditions are created [9,10,11]. The pathogenicity of S. epidermidis is mainly due to its ability to form biofilms on medical devices such as indwelling catheters and implanted devices. S. epidermidis can cause infections of prosthetic joints, vascular grafts, central nervous system shunts, surgical sites, and cardiac devices. Eradication of S. epidermidis infections is difficult because the bacteria in the biofilm are protected from immune system attack and antibacterial treatment. In recent years, S. epidermidis has become the most important cause of nosocomial infections [9,12,13]. It has been reported that resistance against many antibiotics, such as methicillin, erythromycin, ciprofloxacin, and gentamicin, develops in S. epidermidis strains isolated as a nosocomial infection agent today [14,15,16,17].
Cosmetics used by individuals from almost all age groups, have a wide range of products. There are various chemicals in the content of these products. Repeated and long-term exposure to cosmetics, many of which are used daily, can cause various effects on the organism and the microbiome [18,19,20]. Antiperspirants are cosmetic products widely used in society to prevent sweating. Many antiperspirants (aerosol, roll-on, stick, cream) generally used daily, contain aluminum salts for this purpose. Aluminum chlorohydrate is an inorganic salt consisting of complex basic aluminum chloride. Its pH is about 4.5. This compound does not cause skin irritation because it is less acidic compared to other aluminum salts. Studies have reported that exposure to aluminum compounds from cosmetics is associated with chest diseases, breast cancer, and Alzheimer's disease [18,20,21,22]. According to the literature review, no study was found investigating the effects of aluminum chlorohydrate on the skin microbiota.
Recent studies show that some non-antibiotic long-term medications may play a role in the development of antibiotic resistance [2,6]. In addition to non-antibiotic drugs, the possibility that cosmetics used daily may play a role in the development of antibiotic resistance should also be considered. The purpose of this study is to investigate the effects of aluminum chlorohydrate, commonly used in deodorants for its antiperspirant properties, on the development of antibiotic resistance in S. epidermidis, a member of the normal human axillary flora. According to the literature review, this study was the first to demonstrate the effect of aluminum chlorohydrate on the development of antibiotic resistance in S. epidermidis, a member of the skin microbiome.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
Twenty-two (oxacillin, ciprofloxacin) susceptible and one resistant S. epidermidis isolated from the axilla were used. S. epidermidis ATCC 12228 (susceptible strain) was used as standard strain. Antibiotic susceptibilities of all test bacteria were confirmed by the disc diffusion test. In the susceptibility tests, Staphylococcus aureus ATCC 25923 was used as a control [23].
Trypticase Soy Broth (Merck, Germany) and Trypticase Soy Agar (Merck, Germany) were used as initial growth media. Mueller Hinton Agar (MHA) (Merck, Germany) and Mueller Hinton Broth (MHB) (Merck, Germany) were used in antibiotic susceptibility tests. The aluminum chlorohydrate (Sigma-Aldrich, Germany) exposure was performed in Lysogeny Broth (LB) (Merck, Germany). MHA plates containing 0.5 μg ml−1 oxacillin (Sigma-Aldrich, Germany) and 4 μg ml−1 ciprofloxacin (Sigma-Aldrich, Germany) were used to isolate antibiotic-resistant test bacteria. All test bacteria were incubated at 37 °C for 18-24 h.
2.2. Detection of Antibiotic Resistance Genes
The presence of the mecA gene encoding PBP2a synthesis, the gyrA and gyrB genes encoding the DNA gyrase enzyme, the parC and parE genes encoding the topoisomerase IV, and the norA and norB/C genes, which play a role in regulating the efflux pump in the test bacteria, was detected by polymerase chain reaction (PCR) [24,25,26,27,28,29]. Gel electrophoresis was used to detect amplification products, which were visualized using SafeView Classic (ABM, Canada). The size of PCR products was compared with a 100bp DNA ladder (New England BioLabs, USA).
2.3. Determination of Minimum Inhibitory Concentration Values
Before and after exposure to aluminum chlorohydrate, MIC values of test bacteria against oxacillin and ciprofloxacin were determined by the microdilution method [23]. Increase in the MIC values was detected after exposure. Each test bacterium was tested in triplicate.
2.4. Aluminum Chlorohydrate Exposure and Isolation of Resistant Bacteria Against Test Antibiotics
Considering the Scientific Committee on Consumer Safety 2018 guidelines, the daily exposure amount from applying non-spray antiperspirants to the armpits twice a day was set at 1.5 g. The average highest concentration of aluminum chlorohydrate in antiperspirants on the market was assumed to be 20% [30,31]. Based on this information, the daily exposure to aluminum chlorohydrate was determined to be a minimum of 300 mg/L, and the aluminum chlorohydrate exposure concentrations of bacteria were adjusted according to this value. Due to the rapid moisture absorption of aluminum chlorohydrate, the aluminum chlorohydrate solution was freshly prepared with sterile distilled water at each step of the study.
First, the exposure concentration of aluminum chlorohydrate was determined by MIC test [23]. 40 μl suspension of the test bacteria (adjusted to 108-109 cfu ml-1) was added to 3.96 ml LB containing aluminum chlorohydrate (300 mg/L). After incubation at 37 °C for 18-24 h, 40 μl of the test bacteria cultures were transferred to 3.96 ml of fresh LB containing the concentration of aluminum chlorohydrate above-stated and incubated again. This subculturing procedure was repeated continuously for 30 days. On the 10th and 30th days of this procedure, 100 μl of the exposed test bacteria samples were plated on the MHA plates with 0.5 μg ml−1 oxacillin and 4 μg ml−1 ciprofloxacin. After 24 hours of incubation at 37 °C, colonies formed on the MHA plates containing test antibiotics were found to be resistant. Test bacteria subcultured in LB broth without aluminum chlorohydrate was used as the control group [2,3,4,5,6].
2.5. Quantitative Reverse Transcriptase PCR Analysis of Target Genes
The expression levels of the target genes of test bacteria were determined by Quantitative Reverse Transcriptase PCR (RT-qPCR). The expression levels of mecA, gyrA, gyrB, parC, parE, norA, norB, norC genes were compared with control groups [6,26,27,32]. Total RNA was extracted using an RNA isolation kit (FavorPrepTM, Taiwan) according to the manufacturer’s instructions. Total RNA was quantified in each sample using a NanoDrop spectrophotometer (Thermo Scientific, USA). cDNA was synthesized using iScriptTM cDNA reverse transcription kit (Bio-Rad, USA) according to the manufacturer’s instructions. An iTaqTM Universal SYBR® Green Supermix Kit (2x) (Bio-Rad®, USA) was used to determine the expression levels of target genes. RT-qPCR analysis was performed for each 10 µl PCR reaction mixture (5 µl iTaqTM Universal SYBR® Green Supermix 2X, 3 µl ddH2O, 1 µl cDNA template, 0.5 µl forward primer and 0.5 µl reverse primer). Relative gene expression values were calculated by the 2 −ΔΔCT method using an RT2 Profiler PCR Array Data Analysis v3.5 (Qiagen, Hilden, Germany) analysis program. All data were normalized to 16S rRNA housekeeping gene expression levels.
3. Results and Discussion
3.1. Detection of Antibiotic Resistance Genes
Agarose gel electrophoresis images of antibiotic resistance genes are given in Figure 1.
3.2. MIC Values of Aluminum Chlorohydrate
No antibacterial effect of aluminum chlorohydrate was observed against test bacteria in the range of 187.5-24,000 mg L-1. Therefore, the previously reported concentration of 300 mg L-1 determined as the exposure concentration for the test bacteria to aluminum chlorohydrate was considered appropriate.
3.3. Antibiotic Resistance Development After Aluminum Chlorohydrate Exposure
The number of susceptible test bacteria that developed phenotypic resistance to oxacillin and ciprofloxacin on day 10, and day 30 of exposure is shown in Figure 2. The resistant test bacteria are listed in Table 1.
When considering the results of phenotypic resistance development after exposure, it was found that the number of test bacteria that developed resistance to ciprofloxacin increased. The strain 24 (S. epidermidis ATCC 12228), which developed resistance to ciprofloxacin after 30 days of exposure, also developed resistance in the control group, which was subcultured only in the medium for 30 days. When considering the relative expression results between the two test conditions, increased gene expression of norA, norB/C, gyrA, gyrB, parC, and parE was observed in S. epidermidis ATCC 12228 exposed to aluminum chlorohydrate, consistent with phenotypic resistance. This result suggests that the observed ciprofloxacin resistance developed as a consequence of exposure to aluminum chlorohydrate (Figure 3, Table 2).
The development of ciprofloxacin resistance in isolate 14 subcultured only in LB broth without aluminum chlorohydrate for 30 days, which did not develop resistance to ciprofloxacin as a result of exposure, suggests that long-term subculturing also played a role in the development of resistance. The relative expression results of the control group test bacteria, which developed resistance to ciprofloxacin by subculturing only in LB broth without any aluminum chlorohydrate for 30 days, and the control group test bacteria taken on the first day as the starter, support this phenotypic resistance result. In the stated conditions, an increase in norA, norB/C, gyrA, gyrB, parC, and parE gene expressions were observed for test bacteria 14 and 24, consistent with the development of phenotypic resistance to ciprofloxacin (Figure 4, Table 3).
Ciprofloxacin is a bactericidal antibiotic of the fluoroquinolone class used to treat infections such as urinary tract infections and pneumonia. The mechanism of action of ciprofloxacin is inhibition of DNA replication by inhibition of bacterial DNA topoisomerase and DNA gyrase enzymes. The development of resistance to this group of antibiotics occurs through point mutations in genes encoding DNA gyrase and topoisomerase IV or mutations in genes regulating efflux-pump proteins [33,34]. The amino-terminal domains of GyrA or ParC, covalently bound to DNA in an enzyme intermediate are the sites where these resistance mutations are most frequently detected. Quinolone resistance has also been associated with alterations in specific domains of GyrB and ParE; however, these alterations are much less common in resistant clinical bacterial isolates than mutations in GyrA or ParC [35]. In this study, an increase in norA, norB/C, gyrA, gyrB, parC, and parE gene expressions was observed in agreement with phenotypic ciprofloxacin resistance. Although efflux pumps exhibit substrate specificity, most of them can also pump different chemical compounds out of the cell. This situation is effective in the development of multidrug resistance in bacteria. More than ten efflux pump genes have been detected in methicillin-resistant Staphylococcus aureus (MRSA) strains. The most frequently detected norA and norB/C genes are chromosomally encoded efflux genes that are also present in S. epidermidis strains. Demarco et al. (2007) reported that nearly half (54/114 strains) of the S. aureus isolates tested effluxed at least two structurally distinct substrates [36,37,38]. In our study, an increase in the expression levels of norA and norB/C efflux genes was detected. This could play a role in the development of resistance to other antibiotics and lead to the emergence of multidrug resistant strains.
Methicillin resistance occurs in two different phenotypes: homogeneous and heterogeneous. Homogeneous resistance refers to a cell population in which all cells are resistant to high concentrations of methicillin. In heterogeneous resistance, although all cells carry the mecA gene in the bacterial community, only a small minority of bacteria exhibit high levels of methicillin resistance [39]. mecA gene is not expressed in heterogeneous resistant staphylococcal strains, and bacteria that should be resistant may be found to be susceptible in routine susceptibility testing [40]. However, isolates that do not carry the mecA gene have also been reported to be resistant to oxacillin [41]. In our study, oxacillin resistance was observed after 30 days in both groups subcultured in medium only and exposed to aluminum chlorohydrate. PCR revealed that the mecA gene was found only in some of the test bacteria (Figure 1). Since all test bacteria developed resistance to oxacillin after 30 days of exposure, changes in gene expression were determined by sampling in two of them. Test bacteria 14 and 22 were selected for this purpose. The relative expression results for the mecA gene of test bacteria 14 and 22, which had developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate, and the control group test bacteria taken on the first day are shown in Figure 5. Under the stated conditions, an increase in mecA gene expression was observed in test bacteria 14 and 22, consistent with the development of phenotypic resistance to oxacillin (Figure 5, Table 4).
The relative expression results for the mecA gene of test bacteria 14 and 22 that developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate and the control group of test bacteria that developed resistance to oxacillin by 30 days of subculturing only in LB broth without aluminum chlorohydrate are shown in Figure 6. Under the stated conditions, an increase in mecA gene expression was observed in test bacteria 14 and 22, consistent with the development of phenotypic resistance to oxacillin (Figure 6, Table 5). An increase in gene expression of fourfold or more was considered significant.
Since oxacillin resistance was detected in all isolates of the control group subcultured for 30 days only in LB broth without aluminum chlorohydrate, it could not be correlated with the resistance developed after aluminum chlorohydrate exposure. However, the increase in mecA gene expressions of the selected test bacteria that became resistant to oxacillin after exposure compared to the control groups suggests that the observed oxacillin resistance is related to aluminum chlorohydrate exposure.
3.4. Increase in the Minimum Inhibitory Concentration Values of the Test Bacteria
The MIC values of the bacteria before and after exposure to the test antibiotics are shown in Table 6. The LB broth without aluminum chlorohydrate was used as a control.
Considering the MIC values of all test bacteria before and after exposure, it was found that the MIC values of isolates that developed ciprofloxacin resistance as a result of exposure to aluminum chlorohydrate increased 4-32-fold. Although all test bacteria developed resistance to oxacillin as a result of exposure, it was found that the MIC values of most of them increased 4-256-fold.
Thanks to their excellent adaptability, microorganisms adapt to the new ecological environments they encounter. They try to maintain their vitality and generation in these new environments thanks to their resistance mechanisms [42]. Although the improper use of antibiotics is considered the main cause of the development of antibiotic resistance, studies from recent years have shown that some non-antibiotic drugs may also play a role in promoting antibiotic resistance. These drugs include antidepressants. Jin et al. (2018) was the first to report that fluoxetine use induced antibiotic resistance in Escherichia coli. In this study, expression of the multidrug efflux pump genes acrB and acrD increased as a result of a 10 h exposure of E. coli to fluoxetine, while expression levels of the outer membrane porin genes ompF and ompW decreased. The bacterium protected itself from exposure to fluoxetine by expelling the drug from the cell and preventing the drug from entering the cell. It was found that a-30-day exposure to fluoxetine caused an increase in chloramphenicol, amoxicillin, and tetracycline resistance. In addition, these mutants exhibited multiple resistance to fluoroquinolones, aminoglycosides, and beta-lactams. Another study by our research group examined the effects of fluoxetine, sertraline, and amitriptyline, commonly used antidepressants, on the development of antibiotic resistance in clinical Acinetobacter baumannii isolates. The isolates were exposed to fluoxetine, sertraline, and amitriptyline for 30 days, respectively. The bacteria that developed resistance to gentamicin, imipenem, colistin, and ciprofloxacin were isolated, and the expression levels of some antibiotic resistance genes were determined by quantitative reverse transcriptase-PCR. Before and after exposure, MICs of bacteria were determined by the microdilution method. A time-dependent increase in the number of bacteria that developed resistance and increased MICs was observed. After exposure to fluoxetine and sertraline, decreases in efflux and outer membrane porin genes were observed in isolates that developed colistin resistance, whereas increases were observed in isolates that developed ciprofloxacin resistance. These observations suggest that these antidepressants have similar effects on the development of resistance. While the exposure to fluoxetine did not result in the development of resistance to imipenem, it was observed after exposure to sertraline and amitriptyline, and a common decrease in ompA gene expression was observed in these isolates [6]. Wang et al. (2021) reported that nonsteroidal anti-inflammatory drugs (ibuprofen, naproxen, diclofenac), a lipid-lowering drug (gemfibrozil), and a β- blocker (propranolol) accelerated the spread of antibiotic resistance through plasmid-borne bacterial conjugation. In addition to nonantibiotic drugs, the possibility that cosmetics used daily may play a role in the development of antibiotic resistance should also be considered.
The response of microorganisms to chemical changes in their environment has long been one of the most important topics of research. However, concrete data on how the environmental changes that cause stress conditions for microorganisms may contribute to antimicrobial resistance are not yet avaliable. According to the literature review, this study was the first demonstrate the effect of exposure to aluminum chlorohydrate on the development of antibiotic resistance in S. epidermidis, a member of the skin microbiome. Because of the repeated and long-term use of cosmetics, the effect of chemicals on the resistance mechanisms of bacteria in the microbiota should be investigated with more comprehensive phenotypic and genotypic analyzes.
Supplementary Materials
The following supporting information can be downloaded at: Table S1. The sequences of primers used for the Polymerase Chain Reaction.
Author Contributions
Conceptualization, A.A. and M.E.; methodology, A.A.; M.M.; S.S.R.; B.K. and E.S.T; validation, A.A.; M.M.; S.S.R.; B.K. and E.S.T; formal analysis, A.A.; M.M.; S.S.R.; B.K. and E.S.T; investigation, A.A.; M.M.; S.S.R.; B.K. and E.S.T; resources, A.A. and M.M.; data curation, A.A.; M.M.; B.K. and E.S.T; writing—original draft preparation, M.M.; writing—review and editing, A.A.; M.M.; and B.K.; visualization, M.M.; supervision, M.M; project administration, M.M; funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
“This research was funded by Ankara University Scientific Research Council, grant number 20L0237006”.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Acknowledgments
We are very grateful to Prof. Barış Otlu at Inonu University (Malatya, Türkiye) for his support in our study.
Conflicts of Interest
The authors declare no conflict of interest.
Ethical Statements
Ethical approval was received from Ethics Committee of Ankara University Faculty of Medicine (Reference No: I5-196- 19) (14 November 2019).
References
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A review of antibiotic use in food animals: perspective, policy, and potential. Public Health Rep 2012, 127, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Lu, J.; Chen, Z.; Nguyen, S.H.; Mao, L.; Li, J.; Yuan, Z.; Guo, J. Antidepressant fluoxetine induces multiple antibiotics resistance in Escherichia coli via ROS-mediated mutagenesis. Environ Int 2018, 120, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Morehead, M.S.; Scarbrough, C. Emergence of global antibiotic resistance. Prim Care. 2018, 45, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Antibiotic resistance threats in the United States 2019. Available online: https://www.cdc.gov/ drugresistance/pdf/threats-report/2019-ar-threats-report508.pdf (accessed on 15 October 2021).
- World Health Organization. Antibiotic resistance. Available online: https:// www.who.int/news-room/factsheets/detail/antibiotic-resistance (accessed on 23 August 2021).
- Gurpinar, S.S.; Kart, D.; Eryilmaz, M. The effects of antidepressants fluoxetine, sertraline, and amitriptyline on the development of antibiotic resistance in Acinetobacter baumannii. Arch Microbiol 2022, 204, 230. [Google Scholar] [CrossRef] [PubMed]
- Haran, J.P.; McCormick, B.A. Aging, frailty, and the microbiome-how dysbiosis influences human aging and disease. Gastroenterology 2021, 160, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, F.; Ghosh, T.S.; O'toole, P.W. The healthy microbiome-What is the definition of a healthy gut microbiome? Gastroenterology 2021, 160, 483–494. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis--the 'accidental' pathogen. Nat Rev Microbiol 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Brooks, G.F.; Carroll, K.C.; Butel, J.S.; Morse, S.A.; Mietzner, T.A. Jawetz, Melnickk ve Adelberg Tıbbi Mikrobiyoloji. Ç. Dr. Osman Şadi Yenen: Nobel Kitapevleri, Ankara, 2014; p. 167. [Google Scholar]
- Fournière, M.; Latire, T.; Souak, D.; Feuilloley, M.G.J.; Bedoux, G. Staphylococcus epidermidis and Cutibacterium acnes: Two major sentinels of skin microbiota and the influence of cosmetics. Microorganisms 2020, 8, 1752. [Google Scholar] [CrossRef] [PubMed]
- Vuong, C.; Otto, M. Staphylococcus epidermidis infections. Microbes Infect 2002, 4, 481–489. [Google Scholar] [CrossRef]
- Eryilmaz, M.; Gurpinar, S.S. Investigation of the antibacterial efficacy of some commonly used antiseptics in hospitals against biofilm forming and non-biofilm forming Staphylococcus epidermidis strains. J Fac Pharm Ankara 2017, 41, 1–8. [Google Scholar]
- Cerca, N.; Martins, S.; Cerca, F.; Jefferson, K.K.; Pier, G.B.; Oliveira, R.; Azeredo, J. Comparative assessment of antibiotic susceptibility of coagulase-negative Staphylococci in biofilm versus planktonic culture as assessed by bacterial enumeration or rapid XTT colorimetry. J Antimicrob Chemother 2005, 56, 331–336. [Google Scholar] [CrossRef]
- Morgenstern, M.; Erichsen, C.; von Rüden, C.; Metsemakers, W.J.; Kates, S.L.; Moriarty, T.F.; Hungerer, S. Staphylococcal orthopaedic device-related infections in older patients. Injury 2016, 47, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Sabaté Brescó, M.; Harris, L.G.; Thompson, K.; Stanic, B.; Morgenstern, M.; O'Mahony, L.; Richards, R.G.; Moriarty, T.F. Pathogenic mechanisms and host interactions in Staphylococcus epidermidis device-related infection. Front Microbiol 2017, 8, 1401. [Google Scholar] [CrossRef] [PubMed]
- Bouiller, K.; Ilic, D.; Wicky, P.H.; Cholley, P.; Chirouze, C.; Bertrand, X. Spread of clonal linezolid-resistant Staphylococcus epidermidis in an intensive care unit associated with linezolid exposure. Eur J Clin Microbiol Infect Dis 2020, 39, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
- Callewaert, C.; Hutapea, P.; Van de Wiele, T.; Boon, N. Deodorants and antiperspirants affect the axillary bacterial community. Arch Dermatol Res 2014, 306, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Glatz, M.; Jo, J.H.; Kennedy, E.A.; Polley, E.C.; Segre, J.A.; Simpson, E.L.; Kong, H.H. Emollient use alters skin barrier and microbes in infants at risk for developing atopic dermatitis. PLoS One 2018, 13, e0192443. [Google Scholar] [CrossRef] [PubMed]
- Aras, A.; Eryilmaz, M. Microbiological contamination of cosmetic products. J Fac Pharm Ankara 2022, 46, 262–276. [Google Scholar]
- Holzle, E. Antiperspirants; Dermatopharmacology of Topical Preparations, Springer-Verlag: Berlin Heidelberg, 2000; pp. 401–402. [Google Scholar]
- Martini, M.C. Déodorants et antitranspirants [Deodorants and antiperspirants]. Ann Dermatol Venereol 2020, 147, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standartds for Antimicrobial Susceptibility Testing, CLSI supplement M100, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, 2020. [Google Scholar]
- Vandecasteele, S.J.; Peetermans, W.E.; Merckx, R.; Van Eldere, J. Quantification of expression of Staphylococcus epidermidis housekeeping genes with Taqman quantitative PCR during in vitro growth and under different conditions. J Bacteriol 2001, 183, 7094–7101. [Google Scholar] [CrossRef]
- Yamada, M.; Yoshida, J.; Hatou, S.; Yoshida, T.; Minagawa, Y. Mutations in the quinolone resistance determining region in Staphylococcus epidermidis recovered from conjunctiva and their association with susceptibility to various fluoroquinolones. Br J Ophthalmol 2008, 92, 848–851. [Google Scholar] [CrossRef]
- Betanzos-Cabrera, G.; Juárez-Verdayes, M.A.; González-González, G.; Cancino-Díaz, M.E.; Cancino-Díaz, J.C. Gatifloxacin, moxifloxacin, and balofloxacin resistance due to mutations in the gyrA and parC genes of Staphylococcus epidermidis strains isolated from patients with endophthalmitis, corneal ulcers and conjunctivitis. Ophthalmic Res 2009, 42, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Verdayes, M.A.; Parra-Ortega, B.; Hernández-Rodríguez, C.; Betanzos-Cabrera, G.; Rodríguez-Martínez, S.; Cancino-Diaz, M.E.; Cancino-Diaz, J.C. Identification and expression of nor efflux family genes in Staphylococcus epidermidis that act against gatifloxacin. Microb Pathog 2012, 52, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Sully, E.K.; Malachowa, N.; Elmore, B.O.; Alexander, S.M.; Femling, J.K.; Gray, B.M.; DeLeo, F.R.; Otto, M.; Cheung, A.L.; Edwards, B.S.; Sklar, L.A.; Horswill, A.R.; Hall, P.R.; Gresham, H.D. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoS Pathog 2014, 10, e1004174. [Google Scholar] [CrossRef] [PubMed]
- Eladli, M.G.; Alharbi, N.S.; Khaled, J.M.; Kadaikunnan, S.; Alobaidi, A.S.; Alyahya, S.A. Antibiotic-resistant Staphylococcus epidermidis isolated from patients and healthy students comparing with antibiotic-resistant bacteria isolated from pasteurized milk. Saudi J Biol Sci 2019, 26, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Zen, J.M.; Yang, T.H.; Kumar, A.S.; Chen, Y.J.; Hsu, J.C.; Shih, Y. Detection of aluminum chlorohydrate content in antiperspirant deodorants using screen-printed silver electrodes by one drop analysis. Electroanalysis 2009, 21, 2272–2276. [Google Scholar] [CrossRef]
- Scientific Committee on Consumer Safety (SCCS). The SCCS Notes of Guidance for the Testing of Cosmetic Ingredients and Their Safety Evaluation 10th Revision. SCCS/1628/18: 24-25, 2018.
- Kart, D.; Gurpinar, S.S.; Eryilmaz, M. Assessment of the anti-quorum sensing effect of Lactobacillus sp. metabolites on expression levels of QS-related genes in Pseudomonas aeruginosa PAO1. Turk Bull Hyg Exp Biol 2020, 77, 311–318. [Google Scholar] [CrossRef]
- Yilmaz, E. Quinolones. Turkiye Klinikleri J Inf Dis-Special Topics 2017, 10, 99–105. [Google Scholar]
- Thai, T.; Salisbury, B.H.; Zito, P.M. Ciprofloxacin. [Updated 2022 Sep 2]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2022 Jan. Available online: https://www.ncbi.nlm.nih.gov/books/NBK535454/.
- Shariati, A.; Arshadi, M.; Khosrojerdi, M.A.; Abedinzadeh, M.; Ganjalishahi, M.; Maleki, A.; Heidary, M.; Khoshnood, S. The resistance mechanisms of bacteria against ciprofloxacin and new approaches for enhancing the efficacy of this antibiotic. Front Public Health 2022, 10, 1025633. [Google Scholar] [CrossRef] [PubMed]
- DeMarco, C.E.; Cushing, L.A.; Frempong-Manso, E.; Seo, S.M.; Jaravaza, T.A.; Kaatz, G.W. Efflux-related resistance to norfloxacin, dyes, and biocides in bloodstream isolates of Staphylococcus aureus. Antimicrob Agents Chemother 2007, 51, 3235–3239. [Google Scholar] [CrossRef]
- Qin, R.; Xiao, K.; Li, B.; Jiang, W.; Peng, W.; Zheng, J.; Zhou, H. The combination of catechin and epicatechin callate from Fructus Crataegi potentiates beta-lactam antibiotics against methicillin-resistant Staphylococcus aureus (MRSA) in vitro and in vivo. Int J Mol Sci 2013, 14, 1802–1821. [Google Scholar] [CrossRef]
- Bostanmaneshrad, A.; Nowroozi, J.; Eslami, G.; Hashemi, A. The expression of efflux pump genes in methicillin-resistant Staphylococcus aureus (MRSA) strains isolated from blood cultures in Iran. Arch Clin Infect Dis 2020, 15, e99804. [Google Scholar] [CrossRef]
- Sancak, B. Resistance mechanisms of MRSA: Epidemiology in the World and Turkey. Ankem Derg 2012, 26, 38–47. [Google Scholar]
- Çiftci, İ.H.; Altindis, M.; Cetinkaya, Z.; Asik, G.; Aktepe, O.C. Investigation of mecA genes in Staphylococcus strains isolated from clinical samples. Kocatepe Med J 2009, 10, 17–20. [Google Scholar]
- Xu, Z.; Shah, H.N.; Misra, R.; Chen, J.; Zhang, W.; Liu, Y.; Cutler, R.R.; Mkrtchyan, H.V. The prevalence, antibiotic resistance and mecA characterization of coagulase negative Staphylococci recovered from non-healthcare settings in London, UK. Antimicrob Resist Infect Control 2018, 7, 73. [Google Scholar] [CrossRef]
- Tan, Y.S.; Zhang, R.K.; Liu, Z.H.; Li, B.Z.; Yuan, Y.J. Microbial adaptation to enhance stress tolerance. Front Microbiol 2022, 13, 888746. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, J.; Zhang, S.; Li, J.; Mao, L.; Yuan, Z.; Bond, P.L.; Guo, J. Non-antibiotic pharmaceuticals promote the transmission of multidrug resistance plasmids through intra- and intergenera conjugation. ISME J 2021, 15, 2493–2508. [Google Scholar] [CrossRef]
Figure 1.
Agarose gel electrophoresis images of mecA, gyrA, gyrB, parC, parE, norA, norB/C genes (M: DNA Ladder (100 bp), 1-22: The isolates susceptible to oxacillin and ciprofloxacin, 23: The isolate resistant to oxacillin and ciprofloxacin, 24: S. epidermidis ATCC 12228) (a) mecA (1039 bp), (b) gyrA (284 bp), (c) gyrB (250 bp), (d) parC (197 bp), (e) parE (325 bp), (f) norA (434 bp), (g) norB/C (563 bp)
Figure 1.
Agarose gel electrophoresis images of mecA, gyrA, gyrB, parC, parE, norA, norB/C genes (M: DNA Ladder (100 bp), 1-22: The isolates susceptible to oxacillin and ciprofloxacin, 23: The isolate resistant to oxacillin and ciprofloxacin, 24: S. epidermidis ATCC 12228) (a) mecA (1039 bp), (b) gyrA (284 bp), (c) gyrB (250 bp), (d) parC (197 bp), (e) parE (325 bp), (f) norA (434 bp), (g) norB/C (563 bp)

Figure 2.
Phenotypic antibiotic resistance development of susceptible test bacteria on the 10th, and 30th days of the exposure. (a) Aluminum chlorohydrate (300 mg/L) exposure, (b) Control (susceptible test bacteria subcultured in LB broth without aluminum chlorohydrate).
Figure 2.
Phenotypic antibiotic resistance development of susceptible test bacteria on the 10th, and 30th days of the exposure. (a) Aluminum chlorohydrate (300 mg/L) exposure, (b) Control (susceptible test bacteria subcultured in LB broth without aluminum chlorohydrate).
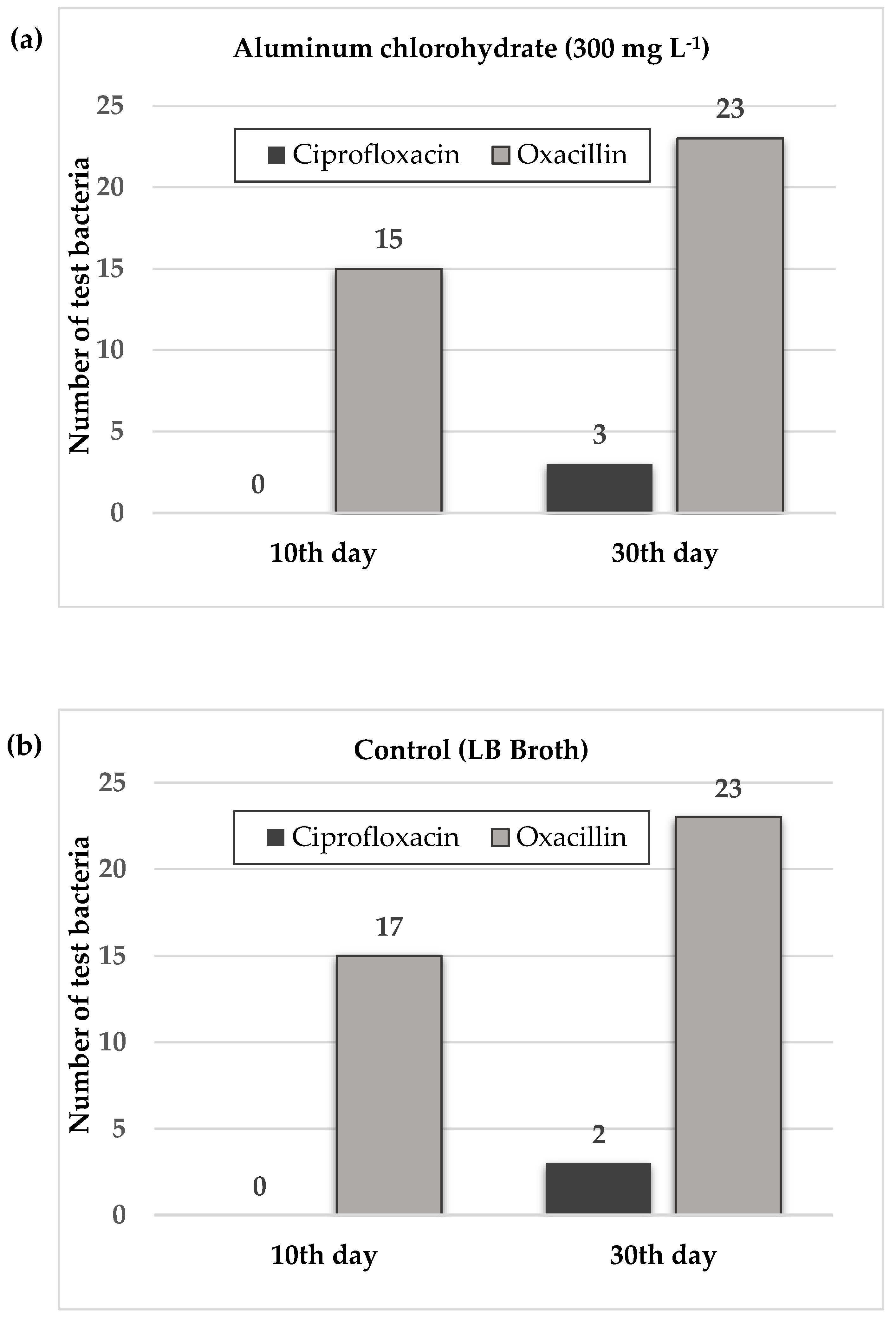
Figure 3.
Comparison of gene expression values of the strain 24 (S. epidermidis ATCC 12228), which developed resistance to ciprofloxacin after 30 days of exposure to aluminum chlorohydrate, and control bacteria that developed resistance to ciprofloxacin by subculturing only in LB broth without aluminum chlorohydrate for 30 days.
Figure 3.
Comparison of gene expression values of the strain 24 (S. epidermidis ATCC 12228), which developed resistance to ciprofloxacin after 30 days of exposure to aluminum chlorohydrate, and control bacteria that developed resistance to ciprofloxacin by subculturing only in LB broth without aluminum chlorohydrate for 30 days.
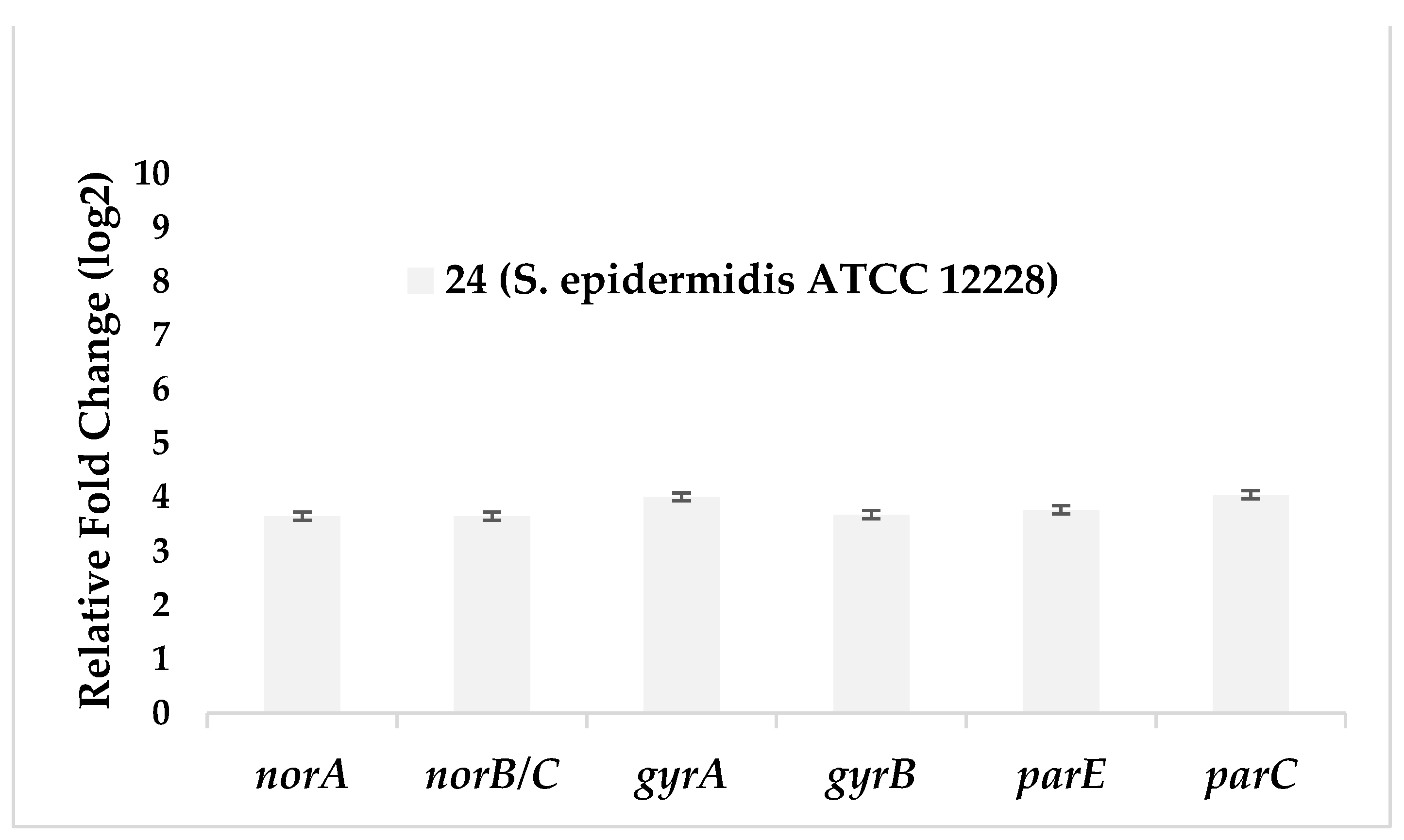
Figure 4.
Comparison of gene expression values of the test bacteria of the control group that developed resistance to ciprofloxacin by subculturing them only in LB broth without aluminum chlorohydrate for 30 days and the test bacteria of the control group that were taken as starter on the first day.
Figure 4.
Comparison of gene expression values of the test bacteria of the control group that developed resistance to ciprofloxacin by subculturing them only in LB broth without aluminum chlorohydrate for 30 days and the test bacteria of the control group that were taken as starter on the first day.
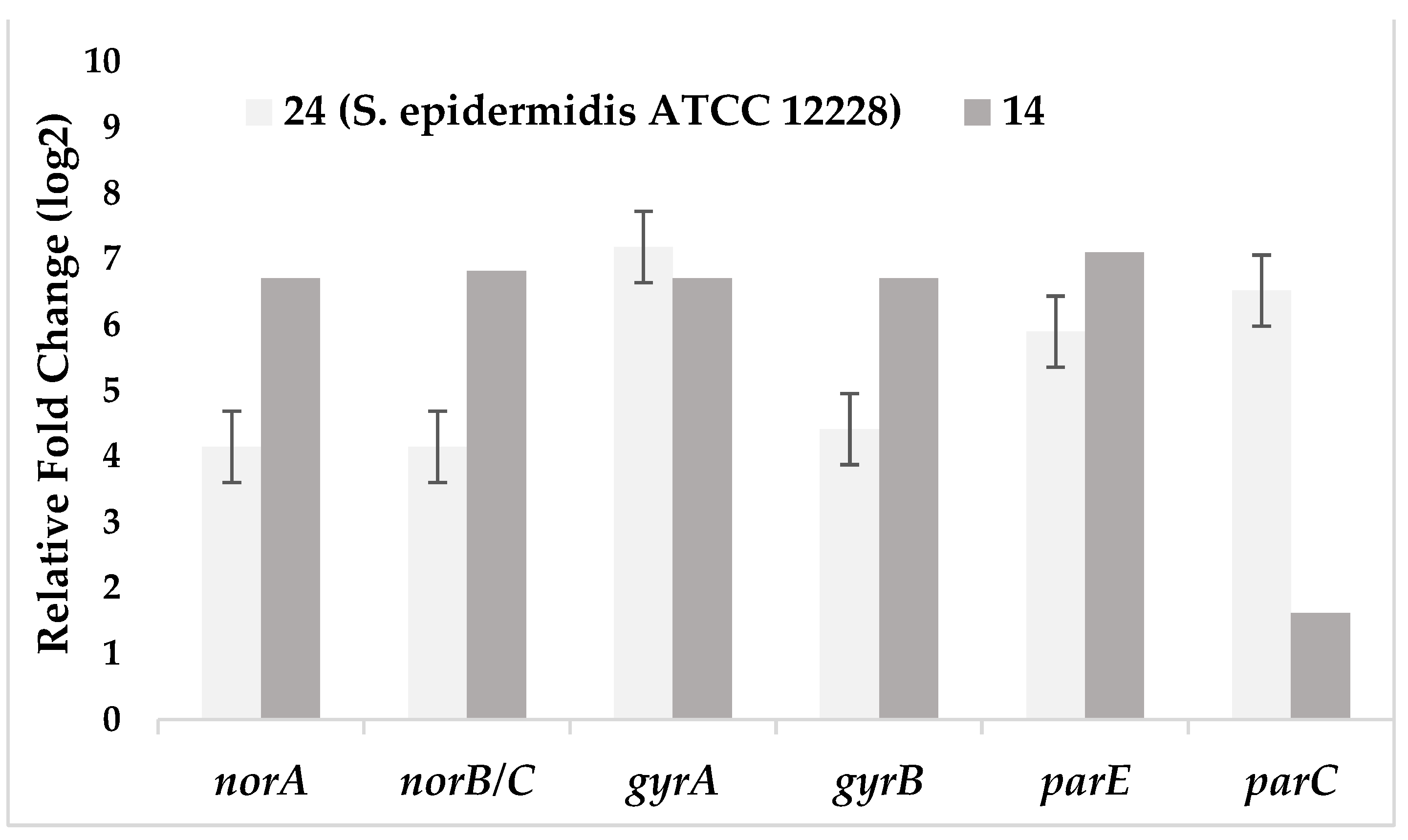
Figure 5.
Comparison of mecA gene expression levels of test bacteria 14 and 22 that developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate and the control group test bacteria taken on the first day as the starter
Figure 5.
Comparison of mecA gene expression levels of test bacteria 14 and 22 that developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate and the control group test bacteria taken on the first day as the starter
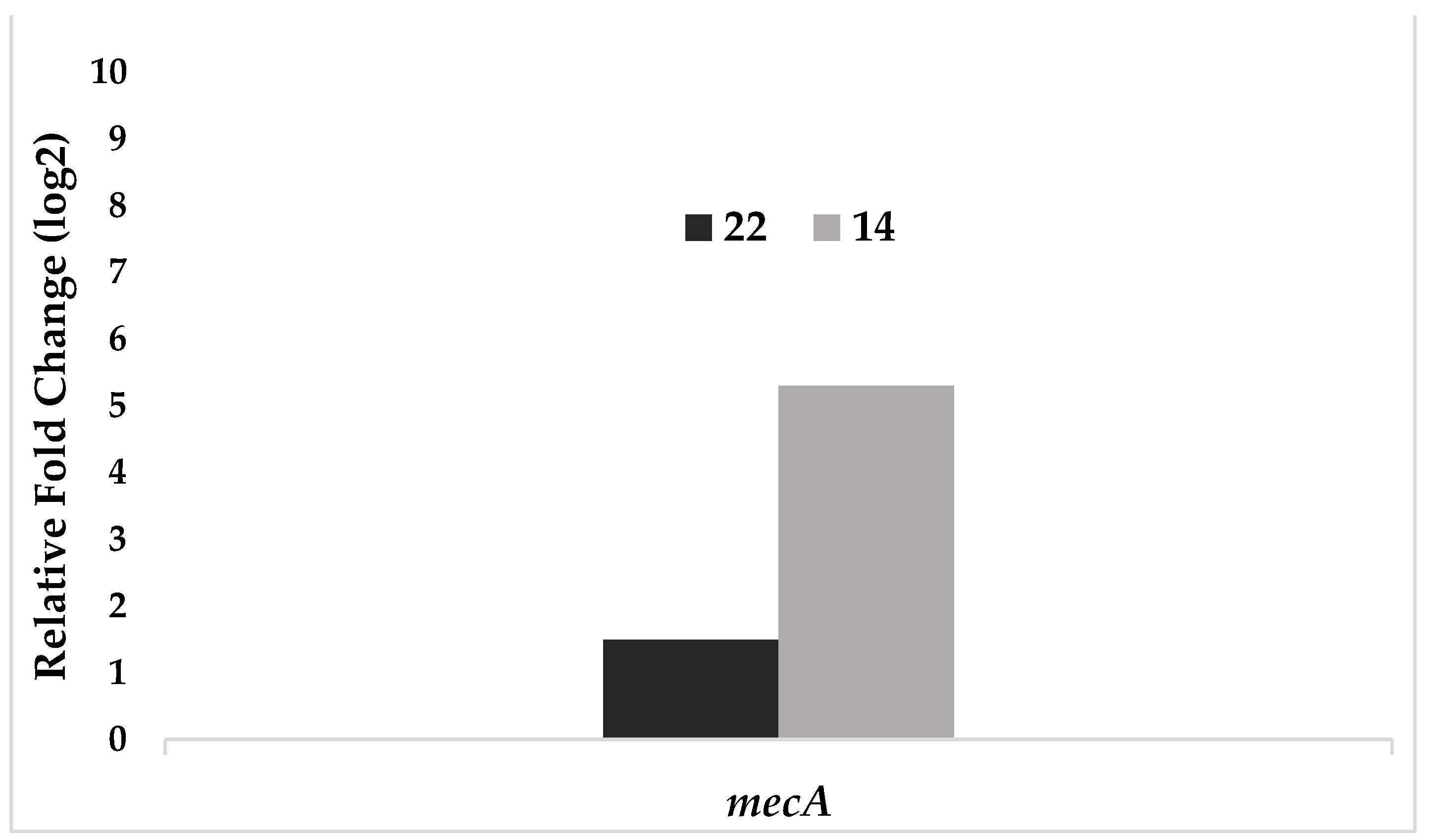
Figure 6.
Comparison of mecA gene expression values of the test bacteria 14 and 22, which developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate, and the control group test bacteria that developed resistance to oxacillin by subculturing only in LB broth without aluminum chlorohydrate for 30 days.
Figure 6.
Comparison of mecA gene expression values of the test bacteria 14 and 22, which developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate, and the control group test bacteria that developed resistance to oxacillin by subculturing only in LB broth without aluminum chlorohydrate for 30 days.
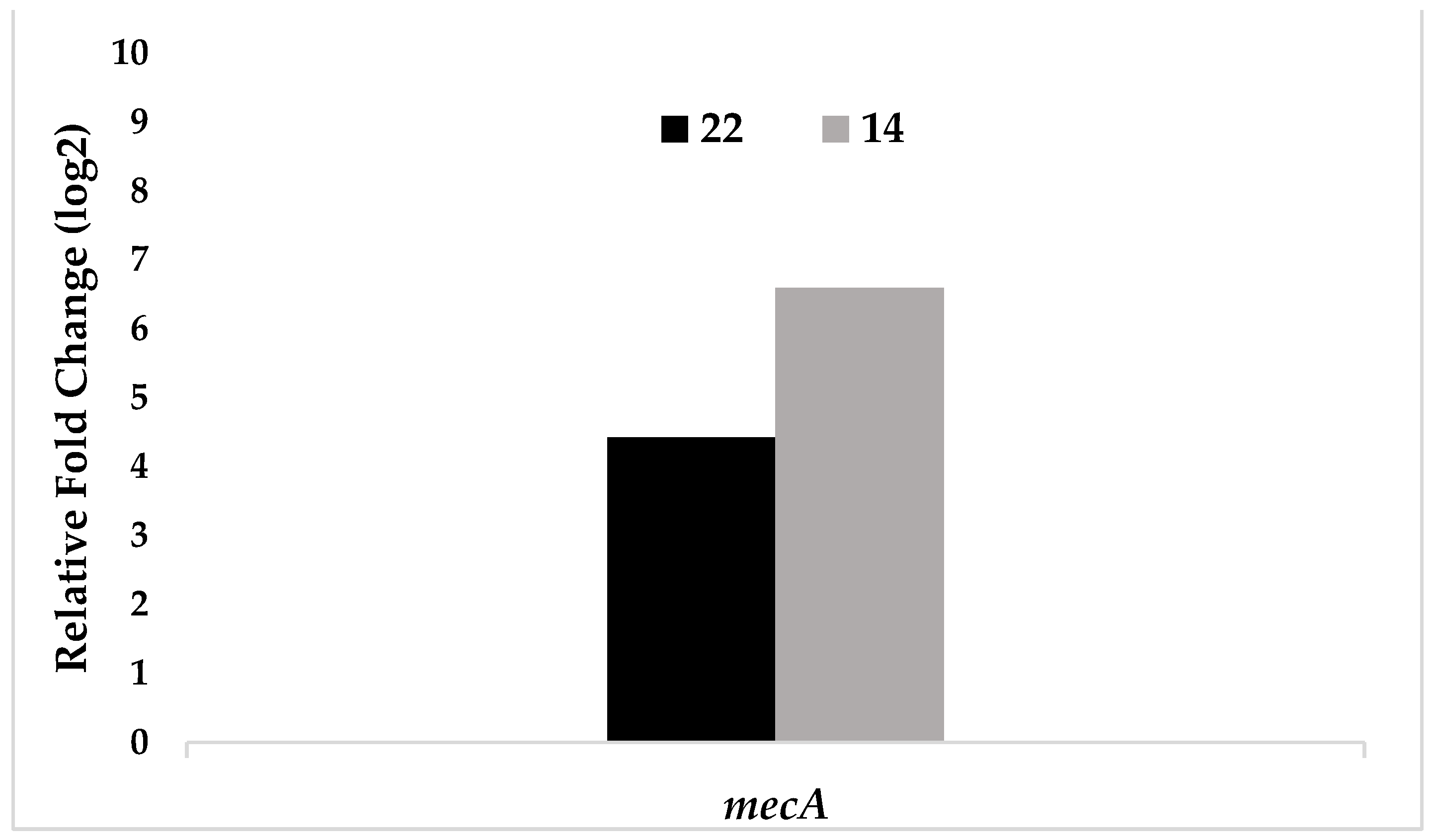

Table 1.
Susceptible test bacteria developed phenotypic resistance on the exposure times.
| Exposure day |
Test antibiotics |
Aluminum chlorohydrate (300 mg L-1) |
Control* |
|---|---|---|---|
| 10th | CIP | - | - |
| OXA | 1, 2, 3, 5, 12, 13, 14, 15, 16, 17, 18, 20, 21, 22, 24 | 1, 2, 3, 4, 6, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 22 | |
| 30th | CIP | 18, 22, 24 | 14, 24 |
| OXA | 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 24 | 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 24 |
1-22: Susceptible isolates, 24: S. epidermidis ATCC 12228. CIP: Ciprofloxacin, OXA: Oxacillin. *Susceptible test bacteria that subcultured in LB broth without aluminum chlorohydrate for 30 days.
Table 2.
Relative fold change of resistance gene expression levels of strain 24 (S. epidermidis ATCC 12228), which developed resistance to ciprofloxacin after 30 days exposure to aluminum chlorohydrate, and control bacteria, which developed resistance to ciprofloxacin only by 30-day subculturing in LB broth without aluminum chlorohydrate.
Table 2.
Relative fold change of resistance gene expression levels of strain 24 (S. epidermidis ATCC 12228), which developed resistance to ciprofloxacin after 30 days exposure to aluminum chlorohydrate, and control bacteria, which developed resistance to ciprofloxacin only by 30-day subculturing in LB broth without aluminum chlorohydrate.
| Test bacteria | norA | norB/C | gyrA | gyrB | parE | parC |
|---|---|---|---|---|---|---|
| 24 (S. epidermidis ATCC 12228) |
3,65 | 3,65 | 4,01 | 3,68 | 3,77 | 4,05 |
Table 3.
Relative fold change of resistance gene expression levels of the control group test bacteria, which developed resistance to ciprofloxacin by subculturing only in LB broth without aluminum chlorohydrate for 30 days, and the control group test bacteria taken on the first day as the starter
Table 3.
Relative fold change of resistance gene expression levels of the control group test bacteria, which developed resistance to ciprofloxacin by subculturing only in LB broth without aluminum chlorohydrate for 30 days, and the control group test bacteria taken on the first day as the starter
| Test bacteria | norA | norB/C | gyrA | gyrB | parE | parC |
|---|---|---|---|---|---|---|
| 24 (S. epidermidis ATCC 12228) |
4,14 | 4,14 | 7,18 | 4,41 | 5,89 | 6,52 |
| 14 | 6,71 | 6,82 | 6,71 | 6,71 | 7,10 | 1,61 |
Table 4.
Relative fold change of mecA gene expression levels of the test bacteria 14 and 22, which developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate, and the control group test bacteria taken on the first day as the starter
Table 4.
Relative fold change of mecA gene expression levels of the test bacteria 14 and 22, which developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate, and the control group test bacteria taken on the first day as the starter
| Test bacteria | mecA |
|---|---|
| 22 | 1,50 |
| 14 | 5,30 |
Table 5.
Relative fold change of mecA gene expression levels of the test bacteria 14 and 22, which developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate, and the control group test bacteria that developed resistance to oxacillin by subculturing only in LB broth without aluminum chlorohydrate for 30 days
Table 5.
Relative fold change of mecA gene expression levels of the test bacteria 14 and 22, which developed resistance to oxacillin after 30 days of exposure to aluminum chlorohydrate, and the control group test bacteria that developed resistance to oxacillin by subculturing only in LB broth without aluminum chlorohydrate for 30 days
| Test bacteria | mecA |
|---|---|
| 22 | 4,42 |
| 14 | 6,59 |
Table 6.
MIC values of the test bacteria before and after exposure to the test antibiotics (µg/ml).
| Test bacteria | Test antibiotics | Pre-exposure | Post-exposure (30th day) |
|---|---|---|---|
| 1 | CIP | 0,125 | 4 |
| 2 | OXA | 2 | 16 |
| 3 | OXA | 16 | >64 |
| 4 | OXA | 16 | >64 |
| 5 | OXA | 0,25 | 32 |
| 6 | OXA | 8 | 32 |
| 7 | OXA | 0,25 | 16 |
| 8 | OXA | 0,25 | 1 |
| 11 | OXA | 1 | 16 |
| CIP | 0,25 | 1 | |
| 12 | OXA | 8 | 32 |
| 13 | CIP | 0,25 | 8 |
| 14 | OXA | 64 | >64 |
| 16 | CIP | 0,125 | 0,5 |
| 17 | OXA | 0,5 | 16 |
| 18 | OXA | 0,25 | 64 |
| CIP | 0,25 | 8 | |
| 19 | OXA | 0,25 | 1 |
| 20 | OXA | 0,25 | 1 |
| CIP | <0,03125 | 0,125 | |
| 21 | OXA | 0,25 | 64 |
| CIP | 0,125 | 4 | |
| 22 | OXA | 2 | 4 |
| CIP | 0,25 | 4 | |
| 23 | OXA | 8 | 64 |
| 24 | OXA | 0,125 | 0,5 |
| CIP | 0,125 | 0,5 |
CIP: Ciprofloxacin, OXA: Oxacillin. 1-22: Susceptible isolates, 23: Resistant isolate, 24: S. epidermidis ATCC 12228.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



