Submitted:
09 March 2023
Posted:
14 March 2023
You are already at the latest version
Abstract
A primary challenge facing conservationists is reconciling the human needs of forest adjacent communities with the needs of conserving forest biodiversity, especially in tropical regions with growing populations of rural poor. Agroforestry has the potential to simultaneously provide for human needs and enhance forest biodiversity, but the complex interactions and feedbacks between the social and natural dimensions are relatively undescribed and thus systematic implementation is rare. The attributes of trees on farms required for human needs and conservation value may conflict. For example, exotic species in monoculture may provide the most economic value for farmers, while relic or planted indigenous tree mixtures may be more valuable for biological conservation. The objective of this study was to explore whether agroforestry practices in a moist tropical forest ecosystem in Kenya can simultaneously provide timber and fuelwood value to small-holder farmers while extending forest tree biodiversity. We described the agroforestry attributes on farms around a tropical forest, assessed the relationship between number and biomass of timber/fuelwood trees and tree biodiversity, and explored the relationships between forest tree diversity attributes and farm tree diversity attributes on a landscape scale using spatial analysis. We found that the diversity and number of trees on farms in this area are extensive yet variable, but that no significant relationship exists between the number of timber/fuelwood trees and tree diversity. This suggests that the two values of agroforestry may not be in conflict, due mainly to the high diversity of trees used for fuelwood. We also found that trees on farms in the larger landscape add to the conservation value of forest tree biodiversity and could be important components in conservation management. If agroforestry is to play an increasingly active role in conserving biodiversity in human-dominated landscapes, particularly in areas of dense subsistence farmer populations, increase recognition needs to be given to farmer’s perception of the value of trees and their selection of what trees to plant or maintain.
Keywords:
Kakamega Rain Forest
; Conservation
; Biodiversity
; Complementarity
; Agroforestry
1. Introduction
One challenge in managing tropical forests is reconciling the extraction needs of the local people with conservation of intact ecosystems. Many tropical forests are in regions with high population density coupled with relative poverty giving rise to a pervasive reliance on subsistence harvest of forest products (e.g. fuel wood) [1] (Meyer and Turner 1992). This common dilemma to reconcile the human needs of forest adjacent communities with the needs of forest biodiversity conservation and preservation of ecosystem services is the primary challenge facing conservationists.
There is growing consensus that trees on farms can play an important role in conserving biodiversity, improving agricultural production, providing fuel and incomes, delivering and extending ecosystem services, and ultimately halting the loss and degradation of forests [2,3,4,5] (Schroth 2004, Jerneck and Olsson 2013, Nyambane 2014, Mbow et al. 2014). In the modern fragmented landscape, it is commonly recognized that the human matrix (e.g. farms and agroforestry) will be increasingly important for biodiversity conservation [6] (Perfecto and Vandermeer 2008), and studies in Africa have shown that successful agrofrorestry systems can contribute to biodiversity conservation of fragmented forest and woodland ecosystems while providing socio-economic benefits to farmers [7,8,9] (Sileshi et al 2007, Kalaba et. al. 2010, Jose 2012).
The FAO defines agroforestry as “land-use systems and technologies where woody perennials are deliberately used on the same land-management units as agricultural crops and/or animals, in some form of spatial arrangement or temporal sequence”. Trees on farms can provide multiple benefits to both the farmer and the ecosystem at large [9,10,11,12] (Mattison 2005, Bhagwat et al. 2008, Jose 2012, Tscharntkea et. al. 2012) but balancing optimal conditions for crops and the extractive needs of the farmer with biodiversity and ecosystem benefits is a challenge [9,13,14] (van Noordwijk et. al. 2016, Jose 2011, Jose 2012). Despite the potential of agroforestry, it remains rarely systematically implemented [13] (van Noordwijk et. al. 2016) and its contribution to livelihoods and forest biodiversity are relatively undescribed. Reasons for this stem from multiple causes, including a myriad of policy adjustments [13] (van Noordwijk et. al. 2016), a constant shift of our conceptualization of agroforestry, and the complex inter-connections and feedbacks between the social and natural dimensions inherent in its implementation. The almost ubiquitous tendency arising from this lack of clarity is to separate the human and natural dimensions; with policy dictating that agricultural development and forestry end up under different dockets, and ecologists ignoring or underestimating the ecological value of agricultural landscapes in conservation [15] (de Foresta et al. 2013). Recently, increasing recognition is given to the value of micro-reserves outside of the primary reserve network [16] (Laguna et al. 2016) giving rise to recommendations for a “top-down” approach to regional reserve planning [17] (Margules et al. 1988) that allows for incorporation of private land holdings into conservation planning. This gives a new impetus to better understand and implement agroforestry systems.
Though trees on farms can simultaneously provide economic value while conserving biodiversity [18,19,20,21](Bugayong 2003, Schroth 2004, Swihart and Moore 2004, Sodhi and Ehrlich 2010), the attributes of trees on farms required for these benefits may conflict: exotic species (e.g. Eucalyptus spp. in the African setting) may provide the most economic value while providing the least ecosystem benefits, while indigenous tree mixtures may be more valuable for biological conservation but provide the least immediate economic value to farmers. The actual composition of trees on farms will depend on a number of factors, but ultimately for agroforestry to be effective, thereby ensuring its adoption, trees in the forest and on farms must complement each other in terms of maintaining biodiversity and providing forestry resources [8,22] (Cannell et. all. 1996, Kalaba et. al. 2010), or at the very least, not be in direct conflict. This is a key challenge in building sustainable and viable agroforestry systems.
Wood fuel continues to be the primary energy source in Africa, with data showing that not only is it used in 77% of African households, but use in rural households in some African countries is as high as 90% and may be increasing [23,24,25] (Mugo and Gathui 2010, Legros et al. 2009, Bonjour et al. 2013). In the western ecoregion of Kenya, for example, there are over 500,000 households each consuming over 4,000 kilograms of wood annually, predominately from forests, but also from trees on farms [26,27] (KNBS 2015, Lung and Espira 2019). While not responsible for large scale forest loss, there is considerable evidence this subsistence wood fuel use is causing complex changes in forest diversity and structure and is leading to irreversible degradation [28,29,30,31] (Lung and Schaab 2007, Kefa 2015, Lung and Espira 2015, Kefa et al. 2017).
The objective of this study was to explore whether agroforestry practices in a moist tropical forest ecosystem can simultaneously provide adequate resources for timber and fuelwood extraction by farmers while extending forest tree biodiversity conservation. First, we described the agroforestry attributes on farms in terms of tree numbers, biomass and diversity. Second, we assessed the relationships between farm attributes (i.e. land size, crop diversity, and presence of cash crop) and agroforestry attributes (i.e. number and biomass of timber/fuelwood trees and tree biodiversity). Third, controlling for the effects of farm attributes, we assessed the relationship between number and biomass of timber/fuelwood trees and tree biodiversity. Lastly, we assessed the relationships between forest tree diversity attributes and farm tree diversity attributes on a landscape scale using spatial analyses. If agroforestry practices can simultaneously provide timber and fuelwood while conserving tree biodiversity, then we should see no changes in tree biodiversity per farm as the number and biomass of timber/fuelwood trees varies. Conversely, if these two are in contrast, then biodiversity should decrease with increasing number of biomass of timber/fuelwood trees.
2. Materials and Methods
The study was conducted on small-holder farms in communities bordering the Kakamega forest in western Kenya (34°75’ E; 0°15’ N) (Figure 1). A large majority (>90%) of the rural population are subsistent farmers with few formal employment opportunities, and with greater than 50% of the population being unemployed and living below the poverty line [26}(KNBS 2015). Most farmers grow a combination of cash crops and subsistence food crops, with the ratio varying depending on season and size of land holding. The primary cash crops are sugar cane and tea, and the primary food crop is maize usually grown with a variety of vegetables and pulses. Almost all rural households use wood for cooking [27] (Lung and Espira 2019). Many farmers maintain trees on their land for a variety of reasons, including cultural beliefs, shade, and fuelwood among others.
On-farm tree assessment was done on the farms of 50 households in the study area. The households were randomly selected from an existing database of over 15,000 households located within 3 kilometers of the forest edge. Each household farm was assessed by a trained field technician and assistant who worked with the farm owner to identify and measure all trees on the farm, to elucidate their use and origin, and to answers questions about farm size, presence of cash crops and crop diversity. All trees were identified to species level (where possible), counted, and their diameter at breast height (DBH) measured. Each tree was also categorized by potential use (e.g. fuelwood, timber, fruit, etc.) and origin (exotic or indigenous). To corroborate use, we also surveyed 20 people collecting fuelwood regarding their preferred fuelwood species. Using DBH we created 5 age-classes of trees (<10 cm, 10-20 cm, 21-30 cm, 21-40 cm, and >40 cm). DBH, along with wood density (ƥ) and an estimate of environmental stress (E) were then used in a pan-tropical allometric equation [32](Chave et al., 2014) to estimate above ground biomass (AGB) for each tree:
AGB = exp[-1.803 – 0.976 + E + 0.976 x lnƥ + 2.673 x ln(DBH) – 0.0299[ln(DBH)]2]
Wood density (ƥ) was obtained from the global wood density database [32] (Chave et al., 2009). When multiple values were available for a species, we used the average the value. Environmental stress (E) values for the regions were obtained from a gridded global layer available at http://chave.ups-tlse.fr/pan-tropical_allometry.htm [32](Chave et al., 2014). For each farm we calculated the total number of trees and total biomass for (1) all trees, (2) all indigenous trees, (3) all trees used for fuelwood/timber, and (4) all indigenous trees used for fuelwood/timber.
To provide a more comprehensive account of forest diversity on farms than any one metric could provide, we calculated four indices of species diversity (species richness, Simpson’s index, Shannon’s index, and evenness) for all tree species (including exotic species) and for just indigenous tree species per farm.
We used a modified point quarter count method to measure species richness, rarity, and diversity at 35 randomly selected 0.5-acre survey locations within the Kakamega Forest [31] (Kefa et al. 2017). At forest survey locations, the standard USDA Forest Service Point Quarter Count Method for surveying forest points was modified to include measurement of a central tree and up to the two largest trees in the plot if those trees were not included in the initial survey. This means that each site included measurements and identities for up to six total trees. On-farm surveys of diversity included a complete census of all trees, and included up to 200 measured trees. Therefore, prior to analysis we rarefied the farm biomass and tree diversity data to the median forest diversity and biomass measurements of 5.4 trees per site using the R Package iNEXT [33] (Hsieh et al. 2016). We then calculated biodiversity using the Shannon Diversity index H’, Species Richness, and RWR on the forest data and rarefied farm data. Biomass for each tree was calculated following the same allometric biomass equations for trees on farms and then summed for each site or rarefied site set of trees.
There were two primary components statistically analyzed within farms – the relationship between farm attributes and tree attributes, and the influence of timber/fuelwood tree numbers on tree biodiversity. We used a multivariate regression general linear model to explore the relationship between three farm attributes (land size, presence of a cash crop, and crop diversity) and five tree attributes (number of total species, number of indigenous species, number of all trees, number of exotic timber/fuelwood trees, and number of all timber/fuelwood trees). We also used a multivariate regression general linear model to analyze the influence of the number of timber/fuelwood species on each diversity index (richness, Simpson’s, Shannon’s, and evenness) calculated for all tree species and for only indigenous tree species. For both statistical components, all predictor variables were compared in a correlation matrix to check for autocorrelation, and all variables in resulting models were checked for multicollinearity using VIF, tolerance calculations, and eigenvalues [34](Field 2013). We used the Durbin-Watson estimation to check for independence of residuals, a case-wise diagnosis to look for bias due to extreme values, residual plots and partial plots to assess homogeneity of variance. All statistical analyses were conducted in SPSS. Unless otherwise stated, all values are reported as mean (+/- standard error).
Complementarity and HotSpot Analyses. We used a complementarity analysis based on rarity weighted richness scores to identify priority conservation areas within our set of forest and farmed sites [35,36,37] (Kirkpatrick 1983, Margules et al. 1988, Albuqurque and Gregory 2017). Complementarity analyses use an iterative process to identify priority sites for conservation in an effort to save all species in the system, or as many species as is possible, in as small an area as possible. For our complementarity analysis we ordered surveyed farm or forest sites in descending order by their RWR scores. We then chose sites with the highest RWR scores until 90%, 95%, and 100% of all tree species identified in the system were included in the set of chosen sites. [38] Albuquerque and Beier (2015) have shown that this application of RWR scores to choose reserve designs is highly effective and efficient at identifying priority areas for conserving species, and [37] Albuquerque and Gregory (2017) have further shown that this method can be applied to large geographic areas and is effective at evaluating gaps in species coverage by existing reserve networks.
We used ArcGIS 10.4 minimum bounding geometry to identify the spatial area encompassed by the sets identified as conserving 90%, 95% and 100% of the species identified in our field surveys. We then calculated the area encompassed by the minimum bounding geometry of each region, and the number of forest and farm sites included in each area. Some farm and forest sites included within the spatial area encompassed by the minimum bounded areas were of lower complementarity (RWR) rank and therefore not used to identify the minimum conservation extent needed to reach species conservation coverage of 90%, 95%, 100% species representation. These areas represent potential redundancy in species coverage.
We used ArcGIS 10.4 to conduct a Gi* HotSpot analysis on the species richness data and the biomass data. Hotspot analysis identifies areas of significantly clustered high × high spatial autocorrelation (HotSpots) and areas of significantly clustered low x low spatial autocorrelation [39] (ColdSpots; Ord and Getis 1995). We performed the Hotspot analysis independently on the farm and forest species richness data sets, and on a pooled dataset that included both the farm and forest diversity data in a single HotSpot analysis. Similarly, we performed HotSpot analyses on the forest and farm biomass datasets and a combined forest and farm dataset. We retained all sites that were clustered at >90% confidence as Hotspots or ColdSpots. We used a Geographically Weighted Regression to test for significant correlations among species richness and biomass, and rarity (RWR) and biomass. For GWR we used an adaptive kernel and search radius.
3. Results
3.1. Tree Attributes Per Farm
We collected data on 50 farms with a mean size of 0.83 (+/- 0.17) and a range from 0.1 to 20.5 acres. All farms were within 3 kilometers of the forest boundary and the total area sampled was 104.35 acres. We found an average of 89.88 (+/- 12.44) trees per farm, ranging from 10 to 385 trees per farm. Trees were located along boundaries, interspersed in crops, located around the home, and in designated woodlots. Forty (40%) percent of farms had a woodlot. We observed an average of 2.22 age classes (range: 1 to 5), with 90.56% of trees on farms in the two smallest age-classes and 4% in age classes over 30 cm DBH. Total tree biomass per farm averaged 10.78 (+/- 2.51) tons and ranged from 0.15 to 108.57 tons. This equates to mean carbon stocks of 5.39 (+/- 0.97) MgC per farm ranging from 0.19 to 136.08 MgC (using carbon fraction of 0.50). The average number of tree species observed on farms was 8.40 (+/- 0.59) and ranged from 2 to 20 species of which 4.20 (+/- 0.44) species, or 50%, were indigenous forest trees (Table 1). Fifty-two (52%) percent of farms had at least one large tree (>30 cm DBH) and 34% had more than one large tree. Thirty percent (30%) of farms had at least one large (>30 cm DBH) indigenous tree. Twenty-eight percent (28%) of farms had multiple age-classes (including at least one large tree) of indigenous trees. The highest number of large trees on a farm was 10 trees with a DBH>30 cm.
In the survey of fuelwood users, of the ten most commonly mentioned fuelwood tree species (Table 2), seven are shared with the ten most abundant trees on farms. All farms had at least one timber or fuelwood tree with Eucalyptus spp. (saligna / grandis) and Cuppressus lusitanica accounting for over 90% of both number and biomass of timber/fuelwood trees. The number and biomass of exotic timber and fuelwood trees averaged 51.04 (+/- 8.29) trees and 5.72 (+/- 1.21) tons per farm respectively which represented just over 50% of total tree number and biomass. The number and biomass of all timber/fuelwood trees averaged 69.28 (+/- 10.47) trees and 7.18 (+/- 1.35) tons per farm, representing three quarters (75%) of total tree number and biomass.
3.2. Tree Attributes of Farm Landscape
We counted 4,494 trees of 64 species (42 indigenous) accounting for 539 tons of biomass within 104.5 acres of farmland surrounding Kakamega forest. This equates to 43.63 trees and 5.23 tons of tree biomass per acre across the farm landscape. In terms of carbon stocks, this equates to mean carbon stocks of 14.375 (+/- 2.4) MgC per acre. In terms of exotic timber/fuelwood trees and all timber/fuelwood trees, biomass per acre across the farm landscape equaled 2.78 and 3.48 tons. These attributes are, of course, delineated by farms and if number of trees and biomass are calculated by farm area then we find 74.75 (+/- 12.08) trees and 4.33 (+/- 0.77) tons of tree biomass per acre. In terms of exotic timber/fuelwood trees and all timber/fuelwood trees, average biomass per acre was 4.02 (0.72) and 5.09 (0.75) tons.
3.3. Tree Assemblages on the Farm Landscape
Just over 73% of all trees on farms were exotic species, with one species, Eucalyptus spp. (saligna /grandis), comprising 46% of all trees counted. In terms of individual trees, timber trees dominated the farm landscape, forming 57% of all individuals counted. Naturally occurring trees (that are unlikely to have been planted) comprised 18% of all trees, fruit trees 12%, culturally important trees 7%, and decorative and medicinal trees 3% each. In terms of diversity, naturally occurring trees comprised 33% of the total species (21) while timber species comprised 28% of the total species (18 species). Fruit trees were the next most diverse, with 11 species (17%), and decorative trees with 9 species (14%). Culturally important trees consistent of 3 species (5%) and medicinal trees 2 species (3%). In terms of biomass, the dominance of exotic species is even more pronounced, with 75% of all biomass on farms being from exotic trees, with Eucalyptus spp. (saligna / grandis) alone accounting for 60% of all biomass measured. However, of the 10 largest trees recorded on farms, only two were exotic species, and the rest were culturally important or relic forest trees.
We also evaluated the relationships between three farm attributes (farm size, crop diversity, and presence of cash crop) and five tree attributes (total number of species, number of indigenous species, total number of trees, number of exotic timber/fuelwood trees, number of all timber/fuelwood trees; Table 3). As expected, we found that Land size was a significant predictor of all tree diversity attributes, explaining ~40% of the variation in tree numbers and 16% of the variation in overall tree diversity.
We similarly found no relationship between the number of timber/fuelwood trees and diversity measures, unless the diversity index also accounted for dominance (e.g. Simpson’s index and evenness), suggesting that dominance of some exotic species may drive overall farm tree diversity (Table 4).
3.4. Complementarity and Hot Spot Analyses
In forest plots, we recorded 58 species of trees accounting for 26.64 (+/- 4.50) tons biomass (range 0 - 46.36 tons) per acre. Of the 58 forest tree species, 6 were unique to the forest, in contrast to the 64 farm tree species of which 11 were unique to farms. Prior to analysis we rarefied the farm biomass and farm tree diversity data to the median forest diversity and biomass measurements of 5.4 trees per site using the R Package iNEXT [33](Hsieh et al. 2016). The rarefied farm data set included 37 total tree species and an average of 4.2 rarefied species per farm and 7 unique farm trees. This suggests that the rarefied farm data set represents an approximate 34% reduction in on-farm tree diversity from the full data set. However, given the different collection methods used in the forest and on farms, rarefaction is necessary for statistically valid comparisons. We calculated RWR, and diversity from the rarefied data sets. In total, farms and forests together had a total RWR score of 80.0997 with an average RWR per site = 0.953 (+/- 0.932) and range = 0 - 5.099, farms alone had a total rarefied RWR of 30.88, with an average RWR per farm site = 0.89 (+/- 0.52) and range = 0 - 2.01, and forest sites alone had a total RWR of 50.198 with an average RWR per site = 1.03 (+/- 1.13) and range = 0.021 - 5.099.
All complimentary analysis results included contributions from farmed sites outside of the gazetted boundary of the Kakamega Forest. The 90% species richness complimentary reserve design included 19 total surveyed sites (12 farm and 7 forest), encompassed a total area of 5,791.7 ha (Figure 2), 33.23% of the maximum RWR (Range in per site RWR 1.1-5.099), and a total estimated biomass of 393,635.54kg. In total this design conserved 62 (90%) of total tree species. The 95% complimentary conservation area includes contributions from 22 farms and 22 forest locations. It encompasses a total 21,791.51ha area and accounts for 64.28 (81.24%) of the total observed RWR score and 4,451,319.6kg of biomass. It conserves 66 (96%) of the total tree species observed across forest and farm surveys (Figure 2). The all- inclusive complimentary design included contributions from 50 farm sites and 33 forest sites. It encompasses 96,966.25 acres of forest and adjacent farm lands and combined RWR score of 75.121 (93.67% of total RWR). This region also has a combined biomass of 4,594,105.7 kg and conserves 100% of the total tree species observed (Figure 2). These complimentary reserve regions represent from 0.25-0.86x the size of the current Kakamega Forest Reserve area.
We found no biomass Hotspots or Coldspots suggesting that high biomass farms and forest sites are more-or-less randomly distributed across the landscape. When we conducted a Hotspot analysis on a pooled species richness dataset that included both farms and reserve sites, we detected no Hotspots or Coldspots. However, when we conducted Hotspot analyses on the individual farm and forest species richness data sets we did detect significant species richness Hotspots and Coldspots within the forest and significant species richness Hotspots among the farm data sets (Figure 2). Quixotically, the farm species richness Hotspots and the forest species richness Hotspots were found on the opposite sides of the forest from one another, such that high diversity farms were found adjacent to lower diversity forest sites and high diversity forest sites were found adjacent to lower diversity farm sites (Figure 2). GWR analysis, detected no spatial correlation between biomass and species richness (r2 = 0.00023; P<0.14), nor did we detect any relationship between biomass and RWR (r2 = -0.013; P<0.50).
4. Discussion
Our results suggest that the tradeoffs between agroforestry and conservation may not be as pronounced as casual observations may suggest. On farms surrounding Kakamega Forest, we found a considerable abundance of trees in terms of numbers, biomass, and diversity, although the variability was large. This is consistent with prior research which documented agroforestry interventions in the region in the 1990’s in response to concerns about forest timber/fuelwood supply in relation to demand [40,41,42,43](Tengnas 1994, Scherr 1995, Paterson 1998, Mercer 2004) in the context of a very dense rural population [26](>500 persons/km2 – KNBS 2015) almost entirely dependent on forest-sourced fuelwood as their primary fuel [27](Lung and Espira 2019). Three descriptive observations from our study provide insight regarding the potential tradeoff between the wood value to farmers and the conservation value to society. First, timber/fuelwood tree species accounted for 90% of tree numbers and biomass. Second, while two exotic timber/fuelwood tree species (Eucalyptus and Cuppressus) dominate numbers and biomass on farms, there are at least five indigenous tree species used for fuelwood commonly found on farms (Croton macrostachyus, Markhamia lutea, Harungana madagascariensis, Maesopsis eminii, Strychnos usambarensis). Third, tree richness averaged 8 species per farm (50% indigenous) and about one third of farms had multiple age-classes of indigenous trees, usually including the largest tree per farm. As such, despite the reliance on fuelwood and the trend towards using trees on farms for fuelwood supply, the presence of large relic indigenous trees on farms and the use of a diversity of trees for timber/fuelwood suggest the possibility that wood value and conservation value are not at complete odds. This is further supported by the observation that indigenous tree diversity on farms (averaging 4 species but as high as 12 species per farm) can be as high as average tree diversity in forest plots [30](7.4 species per 0.31-acre plot: Lung and Espira 2015).
We predicted that if timber/fuelwood and conservation values were at odds, then we would see a negative relationship between the number of timber/fuelwood trees on a farm and tree diversity variables. In other words, planting trees for timber/fuelwood would be at the expense of tree diversity. For most diversity variables, we detected no significant relationship between the number of timber/fuelwood trees and overall tree diversity. Specifically, Shannon diversity index and tree species richness index did not covary with number of timber/fuelwood trees. This effect was present regardless of if we included exotic species or not. Based on calculated coefficients and effect sizes it is unlikely that we failed to detect differences. For example, we detected no significant relationship between the number of all timber/fuelwood trees and the number of indigenous species per farm and the coefficient and effect size for this relationship was 0.009 and 0.030, respectively. These findings, which suggest that wood and conservation values are not always at odds, may be explained by the high diversity of trees used for timber/fuelwood. [31]Kefa et al. (2017) found 47 tree species (34 indigenous) in fuelwood head bundles in the same study region, and of the ten most common trees found on farms, seven were the most commonly mentioned fuelwood species by surveyed wood gatherers (Table 2).
Lastly, we found that on a landscape level, trees on farms added to the biodiversity of natural forests and could be important components in conservation management. The total diversity of trees on farms (64 total species, 42 indigenous) was similar to the tree diversity in adjacent forest. In the 33 plots used in this study, we recorded 58 tree species. [30]Lung and Espira (2015) recorded 65 tree species from 95 plots in adjacent forest. However, this has to be taken in the context that a significant portion of tree diversity on farm land was accounted for by exotic species. Of the 65 species encountered on farms, 25 were exotic (38%) and 6 (9%) were indigenous non-forest trees, and only 34 species of forest trees were found on farms. This is significantly fewer than the 65 species recorded in the forest [30](Lung and Espira 2015). This raises the question whether on-farm tree diversity, consisting of a large assemblage of exotic and non-forest trees, is able to provide the same biodiversity and ecosystem services as indigenous forest.
Our spatial analysis provides some interesting insights. First, the geographically weighted regression results reinforce the notion of no tradeoff in terms of biodiversity and biomass, even taking spatial heterogeneity into account. It also suggests there is no spatial heterogeneity in biomass. We also found no Hotspots when we pooled the farm and forest data sets into one dataset but found offsetting hotspots when the farm and forest data were analyzed separately. In other words, in forest regions that contained Hotspots we detected no corresponding Hotspots in adjacent farm regions, while in forest regions that contained no Hotspots, we detected farm Hotspots in adjacent farm regions. This observation suggests that spatial autocorrelation in one data set offsets the spatial autocorrelation in the other data set. While it is an anecdotal observation it could be explained through the lens of market economics. We know that there is a very high diversity of trees used for fuelwood and timber [29,31] (Kefa 2015, Kefa et al. 2017). In areas where the forest lacks high diversity, we found farms with high diversity of trees. This could be explained as a response by farmers to grow a diversity of trees in response to a low supply by forests. However, this could also be explained by farmers merely conserving high diversity forest remnants. Regardless it further highlights the conservation value of farms in maintaining regional tree species richness.
The complementarity analysis with RWR also allowed us to identify the minimum spatial area necessary to conserve landscape species richness. It is interesting that even when conserving only 90% of the total species richness farm sites were required. This highlights that farms and areas outside of the forest are important components in conserving regional biodiversity. This supports a growing body of literature about the value of micro reserves and areas outside of reserve areas in conservation [44,45] (Laguna et al. 2016, Arenas et al. 2017).
One of the challenges of agroforestry is getting farmers to both plant and maintain trees on their actively used farmland. Though the theory of agroforestry suggests that farmers should seek complementarity of resource capture by trees and crops [46](Cannell et. all. 1996) and provision of forestry resources [8](Kalaba et. al. 2010), a number of our findings suggest that farmers are not actively selecting or maintaining tree biodiversity on their farms, but more likely passively letting biodiversity remain on their farmland. First, the majority of indigenous trees on farms are likely to be relic species or self-dispersed species that are not planted by the farmer. Other than Markhamia lutea, and Zanthoxylum gilletii, species that are of cultural significant (Espira, personal observation), the opportunistic species Croton macrostachyus, Harungana madagascariensis and Bridelia micrantha, that dominate forest regeneration regions, are the most common indigenous trees on farm land. In addition, over 73% of all trees counted were exotic species, with Eucalyptus saligna alone comprising 46% of all trees on farms. This suggests that farmers are making their choices on which trees to plant based primarily on their economic value (Eucalyptus being the quintessential example of such a species) and on cultural considerations rather through an active choice to maintain diversity or to provide agroforestry benefits (e.g. fruit, nitrogen fixing, etc.). Eucalyptus is both fast growing and easily converted to revenue at multiple stages of its growth phase (from firewood to poles to timber). The fact that the largest trees on farms are indigenous forest species suggests that relic trees (left over from forest clearing) also play a vital role in contributing to farmland diversity, and may form the nexus around which pioneer species such as Croton macrostachyus, Harungana madagascariensis and Bridelia micrantha may seed themselves. These results suggest that farmers may be willing to devote a substantial area of their farmland to trees as long as they see a specific benefit of these trees, with cultural and economic benefits being good examples.
If agroforestry is to play an increasingly active role in conserving biodiversity in human-dominated landscapes, particular in areas of dense subsistence farmer populations, increase recognition needs to be given to farmer’s perception of the value of trees and their selection of what trees to plant or maintain. In this study, we found that farmers choose to include trees within their farm landscape even though there is currently no active agroforestry initiative in the area. This suggests that a drive to increase on-farm biodiversity should target species that farmers already prefer or species that can serve the same purpose that farmers perceive their on-farm trees serving. Furthermore, as much as exotics like Eucalyptus spp. are not seen as being ideal for agroforestry, an insistence on their exclusion from any drive for increased trees on farmers is likely to alienate farmers.
Although agroforestry initiatives have historically targeted resource use complementarity (e.g. shade, water use), alternate farm uses (e.g. fodder, nitrogen fixing), biodiversity benefits (e.g. pollination), and food production (e.g. fruit, berries), a further consideration, namely farmer perception, should be given a high priority as well. For this to be successful, agroforestry needs to be approached from an increasingly multi-disciplinary approach, giving increasing weight to economic and social factors. In the meantime, ecologists should continue to study how much biodiversity can benefit from various agroforestry initiatives, what roles farmland can play in conservation, and just how much of ecosystem function can be maintain by heavily altered and fragmented, but nevertheless biologically rich, landscapes.
Funding
This research was completely supported by the authors and has no funding source to declare.
Acknowledgments
We thank the Kenya National Commission for Science and Technology, Kenya Forest Service and Kenya Wildlife Service for granting us permits to undertake this study and the County government of Kakamega County for approving our research permit. .
Conflicts of Interest
The authors declare no conflict of interest exists.
References
- Meyer, W.B.; Turner, B.L Human population growth and global land-use/cover change. Annual review of ecology and systematics. 1992, 23(1):39-61.
- Schroth, G.; da Fonseca, G,A.; Harvey, C.A.; Vasconcelos, H.L.; Gascon, C.; Izac, A.M. editors. Agroforestry and biodiversity conservation in tropical landscapes. Island Press; Washington, DC, USA; 2004.
- Jerneck, A,; Olsson, L. More than trees! Understanding the agroforestry adoption gap in subsistence agriculture: Insights from narrative walks in Kenya. Journal of Rural Studies. 2013, 32:114-25.
- Nyambane, L. K. Influence of farm enterprise diversification on socio-economic status among sugarcane farmers in Bumula sub-county Bungoma county, Kenya. Doctoral dissertation, University of Nairobi, Kenya, 2014.
- Mbow, C.; Van Noordwijk, M.; Luedeling, E.; Neufeldt, H.; Minang, P.A.; Kowero, G. Agroforestry solutions to address food security and climate change challenges in Africa. Current Opinion in Environmental Sustainability, 2014, 6:61-7.
- Perfecto, I.; Vandermeer, J. Biodiversity conservation in tropical agroecosystems: a new conservation paradigm. Annals of the New York Academy of Sciences, 2008, 1134(1), 173-200.
- Sileshi, G.; Akinnifesi, F. K.; Ajayi, O. C.; Chakeredza, S.; Kaonga, M.; & Matakala, P. W. Contributions of agroforestry to ecosystem services in the Miombo eco-region of eastern and southern Africa. African journal of environmental science and technology, 2007,1(4), 68-80.
- Kalaba, K. F.; Chirwa, P.; Syampungani, S.; & Ajayi, C. O. Contribution of agroforestry to biodiversity and livelihoods improvement in rural communities of Southern African regions. Tropical rainforests and agroforests under global change: ecological and socio-economic valuations, 2010, 461-476.
- Jose, S. Agroforestry for conserving and enhancing biodiversity. Agroforestry Systems, 2012. 85, 1-8.
- Mattison, E. H.; & Norris, K. Bridging the gaps between agricultural policy, land-use and biodiversity. Trends in ecology & evolution, 2005, 20(11), 610-616.
- Bhagwat, S. A.; Willis, K. J.; Birks, H. J. B.; & Whittaker, R. J. Agroforestry: a refuge for tropical biodiversity? Trends in ecology & evolution, 2008, 23(5), 261-267.
- Tscharntke, T.; Clough, Y.; Wanger, T. C.; Jackson, L.; Motzke, I.; Perfecto, I.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biological Conservation, 2012, 151(1), 53-59.
- van Noordwijk, M.; Kim, Y. S.; Leimona, B.; Hairiah, K.; & Fisher, L. A. A. Metrics of water security, adaptive capacity, and agroforestry in Indonesia. Current Opinion in Environmental Sustainability, 2016, 21, 1–8. [Google Scholar] [CrossRef]
- Jose, S. Managing native and non-native plants in agroforestry systems. Agroforestry systems, 2011, 83, 101-105.
- De Foresta, H.; Somarriba, E.; Temu, A.; Boulanger, D.; Feuilly, H.; Gauthier, M.; & Taylor, D. Towards the assessment of trees outside forests: a thematic report prepared in the Framework of the Global Forest Resources Assessment 2010. Forest Resources Assessment Working Paper (FAO) (2013).
- Laguna, E.; Fos, S.; Jiménez, J.; & Volis, S. Role of micro-reserves in conservation of endemic, rare and endangered plants of the Valencian region (Eastern Spain). Israel Journal of Plant Sciences, 2016, 63(4), 320-332.
- Margules, C. R.; Nicholls, A. O.; & Pressey, R. L. Selecting networks of reserves to maximise biological diversity. Biological conservation, 1988, 43(1), 63-76.
- Bugayong, L. A. Socioeconomic and environmental benefits of agroforestry practices in a community-based forest management site in the Philippines. In International Conference on Rural Livelihoods, Forests and Biodiversity (pp. 19-23). May 2003.
- Schroth, G.; Harvey, C.A.; G. Vincent, G. Complex agroforests: Their structure, diversity and potential role in landscape conservation. 2004b, Chapter 10, Pp. 227–260 in: Schroth, G., G.A.B. da Fonseca, C.A. Harvey, C. Gascon, H.L. Vasconcelos and A.M.N. Izac (eds) Agroforestry and Biodiversity Conservation in Tropical Landscapes. Island Press, Washington, DC, USA.
- Swihart, R. K.; & Moore, J. E. Conserving biodiversity in agricultural landscapes: model-based planning tools. Purdue University Press. 2004.
- Sodhi, N. S.: & Ehrlich, P. R. (Eds.). Conservation biology for all. Oxford University Press, 2010.
- Cannell, M. G. R.; Van Noordwijk, M.; & Ong, C. K. The central agroforestry hypothesis: the trees must acquire resources that the crop would not otherwise acquire. Agroforestry systems, 1996, 34, 27-31.
- Mugo, F.; & Gathui, T. Biomass energy use in Kenya. A background paper prepared for the International Institute for Environment and Development (IIED) for an international ESPA workshop on biomass energy, 2010, 19-21 October 2010, Parliament House Hotel, Edinburgh. Practical Action, Nairobi, Kenya.
- Legros, G.; Havet, I.; Bruce, N.; & Bonjour, S. The Energy Access Situation in Developing Countries: A Review Focusing on the Least Developed Countries and Sub-Saharan Africa, United Nations Development Programme and World Health Organization, New York, 2009.
- Bonjour, S.; Adair-Rohani, H.; Wolf, J.; Bruce, N. G.; Mehta, S.; Prüss-Ustün, A., ... & Smith, K. R. Solid fuel use for household cooking: country and regional estimates for 1980–2010. Environmental health perspectives, 2013, 121(7), 784-790.
- KNBS (Kenya National Bureau of Statistics). 2015/16 Kenya Integrated Household Budget Survey (KIHBS) – Measuring Wellbeing for Sustainable Development Status Report, 2015.
- Lung, M. A.; & Espira, A. A large-scale, village-level test of wood consumption patterns in a modified traditional cook stove in Kenya. Energy for Sustainable Development, 2019, 49, 11-20.
- Lung, T.; & Schaab, G. Assessing Fragmentation and Disturbance of West Kenyan Rainforests by Means of Remotely Sensed Time Series Data and Landscape Metrics. African Journal of Ecology 2007, 44: 1–16.
- Kefa, C.A. Assessing the impacts of bioenergy extraction and human land use of the biodiversity of Kakamega Tropical Rainforest, Kenya. MSc. Thesis, Bowling Green State University, OH, USA, 2016.
- Lung, M.; Espira, A. The Influence of Stand Variables and Human Use on Biomass and Carbon Stocks of a Transitional African Forest: Implications for Forest Carbon Projects. Forest Ecology and Management, 2015, 351: 36–46.
- Kefa, C. A.; Lung, M.; Espira, A.; Gregory, A. Quantifying the Rate of Subsistence Wood Harvesting from a Tropical Rainforest in Kenya. Oryx, 2017, 5: 1–5.
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M. S.; Delitti, W. B.; ... & Vieilledent, G. Improved allometric models to estimate the aboveground biomass of tropical trees. Global change biology, 2014, 20(10), 3177-3190.
- Hsieh, T. C.; Ma, K. H.; & Chao, A. iNEXT: an R package for rarefaction and extrapolation of species diversity (H ill numbers). Methods in Ecology and Evolution, 2016, 7(12), 1451-145.
- Field, A. Discovering statistics using IBM SPSS statistics. sage. 2013.
- Kirkpatrick, J. B. An iterative method for establishing priorities for the selection of nature reserves: an example from Tasmania. Biological Conservation, 1983, 25(2), 127-134.
- Margules, C. R.; Nicholls, A. O.; Pressey, R. L. Selecting networks of reserves to maximise biological diversity. Biological conservation, 1988, 43(1), 63-76.
- Albuquerque, F. S. D.; Gregory, A. The geography of hotspots of rarity-weighted richness of birds and their coverage by Natura 2000. PLoS One, 2017, 12(4), e0174179.
- Albuquerque, F.; Beier, P. Rarity-weighted richness: a simple and reliable alternative to integer programming and heuristic algorithms for minimum set and maximum coverage problems in conservation planning. PloS one, 2015, 10(3), e0119905.
- Ord, J. K.; Getis, A. Local spatial autocorrelation statistics: distributional issues and an application. Geographical analysis, 1995, 27(4), 286-306.
- Tengnäs, B. ; Agroforestry extension manual for Kenya. World Agroforestry Centre, 1994.
- Scherr, S. J. Economic factors in farmer adoption of agroforestry: patterns observed in Western Kenya. World Development, 1995, 23(5), 787-804.
- Paterson, R. T.; Karanja, G. M.; Nyaata, O. Z.; Kariuki, I. W.; Roothaert, R. L. A review of tree fodder production and utilization within smallholder agroforestry systems in Kenya. Agroforestry systems, 1998, 41, 181-199.
- Mercer, D. E. Adoption of agroforestry innovations in the tropics: a review. Agroforestry systems, 2004, 61, 311-328.
- Laguna, E.; Fos, S.; Jiménez, J.; Volis, S. Role of micro-reserves in conservation of endemic, rare and endangered plants of the Valencian region (Eastern Spain). Israel Journal of Plant Sciences, 2016, 63(4), 320-332.
- Arenas, S.; Cortés A.J.; Mastretta-Yanes, A.; Jaramillo-Correa, J.P. Evaluating the accuracy of genomic prediction for the management and conservation of relictual natural tree populations. Tree Genetics & Genomes, 2021, 17:1-19.
- Cannell, M. G. R.; Noordwijk, M.; Ong, C. K. The central agroforestry hypothesis: the trees must acquire resources that the crop would not otherwise acquire. Agroforestry systems, 1996, 34, 27-31.
Figure 1.
Kakamega Forest Study Area. fragments in dark grey. Study Area. Kakamega Forest fragments in dark grey. Gazetted forest in light grey. Roads shown as solid lines. Households in study as black dots. Inset is Kenya with location of Kakamega Forest.
Figure 1.
Kakamega Forest Study Area. fragments in dark grey. Study Area. Kakamega Forest fragments in dark grey. Gazetted forest in light grey. Roads shown as solid lines. Households in study as black dots. Inset is Kenya with location of Kakamega Forest.
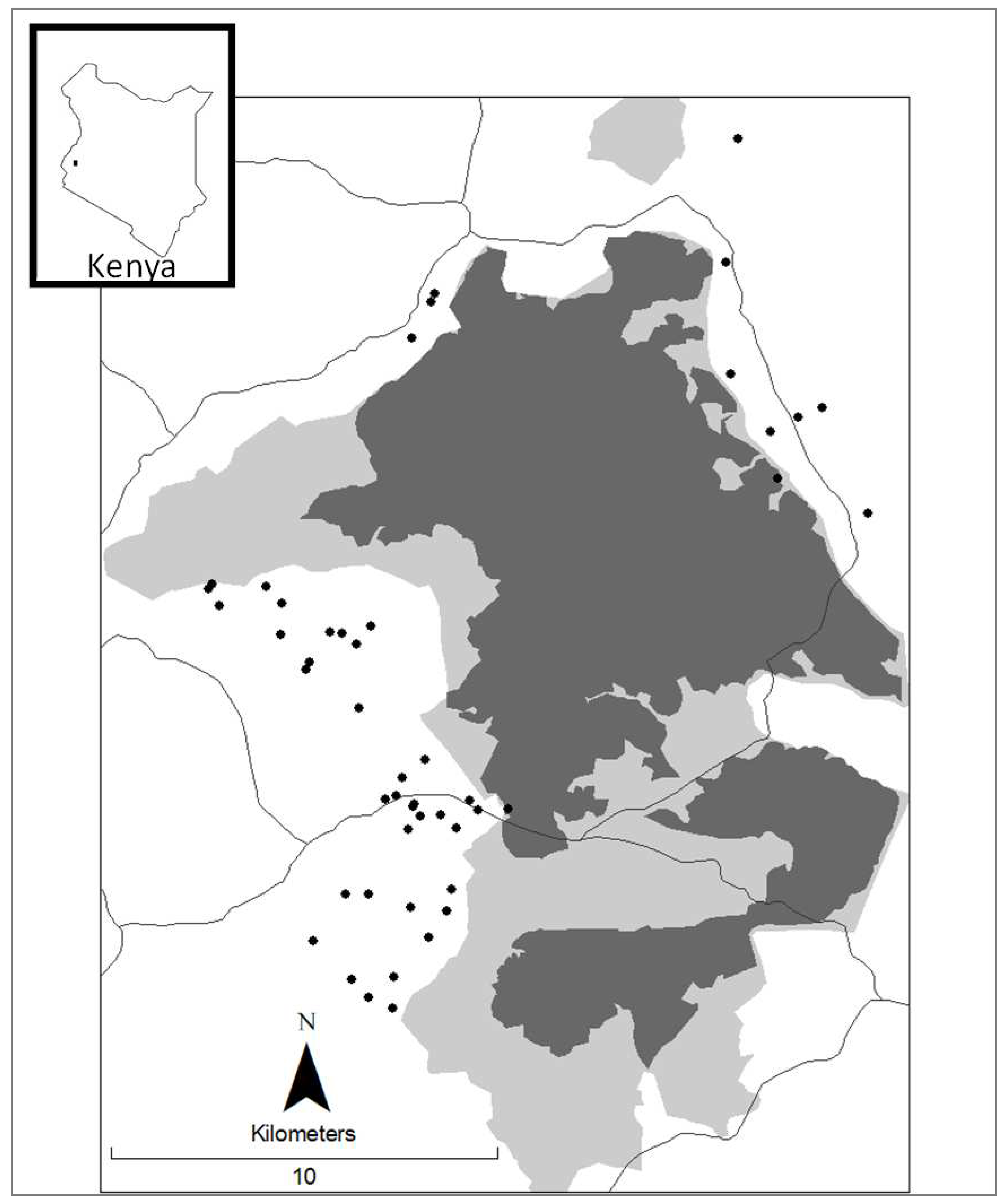
Figure 2.
Map of complimentary reserve boundaries.
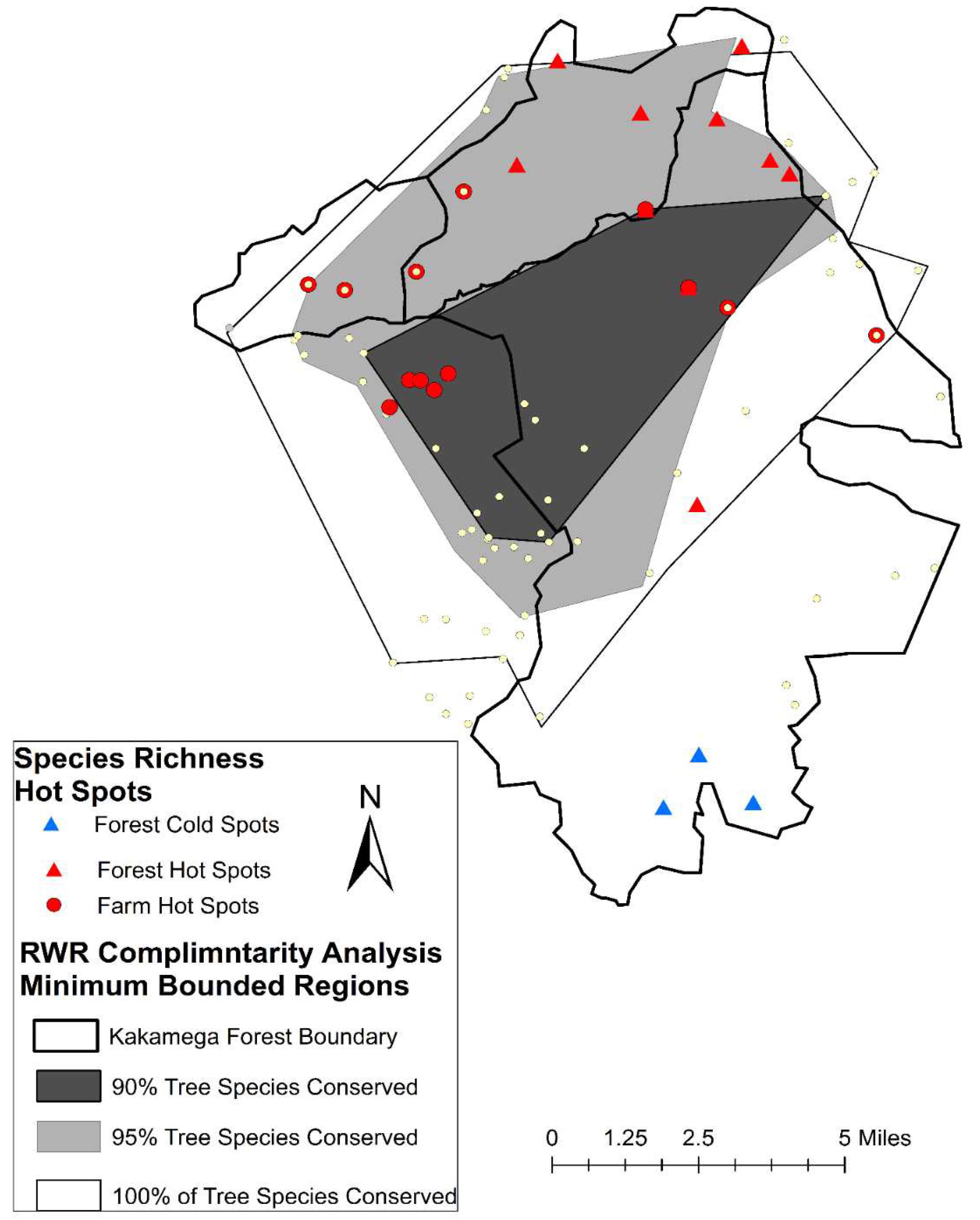

Table 1.
Number and size of trees observed on agroforestry farms.
| Landscape | Per Farm | ||||
|---|---|---|---|---|---|
| # Species | # Species | Simpson’s | Shannon’s | Evenness | |
| All trees | 64 | 8.4 ± 0.59 | 0.35 ± 0.02 | 1.41 ± 0.07 | 0.70 ± 0.02 |
| Indigenous trees | 42 | 4.2 ± 0.44 | 0.46 ± 0.04 | 0.94 ± 0.09 | 0.67 ± 0.04 |
Table 2.
The Most Common Tree Species on Farms and Most Common Fuelwood Species.
| Ten Most Common Species on Farms | % Total Trees | Ten Most Commonly Mentioned Fuelwood Species |
|---|---|---|
| Eucalyptus (grandis / saligna)^ | .464 | Eucalyptus (grandis / saligna) |
| Grevillea robusta | .086 | Croton macrostachyus |
| iCroton macrostachyus | .071 | Strychnos usambarensis |
| iMarkhamia lutea | .062 | Markhamia lutea |
| Cupressus lusitanica | .060 | Cupressus lusitanica |
| iHarungana madagascariensis | .032 | Psidium guajava |
| Pinus patula | .028 | Bridelia micrantha |
| Persea americana | .024 | Persea americana |
| iMaesopsis eminii | .020 | Grevillea robusta |
| iBridelia micrantha | .018 | Diospyros abyssinica |
i = indigenous tree species.
Table 3.
Summary of Multivariate Regression GLM among Farm and Tree Attributes.
| # Species | # Ind. Species | # Exotic Trees | # All Trees | |
|---|---|---|---|---|
| Farm Size | ** (r2=0.16) |
* (r2=0.08) |
*** (r2=0.33) |
*** (r2=0.37) |
| Crop Diversity | * (r2=0.09) |
* (r2=0.08) |
ns | ns |
| Cash Crop | ns |
ns |
ns | ns |
ns = not significant at p<0.05; *= 0.05<p>0.01; ** = 0.01<p>0.001; *** = p<.001.
Table 4.
Summary of Multivariate Regression of just exotic and all timber/fuelwood trees on the residuals of diversity measures from farm size regression.
Table 4.
Summary of Multivariate Regression of just exotic and all timber/fuelwood trees on the residuals of diversity measures from farm size regression.
| Richness | Simpson | Shannon | Evenness | |||||
|---|---|---|---|---|---|---|---|---|
| Indig. | All Trees | Indig. | All Trees | Indig. | All Trees | Indig. | All Trees | |
| # Exotic Trees | ns 0.011 0.050 |
ns 0.010 0.023 |
ns 0.000 0.014 |
* 0.001 0.103 |
ns 0.001 0.022 |
ns -0.001 0.051 |
ns 0.000 0.004 |
** -0.001 0.125 |
| # All Trees | ns 0.009 0.030 |
ns 0.010 0.058 |
ns 0.000 0.017 |
ns 0.000 0.050 |
ns 0.001 0.037 |
ns -0.001 0.017 |
ns 0.000 0.006 |
* -0.001 0.077 |
ns = not significant at p<0.05; * = 0.05<p>0.01; ** = 0.01<p>0.001; First number = βcoefficient from regression; Second number is partial beta squared.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



