
Submitted:
14 March 2023
Posted:
14 March 2023
You are already at the latest version
Abstract
Rice (Oryza sativa L.) is a primary energy food for Asian population. One of the most constraints in rice production is soil salinity because of the rice is very susceptible to salt. Meanwhile, many agricultural land in Asia are in the saline area. It is important to identify and develop salt-tolerant rice varieties that can adapt in Asian climate. By combining morphological, physiological, and biochemical assessments for screening the salt tolerant of 116 Asian rice cultivars was able to classify into tolerant, moderate, and sensitive rice cultivars under salinity stress condition, and also for understanding the salt tolerance mechanisms. The rice cultivars which belong to salt-tolerant including Pokkali from India, TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea. Whereas, salt-sensitive rice varieties like IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea. The salt-tolerant varieties showed signs of tolerance, including lower percent reduction of percentage germination, root length, root fresh weight, shoot length, plant biomass, and chlorophyll content. In order to maintain the cellular osmotic balance under saline condition, the salt-tolerant ones exhibited less membrane damage, lower Na/K ratio, high proline and sugar accumulation, lower malondialdehyde (MDA) and hydrogen peroxide (H2O2). Pokkali from India, TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea are recommended as valuable germplasm resource for Asian rice breeding program in saline agricultural area.
Keywords:
Asian rice
; salt stress
; morphology
; physiology
; biochemistry
1. Introduction
Rice (Oryza sativa L.) is one of the major food source for more than half of the global population. A 100 g of rice contains 78.2 g of carbohydrates, 6.8 g of protein, and provides 345.0 kcal [1]. Most of the global rice is produced and consumed in Asia [2,3,4]. In Asia, small farmers produced almost 90% of the rice which become their source of income [5]. One of the most constraints in rice production is soil salinity because of the rice is very susceptible to salt, especially at the seedling, early vegetative (3 leaf stage), and reproductive stages [6,7,8,9,10,11]. Based on the history, Mesopotamian civilization (now part of Iraq) collapsed due to the salinization in agricultural areas that cause crop failures [12]. Because of the human activities and natural phenomena, soil salinity is increasing. Soil salinity stress generally occur in the rice field area that have improper irrigation and drainage systems, and also in the coastal areas [13,14]. The effects of soil salinity stress also depend on the rice genotypes and the salt concentration. Indica rice is more salt tolerance compared with japonica rice subspecies [15]. Soil salinity is a serious problem in most of the rice-growing areas of Asian countries in the tropics and temperate regions. Most of the agricultural areas in the Asian regions belong to the saline soils [16,17]. Rice belongs to the most susceptible crop to soil salinity stress compared with barley and wheat [18]. Saline soils have Electrical Conductivity (ECe) more than 4 dSm-1 (~40 mM NaCl) because of the present of salts including sodium chloride, bicarbonates, magnesium and calcium sulphates, and also a number of inorganic ions, such as Na+, Ca2+, Mg2+, K+, CO3 2-, HCO3-, SO4 2-, and NO3- [19,20,21]. Sodium chloride (NaCl) is the most abundant salt in the saline soils.
Saline soil condition makes roots difficult to uptake nutrients and water which induce osmotic and ionic stress in the rice plants [22]. Under salinity stress condition, large amounts of Na+ influx into the plant cells and improved the Na+ concentrations in the cytoplasm and vacuole which toxic to the metabolism mechanism and leading to the cell death due to the osmotic and ionic stresses [18,23]. Excessive salt concentration in the plant cells absolutely leads to reduce cell membrane stability, cell wall damage, cytoplasmic degradation, plasmolysis, endoplasmic reticulum damage, accumulation of malate, citrate, and inositol in the leaf, and increase proline concentration, which leading to decrease the grain yield [24]. Recent study found that Pokkali is the salt-tolerant rice cultivar and IR20 belongs to the salt-sensitive cultivar [25]. In the salt-tolerant cultivar maintains a higher K+/Na+ ratio than salt-sensitive cultivar. Excess salts in the soil cause high osmotic pressure outside the roots which reduce the ability of root cells to take up water and nutrients from the soil. In order to adapt in the soil salinity stress, plants cells need to accumulate osmolytes, including proline, glycine, taurine, sugars, inositols, glycerol, sorbitol, etc. The salt-tolerant cultivar accumulates more proline than the salt-sensitive cultivar [26,27].
Salinity stress effects the morphological, physiological, and biochemical characteristics of the rice plants which varies with the growth stages, including reduction in plant height, productive tiller number, biomass, grain yield, filled grain per panicle, grain weight, grain quality, harvest index, photosynthetic activity, and increased Na+ and Cl- uptake to the shoot [28,29,30,31]. Under salinity stress at 3.5 dSm-1 or 50 mM NaCl, the grain yield of rice significantly reduced 90% [32,33]. Meanwhile, rice seedlings die at 10 dSm-1 [34]. In order to cope the soil salinity stress, rice plants develop several mechanisms, such as antioxidants for reactive oxygen species (ROS) detoxification, ion homeostasis, biosynthesis and accumulation of osmolytes for osmo-protection, and programmed cell death [26,35].
Soil salinity tolerance is a quantitative traits which controlled by multiple genes and highly affected by the environmental conditions [36,37,38,39,40]. The screening of rice for salinity tolerance is also complex. Thus, salinity screening under laboratory conditions is more controllable, rapid and accurate than field screening. Under the field screening, environmental conditions such as dynamic climate factors and soil heterogeneity might influence the accuracy of the salinity effects on the rice plants [41]. The potential indicators for salt tolerance screening for morphological, physiological, and biochemical characteristics are germination percentage, germination time, seedling vigor index, root length, shoot length, plant biomass, cell-membrane stability, Na+/K+ ratio, proline content, malondialdehyde (MDA) content, hydrogen peroxide (H2O2) content, sugar content, ethylene content, and chlorophyll content [42,43]. International Rice Research Institute (IRRI) has been released the standard evaluation score for visual salinity injury from 1 to 9 [6]. Salt tolerant is indicated by the lower score (1) and the salt sensitive is denoted by the higher score (9) based on the leaf symptoms, tiller number, and the growth characteristics under salinity conditions. Identification of quantitative trait loci (QTL) and cloning of genes correlated to salt tolerance in rice accelerate developing salt-tolerance rice varieties (Qin et al., 2020). Meanwhile, not many genes associated with the salt tolerance have been isolated and applied in rice breeding programs.
Many QTLs associated with the salinity tolerance have been identified by using mapping populations derived from crosses between salt-tolerant rice varieties and salt-sensitive varieties. The most popular marker in QTL mapping is single nucleotide polymorphism (SNP) [44,45]. A recombinant inbred line (RIL) population that derived from a cross between salt-tolerant Pokkali and salt-sensitive IR29 has been used to identify a major QTL Saltol on chromosome 1 which involved in regulating Na+/K+ homeostasis [46]. Another QTLs that have been identified are qSKC-1, qSNC-7, qSE3, and qST1 which plays an important role in salt tolerance at the different growth stages [26]. Genome-wide association studies (GWAS) accelerates the breeding of salt-tolerant rice varieties because of the availability of rice reference genome and next-generation sequencing (NGS) techniques. GWAS identified more accurate genomic locations associated with the salinity tolerance. A total of twenty-one QTLs and two candidate genes correlated to salinity tolerance were identified by using GWAS with 664 rice varieties [47]. GWAS was also conducted to identify salt-tolerant loci during reproductive stage [48,49]. A number of genes associated with the grain yield under salinity stress condition were also identified by using GWAS with 708 rice genotypes [50].
Analysis of genomic, transcriptomic, proteomics, and metabolomics also important in identifying genes correlated to salinity tolerance in rice. Under salinity stress, a series of changes in the rice plants, including the gene expression, protein content, and metabolite concentrations [51,52]. A number of potential genes associated with the salinity tolerance can be identified by comparing the transcriptome, proteome, and metabolome characteristics of salt-tolerant rice varieties with salt-sensitive varieties under salinity stress versus normal conditions. The most important goal in relieving the soil salinity problem is to identify and develop the rice varieties with high tolerance to salinity stress. Rice genotypes with highly salinity tolerant can be identified by using effective screening methods and provide donor alleles for salt tolerance to develop high salinity tolerant varieties through rice breeding program. By understanding the mechanism of salinity tolerance in the rice plants based on the morphological, physiological, biochemical, and genetics effects, the development of rice varieties with high salinity tolerant by genetic engineering techniques can be accelerated [28,53]. Breeding for rice salt tolerance is a major goal for rice breeders in agricultural areas with saline soil condition to ensure the food sustainability. The objectives of this research is to screen the salt tolerance of Asian rice cultivars based on the morphological, physiological, and biochemical characteristics.
2. Results
2.1. Morphological Responses of Asian Rice Cultivars to Salt Stress at Seedling and Early Vegetative Stages
All of the Asian rice varieties showed normal growth in the control condition. Percentage germination of Asian rice cultivars under salinity condition of 200 mM NaCl showed reduction compared with the normal condition because of the salt caused retardation in plumule and radicle length. Under salinity stress conditions, the Asian rice cultivars had a wide range of percentage germination (Figure 1) and visual symptoms of salt injury at the seedling stage ranging from score 1 (salt tolerant) to score 9 (salt susceptible) (Figure 2). Based on the SES of visual salt injury at the seedling stage, 31 rice cultivars were identified as salt tolerant, 45 cultivars were classified as moderately salt tolerant, and 40 cultivars belong to salt susceptible. The most salt tolerant were Pokkali from India, TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea. Meanwhile, the most salt susceptible rice varieties were IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea.
Another morphological responses to salinity stress condition, including root length, root fresh weight, shoot length, and plant biomass, showed variation among the Asian rice cultivars (Figure 3). In all of the rice cultivars, root length, root fresh weight, shoot length, and plant biomass were reduced under salinity stress condition, and the reduction was high as 59%, 51%, 61%, and 55% respectively. Pokkali from India, TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea showed the lowest reduction for root length, root fresh weight, shoot length, and plant biomass. Meanwhile, IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea had the highest reduction for root length, root fresh weight, shoot length, and plant biomass. All of the Asian rice cultivars displayed root length reduction ranged from 33% to 59%; the range reduction for root fresh weight from 35% to 51%; the shoot length had range reduction from 37% to 61%; and the reduction ranged for plant biomass from 31 to 55%.
Rice plants developed leaf symptoms, such as leaf rolling, yellowing, and necrotic lesions when rice is grown under salinity stress condition. Leaf rolling leads to minimize the water loss by respiration caused by water deficit under salinity stress condition. Salinity stress at the early vegetative stage of rice plants with 3 leaves; leaf rolling, yellowing, necrotic lesions, drying of leaves and other senescence symptoms were observed in all Asian rice cultivars with a wide range of variation (Figure 4). The damage of leaves under salinity stress condition related to the accumulation of Na+ from root to shoot.
Salinity stress condition reduced the total chlorophyll content significantly in the Asian rice cultivars which belong to the salt-susceptible cultivars. The decrease of chlorophyll content in 116 Asian rice cultivars showed a wide range variation (Figure 4). Reduction of chlorophyll content influences the low photosynthetic activity. This result was in line with the other crops, including peas [56], wheat [57], rapeseed [58], and safflower [59].
2.2. The Effect of Salinity Stress on Physiological and Biochemical Characteristics of Asian Rice Cultivars at Seedling and Early Vegetative Stages
Under salinity stress condition, sodium (Na) content in all of the Asian rice cultivars were increased significantly. The highest accumulation of sodium was found in the salt-susceptible rice varieties such as IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea followed by salt-moderate tolerant including Nipponbare from Japan, Padi Tarab from Malaysia, and Ciherang from Indonesia. Meanwhile, the lowest sodium concentration was observed in the shoots of salt-tolerant varieties but had higher potassium (K) accumulation than the sensitive ones, such as in Pokkali from India, TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea. The Na/K ratio in all 116 Asian rice varieties exhibited a wide variation (Figure 5).
Cell membrane stability of the rice plants under salinity stress tolerant was influenced by the osmotic adjustment. With the increasing of Na+ concentration in the cells, water potential inside of the cells is reducing and effect the cell membrane stability. Salt-sensitive rice varieties like IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea accumulated high concentration of Na+ which experience toxicity and cell damage (Figure 6).
In order to maintain the cellular osmotic balance under saline condition, the rice plant cells accumulate compatible solutes or metabolites such as proline, malondialdehyde (MDA), hydrogen peroxide (H2O2), and sugar content. All of the 116 Asian rice cultivars showed fluctuated compatible solutes under salinity stress condition (Figure 7). In the salt-tolerant rice varieties such as Pokkali from India, TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea accumulated higher amount of proline and sugar than the salt-sensitive varieties. Meanwhile, in the salt-tolerant varieties accumulated lower concentration of MDA and H2O2 compared to the sensitive ones.
2.3. Correlation among Morphological, Physiological, and Biochemical Traits under Salt Stress
Correlation of all traits lead us to understand the relationships among traits under salinity stress tolerance in Asian rice cultivars. Instead of considering only morphological traits such as salinity score, percentage germination, root length, root fresh weight, shoot length, plant biomass, plant height, leaf rolling, and chlorophyll content; physiological and biochemical traits, including Na/K ratio, cell membrane stability, proline, MDA, hydrogen peroxide, and sugar could be potential parameters for assessing salinity tolerance mechanisms.
Pearson correlations were calculated among 15 traits of morphological, physiological, and biochemical parameters under salinity stress condition at seedling and early vegetative stages (Table 3). Based on the Pearson correlations, there are 20 correlation pairs which reached significant levels under 200 mM NaCl condition. Salt-tolerant rice cultivars showed higher percentage germination, root length, root fresh weight, shoot length, plant biomass, plant height, and chlorophyll content (r=0.87*, r=0.91*, r=0.93*, r= 0.92*, r=0.85*, r=0.89*, r=87*, respectively). Meanwhile, salt-sensitive rice cultivars with higher salinity score had higher leaf rolling, Na/K ratio, and H2O2 content (r=0.95*, r=0.92*, r=0.91*, r=0.91*, respectively) under salinity stress condition. Meanwhile, salt-tolerant rice cultivars had high proline (r=90*) and sugar content (0.91*). High root length, root fresh weight, and shoot length affected the high plant biomass (r=0.87*, r=0.91*, r=0.92*, respectively). Asian rice cultivars with high Na/K ratio concentration under salinity stress condition was associated with high concentration of MDA and H2O2 (r=0.97*, r=0.96, respectively).

Table 3.
Correlation coefficients among morphological, physiological, and biochemical parameters from 116 Asian rice cultivars under 200 mM NaCl at seedling and early vegetative stages.
Table 3.
Correlation coefficients among morphological, physiological, and biochemical parameters from 116 Asian rice cultivars under 200 mM NaCl at seedling and early vegetative stages.
| SS | PG | RL | RFW | SL | PB | PH | LR | CC | CMS | Na/K | Proline | MDA | H2O2 | Sugar | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
SS PG RL RFW SL PB PH LR CC CMS Na/K Proline MDA H2O2 Sugar |
1 -0.87* -0.91* -0.93* -0.92* -0.85* -0.89* 0.95* -0.87* -0.14 0.92* -0.90* 0.57 0.91* -0.91* |
1 0.61 0.57 0.68 0.73 0.67 0.53 0.56 0.51 -0.47 -0.67 -0.61 -0.73 -0.64 |
1 0.89* 0.83 0.87* 0.88* 0.51 -0.73 0.81 -0.61 -0.58 -0.51 -0.62 -0.64 |
1 0.75 0.91* 0.68 0.42 0.83 0.69 -0.47 -0.53 -0.42 -0.39 -0.54 |
1 0.92* 0.79 0.39 0.74 0.51 -0.68 -0.53 -0.41 -0.36 -0.31 |
1 0.92* 0.41 0.86 0.62 -0.57 -0.41 -0.47 -0.58 -0.64 |
1 0.37 0.79 0.84 -0.62 -0.68 -0.79 -0.61 -0.57 |
1 -0.59 0.68 0.55 0.61 0.69 0.54 0.62 |
1 0.36 -0.27 -0.22 -0.22 -0.27 -3.48 |
1 -0.35 -0.29 -0.41 -0.32 -0.37 |
1 -0.47 0.97* 0.96* -0.51 |
1 -0.39 0.42 0.38 |
1 0.85 0.41 |
1 0.25 |
1 |
* Significant at the p < 0.05. SS: salinity score, PG: percentage germination, RL: root length, RFW: root fresh weight, SL: shoot length, PB: plant biomass, PH: plant height, LR: leaf rolling, CC: chlorophyll content, CMS: cell membrane stability, Na/K: Na+/K+ ratio, Proline: proline content, MDA: malondialdehyde content, H2O2: hydrogen peroxide content, Sugar: sugar content.
3. Discussion
Seedlings grown in salinity stress condition of 116 Asian rice cultivars exhibited different visual symptoms of salt injury. Salinity condition decreased percentage germination of Asian rice cultivars due to the osmotic and ionic stress cause insufficient water absorption which lead to the toxic effects on the seed embryo. Based on the interaction between salinity condition and rice cultivars, among the Asian rice cultivars there is a wide difference in response to salinity stress condition. Salt-tolerant seedlings were distinguished from the salt-sensitive seedlings under salinity stress condition. Our results are consistent with the previous studies which mentioned that different genotypes of barley, cabbage, and Suaeda species showed different responses to salinity condition with regard to percentage germination [60,61,62,63]. Salinity condition decreased the radicle and plumule growth. The retardation of the radicle and plumule length because of the reduction in turgor cells.
Visual salt injury begins with the reduction of effective leaf area. Salt-sensitive rice cultivars had high score for leaf rolling and leads to the decrease of photosynthetic activity. The distribution for root length, root fresh weight, shoot length, and plant biomass across 116 Asian rice cultivars under salinity stress condition showed a wide fluctuations. Root length and shoot length were shorter in salinity condition, compared to the normal condition. Root fresh weight and plant biomass of salt-susceptible cultivars showed higher percent reduction than salt-tolerant cultivars. Lower percent reduction of root length, root fresh weight, shoot length, and plant biomass were recorded in Pokkali from India followed by TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea. On the other hand, higher percent reduction of root length, root fresh weight, shoot length, and plant biomass were exhibited by IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea. Salt-tolerant rice cultivars displayed less growth reduction than salt-sensitive cultivars under salinity stress condition.
Cell membrane stability of 116 Asian rice cultivars were affected by the salinity stress condition. In the salt-sensitive rice cultivars, the cell membrane structure was damaged by the salt which increases the membrane permeability, and destroy the plasma membrane; as a result, the plant growth reduced [64]. In order to maintain the Na/K balance in the shoot, the plant absorb more K and exclude the toxic Na [39,65]. In this study, salt-tolerant rice cultivars showed the ability to absorb more K than Na in order to maintain the Na/K balance in the shoot. According to Ponnamperuma [66], K concentration in the shoot has a positive correlation with salinity tolerance because of the K is important in stomatal functions. Gregorio and Senadhira [39] also reported that salt-tolerant rice cultivars had higher K concentration and lower Na content in the shoot. This Na/K balances and maintains of low Na/K ratio are part of the salt tolerance mechanisms and could be promising criterion for salt-tolerant selection [15,67].
Variation of chlorophyll content in 116 Asian rice varieties can be used as a potential salinity stress indicator because chlorophyll content reduced in the salt-sensitive rice varieties under salinity stress condition. The salt-sensitive rice varieties, including IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea showed lower chlorophyll content compared to the salt-tolerant varieties. Salinity stress condition causes oxidative stress to the chloroplast that reduced the size and number of chloroplast in the leaves by inhibiting the synthesis of chloroplast [69,70,71]. Chlorophyllase enzyme activity which degrade the chlorophyll increases under salinity stress condition that lead the decrease of photosynthetic activity [16].
Salt-tolerant rice varieties accumulated high proline under salinity stress condition [72,73]. In this study, salt-tolerant varieties such as Pokkali from India followed by TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea also showed higher proline concentration compared to the salt-sensitive varieties. These results were consistent with Ghosh et al. [74] who found that salt-tolerant rice cultivars like Nonabokra and Pokkali under salinity stress condition displayed the high proline concentration in seedling stage. Under salinity stress condition, proline plays an important role in protection the protein against denaturation by regulate the redox potential and acts as source of nitrogen and carbon for post salt stress recovery in the rice plants [75,76,77].
Under salinity stress condition, malondialdehyde (MDA) is increasing in the salt-sensitive rice cultivars such as IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea. MDA represents the biological membrane damage because MDA is the primary product of decomposition process from unsaturated fatty acids due to oxidative stress under 200 mM NaCl [76,78,79,80,81,82,83,84]. The amount of hydrogen peroxide (H2O2) showed a wide variation in 116 Asian rice cultivars under salinity stress condition. Salt-tolerant rice varieties like Pokkali from India followed by TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea exhibited lower H2O2 than in the salt-sensitive ones. Sugar content of salt-tolerant rice varieties showed significantly increase under salinity stress condition. These results consistent with the previous study by Chang et al. [85] that found many sugars like raffinose, glucose, fructose, sucrose, galactose, mannose, ribose, xylose, melibiose, galactitol, mannitol, rhamnose, ribose, and erythritol were increasing in the leaves of the salt-tolerant rice cultivars such as Fatmawati and Dendang under salinity stress condition. The accumulation of sugars under salinity stress condition is to escape from the osmotic stress [86,87,88,89,90].
At the seedling and early vegetative stages, salt-tolerant rice varieties showed higher morphological characteristics such as percentage germination, root length, root fresh weight, shoot length, plant biomass, plant height, and chlorophyll content; and also higher physiological and biochemical traits like cell membrane stability, proline, and sugar content. Meanwhile, salt-sensitive ones had high leaf rolling, Na/K ratio, H2O2, and MDA content. These results were supported by the previous studies such as Peng et al. [91], Zhang et al. [92], Dwiningsih et al., [93], and Bhowmik et al. [10]. Many studies indicated that all of the growth stages in rice plants under salinity stress condition showed different responses to salt [94]. The most sensitive stage to salt is during fertilization and flowering, followed by early vegetative stage, germination, and maturity [95,96,97]. The tolerance responses of rice cultivars to the salinity stress condition may become the foundation of breeding tolerant rice cultivars in the saline agricultural area.
4. Materials and Methods
4.1. Plant Materials
A total of 116 rice genotypes originally from Asian countries (Table 1) were screened for salinity tolerance at the seedling stage and early vegetative stage (3 leaf stage). Seeds were sterilized with 10% (v/v) NaClO for 15 minutes and washed with distilled water. Pokkali was used as the salt-tolerant standard check and IR29 was used as salt-sensitive check [54].
4.2. Salinity Screening at Seedling and Early Vegetative Stage
Concentration of salt (NaCl) used in this experiment was 200 mM. Ten seeds of each rice genotype were germinated in the petri dishes containing blotting paper and were treated with 200 mM NaCl and kept in the incubator at 30oC. For the control condition, seeds were germinated with distilled water. After 12 days of salt treatment, germinated seedlings were transferred to a hydroponic system containing Yoshida’s medium [55] with 200 mM NaCl. Rice plants were grown in a growth-chamber at temperature 28oC/24oC for day/night with 65% humidity, and light intensity 500 µEm-2s-1 until early vegetative stage (3 leaf stage). Morphological, physiological, and biochemical responses, including root length, root fresh weight, shoot length, plant biomass, leaf rolling, chlorophyll content, Na+/K+ ratio, cell-membrane stability, proline content, malondialdehyde (MDA) content, hydrogen peroxide (H2O2) content, and sugar content were measured at early vegetative stage (3 leaf stage).
4.3. Measurement of Morphological, Physiological, and Biochemical Traits
4.3.1. Determination of Percentage Germination and Standard Evaluation Score for Salt Injury
The indicator for germinated seeds when the radicle had protruded through the seed coat, hypocotyl was extended, and cotyledon was unfolded. Percentage germination and visual symptoms of salt injury at the seedling stage was evaluated based on the standard evaluating score at 12 days of salt treatment (Table 2) [6].
4.3.2. Measurement of Root Length, Root Fresh Weight, Shoot Length, and Plant Biomass
Root length was measured from maximum length of the root for each rice plant at early vegetative stage (3 leaf stage). Whole weight of the roots of each rice plant was determined as the root fresh weight. Shoot length was measured from the ground surface to the tallest leaf tip with a ruler. The roots and shoot of each rice plant were weighted fresh and determined as plant biomass.
4.3.3. Determination of Leaf Rolling
The leaf rolling score on the three leaves of each plant was identified at early vegetative stage (3 leaf stage) based on the standard evaluation system for rice [6]. The range in score is from 1 to 5, 1 indicating unrolled leaves and fully turgid, 2 indicating leaves are folded (Deep-V-shaped), 3 indicating leaves are fully cupped (U-shaped), 4 indicating leaves margins touching (O-shaped), and 5 indicating completely rolled leaves.
4.3.4. Analysis of Chlorophyll Content
Chlorophyll content of the fully expanded leaves on the top of each plant was measured by using Soil and Plant Analyzer Development (SPAD)-502 Plus Chlorophyll Meter (Spectrum Technologies, USA). Leaf was inserted into the sample slot of the SPAD in such a way to avoid the midrib and five reading was measured for each leaf.
4.3.5. Measurement of Na/K Ratio
The ratio of sodium (Na) and potassium (K) concentration in the root and shoot were measured for each rice genotype grown under salinity condition of 200 mM at early vegetative stage (3 leaf stage). Each rice plant was rinsed with the distilled water and then dried at 65oC for 2 days. The dried tissue sample for each rice genotype was ground by mortar and pestle. A total of 100 mg of ground sample were digested in 3 ml hydrogen peroxide and 5 ml of nitric acid for 3 hours at 152−155oC. Then, the digested sample was diluted to a final volume of 12.5 ml. The concentration of Na and K were quantified by using a flame photometer (ANA-135, Tokyokoden, Tokyo, Japan). The estimated concentration of Na and K were calculated based on the standard curve.
4.3.6. Measurement of Cell Membrane Stability
Leaf cell membrane stability (CMS) of each rice genotype under salinity condition was determined by using this equation:
CMS (%) = 1 – (E1/E2) × 100
Leaf samples were washed with distilled water and then, placed in 10 ml of deionized water at 10oC for 18 hours. Next, the samples were heated at 52oC for 1 hour in a water bath. In order to diffuse the electrolytes from leaf tissue to aqueous media, the samples were incubated at 10oC for 24 hours. The samples were shaken and initial conductance (E1) was measured for each sample. All of the samples were then autoclaved at 121oC and 0.10 MPa for 15 minutes in order to kill the plant tissue and release the electrolytes. The samples were placed in incubator at 25oC for cooling down, then the samples were shaken, and the final conductance (E2) was measured.
4.3.7. Measurement of Proline Content
A total of 0.5 g fresh leaf sample of each rice genotype was diluted in 10 ml of 3% aqueous sulfosalicylic acid and centrifuged for 1 minute at 3000 rpm. The 2 ml of supernatant was reacted with 2 ml of glacial acetic acid and 2 ml of ninhydrin acid at 100oC for 1 hour. Exactly 2 ml of Toluene was used to extract the chromophore. Absorbance of the chromophore was measured at 520 nm by using Genesys 10-s UV/Vis Spectrophotometer (Thermo Spectronic, Waltham, MA, USA) and Toluene as the blank. A standard curve for proline content was quantified by using purified proline (Sigma Aldrich, Melbourne, VIC, Australia). Proline content of each rice genotype was calculated by using the following formula.
((µg proline/mL × mL toluene)/115.5 µg/µmole) × (g sample/5) = µmoles proline gram FW−1
4.3.8. Measurement of Malondialdehyde (MDA) Content
Exactly 0.5 g of rice leaf from each rice genotype was cut and homogenized with 1.5 ml of a 0.5% (w/v) thiobarbituric acid solution that consisting of 20% (w/v) trichloroacetic acid and 1.5 ml of distilled water. The solution was heated for 25 minutes at 95oC, and then the reaction was stopped by placing the samples on ice. Next, the solution was centrifuged, absorbance of the supernatant was measured at 532 and 600 nm. The extinction coefficient of mM-1 cm-1 was used to calculate the MDA content and expressed as nanomoles per gram (nmol/g) of fresh weight.
4.3.9. Measurement of Hydrogen Peroxide (H2O2) Content
About 0.1 g of leaf from each rice genotype was diluted in 3 ml of 5% (w/v) trichloroacetic acid and incubated for 3 hours at 4oC. Next, 1 ml of FOX reagent was added to the 0.2 ml of supernatant of the sample, then mixed and incubated for 15 minutes at 25oC. Absorbance of the solution was read at 560 nm. The H2O2 content was expressed as micromoles per gram (µmol/g) of fresh weight.
4.3.10. Measurement of Sugar Content
Leaf sample from each rice genotype was ground and 1.0 g of ground sample was added to the 1 ml of distilled water. Then, 1 ml Anthrone reagent was added to the suspension and incubated for 8 minutes at 25oC. Absorbance of the solution was read at 630 nm. The content of soluble sugar in each rice genotype was calculated by using a standard graph and expressed in milligram per gram (mg/g) of fresh weight.
4.4. Statistical Analysis
The experiment of salinity screening was conducted in a randomized completely block design (RCBD) with five replications. The salinity treatment and control conditions were compared by Least Significant Difference (LSD) test at 0.05 probability level.
5. Conclusions
Combining morphological, physiological, and biochemical assessments for screening the salt tolerant of 116 Asian rice cultivars was able to classify into tolerant, moderate, and sensitive rice cultivars under salinity stress condition, and also for understanding the salt tolerance mechanisms. The rice cultivars which belong to salt-tolerant including Pokkali from India followed by TCCP 266 from Philippines, IR 45427 also from Philippines, and Namyang 7 from Korea. Whereas, salt-sensitive rice varieties like IR29 from Philippines, IR58 also from Philippines, Daegudo from Korea, and Guweoldo also from Korea. The identified salt-tolerant rice varieties potentially provide a valuable germplasm resource in the breeding programs for developing salt-tolerant rice.
Author Contributions
Conceptualization, Y.D., M.A., and J.A.; methodology, Y.D., J.A., and M.A.; software, J.A., M.A., and Y.D.; validation, Y.D., J.A., and M.A.; formal analysis, M.A., J.A., and Y.D.; writing – original draft preparation, Y.D., J.A., and M.A. All authors have read and agreed to the published version of the manuscript.
Funding
Not Applicable.
Data Availability Statement
Not Applicable.
Acknowledgments
This project was supported by Researchers Supporting Project number (RSP2023R193) King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gopalan, C.; Rama Sastri, B.V.; Balasubramanian, S. Nutritive Value of Indian Foods. National Institute of Nutrition (NIN), ICMR, India 2007.
- Khush, G.S.; Virk, P.S. Rice Breeding: Achievements and Future Strategies. Crop Improv. 2000, 27, 115-144.
- GRiSP. Rice Almanac. Fourth ed. International Rice Research Institute, Los Banos, Philippines 2013.
- Dwiningsih, Y.; Kumar, A.; Thomas, J.; Ruiz, C.; Alkahtani, J.; Al-hashimi, A.; Pereira, A. Identification of Genomic Regions Controlling Chalkiness and Grain Characteristics in a Recombinant Inbred Line Rice Population Based on High-Throughput SNP Markers. Genes 2021, 12, 11, 1690. [CrossRef]
- ANU. Technological Transformation of Productivity, Profitability and Sustainability: Rice. In The First Ten K R Narayanan Orations: Essays by Eminent Persons on the Rapidly Transforming Indian Economy. Australia South Asia Research Centre The Australian National University 2006. http://epress.anu.edu.au/narayanan/mobile_ devices/pr01.html.
- Gregorio, G.B. Tagging Salinity Tolerant Genes in Rice Using Amplified Fragment Length Polymorphism (AFLP). Ph.D. dissertation. University of the Philippines Los Baños College, Laguna, Philippines 1997, 118.
- Abdullah, Z.; Khan, M.A.; Flowers, T.J. Causes of Sterility in Seed Set of Rice under Salinity Stress. J. Agron. Crop. Sci. 2001, 187, 25–32. [CrossRef]
- Dwiningsih, Y. Molecular genetic analysis of drought resistance and productivity traits of rice genotypes. University of Arkansas, Fayetteville, USA. 2020.
- Aref, F.; Rad, H.E. 2012. Physiological Characterization of Rice under Salinity Stress during Vegetative and Reproductive Stages. Indian J. Sci. Technol. 2012, 5, 2578-2586.
- Bhowmik, S.K.; Titov, S.; Islam, M.M.; Siddika, A.; Sultana, S.; Haque, M. Phenotypic and genotypic screening of rice genotypes at seedling stage for salt tolerance. African Journal of Biotechnology 2009, 8, 23, 6490-6494.
- Dwiningsih, Y.; Kumar, A.; Thomas, J.; Yingling, S.; Pereira, A. Molecular genetic analysis of drought resistance and productivity in US rice cultivars. Plant and Animal Genome XXVII Conference (January 12-16, 2019). 2019.
- Bado, S.; Forster, B.P.; Ghanim, A.; Jankowicz-Cieslak, J.; Berthold, G.; Luxiang, L. Protocols for pre-field screening of mutants for salt tolerance in rice, wheat and barley. Springer 2016. [CrossRef]
- Ueda, A.; Yahagi, H.; Fujikawa, Y.; Nagaoka, T.; Esaka, M.; Calcaño, M.; González, M.; Martich, J.; Saneoka, H. Comparative physiological analysis of salinity tolerance in rice. Soil Science and Plant Nutrition 2013, 59, 6, 896-903. [CrossRef]
- Ge, X.; Khan, Z.I.; Chen, F.; Akhtar, M.; Ahmad, K.; Ejaz, A.; Ashraf, M.A.; Nadeem, M.; Akhtar, S.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. A study on the contamination assessment, health risk and mobility of two heavy metals in the soil-plants-ruminants system of a typical agricultural region in the semi-arid environment. Environmental Science and Pollution Research 2022, 29,14584–14594. [CrossRef]
- Lee, K.S.; Choi, W.Y.; Ko, J.C.; Kim, T.S.; Gregorio, G.B. Salinity tolerance of japonica and indica rice (Oryza sativa L.) at the seedling stage. Planta 2003, 216, 1043–1046. [CrossRef]
- Kordrostami, M.; Rabiei, B.; Kumleh, H. Biochemical, physiological and molecular evaluation of rice cultivars differing in salt tolerance at the seedling stage. Physiol Mol Biol Plants 2017, 23, 3, 529–544. [CrossRef]
- Maqsood, A.; Khan, Z.I.; Ahmad, K.; Akhtar, S.; Ashfaq, A.; Malik, I.S.; Sultana, R.; Nadeem, M.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M. Quantitative evaluation of zinc metal in meadows and ruminants for health assessment: implications for humans. Environmental Science and Pollution Research 2022, 29, 15, 21634–21641. [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [CrossRef]
- Hoang, T.M.L.; Williams, B.; Khanna, H.; Dale, J.; Mundree, S.G. Physiological basis of salt stress tolerance in rice expressing the antiapoptotic gene SfIAP. Funct. Plant Biol. 2014, 41, 1168–1177. [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants. Academic Press, London. 1995.
- Dwiningsih, Y.; Thomas, J.; Kumar, A.; Gupta, C.; Ruiz, C.; Yingling, S.; Crowley, E.; Pereira, A. Molecular genetic analysis of drought resistance and productivity mechanisms in rice. Plant and Animal Genome XXVIII Conference, January 11-15, 2020.
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499.
- Dwiningsih, Y.; Kumar, A.; Thomas, J.; Ruiz, C.; Alkahtani, J.; Baisakh, N.; Pereira, A. Quantitative trait loci and candidate gene identification for chlorophyll content in RIL rice population under drought conditions. Indonesian Journal of Natural Pigments 2021, 3, 2, 54-64. [CrossRef]
- Sahi, C.; Singh, A.; Kumar, K. Salt stress response in rice: genetics, molecular biology and comparative genomics. Funct. Integr. Genomics 2006, 6, 263–284.
- Kavitha, P.G.; Miller, A.J.; Mathew, M.K.; Maathuis, F.J.M. Rice cultivars with differing salt tolerance contain similar cation channels in their root cells. Journal of Experimental Botany 2012, 63, 8, 3289–3296. [CrossRef]
- Hoang, T.; Tran, T.; Nguyen, T.; Williams, B.; Wurm, P.; Bellairs, S.; Mundree. Improvement of Salinity Stress Tolerance in Rice: Challenges and Opportunities. Agronomy 2016, 6, 54. [CrossRef]
- Sitrarasi, R.; Nallal, U.M.; Razia, M.; Chung, W.J.; Shim, J.; Chandrasekaran, M.; Dwiningsih, Y.; Rasheed, R.A.; Alkahtani, J.; Elshikh, M.S.; Debnath, O.; Ravindran, B. Inhibition of multi-drug resistant microbial pathogens using an ecofriendly root extract of Furcraea foetida silver nanoparticles. Journal of King Saud University-Science 2022, 34, 2, 101794. [CrossRef]
- De Leon, T.B.; Linscombe, S.; Gregorio, G.; Subudhi, P.K. Genetic variation in Southern USA rice genotypes for seedling salinity tolerance. Front. Plant Sci. 2015, 6, 374. [CrossRef]
- Mohammadi-Nejad, G.; Singh, R.K.; Arzanic, A.; Rezaiec, A.M.; Sabourid, H.; Gregorio, G.B. Evaluation of Salinity Tolerance in Rice Genotypes. Int. J. Plant Prod. 2010, 4, 199–207.
- Morales, S.G.; Tellez, L.I.T.; Merino, F.C.G.; Caldana, C.; Victoria, D.E.; Cabrera, B.E.H. Growth, Photosynthetic Activity, and Potassium and Sodium Concentration in Rice Plants under Salt Stress. Acta Sci. Agron. 2012, 34, 317-324. [CrossRef]
- Dwiningsih, Y.; Thomas, J.; Kumar, A.; Gupta, C.; Gill, N.; Ruiz, C.; Alkahtani, J.; Baisakh, N.; Pereira A. Identification of QTLs and Candidate Loci Associated with Drought-Related Traits of the K/Z RIL Rice Population. Research Square 2022. [CrossRef]
- Asch, F.; Dingkuhn, M.; Dorffling, K.; Miezan, K. Leaf K/Na ratio predicts salinity induced yield loss in irrigated rice. Euphytica 2000, 113, 109–118. [CrossRef]
- Yeo, A.R.; Flowers, T.J. Salinity resistance in rice (Oryza Sativa L.) and a pyramiding approach to breeding varieties for saline soils. Aust. J. Plant Physiol. 1986, 13, 161–173. [CrossRef]
- Munns, R.; James, R.A.; Lauchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [CrossRef]
- Dwiningsih, Y.; Alkahtani, J. Genetics, Biochemistry and Biophysical Analysis of Anthocyanin in Rice (Oryza sativa L.). Advance Sustainable Science, Engineering and Technology (ASSET) 2022, 4, 1. [CrossRef]
- Ferreira, L.J.; Azevedo, V.; Maroco, J.; Oliveira, M.M.; Santos, A.P. Salt Tolerant and Sensitive Rice Varieties Display Differential Methylome Flexibility under Salt Stress. PLoS ONE 2015, 10, 5, e0124060. [CrossRef]
- Flowers, T.J. Improving crop salt tolerance. J. Exp. Bot. 2004, 55, 307–319. [CrossRef]
- Koyama, M.L.; Levesley, A.; Koebner, R.; Flowers, T.J.; Yeo, A.R. Quantitative trait loci for component physiological traits determining salt tolerance in rice. Plant Physiol. 2001, 125, 406–422. [CrossRef]
- Gregorio, G.B.; Senadhira, D. Genetic analysis of salinity tolerance in rice (Oryza sativa L.). Theor. Appl. Genet. 1993, 86, 333–338. [CrossRef]
- Bashir, S.; Gulshan, A.B.; Iqbal, J.; Husain, A.; Alwahibi, M.S.; Alkahtani, J.; Dwiningsih, Y.; Bakhsh, A.; Ahmed, N.; Khan, M.J.; Ibrahim, M.; Diao, Z.H. Comparative role of animal manure and vegetable waste induced compost for polluted soil restoration and maize growth. Saudi Journal of Biological Sciences, 2021, 28, 4, 2534-2539. [CrossRef]
- Ali, M.; Yeasmin, L.; Gantait, S.; Goswami, R.; Chakraborty, S. Screening of rice landraces for salinity tolerance at seedling stage through morphological and molecular markers. Physiol Mol Biol Plants 2014, 20, 4, 411–423. [CrossRef]
- Flowers, T.J.; Yeo, A.R. Breeding for salinity resistance in crop plants: where next? Aust J Plant Physiol. 1995, 22, 875–884.
- Ali, M.H.; Khan, M.I.; Bashir, S.; Azam, M.; Naveed, M.; Qadri, R.; Bashir, S.; Mehmood, F.; Shoukat, M.A.; Li, Y.; Alkahtani, J.; Elshikh, M.S.; Dwiningsih, Y. Biochar and Bacillus sp. MN54 Assisted Phytoremediation of Diesel and Plant Growth Promotion of Maize in Hydrocarbons Contaminated Soil. Agronomy 2021, 11, 9, 1795. [CrossRef]
- Qin, H.; Li, Y.; Huang, R. Advances and Challenges in the Breeding of Salt-Tolerant Rice. Int. J. Mol. Sci. 2020, 21, 8385. [CrossRef]
- Dwiningsih Y.; Rahmaningsih M.; Alkahtani J. Development of single nucleotide polymorphism (SNP) markers in tropical crops. Advance Sustainable Science, Engineering and Technology (ASSET) 2020, 2, 2. [CrossRef]
- Thomson, M.J.; de Ocampo, M.; Egdane, J.; Rahman, M.A.; Sajise, A.G.; Adorada, D.L.; Tumimbang-Raiz, E.; Blumwald, E.; Seraj, Z.I.; Singh, R.K.; et al. Characterizing the saltol quantitative trait locus for salinity tolerance in Rice. Rice 2010, 3, 148–160. [CrossRef]
- Yuan, J.; Wang, X.; Zhao, Y.; Khan, N.U.; Zhao, Z.; Zhang, Y.; Wen, X.; Tang, F.; Wang, F.; Li, Z. Genetic basis and identification of candidate genes for salt tolerance in rice by GWAS. Sci. Rep. 2020, 10, 9958. [CrossRef]
- Lekklar, C.; Pongpanich, M.; Suriya-arunroj, D.; Chinpongpanich, A.; Tsai, H.; Comai, L.; Chadchawan, S.; Buaboocha, T. Genome-wide association study for salinity tolerance at the flowering stage in a panel of rice accessions from Thailand. BMC Genom. 2019, 20, 76. [CrossRef]
- Adil, M.; Bashir, S.; Bashir, S.; Aslam, Z.; Ahmad, N.; Younas, T.; Asghar, R.M.A.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front. Plant Sci. 2022, 13, 932861. [CrossRef]
- Liu, C.; Chen, K.; Zhao, X.Q.; Wang, X.Q.; Shen, C.C.; Zhu, Y.J.; Dai, M.L.; Qiu, X.J.; Yang, R.W.; Xing, D.Y.; et al. Identification of genes for salt tolerance and yield-related traits in rice plants grown hydroponically and under saline field conditions by genome-wide association study. Rice 2019, 12, 88. [CrossRef]
- Mansuri, R.M.; Shobbar, Z.S.; Jelodar, N.B.; Ghaffari, M.R.; Nematzadeh, G.A.; Asari, S. Dissecting molecular mechanisms underlying salt tolerance in rice: A comparative transcriptional profiling of the contrasting genotypes. Rice 2019, 12, 13. [CrossRef]
- Alkahtani, J.; Elshikh, M.S.; Dwiningsih, Y.; Rathi, M.A.; Sathya, R.; Vijayaraghavan, P. In-vitro antidepressant property of methanol extract of Bacopa monnieri. Journal of King Saud University – Science 2022, 34, 102299. [CrossRef]
- Alshiekheid, M.A.; Dwiningsih, Y.; Sabour, A.A.; Alkahtani, J. Phytochemical Composition and Antibacterial Activity of Zingiber cassumunar Roxb. against Agricultural and Foodborne Pathogens. Preprint 2022. [CrossRef]
- Pathan, M.S.; Nguyen, H.T.; Subudhi, P.K.; Courtois, B. Molecular Dissection of Abiotic Stress Tolerance in Sorghum and Rice. In: “Physiology and Biotechnology Integration for Plant Breeding”, (Eds.): H. T. Nguyen, H. T. and Blum, A. Marcel Dekker Inc., New York, USA, 2004, 525-570.
- Yoshida, S.; Forno, D.A.; Cock, J.H.; Gomez, K.A. Laboratory manual for physiological studies of rice. IRRI, Las Banos, Laguna, 1976, 83.
- Yildirim, B.; Yaser, F.; Ozpay, T.; Ozpay, D.; Turkozu, D.; Terziodlu, O.; Tamkoc, A. Variations in response to salt stress among field pea genotypes (Pisum sativum sp. arvense L.). J Anim Vet Adv. 2008, 7, 907–910.
- Raza, S.H.; Athar, H.U.R.; Ashraf, M. Influence of exogenously applied glycinebetaine on the photosynthetic capacity of two differently adapted wheat cultivars under salt stress. Pak J Bot. 2006, 38, 341–351.
- Mukhtar, E.; Siddiqi, K.; Bhatti, K.H.; Nawaz, K.; Hussain, K. Gas exchange attributes can be valuable selection criteria for salinity tolerance in canola cultivars (Brassica napus L.). Pak J Bot. 2013, 45, 35–40.
- Siddiqi, E.H.; Ashraf, M.; Hussain, M.; Jamil, A. Assessment of intercultivar variation for salt tolerance in safflower (Carthamus tinctorius L.) using gas exchange characteristics as selection criteria. Pak J Bot. 2009, 41, 2251–2259.
- Mano, Y.; Takeda, K. Diallel analysis of salt tolerance at germination and the seedling stage in barley (Hordeum vulgare L.). Breeding Sci. 1997, 47, 203-209. [CrossRef]
- Jamil, M.; Lee, K.B.; Jung, K.Y.; Lee, D.B.; Han, M.S.; Rha, E.S. Salt stress inhibits germination and early seedling growth in cabbage (Brassica oleracea capitata L.). Pakistan J. Biol. Sci. 2007, 10, 6, 910-914. [CrossRef]
- Guan, B.; Yu, J.; Lu, Z.; Japhet, W.; Chen, X.; Xie, W. Salt tolerance in two Suaeda species: seed germination and physiological response. Asian J. Plant Sci. 2010, 9, 4, 194-199. [CrossRef]
- Dwiningsih, Y.; Alkahtani, J. Phenotypic Variations, Environmental Effects and Genetic Basis Analysis of Grain Elemental Concentrations in Rice (Oryza sativa L.) for Improving Human Nutrition. Preprints 2022, 2022090263. [CrossRef]
- Zhang, R.; Hussain, S.; Wang, Y.; Liu, Y.; Li, Q.; Chen, Y.; Wei, H.; Gao, P.; Dai, Q. Comprehensive Evaluation of Salt Tolerance in Rice (Oryza sativa L.) Germplasm at the Germination Stage. Agronomy 2021, 11, 1569. [CrossRef]
- Dwiningsih, Y.; Alkahtani, J. Rojolele: a Premium Aromatic Rice Variety in Indonesia. Preprints 2022, 2022100373. [CrossRef]
- Ponnamperuma, F.N. Role of cultivar tolerance in increasing rice production in saline lands. In: Staples RC, Toenniessen GH (eds) Salinity tolerance in plants. Strategies for crop improvement. Wiley-Interscience, New York, 1984, 255–271.
- Dwiningsih, Y.; Al-Kahtani, J. Genome-Wide Association Study of Complex Traits in Maize Detects Genomic Regions and Genes for Increasing Grain Yield and Grain Quality. Advance Sustainable Science Engineering and Technology 2022, 4, 2, 0220209. [CrossRef]
- Ashraf, M.; Harris, P. Photosynthesis under stressful environments: an overview. Photosynthetica 2013, 51, 163–190. [CrossRef]
- Khafagy, M.A.; Arafa, A.A.; El-Banna, M.F. Glycinebetaine and ascorbic acid can alleviate the harmful effects of NaCl salinity in sweet pepper. Aust J Crop Sci. 2009, 3, 257–267.
- Santos, V.C. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci Hortic. 2004, 130, 93–99.
- Lone, J.; Shikari, A.; Sofi, N.; Ganie, S.; Sharma, M.; Sharma, M.; Kumar, M.; Saleem, M.H.; Almaary, K.S.; Elshikh, M.S.; Dwiningsih, Y. Screening technique based on seed and early seedling parameters for cold tolerance of selected F2-derived F3 rice genotypes under controlled conditions. Sustainability 2022, 14, 14, 8447. [CrossRef]
- Kishor, P.B.; Sangam, S.; Amrutha, R.N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance. Curr Sci. 2005, 88, 424–438.
- Dwiningsih, Y.; Kumar, A.; Thomas, J.; Gupta, C.; Ruiz, C.; Alkahtani, J.; Baisakh, N.; Pereira, A. Identification and expression of abscisic acid-regulated genes in US RIL rice population under drought conditions. In 82nd Meeting of Southern Section of the American Society of Plant Biologists. 2021.
- Ghosh, N.; Adak, M.K.; Ghosh, P.D.; Gupta, S.; Sen Gupta, D.N.; Mandal, C. Differential responses of two rice varieties to salt stress. Plant Biotechnol Rep. 2011, 5, 89–103. [CrossRef]
- Fariduddin, Q.; Khalil, R.R.A.E.; Mir, B.A.; Yusuf, M.; Ahmad, A. 24-Epibrassinolide regulates photosynthesis, antioxidant enzyme activities and proline content of Cucumis sativus under salt and/ or copper stress. Environ Monit Assess. 2013, 185, 7845–7856.
- Saha, P.; Chatterjee, P.; Biswas, A.K. NaCl pretreatment alleviates salt stress by enhancement of antioxidant defense system and osmolyte accumulation in mungbean (Vigna radiata L. Wilczek). Indian J Exp Biol. 2010, 48, 593–600.
- Dwiningsih, Y.; Kumar, A.; Thomas, J.; Yingling, S.; Pereira, A. Identification of QTLs associated with drought resistance traits at reproductive stage in K/Z RILs rice population. In 5th Annual Meeting of the Arkansas Bioinformatics Consortium AR-BIC. 2020.
- Abu-Muriefah, S.S. Effect of sitosterol on growth, metabolism and protein pattern of pepper (Capsicum Annuum L.) plants grown under salt stress conditions. Int J Agric Crop Sci. 2015, 8, 94–106.
- Azuma, R.; Ito, N.; Nakayama, N.; Suwa, R.; Nguyen, N.T.; Larrinaga Mayoral, J.A.; Esaka, M.; Fujiyama, H.; Saneoka, H. Fruits are more sensitive to salinity than leaves and stems in pepper plants (Capsicum annuum L.). Sci Hortic. 2010, 125, 171–178. [CrossRef]
- Dwiningsih, Y.; Kumar, A.; Thomas, J.; Yingling, S.; Pereira, A. Molecular genetic analysis of drought resistance and productivity in K/Z RIL rice population. Arkansas Bioinformatics Consortium. 2019.
- Stepien, P.; Klobus, G. Antioxidant defense in the leaves of C3 and C4 plants under salinity stress. Physiol Plantarum 2005, 125, 31–40. [CrossRef]
- Sudhakar, C.; Lakshmi, A.; Giridarakumar, S. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Sci. 2001, 161, 613–619. [CrossRef]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ Exp Bot. 2003, 49, 69–76. [CrossRef]
- Dwiningsih, Y.; Thomas, J.; Kumar, A.; Gupta, C.; Crowley, E.; Ruiz, C.; Pereira, A. Drought stress response in US recombinant inbred line of rice population. In National Science Foundation (NSF) 26th National Conference 2019, 26, 76, 127.
- Chang, J.; Cheong, B.; Natera, S.; Roessner, U. Morphological and metabolic responses to salt stress of rice (Oryza sativa L.) cultivars which differ in salinity tolerance. Plant Physiology and Biochemistry, 2019, 144, 427–435. [CrossRef]
- Guo, R.; Yang, Z.Z.; Li, F.; Yan, C.R.; Zhong, X.L.; Liu, Q.; Xia, X.; Li, H.R.; Zhao, L. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress. BMC Plant Biol. 2015, 15. [CrossRef]
- Dwiningsih, Y.; Thomas, J.; Kumar, A.; Gupta, C.; Yingling, S.; Basu, S.; Pereira, A. Circadian expression patterns of the HYR gene. Arkansas Bioinformatics Consortium 2018, 7, 11, 34.
- Rock, W.R.L. Artificial Intelligence (AI) in Arkansas (AR). 2020.
- Ismanto, A.; Hadibarata, T.; Widada, S.; Indrayanti, E.; Ismunarti, D.; Safinatunnajah, N.; Kusumastuti, W.; Dwiningsih, Y.; Alkahtani, J. Groundwater contamination status in Malaysia: level of heavy metal, source, health impact, and remediation technologies. Bioprocess and Biosystems Engineering 2022. [CrossRef]
- Dwiningsih, Y.; Kumar, A.; Thomas, J.; Pereira, A. Identification drought-tolerance rice variety for reducing climatic impacts on rice production. In Fulbright Enrichment Seminar Climate Change, Estes Park, Colorado, USA. 2017.
- Peng, S.; Cassman, K.G.; Virmani, S.S.; Sheehy, J.; Khush, G.S. Yield potential trends of tropical rice since the release of IR8 and the challenge of increasing rice yield potential. Crop Sci. 1999, 39, 1552-1559. [CrossRef]
- Zhang, Z.H.; Li, P.; Wang, L.X.; Hu, Z.L.; Zhu, L.H.; Zhu, Y.G. Genetic dissection of the relationships of biomass production and partitioning with yield and yield related traits in rice. Plant Sci. 2004, 167, 1-8.
- Dwiningsih, Y.; Alkahtani, J. Agronomics, Genomics, Breeding and Intensive Cultivation of Ciherang Rice Variety. Preprints 2022, 2022110489. [CrossRef]
- Dwiningsih, Y.; Alkahtani, J. Potential of Pigmented Rice Variety Cempo Ireng in Rice Breeding Program for Improving Food Sustainability. Preprints 2023, 2023020039. [CrossRef]
- IRRI [International Rice Research Institute]. Annual report for 1967. Manila (Philippines): International Rice Research Institute 1967, 308.
- Dwiningsih, Y.; Kumar, A.; Thomas, J.; Gupta, C.; Ruiz, C.; Baisakh, N.; Pereira, A. QTLs analysis and identification of candidate genes for flag leaf characteristics related to grain yield in US RIL rice population under drought conditions. In ASA, CSSA, SSSA International Annual Meeting. ASA-CSSA-SSSA. 2021.
- Pearson, G.A.; Ayers, S.D.; Eberhard, D.L. Relative salt tolerance of rice during germination and early seedling development. Soil Sci. 1966, 102, 151-156. [CrossRef]
Figure 1.
Percentage germination of 116 Asian rice cultivars under salinity stress condition 200 mM NaCl.
Figure 1.
Percentage germination of 116 Asian rice cultivars under salinity stress condition 200 mM NaCl.
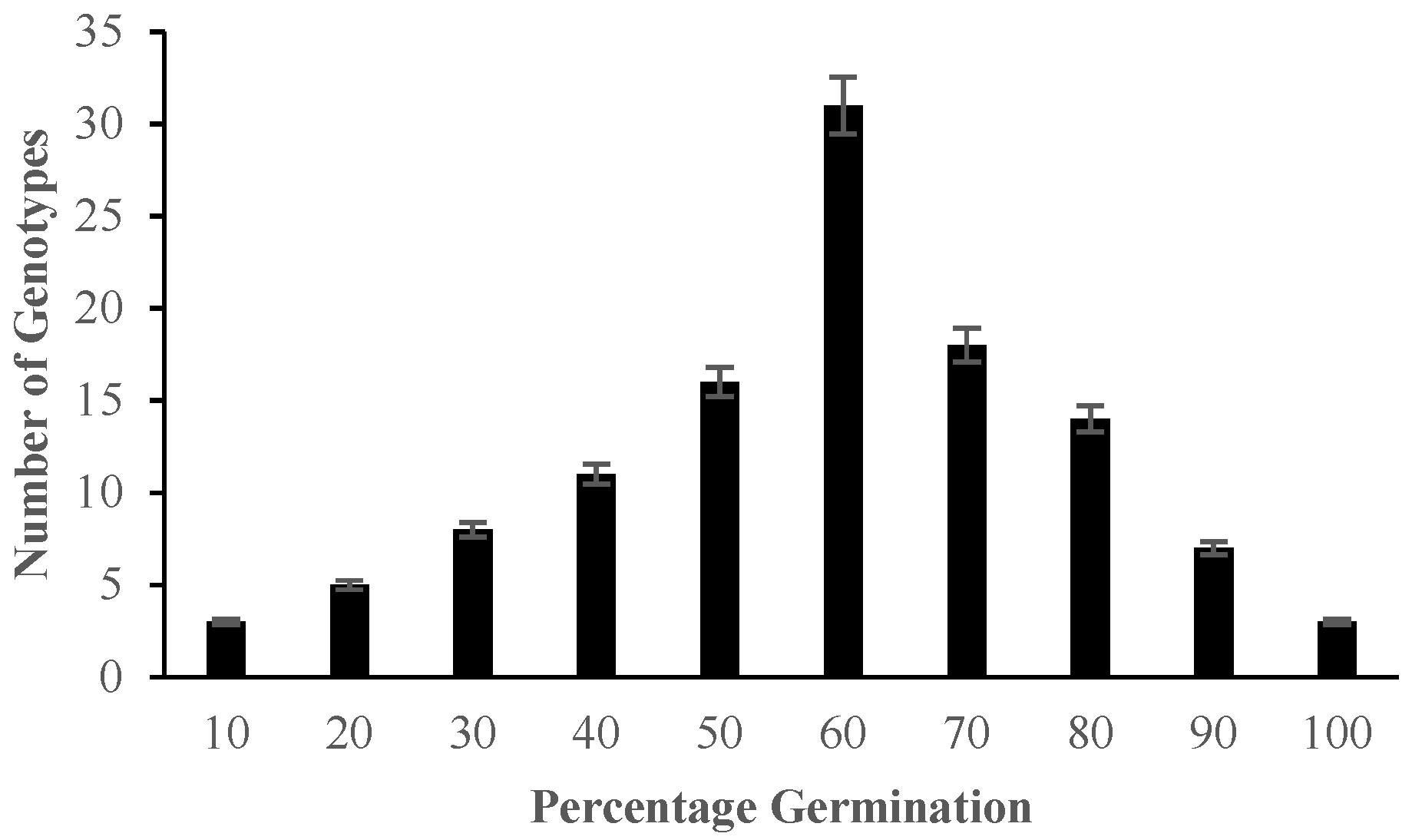
Figure 2.
Standard evaluation score (SES) of visual salt injury at seedling stage of 116 Asian rice cultivars.
Figure 2.
Standard evaluation score (SES) of visual salt injury at seedling stage of 116 Asian rice cultivars.
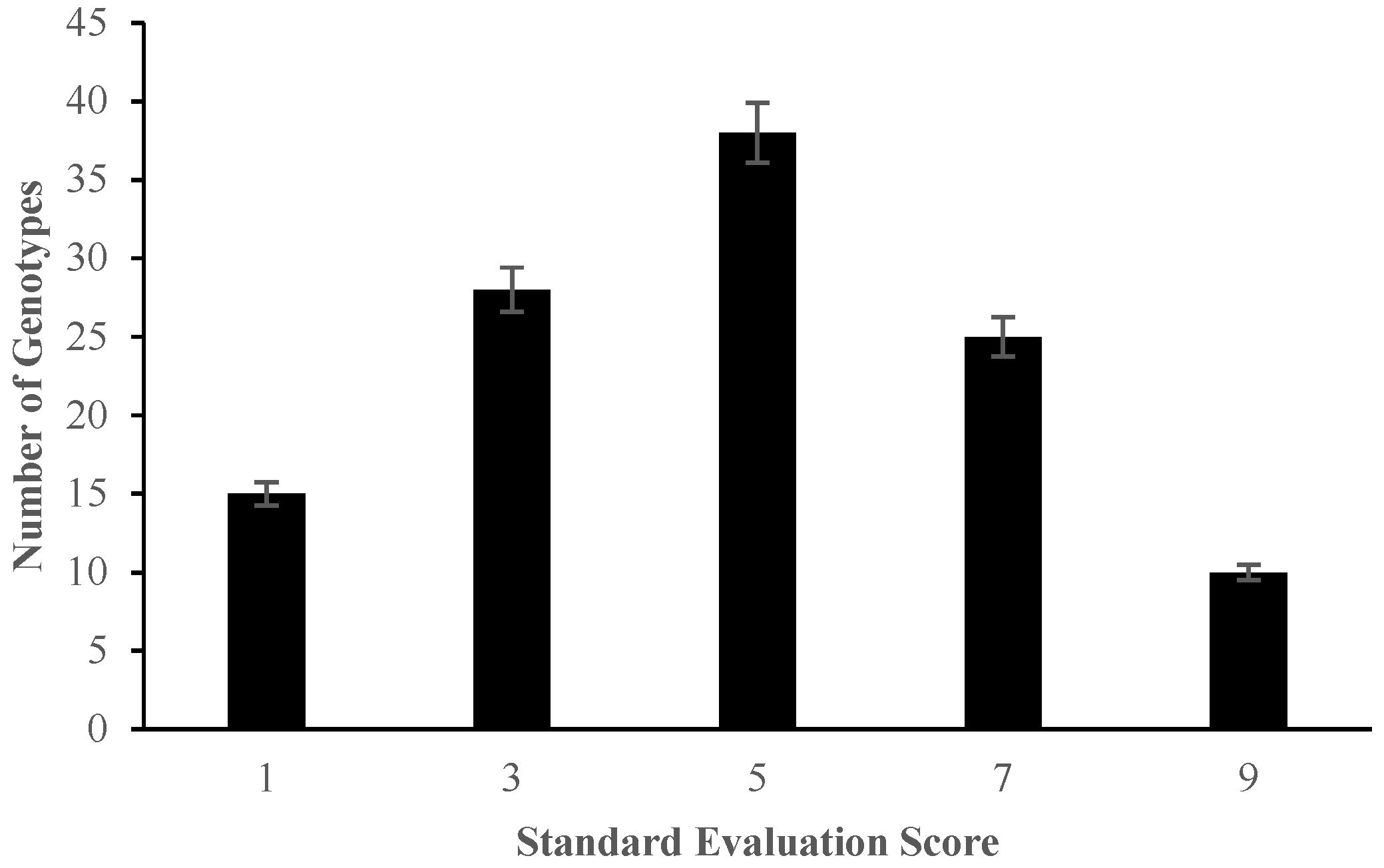
Figure 3.
Effect of salinity stress condition on root length (a), root fresh weight (b), shoot length (c), and plant biomass (d) of 116 Asian rice cultivars.
Figure 3.
Effect of salinity stress condition on root length (a), root fresh weight (b), shoot length (c), and plant biomass (d) of 116 Asian rice cultivars.
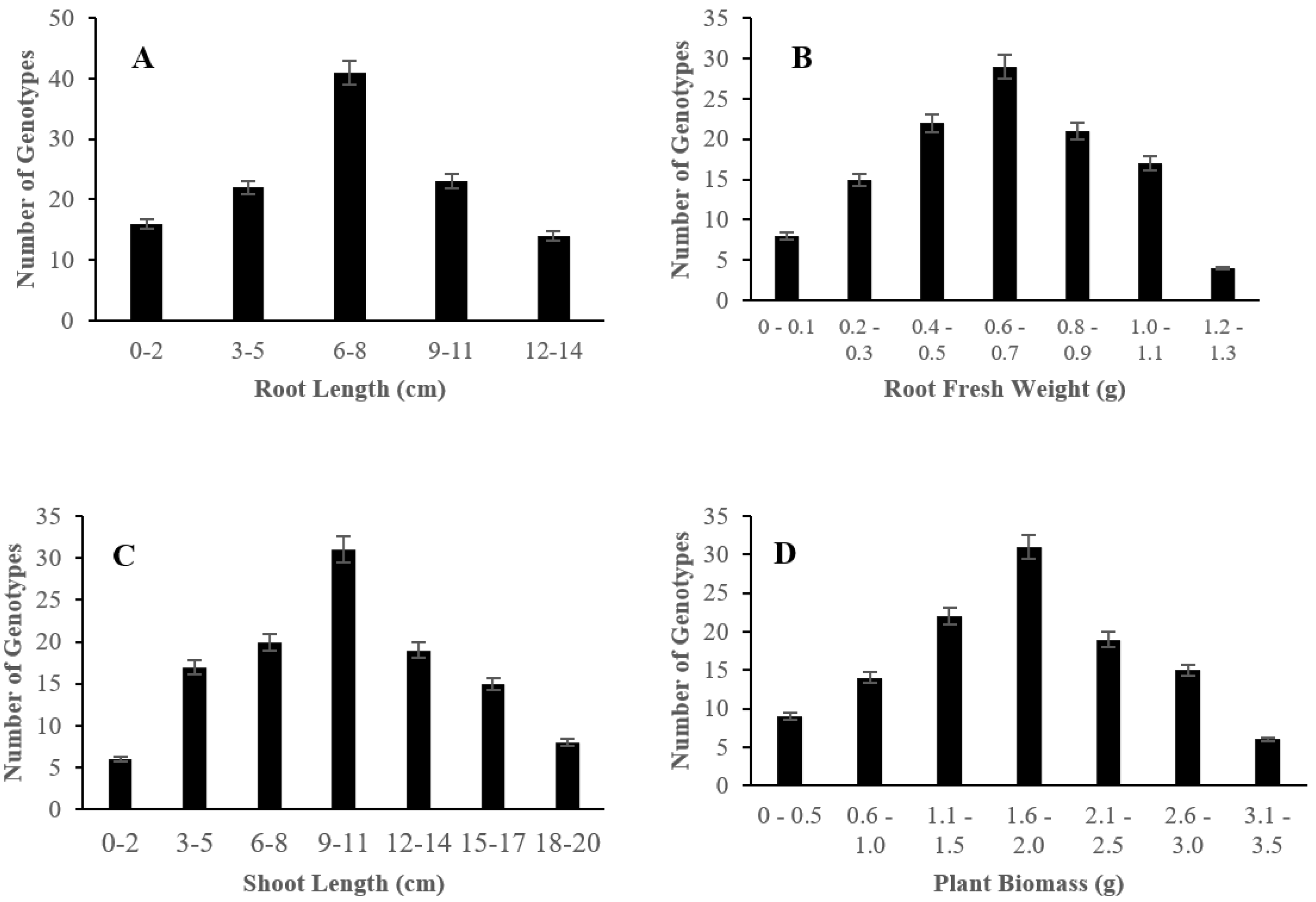
Figure 4.
Leaf symptoms of leaf rolling (a) and chlorophyll content (b) from 116 Asian rice cultivars under salinity stress condition.
Figure 4.
Leaf symptoms of leaf rolling (a) and chlorophyll content (b) from 116 Asian rice cultivars under salinity stress condition.
Figure 5.
Na/K ratio of 116 Asian rice varieties under salinity stress condition.
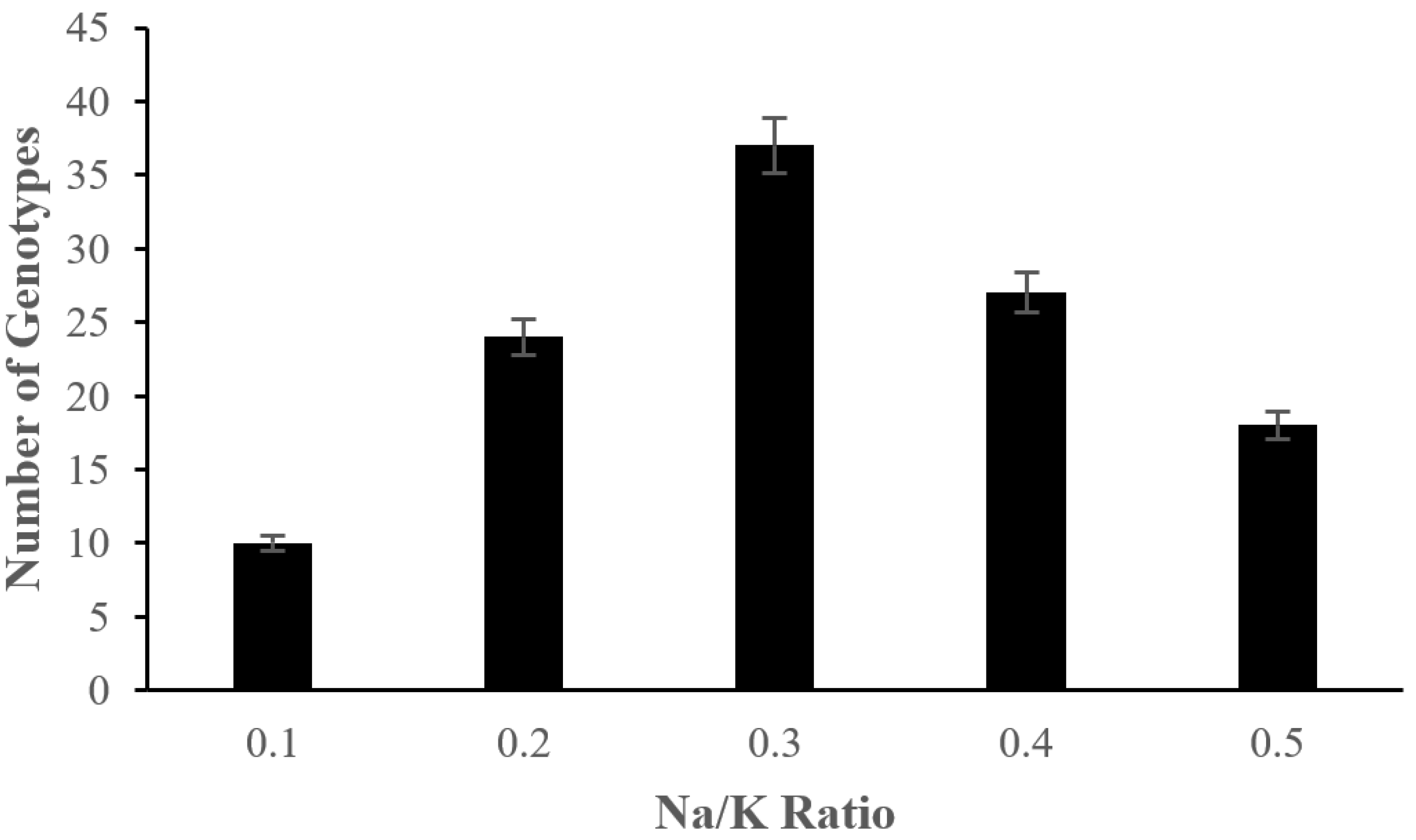
Figure 6.
Effect of salinity stress condition on cell membrane stability of 116 Asian rice varieties.
Figure 6.
Effect of salinity stress condition on cell membrane stability of 116 Asian rice varieties.
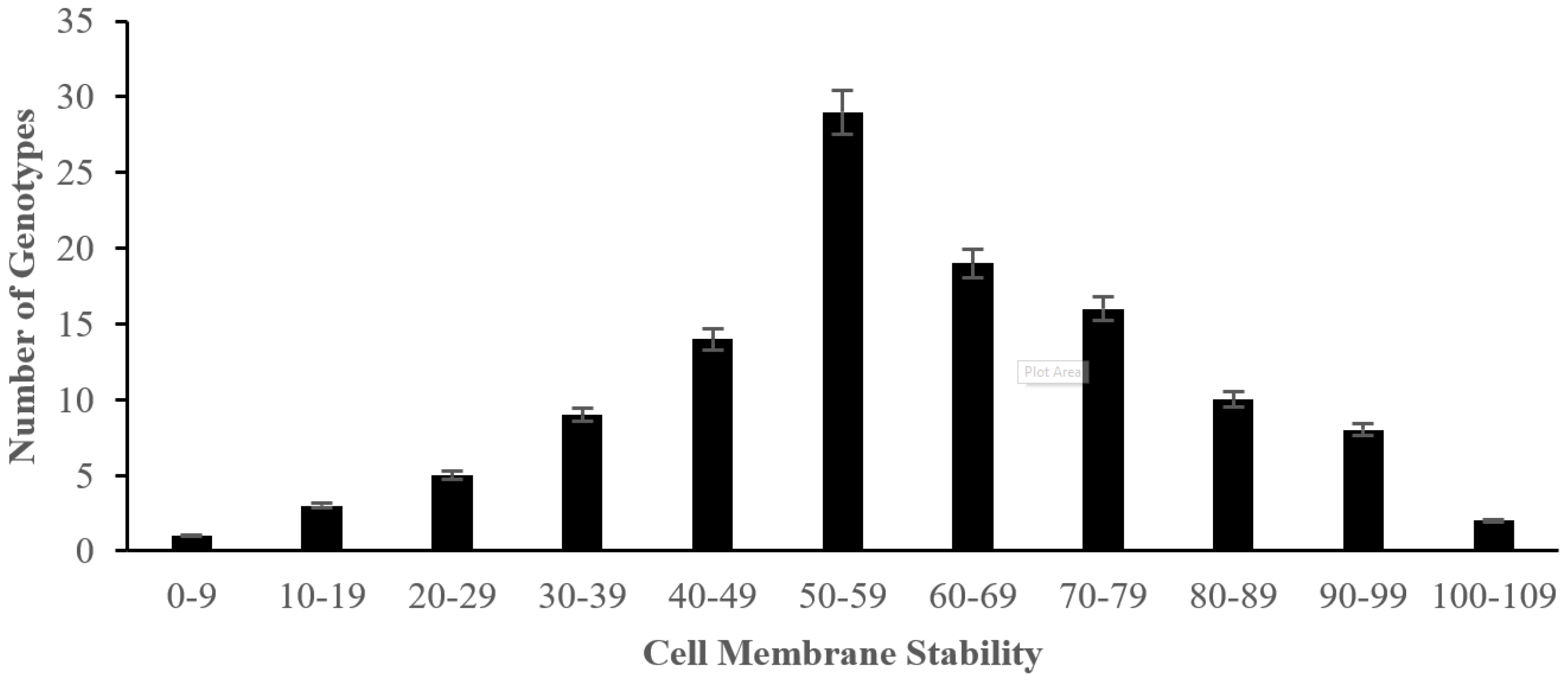
Figure 7.
Accumulation of proline (a), malondialdehyde (MDA) (b), hydrogen peroxide (H2O2) (c), sugar (d) in 116 Asian rice varieties under salinity stress condition.
Figure 7.
Accumulation of proline (a), malondialdehyde (MDA) (b), hydrogen peroxide (H2O2) (c), sugar (d) in 116 Asian rice varieties under salinity stress condition.
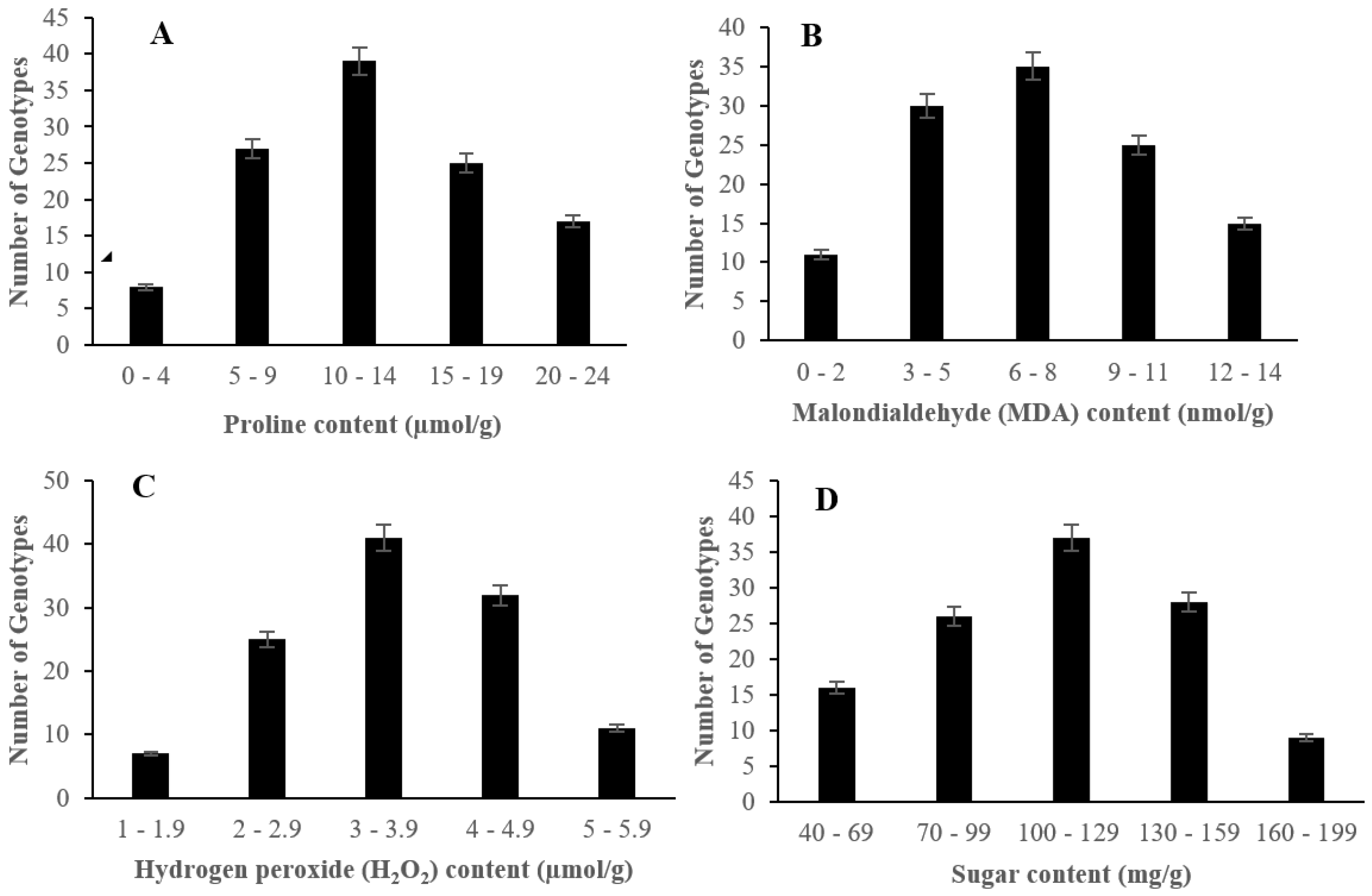
Table 1.
Rice varieties for salt stress tolerance screening.
| # | Rice Varieties | Country | Sub-population |
|---|---|---|---|
| 1 | Karang Serang | Indonesia | TRJ |
| 2 | Rojolele | Indonesia | TRJ |
| 3 | Cempo Ireng | Indonesia | TRJ |
| 4 | Ciherang | Indonesia | IND |
| 5 | Mayang Khang | Indonesia | IND |
| 6 | Sipirasikkam | Indonesia | TRJ |
| 7 | Mitak | Indonesia | TRJ |
| 8 | Dara | Indonesia | AUS |
| 9 | B805D-Mr-16-8-3 | Indonesia | IND |
| 10 | Tia Bura | Indonesia | TRJ |
| 11 | C 5560 | Thailand | TEJ/TRJ |
| 12 | Nam Dawk Mai | Thailand | IND |
| 13 | Bkn 6987-68-14 | Thailand | IND |
| 14 | Td 70 | Thailand | IND |
| 15 | Cntlr80076-44-1-1-1 | Thailand | IND |
| 16 | Nahng Sawn | Thailand | IND |
| 17 | Quinimpol | Philippines | TRJ |
| 18 | TCCP 266 | Philippines | IND |
| 19 | IR 4482-5-3-9-5 | Philippines | IND |
| 20 | IR 45427 | Philippines | IND |
| 21 | IR 9660-48-1-1-2 | Philippines | IND |
| 22 | IR 238 | Philippines | IND |
| 23 | IR 2061-214-2-3 | Philippines | IND |
| 24 | IR 2462 | Philippines | IND |
| 25 | IR 58614-B-B-8-2 | Philippines | IND |
| 26 | Pakkali | Philippines | ARO |
| 27 | IR64 | Philippines | IND |
| 28 | IR58 | Philippines | IND |
| 29 | IR29 | Philippines | IND |
| 30 | Taichu Mochi 59 | Taiwan | TRJ |
| 31 | Ai Chueh Ta Pai Ku | Taiwan | IND |
| 32 | Ragasu | Taiwan | TEJ/TRJ |
| 33 | Tobura | Taiwan | TEJ/TRJ |
| 34 | Kao Chio Lin Chou | Taiwan | IND |
| 35 | Taino 38 | Taiwan | IND/AUS |
| 36 | Nanton No. 131 | Taiwan | TRJ/(admix) |
| 37 | Hsin Hsing Pai Ku | Taiwan | IND |
| 38 | Tainung 45 | Taiwan | IND |
| 39 | Ao Chiu 2 Hao | China | IND |
| 40 | Chun 118-33 | China | IND |
| 41 | Kin Shan Zim | China | IND |
| 42 | Pan Ju | China | IND |
| 43 | Kechengnuo No. 4 | China | IND |
| 44 | 4484 | China | IND |
| 45 | 4595 | China | IND |
| 46 | You-I B | China | IND |
| 47 | Chunjiangzao No. 1 | China | TEJ |
| 48 | Shimizu Mochi | Japan | TEJ |
| 49 | Norin 11 | Japan | TEJ |
| 50 | Tamanishiki | Japan | TEJ |
| 51 | Niwahutaw Mochi | Japan | TEJ |
| 52 | Somewake | Japan | TEJ |
| 53 | A 5 | Japan | TEJ |
| 54 | C.B. Ii | Japan | AUS |
| 55 | Fujisaka 5 | Japan | IND |
| 56 | Nipponbare | Japan | TEJ |
| 57 | Khao Phoi | Laos | TEJ/TRJ |
| 58 | Khao Luang | Laos | TRJ/(admix) |
| 59 | Padi Pohon Batu | Malaysia | TRJ |
| 60 | Acheh | Malaysia | IND |
| 61 | Mahsuri | Malaysia | IND |
| 62 | Padi Tarab Arab | Malaysia | TRJ |
| 63 | Nc 1/536 | Pakistan | AUS |
| 64 | Red | Pakistan | AUS/(Admix) |
| 65 | Santhi 990 | Pakistan | IND/AUS |
| 66 | Daudzai Field Mix | Pakistan | AUS |
| 67 | Jp 5 | Pakistan | IND/AUS |
| 68 | Won Son Zo No. 11 | Korea | IND |
| 69 | Daegudo | Korea | TEJ |
| 70 | Guweoldo | Korea | TEJ |
| 71 | Namyang 7 | Korea | TEJ |
| 72 | Yong Chal Byo | Korea_ South | TEJ |
| 73 | Thang 10 | Vietnam | IND |
| 74 | Cm1_ Haipong | Vietnam | IND |
| 75 | Nang Bang Bentre | Vietnam | AUS |
| 76 | Lua Chua Chan | Vietnam | TRJ |
| 77 | Soc Nau | Vietnam | IND |
| 78 | Heo Trang | Vietnam | IND |
| 79 | Pd 46 | Sri Lanka | IND |
| 80 | Bakiella 1 | Sri Lanka | IND |
| 81 | Gallawa | Sri Lanka | AUS |
| 82 | Ittikulama | Sri Lanka | AUS |
| 83 | Karayal | Sri Lanka | AUS |
| 84 | Amane | Sri Lanka | IND |
| 85 | Thavalu | Sri Lanka | AUS |
| 86 | Patnai 6 | Myanmar | AUS |
| 87 | Buphopa | Myanmar | TEJ/TRJ |
| 88 | Kaukkyi Ani | Myanmar | TRJ |
| 89 | A100943-R | Myanmar | AUS |
| 90 | Nsgc 5953 | Myanmar | IND |
| 91 | A 36-3 | Myanmar | IND |
| 92 | Jumli Dhan | Nepal | TEJ/TRJ |
| 93 | N-2703 | Nepal | AUS |
| 94 | Bhim Dhan | Nepal | TEJ/TRJ |
| 95 | Juppa | Nepal | IND |
| 96 | Tauli | Nepal | AUS |
| 97 | Darmali | Nepal | TEJ/ARO |
| 98 | Dhan | Nepal | IND |
| 99 | Tchampa | Iran | AUS |
| 100 | Phudugey | Bhutan | AUS |
| 101 | Jyanak | Bhutan | TEJ/TRJ |
| 102 | Wir 3039 | Tajikistan | TEJ |
| 103 | Ak Tokhum | Azerbaijan | ARO |
| 104 | Gasym Hany | Azerbaijan | ARO |
| 105 | Celiaj | Azerbaijan | TEJ |
| 106 | Shimla Early | Iraq | IND/AUS |
| 107 | A 152 | Bangladesh | IND/TRJ |
| 108 | Dnj 179 | Bangladesh | AUS |
| 109 | Dj 24 | Bangladesh | AUS |
| 110 | Dj 102 | Bangladesh | AUS |
| 111 | Dnj 121 | Bangladesh | AUS |
| 112 | Aswina 330 | Bangladesh | AUS |
| 113 | Tranoeup Beykher | Cambodia | IND |
| 114 | Srav Prapay | Cambodia | IND |
| 115 | Simpor | Brunei | TRJ |
| 116 | Pokkali | India | IND |
* IND = indica; TEJ = temperate japonica; TRJ = Tropical japonica; ARO = aromatic; AUS = aus.
Table 2.
Standard evaluation score (SES) of visual salt injury at seedling stage [6].
Table 2.
Standard evaluation score (SES) of visual salt injury at seedling stage [6].
| Score | Observation | Tolerance |
|---|---|---|
| 1 3 5 7 9 |
Normal growth, no leaf symptoms Nearly normal growth, but leaf tips or few leaves whitish and rolled Growth severely retarded; most leaves rolled; only a few are elongating Complete cessation of growth; most leaves dry; some plants dying Almost all plants dead or dying |
Highly tolerant Tolerant Moderately tolerant Susceptible Highly susceptible |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



