
Submitted:
14 March 2023
Posted:
15 March 2023
You are already at the latest version
Abstract
Coastal lagoons are dynamic transitional water ecosystems hosting valuable biological communities, including rich and diverse macrophyte assemblages. Aquatic macrophytes must cope with large fluctuations of environmental conditions on a spatial and seasonal scale. Salinity is one of the most variable parameters, changing from nearly freshwater to hypersalinity, and it is known to have a strong influence on the composition and structure of macrophyte assemblages. This study is focused on the effect of salinity on macrophyte communities of the eight most important coastal lagoons of Apulia (south-eastern Mediterranean Sea). A set of eleven transitional water body types (sensu Water Framework Directive) were allocated in a range of meso- to hyperhaline lagoons. Macrophyte sampling was carried out between 2011 and 2019 and a total of 324 samples (18 sampling stations x 2 seasons x 9 years) was analyzed. Then, macrophyte occurrence in each transitional water body (T-WB) was expressed as frequency values (%) and assemblages were compared to assess any similarity in relation to four salinity classes (mesohaline, polyhaline, euhaline, hyperhaline). Species richness varied according to the salinity class, being much higher in polyhaline and euhaline T-WBs and strongly decreasing at the extremes of salinity range (mesohaline and hyperhaline T-WBs). Moreover, statistical analysis showed a high resemblance of macrophyte assemblages of T-WBs within the same salinity class, which shared a great number of species. Four distinct macrophyte communities were distinguished, reflecting the salinity conditions of different T-WB types and confirming the effectiveness of a lagoon typology based on this descriptor.
Keywords:
transitional water ecosystems
; coastal lagoons
; biodiversity
; typology
; aquatic vegetation
; abiotic factors
; benthic communities
; conservation
; environmental variability
1. Introduction
Transitional waters, including coastal lagoons, saline lakes, river estuaries and deltas, are dynamic and heterogeneous ecosystems placed at the interface between water and terrestrial environments receiving variable amounts of freshwater [1,2]. Due to their geomorphological and hydrological features, such as the shallow depth, partial confinement, limited extension, variation of freshwater inputs and the exchange flows with marine waters, these environments frequently undergo strong fluctuations in physical and chemical parameters such as salinity, temperature, turbidity, dissolved oxygen, and pH [3]. Moreover, transitional waters are characterized by high trophic inputs and rapid biogeochemical cycles, being highly sensitive to eutrophication events [4,5,6]. Such conditions select peculiar macrophyte communities well-adapted to the variability of abiotic factors [7]. Besides opportunistic species, these ecosystems are naturally colonized by aquatic angiosperms and macroalgae with remarkable ecological value [8,9]. Submerged macrophytes represent important primary producers and many species are considered ecosystem engineers, providing several ecosystem goods and services [10], including substrate stabilization, erosive processes reduction [11], acidification and eutrophication mitigation, as well as waters and atmosphere oxygenation [12]. Healthy macrophyte associations host rich and diverse communities of aquatic invertebrates, fish and birds which rely on these habitats as spawning, nursery, feeding, and refuge areas [13,14]. Among transitional waters, coastal lagoons are listed as priority habitats in Annex I of the Habitat Directive (92/43/EEC) [15] and are protected by several national and international agreements (Natura 2000 network; Ramsar convention). Hence, conservation and sustainable management of such habitats are strongly advocated [16]. Moreover, macrophytes can be used as ecological indicators to assess the quality of water bodies due to their ability to respond to environmental changes [17,18,19,20]. As a consequence, they are among the biological elements proposed in the European Water Framework Directive (WFD; 2000/60/CE) [21] as quality elements for the classification of the ecological status of transitional water bodies (T-WBs). In Italy, the use of the Macrophyte Quality Index (MaQI) [22,23,24] is currently compulsory under national law (Italian Ministerial Decree 260/2010).
Although the crucial role of macrophytes in transitional waters is well-known, there is still scarce knowledge about the species’ ecological traits and their responses to the main abiotic factors. Salinity is known to be one of the most variable parameters in T-WBs, and it can be a key factor in modelling the biodiversity of macrophyte assemblages [7,25,26,27]. Some T-WBs are nearly fresh, with mean salinity below 1 PSU [28], while hypersaline transitional waters can reach mean salinity values of 75 PSU [29,30]. Moreover, within the same T-WB, salinity can also show a great range of variability during the year, even reaching a difference of 100 PSU between winter and summer [31]. The presence of a strong genetic adaption of species to salinity regimes of transitional waters has been demonstrated by previous studies, proving the crucial role of this abiotic factor in determining macrophyte distribution [32,33,34]. However, there is still a lack of knowledge about the presence of possible analogies in macrophyte species composition of T-WBs with similar salinity ranges.
In the Mediterranean ecoregion, four types of transitional waters (estuaries, deltas, microtidal and non-tidal coastal lagoons) are present and, according to WFD, a typological scheme was defined by each country. Coastal lagoons were distinguished on the base of their shape (as surface area), tidal range and salinity, as indicated in System B of WFD, in the Italian lagoon typology [35]. Salinity thresholds were derived from the Venice System [36]. In particular, the Italian lagoon types were classified on two levels of tidal range (> or < 50 cm), two of surface area (>2.5 km2 or between 0.5 and 2.5 km2) and five of water salinity (oligohaline: <5 PSU, mesohaline: 5-20 PSU, polyhaline: 20-30 PSU, euhaline: 30-40 PSU, hyperhaline: >40 PSU) [35,37], according to the indications given by the European Common Implementation Strategy (CIS) working groups (CIS Guidance n. 5).
In the Mediterranean context, more than 130 transitional water systems are present in Italy, with a surface area greater than 1,700 km2 [38]. The largest coastal lagoons are located along the northern Adriatic coasts, but the highest number is concentrated in the southern part of the Italian peninsula. Our work is focused on the salinity influence on macrophyte species composition of the eight most important and wider transitional basins of the Apulian region (south of Italy) [39]. The study falls within the monitoring and assessment of the ecological status of T-WBs by MaQI, as required by Italian legislation (Italian Ministerial Decree 260/2010). All the investigated T-WBs are located in non-tidal coastal lagoons of small and medium size, distributed into four salinity classes, from mesohaline to hyperhaline [40]. We assessed the presence of any relation among macrophyte assemblages in T-WBs of the same salinity class, as well as their species richness, aiming at proving the role of salinity in structuring macrophyte communities. The validity of the Apulian lagoon typology, mainly based on the salinity descriptor, was also checked as a case study in the Mediterranean ecoregion.
2. Materials and Methods
The study was based on the identification and classification of the coastal lagoons of Apulia following the current Italian legislation (Italian Ministerial Decree 131/08). According to it, the investigated area included eight lagoons, two for each salinity class: Cesine (CE) and Torre Guaceto (TG) classified as mesohaline, Lesina (LE) and Varano (VA) as polyhaline, Porto Cesareo (PC) and Mar Piccolo (MP) as euhaline, Margherita di Savoia, only for the part known as “Vasche Evaporanti – Lago Salpi” (LS), and Punta della Contessa (PU) as hyperhaline (Figure 1). A total of 11 T-WBs were considered. In almost all cases, each lagoon comprised a single T-WB, with the exception of LE, divided into three different T-WBs (LE1, LE2, LE3) and MP divided into two T-WBs corresponding to two well-distinct inlets, called First and Second Inlet (MP1 and MP2, respectively).
Macrophyte sampling was carried out from 2011 to 2019, during spring and autumn, for a total of 18 seasonal surveys. Sampling stations were distributed taking into account morphology, spatial heterogeneity and extension of each T-WB (Figure 1; Table 1). Only one station was chosen as representative of the hyperhaline basins (PU and LS), which are choked lagoons with homogeneous ecological conditions. Two stations were sampled in TG, a small basin almost entirely dominated by Chara baltica (Hartman) Bruzelius. Three stations were placed in CE, inside the main perennial pond called Pantano Grande (0.68 km2). Four stations were selected to cover PC, a bay in open connection with the sea, while, to characterize the larger basins of LE, VA and MP a higher number of stations was required (ten, eight and seven, respectively). In detail, for the different T-WBs of LE and MP, it ranged between three (in LE2, LE3, MP1) and four stations (in LE1 and MP2). The southern part of VA was not investigated due to the lack of vegetation (Bottalico, pers. obs.) In total, 324 samples (18 stations x 2 seasons x 9 years) were analyzed. Samples were collected according to the monitoring protocols by the Italian Institute for Environmental Protection and Research [41] and as required by MaQI application [22,23]. Macrophytes were preserved in 4% buffered formalin/seawater and determined at specific and intra-specific levels in the laboratory by means of a Leica MZ 7.5 stereo-microscope (Leica, Wetzlar, Germany) and a light microscope Olympus BX-40 (Olympus, Melville, USA). For morphological observations, sections of thalli were obtained by free-hand cutting or with a DSK-1000 vibratome (Dosaka, Kyoto, Japan). Some samples of uncertain identification were also preserved in silica gel for molecular analyses. The up-to-date nomenclature of the identified taxa followed AlgaeBase [42].
Macrophyte occurrence in each water body was expressed as frequency values (%), representing the ratio between the number of samplings where the species were found and the total number of samplings carried out in a given T-WB. Then, macrophyte assemblages were compared to assess any relation between species presence and abundance with salinity classes. To test the salinity influence on macrophyte communities, Cluster Analysis and non-Metric Multidimensional Scaling (n-MDS) were performed using the software Past 4.03, based on the Bray-Curtis dissimilarity matrix. The non-parametric ANOSIM (Analysis of Similarities) was tested to verify the data significance (p-values and R-values). Then, the SIMPER test (Similarity Percentages) was run using the software Primer 5 (version 5.2.9) to evaluate the contribution of each species (%) to the similarity among T-WB groups identified by clustering (cut-off at 90%). The Species Richness index (S), representing the total number of species counted in each T-WB, was also calculated.
3. Results
A total of 171 species belonging to Magnioliophyta (3.5%), Charophyta (3.5%), Ochrophyta-Phaeophyceae (8%), Chlorophyta (33%) and Rhodophyta (52%) were identified (Table 2). A considerable part of the species was only sporadically found in one or few T-WBs, with very low frequency. On the other hand, several species of Chlorophyta, mainly Ulvales and Cladophorales, stably colonized different T-WBs. Some taxa also resulted to be very abundant in certain circumstances, acting as structuring species. This is the case of some species of phanerogams, such as Cymodocea nodosa (Ucria) Ascherson, Ruppia spiralis Linnaeus ex Dumortier and Zostera noltei Hornemann, as well as perennial brown algae, such as Gongolaria barbata (Stackhouse) Kuntze, which colonized wide areas in the presence of salinity values suiting with their ecological traits.
Species Richness (S) of macrophyte assemblages varied according to the salinity class of T-WBs (Figure 2a,b). Salinity values at the end of the range generally reflected a scarce macrophyte diversity, as in the case of the mesohaline T-WBs of TG and CE and the hyperhaline PU. LS, likewise hyperhaline, showed a moderate macrophyte diversity mainly represented by opportunistic species. On the other hand, macrophyte assemblages proved to be much more diverse in polyhaline (LE1, LE2, LE3 and VA) and euhaline T-WBs (MP1, MP2 and PC).
Cluster Analysis showed that T-WBs within the same salinity class had remarkable analogies in macrophyte species composition and frequency, forming four well-distinct groups with a high level of similarity, especially in the case of hyperhaline, mesohaline and polyhaline T-WBs (Figure 3a). On a smaller scale, Cluster Analysis showed the existence of two groups of T-WBs. In fact, clustering highlighted a certain similarity (~17%) between macrophyte assemblages occurring at both the ends of the salinity range (mesohaline and hyperaline T-WBs), as well as between those of polyhaline and euhaline T-WBs (similarity ~16%). The n-MDS confirmed the presence of four distinct groups corresponding to the four salinity classes, with a stress value of 0.14 (Figure 3b).
The role of the salinity regime in influencing macrophyte composition was confirmed by ANOSIM, which gave back a Global R-value of ~0.95 and a p-value of 0.002, attesting to highly significant differences among assemblages (Table 3). However, although pairwise ANOSIM revealed very high R-values, only the comparison between euhaline and polyhaline T-WBs showed to be strictly significant (p-value < 0.005), probably because of the scarce number of species occurring in hyperaline and mesohaline T-WBs, as well as due to the presence of several eurytherm and euryhaline species found in most of the analyzed T-WBs.
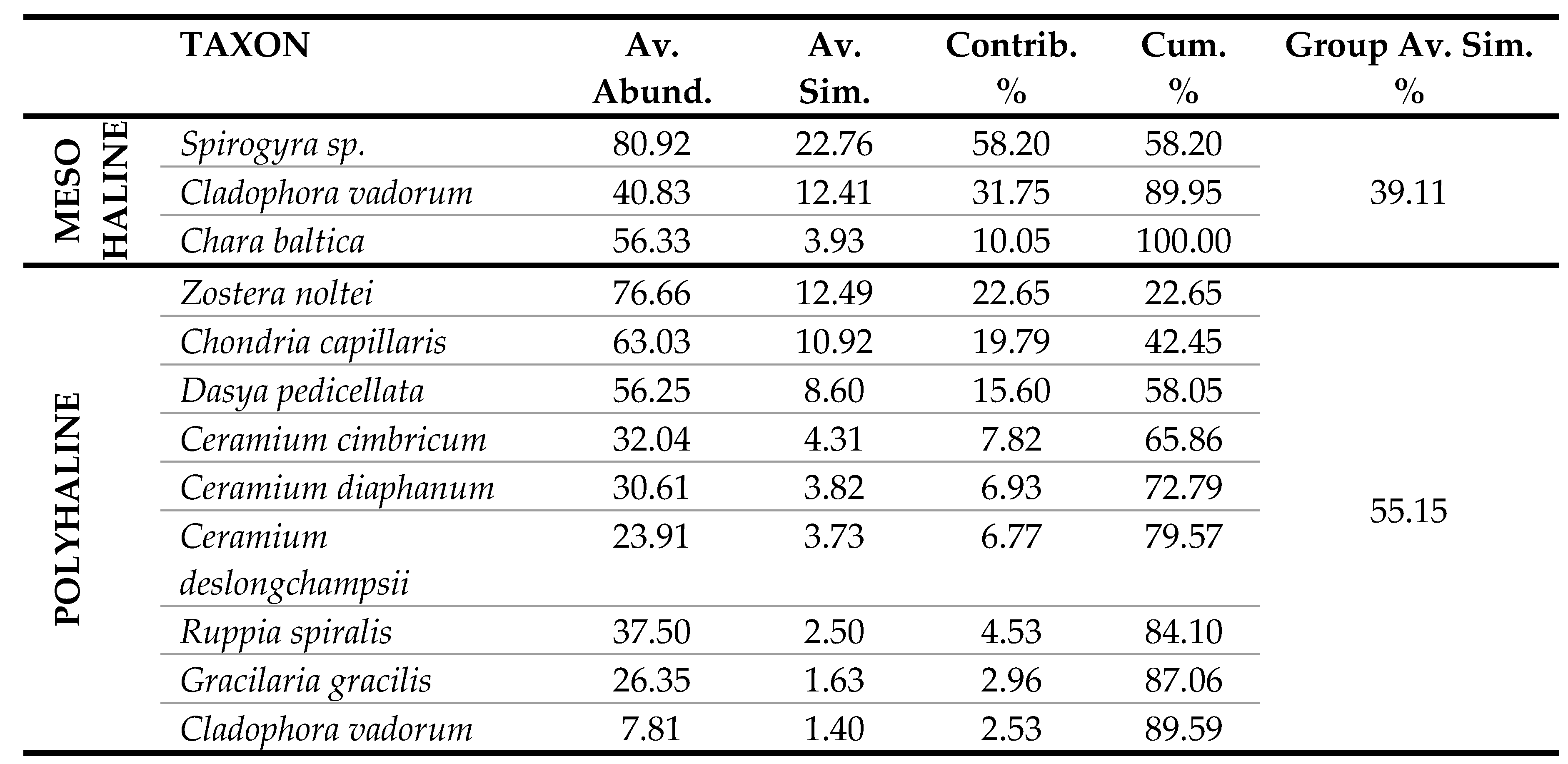
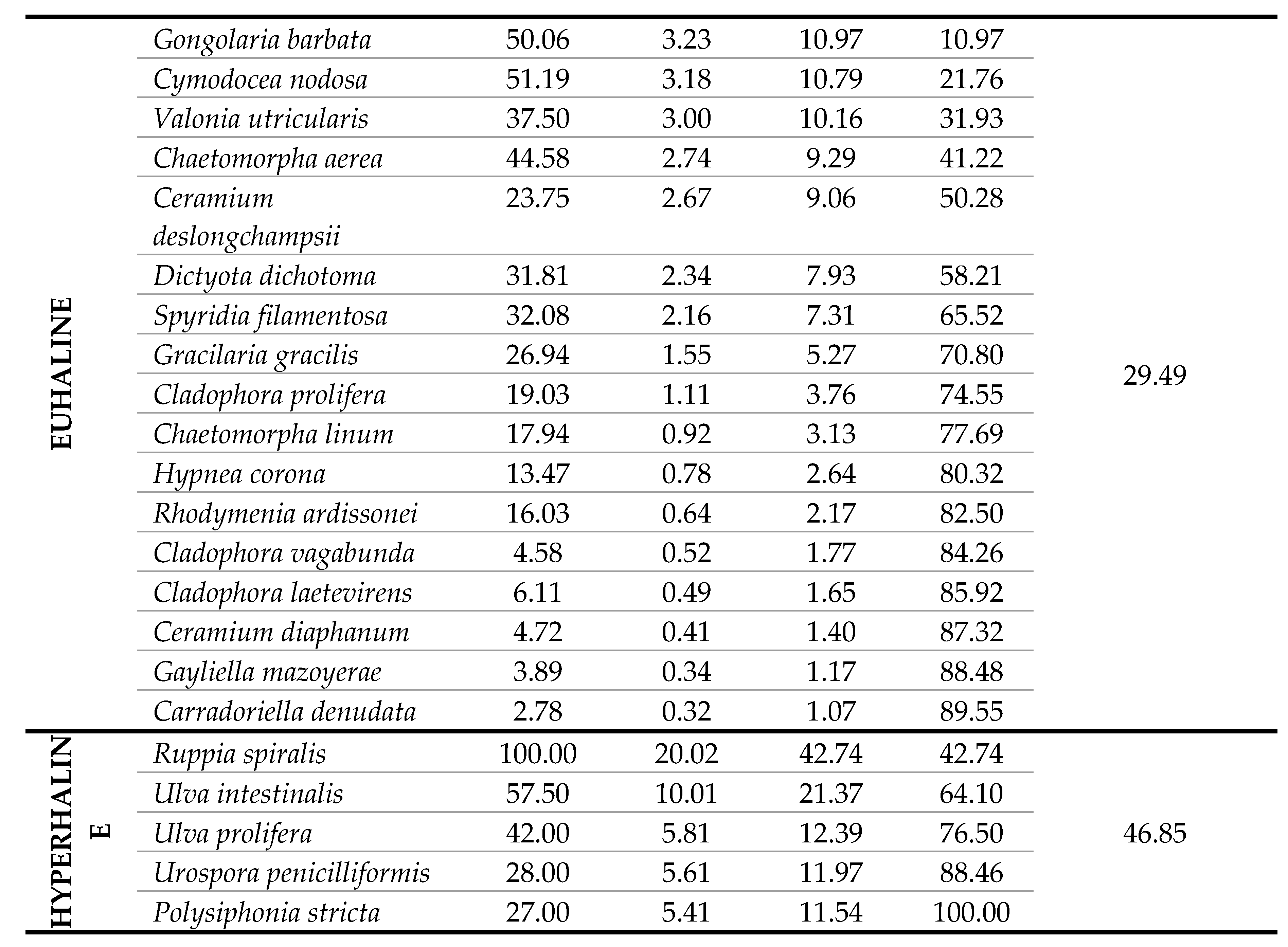
The polyhaline T-WBs of LE1, LE2, LE3 and VA showed the highest Group Average Similarity (55.15%), highly resembling in terms of species composition and frequency (Table 4). The hyperaline T-WBs of LS and PU showed a high Group Average Similarity of 46.85%, given by the restricted number of species. The mesohaline T-WBs of CE and TG shared few taxa with a Group Average Similarity of 39.11%, while the euhaline T-WBs, including PC, MP1 and MP2, showed a great number of common species but with a Group Average Similarity of 29.49%. Macrophyte composition and species mean abundance of the mesohaline T-WBs type was dominated by Charophyta. In fact, almost 70% of the similarity was due to the contribution of a species of Spirogyra (only identified at a generic level) and C. baltica, both characteristic of lower salinity waters. About 32% of this similarity was contributed by Cladophora vadorum (Areschoug) Kützing, a species with a wide distribution in lagoonal environments. Macrophyte composition of the polyhaline T-WBs type was dominated by the angiosperm Z. noltei and by different species of Ceramiales such as Chondria capillaris (Hudson) M.J. Wynne, Dasya pedicellata (C. Agardh) C. Agardh and various Ceramium spp. which, together, contributed up to about 80% of the total similarity within this type. In the hyperaline T-WBs type, macrophyte composition was dominated by R. spiralis and two species of Ulvales, Ulva intestinalis Linnaeus and U. prolifera O.F. Müller, reaching a total of 76.5% of the similarity within this type. Macrophyte composition of the euhaline T-WBs type was composed by an array of heterogeneous taxa, not only typical of marine waters, such as some brown algae belonging to Fucales and Dictyotales, but also more ubiquitous species, mainly belonging to Ceramiales and Cladophorales (Table 4).
4. Discussion
The definition of surface T-WB types is the first milestone in the implementation of the WFD (Section 1.2, Annex II) [21] to enable the establishment of appropriate reference conditions and to make a valid assessment of the ecological status of T-WBs. However, at the European level, many difficulties have been encountered in defining a general typology for transitional waters [43]. In particular, in the Mediterranean ecoregion, typologies based on different descriptors have been proposed e.g., [37,43,44,45,46,47], but a common consensus is still lacking. An approach based on the composition of macrophyte assemblages was used in this study to test the validity of the Apulian transitional water types, considering that they are representative of other Mediterranean ecosystems. Our results highlighted that a typology based on salinity ranges accounts for the variability of macrophyte assemblages, as required by WFD. Salinity proved to be the main environmental factor driving both the distribution and the composition of macrophyte communities, confirming previous observations in other areas [7,10,48,49,50,51,52,53,54,55,56]. In fact, although several geomorphological and hydrological factors can also play a determinant role in modelling macrophyte assemblages [57,58], salinity regime well-explained the variability found in the T-WBs along the gradient of salinity. This agrees with Pèrez-Ruzafa et al. [1], which had also identified the salinity regime as an influential factor, highly affecting sessile organisms such as macrophytes. Despite the differences in morphology, depth and extension, as well as spatial distances among T-WBs, those within the same salinity class showed comparable macrophyte communities in most of the cases. The mesohaline community was mainly represented by the Charophyta Spirogyra sp. and C. baltica, which showed high selectivity for the mesohaline T-WBs (TG and CE), being only found in both the basins although they represent quite different environments in terms of physico-chemical features. In particular, the genus Spirogyra confirmed its preference for nearly freshwater habitats [59], while C. baltica showed the ability to tolerate brackish waters with salinity up to 15 PSU [60].
On the other hand, geomorphologically and hydrologically similar coastal lagoons, but of different salinity classes, showed significant differences in their macrophyte composition and once again salinity played a crucial role in influencing macrophyte assemblages, as well as in the classification of lagoon types. The polyhaline community was represented by the dominance of Z. noltei, a species considered euryhaline [61], which formed monospecific beds or coexisted with R. spiralis. This angiosperm also characterized macrophyte assemblages of meso- to polyhaline waters of the Rodia lagoon in Western Greece [27]. The macrophyte community of the euhaline type was mainly represented by the euryhaline C. nodosa [51] and other typical marine species, such as the brown algae G. barbata and Dictyota dichotoma (Hudson) J.V. Lamouroux, the green Valonia utricularis (Roth) C. Agardh and the red Spyridia filamentosa (Wulfen) Harvey, well adapted to a salinity range which is almost the same of the open sea. Among the most abundant species, G. barbata and C. nodosa proved to be widely spread in the euhaline T-WBs (PC, MP1 and MP2), forming secondary substrates for several species and enhancing biodiversity. Even though harboring some common marine taxa, the euhaline coastal lagoons examined in this study displayed the lowest average similarity and some differences in macrophyte spectra composition that can be related to morphological aspects, such as the shape of the lagoons, the depth of the basins and the sea/lagoon exchange features. For instance, PC is an almost elliptical bay (2 km2), only partially delimited by a narrow peninsula which leaves a permanent connection with the open sea through a channel system about 700 m wide; its average depth is about 1 m [62]. MP is over ten times larger than PC, with a surface area of 20.72 km2, but it is a semi-enclosed basin divided into First Inlet (MP1 T-WB) and Second Inlet (MP2 T-WB), with a maximum depth of 12 and 8 m, respectively. However, only the First Inlet is in communication with the Ionian Sea through two channels, thus causing a reduced water exchange [63]. Effectively, PC was almost exclusively colonized by marine taxa, most of which shared with the adjacent open sea assemblages [64], indicating constant species recruitment from these habitats. On the contrary, MP1 and, especially, MP2 were also inhabited by species of more confined environments, such as Chaetomorpha aerea (Dillwyn) Kützing and Ulva rigida C. Agardh. The macrophyte community characterizing the hyperhaline type was mainly represented by R. spiralis. The genus Ruppia has long suffered a chaotic taxonomy, but recently the three species R. maritima Linnaeus, R. cirrhosa (Petagna) Grande and R. spiralis Linnaeus ex Dumortier have been considered as fully independent taxa [65]. These authors underlined that many Mediterranean records of R. cirrhosa can actually be referred to R. spiralis, a more common European species occurring in brackish habitats. Our specimens matched the specific characteristics of this species; therefore, they are here reported as R. spiralis. This species can be considered as the most “marine” Ruppia in Europe [65], with broader salt tolerance. In addition to the main species characterizing the different macrophyte communities, some opportunistic taxa revealed to be well adapted to salinity variations and occurred in all the salinity ranges. They mainly belonged to the orders Ulvales and Cladophorales (Chlorophyta) and Ceramiales (Rhodophyta).
In transitional waters, other factors, such as trophic status and water oxygenation may drive the macrophyte community structure [7]. However, the TWEAM multi-index method for the eutrophication assessment in these ecosystems [66] classified over 60% of the water bodies examined in this study in oligothropic status, except for LE1, MP1, MP2 and TG which resulted mesotrophic.
Species Richness index was closely associated with salinity. Macrophyte diversity was the highest in euhaline T-WBs, which had salinity levels similar to marine waters, started to decrease in polyhaline T-WBs and reached minimum values at the two ends of the salinity range. This declining trend in the number of taxa, depending on too low or too high salinity, was already reported in Mediterranean coastal lagoons [19,52,67] and in the Baltic Sea [56,68] and it was confirmed in the present study. The lowest species richness recorded in mesohaline and hyperhaline T-WBs, where salinity was quite far from marine waters, reflected stress conditions, with fewer species tolerating these extremes. The strongest reduction was observed in Rhodophyta, for which the number of species consistently dropped along the salinity gradient (75 in euhaline T-WBs vs 5 and 2 in hyperhaline and mesohaline T-WBs, respectively), with a reduction between 93 and 97%. The percentage of decrease in species number was still quite high in the Ochrophyta-Phaeophyceae (75–88%) and the lowest was displayed by Chlorophyta (49–77%). This pattern was also evident at order level. For example, some typical marine taxa, belonging to Corallinales or Gelidiales orders, not only were found in euhaline T-WBs but were also able to tolerate the lower salinity of polyhaline T-WBs; they completely disappeared in mesohaline and hyperhaline T-WBs, that were almost exclusively inhabited by opportunistic Ceramiales. The salinity ranges in these environments prevented large colonization by red algae and favoured the settlement of Chlorophyta, mainly Cladophorales and Ulvales, the best adapted to low-salinity and hyper-saline conditions [69,70]. Among Ochrophyta-Phaeophyceae, likewise, well-structured Dictyotales and Fucales species were found in euhaline and polyhaline T-WBs, while very few opportunistic species of Ectocarpales were the only present in the other T-WBs.
5. Conclusions
Macrophyte assemblages of T-WBs showed to be influenced by salinity both in terms of species composition and richness, testifying to the crucial role of this abiotic factor in modelling such communities. However, even though the macrophyte variability found in T-WBs was well-explained by the salinity regime, other abiotic factors may contribute to structuring these assemblages and further studies should be addressed to understand their importance in influencing macrophyte presence and distribution.
This study also contributed to implementing the knowledge about the macrophyte assemblages of the most important T-WBs of the Apulian region, whose biodiversity was still scarcely known in most of the cases. The assessment of macrophyte diversity and the structure of their assemblages in transitional waters is a key point for the development and implementation of management strategies, which represent one of the main goals of WFD. At the same time, the periodic monitoring of the conservation status of these ecosystems, based on biological indicators such as macrophytes, allows to understand the effectiveness of such management actions, with the aim to reach and/or maintain the Good Ecological Status claimed by WFD.
Author Contributions
Conceptualization, A.T., F.M., and A.B.; methodology, A.T., A.L., F.M., N.U. and A.B.; software, A.T, F.M. and A.B.; validation A.T, F.M., G.C. and A.B.; formal analysis, A.T, A.L., F.M., N.U. and A.B.; investigation, A.L., N.U and A.B.; resources, A.B.; data curation, A.T, F.M. and A.B.; original draft preparation, A.T. and A.B.; writing—review and editing, A.T., A.L., F.M., G.C., N.U and A.B; visualization, A.T and A.B; supervision, A.B., project administration, A.B., funding acquisition, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Apulian Regional Agency for the Environmental Prevention and Protection, Research Agreement “Monitoring of Biological Quality Elements (BQEs) macroalgae and phanerogams in the Apulian transitional waters. CUP: B31G22000110002.
Institutional Review Board Statement
Not Applicable.
Data Availability Statement
The dataset generated during and/or analyzed for the current study is available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M.; Pérez-Marcos, M. Coastal lagoons: “transitional ecosystems” between transitional and coastal waters. J. Coast. Conserv. 2011, 15, 369–392. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Pérez-Ruzafa, I.M.; Newton, A.; Marcos, C. Coastal lagoons: environmental variability, ecosystem complexity and goods and services uniformity. In Coasts and Estuaries, the Future; Wolanski, E., Day, J., Elliott, M., Ramesh, R., Eds.; Elsevier: New York, USA, 2019; pp. 253–276. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Sigovini, M.; Ghirardini, A.V. A review of terms and definitions to categorize estuaries, lagoons and associated environments. Mar. Freshwater Res. 2009, 60, 497–509. [Google Scholar] [CrossRef]
- Petihakis, G.; Triantafyllou, G.; Koutsoubas, D.; Allen, I.; Dounas, C. Modelling the annual cycles of nutrients and phytoplankton in a Mediterranean lagoon (Gialova, Greece). Mar. Environ. Res. 1999, 48, 37–58. [Google Scholar] [CrossRef]
- Plus, M.; Chapelle, A.; Lazure, P.; Auby, I.; Levavasseur, G.; Verlaque, M.; Belsher, T.; Deslous-Paoli, J.-M.; Zaldívar, J.-M.; Murray, C.N. Modelling of oxygen and nitrogen cycling as a function of macrophyte community in the Thau lagoon. Cont. Shelf Res. 2003, 23, 1877–1898. [Google Scholar] [CrossRef]
- Pastres, R.; Solidoro, C.; Ciavatta, S.; Petrizzo, A.; Cossarini, G. Long-term changes of inorganic nutrients in the Lagoon of Venice (Italy). J. Marine Syst. 2004, 51, 179–189. [Google Scholar] [CrossRef]
- Le Fur, I. .; Wit, R.D.; Plus, M.; Oheix, J.; Simier, M.; Ouisse, V. Submerged benthic macrophytes in Mediterranean lagoons: distribution patterns in relation to water chemistry and depth. Hydrobiologia 2018, 808, 175–200. [Google Scholar] [CrossRef]
- Viaroli, P.; Bartoli, M.; Giordani, G.; Naldi, M.; Orfanidis, S.; Zaldivar, J.M. (2008). Community shifts, alternative stable states, biogeochemical controls and feedbacks in eutrophic coastal lagoons: a brief overview. Aquat. Conserv. 2008, 18, S105–S117. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Ghetti, P.F. Rapid Quality Index (R-MaQI), based mainly on macrophyte associations, to assess the ecological status of Mediterranean transitional environments. Chem. Ecol. 2007, 23, 493–503. [Google Scholar] [CrossRef]
- Christia, C.; Tziortzis, I.; Fyttis, G.; Kashta, L.; Papastergiadou, E. A survey of the benthic aquatic flora in transitional water systems of Greece and Cyprus (Mediterranean Sea). Bot. Mar. 2011, 54, 169–178. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Marcomini, A. Sedimentation rates and erosion processes in the lagoon of Venice. Environ. Int. 2005, 31, 983–992. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Agostini, S.; Marchand, B.; Pergent, G. Temporal and spatial changes of seagrass meadows in a Mediterranean coastal lagoon. Oceanol. Acta 2003, 25, 297–302. [Google Scholar] [CrossRef]
- EEC. Council Directive 92/43/EEC on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Comm. July 1992. [Google Scholar]
- Garrido, J.; Pérez-Bilbao, A.; Benetti, C.J. Biodiversity and Conservation of Coastal Lagoons. In Ecosystems Biodiversity; Grillo, O., Venora, G., Eds.; InTech: Vienna, Austria, 2011; pp. 1–28. [Google Scholar]
- Orfanidis, S.; Panayotidis, P.; Stamatis, N. An insight to the ecological evaluation index (EEI). Ecol. Indic. 2003, 3, 27–33. [Google Scholar] [CrossRef]
- García-Sánchez, M.; Pérez-Ruzafa, I.M.; Marcos, C.; Pérez-Ruzafa, A. Suitability of benthic macrophyte indices (EEI, E-MaQI and BENTHOS) for detecting anthropogenic pressures in a Mediterranean coastal lagoon (Mar Menor, Spain). Ecol. Indic. 2012, 19, 48–60. [Google Scholar] [CrossRef]
- Orfanidis, S.; Dencheva, K.; Nakou, K.; Tsioli, S.; Papathanasiou, V.; Rosati, I. Benthic macrophyte metrics as bioindicators of water quality: towards overcoming typological boundaries and methodological tradition in Mediterranean and Black Seas. Hydrobiologia 2014, 740, 61–78. [Google Scholar] [CrossRef]
- Tziortzis, I.; Kadis, K.; Papastegiadou, E. Use of Macrophyte assemblages for the ecological evaluation of two coastal lagoons of Greece according to WFD 2000/60/EC. J. Wetl. Biodivers. 2014, 4, 97–111. [Google Scholar]
- EEC. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 on establishing a framework for community action in the field of water policy. J. Eur. Commun. 2000, L327, 1–72. [Google Scholar]
- Sfriso, A.; Facca, C.; Ghetti, P.F. Validation of the Macrophyte Quality Index (MaQI) set up to assess the ecological status of Italian marine transitional environments. Hydrobiologia 2009, 617, 117–141. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Bonometto, A.; Boscolo, R. Compliance of the macrophyte quality index (MaQI) with the WFD (2000/60/EC) and ecological status assessment in transitional areas: The Venice lagoon as study case. Ecol. Indic. 2014, 46, 536–547. [Google Scholar] [CrossRef]
- ISPRA, 2010. Implementazione della Direttiva 2000/60/CE - Linea guida per l’applicazione dell’indice MaQI.
- Wilkinson, M. Benthic estuarine algae and their environment: a review. In The Shore Environment: Methods and Ecosytems; Price, J.H., Irvine, D.E.G., Farnham, W.F., Eds.; Academic Press: London, UK, 1980; Volume 2, pp. 425–486. [Google Scholar]
- Russell, G. Salinity and seaweed vegetation. In The physiological vegetation of amphibious and intertidal plants; Crawford, R.M., Ed.; Blackwell: Oxford, UK, 1987; pp. 32–35. [Google Scholar]
- Christia, C.; Giordani, G.; Papastergiadou, E. Environmental Variability and Macrophyte Assemblages in Coastal Lagoon Types of Western Greece (Mediterranean Sea). Water 2018, 10, 151. [Google Scholar] [CrossRef]
- Ferrarin, F.; Rapaglia, J.; Zaggia, L.; Umgiesser, G.; Zuppi, G.M. Coincident application of a mass balance of radium and a hydrodynamic model for the seasonal quantification of groundwater flux into the Venice Lagoon, Italy. Mar. Chem. 2008, 10, 179–188. [Google Scholar] [CrossRef]
- Krumgalz, B.S.; Hornung, H.; Oren, O.H. The study of a natural hypersaline lagoon in a desert area (the Bardawil Lagoon in Northern Sinai). Estuar. Coast. Mar. Sci. 1980, 10, 403–415. [Google Scholar] [CrossRef]
- Boutron, O.; Paugam, C.; Luna-Laurent, E.; Chauvelon, P.; Sous, D.; Rey, V.; Meulé, S.; Chérain, Y.; Cheiron, A.; Migne, E. Hydro-Saline Dynamics of a Shallow Mediterranean Coastal Lagoon: Complementary Information from Short and Long Term Monitoring. J. Mar. Sci. Eng. 2021, 9, 701. [Google Scholar] [CrossRef]
- Silva, E.I.L.; Katupotha, J.; Amarasinghe, O.; Manthrithilake, H.; Ariyaratna, R. Lagoons of Sri Lanka: From the Origins to the Present; International Water Management Institute: Colombo, Sri Lanka, 2013. [Google Scholar]
- Bamber, R.N.; Batten, S.D.; Sheader, M.; Bridgwater, M.D. On the ecology of brackish water lagoons in Great Britain. Aquat. Conserv. Mar. Freshw. Ecosyst. 1992, 2, 65–94. [Google Scholar] [CrossRef]
- Kristiansen, A.; Pedersen, P.M.; Moseholm, L. Salinity-temperature effects on growth and reproduction of Scytosiphon lomentaria (Fucophyceae) along the salinity gradient in Danish waters. Phycologia 1994, 33, 444–454. [Google Scholar] [CrossRef]
- Düwel, L. Experimental studies on macroalgae along the salinity gradient in the Baltic Sea area. Ph.D. Dissertation, Botanical Institute, Faculty of Science, University of Copenhagen, 2001. [Google Scholar]
- ISPRA, 2007. Guida alla tipizzazione dei corpi idrici di transizione ed alla definizione delle condizioni di riferimento ai sensi della direttiva 2000/60/CE.
- Battaglia, B. Final resolution of the symposium on the classification of brackish waters. Archo Oceanography Limnology 1959, 11, 243–248. [Google Scholar]
- Basset, A.; Sabetta, L.; Fonnesu, A.; Mouillot, D.; Do Chi, T.; Viaroli, P.; Giordani, G.; Reizopoulou, S.; Abbiati, M.; Carrada, G.C. Typology in Mediterranean transitional waters: new challenges and perspectives. Aquat. Conserv. 2006, 16, 441–455. [Google Scholar] [CrossRef]
- Cecere, E.; Petrocelli, A.; Izzo, G.; Sfriso, A. Flora and vegetation of the Italian transitional water systems (pp. 278). In Proceedings of the 4th Mediterranean Symposium on Marine Vegetation, Yasmine-Hammamet, Tunisia, 2–4 December 2010. [Google Scholar]
- Varvaglione, B.; Sabetta, L.; Basset, A. Tra Terra e Mare. Ecoguida alla Scoperta Delle Lagune e dei Laghi Costieri in Puglia; Università degli Studi di Lecce: Lecce, Italy, 2006. [Google Scholar]
- Apulian Region. Regional Water Protection Plan, Water Protection Service, General Report Area (In Italian), 2009.
- ISPRA, 2011. Protocols for sampling and determination of the biological and the physicochemical quality in the framework of the transitional water monitoring programs ex 2000/60/EC.
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway, 2022. https://www.algaebase.org; searched on 06 March 2023. 06 March.
- Lucena-Moya, P.; Pardo, I.; Álvarez, M. Development of a typology for transitional waters in the Mediterranean ecoregion: The case of the islands. Estuar. Coast. Shelf Sci. 2009, 82, 61–72. [Google Scholar] [CrossRef]
- Carstens, M.; Claussen, U.; Bergemann, M.; Gaumert, T. Transitional waters in Germany: The Elbe estuary as an example. Aquat. Conserv. 2004, 14(1), 81–92. [Google Scholar] [CrossRef]
- Boix, D.; Gascón, S.; Sala, J.; Martinoy, M.; Gifre, J.; Quintana, X.D. A new index of water quality assessment in Mediterranean wetlands based on crustacean and insect assemblages: the case of Catalunya (NE Iberian peninsula). Aquat. Conserv. 2005, 15, 635–651. [Google Scholar] [CrossRef]
- Ferreira, J.G.; Nobre, A.M.; Simas, T.C.; Silva, M.C.; Newton, A.; Bricker, S.B.; Wolff, W.J.; Stacey, P.E.; Sequeira, A. A methodology for defining homogeneous water bodies in estuaries e application to the transitional systems of the EU water Framework Directive. Estuar. 2006, 66, 468–482. [Google Scholar] [CrossRef]
- Kagalou, I.; Leonardos, I. Typology, classification and management issues of Greek lakes: implication of the Water Framework Directive (2000/60/EC). Environ. 2009, 150, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Por, F.D. A classification of hypersaline waters, based on trophic criteria. Mar. Ecol. 1980, 1, 121–131. [Google Scholar] [CrossRef]
- Adams, J.B.; Knoop, W.T.; Bate, G.C. The distribution of estuarine macrophytes in relation to freshwater. Bot. Mar. 1992, 35, 215–226. [Google Scholar] [CrossRef]
- Menéndez, M.; Hernandez, O.; Comin, F.A. Spatial distribution and ecophysiological characteristics of macrophytes in a Mediterranean coastal lagoon. Estuar. Coast. Shelf Sci. 2002, 55, 403–413. [Google Scholar] [CrossRef]
- Greve, T.M.; Binzer, T. Which factors regulate seagrass growth and distribution? In: Borum, J., C.M. Duarte, D. Krause-Jensen & T. M. Greve (eds), European Seagrasses an Introduction to Monitoring and Management. The M&Ms Project, Hillerød: 19–23, 2004.
- Casagranda, C.; Boudouresque, C.F. Biomass of Ruppia cirrhosa and Potamogeton pectinatus in a Mediterranean brackish lagoon, Lake Ichkeul, Tunisia. Fundam. 2007, 168, 243. [Google Scholar] [CrossRef]
- Christia, C. , Papastergiadou, E.S. Spatial and temporal variations of aquatic macrophytes and water quality in six coastal lagoons of western Greece. Belg. J. Bot. 2007, 39–50. [Google Scholar]
- Obrador, B.; Moreno-Ostos, E.; Pretus, J.L. A Dynamic Model to Simulate Water Level and Salinity in a Mediterranean Coastal Lagoon. Estuaries and Coasts 2008, 31, 1117–1129. [Google Scholar] [CrossRef]
- Antunes, C.; Correia, O.; da Silva, J.M.; Cruces, A.; da Conceição Freitas, M.; Branquinho, C. Factors involved in spatiotemporal dynamics of submerged macrophytes in a Portuguese coastal lagoon under Mediterranean climate. Estuar., Coast. and Shelf Sci. 2012, 110, 93–100. [Google Scholar] [CrossRef]
- Schubert, H.; Feuerpfeil, P.; Marquardt, R.; Telesh, I.; Skarlato, S. Macroalgal diversity along the Baltic Sea salinity gradient challenges Remane’s species-minimum concept. Mar. Pollut. 2011, 62, 1948–1956. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ruzafa, A.; Fernández, A.I.; Marcos, C.; Gilabert, J.; Quispe, J.I.; García-Charton, J.A. Spatial and temporal variations of hydrological conditions, nutrients and chlorophyll in a Mediterranean coastal lagoon (Mar Menor, Spain). Hydrobiologia 2005, 550, 11–27. [Google Scholar] [CrossRef]
- Franco, A.; Franzoi, P.; Torricelli, P. Structure and functioning of Mediterranean lagoon fish assemblages: a key for the identification of water body types. Estuar., Coast. and Shelf Sci. 2008, 79, 549–558. [Google Scholar] [CrossRef]
- Graham, L.; Graham, J.; Wilcox, L. Algae; Pearson Education, Inc.: San Francisco, 2009. [Google Scholar]
- Boegle, M.G.; Schneider, S.; Mannschreck, B.; Melzer, A. Differentiation of Chara intermedia and C. baltica compared to C. hispida based on morphology and amplified fragment length polymorphism. Hydrobiologia 2007, 586, 155–166. [Google Scholar] [CrossRef]
- den Hartog, C. The Seagrasses of the World; North Holland: Amsterdam, 1970. [Google Scholar]
- Aiello, G.; Barra, D.; Coppa, M.G.; Valente, A.; Zeni, F. Recent infralittoral foraminiferida and ostracoda from the porto Cesareo Lagoon (Ionian Sea, Mediterranean). Boll. Soc. Paleontol. 2006, 45, 1. [Google Scholar]
- Cardellicchio, N.; Annicchiarico, C.; Di Leo, A.; Giandomenico, S.; Spada, L. The Mar Piccolo of Taranto: an interesting marine ecosystem for the environmental problems studies. Environ. 2016, 23, 12495–12501. [Google Scholar] [CrossRef] [PubMed]
- Cecere, E.; Petrocelli, A.; Saracino, O.D. Biodiversity of phytobenthic communities in the marine reserve of Porto Cesareo. Biol. Mar. Medit. 2005, 2005 12, 78–87. [Google Scholar]
- den Hartog, C.; Triest, L. A profound view and discourse on the typification and status of three confused taxa: Ruppia maritima, R. spiralis and R. cirrhosa. Bot. Mar. 2020, 63, 229–239. [Google Scholar] [CrossRef]
- Bonometto, A.; Ponis, E.; Cacciatore, F.; Riccardi, E.; Pigozzi, S.; Parati, P.; Novello, M.; Ungaro, N.; Acquavita, A.; Manconi, P.; et al. A New Multi-Index Method for the Eutrophication Assessment in Transitional Waters: Large-Scale Implementation in Italian Lagoons. Environments 2022, 9, 41. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Bon, D.; Buosi, A. Macrophytes and ecological status assessment in the Po delta transitional systems, Adriatic Sea (Italy). Application of Macrophyte Quality Index (MaQI). Acta Adriat. 2016, 57, 209–225. [Google Scholar]
- Wallentinus, I. The Baltic Sea. Ecosys. World 1991, 24, 83–108. [Google Scholar]
- Hayakawa, Y.I.; Ogawa, T.; Yoshikawa, S.; Ohki, K.; Kamiya, M. Genetic and ecophysiological diversity of Cladophora (Cladophorales, Ulvophyceae) in various salinity regimes. Phycological Res. 2012, 60, 86–97. [Google Scholar] [CrossRef]
- Rybak, A.S. Species of Ulva (Ulvophyceae, Chlorophyta) as indicators of salinity. Ecol. Indic. 2018, 85, 253–261. [Google Scholar] [CrossRef]
Figure 1.
Map of the study area showing its position in the Mediterranean Sea, the location of the transitional water basins and the sampling stations placed in each transitional water body (sensu WFD).
Figure 1.
Map of the study area showing its position in the Mediterranean Sea, the location of the transitional water basins and the sampling stations placed in each transitional water body (sensu WFD).
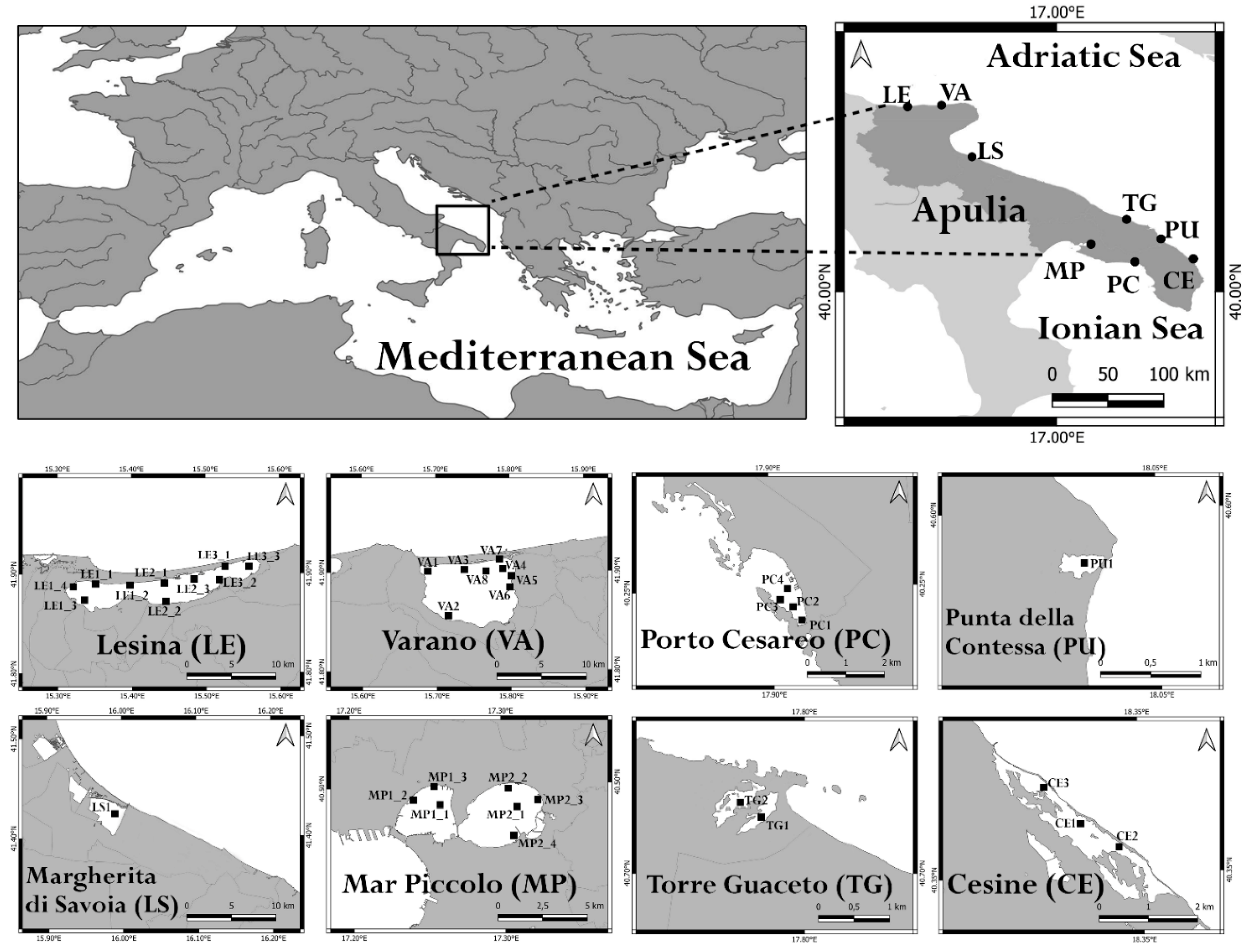
Figure 2.
Trend of the species richness (i.e., number of species) in relation to the salinity level of each T-WB considered in the study (a) and total number of species counted for each of the four classes (b).
Figure 2.
Trend of the species richness (i.e., number of species) in relation to the salinity level of each T-WB considered in the study (a) and total number of species counted for each of the four classes (b).
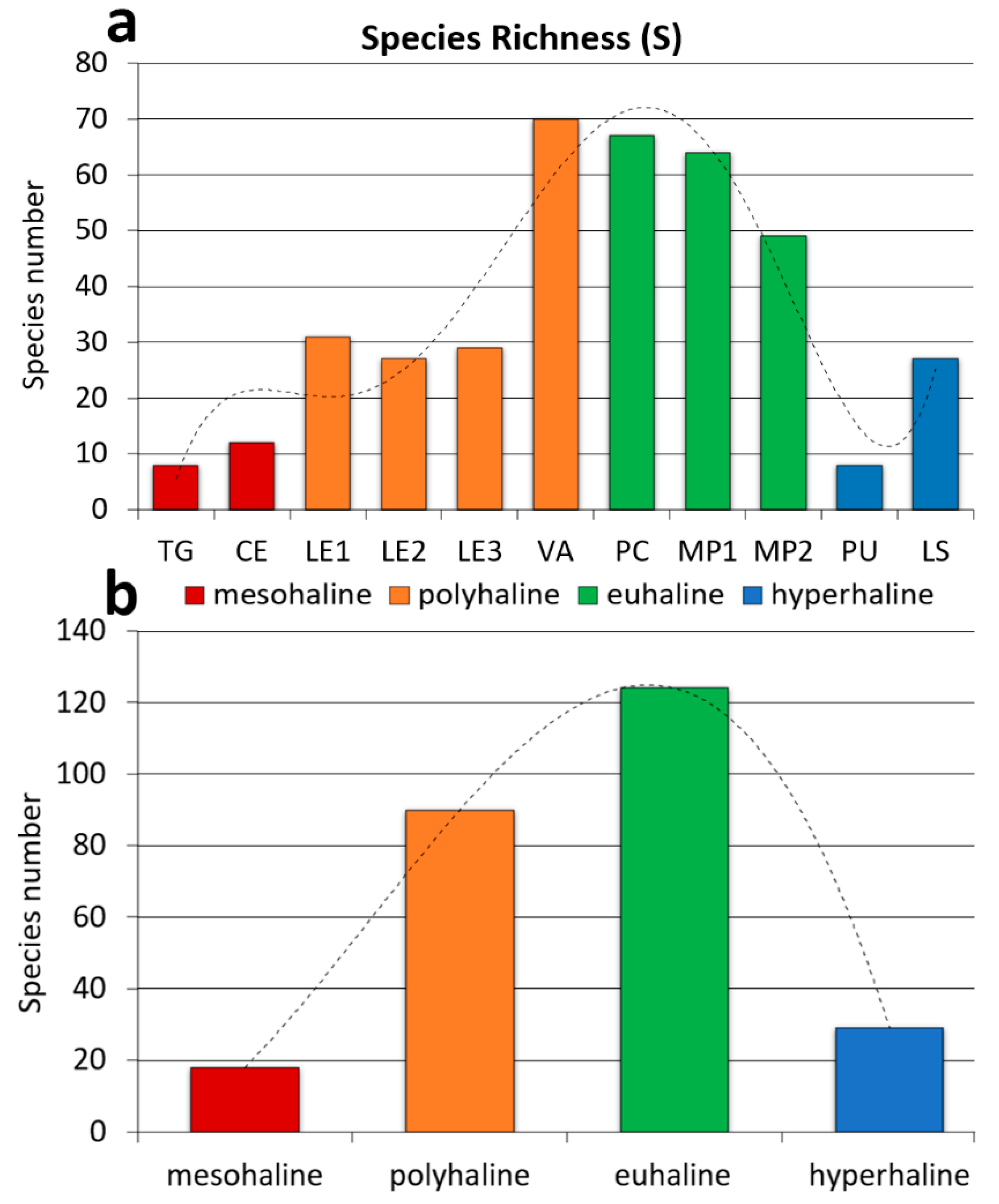
Figure 3.
Results of Cluster Analyses (a) and non-Metric Multidimensional Scaling (n-MDS) (b) based on Bray-Curtis dissimilarities between T-WBs of the four salinity classes.
Figure 3.
Results of Cluster Analyses (a) and non-Metric Multidimensional Scaling (n-MDS) (b) based on Bray-Curtis dissimilarities between T-WBs of the four salinity classes.

Table 1.
Coordinates, extension (km2), depth range (m) and salinity class (PSU) of each T-WB.
| WB | Coordinates | Surface area (km2) | Depth (m) | Salinity Class (PSU)* |
|---|---|---|---|---|
| Torre Guaceto (TG) | 40°42′51.34” N – 17°47′42.91” E | 1.2 | 0.4 – 0.6 | Mesohaline (5–20) |
| Cesine (CE) | 40°21′33.46” N – 18°20′09.30” E | 0.7 | 0.2 – 0.8 | Mesohaline (5 – 20) |
| Lesina (LE1) | 41°53′12.64” N – 15°21′15.65” E | 18 | 0.7 – 2 | Polyhaline (20 – 30) |
| Lesina (LE2) | 41°53′01.23” N – 15°27′20.15” E | 17 | 0.7 – 2 | Polyhaline (20 – 30) |
| Lesina (LE3) | 41°53′57.20” N – 15°31′00.45” E | 16 | 0.7 – 2 | Polyhaline (20 – 30) |
| Varano (VA) | 41°52′43.65” N – 15°44′35.42” E | 60.5 | 0.5 – 5 | Polyhaline (20 – 30) |
| Porto Cesareo (PC) | 40°14′31.80” N – 17°54′32.82” E | 2 | 0.3 – 5 | Euhaline (30 – 40) |
| Mar Piccolo (MP1) | 40°29′19.68” N – 17°15′29.51” E | 9.7 | 0.5 – 12 | Euhaline (30 – 40) |
| Mar Piccolo (MP2) | 40°29′22.92” N – 17°18′29.18” E | 11 | 0.5 – 12 | Euhaline (30 – 40) |
| Margherita di Savoia (LS) | 41°25′27.34” N – 15°59′53.29” E | 45 | 0.4 – 0.6 | Hyperhaline (> 40) |
| Punta della Contessa (PU) | 40°35′42.31” N – 18°02′30.05” E | 2 | 0.4 – 1 | Hyperhaline (> 40) |
*for the first identification of T-WBs according to the Italian Ministerial Decree 131/08.
Table 2.
Taxonomic list of the species found during the monitoring periods (2011-2019), grouped in orders inside each phylum. The table reports the frequency values abundance (%) of the taxa in the four different types of T-WBs according to salinity classes (HH: hyperhaline; EH: euhaline; PH: polyhaline; MH: mesohaline). +: 1–24%; ++: 25–49%; +++: 50–74%; ++++: 75–100%:.
Table 2.
Taxonomic list of the species found during the monitoring periods (2011-2019), grouped in orders inside each phylum. The table reports the frequency values abundance (%) of the taxa in the four different types of T-WBs according to salinity classes (HH: hyperhaline; EH: euhaline; PH: polyhaline; MH: mesohaline). +: 1–24%; ++: 25–49%; +++: 50–74%; ++++: 75–100%:.
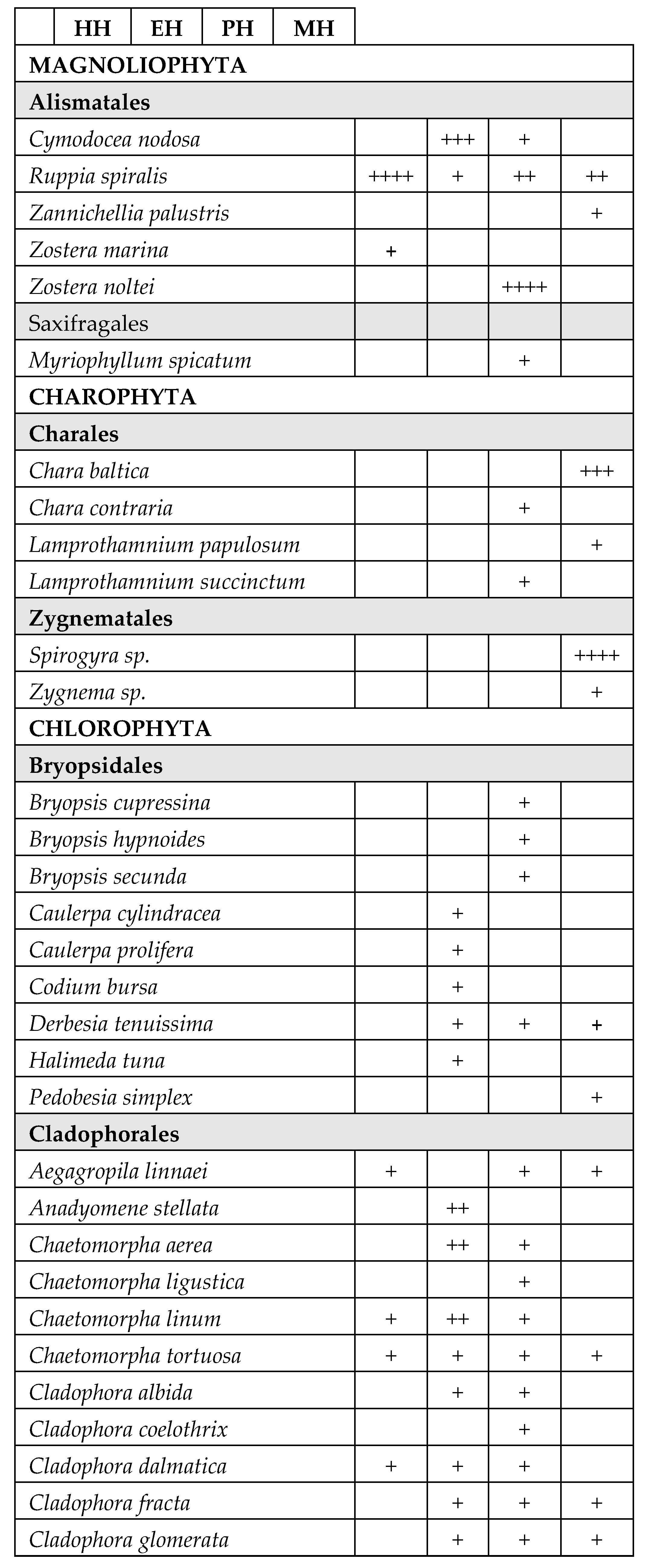
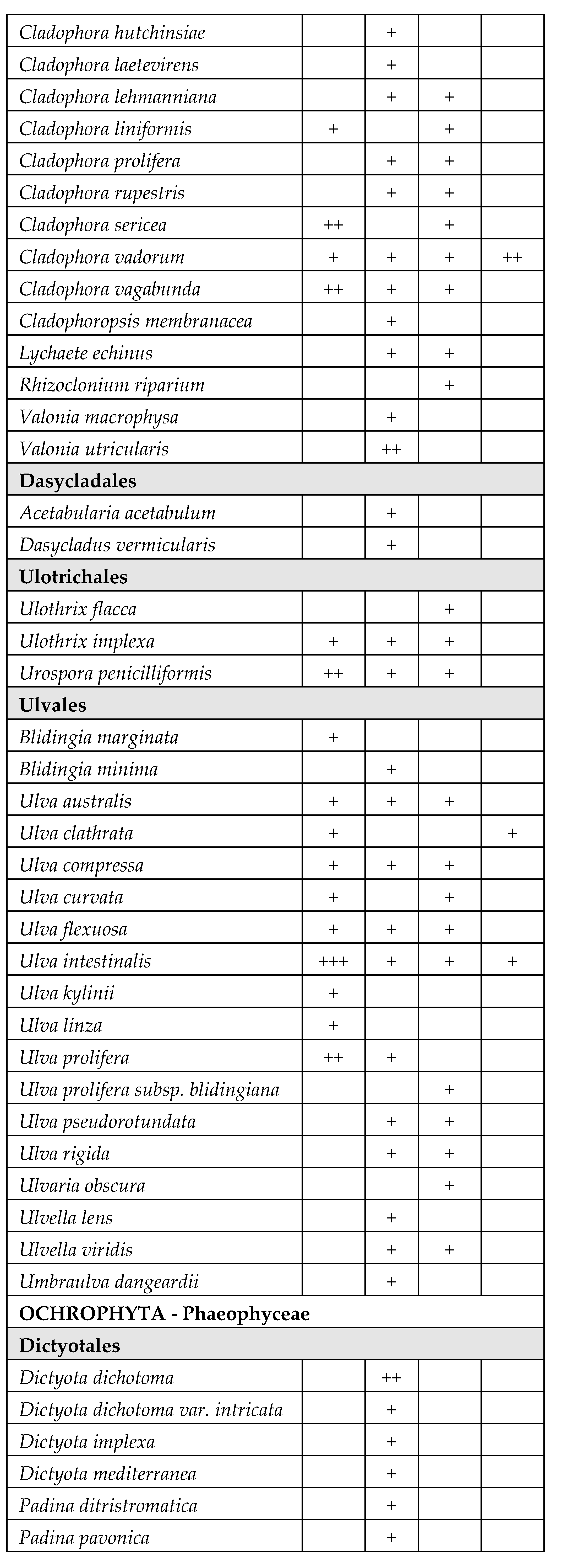
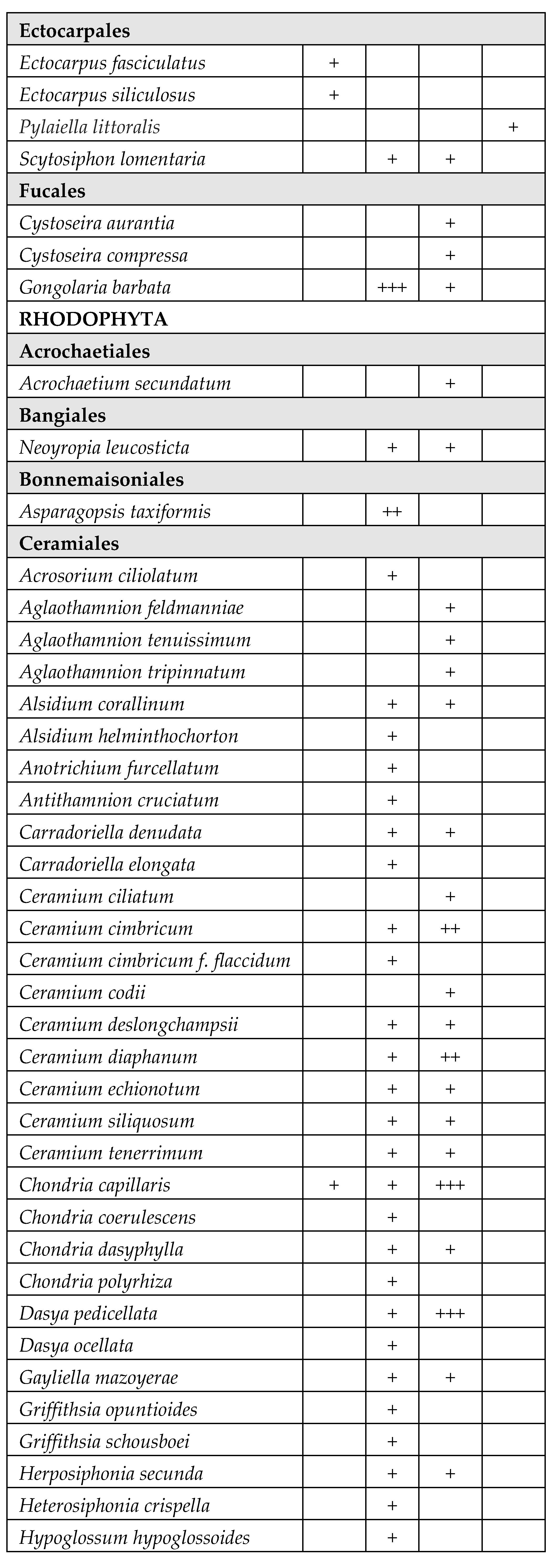
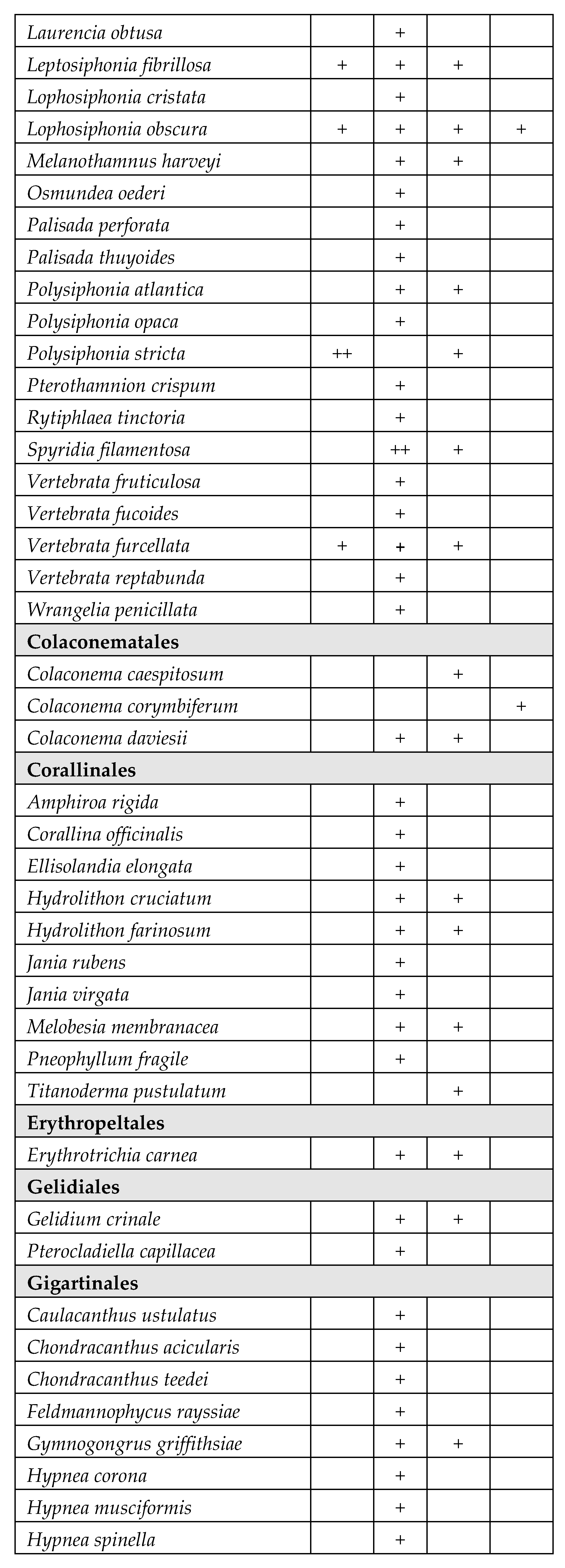
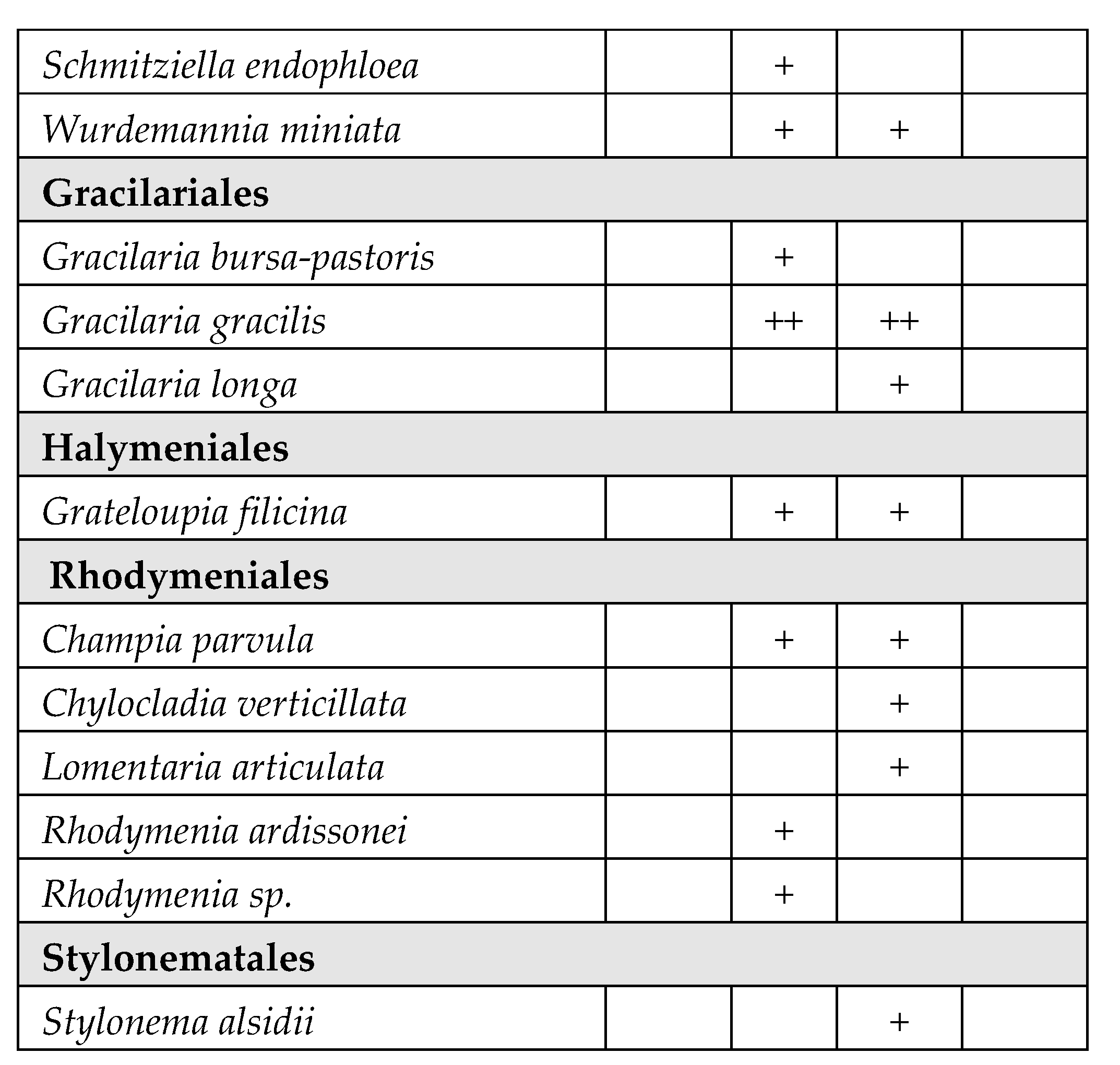
Table 3.
Pairwise and global results of the ANOSIM test carried out to verify the data significance.
Table 3.
Pairwise and global results of the ANOSIM test carried out to verify the data significance.
| pairwise ANOSIM | R-values | p-values |
|---|---|---|
| hyperhaline vs euhaline | 1 | 0.1019 |
| hyperhaline vs polyhaline | 1 | 0.0649 |
| hyperhaline vs mesohaline | 1 | 0.3290 |
| euhaline vs polyhaline | 0.9630 | 0.0278* |
| euhaline vs mesohaline | 1 | 0.0986 |
| polyhaline vs mesohaline | 1 | 0.0659 |
| Global ANOSIM | R-values | p-values |
| 0.9545 | 0.0002*** |
* p < 0.05; *** p < 0.001.
Table 4.
SIMPER results showing the average abundance, the average similarity, the percentage contribution of each species in similarity among the four salinity classes, the percentage cumulative similarity and the average similarity of the groups.
Table 4.
SIMPER results showing the average abundance, the average similarity, the percentage contribution of each species in similarity among the four salinity classes, the percentage cumulative similarity and the average similarity of the groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



