
Submitted:
28 March 2023
Posted:
29 March 2023
You are already at the latest version
Abstract
Phylogenetics is a powerful tool for understanding the evolutionary history of organisms and for informing conservation and management of species. Among the strepsirrhine primates, the slender lorises are a threatened genus of small, nocturnal animals confined to India and Sri Lanka. The grey slender loris (Loris lydekkerianus) is divided into several subspecies based on the morphological and geographical variation but not supported by molecular data. We investigated the genetic basis of taxonomic and biogeographic variation as well as the phylogenetic divergence of two subspecies of the grey slender loris in southern India: the Mysore slender loris (Loris lydekkerianus ssp. lydekkerianus) and the Malabar slender loris (Loris lydekkerianus ssp. malabaricus). We sequenced and assembled the whole mitochondrial genomes of three representative individuals from their distribution in southern India and compared them with publicly available mitogenomes of other lorises. We found that the two Indian subspecies vary by 2.09% in the COX1 and CYTB gene regions and form distinct monophyletic clades that diverged about 1.049 million years ago. Our results support the morphological classification of these two subspecies in southern India and have implications for their conservation and management in captivity and in the wild.
Keywords:
Grey slender loris
; Mysore slender loris
; Malabar slender loris
; Phylogenetics
1. Introduction
Slender lorises are one of the two genera of extreme nocturnal primates that inhabit India [1]. They belong to the family Lorisidae, which also includes slow lorises, pottos, and angwantibos. Slender lorises are confined to India and Sri Lanka, where they inhabit dry to moist and lowland to montane forests [2]. Slender lorises are characterized by their small size, long limbs, vestigial tail, large eyes, and slow locomotion. They are adapted for arboreal life, using their opposable thumbs and toes to grasp branches and their binocular vision to judge distances. They feed mainly on insects but also consume fruits, flowers, gums, and other plant materials [3,4]. They have a variety of vocalizations that may help them avoid predators and communicate with conspecifics [5]. Slender lorises are divided into two species: the grey slender loris (Loris lydekkerianus) found in southern India and Sri Lanka and the red slender loris (Loris tardigradus) found only in Sri Lanka [6]. Both species show high phenotypic variation in fur color, body size, and cranial morphology, leading to the recognition of several subspecies, most of which are refuted by molecular studies [7]. The Mysore slender loris (Loris lydekkerianus ssp. lydekkerianus) and the Malabar slender loris (Loris lydekkerianus ssp. malabaricus), which live in the dry and wet forests of the Eastern and Western Ghats, respectively, are the two subspecies that have been recognized thus far in southern India [8]. There are several regions in their distribution where slender lorises face serious threats to their existence such as habitat loss due to deforestation and urbanization, electrocution on live wires, road accidents, pet trade, and illegal poaching for traditional medicine and black magic [9]. The IUCN Red List of Threatened Species classifies the Mysore Slender Loris [10] and Malabar Slender Loris [11] as "Near Threatened" and they are listed under Schedule I of the Indian Wild Life (Protection) Act, 1972. Recently, Tamil Nadu became the first Indian state to notify a sanctuary for slender lorises spanning 11,806 hectares which is crucial for protecting their habitat and ensuring the survival of this unique primate species.
The Mysore and Malabar subspecies of slender loris were delineated based on their geographic distribution and morphological differences [6,8]. The Mysore slender loris has a reddish-brown coat, a prominent white stripe on its forehead, compared to the Malabar slender loris, which has a greyish-brown coat, a less distinct forehead stripe [6,12]. The relative distribution of the two subspecies as well as their comparative densities and the extent of overlap between their distributions have been very well established [8]. The Mysore slender loris is found in the Eastern Ghats and eastern foothills of the southern Western Ghats, while the Malabar slender loris is confined to the western slope of the entire Western Ghats [8]. The Mysore slender loris prefers dry deciduous forests with moderate canopy cover and high tree density, while the Malabar slender loris prefers moist evergreen forests with high canopy cover and low tree density [8]. Their distributions overlap along the southern ridges of the Western Ghats, where hybridization may occur. The Mysore slender loris is relatively larger (ca. 260 g) than Malabar slender loris (ca. 180 g) [12]. The diet and feeding behavior of these subspecies varies depending on their habitat types and availability of food resources [4]. The reproductive biology and social system of the Mysore subspecies is influenced by factors such as seasonality, food availability, predation risk, and population density. They also have a seasonal breeding cycle that coincides with periods of high food availability [13]. But no such information on the reproductive biology of the Malabar subspecies is available. Behavioral studies on lorises have always been more challenging than on relatively large, diurnal, and group-living primates such as macaques and langurs because they are nocturnal, small in size, and mostly semi-gregarious. Considering these two subspecies geographically occupy two distinct landscapes and habitat conditions, and further very distinct in morphology and body size, understanding their evolutionary history and genetic difference becomes vital to address their conservation status and management issues.
Therefore, the main objective of this study is to investigate the phylogenetic relationship and genetic divergence between the Mysore and Malabar subspecies of slender loris in southern India. To achieve this, we sequenced and assembled the whole mitochondrial sequences from three representative samples. We aligned these sequences with the publicly available sequences of other lorises and constructed phylogenetic trees. We estimated the sequence divergence and divergence time between the two subspecies and found that the phylogenetic analysis supports the morphological and geographical delineation of the Malabar and Mysore slender lorises. The results of this study will contribute to the understanding of the biogeography and speciation processes of these threatened lorises and provide crucial insights for their conservation management.
2. Materials and Methods
2.1. Sample Collection, DNA Extraction, and Sequencing
Sample collection was done according to the guidelines of the Animal ethics committees of the CSIR-Centre for Cellular and Molecular Biology and Salim Ali Centre for Ornithology and Natural History. Necessary permissions for sample collection were obtained from the Central Zoo Authority of India, Ministry of Environment, Forests and Climate Change, Government of India, vide Ref. No. 9-2/2005-CZA(M) Vol III. Rescued lorises of known wild origin within the IUCN designated ranges (Figure 1) that were captive in Mysore and Hyderabad Zoos were the sources of our samples. Blood samples were collected in EDTA vacutainers by qualified Zoo veterinarians from three representative individuals of Loris lydekkerianus ssp. lydekkerianus (N=2) and Loris lydekkerianus ssp. malabaricus (N=1). We used the Qiagen DNeasy Blood and Tissue Kit to isolate the high-quality genomic DNA from the blood samples. The quality and quantity of genomic DNA were measured using Nanodrop and Qubit 4. We constructed whole genome libraries using the Truseq PCR-free library preparation kit according to Illumina’s protocols. Briefly, 1ug of genomic DNA was sheared to approximately 350 bp using the Covaris ultrasonicator. The fragmented DNA was then end-repaired and blunt-end ligated with sequencing adapters containing unique dual indices from IDT. The library was then size-selected using SPRI beads and verified on the Agilent fragment analyzer. The cleaned-up libraries were finally quantified in qPCR using the standards and Illumina adapter-specific primers from the Roche library quantification kit. Libraries having good concentration were pooled along with other samples and sequenced on the Illumina Novaseq 6000 platform for 300 cycles in paired-end mode.
2.2. Mitochondrial Genome Assembly
We demultiplexed the base call files to separate the 3 samples with the dual-indexed barcodes using the BCL2FASTQ tool from Illumina. Raw reads were quality-filtered with a Phred quality score threshold of 15 using FASTP v0.20 [14]. We subsampled 10 million quality filtered reads to de novo assemble the circular mitochondrial genomes of all 3 samples using GetOrganelle v1.7.1 [15]. We then annotated all the mitogenomes using MITOS2 [16] with the Refseq 89 Metazoa reference mitochondrial database and the Vertebrate Mitochondrial genetic code. All the coding and non-coding genes were extracted from the mitochondrial genomes using the annotations. The pairwise sequence similarity in the full-length COX1 and CYTB genes was calculated using Clustal Omega with the “distmat” flag [17].
2.3. Phylogenetic Analyses
To build the phylogeny, along with our samples we used the NCBI RefSeq mitochondrial sequences from strepsirrhines namely, Loris lydekkerianus, Loris tardigradus, Nycticebus coucang, Nycticebus bengalensis, Galago senegalensis, and Lemur catta. We aligned the 13 coding and 2 non-coding ribosomal RNA genes individually from the assembled mitochondrial genomes and reference sequences using the MUSCLE algorithm in MEGA7 [18] and checked for the presence of any sequencing errors or frameshifts for codon position. We then concatenated all the gene alignments and used the corrected Akaike information criterion (AICc) values from PartitionFinder2 [19] to identify the optimum nucleotide substitution model for each partition when building a phylogenetic tree (TRN, JC, HKY, GTR, GTR+I, TRN+I, GTR+G, HKY+G, HKY+I, K80). We built the maximum likelihood (ML) tree based using the best-fitting substitution model using IQ-TREE [20] with 1000 times bootstrapping. The ML tree was visualized in iTOL [21]. We utilized BEAST2.5 [22] to create a Bayesian inference-based divergence time tree using the same concatenated alignment of 13 coding and 2 non-coding genes from the complete mitochondrial genomes. We chose two calibration points:
1) We calibrated the crown node of galagos with 38 mya based on the age of the fossil Saharagalago misrensis (PaleoDB collection 67706) [23]. We applied a normal distribution at 40 Mya (SD = 0.04; 95% range: 36–43)
2) We calibrated the crown node of slow lorises with 13.82 mya based on the age of the fossil Nycticebus linglom (PaleoDB collection 48126) [24]. We applied a normal distribution at 14 Mya (SD = 0.05; 95% range: 9–17)
For all the partitions, we created a relaxed lognormal clock and employed a Birth-Death process using prior distributions. To get to the final tree, we ran for 40 million generation runs, sampling every 4,000th generation using TreeAnnotator [25] with a 10% burn-in. We verified that all the ESS values were over 200 in Tracer 1.7 [26] and visualized the tree in Figtree [27].
3. Results
3.1. Phylogenetics Supports the Morphological Delineation of Mysore and Malabar Slender Lorises
To investigate the genetic differences between the two subspecies of slender loris in southern India, we first assembled three new circular mitochondrial genomes with an average length of 16,771 bp from two samples of the Mysore slender loris (Loris lydekkerianus ssp. lydekkerianus) and one sample of Malabar slender loris (Loris lydekkerianus ssp. malabaricus). We annotated the mitochondrial genomes along with published reference sequences and obtained the full-length sequences of 13 protein-coding genes and 2 ribosomal RNA genes. To check the variation in the nucleotide sequence within the Loris genus, we estimated the pairwise sequence similarity in the COX1 and CYTB regions spanning 2,682 bp between all six Loris samples (Table 1). We observed the highest average sequence variation of 2.82% (S.D. 0.16) between the 4 sequences of the grey slender loris (Loris lydekkerianus) and the 2 sequences of the red slender loris (Loris tardigradus) as they belong to two different species within the Loris genus. While there was no sequence variation found within the 2 sequences of red slender loris, there was considerable variation within the 4 sequences of grey slender loris contributed by the difference between the two subspecies. We found about 2.09% variation in the COX1 and CYTB sequences of the Mysore and Malabar subspecies of the grey slender loris.
To understand the phylogenetic relationships of the two subspecies with other lorises, we constructed a maximum likelihood tree from 13 coding and 2 non-coding genes. Along with our 3 samples, we included reference mitochondrial sequences from 2 species of slender lorises (L. lydekkerianus, L. tardigradus) and 2 species of slow lorises (N. bengalensis, N. coucang) along with Galago and Lemur as outgroups (Figure 2). The phylogenetic tree recapitulates the broad evolutionary relationships of slender lorises with slow lorises and the outgroups. It reveals an interesting pattern within the clade of slender lorises where the Mysore subspecies (L. l. lydekkerianus) clusters with the reference sequence of grey slender loris to form a monophyletic clade and the Malabar subspecies (L. l. malabaricus) forms a separate clade with very strong statistical support. We noted that the Malabar slender loris appears more closely related to the red slender loris (L. tardigradus), albeit with a very small branch length (Figure 2). To estimate the divergence time between the two subspecies and other lorises, we constructed a fossil-calibrated bayesian tree (Figure 3). Our data suggests that the split between the grey slender loris (L. lydekkerianus) and red slender loris (L. tardigradus) occurred approximately 1.087 million years ago (mya). This was immediately followed by diversification of the Mysore (L. l. lydekkerianus) and Malabar (L. l. malabaricus) subspecies of grey slender loris at around 1.049 mya (PP = 0.99) (Figure 3).
4. Discussion
Our results from the phylogenetic analyses based on the mitochondrial sequences show that the Mysore and Malabar subspecies have about 2.09% nucleotide sequence variation in the full-length COX1 and CYTB genes and form distinct monophyletic clades in the phylogenetic tree that diverged about 1.049 million years ago. This data supports the morphological and geographical delineation of Mysore slender loris (Loris lydekkerianus ssp. lydekkerianus) and Malabar slender loris (Loris lydekkerianus ssp. malabaricus) [6,8,12].
The genetic differences and divergence time between the Mysore and the Malabar subspecies are surprisingly high, which is not very common between primate subspecies. The level of genetic divergence observed between these two subspecies indicates that they have been evolving independently for a significant period of time. This is also corroborated by the results from the molecular dating analysis which estimates that the split between these two subspecies occurred over a million years ago. This divergence time estimate is supported by a number of molecular markers and is consistent with previous studies on the evolutionary history of this genus [28]. Understanding the genetic structure and variation of species is crucial for the scientific management of threatened species and their eventual recovery. The findings of this study have important implications for the conservation and management of the grey slender loris in India. With a clearer understanding of the genetic differences between these two subspecies, it will be possible to more accurately identify and classify individual animals, which will in turn facilitate the development of an effective conservation breeding program. Such a program can be particularly beneficial for species like the slender loris that are threatened by habitat loss and fragmentation, and whose populations have been declining in recent years [12,29].
The main limitation of this study is the small sample size due to the requirement of whole mitochondrial genomes for accurate molecular dating. More samples from the Malabar slender loris could better resolve the phylogenetic tree and nuclear markers could also be used to further confirm our findings. Comprehensive genome sequencing of all the subspecies of slender loris would also help to understand the genomic basis of morphological differences between these subspecies that have adapted to their respective niches for over one million years.
5. Conclusions
In summary, this study provides strong molecular evidence for the genetic divergence and distinctiveness of the Mysore slender loris (Loris lydekkerianus ssp. lydekkerianus) and the Malabar slender loris (Loris lydekkerianus ssp. malabaricus). The sequence analysis, phylogenetic analyses, and molecular dating suggest that the Mysore and Malabar subspecies are genetically distinct and have been evolving independently for a significant period of time and support their morphological and geographical delineation. The high level of genetic divergence between the two subspecies highlights the importance of preserving the genetic diversity of these subspecies and underscores the need for more efforts to conserve them in the wild. We recommend that both the Mysore and Malabar subspecies of grey slender loris should be considered as distinct conservation units and protected accordingly as each subspecies represents a unique evolutionary lineage that deserves separate recognition and protection.
Author Contributions
G.U. and H.N.K. conceived the idea for this study and collected the samples. S.M. generated the data; V.T. and S.M. analyzed and interpreted the data and drafted the manuscript. G.U. and H.N.K. edited the manuscript. All authors have read and agreed to the final version of the manuscript.
Funding
V.T. was supported by a CSIR PhD fellowship and S.M. was supported by a DBT-BINC PhD fellowship from the Govt. of India. GU was supported by CSIR grant.
Data Availability Statement
The three whole mitochondrial genome sequences generated in this study will be deposited to the NCBI database and the accession numbers will be provided after acceptance.
Acknowledgments
We are grateful to Mihir Trivedi for providing useful suggestions for the phylogenetic analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nekaris, K. a. I. Extreme Primates: Ecology and Evolution of Asian Lorises. Evol. Anthropol. Issues News Rev. 2014, 23, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Singh, M.; Kumara, H.N.; Kumar, S.; Gnanaoliu, S.D.; Sasi, R. A Review of Research on the Distribution, Ecology, Behaviour, and Conservation of the Slender Loris Loris Lydekkerianus (Mammalia: Primates: Lorisidae) in India. J. Threat. Taxa 2021, 13, 19540–19552. [Google Scholar] [CrossRef]
- Nekaris, K.A.I.; Rasmussen, D.T. Diet and Feeding Behavior of Mysore Slender Lorises. Int. J. Primatol. 2003, 24, 33–46. [Google Scholar] [CrossRef]
- Radhakrishna, S.; Kumara, H.N. Behavioural Variation in the Mysore Slender Loris Loris Lydekkerianus Lydekkerianus. Curr. Sci. 2010, 99, 1226–1232. [Google Scholar]
- Radhakrishna, S.; Singh, M. Social Behaviour of the Slender Loris (Loris Tardigradus Lydekkerianus). Folia Primatol. (Basel) 2002, 73, 181–196. [Google Scholar] [CrossRef]
- Groves, C.P. Primate Taxonomy; Smithsonian Institution Press, 2001.
- Nijman, V.; Robbins, T.; Maddock, A.J.; Ang, A. Molecular Phylogeny, Taxonomy and Conservation of Slender Lorises. Primate Conserv. 2020, 34, 143–151. [Google Scholar]
- Kumara, H.N.; Singh, M.; Irfan-Ullah, M.; Kumar, S. Status, Distribution, and Conservation of Slender Lorises in India. In Leaping Ahead: Advances in Prosimian Biology; Masters, J., Gamba, M., Génin, F., Eds.; Developments in Primatology: Progress and Prospects; Springer: New York, NY, 2013; pp. 343–352. ISBN 978-1-4614-4511-1. [Google Scholar]
- Dittus, W.; Singh, M.; Kumara, H.N.; Kumar, A. IUCN Red List of Threatened Species: Loris Lydekkerianus. IUCN Red List Threat. Species 2022. [Google Scholar]
- Kumara, H.N.; Nekaris, A.; Singh, M. IUCN Red List of Threatened Species: Loris Lydekkerianus Ssp. Lydekkerianus. IUCN Red List Threat. Species 2022. [Google Scholar]
- Kumara, H.N.; Nekaris, A.; Singh, M. IUCN Red List of Threatened Species: Loris Lydekkerianus Ssp. Malabaricus. IUCN Red List Threat. Species 2022. [Google Scholar]
- Kumara, H.N.; Singh, M.; Kumar, S. Distribution, Habitat Correlates, and Conservation of Loris Lydekkerianus in Karnataka, India. Int. J. Primatol. 2006, 27, 941–969. [Google Scholar] [CrossRef]
- Radhakrishna, S.; Singh, M. Reproductive Biology of the Slender Loris (Loris Lydekkerianus Lydekkerianus). Folia Primatol. (Basel) 2004, 75, 1–13. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A Fast and Versatile Toolkit for Accurate de Novo Assembly of Organelle Genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de Novo Metazoan Mitochondrial Genome Annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. The Clustal Omega Multiple Alignment Package. In Multiple Sequence Alignment: Methods and Protocols; Katoh, K., Ed.; Methods in Molecular Biology; Springer US: New York, NY, 2021; pp. 3–16. ISBN 978-1-07-161036-7. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLOS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed]
- Seiffert, E.R.; Simons, E.L.; Attia, Y. Fossil Evidence for an Ancient Divergence of Lorises and Galagos. Nature 2003, 422, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Harrison, T. Later Tertiary Lorisiformes. In Cenozoic Mammals of Africa; Werdelin, L., Ed.; University of California Press, 2010; p. 0 ISBN 978-0-520-25721-4.
- Helfrich, P.; Rieb, E.; Abrami, G.; Lücking, A.; Mehler, A. TreeAnnotator: Versatile Visual Annotation of Hierarchical Text Relations. In Proceedings of the Proceedings of the Eleventh International Conference on Language Resources and Evaluation (LREC 2018); European Language Resources Association (ELRA): Miyazaki, Japan, May 2018. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree v1. 4.2, a Graphical Viewer of Phylogenetic Trees 2014.
- Finstermeier, K.; Zinner, D.; Brameier, M.; Meyer, M.; Kreuz, E.; Hofreiter, M.; Roos, C. A Mitogenomic Phylogeny of Living Primates. PLOS ONE 2013, 8, e69504. [Google Scholar] [CrossRef] [PubMed]
- Kumara, H.N.; Sasi, R.; Chandran, S.; Radhakrishna, S. Distribution of the Grey Slender Loris (Loris Lyddekerianus Cabrera, 1908) in Tamil Nadu, Southern India. Folia Primatol. (Basel) 2016, 87, 291–302. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
IUCN range of the grey slender loris subspecies in southern India and Sri Lanka.
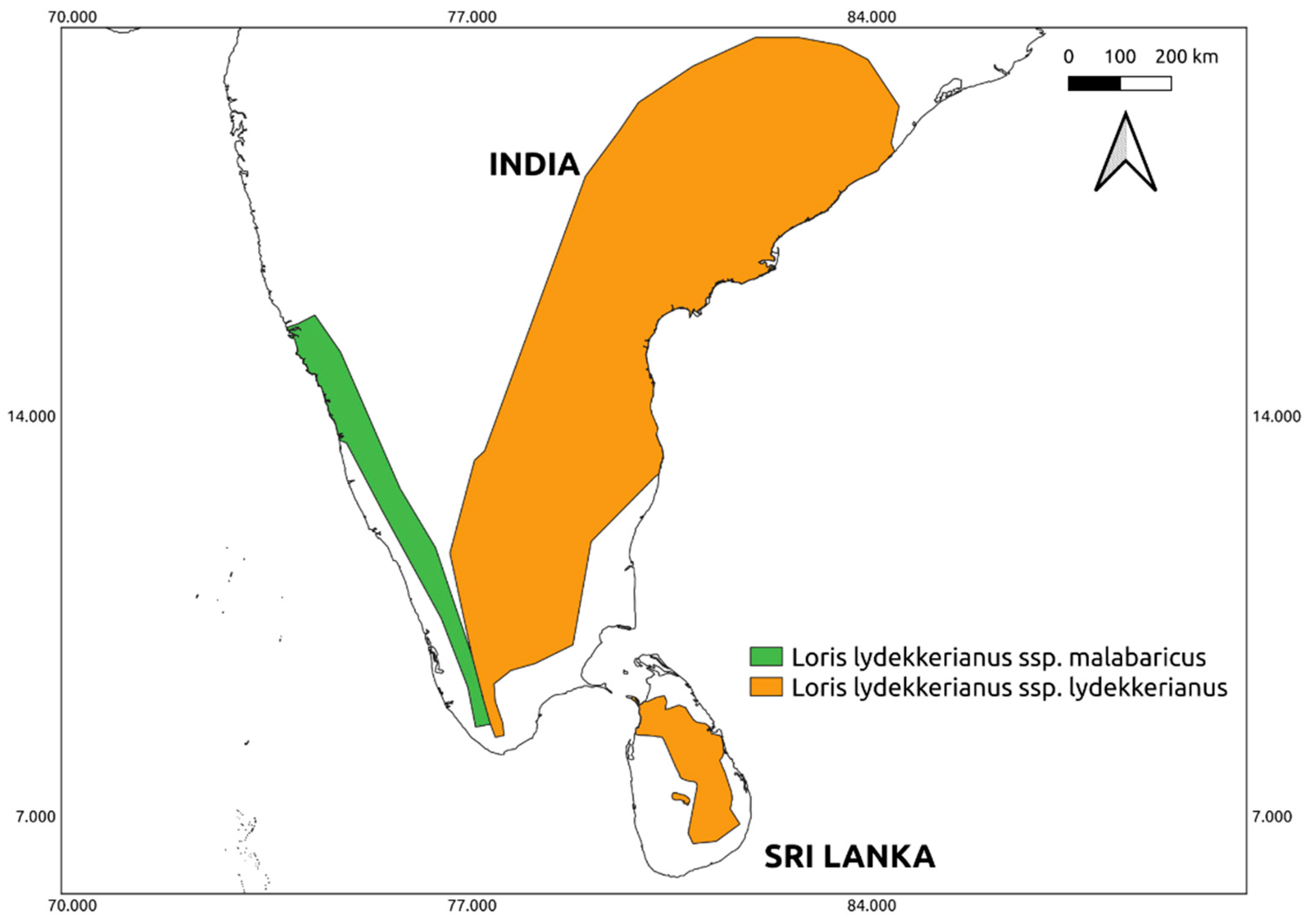
Figure 2.
Maximum likelihood tree based on 13 coding and 2 ribosomal RNA genes from whole mitochondrial genomes. The bootstrap values are denoted at the nodes. .
Figure 2.
Maximum likelihood tree based on 13 coding and 2 ribosomal RNA genes from whole mitochondrial genomes. The bootstrap values are denoted at the nodes. .
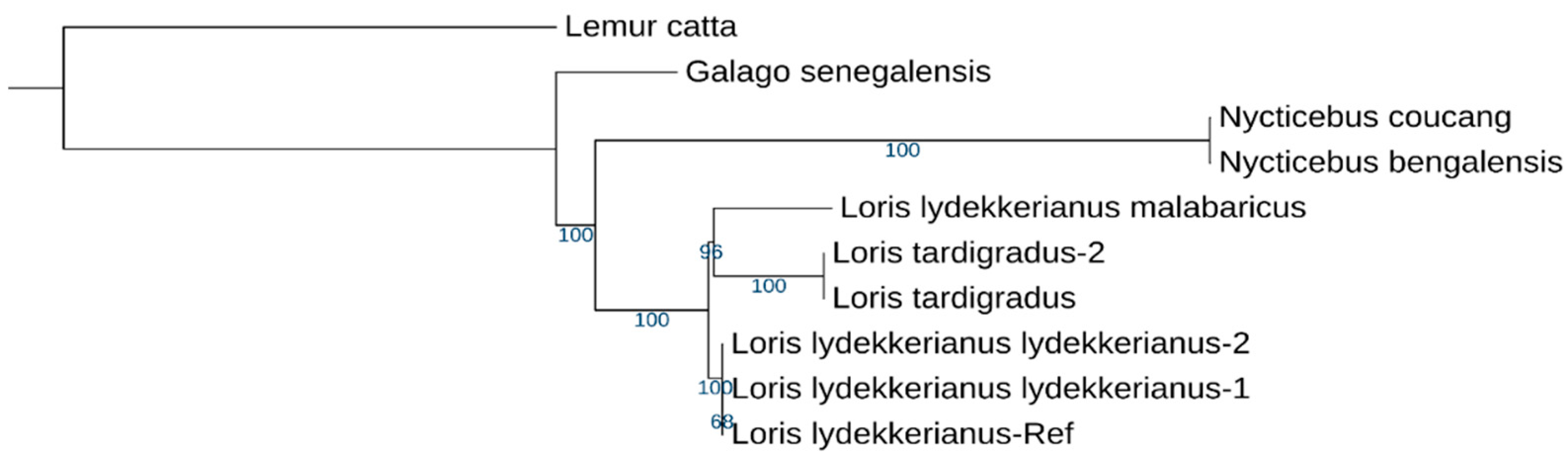
Figure 3.
Fossil-calibrated Bayesian inference tree showing the divergence time estimates in Mya. Bars indicate 95% CI.
Figure 3.
Fossil-calibrated Bayesian inference tree showing the divergence time estimates in Mya. Bars indicate 95% CI.
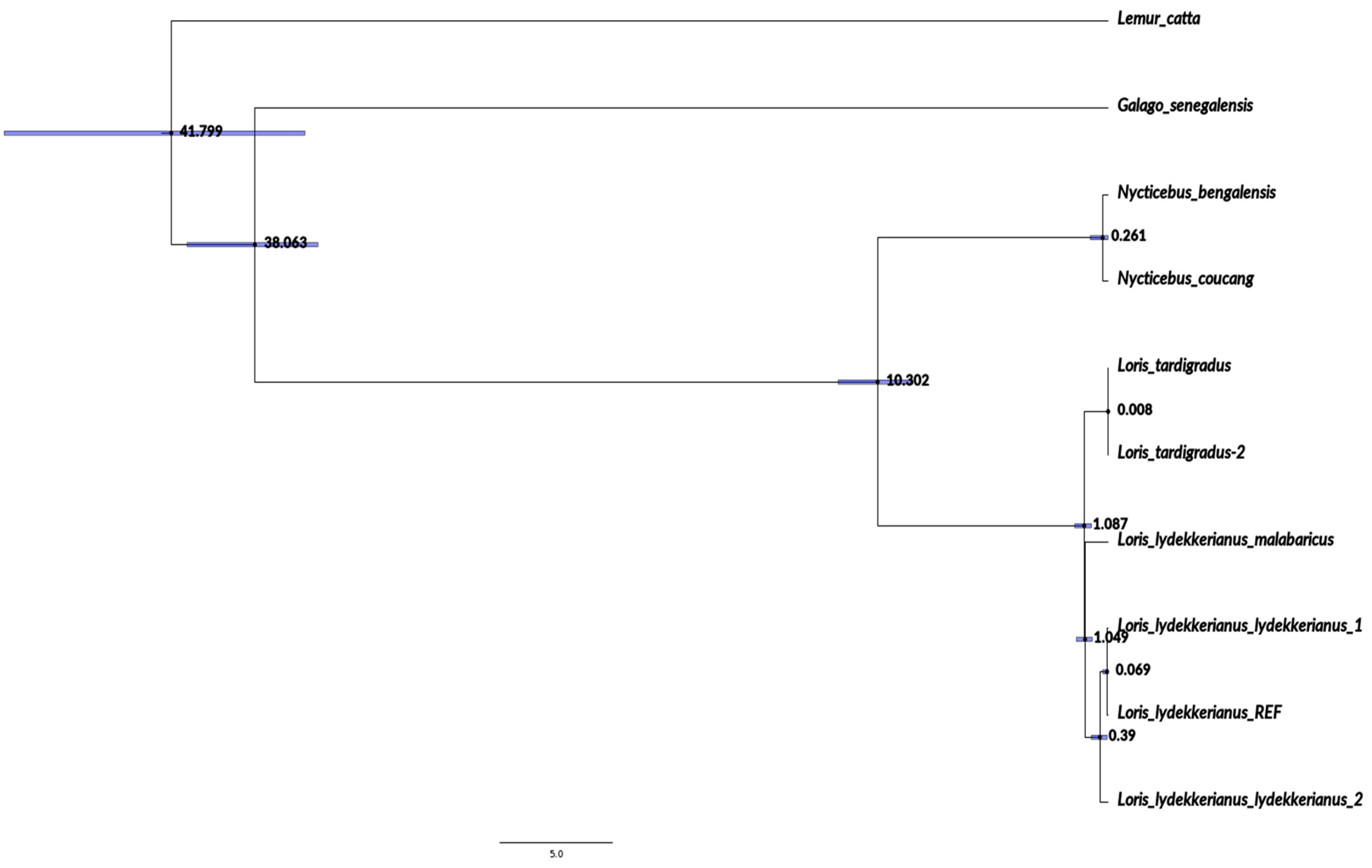

Table 1.
Sequence similarity matrix based on COX1 and CYTB genes in the mitochondrial genome.
| Sample |
Loris tardigradus |
Loris tardigradus-2 |
Loris lydekkerianus malabaricus |
Loris lydekkerianus lydekkerianus-1 |
Loris lydekkerianus lydekkerianus-2 |
Loris lydekkerianus-Ref |
|---|---|---|---|---|---|---|
| Loris tardigradus | - | 100 | 97.42 | 97.18 | 97.05 | 97.10 |
| Loris tardigradus-2 | 100 | - | 97.42 | 97.18 | 97.03 | 97.10 |
| Loris lydekkerianus malabaricus | 97.42 | 97.42 | - | 97.91 | 97.91 | 97.91 |
| Loris lydekkerianus lydekkerianus-1 | 97.18 | 97.18 | 97.91 | - | 99.84 | 99.92 |
| Loris lydekkerianus lydekkerianus-2 | 97.03 | 97.03 | 97.91 | 99.84 | - | 99.84 |
| Loris lydekkerianus-Ref | 97.10 | 97.10 | 97.91 | 99.92 | 99.84 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



