
Submitted:
31 March 2023
Posted:
03 April 2023
You are already at the latest version
Abstract
ATP-dependent chromatin remodeling complexes are involved in nucleosomes sliding, eviction and/or histone variants incorporation into chromatin to facilitate several cellular and biological processes, including DNA transcription, replication and repair. The DOM/TIP60 chromatin remodeling complex of Drosophila melanogaster contains 18 subunits, including the DOMINO (DOM), an ATPase that catalyzes the ex-change of the canonical H2A with its variant (H2A.V); and TIP60, a lysine-acetyltransferase that acetylates H4, H2A and H2A.V histones. In the last decade, different experimental evidence showed that ATP-dependent chromatin remodeling factors, in addition to their role in chromatin organization, have a functional relevance in cell division. In particular, emerging studies suggested direct roles of ATP-dependent chromatin remodeling complex subunits in controlling mitosis and cytokinesis in both humans and D. melanogaster. However, little is known about their possible involvement during meiosis Meiotic chromosomes non-disjunction led to aneuploid offspring, which are often inviable/poorly viable or sterile due to gene dosage imbalance. Therefore, studying the role of DOM/TIP60 complex in D. melanogaster meiosis can provide new insights on our understanding of the molecular mechanisms underlying cell division control in gametogenesis.
Keywords:
TIP60
; DOMINO
; ATPase
; Drosophila male meiosis
; chromatin remodeling
; cell division
; epigenetics
1. Introduction
ATP-dependent chromatin remodeling complexes use ATP hydrolysis to carry out nucleosomes sliding, eviction or incorporation of histone variants [1,2]. These events are required for several cellular and biological processes, including DNA transcription, replication and repair [3].
In Drosophila melanogaster, the DOM/TIP60 chromatin remodeling complex belongs to the INO80 family and it contains 18 subunits that provide a multitude of functions [1,4]. Major subunits of the DOM/TIP60 complex are DOMINO (DOM), an ATPase of the SWI2/SNF2 type that catalyzes the chromatin remodeling activity exchanging canonical H2A with its H2A variant (H2A.V), and TIP60, a lysine-acetyltransferase that acetylates H4, H2A and H2A.V histones [5,6,7,8,9,10]. In addition, the D. melanogaster DOM/TIP60 complex includes ACT87E, BAP55, BRD8, DOMINO (A and B), DMAP1, EAF6, E(PC), GAS41, ING3, MRG15, MRGBP, NIPPED-A, PONTIN, REPTIN, and YL1 subunits [11,12,13].
In the last decade, several lines of evidence indicated that ATP-dependent chromatin remodelling factors, in addition to their role in chromatin regulation, have a functional relevance in mitotic cell division [14,15,16,17,18,19,20,21,22,23,24]. A genome-wide RNAi screening in D. melanogaster Schneider 2 (S2) cells identified multiple members of several chromatin remodeling complexes as potential novel regulators of cell cycle. Specifically, DOM and YETI have been found near APC/C (anaphase-promoting complex/cyclosome) in a mitotic index-based network suggesting that these components interact each other [25]. Accordingly, the in vivo expression of CFDP1, the human ortholog of YETI, in D. melanogaster determines the formation of inactive heterodimers producing a strong dominant negative effect which affect cell proliferation and differentiation [24]. In the opposite way, the expression of YETI in HeLa cells decrease the mitotic index with an impairment of cell cycle progression [26,27]. Additional evidence has shown that RNAi-mediated depletion of BAP55 increases the incidence of multinucleated cells and results in spindle assembly defects [28]. More recently, DOM-A and MRG15 [15,29] were found to localize on both centrosomes and midbody, and their depletion lead to mitotic and cytokinesis defects in D. melanogaster S2 cells [14]. Similarly, TIP60 and YETI were found to the midbody too, and their knockdown lead to cytokinesis failure [15]. Moreover, the Domino gene products (DOM-A and DOM-B) are required for the normal asymmetric neuroblast (NB) division and they contribute, together with others TIP60 complex subunits, to NB maintenance and polarity [30]. Taken together, this evidence suggests a specific role for chromatin regulators during mitosis.
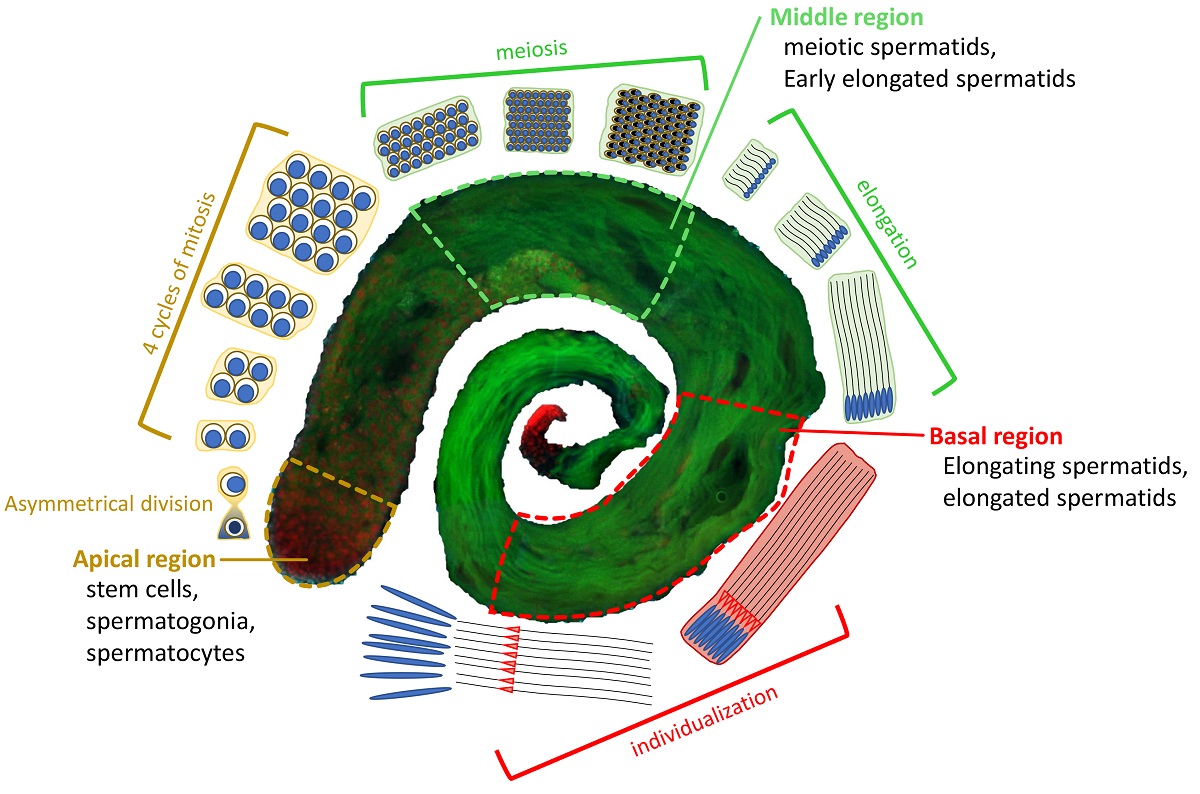
In contrast, little is known about the involvement of TIP60 chromatin remodeling complex during meiosis and spermatogenesis. In D. melanogaster, sperm cells production initiates from the asymmetric cell division of gonial stem cells (GSCs), generating a self-renewed GSC and a gonialblast cell (GC), which undergoes four-round mitosis as the transit-amplifying spermatogonia. After mitosis, 16 interconnected germ cells enter meiosis with a prolonged G2-phase as spermatocytes followed by two rounds of meiotic divisions, in which first, homologous chromosomes (meiosis I), and later sister chromatids (meiosis II) are segregated [31,32,33]. Haploid cells (called spermatids) subsequentially undergo nuclear elongation, compaction-protamination and individualization (Figure 1).
The challenge of meiosis is then to segregate both homologous chromosomes and sister chromatids accurately so that each gamete receives exactly one copy of each chromosome. When meiotic chromosomes do not properly segregate aneuploid gametes are generated, which can led to the formation of inviable or poorly viable zygotes due to gene dosage imbalance [34].
In this context, sporadic studies indicated an involvement of DOM/TIP60 complex subunits in meiotic cell divisions. Cenci, et al. [35] have reported failure of cytokinesis in meiotic cells of Yeti mutants. It has also been found that REPTIN and PONTIN function as dynein cytoplasmic assembly factors, highlighting their role in sperm motility and male fertility of D. melanogaster [36]. Moreover, E(PC) and NIPPED-A were found to play crucial roles during the early stages of fly germ cell development [37,38]. In particular, E(PC) promotes mitosis-to-meiosis transition in D. melanogaster male germline lineage [39].
The DOM/TIP60 complex was also identified as regulator in double-strand breaks (DSBs) repair response during oocytes development [40]. This process involves dynamic changes in chromatin structure with a continuously H2A.V/γH2A.V turnover at the break site until repair is finished. A complete absence of H2A.V, both phosphorylated and unphosphorylated, has been observed in MRG15 mutant germline clones suggesting that the MRG15 subunit is required for γH2A.V exchange during meiotic prophase [41]. Thus, a meiotic role for the DOM/TIP60 chromatin remodeling complex is worth exploring.
To deep our understanding of the roles played by DOM/TIP60 complex in D. melanogaster meiosis, the subcellular distribution of DOM/TIP60 complex subunits was studied. We found that the subunits under investigation (BAP55::HA, DMAP1::HA, DOMINO-A, MRG15, TIP60::HA, YETI, YL1::GFP) localized to sites of meiotic apparatus. Most notably, the down-regulation of 12 subunits under investigation (BAP55, DOMINO, DMAP1, EAF6, E(PC), GAS41, MRG15, MRGBP, PONTIN, REPTIN, YE
TI and YL1) affected chromosome segregation and cytokinesis. Based on these results, we hypothesized that DOM/TIP60 complex play crucial roles in different steps of male meiosis progression.
2. Materials and Methods
2.1. Fly stocks and genetics
All the stocks used in this work (Table S1) were raised on standard Bloomington formulation Drosophila medium at 25°C. w1; P{w+, Ubq11>EGFP::alphaTub84B}, P{w[+mC]=His2Av-mRFP1}; P{w+, bamP>GAL4VP16} (shortened w1; EGFP::αTub, H2A.V::mRFP; bam>Gal4) and w1; P{w+, Ubq11>EGFP::alphaTub84B}, P{Ubi-RFP-spd-2}; P{w+, bamP>GAL4VP16} (shortened w1; EGFP::αTub, Spd2::mRFP; bam>Gal4) were obtained by genetic recombination crosses between stocks carrying single transgene (Table S1).
2.2. Expression of tagged remodelers by using UAS/Gal4 system
Drosophila stocks for expressing BAP55::HA, DMAP1::HA and Tip60::HA proteins (UAS>HA-tagged) were purchased from FlyORF [42,43]. Virgins females carrying the UAS>[remodeler]::HA construct were crossed with males carrying the αTubulin84B>Gal4-VP16/TSTL, CyO:TM6B, Tb ubiquitous driver to trigger the expression of the HA-fused protein in all tissues. As negative control, virgin females w1 were crossed with males from the same driver stocks (αTubulin84B>Gal4-VP16/TSTL, CyO:TM6B, Tb) and males with only the driver were took in consideration for the analysis. Testes from F1 generation of adult males expressing or not (negative control) the HA-fused remodeller (UAS>[remodeller]::HA/αTubulin84B>Gal4) were dissected and fixed for immunofluorescence analysis by using anti-HA antibodies. The RNAi-mediated down-regulation of the targeted proteins in testes was performed with the bag of marbles-GAL4 (bam-GAL4), a spermatogonial specific driver [42,44]. In the case of UAS-Domino RNAi transgene (VDRC line 7787), the expressed shRNA induces the simultaneous silencing of both Domino transcripts coding for DOM-A and DOM-B isoforms.
2.3. Cytological analyses and Immunofluorescence
Testes of 1-day old adult males were dissected in TIB (testis isolation buffer): 183 mM KCl, 47 mM NaCl, 10 mM Tris pH 6.8.
Spermatids of fresh testes were analysed by a phase contrast objective of Nikon Eclipse 50i epifluorescence microscope to evaluate cytokinesis defects.
For fixed preparations, testes were placed in 8 μl of TIB and squashed on a slide, overlaid with a coverslip, and frozen in liquid nitrogen. Tissue was dehydrated in cold ethanol for 10’, then fixed for 7’ in 4% paraformaldehyde in phosphate buffered saline (PBS). Fixation was followed by a 30’ PBTx-DOC (0.3% Triton-X and 0.3% sodium deoxycholate in PBS) permeabilization, 10’ staining with 1 μg/ml DAPI (4,6-diamidino-2-phenylindole) in PBS and mounting in anti-fade medium (DABCO, Sigma). For immunofluorescence analyses, after permeabilization, slides were incubated with blocking solution (0.1% Triton X-100 and 5% FBS in PBS) for 30’ at room temperature in a moist chamber and then they were incubated overnight with primary antibodies dilutions (Table S2) at 4°C, washed and incubated with the secondary antibodies for 1h at room temperature in a moist chamber. Nuclei were stained with DAPI as described above. Testes preparations from a minimum of 3 controls and 3 RNAi-induced independent experiments were examined for each assay. Fluorescent cysts and spermatids were observed with a Nikon Eclipse 50i epifluorescence microscope equipped with a CCD camera. Images were acquired with NIS-Elements software provided by Nikon and processed using Adobe Photoshop (Adobe Systems, Mountain View, CA) and ImageJ software (http://rsbweb.nih.gov/ij/).
2.4. Statistical Analysis
Data analyses were performed using the GraphPad Prism software (GraphPad Software, Inc., La Jolla, CA, USA). All results are expressed as mean ±SD values from three independent replicate experiments. P value of less than 0.05 (*P < 0.05, compared with the control group) were considered to be statistically significant by using two-tailed Fisher’s exact test.
2.5. Bioinformatic analysis
3. Results
3.1. The subunits of DOM/TIP60 chromatin remodeling complex localize to the meiotic apparatus
Using immunofluorescence microscopy (IFM), we investigate the subcellular localization of 12 subunits of the Tip60 chromatin remodeling complex in meiotic cell division of D. melanogaster testes.
We have found that DOM-A and MRG15 show centrosomal localization (Figure 2B), while YETI appears to localize along the microtubular structure of the spindle (Figure 2B). To depict the meiotic localization of subunits for which no antibodies was available, the expression of BAP55::HA, DMAP1::HA and TIP60::HA proteins was induced by using the UAS-Gal4 binary expression system (see Materials and Methods) [43,48,49].
As shown in Figure 2C, the signals of BAP55::HA and DMAP1::HA fusion proteins were found to centrosomal structures, while the Tip60::HA signal localized along the meiotic spindle. Any specific signal is lost in negative control. To investigate the localization of the YL1 subunit, we used a stock expressing GFP::YL1-tagged protein under the native promoter. Using anti-GFP antibodies (Table S2), the GFP::YL1 signal showed a centrosomal localization, which is absent in the w1 negative controls (Figure 2D). Taken together, these data suggest a collective relocation of four DOM/Tip60 remodeling subunits from chromatin to meiotic apparatus.
3.2. RNAi depletion of DOM/TIP60 complex subunits
To study the knockdown effects of DOM/TIP60 chromatin remodeling complex subunits in male meiosis, specific RNA interference was activated by bag of marbles-GAL4 (bam-GAL4), a spermatogonial specific driver [43,44]. The eGFP::αTubulin and mRFP::H2A.V transgenes were used to fluorescently mark spindle and chromatin, respectively. Specifically, eGFP::αTubulin, mRFP::H2A.V; bam˃Gal4 virgin females were crossed to males from 12 RNAi stocks carrying interfering short-harpin to down-regulate the following DOM/TIP60 subunits: BAP55, DMAP1, DOMINO, E(PC), EAF6, GAS41, MRG15, PONTIN, REPTIN, TIP60, YETI, and YL1 (Table S1). The F1 progeny of these crosses was analysed for chromosome segregation and chromatin integrity. The efficiency of RNAi constructs was already checked by semi-quantitative PCR [4]. Following RNAi-mediated depletion of the subunits under investigation induced in testes, we found chromatin integrity defects (CID) including condensation and chromosome segregation issues with loss of chromatin fragments (Figure 3 A,B and Table 1). In agreement with the canonical function of DOM/TIP60 chromatin remodeling complex, we also found that the knockdown of some subunits (BAP55, DMAP1, EAF6, PONTIN, REPTIN, TIP60 and YETI) generates H2A.V mislocalization (HM), (Figure 3 C,D and Table 2). Similar defects were showed as a consequence of a loss of YETI protein during mitosis [35].
Taken together, these data suggested a role of DOM/TIP60 remodeling subunits in maintaining genome integrity not only in interphase but also during meiotic division.
3.3. RNAi-mediated depletion of TIP60 subunits affects spindle integrity
To further explore the possibility that DOM/TIP60 complex subunits could have a specific role in meiotic division, we analysed young RNAi-knockdown males harbouring.
eGFP::αTubulin, mRFP::H2A.V. As shown in Figure 4 A,B and Table 3, RNAi knockdown of DOM/Tip60 complex subunits lead to spindle morphology alteration, with the exception of BAP55 RNAi. In particular, the abnormal spindle defect can either be milder, as shown for DMAP1 RNAi in which the spindle structure is affected but a centrosomal signal is still perceptible, or stronger, as shown for GAS41 RNAi in which both spindle fibers and centrosomes are no longer distinguishable. In addition, in some cases (DOMINO, GAS41, MRG15 PONTIN, REPTIN, TIP60 and YL1), cells exhibited more of two spindle poles.
To investigate the occurrence of multipolar spindle defects as a consequence of RNAi-knockdown of DOM/TIP60 subunits, we have recombined the Spd2::mRFP centrosomal fluorescent marker with spindle marker (EGFP::αTub) and coupled with and Bam>Gal4 driver to achieve RNAi of DOM/TIP60 complex subunits in testes. Then, females w*; EGFP::αTub, Spd2::mRFP; bam>Gal4 were crossed to homozygous UAS>RNAi males and the testes of F1 progeny, carrying the three transgenes, were analysed. As shown in Figure 4 C,D and Table 4, the analysis of squash preparations knockdown showed the occurrence of a high percentage of multipolar meiotic spindles in DOM, GAS41, MRG15, PONTIN, REPTIN, TIP60 and YL1 depleted testis. Taken together, the results of these experiments suggested a requirement of DOM/TIP60 complex subunits in the maintenance of a proper spindle structure during meiotic division.
3.4. Cytokinesis Defects
We extended our analysis to cytokinesis, the crucial step in cell division giving rise to the two daughter cells following the final cut of the cytoplasmic bridge. Homozygous UAS>RNAi males were crossed to w1;; bam>Gal4 females and fresh testes squash preparations from the F1 progeny were analysed for cytokinesis defects during the two meiotic divisions, using phase-contrast microscopy. As shown in Figure 5A and Table 5, in the control experiment, wt spermatids physiologically showed one nucleus and one Nebenkern (mitochondrial derivative) with comparable sizes (Nu/Nk = 1/1). By contrast, in RNAi -treated samples, a significant percentage of spermatids containing 2 or 4 nuclei with only a bigger Nebenkern (Nu/Nk = 2/1 or 4/1) was observed. In most cases the aberrant ratio found was Nu/Nk = 2/1, suggesting that defective cytokinesis primarily occurs during the first meiotic division.
4. Discussion
We have recently found that subunits of SRCAP and p400/Tip60 complexes are recruited to mitotic apparatus in HeLa and MRC5 cells to ensure proper cell division [14,15]. Similarly, we have found that DOM-A, MRG15, TIP60, and YETI, four subunits of the evolutionary related DOM/TIP60 complex are recruited to mitotic apparatus and midbody with their depletion affecting both mitosis and cytokinesis in D. melanogaster S2 cells [14,15].
Here we provided evidence that a similar phenomenon may also occurs in vivo during the meiotic divisions in D. melanogaster males.
Previous studies have been reported that Drosophila H2A.V is essential to maintain chromosome structure in mitosis, and it might be also involved in both chromosome segregation and organization of kinetochore-driven k-fibers [50]. These defects are in line with chromatin integrity defects showed in Figure 2. In fact, the knock-down of BAP55, DMAP1, EAF6, PONTIN, REPTIN, TIP60 and YETI impaired with H2A.V localization in spermatocytes. However, while DOM-B and DOM-A isoforms play roles in H2A.V incorporation and removal, respectively during D. melanogaster oogenesis [50], in our experiments their depletion did not affect the H2A.V localization during male meiosis [50]. On the other hand, DOM-B and DOM-A seem to be crucial to prevent specific cell division defects such as abnormal spindle morphology, multipolar spindles and multinucleated cells (Figure 3,Figure 4,Figure 5/Table 1,Table 2,Table 3,Table 4,Table 5). While multipolar spindle formation was observed only following the depletion of a subset of subunits, abnormal spindle morphology and cytokinesis defects were found for all the tested subunits.
The different extent and quality of defects observed following depletion of the tested subunits may be ascribed to a different efficiency of the siRNA lines used in this work. It is also possible that the lack of certain subunits may have a milder impact on meiotic division compared to others.
Together, these results suggested that the entire complex play a crucial role in the meiotic spindle assembly possibly participating to microtubule polymerization and/or stabilization. Moreover, the observed meiotic defects strongly suggest that most of the subunits of DOM/TIP60 remodeling complex can cooperate in the control of several steps of meiotic cell division possibly maintaining their interactions during their relocation. Spermatogenesis is a finely regulated process generating highly polarized motile sperms (1.8 mm long) from small round cells (approximately 12 μm in diameter). This process can be dramatically affected by the failure of meiotic divisions. Our preliminary data suggest that RNAi depletion of some DOM/TIP60 complex subunits negatively impacts the physiological elongation of D. melanogaster sperms, thus affecting male fertility (data not shown). This matter gains importance in the light of the high level of sequence similarity between subunits of DOM/TIP60 complex in D. melanogaster and humans (Figure 6). In this perspective, haploinsufficiency mutation of human TIP60 subunits, not only can predispose to genetic instability and cancer onset [14,15], but may also affect meiosis and gametogenesis, thus reducing individual fertility.
5. Conclusions
In summary, these results highlight the intriguing possibility that subunits of the DOM/TIP60 complex, apart from the canonical functions in chromatin regulation, can maintain their interaction during the relocation to the meiotic apparatus and play essential roles in meiotic cell division to regulate cell cycle progression, centrosome function, and spindle organization/function.
Collectively, our results lead to conclude that the subunits of DOM/TIP60 complex performs extra-chromatin functions not only in mitosis, as shown by Messina et al., (2022), but also during the meiotic cell division. In this view, cell division dysfunctions, cancer and infertility appear to be closely interlinked, an aspect that deserves to be further explored by future studies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, P.D. and G.M.; methodology, P.D. and G.M.; software, Y.P. and D.F.; validation, Y.P., G.F. and D.F.; formal analysis, G.M.; investigation, Y.P., G.F. and D.F.; resources, P.D. and G.M.; data curation, Y.P., P.D. and G.M.; writing—original draft preparation, Y.P., G.F., D.F. and G.M.; writing—review and editing, Y.P., P.D. and G.M.; visualization, M.L.; supervision, P.D. and G.M.; project administration, G.M.; funding acquisition, G.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MIUR-PRIN 2017 (PD) and Pasteur Institute of Italy – Fondazione Cenci Bolognetti, “Teresa Ariaudo Research Program 2018” (GM).
Acknowledgments
We are grateful to Renata Basto for gift w1; P{w+, Ubq11>EGFP::alphaTub84B}/CyO stock; Dennis M McKearin for gift w1;; P{w+, bamP>GAL4VP16}III stocks; Jordan W. Raff for w1; P{Ubi-RFP-spd-2}/CyO stock; and Giovanni Bosco for gift of anti-MRG15 antibody.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Clapier, C.R.; Cairns, B.R. The biology of chromatin remodeling complexes. Annual review of biochemistry 2009, 78, 273–304. [Google Scholar] [CrossRef]
- Yodh, J. ATP-Dependent Chromatin Remodeling. Advances in experimental medicine and biology 2013, 767, 263–295. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Demajo, S.; Jain, P.; Di Croce, L. Combinatorial assembly and function of chromatin regulatory complexes. Epigenomics 2011, 3, 567–580. [Google Scholar] [CrossRef]
- Prozzillo, Y.; Cuticone, S.; Ferreri, D.; Fattorini, G.; Messina, G.; Dimitri, P. In Vivo Silencing of Genes Coding for dTip60 Chromatin Remodeling Complex Subunits Affects Polytene Chromosome Organization and Proper Development in Drosophila melanogaster. International journal of molecular sciences 2021, 22, 4525. [Google Scholar] [CrossRef] [PubMed]
- Kusch, T.; Florens, L.; Macdonald, W.H.; Swanson, S.K.; Glaser, R.L.; Yates, J.R., 3rd; Abmayr, S.M.; Washburn, M.P.; Workman, J.L. Acetylation by Tip60 is required for selective histone variant exchange at DNA lesions. Science 2004, 306, 2084–2087. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, G.; Shen, X.; Landry, J.; Wu, W.H.; Sen, S.; Wu, C. ATP-driven exchange of histone H2AZ variant catalyzed by SWR1 chromatin remodeling complex. Science 2004, 303, 343–348. [Google Scholar] [CrossRef]
- Keogh, M.C.; Mennella, T.A.; Sawa, C.; Berthelet, S.; Krogan, N.J.; Wolek, A.; Podolny, V.; Carpenter, L.R.; Greenblatt, J.F.; Baetz, K.; et al. The Saccharomyces cerevisiae histone H2A variant Htz1 is acetylated by NuA4. Genes Dev 2006, 20, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, L.; Lambert, J.P.; Gerdes, M.; Al-Madhoun, A.S.; Skerjanc, I.S.; Figeys, D.; Baetz, K. Functional dissection of the NuA4 histone acetyltransferase reveals its role as a genetic hub and that Eaf1 is essential for complex integrity. Mol Cell Biol 2008, 28, 2244–2256. [Google Scholar] [CrossRef]
- Morillo Prado, J.R.; Srinivasan, S.; Fuller, M.T. The histone variant His2Av is required for adult stem cell maintenance in the Drosophila testis. PLoS Genet 2013, 9, e1003903. [Google Scholar] [CrossRef]
- Borner, K.; Becker, P.B. Splice variants of the SWR1-type nucleosome remodeling factor Domino have distinct functions during Drosophila melanogaster oogenesis. Development 2016, 143, 3154–3167. [Google Scholar] [CrossRef]
- Messina, G.; Damia, E.; Fanti, L.; Atterrato, M.T.; Celauro, E.; Mariotti, F.R.; Accardo, M.C.; Walther, M.; Verni, F.; Picchioni, D.; et al. Yeti, an essential Drosophila melanogaster gene, encodes a protein required for chromatin organization. J Cell Sci 2014, 127, 2577–2588. [Google Scholar] [CrossRef] [PubMed]
- Prozzillo, Y.; Delle Monache, F.; Ferreri, D.; Cuticone, S.; Dimitri, P.; Messina, G. The True Story of Yeti, the "Abominable" Heterochromatic Gene of Drosophila melanogaster. Frontiers in physiology 2019, 10, 1093. [Google Scholar] [CrossRef] [PubMed]
- Messina, G.; Celauro, E.; Atterrato, M.T.; Giordano, E.; Iwashita, S.; Dimitri, P. The Bucentaur (BCNT) protein family: a long-neglected class of essential proteins required for chromatin/chromosome organization and function. Chromosoma 2015, 124, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Messina, G.; Prozzillo, Y.; Delle Monache, F.; Santopietro, M.V.; Atterrato, M.T.; Dimitri, P. The ATPase SRCAP is associated with the mitotic apparatus, uncovering novel molecular aspects of Floating-Harbor syndrome. BMC Biol 2021, 19, 184. [Google Scholar] [CrossRef] [PubMed]
- Messina, G.; Prozzillo, Y.; Monache, F.D.; Santopietro, M.V.; Dimitri, P. Evolutionary conserved relocation of chromatin remodeling complexes to the mitotic apparatus. BMC Biol 2022, 20, 172. [Google Scholar] [CrossRef] [PubMed]
- Messina, G.; Prozzillo, Y.; Bizzochi, G.; Marsano, R.M.; Dimitri, P. The Green Valley of Drosophila melanogaster Constitutive Heterochromatin: Protein-Coding Genes Involved in Cell Division Control. 2022, 11, 3058. [Google Scholar]
- Messina, G.; Celauro, E.; Marsano, R.M.; Prozzillo, Y.; Dimitri, P. Epigenetic Silencing of P-Element Reporter Genes Induced by Transcriptionally Active Domains of Constitutive Heterochromatin in Drosophila melanogaster. 2023, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Marsano, R.M.; Giordano, E.; Messina, G.; Dimitri, P. A New Portrait of Constitutive Heterochromatin: Lessons from Drosophila melanogaster. Trends Genet 2019, 35, 615–631. [Google Scholar] [CrossRef]
- Gartner, W.; Rossbacher, J.; Zierhut, B.; Daneva, T.; Base, W.; Weissel, M.; Waldhausl, W.; Pasternack, M.S.; Wagner, L. The ATP-dependent helicase RUVBL1/TIP49a associates with tubulin during mitosis. Cell motility and the cytoskeleton 2003, 56, 79–93. [Google Scholar] [CrossRef]
- Sigala, B.; Edwards, M.; Puri, T.; Tsaneva, I.R. Relocalization of human chromatin remodeling cofactor TIP48 in mitosis. Exp Cell Res 2005, 310, 357–369. [Google Scholar] [CrossRef]
- Park, E.J.; Hur, S.K.; Lee, H.S.; Lee, S.A.; Kwon, J. The human Ino80 binds to microtubule via the E-hook of tubulin: implications for the role in spindle assembly. Biochem Biophys Res Commun 2011, 416, 416–420. [Google Scholar] [CrossRef]
- Ducat, D.; Kawaguchi, S.; Liu, H.; Yates, J.R., 3rd; Zheng, Y. Regulation of microtubule assembly and organization in mitosis by the AAA+ ATPase Pontin. Mol Biol Cell 2008, 19, 3097–3110. [Google Scholar] [CrossRef]
- Zhang, S.M.; Song, M.; Yang, T.Y.; Fan, R.; Liu, X.D.; Zhou, P.K. HIV-1 Tat impairs cell cycle control by targeting the Tip60, Plk1 and cyclin B1 ternary complex. Cell Cycle 2012, 11, 1217–1234. [Google Scholar] [CrossRef] [PubMed]
- Gentili, C.; Castor, D.; Kaden, S.; Lauterbach, D.; Gysi, M.; Steigemann, P.; Gerlich, D.W.; Jiricny, J.; Ferrari, S. Chromosome Missegregation Associated with RUVBL1 Deficiency. PLoS One 2015, 10, e0133576. [Google Scholar] [CrossRef]
- Billmann, M.; Horn, T.; Fischer, B.; Sandmann, T.; Huber, W.; Boutros, M. A genetic interaction map of cell cycle regulators. Mol Biol Cell 2016, 27, 1397–1407. [Google Scholar] [CrossRef]
- Messina, G.; Atterrato, M.T.; Fanti, L.; Giordano, E.; Dimitri, P. Expression of human Cfdp1 gene in Drosophila reveals new insights into the function of the evolutionarily conserved BCNT protein family. Scientific reports 2016, 6, 25511. [Google Scholar] [CrossRef]
- Messina, G.; Atterrato, M.T.; Prozzillo, Y.; Piacentini, L.; Losada, A.; Dimitri, P. The human Cranio Facial Development Protein 1 (Cfdp1) gene encodes a protein required for the maintenance of higher-order chromatin organization. Scientific reports 2017, 7, 45022. [Google Scholar] [CrossRef] [PubMed]
- Echard, A.; Hickson, G.R.; Foley, E.; O'Farrell, P.H. Terminal cytokinesis events uncovered after an RNAi screen. Curr Biol 2004, 14, 1685–1693. [Google Scholar] [CrossRef] [PubMed]
- Scacchetti, A.; Schauer, T.; Reim, A.; Apostolou, Z.; Campos Sparr, A.; Krause, S.; Heun, P.; Wierer, M.; Becker, P.B. Drosophila SWR1 and NuA4 complexes are defined by DOMINO isoforms. eLife 2020, 9. [Google Scholar] [CrossRef]
- Rust, K.; Tiwari, M.D.; Mishra, V.K.; Grawe, F.; Wodarz, A. Myc and the Tip60 chromatin remodeling complex control neuroblast maintenance and polarity in Drosophila. EMBO J 2018, 37. [Google Scholar] [CrossRef] [PubMed]
- Page, S.L.; Hawley, R.S. Chromosome choreography: the meiotic ballet. Science 2003, 301, 785–789. [Google Scholar] [CrossRef]
- Petronczki, M.; Siomos, M.F.; Nasmyth, K. Un menage a quatre: the molecular biology of chromosome segregation in meiosis. Cell 2003, 112, 423–440. [Google Scholar] [CrossRef]
- Frappaolo, A.; Piergentili, R.; Giansanti, M.G. Microtubule and Actin Cytoskeletal Dynamics in Male Meiotic Cells of Drosophila melanogaster. Cells 2022, 11. [Google Scholar] [CrossRef]
- McKee, B.D.; Yan, R.; Tsai, J.H. Meiosis in male Drosophila. Spermatogenesis 2012, 2, 167–184. [Google Scholar] [CrossRef]
- Cenci, G.; Belloni, G.; Dimitri, P. 1(2)41Aa, a heterochromatic gene of Drosophila melanogaster, is required for mitotic and meiotic chromosome condensation. Genet Res 2003, 81, 15–24. [Google Scholar] [CrossRef]
- Zur Lage, P.; Stefanopoulou, P.; Styczynska-Soczka, K.; Quinn, N.; Mali, G.; von Kriegsheim, A.; Mill, P.; Jarman, A.P. Ciliary dynein motor preassembly is regulated by Wdr92 in association with HSP90 co-chaperone, R2TP. J Cell Biol 2018, 217, 2583–2598. [Google Scholar] [CrossRef]
- Feng, L.; Shi, Z.; Chen, X. Enhancer of polycomb coordinates multiple signaling pathways to promote both cyst and germline stem cell differentiation in the Drosophila adult testis. PLoS Genet 2017, 13, e1006571. [Google Scholar] [CrossRef]
- McCarthy, A.; Deiulio, A.; Martin, E.T.; Upadhyay, M.; Rangan, P. Tip60 complex promotes expression of a differentiation factor to regulate germline differentiation in female Drosophila. Mol Biol Cell 2018, 29, 2933–2945. [Google Scholar] [CrossRef]
- Feng, L.; Shi, Z.; Xie, J.; Ma, B.; Chen, X. Enhancer of polycomb maintains germline activity and genome integrity in Drosophila testis. Cell Death Differ 2018, 25, 1486–1502. [Google Scholar] [CrossRef]
- Baldi, S.; Becker, P.B. The variant histone H2A.V of Drosophila-three roles, two guises. Chromosoma 2013, 122, 245–258. [Google Scholar] [CrossRef]
- Joyce, E.F.; Pedersen, M.; Tiong, S.; White-Brown, S.K.; Paul, A.; Campbell, S.D.; McKim, K.S. Drosophila ATM and ATR have distinct activities in the regulation of meiotic DNA damage and repair. J Cell Biol 2011, 195, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Bischof, J.; Bjorklund, M.; Furger, E.; Schertel, C.; Taipale, J.; Basler, K. A versatile platform for creating a comprehensive UAS-ORFeome library in Drosophila. Development 2013, 140, 2434–2442. [Google Scholar] [CrossRef]
- Bischof, J.; Sheils, E.M.; Bjorklund, M.; Basler, K. Generation of a transgenic ORFeome library in Drosophila. Nat Protoc 2014, 9, 1607–1620. [Google Scholar] [CrossRef]
- Consortium, T.U. UniProt: the universal protein knowledgebase in 2021. Nucleic Acids Research 2020, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res 2019, 47, W636–W641. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: recent updates, new developments and status in 2020. Nucleic Acids Research 2020, 49, D458–D460. [Google Scholar] [CrossRef]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. DOG 1.0: illustrator of protein domain structures. Cell research 2009, 19, 271–273. [Google Scholar] [CrossRef]
- Duffy, J.B. GAL4 system in Drosophila: a fly geneticist's Swiss army knife. Genesis 2002, 34, 1–15. [Google Scholar] [CrossRef]
- Elliott, D.A.; Brand, A.H. The GAL4 system : a versatile system for the expression of genes. Methods Mol Biol 2008, 420, 79–95. [Google Scholar] [CrossRef]
- Verni, F.; Cenci, G. The Drosophila histone variant H2A.V works in concert with HP1 to promote kinetochore-driven microtubule formation. Cell Cycle 2015, 14, 577–588. [Google Scholar] [CrossRef]
Figure 1.
Overview of D. melanogaster spermatogenesis. At the Apical region of a testis, a hub of germline stem cells (GSCs) divides asymmetrically into two cells: a daughter stem cell and a differentiating gonialblast (GB). The GB goes through four cycles of mitosis to form a cyst of 16 primary spermatocytes. Primary spermatocytes will proceed through meiosis resulting in the generation of 64 roundish haploid spermatids (Middle region). The latter undergo elongation processes characterized by changes in nuclear shape and chromatin condensation to form indi-vidualized mature sperm (Basal region), which are stored to the seminal vesicle until fertilization. EGFP::α-Tubulin84D in green, and H2A.V::mRFP in red.
Figure 1.
Overview of D. melanogaster spermatogenesis. At the Apical region of a testis, a hub of germline stem cells (GSCs) divides asymmetrically into two cells: a daughter stem cell and a differentiating gonialblast (GB). The GB goes through four cycles of mitosis to form a cyst of 16 primary spermatocytes. Primary spermatocytes will proceed through meiosis resulting in the generation of 64 roundish haploid spermatids (Middle region). The latter undergo elongation processes characterized by changes in nuclear shape and chromatin condensation to form indi-vidualized mature sperm (Basal region), which are stored to the seminal vesicle until fertilization. EGFP::α-Tubulin84D in green, and H2A.V::mRFP in red.
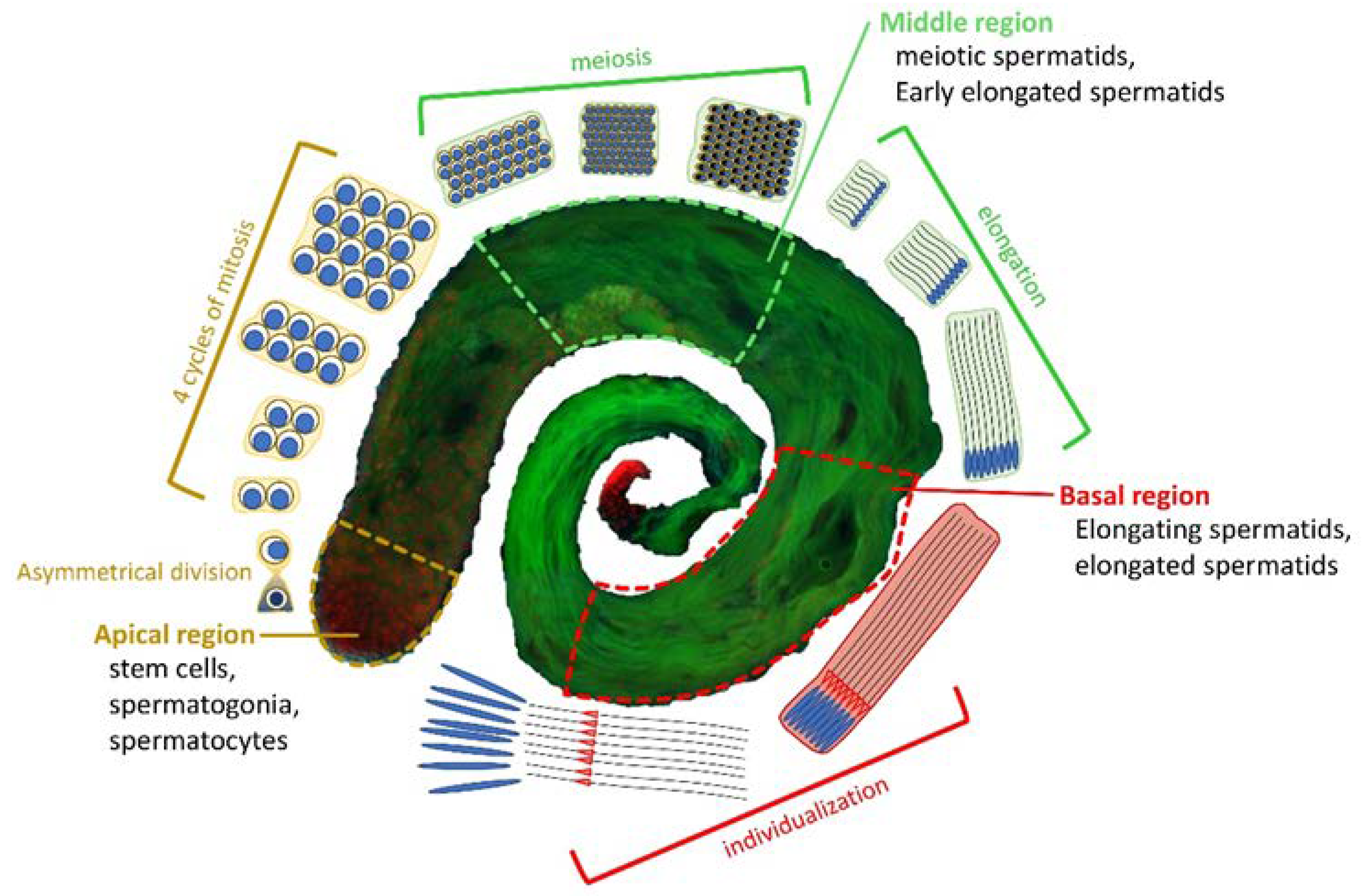
Figure 2.
Dynamic localization of DOM/Tip60 chromatin remodeling complex subunits in meiosis. (A) Graphical representation of DOMINO/DOM/Tip60 remodeling complex. Subunits are not in scale. (B, C, D) Immuno-localization of DOMINO-A, MRG15, Yeti, BAP55, DMAP1, TIP60 and YL1. Testes of young adult, 1-3 days from EGFP::αTub; Bam>Gal4 crossed with UAS>protein-HA tag and UAS>protein-GFP tag, were stained with specific-antibody (in red) while αTubulin is endogenously fluorescent (in green). DNA is stained with DAPI (in blue). DOMINO-A, MRG15, BAP55, DMAP1 and YL1 showed a specific localization to centrosomes, YETI showed a spindle localization while Tip60 showed a signal along the mitotic spindle.
Figure 2.
Dynamic localization of DOM/Tip60 chromatin remodeling complex subunits in meiosis. (A) Graphical representation of DOMINO/DOM/Tip60 remodeling complex. Subunits are not in scale. (B, C, D) Immuno-localization of DOMINO-A, MRG15, Yeti, BAP55, DMAP1, TIP60 and YL1. Testes of young adult, 1-3 days from EGFP::αTub; Bam>Gal4 crossed with UAS>protein-HA tag and UAS>protein-GFP tag, were stained with specific-antibody (in red) while αTubulin is endogenously fluorescent (in green). DNA is stained with DAPI (in blue). DOMINO-A, MRG15, BAP55, DMAP1 and YL1 showed a specific localization to centrosomes, YETI showed a spindle localization while Tip60 showed a signal along the mitotic spindle.
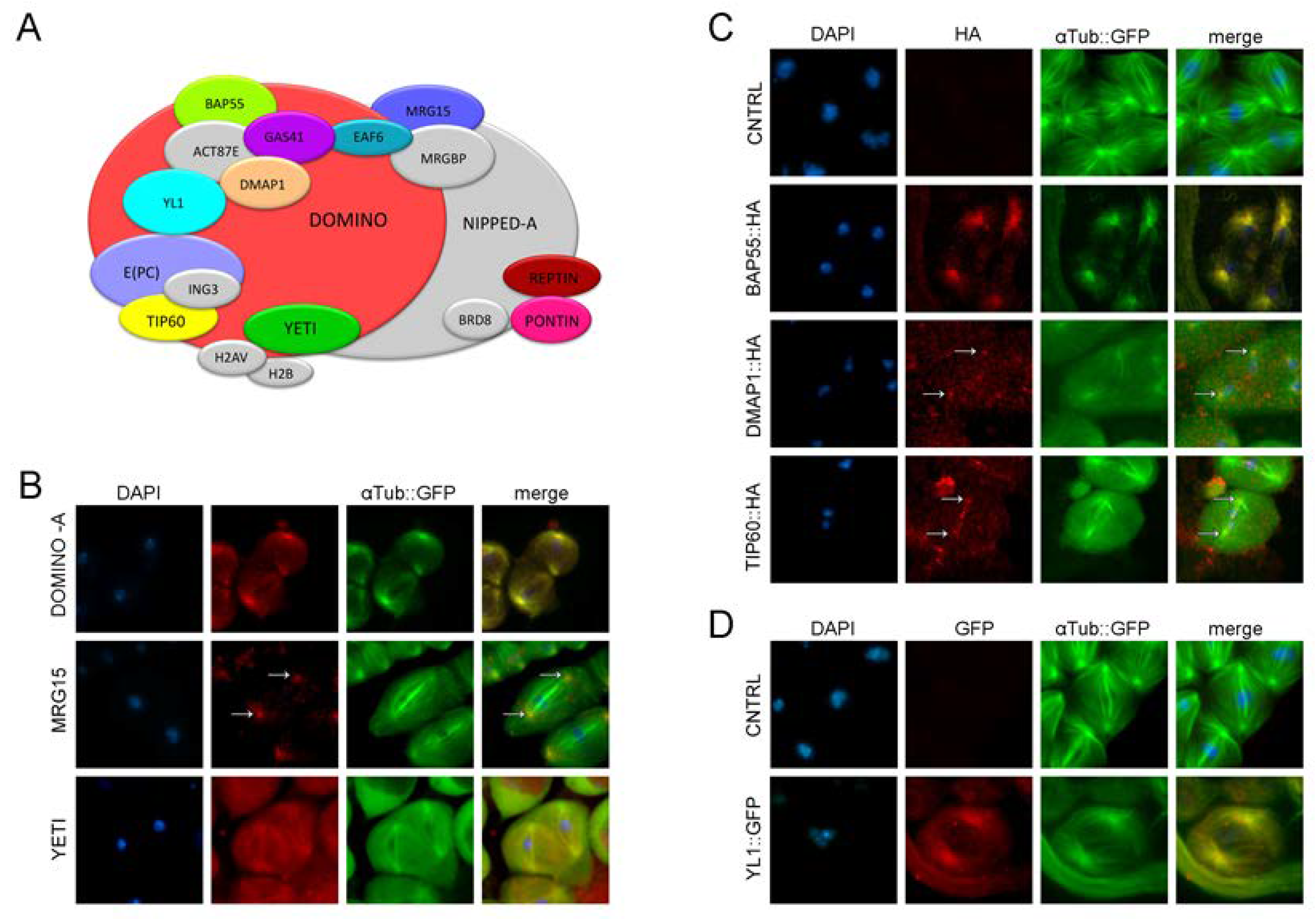
Figure 3.
Chromatin integrity defects (CID) and H2A.V mislocalization (HM) defects induced by RNAi in meiosis. Cytological analysis of testis squashes preparation from EGFP::αTub, H2A.V::mRFP; Bam>Gal4 driver crossed with specific remodelers RNAi construct. DNA is stained with DAPI (in blue), EGFP::αTub (in green) and H2A.V::mRFP (in red). (A) For Tip60, MRG15 and E(pc) the white arrow indicates chromatin fragments probably loss during segregation. (B) Quantification analysis of CID after RNAi knockdown effects activated by the EGFP::αTub, H2A.V::mRFP; Bam>Gal4 driver. n= number of analysed cells: Mock (n=72), BAP55 (n=128), DMAP1 (n=260), DOMINO (n=98), E(PC) (n=113), EAF6 (n=289), GAS41 (n=196), MRG15 (n=82), PONTIN (n=281), REPTIN (n=339), TIP60 (n=116), YETI (n=79) and YL1 (n=102). (C) H2A.V mislocalization is reported for YETI, PONTIN and EAF6 as a widespread nuclear signal compared to Control sample. (D) Quantification analysis of HM after RNAi knockdown effects activated by the EGFP::αTub, H2A.V::mRFP; Bam>Gal4 driver. n= number of analysed cells: Mock (n=148), BAP55 (n=138), DMAP1 (n=132), DOMINO (n=64), E(PC) (n=124), EAF6 (n=184), GAS41 (n=195), MRG15 (n=70), PONTIN (n=119), REPTIN (n=85), TIP60 (n=43), YETI (n=154) and YL1 (n=82). The statistical analysis is performed by using two-tailed Fisher’s exact test (* = Pvalue = 0,05, ** = Pvalue ≤ 0,005, ***= Pvalue ≤ 0,0005).
Figure 3.
Chromatin integrity defects (CID) and H2A.V mislocalization (HM) defects induced by RNAi in meiosis. Cytological analysis of testis squashes preparation from EGFP::αTub, H2A.V::mRFP; Bam>Gal4 driver crossed with specific remodelers RNAi construct. DNA is stained with DAPI (in blue), EGFP::αTub (in green) and H2A.V::mRFP (in red). (A) For Tip60, MRG15 and E(pc) the white arrow indicates chromatin fragments probably loss during segregation. (B) Quantification analysis of CID after RNAi knockdown effects activated by the EGFP::αTub, H2A.V::mRFP; Bam>Gal4 driver. n= number of analysed cells: Mock (n=72), BAP55 (n=128), DMAP1 (n=260), DOMINO (n=98), E(PC) (n=113), EAF6 (n=289), GAS41 (n=196), MRG15 (n=82), PONTIN (n=281), REPTIN (n=339), TIP60 (n=116), YETI (n=79) and YL1 (n=102). (C) H2A.V mislocalization is reported for YETI, PONTIN and EAF6 as a widespread nuclear signal compared to Control sample. (D) Quantification analysis of HM after RNAi knockdown effects activated by the EGFP::αTub, H2A.V::mRFP; Bam>Gal4 driver. n= number of analysed cells: Mock (n=148), BAP55 (n=138), DMAP1 (n=132), DOMINO (n=64), E(PC) (n=124), EAF6 (n=184), GAS41 (n=195), MRG15 (n=70), PONTIN (n=119), REPTIN (n=85), TIP60 (n=43), YETI (n=154) and YL1 (n=82). The statistical analysis is performed by using two-tailed Fisher’s exact test (* = Pvalue = 0,05, ** = Pvalue ≤ 0,005, ***= Pvalue ≤ 0,0005).
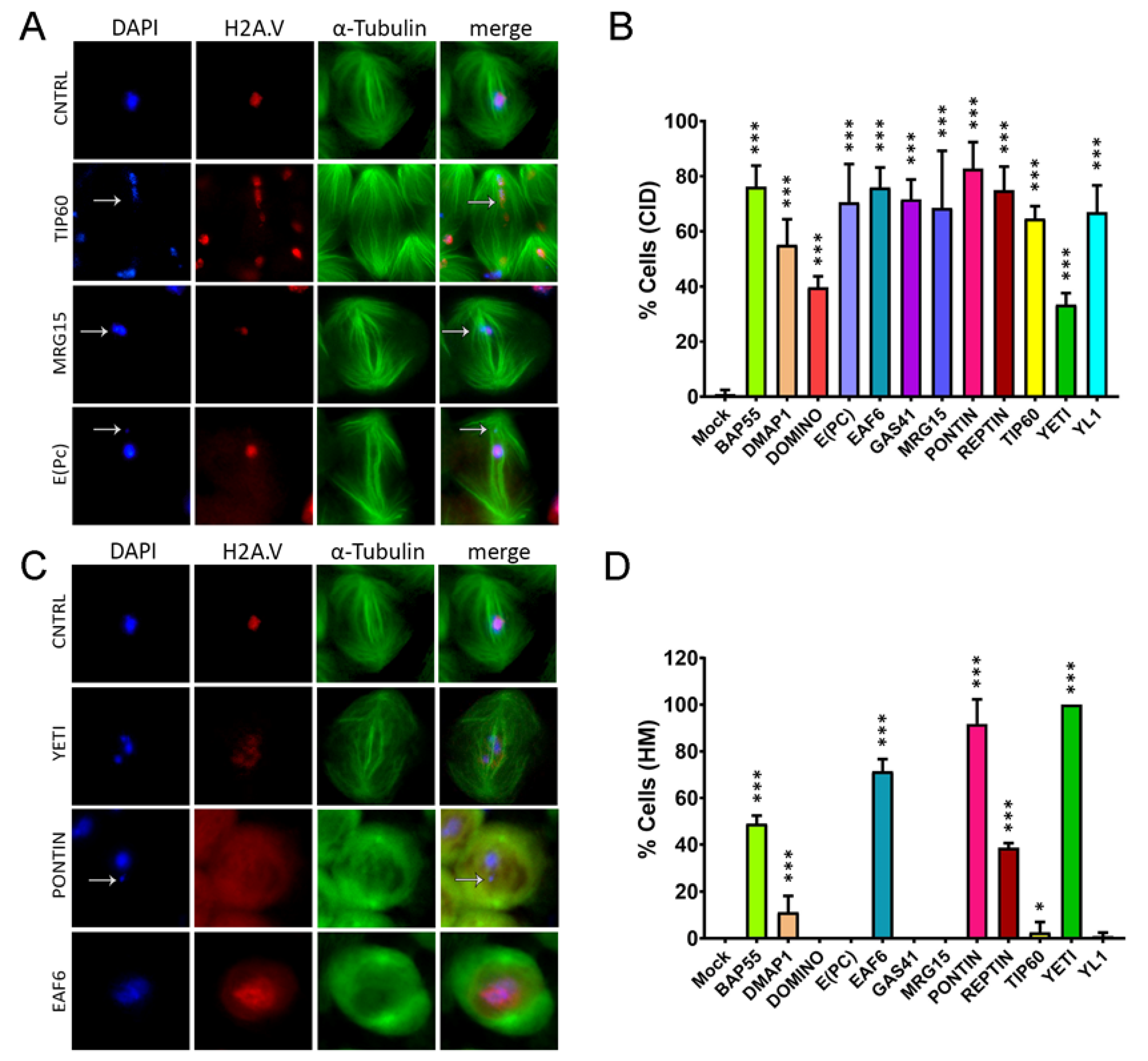
Figure 4.
Abnormal spindle morphology (ASM) and Multipolar spindle (MS) defects induced by RNAi in meiosis. Testis squashes preparation from Bam>Gal4 driver strain crossed with specific subunit RNAi construct. DNA is stained with DAPI (in blue), EGFP::αTub (in green) and H2A.V::mRFP (in red). (A) Early chromatin decondensation effects are shown for DMAP1 knockdown, with also a widespread signal for DNA and H2A.V, and for GAS41 knockdown with also detached chromatin fragments from the central plate. B) Quantification analysis of AS after RNAi knockdown effects activated by the EGFP::αTub, H2A.V::mRFP; Bam>Gal4 driver. n= number of analysed cells: Mock (n=85), BAP55 (n=74), DMAP1 (n=58), DOMINO (n=113), E(PC) (n=28), EAF6 (n=32), GAS41 (n=31), MRG15 (n=68), PONTIN (n=53), REPTIN (n=69), TIP60 (n=65), YETI (n=87) and YL1 (n=63). C) Alteration of spindle structure, here shown for DOMINO and TIP60 subunits from squashed testis of young adult flies, 1-3 days, from EGFP::αTub, Spd2::mRFP; Bam>Gal4 driver crossed with specific subunit RNAi construct. DNA is stained with DAPI (in blue), EGFP::αTub (in green) and Spd2::mRFP (in red). White arrows indicate two centrosomes in the control sample while in the interfered samples for TIP60 and DOMINO multiple centrosomes are noticeable. D) Quantification analysis of MS after RNAi knockdown effects activated by the EGFP::αTub, Spd2::mRFP; Bam>Gal4 driver. n= number of analysed cells: Mock (n=199), BAP55 (n=131), DMAP1 (n=246), DOMINO (n=328), E(PC) (n=189), EAF6 (n=147), GAS41 (n=181), MRG15 (n=96), PONTIN (n=119), REPTIN (n=152), TIP60 (n=162), YETI (n=130) and YL1 (n=192). The statistical analysis is performed by using two-tailed Fisher’s exact test (* = Pvalue = 0,05, ** = Pvalue ≤ 0,005, ***= Pvalue ≤ 0,0005).
Figure 4.
Abnormal spindle morphology (ASM) and Multipolar spindle (MS) defects induced by RNAi in meiosis. Testis squashes preparation from Bam>Gal4 driver strain crossed with specific subunit RNAi construct. DNA is stained with DAPI (in blue), EGFP::αTub (in green) and H2A.V::mRFP (in red). (A) Early chromatin decondensation effects are shown for DMAP1 knockdown, with also a widespread signal for DNA and H2A.V, and for GAS41 knockdown with also detached chromatin fragments from the central plate. B) Quantification analysis of AS after RNAi knockdown effects activated by the EGFP::αTub, H2A.V::mRFP; Bam>Gal4 driver. n= number of analysed cells: Mock (n=85), BAP55 (n=74), DMAP1 (n=58), DOMINO (n=113), E(PC) (n=28), EAF6 (n=32), GAS41 (n=31), MRG15 (n=68), PONTIN (n=53), REPTIN (n=69), TIP60 (n=65), YETI (n=87) and YL1 (n=63). C) Alteration of spindle structure, here shown for DOMINO and TIP60 subunits from squashed testis of young adult flies, 1-3 days, from EGFP::αTub, Spd2::mRFP; Bam>Gal4 driver crossed with specific subunit RNAi construct. DNA is stained with DAPI (in blue), EGFP::αTub (in green) and Spd2::mRFP (in red). White arrows indicate two centrosomes in the control sample while in the interfered samples for TIP60 and DOMINO multiple centrosomes are noticeable. D) Quantification analysis of MS after RNAi knockdown effects activated by the EGFP::αTub, Spd2::mRFP; Bam>Gal4 driver. n= number of analysed cells: Mock (n=199), BAP55 (n=131), DMAP1 (n=246), DOMINO (n=328), E(PC) (n=189), EAF6 (n=147), GAS41 (n=181), MRG15 (n=96), PONTIN (n=119), REPTIN (n=152), TIP60 (n=162), YETI (n=130) and YL1 (n=192). The statistical analysis is performed by using two-tailed Fisher’s exact test (* = Pvalue = 0,05, ** = Pvalue ≤ 0,005, ***= Pvalue ≤ 0,0005).
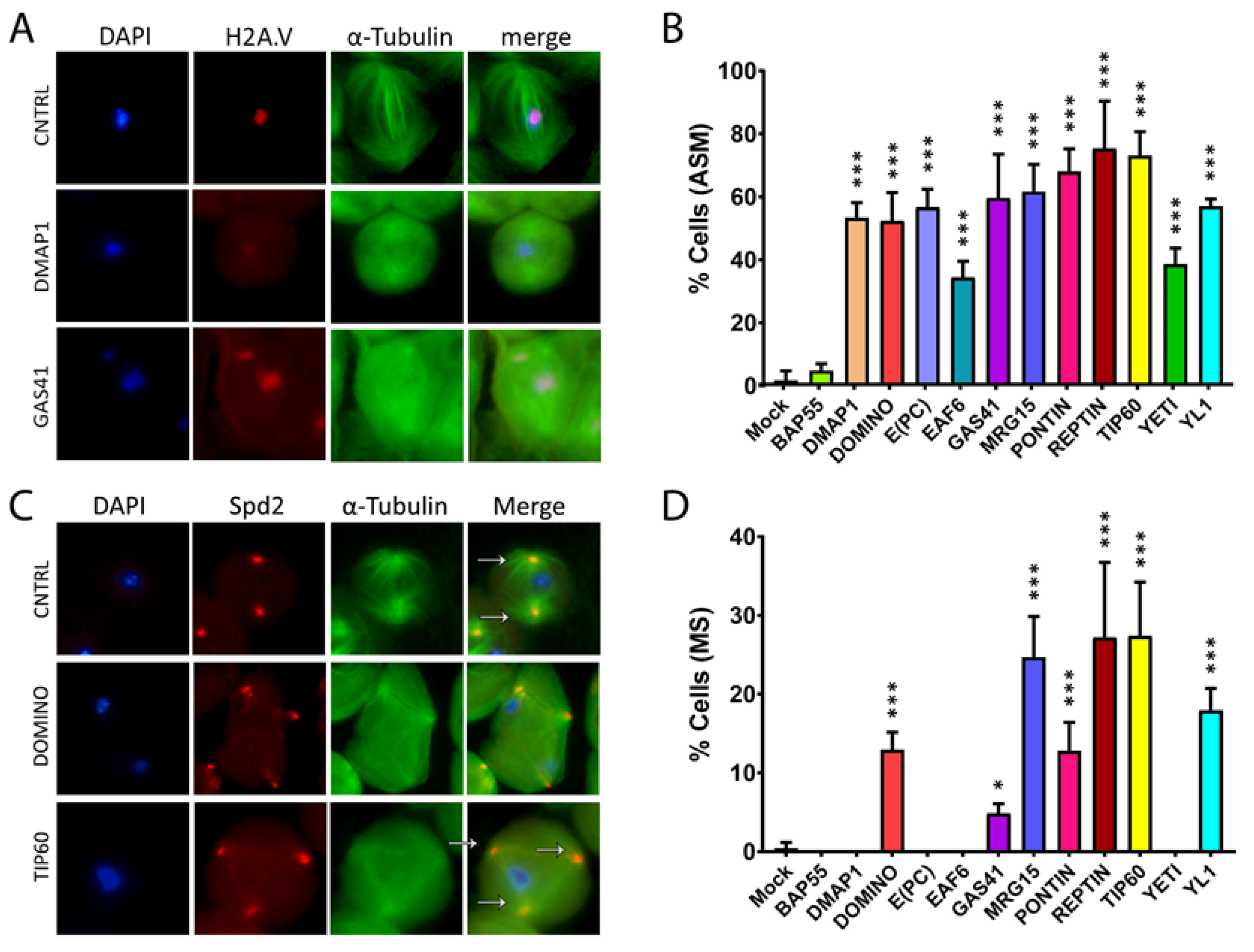
Figure 5.
Cytokinesis defects (CD) induced by RNAi-knockdown of DOM/TIP60 complex subunits in meiosis. A) Difference between normal onion-stage cells, in which the ratio between Nucleus (lighter grey circles) and nebenkern (black circle) is equal to 1:1 with same volume (on the right), while in case of cytokinesis defect the ratio become 2:1 or more and the nucleus volume become smaller than the Nebenkern (on the left). Phase contrast microscopy. B) Quantification analysis of CD after RNAi knockdown effects activated by the Bam>Gal4 driver. n= number of analysed cells: Mock (n=1231), BAP55 (n=772), DMAP1 (n=1327), DOMINO (n=1061), E(PC) (n=595), EAF6 (n=1118), GAS41 (n=1242), MRG15 (n=993), PONTIN (n=922), REPTIN (n=968), TIP60 (n=1013), YETI (n=940) and YL1 (n=1228). The statistical analysis is performed by using two-tailed Fisher’s exact test (* = Pvalue = 0,05, ** = Pvalue ≤ 0,005, ***= Pvalue ≤ 0,0005).
Figure 5.
Cytokinesis defects (CD) induced by RNAi-knockdown of DOM/TIP60 complex subunits in meiosis. A) Difference between normal onion-stage cells, in which the ratio between Nucleus (lighter grey circles) and nebenkern (black circle) is equal to 1:1 with same volume (on the right), while in case of cytokinesis defect the ratio become 2:1 or more and the nucleus volume become smaller than the Nebenkern (on the left). Phase contrast microscopy. B) Quantification analysis of CD after RNAi knockdown effects activated by the Bam>Gal4 driver. n= number of analysed cells: Mock (n=1231), BAP55 (n=772), DMAP1 (n=1327), DOMINO (n=1061), E(PC) (n=595), EAF6 (n=1118), GAS41 (n=1242), MRG15 (n=993), PONTIN (n=922), REPTIN (n=968), TIP60 (n=1013), YETI (n=940) and YL1 (n=1228). The statistical analysis is performed by using two-tailed Fisher’s exact test (* = Pvalue = 0,05, ** = Pvalue ≤ 0,005, ***= Pvalue ≤ 0,0005).
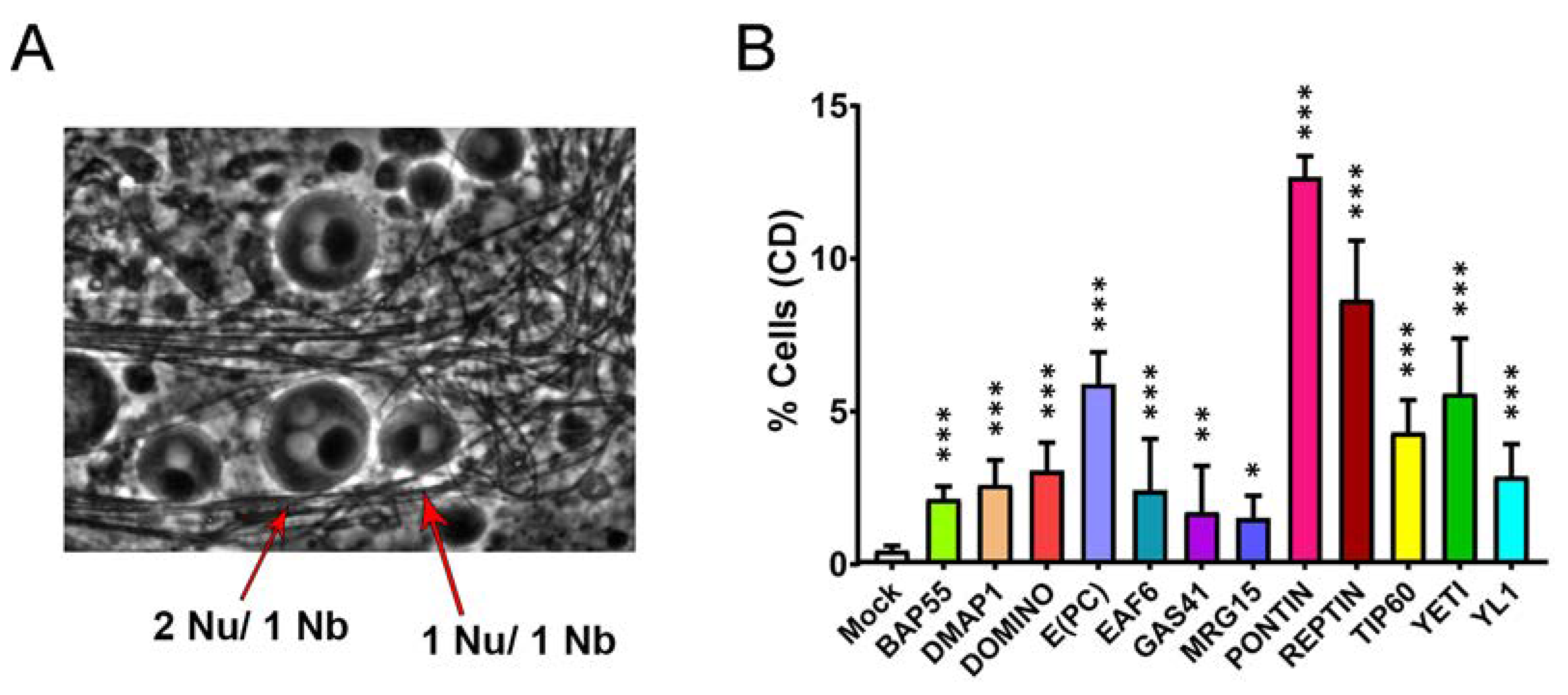
Figure 6.
Schematic representation of DOM/TIP60 chromatin remodeling subunits. Functional domains are coloury defined and details about their conservation in human orthologues are described in Table 6. Dimensions of proteins and domains are in scale.
Figure 6.
Schematic representation of DOM/TIP60 chromatin remodeling subunits. Functional domains are coloury defined and details about their conservation in human orthologues are described in Table 6. Dimensions of proteins and domains are in scale.
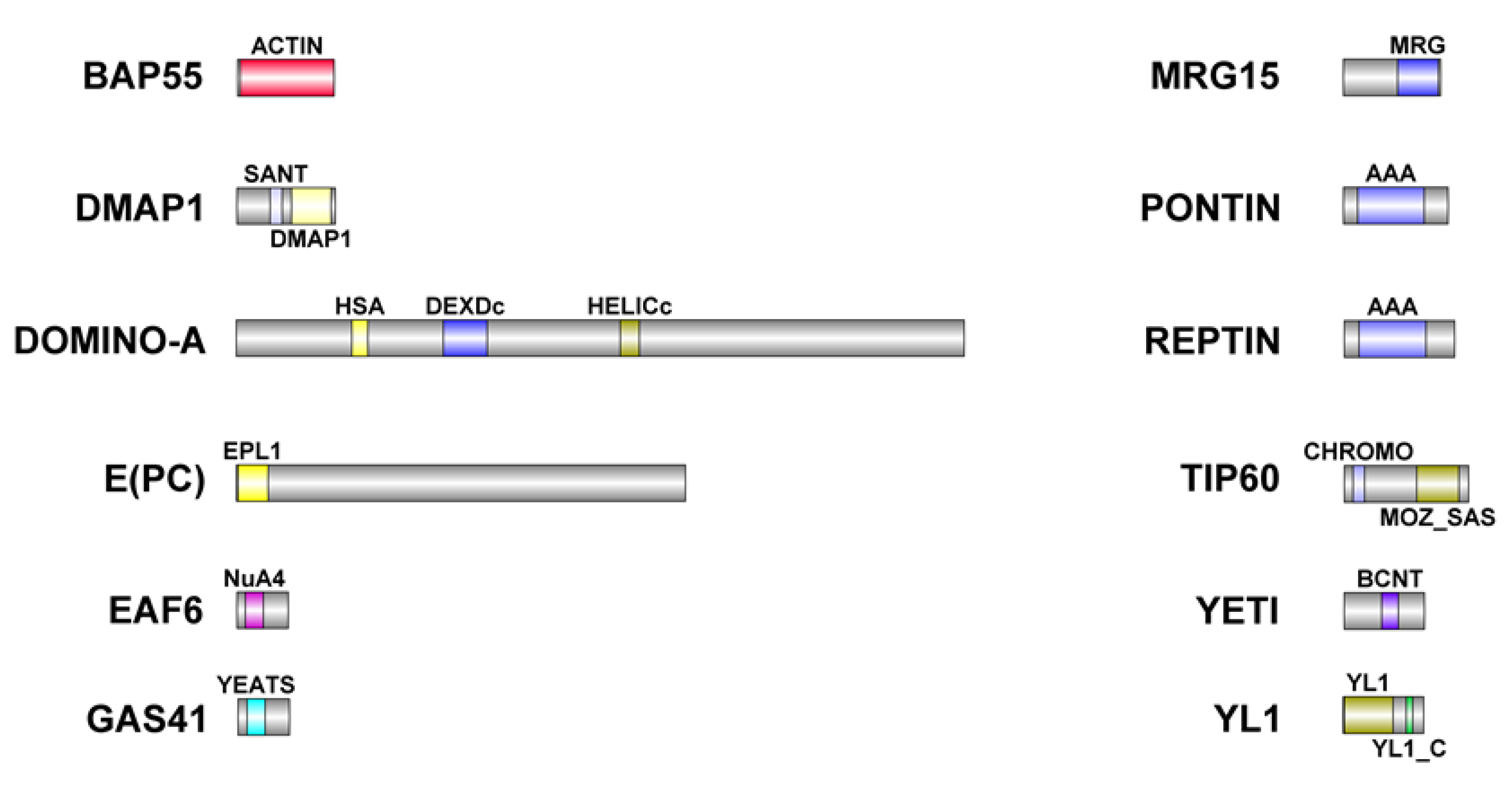

Table 1.
Quantification of chromatin integrity defects (CID).
| Control. | BAP55 | DMAP1 | DOMINO | E(PC) | EAF6 | GAS41 |
|---|---|---|---|---|---|---|
| 0.90 ± 1.56 | 76.20 ± 7.64 | 55.14 ± 9.23 | 39.69 ± 4.00 | 70.47 ± 13.93 | 75.96 ± 7.20 | 71.59 ± 7.21 |
| MRG15 | PONTIN | REPTIN | TIP60 | YETI | YL1 | |
| 68.49 ± 20.73 | 82.80 ± 9.58 | 74.88 ± 8.63 | 64.63 ± 4.50 | 33.37 ± 4.23 | 66.93 ± 9.64 |
Table 2.
H2A.V mislocalization defect (HM).
| Control | BAP55 | DMAP1 | DOMINO | E(PC) | EAF6 | GAS41 |
|---|---|---|---|---|---|---|
| 0 ± 0 | 0 ± 0 | 7.69 ± 4.44 | 0 ± 0 | 0 ± 0 | 71.41 ± 5.27 | 0 ± 0 |
| MRG15 | PONTIN | REPTIN | TIP60 | YETI | YL1 | |
| 0 ± 0 | 91.79 ± 10.47 | 38.66 ± 2.01 | 2.56 ± 4.44 | 100 ± 0 | 0.90 ± 1.56 |
Table 3.
Aberrant Spindle Morphology (ASM).
| Control | BAP55 | DMAP1 | DOMINO | E(PC) | EAF6 | GAS41 |
|---|---|---|---|---|---|---|
| 1.75 ± 3.04 | 4.74 ± 2.26 | 53.42 ± 4.74 | 52.37 ± 8.98 | 56.67 ± 5.77 | 34.44 ± 5.09 | 59.60 ± 13.94 |
| MRG15 | PONTIN | REPTIN | TIP60 | YETI | YL1 | |
| 61.69 ± 8.63 | 68.14 ± 7.09 | 75.32 ± 15.10 | 73.04 ± 7.65 | 38.53 ± 5.18 | 56.98 ± 2.36 |
Table 4.
Multipolar Spindle (MS).
| Control | BAP55 | DMAP1 | DOMINO | E(PC) | EAF6 | GAS41 |
|---|---|---|---|---|---|---|
| 0.43 ± 0.74 | 0 ± 0 | 0 ± 0 | 12.93 ± 2.25 | 0 ± 0 | 0 ± 0 | 4.84 ± 1.25 |
| MRG15 | PONTIN | REPTIN | TIP60 | YETI | YL1 | |
| 24.72 ± 5.14 | 12.81 ± 3.60 | 27.18 ± 9.54 | 27.37 ± 6.88 | 0 ± 0 | 17.93 ± 2.81 |
Table 5.
Cytokinesis defects (CD).
| Control | BAP55 | DMAP1 | DOMINO | E(PC) | EAF6 | GAS41 |
|---|---|---|---|---|---|---|
| 0.44 ± 0.16 | 2.13 ± 0.41 | 2.60 ± 0.81 | 3.05 ± 0.93 | 5.89 ± 1.04 | 2.41 ± 0.63 | 1.70 ± 1.51 |
| MRG15 | PONTIN | REPTIN | TIP60 | YETI | YL1 | |
| 1.52 ± 0.72 | 12.68 ± 0.68 | 8.64 ± 1.94 | 4.31 ± 1.07 | 5.59 ± 1.80 | 2.87 ± 1.06 |
Table 6.
Conserved domains in DOM/TIP60 remodeling complex subunits.
| D. melanogaster | H. sapiens | Domain | Identity (%) | Similarity (%) |
|---|---|---|---|---|
| BAP55 (425aa) | ACTL6A (429aa) | ACTIN | 54 | 71,2 |
| DMAP1 (433aa) | DMAP1 (467aa) | SANT | 55,6 | 79,6 |
| DMAP1 | 47,4 | 68,6 | ||
| DOMINO-A (3198aa) | SRCAP (3230aa) | HSA | 47,2 | 72,2 |
| DEXDc | 86,1 | 93,3 | ||
| HELICc | 91,7 | 95,2 | ||
| E(PC) (1974aa) | EPC1 (834aa) | EPL1 | 1,4 | 2,8 |
| EPC2 (807aa) | EPL1 | 5,7 | 17,8 | |
| EAF6 (225aa) | MEAF6 (191aa) | NuA4 | 77,5 | 90 |
| GAS41 (227aa) | YEATS (227aa) | YEATS | 77,8 | 86,4 |
| MRG15 (424aa) | MORF4L1 (362aa) | MRG | 51,4 | 71,8 |
| Tudor-knot | 46,3 | 59,3 | ||
| PONTIN (456aa) | RUVBL1 (456aa) | AAA | 85,5 | 92,1 |
| REPTIN (481aa) | RUVBL2 (467aa) | AAA | 82,3 | 93,2 |
| TIP60 (543aa) | KAT5 (513aa) | CHROMO | 59,6 | 82,7 |
| MOZ_SAS | 81,1 | 86,5 | ||
| YETI (241aa) | CFDP1 (299aa) | BCNT | 49,3 | 76 |
| YL1 (351aa) | VPS72 (364aa) | YL1 | 46,6 | 60,2 |
| YL1_C | 50 | 70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



