
Submitted:
02 April 2023
Posted:
03 April 2023
You are already at the latest version
Abstract
To meet the growing interest in the development of innovative biological nutrient removal (BNR) alternatives for energy neutrality, resource recovery, and decarbonization, the adsorption/bio-oxidation (A/B) process has been widely studied for carbon capture and shortcut nitrogen (N) removal, while limited research has focused on incorporating enhanced biological phosphorus (P) removal (EBPR), mainly due to the differential carbon usage characteristics of functionally relevant microorganisms. Here, a full-scale pilot testing with an integrated system combining A-stage high-rate activated sludge (HRAS) with B-stage partial (de)nitrification/anammox and side-stream EBPR (HRAS-P(D)N/A-S2EBPR) was conducted treating real municipal wastewater. The results demonstrated that, despite the low influent carbon load, the B-stage P(D)N-S2EBPR system achieved effective and stable P removal performance, especially when the volatile fatty acid (VFA) load from A-stage was elevated. Sludge fermentation in both A-stage and B-stage promoted carbon redistribution and likely provided more competitive advantages for ammonium-oxidizing bacteria and polyphosphate accumulating organisms, leading to carbon-efficient shortcut N removal mainly through partial nitrification pathway and influent carbon-independent EBPR simultaneously. Exposure to high VFA levels was considered a potential selection factor for the suppression of nitrite-oxidizing bacteria in the system. The involvement of internal carbon-accumulating organisms would potentially play an important role in endogenous denitrification. This study provided new insights into the effects of incorporating side-stream EBPR into the A/B process on microbial ecology, metabolic activities, and system performance.
Keywords:
side-stream EBPR
; shortcut nitrogen removal
; partial nitrification
; endogenous denitrification
; microbial ecology
1. Introduction
The motivation for energy neutrality, resource recovery, and decarbonization in the water sector has led to the exploration of novel wastewater treatment schemes towards carbon capture, energy recovery, and efficient nutrient removal and recovery with less energy/external carbon requirements and carbon emissions (Ren and Pagilla 2022, Wan et al. 2016). At present, several energy-efficient and even energy-positive technologies, such as shortcut nitrogen (N) removal processes (Lackner et al. 2014, Ma et al. 2016), coupled aerobic-anoxic nitrous decomposition operation (CANDO) (Scherson et al. 2014), microalgae cultivation (Chen et al. 2015), and anaerobic membrane bioreactors (Smith et al. 2012), have been proposed to replace traditional biological N removal processes (i.e., nitrification-denitrification) in municipal wastewater treatment plants (WWTPs) that are typically energy intensive and require additional carbon. Among them, the adsorption/bio-oxidation (A/B) process, where A-stage diverts most organic matter to anaerobic digestion for carbon capture and bioenergy recovery, while the subsequent B-stage aims for carbon- and energy-efficient N removal, is one of the promising engineering innovations that can meet the needs of energy and nutrient management (Guthi et al. 2022, Liu et al. 2019). Specifically, at the B-stage, the partial nitrification/anammox (PN/A) process, which relies on the cooperation of ammonium-oxidizing bacteria (AOB) and anaerobic AOB (AnAOB) and the suppression of nitrite-oxidizing bacteria (NOB) for completely autotrophic N removal via nitrite pathway, has attracted extensive attentions due to its largely reduced demands for aeration energy, external carbon, and waste sludge treatment (Cao et al. 2017, Ma et al. 2016). Side-stream PN/A has been widely studied and employed in many WWTPs for high-strength digestor centrate treatment, while mainstream PN/A application is still limited and faces challenges in stability (Cao et al. 2017, Lackner et al. 2014, Li et al. 2020b). Alternatively, partial denitrification/anammox (PDN/A) was demonstrated as a more practically viable route for mainstream anammox, although more studies are still needed for its full-scale demonstration and optimization (Ali et al. 2021, Zhang et al. 2019).
However, most of the shortcut N processes, such as PN/A or PDN/A, focused primarily on N removal via nitrite, and did not incorporate and/or optimize for biological phosphorus (P) removal, particularly enhanced biological P removal (EBPR). To date, only a few studies have attempted to incorporate mainstream PN/A with EBPR (Cao et al. 2016, Jia et al. 2023), which is usually due to the presumed incompatibility between these two processes (Roots et al. 2020, Yang et al. 2017). For instance, a favorable carbon-to-phosphorus (C/P) ratio and alternating anaerobic-aerobic/anoxic conditions are required for the enrichment of polyphosphate (polyP) accumulating organisms (PAOs) or denitrifying PAOs (DPAOs) performing EBPR, which are hardly satisfied in the B-stage PN/A process with limited influent organics from A-stage and deficiency/absence of deep anaerobic zone. The supplemental carbon sources added to the B-stage influent for EBPR, however, would potentially compromise NOB suppression, undermining the stability of shortcut N removal performance (Jia et al. 2023, Li et al. 2020b). Recently, the successful implementation of side-stream EBPR (S2EBPR) processes, which involve a side-stream anaerobic reactor for simultaneous PAO enrichment and volatile fatty acid (VFA) generation via hydrolysis and fermentation of return activated sludge (RAS) and/or mixed liquor, has promised the potential to enable stable EBPR performance with more flexible configurations without direct dependence on influent carbon (Barnard et al. 2017, Gu et al. 2019, Onnis-Hayden et al. 2020, Wang et al. 2019). Compared to conventional EBPR, S2EBPR offers unique conditions that favor PAOs over GAOs, which holds the key to EBPR performance and stability, by introducing a more complex composition of VFAs (i.e., a mixture of acetate and propionate) that leads to higher PAO diversity (Onnis-Hayden et al. 2020, Srinivasan et al. 2021, Wang et al. 2019), and consequently, different P release/uptake kinetics. In addition, extended anaerobic conditions in S2EBPR further give PAOs competitive advantages due to their distinct maintenance and delayed decay associated with the versatility of metabolic pathways and the possession of multiple intracellular polymers (i.e., PHA, glycogen, and polyP) (Li et al. 2020a). Furthermore, S2EBPR allows for more flexible carbon supplement and improved carbon utilization efficiency, and thus EBPR can be achieved for systems under previously considered unfavorable C/P ratios (Sabba et al. 2023, Wang et al. 2019). Therefore, we hypothesized that the S2EBPR process could potentially be incorporated into the A/B process for achieving shortcut N removal and efficient P removal/recovery simultaneously.
Recently, a full-scale pilot testing with the incorporation of S2EBPR into an A/B process, defined as an integrated system combining A-stage high-rate activated sludge (HRAS) with B-stage partial (de)nitrification/anammox and side-stream EBPR (i.e., HRAS-P(D)N/A-S2EBPR), was conducted at the Chesapeake-Elizabeth Sewage Treatment Plant (Virginia Beach, VA, USA) to assess the feasibility of this newly established system (Figure 1). To achieve nitrite accumulation, a series of continuous stirred-tank reactors (CSTRs) were intermittently aerated at the same interval and duration via ammonia versus NOx–-N (nitrite+nitrate) (AvN) control strategy (Regmi et al. 2014). To enhance P removal, the S2EBPR reactor was operated in a side-stream RAS fermentation with supplemental carbon (SSRC) configuration (Wang et al. 2019), with A-stage wasted activated sludge (WAS) fermentate as a supplement. To the best of our knowledge, this was the first demonstration and comprehensive investigation of a pilot-scale HRAS-P(D)N/A-S2EBPR system for mainstream shortcut N removal and side-stream EBPR treating real mainstream municipal wastewater. The unique configuration and operating conditions are expected to exert distinct selection forces from conventional EBPR on the microbial ecology of the system. However, several fundamental issues related to processing design and operation, as well as the underlying mechanisms, remain unresolved regarding the coupling of A/B process with S2EBPR, including 1) How would operating conditions in the S2EBPR reactor (e.g., sludge retention time (SRT), hydraulic retention time (HRT), oxidation-reduction potential (ORP), etc.), which differ from the conventional EBPR, affect AOB/NOB growth and activity? 2) What effects would intermittently aerated CSTRs at varying dissolved oxygen (DO) concentrations have on PAOs/DPAOs and their competitors, glycogen accumulating organisms (GAOs)/denitrifying GAOs (DGAOs)? 3) Is there any effect on the functionally relevant microorganisms, particularly PAOs/DPAOs, GAOs/DGAOs, AOB, NOB, and denitrifying bacteria (DNB), when the A-stage WAS fermentate was introduced into the S2EBPR reactor as the additional carbon load?
Therefore, this study aimed to comprehensively and systematically evaluate the feasibility, microbial ecology, and relevant mechanisms involved in the incorporation of S2EBPR into the novel HRAS-P(D)N/A-S2EBPR system. In addition to routine monitoring of influent and effluent water quality, mass balances of carbon, N, and P, metabolic activities batch tests, and microbial community analyses via 16S rRNA gene amplicon sequencing and single-cell Raman micro-spectroscopy (SCRS) were performed. The outcome will improve our fundamental understanding of this unique HRAS-P(D)N/A-S2EBPR process, the associated microbial community structure and phenotypes, and elucidate the key factors and mechanisms that govern process optimization.
2. Materials and methods
2.1. Pilot plant setup and operating conditions
The pilot plant is located at the Chesapeake-Elizabeth Sewage Treatment Plant (Virginia Beach, VA, USA) operated by Hampton Roads Sanitation District (HRSD). Details of this pilot plant were reported by Klaus (2019). The plant was operated in an A/B process (Figure 1), receiving pre-screened and temperature-controlled municipal wastewater at 20℃ and a flow rate of 3.2 m3/d (Printz 2019). The A-stage consists of three HRAS reactors in series with a total volume of 511 L, followed by an intermediate clarifier. The supernatant of the A-stage clarifier flows into the B-stage as its influent. The B-stage comprises a 53 L anaerobic reactor and four CSTRs in series with a total volume of 600 L, operated under AvN intermittent aeration control. A 174 L side-stream anaerobic reactor (S2EBPR) was introduced in B-stage. A fraction of RAS (~20–30%) from the B-stage clarifier was split into S2EBPR for sludge fermentation and PAO enrichment, with A-stage WAS fermentate as supplemental carbon. The S2EBPR is intermittently mixed for a duration of 1 min every 3 h, yielding an estimated side-stream SRT (SRTSS) of ~7–10 h and a side-stream HRT (HRTSS) of ~3.7 h. The effluent from the B-stage clarifier then passes through a 340 L anammox moving bed biofilm reactor (MBBR) loaded with K3 biofilm carriers (AnoxKaldnes, Sweden) for accomplishing shortcut N removal. The whole process is named HRAS-P(D)N/A-S2EBPR, while the focused processes in this study, including mainstream anaerobic reactor and CSTRs for shortcut N removal via PN and/or PDN, coupled with the side-stream anaerobic reactor for EBPR, is defined as the P(D)N-S2EBPR system.
The pilot testing period for this study was conducted from June 13th, 2019 to March 26th, 2020. The long-term operation was divided into three phases based on the major operational differences in total carbon loading level and corresponding performance: Phase 1 (start-up at low carbon load, Day 1–68), Phase 2 (nitrite accumulation at medium carbon load, Day 69–194), and Phase 3 (nitrite accumulation and EBPR at high carbon load, Day 195–288). Detailed influent and operating parameters of the system were summarized in Table S1. The performance of the system during the pilot testing period was routinely monitored by measuring chemical oxygen demand (COD), soluble COD (sCOD), total phosphorus (TP), orthophosphate (PO43–-P), total Kjeldahl nitrogen (TKN), nitrate-nitrogen (NO3–-N), nitrite-nitrogen (NO2–-N), ammonium-nitrogen (NH4+-N), etc. in the influent, effluent, and each zone of the system. The sampling parameters, sites, frequency, and dates are summarized in Table S2. The nitrite accumulation ratio (NAR) was calculated by the ratio of effluent NO2–-N to the sum of effluent NO2–-N and NO3–-N.
2.2. Metabolic activity batch tests
To evaluate the EBPR activity, anaerobic-aerobic batch tests were conducted on-site weekly following the previously described protocol (Gu et al. 2008). Briefly, fresh activated sludge withdrawn from the 4th aerobic tank at B-stage (CSTR-4) was initially aerated for 1 h to remove residual organics. Sodium acetate (HAc) of ~100 mg COD/L was then added and the sludge was continuously purged with nitrogen gas to maintain an anaerobic condition (DO < 0.01 mg/L). After 2 h of anaerobic conditions, the sludge was bubbled with air to obtain an aerobic condition (DO > 2.0 mg/L) for 3 h. The pH and temperature in the batch test were maintained at 7.0 and 20°C, respectively. Samples were collected every 30 min, filtered by 0.45 μm filter, and then analyzed for sCOD, PO43–-P, NH4+-N, NO2–-N, and NO3–-N, etc. To investigate the effect of different carbon sources on PAO activity, activated sludge samples and A-stage WAS fermentate (fermentate) in the P(D)N-S2EBPR system were collected at the end of the pilot testing and immediately shipped to Cornell lab. Either fermentate or HAc was utilized as the carbon source and added at the beginning of the anaerobic-aerobic batch test. Simultaneously, the presence of denitrifying PAOs (DPAOs) was evaluated by carrying out an anaerobic-anoxic batch test with fermentate and NO3–-N as carbon source and electron acceptor, respectively. DPAO activity and fraction of DPAOs over total PAOs were calculated according to previous literature (Ferrentino et al. 2018, Hu et al. 2003).
To evaluate the activities of nitrifying bacteria (i.e., AOB and NOB), ex-situ nitrification batch tests were routinely carried out. Briefly, ~4 L pre-aerated activated sludge from CSTR-4 was spiked with ammonium chloride at a concentration of 25 mg N/L. The pH and DO in the batch test were maintained at ~7.0 and ~3–5 mg/L, respectively. To evaluate the activities of ordinary denitrifiers (i.e., DNB), ~2 L RAS from B-stage clarifier was mixed with A-stage effluent of ~100–150 mg COD/L and NO3–-N of 20 mg N/L. Samples were taken every 15 min for a total of 1 h, and then analyzed for NH4+-N, NO2–-N, NO3–-N, and sCOD concentrations. AOB, NOB, and DNB activities were calculated by linear regression as the NOx–-N production rate, NO3–-N production rate, and NO3–-N reduction rate, respectively.
2.3. Microbial community analysis
The B-stage microbial community was periodically analyzed via 16S rRNA gene amplicon sequencing following the method described by Wang et al. (2019). Briefly, genomic DNA was extracted using the DNeasy PowerSoil Pro Kit (Qiagen, Germantown, MD, USA). The extracted DNA was amplified targeting the V4 region of the 16S rRNA gene using the primers 515F (5'-GTGCCAGCMGCCGCGGTAA-3') and 806R (5'-GGACTACHVGGGTWTCTAAT-3') and the amplicons were sequenced on the Illumina MiSeq platform using the paired-end (2 × 250 bp) option in the Microbial Analyses, Resources, and Service (MARS) facility at the University of Connecticut (Storrs, CT, USA). Raw reads were preprocessed following the standard operating procedure by Mothur software (Schloss et al. 2009). High-quality reads were then clustered at a 97% similarity to obtain the operational taxonomic units (OTUs) and annotated based on the Silva database (Quast et al. 2012). Non-metric multidimensional scaling (NMDS) with distance bray was plotted for the microbial community analysis using the meta MDS function from the vegan package (R Core Team 2013).
2.4. Single-cell Raman micro-spectroscopy-based phenotyping
To quantify PAOs and GAOs based on their phenotypic characteristics in the microbial community, activated sludge samples were collected at the start of anaerobic phase (t = 0 min), end of anaerobic phase (t = 120 min), and end of aerobic phase (t = 300 min) of the anaerobic-aerobic batch tests and prepared for single-cell Raman micro-spectroscopy (SCRS) analysis, as described previously (Majed et al. 2012, Wang et al. 2021). The detailed SCRS analysis protocol related to sample pretreatment, data acquisition, and processing was described in the Supplementary Information.
2.5. Chemical analyses
The concentrations of COD, PO43–-P, NO3–-N, NO2–-N, NH4+-N, mixed liquor suspended solids (MLSS), and mixed liquor volatile suspended solids (MLVSS) were analyzed according to the Standard Methods (APHA. 2012) or USEPA methods or using HACH kits (HACH, Loveland, CO, USA). VFAs, including acetic, propionic, n-butyric, isobutyric, n-valeric, isovaleric, and caproic acids, were analyzed using high-performance liquid chromatography (HPLC). Glycogen was extracted (0.9M HCl and 3 h of digestion time) and determined using a liquid chromatography-mass spectrometer (LC-MS) (Thermo Scientific, Waltham, MA, USA). Poly-β-hydroxyalkanoates (PHAs), including poly-β-hydroxybutyrate (PHB), poly-β-hydroxyvalerate (PHV), and poly-β-hydroxy-2-methylvalerate (PH2MV), were extracted (3% sulfuric acid and 3 h of digestion time) and determined by a gas chromatography-mass spectrometry (GC-MS) (Agilent, Santa Clara, CA, USA) (Lanham et al. 2013).
2.6. Statistics
All the tests were performed at least in duplicate. The one-way analysis of variance (ANOVA) and the least significant difference (LSD) post hoc tests were used to test the significance among different samples using Matlab R2014a (Mathworks, Natick, MA, USA).
3. Results and Discussion
3.1. Performance of the P(D)N-S2EBPR system
The performance of the P(D)N-S2EBPR system during the pilot testing period is summarized in Table S3. As shown in Figure 2A, the P(D)N-S2EBPR system obtained notably better and more stable P removal performance in Phase 3 (high carbon load) compared to Phase 1 (low carbon load) and 2 (medium carbon load). The median effluent PO43–-P concentration in Phase 1 was 2.28 mg/L, with only 16% of the effluent less than 1.0 mg/L (Figure 2B). Afterward, the effluent PO43–-P gradually decreased in Phase 2, with a median concentration of 1.81 mg/L and 18% of the effluent less than 1.0 mg/L. On the contrary, the median effluent PO43–-P concentration in Phase 3 largely decreased to 0.28 mg/L, with 98% and 84% of the effluent lower than 1.0 and 0.5 mg/L, respectively. Accordingly, the PO43–-P removal efficiency in Phase 3 achieved 87±9%, while the average removal efficiency in Phase 1 and 2 were less than 50% (Table S3).
As shown in Figure 3, the average removal efficiency of total inorganic nitrogen (TIN) was 54% in Phase 1, while gradually increased to ~60% in Phase 2 and ~75% in Phase 3. The improved TIN removal is likely due to the increased carbon load resulting in more denitrification capacity. Obvious nitrite accumulation was detected in Phase 2 and 3, with effluent NO2–-N concentration increasing to ~6.0 mg/L and NAR increasing to ~95% during the transition between the two phases (Figure S1). However, both effluent NO2–-N and NAR decreased gradually to ~2.0 mg/L and ~20% by the end of Phase 3. Nevertheless, the significant fluctuations in N species in the system did not appear to negatively affect EBPR, particularly in Phase 3. Collectively, these results indicate that the studied P(D)N-S2EBPR system can achieve effective P removal and NO2–-N accumulation simultaneously, although further optimization are needed for better performance stability.
3.2. Mass balance of the P(D)N-S2EBPR system
3.2.1. Mass balance of carbon and impact of carbon load and composition on S2EBPR
The mass balance of sCOD in the P(D)N-S2EBPR system during the pilot testing period was performed. The influent sCOD concentration generally increased from ~100 mg/L in Phase 1 to ~140 mg/L in Phase 3 (Figure S2A). As a result, the influent sCOD loading level was 300.6 g/d in Phase 1, 355.0 g/d in Phase 2, and 450.6 g/d in Phase 3 (Figures S2B and S3). The corresponding influent sCOD/PO43–-P mass ratio was 39.6, 41.7, and 52.2 for these three phases (Figure S2C and Table S1). A sufficient carbon supply has been considered a key to successful EBPR processes (Gu et al. 2008, Oehmen et al. 2007). For our pilot testing process, most of the carbon in raw wastewater was captured in the A-stage HRAS reactors, resulting in a fairly low influent sCOD concentration and limited carbon load to the B-stage. Additionally considering the rather short mainstream anaerobic HRT (~0.2 h), the influent carbon would not be readily consumed by PAOs for building up sufficient energy reserves (e.g., PHAs) to perform aerobic/anoxic P uptake.
The introduction of VFA-rich A-stage WAS fermentate to B-stage system allowed more efficient carbon capture from raw wastewater and re-direction to the S2EBPR reactor, which would be beneficial for PAO activity. As shown in Figure S2D, the total VFA concentration in fermentate increased substantially from ~1000 mg COD/L in Phase 2 to ~1900 mg COD/L in Phase 3. Correspondingly, the side-stream VFA/PO43–-P mass ratio, which is based on the VFA and PO43–-P loads from fermentate and RAS to S2EBPR, increased significantly from ~14.5 to ~49.3 (Figure S2C and Table S1). According to some previous studies on influent C/P ratios in EBPR systems (Gu et al. 2008, Li 2017, Majed and Gu 2020, Schuler and Jenkins 2003), the minimum HAc to P ratio was found to be 8 to 10, while the appropriate ratio for satisfying P removal was generally in the range of 20 to 50. As shown in Figure S2E, a minimum fermentate VFA dose of ~15 mg COD/mg P was observed for achieving effluent PO43–-P concentration below 0.5 mg/L (Klaus et al. 2019). Therefore, A-stage WAS fermentate supplement allows more controllable and easier optimization of the S2EBPR in this P(D)N-S2EBPR system. In addition, the calculated side-stream VFA/PO43–-P ratio, which did not account for the contribution of B-stage RAS fermentation, fluctuated greatly between 15.0 and 82.7 in Phase 3 (Figure S2D,E). Based on mass balance and stoichiometric ratio, the internal VFA supply via RAS fermentation in S2EBPR was estimated to be ~90 g COD/d, which is less than the external VFA supply via A-stage WAS fermentation (~165 g COD/d) but potentially substantial for achieving efficient and stable EBPR.
For the typical S2EBPR process in side-stream RAS fermentation (SSR) configurations without supplemental carbon addition, a 16–48 h HRTSS was recommended to ensure sufficient VFA production via RAS fermentation (Gu et al. 2019). In the alternative SSR with supplemental carbon (SSRC) configurations, additional carbon flow (e.g., sludge fermentate) was added to the side-stream reactor to increase the C/P ratio (Wang et al. 2019), with a smaller S2EBPR volume and shorter HRTSS of 1–4 h (Gu et al. 2019, Onnis-Hayden et al. 2020). In our P(D)N-S2EBPR system, ~20–30% RAS and A-stage WAS fermentate were blended in the S2EBPR reactor, yielding a HRTSS of ~3.7 h (Table S1). Such a short HRTSS is usually considered insufficient for RAS hydrolysis/fermentation (Barnard et al. 2017). To enhance fermentation for VFA production, intermittent mixing for a duration of 1 min every 3 h in S2EBPR was adopted, resulting in the sedimentation of a part of the sludge, and the consequent decoupling of SRTSS and HRTSS. The extended anaerobic conditions with estimated SRTSS of ~7–10 h (Table S1) would facilitate the fermentative organisms for enhanced RAS fermentation and extra VFA generation. Furthermore, the complex composition of the produced VFAs with a higher propionate (HPr) to HAc ratio of 1.69 (Figure S2D) than those from conventional influent (mostly HAc) would lead to kinetically more favorable conditions for PAOs than their competitors (e.g., GAOs and OHOs) (Oehmen et al. 2006) and higher PAO diversity, thus enabling a shift in carbon flux and distribution for efficient and stable EBPR (Albertsen et al. 2015, Majed and Gu 2020, Marang et al. 2018).
3.2.2. Mass balance of phosphorus and nitrogen
Mass balances of P and N were calculated based on the concentrations of PO43–-P and N species, and the flow rate in each reactor. According to the results of P mass balance, the total input PO43–-P load from influent (~8 g/d) and fermentate (~3 g/d) were similar in different phases, while the effluent PO43–-P load was much lower in Phase 3 (1.1 g/d) than in the other phases (~7 g/d) (Figure 4). During Phase 1 and 2, the sCOD load in fermentate was low (~200 g/d) and the conditions in S2EBPR were likely unsuitable for RAS fermentation, leading to lower P release in S2EBPR (~45 g/d) (Figure 4A,B). While in Phase 3, a greater P release amount of 80.6 g/d was observed in S2EBPR (Figure 4C), which is likely associated with more substrate uptake from A-stage WAS and B-stage RAS fermentation (see Section 3.2.1). As a result of the combined P release in S2EBPR and the mainstream anaerobic reactor, a much higher total P release was observed in Phase 3 (105.7 g/d) compared to the release in Phase 1 (39.2 g/d). The results showed that PAOs received sufficient carbon sources primarily in the side-stream (~76%) to synthesize intracellular storage polymers and subsequently perform effective P uptake. The influent carbon could therefore be mainly utilized by DNB for denitrification, which is consistent with the improved TIN removal efficiency in Phase 3 (Table S3). The S2EBPR in turn received lower NOx–-N loads from diverted RAS with a higher proportion of NO2–-N over NO3–-N compared to the other phases (Figure S4), which would further lead to reduced carbon requirements for NOx–-N reduction in the side-stream and benefit PAOs. Consequently, a high amount of P uptake was obtained in the CSTRs (117.4 g/d in total) with ~98% occurring in the first two CSTRs. Eventually, the elevated P release and uptake amount in Phase 3 yielded a higher net P removal of 12.3 g/d and removal efficiency of 91%.
As shown in Figure S4, noticeable amounts of NO2–-N were produced in the intermittently aerated CSTRs during Phase 2 and 3 (14.3 and 23.8 g/d, respectively), which is conducive to the activity of AnAOB in the downstream anammox MBBR for final polishing of the effluent. Meanwhile, simultaneous and effective P and TIN removal occurred in CSTRs (Figure 2A and Figure 3). Since most of the readily degradable carbon inputs would have been consumed in the preceding anaerobic reactor, the received sCOD in CSTRs, with an average concentration of 47.6 mg/L in Phase 3, was very limited for heterotrophic denitrification by DNB. Under such external carbon-deficient and alternating aerobic-anoxic conditions, DPAOs would likely be activated and make an important contribution to the effective P uptake and denitrification, owing to their abilities to utilize intracellular polymers (e.g., PHA, glycogen) as internal carbon source and use either NOx–-N or O2 as terminal electron acceptors for P uptake (Sun et al. 2020, Wang et al. 2016). The DPAO enrichment has been confirmed by our microbial activity and population results (Section 3.3.3 and Section 3.4), which is probably attributed to the distinct S2EBPR configuration. The intermittent mixing strategy in S2EBPR allowed sludge sedimentation with an extended anaerobic SRTSS for PAO enrichment. While after the mixer was turned on, the bottom sludge would be more thoroughly mixed with NOx–-N from RAS to form an anoxic environment. The long-term exposure to alternating anaerobic-anoxic conditions in S2EBPR would offer the possibility for DPAO selection (Sun et al. 2020), thus enabling efficient nutrient removal performance in the mainstream with minimized influent carbon demands. Notably, a small amount of NH4+-N production (~6 g/d) was observed in S2EBPR, which should be derived from organic matter degradation and/or cell lysis (Wang et al. 2019). However, the additional NH4+-N production in S2EBPR constitutes only ~5% of the total TIN input and thus would have a limited impact on the overall N removal performance, specifically considering the substantial benefits derived from extra VFA production (~90 g COD/d) and PAO/DPAO enrichment (see Section 3.2.1).
3.3. Metabolic activities of functionally relevant microorganisms
3.3.1. Temporal dynamics of EBPR activities
To evaluate the EBPR metabolic activities of the activated sludge in the P(D)N-S2EBPR system, a set of P release and uptake batch tests were routinely conducted during the pilot testing period. Corresponding to the improved P removal performance (Figure 2A), the activated sludge showed a higher specific P release rate (Prel) and specific P uptake rate (Pup) in Phase 3 (18.8±7.4 and 8.3±6.0 mg P/(g VSS·h), respectively) than those in the other phases (~7.5 and ~3.5 mg P/(g VSS·h), respectively) (Table 1 and Figure S5), which is also correlated with the elevated total P content of the sludge (TP/MLVSS, increased from 2.5% to 3.6%). It is worth noting that the P release to HAc uptake (P/HAc) ratio was 0.37–0.83 P-mol/C-mol during the pilot testing, with an average value of ~0.6 P-mol/C-mol which is close to the theoretical ratios for different PAO biochemical pathways (0.50 to 0.75 P-mol/C-mol) (Schuler and Jenkins 2003, Smolders et al. 1994). Such high P/HAc ratios were typically observed in PAO-dominated systems with higher carbon utilization efficiency for P removal and relatively lower GAO abundance (Gu et al. 2008), which is consistent with our microbial population results (Section 3.4). However, in conventional EBPR systems, low GAO abundance may be associated with insufficient carbon available for growth, which would result in lower performance stability due to potential process disturbances (Ahn et al. 2006). Nevertheless, our P(D)N-S2EBPR system achieved efficient carbon utilization by PAOs and stable EBPR performance despite low influent carbon (69 to 208 mg COD/L), probably due to the suitable VFA composition and carbon redirection/redistribution to PAOs over GAOs via A-stage WAS and B-stage RAS fermentation.
3.3.2. EBPR kinetic rates and stoichiometric ratios
To further evaluate the EBPR activities in Phase 3 with effective biological nutrient removal (BNR) performance, a set of ex-situ P release and uptake batch tests using different carbon sources and electron acceptors was performed. As shown in Figure S6, typical EBPR profiles were observed in all the tests. Considering the conventional batch test fed with HAc can only reflect the activities of Accumulibacter-like PAOs that are active in the presence of HAc, the carbon source in the batch test was switched to the real influent of S2EBPR (i.e., A-stage WAS fermentate) to indicate EBPR metabolisms in the practice. As summarized in Table 2, although the specific P release rate (Prel) was lower in the fermentate feeding test than in the HAc feeding test, the amounts of P release and uptake were relatively higher. The extent of P release induced by different carbon sources has been used as an indicator reflecting PAO community composition (Qiu et al. 2019). The additional amount of P release in the fermentate feeding batch test is likely associated with the other PAOs with differentiated substrate uptake capabilities and preferences. For example, Tetrasphaera PAOs, which are capable of taking up complex carbon sources (e.g., sugars, amino acids, etc.), have been considered as important as Accumulibacter PAOs for P removal in full-scale EBPR facilities (Marques et al. 2017, Nielsen et al. 2019, Singleton et al. 2022). Moreover, Dechloromonas has also been recognized as another important PAOs, exhibiting similar physiology as Accumulibacter yet more diverse substrate utilization profiles as Tetrasphaera (Petriglieri et al. 2021). The involvement of Dechloromonas and Tetrasphaera PAOs throughout the pilot testing period has been confirmed by our results of microbial population analysis (Section 3.4).
The anaerobic P release to VFA uptake ratio (P/VFA) in fermentate and HAc feeding batch tests is 0.16 and 0.25 P-mol/C-mol, respectively (Table 2), which are considerably lower than the values predicted in the PAO models (0.37–0.50 P-mol/C-mol) (Hesselmann et al. 2000, Smolders et al. 1995) and some EBPR systems (Lanham et al. 2013, Onnis-Hayden et al. 2020). Relatively low P/VFA ratios have also been reported previously in some EBPR systems with good P removal performance, such as in an Accumulibacter clade II culture exhibiting a mixed PAO-GAO metabolism (0.22 P-mol/C-mol) (Welles et al. 2015), as well as in an S2EBPR facility run in an unmixed in-line MLSS fermentation (UMIF) configuration (0.16 P-mol/C-mol) (Onnis-Hayden et al. 2020). The relatively lower P/VFA ratio, when fermentate was fed instead of HAc, might also be related to the involvement of specific heterotrophic organisms including some specific GAOs that can anaerobically assimilate various organic compounds in the fermentate without reliance on polyP hydrolysis for energy generation.
Under the anaerobic phase, the PHA production to acetate uptake ratios (PHA/VFA) observed in the batch tests (~0.2 C-mol/C-mol) were much lower than the values in the PAO/GAO models (0.90–1.86 C-mol/C-mol), indicating the occurrence of some heterotrophic organisms including specific PAOs or GAOs that do not produce PHAs as their carbon and energy storage polymer (McIlroy et al. 2018, Wang et al. 2019). Consistent with the previous observation in Accumulibacter-enriched culture (Smolders et al. 1994), PHB was the most abundant fraction of PHA when HAc was fed as the sole carbon source (Figure S6A). Contrarily, a pronouncedly high PHV production to PHB production ratio (PHV/PHB) of 3.16 was observed when fermentate was fed (Table 2), which is probably related to the complex composition of fermentate with a higher HPr/HAc ratio (Figure S2D). In addition, a high PHV fraction in generated PHAs was also observed in PAO-enriched systems fed with amino acids (Marques et al. 2017, Zengin et al. 2011). Under the subsequent aerobic phase, the P uptake to PHA utilization ratios (P/PHA) in the batch tests (0.75–0.82 P-mol/C-mol) was higher than the predicted values for the PAO model (0.41 P-mol/C-mol) yet comparable to those observed in the full-scale EBPR facilities (Lanham et al. 2013, Qiu et al. 2019). Overall, the deviations in EBPR kinetics and stoichiometric values from those typically observed in conventional EBPRs suggested that there are more diverse microbial populations and that some specific PAOs/GAOs are involved with other storage polymers rather than PHAs, which require further investigation.
3.3.3. DPAO activity
As discussed in Section 5.2.2, the alternating anaerobic-anoxic conditions in S2EBPR would facilitate DPAO enrichment and intracellular carbon storage, while the alternating aerobic-anoxic conditions in mainstream CSTRs would activate internal carbon-driven denitrification and P uptake by DPAOs. Consistently, the results in the NO3–-N fed anaerobic-anoxic batch test (Figure S6E,F) suggested the presence of DPAO activity. The anoxic P uptake was 29% of the aerobic P uptake (Table 2), indicating a relatively lower DPAO fraction in terms of NO3–-N consumption (Hu et al. 2003, Lanham et al. 2018). The denitrification capability of DPAOs to use NO3–-N as an external electron acceptor for anoxic P uptake is still a matter of debate, while there seems to be a consensus that most DPAOs (e.g., Accumulibacter and Dechloromonas species) can perform denitrifying P removal via NO2–-N pathway (Petriglieri et al. 2021, Saad et al. 2016, Zeng et al. 2017). Therefore, the nitrite accumulation and alternating aerobic-anoxic conditions in the CSTRs of P(D)N-S2EBPR system would potentially promote DPAO activity, allowing more carbon and energy-efficient biological nutrient removal through simultaneous nitrification-denitrification via NO2–-N and P removal (Regmi et al. 2022). In addition, simultaneous NO2–-N reduction (denitritation) and P uptake occurred mostly at the end of the anoxic phase (Figure S6E,F), while rapid NO3–-N reduction and NO2–-N production were observed in the first 100 min (Figure S6F), indicating the potential involvement of other internal carbon-driven denitrifiers without contributing to P removal (e.g., DGAOs). Furthermore, the anoxic P/PHA ratio in the batch test is 0.37 P-mol/C-mol, which is relatively lower than reported in lab-scale DPAO culture (0.46 P-mol/C-mol) (Kuba et al. 1996), suggesting a mixed DPAO-DGAO metabolism with PHA as internal carbon sources for endogenous NOx–-N reduction. Further studies in the future are warranted for determining the contribution of DPAOs to combined N and P removal within the P(D)N-S2EBPR system, and for understanding the cooperation and competition relationships among different functionally relevant microorganisms.
3.4. Microbial community analysis revealed higher PAO diversity and unknown candidate PAOs
To compare the changes in microbial communities in the P(D)N-S2EBPR system, 16S rRNA gene amplicon sequencing was conducted throughout the pilot testing. For a better analysis of the microbial community compositions, Microbial Database for Activated Sludge (MiDAS) database (McIlroy et al. 2015) was utilized by linking the identified microorganisms to their functional information in the activated sludge systems. As shown in Figure S7, known PAOs, including Accumulibacter, Tetrasphaera, and Dechloromonas, were detected in the P(D)N-S2EBPR system. The relative abundance of total known PAOs gradually increased from 2.42±0.10% in Phase 1 to 4.08±2.83% in Phase 3a, and further increased to 5.97±2.92% in Phase 3b (Figure 5A). Accumulibacter is the dominant PAOs in Phase 3, accounting for 61% of the total known PAOs, which is probably due to the increased VFA level in fermentate (see Section 3.2.1) that provided suitable carbon sources for Accumulibacter. Meanwhile, the relative abundance of Tetrasphaera also significantly increased from ~0.12% in Phase 1 to ~1.01% in Phase 3, which could be related to the conditions in S2EBPR that benefited the fermentative organisms including Tetrasphaera (Wang et al. 2019). Besides the three canonical PAOs (Zhao et al. 2022), members belonging to other genera with fairly high fractions in the microbial community have also been proposed previously to behave as PAOs in EBPR systems (Ge et al. 2015, Stokholm-Bjerregaard et al. 2017), such as Comamonadaceae (10.28±5.89%) (Figure S7). Their metabolisms and ecological niches in the complex microbial communities, as well as their potential contributions for BNR in the P(D)N-S2EBPR system, still require further investigations, with some advanced tools, e.g., SCRS-based analyses linking phenotypic and phylogenetic traits at a finer resolution (Fernando et al. 2019, Li et al. 2018, Wang et al. 2021).
Known GAOs, including Competibacter and Propionivibrio, were detected in the system, and the vast majority (~97%) are Competibacter. A decrease in the relative abundance of total known GAOs was observed (from 3.13±1.20% in Phase 1 to 1.91±0.89% in Phase 2) (Figure 5B), which has also been observed in previous S2EBPR studies (Onnis-Hayden et al. 2020, Wang et al. 2019). It suggests that the conditions in the P(D)N-S2EBPR system (e.g., the combination of side-stream RAS fermentation) could provide competitive advantages to PAOs over GAOs, as discussed in Section 3.2.1. Subsequently, total GAO abundance increased to 2.83±1.19% in Phase 3 due to elevated carbon load, while PAO proliferation was observed with a higher abundance up to 9.95%, leading to a higher relative PAO-to-GAO abundance ratio of 1.7 compared to Phase 1 and 2 (0.8 and 1.0, respectively). Similar to previous EBPR studies (Gu et al. 2008), a linear correlation between PAO/GAO abundance ratios and P/HAc ratios was observed in our system (r = 0.789, p < 0.01) (Figure S8A). This suggests that, despite increased GAO abundance, good and stable P removal performance/activity can be maintained, as conditions in the P(D)N-S2EBPR system are more favorable for PAOs to preferentially absorb carbon for efficient EBPR activity, leaving only the spare carbon for GAOs (Gu et al. 2008). In addition, the increased external/internal carbon inputs via A-stage WAS and B-stage RAS fermentation provided sufficient organic load for PAOs/GAOs and thus mitigated competition, as observed in some other EBPR systems treating high-strength wastewater (Feng et al. 2022, Yuan et al. 2021).
With the assistance of the MiDAS database (Dueholm et al. 2022), a total of 14 genera were classified as denitrifiers with NOx–-N as electron acceptors (Figure 5B). Among the known denitrifiers, Denitratisoma, Nitrospira, Dechloromonas, Sulfuritalea, Azospira, and Haliangium were predominated, accounting for > 80% of the total denitrifying populations. The nitrite reductase (nirK) genes identified in a Nitrospira NOB genome indicated the NO2–-N reduction capacity of this organism, yet has not been experimentally demonstrated (Lücker et al. 2010). As mentioned above, the relative abundance of Nitrospira largely decreased during the testing, leading to a declined fraction of Nitrospira in the total denitrifying populations (from ~41% in Phase 1 to ~8% in Phase 3). Oppositely, the fraction of Dechloromonas, a canonical PAO harboring full or PDN pathways with intracellular reserves as internal carbon sources (Petriglieri et al. 2021), increased from ~12% of total denitrifiers in Phase 1 to ~23% in Phase 3. This suggests that the conditions in S2EBPR would have promoted DPAO selection and made great contributions to denitrification, as discussed in Section 3.2.2 and Section 3.3.3. The relative abundance of total known denitrifiers decreased significantly from ~10.16% in Phase 1 to ~3.90% at the end of Phase 2, and then maintained at ~3.77% in Phase 3. Changes in denitrifying populations may be related to the still limited external carbon available for denitrifiers in mainstream receiving A-stage effluent (100–200 mg/L sCOD), compared to the traditional BNR processes receiving raw municipal wastewater without carbon capture. This would provide a competitive advantage to specific denitrifiers that are capable of using internal carbon sources (e.g., DPAOs, DGAOs, or other unknown microorganisms) over conventional DNB. In addition, the switched electron acceptor sources (from NO3–-N in Phase 1 to NO2–-N in Phase 2 and 3) would also have an impact on the composition and abundance of denitrifiers.
3.5. Quantification and phenotypic profiling of PAOs via SCRS analysis
We employed SCRS analysis for the quantification of total PAO and GAO phenotypes (Fernando et al. 2019, Majed et al. 2012, Wang et al. 2020). The SCRS results showed that, at the end of the aerobic phase in the P release and uptake batch test, 24.5−31.1% of cells exhibited PAO phenotypes (Figure 6A), which could be considered active PAOs in the sample, and the number was considerably higher than the relative abundance of total known PAOs (i.e., Accumulibacter, Tetrasphaera, and Dechloromonas) determined by 16S rRNA gene amplicon sequencing (0.7−9.9%), suggesting diverse and unknown PAOs. It is also recognized that some cells that contain polyP may not exert EBPR activities. Figure 6B shows the changes in average poly-P, glycogen, and PHA contents in PAO cells within a batch test for each phase. At the end of the aerobic phase, the intracellular polyP content (expressed as Raman intensity ratio of polyP to amide I band) in Phase 3 (3.68±0.79) was higher than that in Phase 1 (2.06±0.43) and Phase 2 (2.39±0.47), which is consistent with the better P removal performance observed in Phase 3 (see Section 3.1). Furthermore, the differences in polyP content at the beginning and end of the anaerobic/aerobic phase are also higher in Phase 3, suggesting a higher EBPR activity in terms of polyP utilization and formation, as observed in Section 3.3.1.
3.6. Quantification of intracellular polymers indicated carbon flux at population levels
The SCRS-determined relative abundance of total active GAOs was 2.9−5.9%, also slightly higher than the sequencing-quantified relative abundance of total known GAOs (0.6−5.1%) (Figure 5A). Consistent with the observed relationship between P/HAc and the sequencing-based PAO/GAO abundance ratio (Figure S8A), the SCRS-determined PAO/GAO ratio was also positively correlated with the P/HAc ratio (r = 0.992, p < 0.05) (Figure S8B). It again demonstrated that the P(D)N-S2EBPR system could achieve good P removal despite low influent carbon input, due to the unique process conditions that ensure efficient carbon utilization for EBPR with less carbon competition, lower ORP, and GAO suppression.
Notably, a fraction of cells with PHA peaks but no polyP and glycogen peaks were persistently observed throughout the P release and uptake batch test, especially at the end of the anaerobic phase (up to 13.0% of total cells in Phase 3). We termed this cellular phenotype as PHA-producing bacteria (PPB). As shown in Figure 6C, the PHA content in PPB at the end of the anaerobic phase (0.9−1.56) was much higher than that in either PAOs (0.10−0.19) or GAOs (0.00−0.76). It is speculated that PPB could be acclimated and enriched under the external carbon-deficient conditions in the P(D)N-S2EBPR system due to their ability to synthesize PHA bioplastics as internal sources of carbon and energy (Fra-Vázquez et al. 2019). The physiological characteristics, ecological niches, and potential functional roles (e.g., endogenous denitrification) of PPB in the P(D)N-S2EBPR system are unclear. Nevertheless, the SCRS-revealed results provided additional evidence of the diverse metabolic profiles in the complex systems that are related to the involvement of unknown microorganisms (e.g., PAOs, GAOs, PPB) or metabolism shift under certain operating conditions (Acevedo et al. 2012, Kolakovic et al. 2021). Further studies are warranted to better understand the coexistence and synergy among various functionally relevant microorganisms in the system.
4. Conclusions
This is the first study that demonstrated the successful incorporation of S2EBPR into a A/B process in an integrated pilot system combining A-stage HRAS with B-stage P(D)N/A-S2EBPR for simultaneous EBPR and shortcut N removal. The following conclusions are obtained:
- Integrated and efficient P removal and shortcut N removal were achieved simultaneously in our pilot HRAS-P(D)N/A-S2EBPR system, which consists of influent carbon capture in A-stage and re-direction to S2EBPR for biological P removal, and B-stage shortcut N removal followed by anammox, successfully demonstrating the feasibility and reliability of the system coupling mainstream P(D)N/A with side-stream EBPR to achieve carbon-efficient shortcut N removal and influent carbon-independent P removal.
- EBPR activities and stoichiometric evaluation of a novel HRAS-P(D)N/A-S2EBPR system revealed deviations for P/VFA, PHA/VFA, PHV/PHB, and P/PHA ratios from conventional BEPR systems, suggesting the presence of unique microbial populations, activities, and mechanisms in this system.
- Results of 16S rRNA gene amplicon sequencing showed an increase in total known PAOs and relative PAO-to-GAO abundance ratio with elevated VFA levels, and Accumulibacter is the dominant PAO. Changes in denitrifying populations indicated a potentially higher contribution of DPAOs, DGAOs, or other unknown microorganisms to denitrification.
- SCRS analysis identified specific microorganisms and phenotypes, such as various unidentified PAOs and PHA-producing bacteria, which would play non-negligible roles in the system performance (e.g., enhanced P removal and endogenous denitrification).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
This research was funded by the National Natural Science Foundation of China (No. 52070156) and the Scientific Research Project for Returned Overseas Scholars in Shaanxi Province of China.
References
- Acevedo, B., Oehmen, A., Carvalho, G., Seco, A., Borrás, L. and Barat, R. (2012) Metabolic shift of polyphosphate-accumulating organisms with different levels of polyphosphate storage. Water research 46(6), 1889-1900. [CrossRef]
- Ahn, C.H., Park, J.K. and Kim, K.S. (2006) Microbial Adaptability to Organic Loading Changes in an Enhanced Biological Phosphorus Removal Process. Journal of Environmental Engineering 132(8), 909-917. [CrossRef]
- Albertsen, M., Karst, S.M., Ziegler, A.S., Kirkegaard, R.H. and Nielsen, P.H. (2015) Back to Basics – The Influence of DNA Extraction and Primer Choice on Phylogenetic Analysis of Activated Sludge Communities. PLOS ONE 10(7), e0132783. [CrossRef]
- Ali, P., Zalivina, N., Le, T., Riffat, R., Ergas, S., Wett, B., Murthy, S., Al-Omari, A., deBarbadillo, C., Bott, C. and De Clippeleir, H. (2021) Primary sludge fermentate as carbon source for mainstream partial denitrification–anammox (PdNA). 93(7), 1044-1059. [CrossRef]
- APHA. (2012) Standard Methods for the Examination of Water and Wastewater 14ed, APHA American Public Health Association.
- Barnard, J.L., Dunlap, P. and Steichen, M. (2017) Rethinking the Mechanisms of Biological Phosphorus Removal. Water Environment Research 89(11), 2043-2054. [CrossRef]
- Cao, Y., Kwok, B.H., van Loosdrecht, M.C.M., Daigger, G.T., Png, H.Y., Long, W.Y., Chye, C.S. and Ghani, Y.A.B.D. (2016) The occurrence of enhanced biological phosphorus removal in a 200,000 m3/day partial nitration and Anammox activated sludge process at the Changi water reclamation plant, Singapore. Water Science and Technology 75(3), 741-751. [CrossRef]
- Cao, Y., van Loosdrecht, M.C.M. and Daigger, G.T. (2017) Mainstream partial nitritation–anammox in municipal wastewater treatment: status, bottlenecks, and further studies. Applied Microbiology and Biotechnology 101(4), 1365-1383. [CrossRef]
- Chen, G., Zhao, L. and Qi, Y. (2015) Enhancing the productivity of microalgae cultivated in wastewater toward biofuel production: A critical review. Applied Energy 137, 282-291. [CrossRef]
- Daims, H., Lebedeva, E.V., Pjevac, P., Han, P., Herbold, C., Albertsen, M., Jehmlich, N., Palatinszky, M., Vierheilig, J., Bulaev, A., Kirkegaard, R.H., von Bergen, M., Rattei, T., Bendinger, B., Nielsen, P.H. and Wagner, M. (2015) Complete nitrification by Nitrospira bacteria. Nature 528(7583), 504-509. [CrossRef]
- Dueholm, M.K.D., Nierychlo, M., Andersen, K.S., Rudkjøbing, V., Knutsson, S., Arriaga, S., Bakke, R., Boon, N., Bux, F., Christensson, M., Chua, A.S.M., Curtis, T.P., Cytryn, E., Erijman, L., Etchebehere, C., Fatta-Kassinos, D., Frigon, D., Garcia-Chaves, M.C., Gu, A.Z., Horn, H., Jenkins, D., Kreuzinger, N., Kumari, S., Lanham, A., Law, Y., Leiknes, T., Morgenroth, E., Muszyński, A., Petrovski, S., Pijuan, M., Pillai, S.B., Reis, M.A.M., Rong, Q., Rossetti, S., Seviour, R., Tooker, N., Vainio, P., van Loosdrecht, M., Vikraman, R., Wanner, J., Weissbrodt, D., Wen, X., Zhang, T., Nielsen, P.H., Albertsen, M., Nielsen, P.H. and Mi, D.A.S.G.C. (2022) MiDAS 4: A global catalogue of full-length 16S rRNA gene sequences and taxonomy for studies of bacterial communities in wastewater treatment plants. Nature Communications 13(1), 1908. [CrossRef]
- Feng, X., Qian, Y., Xi, P., Cao, R., Qin, L., Zhang, S., Chai, G., Huang, M., Li, K., Xiao, Y., Xie, L., Song, Y. and Wang, D. (2022) Partial Nitrification and Enhanced Biological Phosphorus Removal in a Sequencing Batch Reactor Treating High-Strength Wastewater. International Journal of Environmental Research and Public Health 19(9), 5653. [CrossRef]
- Fernando, E.Y., McIlroy, S.J., Nierychlo, M., Herbst, F.-A., Petriglieri, F., Schmid, M.C., Wagner, M., Nielsen, J.L. and Nielsen, P.H. (2019) Resolving the individual contribution of key microbial populations to enhanced biological phosphorus removal with Raman–FISH. The ISME Journal 13(8), 1933-1946. [CrossRef]
- Ferrentino, R., Langone, M., Villa, R. and Andreottola, G. (2018) Strict anaerobic side-stream reactor: effect of the sludge interchange ratio on sludge reduction in a biological nutrient removal process. Environmental Science and Pollution Research 25(2), 1243-1256. [CrossRef]
- Fra-Vázquez, A., Santorio, S., Palmeiro-Sánchez, T., Val del Río, Á. and Mosquera-Corral, A. (2019) PHA accumulation of a mixed microbial culture co-exists with ammonia partial nitritation. Chemical Engineering Journal 360, 1255-1261. [CrossRef]
- Ge, H., Batstone, D.J. and Keller, J. (2015) Biological phosphorus removal from abattoir wastewater at very short sludge ages mediated by novel PAO clade Comamonadaceae. Water research 69, 173-182. [CrossRef]
- Gu, A.Z., Saunders, A., Neethling, J., Stensel, H. and Blackall, L. (2008) Functionally relevant microorganisms to enhanced biological phosphorus removal performance at full-scale wastewater treatment plants in the United States. Water Environment Research 80(8), 688-698. [CrossRef]
- Gu, A.Z., Tooker, N.B., Onnis-Hayden, A., Wang, D., Srinivasan, V., Li, G., Takács, I. and Vargas, E. (2019) Optimization and design of a side-stream EBPR process as a sustainable approach for achieving stable and efficient phosphorus removal.
- Guthi, R.-S., Tondera, K., Gillot, S., Buffière, P., Boillot, M. and Chazarenc, F. (2022) A-Stage process – Challenges and drawbacks from lab to full scale studies: A review. Water research 226, 119044. [CrossRef]
- Hesselmann, R.P.X., von Rummell, R., Resnick, S.M., Hany, R. and Zehnder, A.J.B. (2000) Anaerobic metabolism of bacteria performing enhanced biological phosphate removal. Water research 34(14), 3487-3494. [CrossRef]
- Hu, J.Y., Ong, S.L., Ng, W.J., Lu, F. and Fan, X.J. (2003) A new method for characterizing denitrifying phosphorus removal bacteria by using three different types of electron acceptors. Water research 37(14), 3463-3471. [CrossRef]
- Jia, Z., Yuan, Q., Roots, P., Sabba, F., Rosenthal, A.F., Kozak, J.A. and Wells, G.F. (2023) Partial Nitritation/Anammox and biological phosphorus removal integration in a single bioreactor under mainstream conditions. Bioresource Technology 373, 128714. [CrossRef]
- Klaus, S., Printz, K., McCullough, K., Srinivasan, V., Wang, D., He, P., De Clippeleir, H., Gu, A. and Bott, C. (2019) Integrating BioP and Shortcut Nitrogen Removal via RAS Fermentation and Partial Denitrification/Anammox, Water Environment Federation. [CrossRef]
- Klaus, S.A. (2019) Intensification of Biological Nutrient Removal Processes, Virginia Tech.
- Kolakovic, S., Freitas, E.B., Reis, M.A.M., Carvalho, G. and Oehmen, A. (2021) Accumulibacter diversity at the sub-clade level impacts enhanced biological phosphorus removal performance. Water research 199, 117210. [CrossRef]
- Kuba, T., Murnleitner, E., van Loosdrecht, M.C.M. and Heijnen, J.J. (1996) A metabolic model for biological phosphorus removal by denitrifying organisms. Biotechnology and Bioengineering 52(6), 685-695. [CrossRef]
- Lackner, S., Gilbert, E.M., Vlaeminck, S.E., Joss, A., Horn, H. and van Loosdrecht, M.C.M. (2014) Full-scale partial nitritation/anammox experiences – An application survey. Water research 55, 292-303. [CrossRef]
- Lanham, A.B., Oehmen, A., Carvalho, G., Saunders, A.M., Nielsen, P.H. and Reis, M.A.M. (2018) Denitrification activity of polyphosphate accumulating organisms (PAOs) in full-scale wastewater treatment plants. Water Science and Technology 78(12), 2449-2458. [CrossRef]
- Lanham, A.B., Oehmen, A., Saunders, A.M., Carvalho, G., Nielsen, P.H. and Reis, M.A.M. (2013) Metabolic versatility in full-scale wastewater treatment plants performing enhanced biological phosphorus removal. Water research 47(19), 7032-7041. [CrossRef]
- Li, G., Tooker, N.B., Wang, D., Srinivasan, V., Barnard, J.L., Russell, A., Takacs, I., Bott, C., Dobrowski, P., Onnis-Hayden, A. and Gu, A.Z. (2020a) Modeling Side-Stream Enhanced Biological Phosphorus Removal (S2EBPR) System Using Agent-based Model with Adaptive Maintenance, Decay and TCA Metabolism. 2020.2011.2018.387589. [CrossRef]
- Li, J., Li, J., Peng, Y., Wang, S., Zhang, L., Yang, S. and Li, S. (2020b) Insight into the impacts of organics on anammox and their potential linking to system performance of sewage partial nitrification-anammox (PN/A): A critical review. Bioresource Technology 300, 122655. [CrossRef]
- Li, Y. (2017) Link Phylogenetic and Phenotypic Characteristics to Reveal Factors Governing Enhanced Biological Phosphorus Removal (EBPR) Process Performance and Stability, Northeastern University.
- Li, Y., Cope, H.A., Rahman, S.M., Li, G., Nielsen, P.H., Elfick, A. and Gu, A.Z. (2018) Toward Better Understanding of EBPR Systems via Linking Raman-Based Phenotypic Profiling with Phylogenetic Diversity. Environmental science & technology 52(15), 8596-8606. [CrossRef]
- Liu, Y., Gu, J. and Zhang, M. (2019) AB processes: Towards energy self-sufficient municipal wastewater treatment, IWA publishing.
- Lücker, S., Wagner, M., Maixner, F., Pelletier, E., Koch, H., Vacherie, B., Rattei, T., Damsté, J.S., Spieck, E., Le Paslier, D. and Daims, H. (2010) A Nitrospira metagenome illuminates the physiology and evolution of globally important nitrite-oxidizing bacteria. Proc Natl Acad Sci U S A 107(30), 13479-13484. [CrossRef]
- Ma, B., Wang, S., Cao, S., Miao, Y., Jia, F., Du, R. and Peng, Y. (2016) Biological nitrogen removal from sewage via anammox: Recent advances. Bioresource Technology 200, 981-990. [CrossRef]
- Majed, N., Chernenko, T., Diem, M. and Gu, A.Z. (2012) Identification of functionally relevant populations in enhanced biological phosphorus removal processes based on intracellular polymers profiles and insights into the metabolic diversity and heterogeneity. Environmental science & technology 46(9), 5010-5017. [CrossRef]
- Majed, N. and Gu, A.Z. (2020) Phenotypic dynamics in polyphosphate and glycogen accumulating organisms in response to varying influent C/P ratios in EBPR systems. Science of The Total Environment 743, 140603. [CrossRef]
- Marang, L., van Loosdrecht, M.C.M. and Kleerebezem, R. (2018) Enrichment of PHA-producing bacteria under continuous substrate supply. New Biotechnology 41, 55-61. [CrossRef]
- Marques, R., Santos, J., Nguyen, H., Carvalho, G., Noronha, J.P., Nielsen, P.H., Reis, M.A.M. and Oehmen, A. (2017) Metabolism and ecological niche of Tetrasphaera and Ca. Accumulibacter in enhanced biological phosphorus removal. Water research 122, 159-171. [CrossRef]
- McIlroy, S.J., Onetto, C.A., McIlroy, B., Herbst, F.-A., Dueholm, M.S., Kirkegaard, R.H., Fernando, E., Karst, S.M., Nierychlo, M., Kristensen, J.M., Eales, K.L., Grbin, P.R., Wimmer, R. and Nielsen, P.H. (2018) Genomic and in Situ Analyses Reveal the Micropruina spp. as Abundant Fermentative Glycogen Accumulating Organisms in Enhanced Biological Phosphorus Removal Systems. Frontiers in microbiology 9. [CrossRef]
- McIlroy, S.J., Saunders, A.M., Albertsen, M., Nierychlo, M., McIlroy, B., Hansen, A.A., Karst, S.M., Nielsen, J.L. and Nielsen, P.H. (2015) MiDAS: the field guide to the microbes of activated sludge. Database 2015. [CrossRef]
- Nielsen, P.H., McIlroy, S.J., Albertsen, M. and Nierychlo, M. (2019) Re-evaluating the microbiology of the enhanced biological phosphorus removal process. Current Opinion in Biotechnology 57, 111-118. [CrossRef]
- Oehmen, A., Lemos, P.C., Carvalho, G., Yuan, Z., Keller, J., Blackall, L.L. and Reis, M.A.M. (2007) Advances in enhanced biological phosphorus removal: From micro to macro scale. Water research 41(11), 2271-2300. [CrossRef]
- Oehmen, A., Saunders, A.M., Vives, M.T., Yuan, Z. and Keller, J. (2006) Competition between polyphosphate and glycogen accumulating organisms in enhanced biological phosphorus removal systems with acetate and propionate as carbon sources. Journal of Biotechnology 123(1), 22-32. [CrossRef]
- Onnis-Hayden, A., Srinivasan, V., Tooker, N.B., Li, G., Wang, D., Barnard, J.L., Bott, C., Dombrowski, P., Schauer, P., Menniti, A., Shaw, A., Stinson, B., Stevens, G., Dunlap, P., Takács, I., McQuarrie, J., Phillips, H., Lambrecht, A., Analla, H., Russell, A. and Gu, A.Z. (2020) Survey of full-scale sidestream enhanced biological phosphorus removal (S2EBPR) systems and comparison with conventional EBPRs in North America: Process stability, kinetics, and microbial populations. Water Environment Research 92(3), 403-417. [CrossRef]
- Petriglieri, F., Singleton, C., Peces, M., Petersen, J.F., Nierychlo, M. and Nielsen, P.H. (2021) “Candidatus Dechloromonas phosphoritropha” and “Ca. D. phosphorivorans”, novel polyphosphate accumulating organisms abundant in wastewater treatment systems. The ISME Journal 15(12), 3605-3614. [CrossRef]
- Printz, K.E. (2019) The Investigation of Nitrite Accumulation and Biological Phosphorus Removal in an Intermittently Aerated Process Combining Shortcut Nitrogen Removal and Sidestream Biological Phosphorus Removal, Virginia Tech.
- Qiu, G., Zuniga-Montanez, R., Law, Y., Thi, S.S., Nguyen, T.Q.N., Eganathan, K., Liu, X., Nielsen, P.H., Williams, R.B.H. and Wuertz, S. (2019) Polyphosphate-accumulating organisms in full-scale tropical wastewater treatment plants use diverse carbon sources. Water research 149, 496-510. [CrossRef]
- Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J. and Glöckner, F.O. (2012) The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research 41(D1), D590-D596. [CrossRef]
- R Core Team (2013) R: A language and environment for statistical computing.
- Regmi, P., Miller, M.W., Holgate, B., Bunce, R., Park, H., Chandran, K., Wett, B., Murthy, S. and Bott, C.B. (2014) Control of aeration, aerobic SRT and COD input for mainstream nitritation/denitritation. Water research 57, 162-171. [CrossRef]
- Regmi, P., Sturm, B., Hiripitiyage, D., Keller, N., Murthy, S. and Jimenez, J. (2022) Combining continuous flow aerobic granulation using an external selector and carbon-efficient nutrient removal with AvN control in a full-scale simultaneous nitrification-denitrification process. Water research 210, 117991. [CrossRef]
- Ren, Z.J. and Pagilla, K. (2022) Pathways to Water Sector Decarbonization, Carbon Capture and Utilization, IWA Publishing.
- Roots, P., Rosenthal, A., Wang, Y., Sabba, F., Jia, Z., Yang, F., Zhang, H., Kozak, J. and Wells, G. (2020) Pushing the limits of solids retention time for enhanced biological phosphorus removal: process characteristics and Accumulibacter population structure. Water Science and Technology 82(8), 1614-1627. [CrossRef]
- Roots, P., Wang, Y., Rosenthal, A.F., Griffin, J.S., Sabba, F., Petrovich, M., Yang, F., Kozak, J.A., Zhang, H. and Wells, G.F. (2019) Comammox Nitrospira are the dominant ammonia oxidizers in a mainstream low dissolved oxygen nitrification reactor. Water research 157, 396-405. [CrossRef]
- Saad, S.A., Welles, L., Abbas, B., Lopez-Vazquez, C.M., van Loosdrecht, M.C.M. and Brdjanovic, D. (2016) Denitrification of nitrate and nitrite by ‘Candidatus Accumulibacter phosphatis’ clade IC. Water research 105, 97-109. [CrossRef]
- Sabba, F., Farmer, M., Jia, Z., Di Capua, F., Dunlap, P., Barnard, J., Qin, C.D., Kozak, J.A., Wells, G. and Downing, L. (2023) Impact of operational strategies on a sidestream enhanced biological phosphorus removal (S2EBPR) reactor in a carbon limited wastewater plant. Science of The Total Environment 857, 159280. [CrossRef]
- Scherson, Y.D., Woo, S.-G. and Criddle, C.S. (2014) Production of Nitrous Oxide From Anaerobic Digester Centrate and Its Use as a Co-oxidant of Biogas to Enhance Energy Recovery. Environmental science & technology 48(10), 5612-5619. [CrossRef]
- Schloss, P.D., Westcott, S.L., Ryabin, T., Hall, J.R., Hartmann, M., Hollister, E.B., Lesniewski, R.A., Oakley, B.B., Parks, D.H., Robinson, C.J., Sahl, J.W., Stres, B., Thallinger, G.G., Horn, D.J.V. and Weber, C.F. (2009) Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Applied and environmental microbiology 75(23), 7537-7541. [CrossRef]
- Schuler, A.J. and Jenkins, D. (2003) Enhanced Biological Phosphorus Removal from Wastewater by Biomass with Different Phosphorus Contents, Part I: Experimental Results and Comparison with Metabolic Models. Water Environment Research 75(6), 485-498. [CrossRef]
- Singleton, C.M., Petriglieri, F., Wasmund, K., Nierychlo, M., Kondrotaite, Z., Petersen, J.F., Peces, M., Dueholm, M.S., Wagner, M. and Nielsen, P.H. (2022) The novel genus, ‘Candidatus Phosphoribacter’, previously identified as Tetrasphaera, is the dominant polyphosphate accumulating lineage in EBPR wastewater treatment plants worldwide. The ISME Journal 16(6), 1605-1616. [CrossRef]
- Smith, A.L., Stadler, L.B., Love, N.G., Skerlos, S.J. and Raskin, L. (2012) Perspectives on anaerobic membrane bioreactor treatment of domestic wastewater: A critical review. Bioresource Technology 122, 149-159. [CrossRef]
- Smolders, G.J.F., van der Meij, J., van Loosdrecht, M.C.M. and Heijnen, J.J. (1994) Model of the anaerobic metabolism of the biological phosphorus removal process: Stoichiometry and pH influence. Biotechnology and Bioengineering 43(6), 461-470. [CrossRef]
- Smolders, G.J.F., van der Meij, J., van Loosdrecht, M.C.M. and Heijnen, J.J. (1995) A structured metabolic model for anaerobic and aerobic stoichiometry and kinetics of the biological phosphorus removal process. Biotechnology and Bioengineering 47(3), 277-287. [CrossRef]
- Srinivasan, V.N., Li, G., Wang, D., Tooker, N.B., Dai, Z., Onnis-Hayden, A., Bott, C., Dombrowski, P., Schauer, P., Pinto, A. and Gu, A.Z. (2021) Oligotyping and metagenomics reveal distinct Candidatus Accumulibacter communities in side-stream versus conventional full-scale enhanced biological phosphorus removal (EBPR) systems. Water research 206, 117725. [CrossRef]
- Stokholm-Bjerregaard, M., McIlroy, S.J., Nierychlo, M., Karst, S.M., Albertsen, M. and Nielsen, P.H. (2017) A Critical Assessment of the Microorganisms Proposed to be Important to Enhanced Biological Phosphorus Removal in Full-Scale Wastewater Treatment Systems. Frontiers in microbiology 8. [CrossRef]
- Sun, Y., Peng, Y., Zhang, J., Li, X., Zhang, Q. and Zhang, L. (2020) Effect of endogenous metabolisms on survival and activities of denitrifying phosphorus removal sludge under various starvation conditions. Bioresource Technology 315, 123839. [CrossRef]
- Wan, J., Gu, J., Zhao, Q. and Liu, Y. (2016) COD capture: a feasible option towards energy self-sufficient domestic wastewater treatment. Scientific Reports 6(1), 25054. [CrossRef]
- Wang, D., He, P., Wang, Z., Li, G., Majed, N. and Gu, A.Z. (2020) Advances in single cell Raman spectroscopy technologies for biological and environmental applications. Current Opinion in Biotechnology 64, 218-229. [CrossRef]
- Wang, D., Li, Y., Cope, H.A., Li, X., He, P., Liu, C., Li, G., Rahman, S.M., Tooker, N.B., Bott, C.B., Onnis-Hayden, A., Singh, J., Elfick, A., Marques, R., Jessen, H.J., Oehmen, A. and Gu, A.Z. (2021) Intracellular polyphosphate length characterization in polyphosphate accumulating microorganisms (PAOs): Implications in PAO phenotypic diversity and enhanced biological phosphorus removal performance. Water research 206, 117726. [CrossRef]
- Wang, D., Tooker, N.B., Srinivasan, V., Li, G., Fernandez, L.A., Schauer, P., Menniti, A., Maher, C., Bott, C.B. and Dombrowski, P. (2019) Side-stream enhanced biological phosphorus removal (S2EBPR) process improves system performance-A full-scale comparative study. Water research 167, 115109. [CrossRef]
- Wang, X., Wang, S., Zhao, J., Dai, X., Li, B. and Peng, Y. (2016) A novel stoichiometries methodology to quantify functional microorganisms in simultaneous (partial) nitrification-endogenous denitrification and phosphorus removal (SNEDPR). Water research 95, 319-329. [CrossRef]
- Welles, L., Tian, W.D., Saad, S., Abbas, B., Lopez-Vazquez, C.M., Hooijmans, C.M., van Loosdrecht, M.C.M. and Brdjanovic, D. (2015) Accumulibacter clades Type I and II performing kinetically different glycogen-accumulating organisms metabolisms for anaerobic substrate uptake. Water research 83, 354-366. [CrossRef]
- Yang, Y., Zhang, L., Shao, H., Zhang, S., Gu, P. and Peng, Y. (2017) Enhanced nutrients removal from municipal wastewater through biological phosphorus removal followed by partial nitritation/anammox. Frontiers of Environmental Science & Engineering 11(2), 8. [CrossRef]
- Yuan, Z., Kang, D., Li, G., Lee, J., Han, I., Wang, D., Zheng, P., Reid, M.C. and Gu, A.Z. (2021) Combined Enhanced Biological Phosphorus Removal (EBPR) and Nitrite Accumulation for Treating High-strength Wastewater. bioRxiv, 2021.2001.2016.426983. [CrossRef]
- Zeng, W., Bai, X., Guo, Y., Li, N. and Peng, Y. (2017) Interaction of “Candidatus Accumulibacter” and nitrifying bacteria to achieve energy-efficient denitrifying phosphorus removal via nitrite pathway from sewage. Enzyme and Microbial Technology 105, 1-8. [CrossRef]
- Zengin, G.E., Artan, N., Orhon, D., Satoh, H. and Mino, T. (2011) Effect of aspartate and glutamate on the fate of enhanced biological phosphorus removal process and microbial community structure. Bioresource Technology 102(2), 894-903. [CrossRef]
- Zhang, M., Wang, S., Ji, B. and Liu, Y. (2019) Towards mainstream deammonification of municipal wastewater: Partial nitrification-anammox versus partial denitrification-anammox. Science of The Total Environment 692, 393-401. [CrossRef]
- Zhao, W., Bi, X., Peng, Y. and Bai, M. (2022) Research advances of the phosphorus-accumulating organisms of Candidatus Accumulibacter, Dechloromonas and Tetrasphaera: Metabolic mechanisms, applications and influencing factors. Chemosphere 307, 135675. [CrossRef]
Figure 1.
Schematic flow diagrams of the pilot plant run in a HRAS-P(D)N/A-S2EBPR configuration and the P(D)N-S2EBPR system (marked by the red dashed box) at the Chesapeake-Elizabeth Sewage Treatment Plant (Virginia Beach, VA, USA). HRAS: high-rate activated sludge; CSTRs: continuous stirred-tank reactors; S2EBPR: side-stream biological P remover; RAS: return activated sludge; WAS: waste activated sludge; MBBR: moving bed biofilm reactor.
Figure 1.
Schematic flow diagrams of the pilot plant run in a HRAS-P(D)N/A-S2EBPR configuration and the P(D)N-S2EBPR system (marked by the red dashed box) at the Chesapeake-Elizabeth Sewage Treatment Plant (Virginia Beach, VA, USA). HRAS: high-rate activated sludge; CSTRs: continuous stirred-tank reactors; S2EBPR: side-stream biological P remover; RAS: return activated sludge; WAS: waste activated sludge; MBBR: moving bed biofilm reactor.
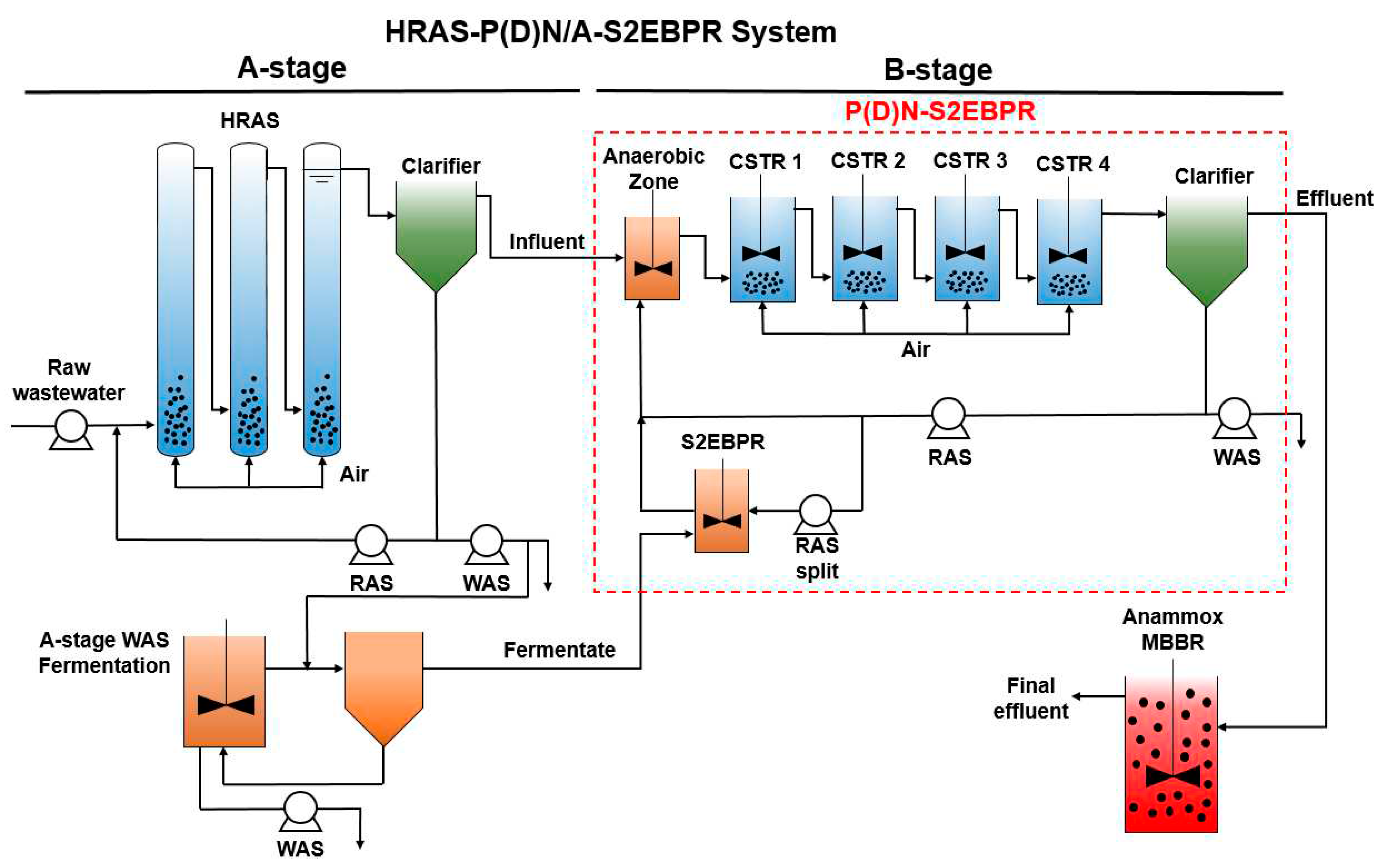
Figure 2.
Biological P removal performance during the pilot testing: (A) influent PO43–-P, effluent PO43–-P, and PO43–-P removal efficiency; and (B) cumulative relative frequency distribution for effluent PO43–-P during different phases. The effluent PO43–-P data shown in the summary table are in mg/L.
Figure 2.
Biological P removal performance during the pilot testing: (A) influent PO43–-P, effluent PO43–-P, and PO43–-P removal efficiency; and (B) cumulative relative frequency distribution for effluent PO43–-P during different phases. The effluent PO43–-P data shown in the summary table are in mg/L.
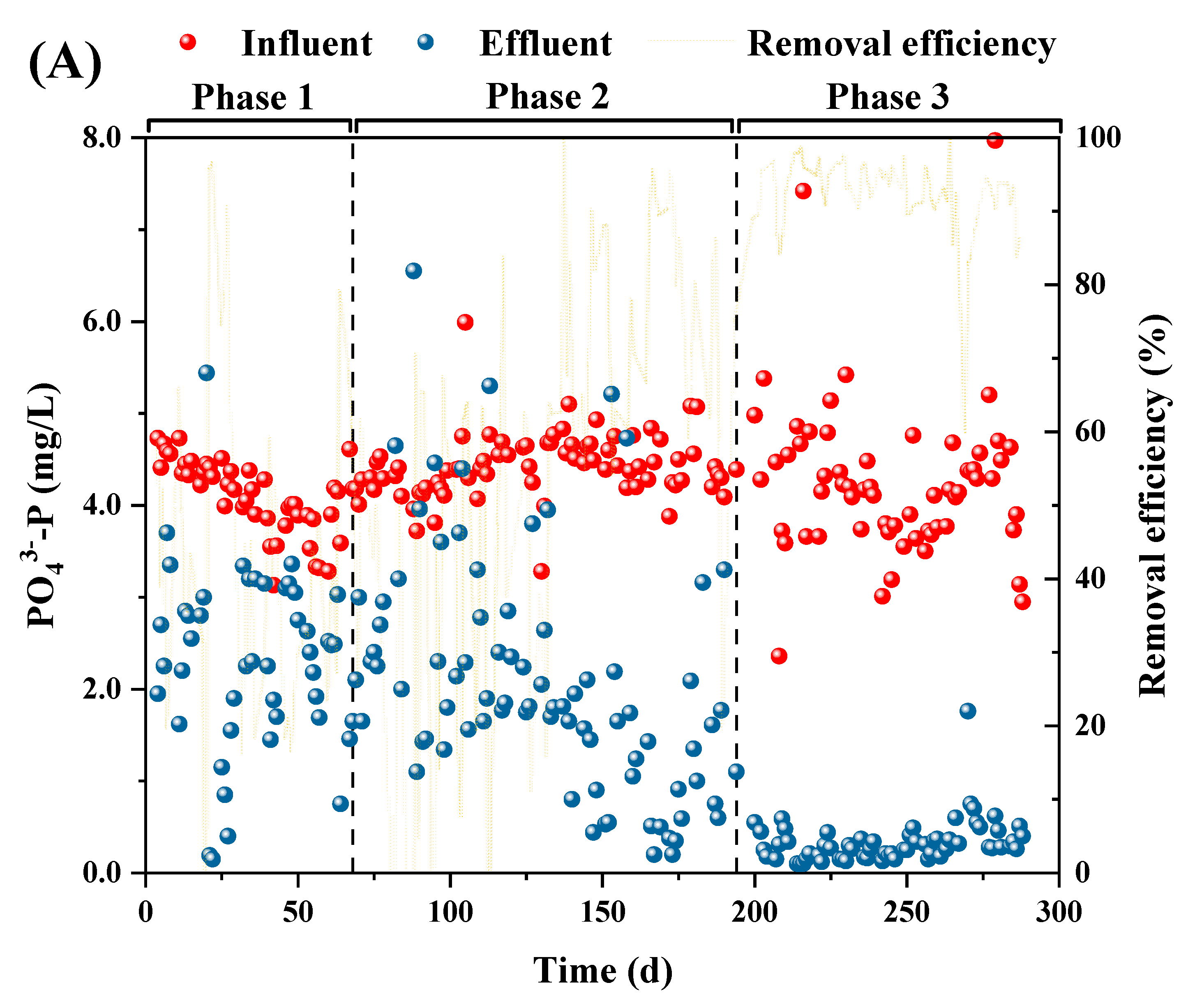
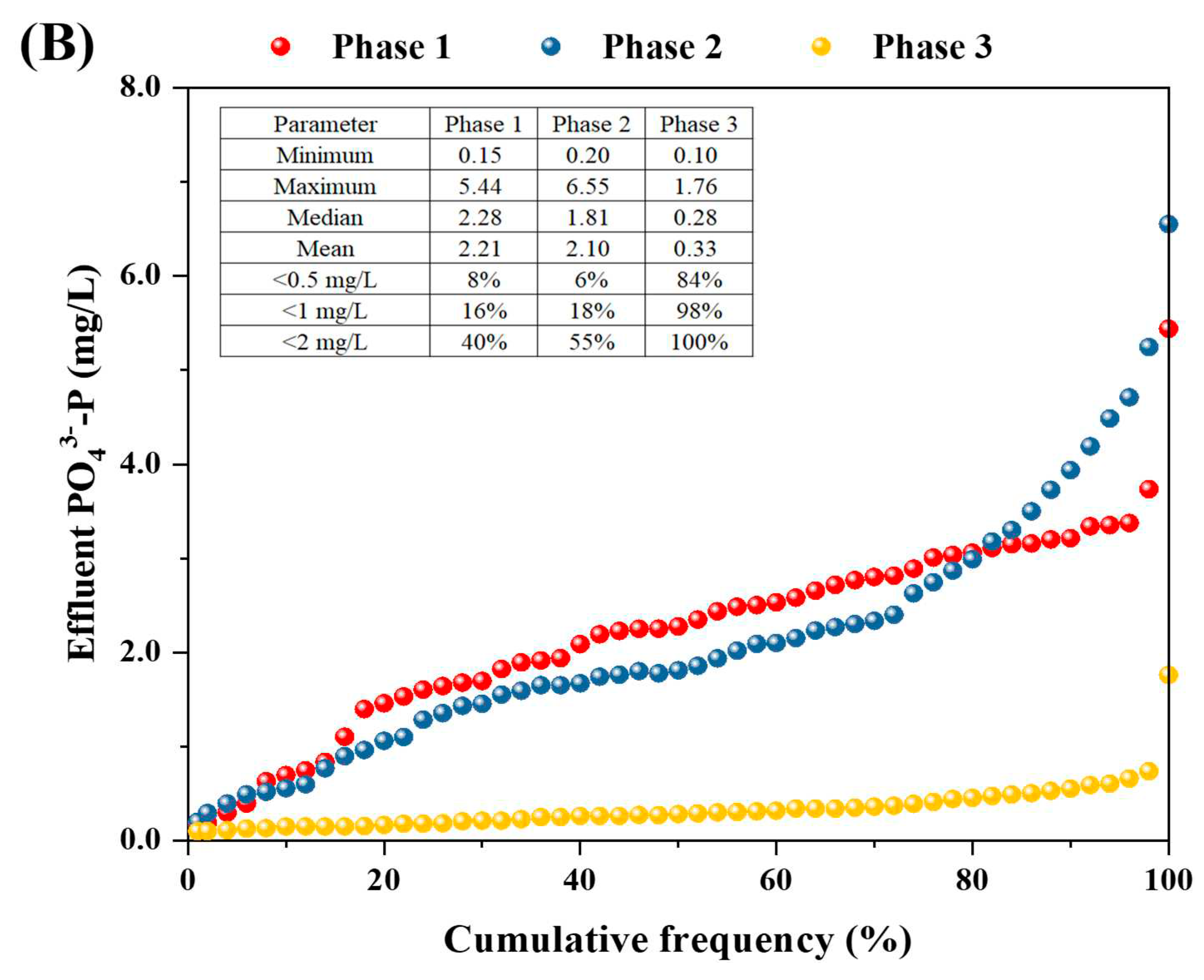
Figure 3.
Biological N removal performance during the pilot testing: influent TIN, effluent TIN, and TIN removal efficiency. TIN: total inorganic nitrogen (sum of NO3–-N, NO2–-N and NH4+-N).
Figure 3.
Biological N removal performance during the pilot testing: influent TIN, effluent TIN, and TIN removal efficiency. TIN: total inorganic nitrogen (sum of NO3–-N, NO2–-N and NH4+-N).
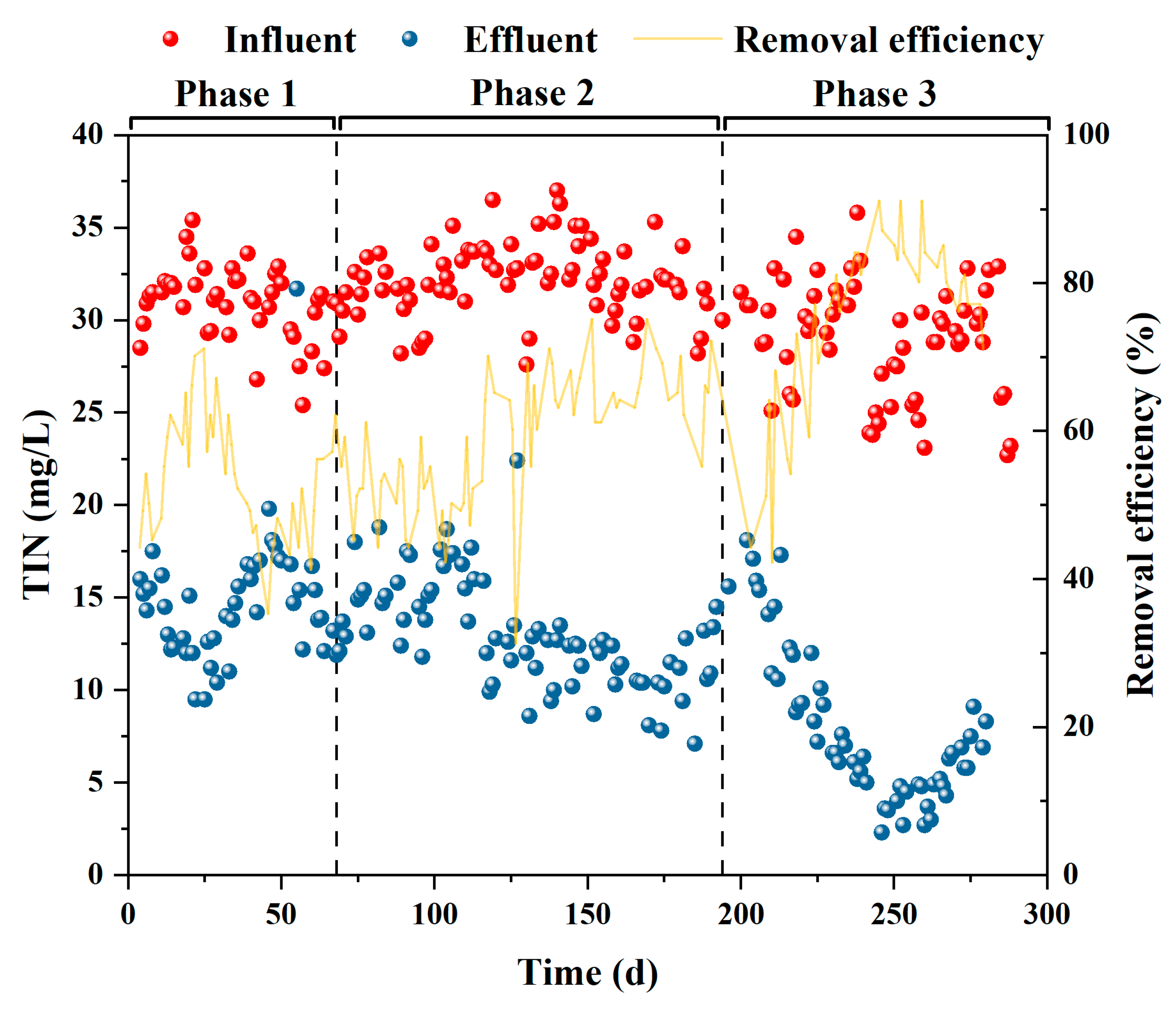
Figure 4.
Fate of PO43–-P in (A) Phase 1, (B) Phase 2, and (C) Phase 3. Value with a plus or minus sign represents the PO43–-P release and uptake, respectively. Value in brackets represents the percentage of PO43–-P in each unit compared to the total input. AN: anaerobic reactor; AE: intermittently aerated continuous stirred-tank reactors; S2EBPR: side-stream biological P remover; RAS: return activated sludge; WAS: calculated P removal via waste activated sludge. Units are in g/d.
Figure 4.
Fate of PO43–-P in (A) Phase 1, (B) Phase 2, and (C) Phase 3. Value with a plus or minus sign represents the PO43–-P release and uptake, respectively. Value in brackets represents the percentage of PO43–-P in each unit compared to the total input. AN: anaerobic reactor; AE: intermittently aerated continuous stirred-tank reactors; S2EBPR: side-stream biological P remover; RAS: return activated sludge; WAS: calculated P removal via waste activated sludge. Units are in g/d.
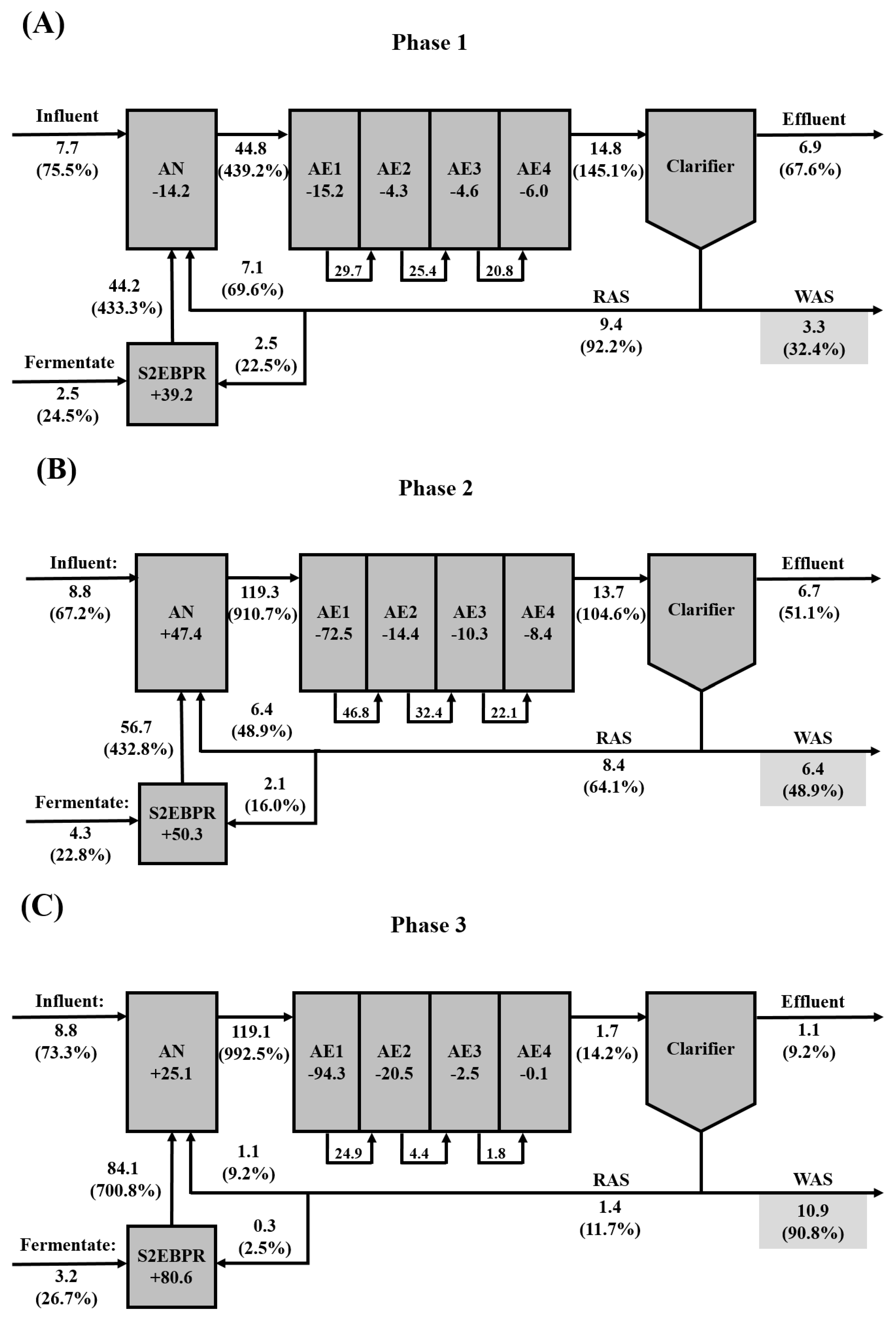
Figure 5.
The 16S rRNA gene amplicon sequencing-determined relative abundance of key functionally relevant microorganisms during the pilot testing: (A) known PAOs (i.e., Accumulibacter, Tetrasphaera, and Dechloromonas) and total known GAOs; (B) Nitrosomonas AOB, Nitrospira NOB, and known denitrifiers.
Figure 5.
The 16S rRNA gene amplicon sequencing-determined relative abundance of key functionally relevant microorganisms during the pilot testing: (A) known PAOs (i.e., Accumulibacter, Tetrasphaera, and Dechloromonas) and total known GAOs; (B) Nitrosomonas AOB, Nitrospira NOB, and known denitrifiers.
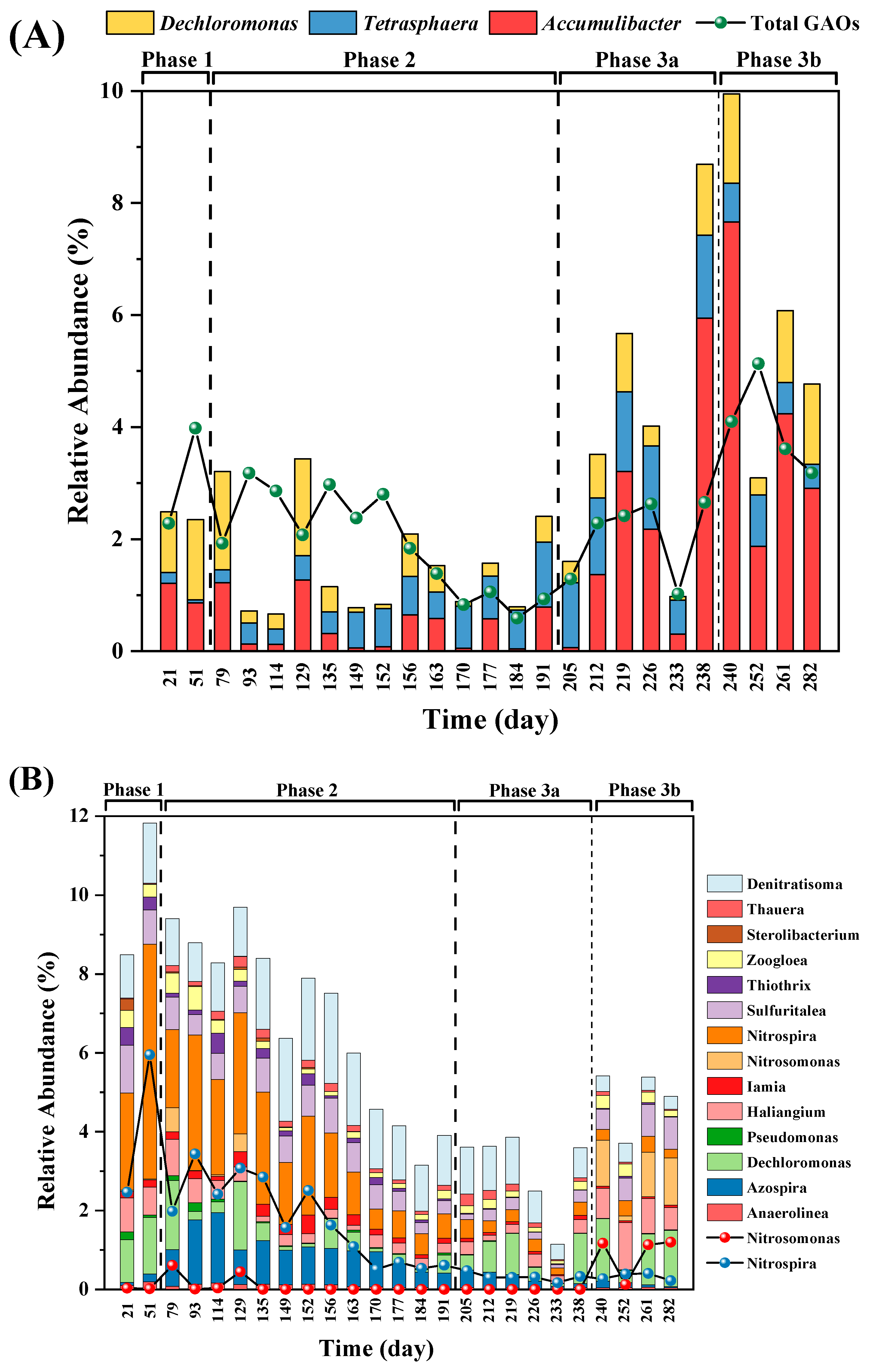
Figure 6.
The single-cell Raman spectroscopy-determined EBPR phenotypes within a P release and uptake batch test for each phase (error bars represent the standard error): (A) relative abundance of PAOs, GAOs and PPOs; (B) polyP, PHA and glycogen contents in PAOs; and (D) PHA content in PAOs, GAOs and PPOs. PPOs: PHA-producing organisms.
Figure 6.
The single-cell Raman spectroscopy-determined EBPR phenotypes within a P release and uptake batch test for each phase (error bars represent the standard error): (A) relative abundance of PAOs, GAOs and PPOs; (B) polyP, PHA and glycogen contents in PAOs; and (D) PHA content in PAOs, GAOs and PPOs. PPOs: PHA-producing organisms.
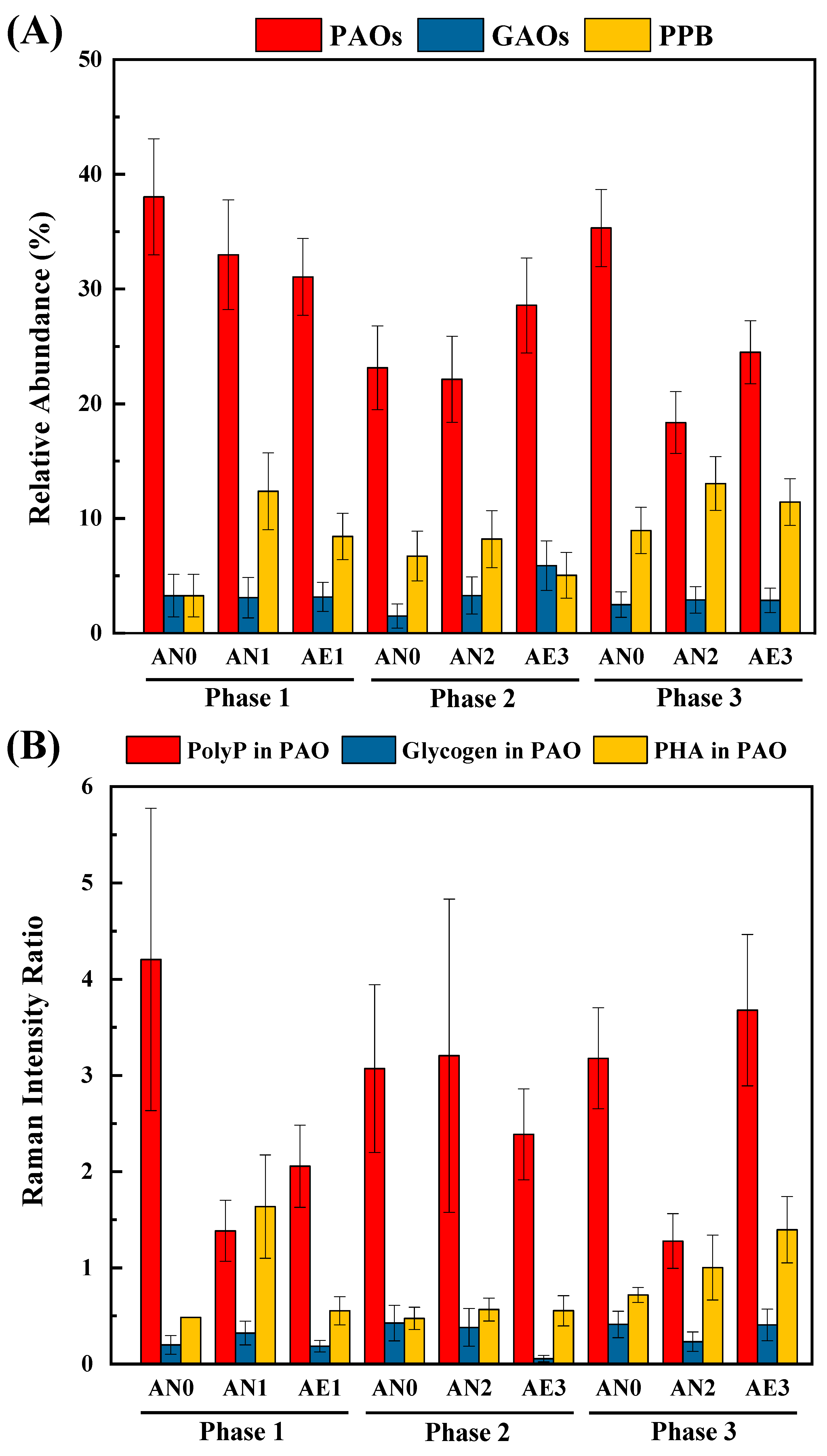
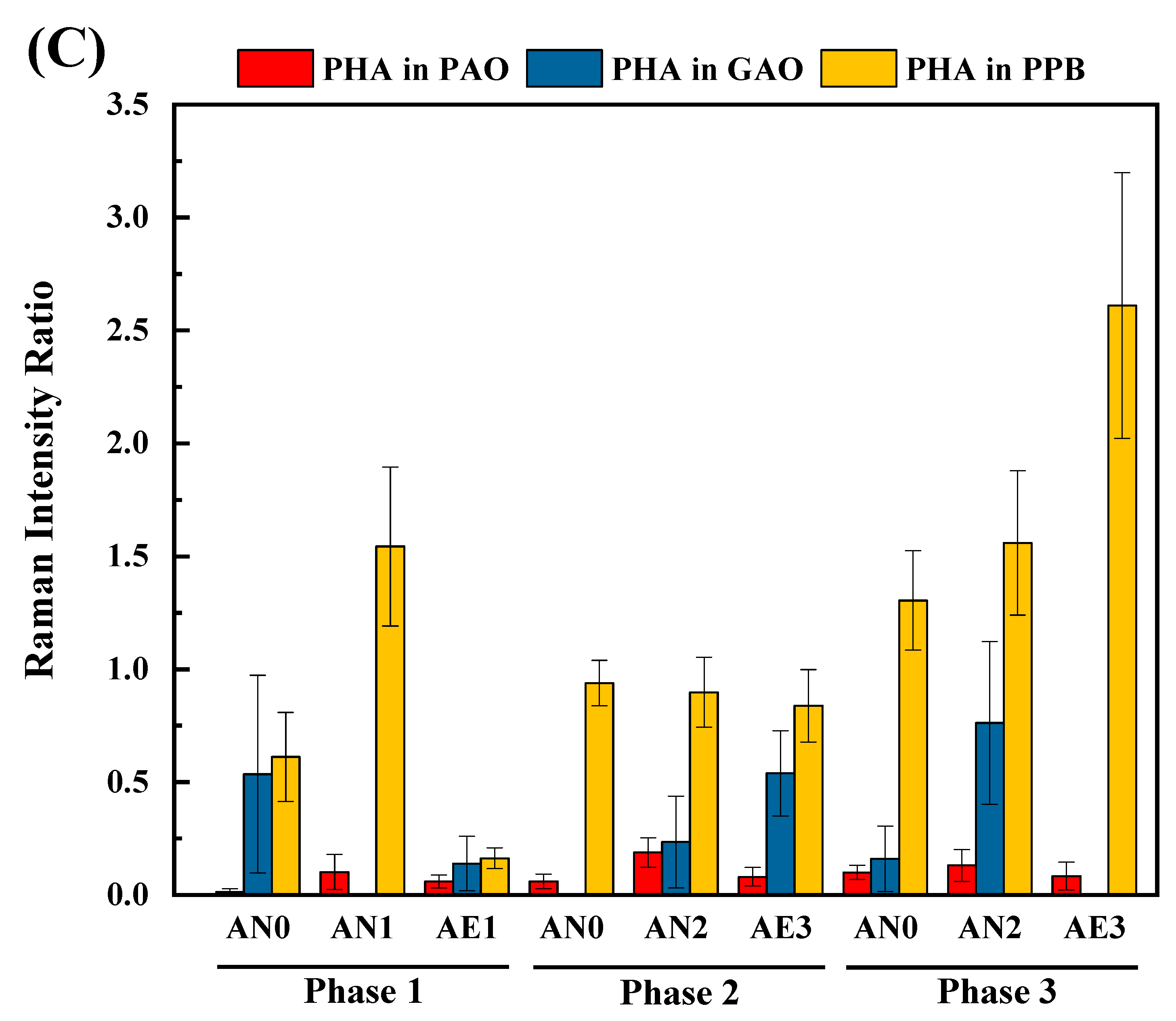

Table 1.
Specific kinetic rates and stoichiometric ratios observed in the ex situ P release and uptake batch tests during the pilot testing period, and comparison with other EBPR systems reported in the literature.
Table 1.
Specific kinetic rates and stoichiometric ratios observed in the ex situ P release and uptake batch tests during the pilot testing period, and comparison with other EBPR systems reported in the literature.
| Parameter | Phase 1 | Phase 2 | Phase 3 | S2EBPR systemsa | EBPR systemsb |
| Prel [mg P/(g VSS·h)] | 7.7±2.6 | 7.2±2.4 | 18.8±7.4 | 2.9–7.0 | 3.2–31.9 |
| Pup [mg P/(g VSS·h)] | 3.9±1.1 | 3.2±1.3 | 8.3±6.0 | 0.6–2.6 | 0.7–19.2 |
| Pup/Prel | 0.5±0.1 | 0.4±0.1 | 0.4±1.1 | 0.2–0.5 | 0.2–0.7 |
| HAcup [mg HAc/(g VSS·h)] | 14.4±4.1 | 10.3±3.5 | 29.1±11.1 | 7.7–24.9 | 9.0–47.0 |
| P/HAc (P-mol/C-mol) | 0.5±0.1 | 0.7±0.1 | 0.6±0.1 | 0.2–1.3 | 0.1–1.1 |
Prel: specific P release rate; HAcup: specific HAc uptake rate; Pup: specific P uptake rate; P/HAc: P release to HAc uptake ratio. a: Refer to Gu et al. (2008), Lanham et al. (2013), and Onnis-Hayden et al. (2020); b: Refer to Kuba et al. (1996), Gu et al. (2008), He et al. (2008), and López-Vázquez et al. (2008).
Table 2.
Specific kinetic rates and stoichiometric ratios observed in the ex situ P release and uptake batch tests using different carbon sources and electron acceptors with the activated sludge collected at the end of Phase 3, and comparison with other full-scale EBPR systems reported in the literature.
Table 2.
Specific kinetic rates and stoichiometric ratios observed in the ex situ P release and uptake batch tests using different carbon sources and electron acceptors with the activated sludge collected at the end of Phase 3, and comparison with other full-scale EBPR systems reported in the literature.
| Carbon source | This study | Previous EBPR studies | ||
| Fermentate | HAc | Real wastewater | HAc | |
| Anaerobic results | ||||
| P release (mg P/g VSS) | 10.9 | 8.6 | 3.4–16.7a | 5.1–24.3b |
| Prel [mg P/(g VSS·h)] | 6.6 | 7.8 | – | 3.2–31.9c |
| VFAup [mg VFA/(g VSS·h)] | 38.2 | 37.9 | – | 9.0–47.0d,e |
| P/VFA (P-mol/C-mol) | 0.16 | 0.25 | 0.63–1.00a | 0.11–1.30c,f |
| PHA/VFA (C-mol/C-mol) | 0.21 | 0.23 | 1.20–1.39a | 0.63–2.10a,c |
| PHV/PHB (C-mol/C-mol) | 3.16 | 0.11 | 0.63–0.86a | 0.00–0.26g,h |
| Anoxic results | ||||
| P uptake (mg P/g VSS) | 2.5 | – | – | – |
| Pup [mg P/(g VSS·h)] | 1.2 | – | – | 0.0–5.9d |
| NRR [mg N/(g VSS·h)] | 1.3 | – | – | 1.0–10.0i,j |
| P/PHA (P-mol/C-mol) | 0.37 | – | – | 0.46e |
| Aerobic results | ||||
| P uptake (mg P/g VSS) | 8.6 | 7.7 | 3.6–18.2a | 4.1–15.0b |
| Pup [mg P/(g VSS·h)] | 4.4 | 3.8 | – | 0.7–19.2c,d |
| P/PHA (P-mol/C-mol) | 0.75 | 0.82 | 0.71–0.90a | 0.20–3.68c,k |
Note: activated sludge samples and fermentate were collected, preserved, and transported from the pilot plant in Virginia Beach, VA to Cornell lab in Ithaca, NY for the batch tests. HAc: acetate; Prel: maximum specific P release rate; VFAup: maximum specific VFA uptake rate; P/VFA: P release to VFA uptake ratio; PHA/VFA: PHA production to VFA uptake ratio; PHV/PHB: PHV production to PHB production ratio; Pup: maximum specific P uptake rate; NRR: maximum specific NOx reduction rate; P/PHA: P uptake to PHA utilization ratio. a: Qiu et al. (2019); b: Gu et al. (2008); c: Onnis-Hayden et al. (2020); d: López-Vázquez et al. (2008); e: Kuba et al. (1996); f: Lanham et al. (2013); g: Smolders et al. (1994a); h: Hesselmann et al. (2000); i: Henze (1991); j: Kujawa and Klapwijk (1999); k: Smolders et al. (1994b).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



