
Submitted:
01 April 2023
Posted:
03 April 2023
You are already at the latest version
Abstract
The Chromogranin A (CgA)-derived peptide Catestatin (CST: hCgA352-372) is highly conserved (>90% in 90% species) in mammalian species. The highest-evolved primates show 58.8% similarity with the lowest-evolved monotremes. CST was initially identified as a physiological brake in catecholamine secretion by inhibiting nicotinic-cholinergic signaling. CST also inhibits desensitization of catecholamine secretion, indicating that CST can act both as a cholinergic antagonist (short-term) and as a cholinergic agonist (long-term). The long-term effect sustains catecholamine secretion during stressful situations. CST is now established as a pleiotropic hormone: it affects the cardiovascular system by lowering blood pressure and cardiac contractility, enhancing baroreflex sensitivity, increasing heart rate variability, and promoting angiogenesis; and it increases insulin sensitivity by reducing inflammation, inhibiting hepatic glucose production, attenuating endoplasmic reticulum stress, and inducing glycogen production. The present review will highlight the important direct and indirect effects of CST, CST1-15 (aka cateslytin), D-CST1-15 (where L-amino acids were changed to D-amino acids), and human variants of CST (Gly364Ser-CST and Pro370Leu-CST) on microbial growth inhibition and their potential as therapy for antibiotic-resistant pathogenic microbes.
Keywords:
Chromogranin A
; catestatin
; gut microbiome
; antimicrobial peptide
; cell permeable peptide
1. Introduction
Chromogranin A (CgA), the acidic and secretory proprotein [1,2,3], is proteolytically cleaved to generate several biologically important peptides including Catestatin (CST: hCgA352-372) [4,5,6,7,8,9,10,11,12]. The 21 amino acid peptide CST was identified in 1997 as a physiologic brake in catecholamine secretion that acts by non-competitive inhibition of nicotinic-cholinergic signaling [5,13,14,15,16,17]. Initial studies were conducted in rat pheochromocytoma cells and primary cultures of bovine chromaffin cells with bovine CST (bCgA344-364) [5]. In 1999, CST was shown to inhibit desensitization of catecholamine secretion [18]. CST blocks nicotinic desensitization of catecholamine release, which might sustain catecholamine release when there is increased sympathetic outflow, such as during stressful situations. Thus, CST can act both as an antagonist to nicotinic-cholinergic transmission (short-term) as well as a partial agonist to nicotinic-cholinergic receptor (long-term). Alanine substitution mutants of bovine CST revealed crucial roles for Arg351, Arg353, and Arg358 in inhibiting nicotine-induced catecholamine secretion [19]. Further studies with bovine CST using serial single amino acid truncations or single residue substitution by alanine identified the N-terminal 15 amino acids (bCST344-358) as crucial both for nicotine-induced catecholamine secretion and desensitization [14]. Subsequently, in 2003, the anti-adrenergic effects of CST were demonstrated in mice [20]. The substantial (63%) β-sheet structure of CST in a hydrophobic environment was revealed in circular dichroism studies [19]. Electrophysiological studies on the interaction between CST and several combinations of neuronal acetylcholine receptor (nAChR) subunits expressed in oocytes revealed CST as a potent and reversible blocker of the nAChR, without significant discrimination among different nAChR subtypes (α7, α3β4, α3β2, and α4β2) tested [21]. The above studies were also shown by the molecular modeling of nAChRs and their interaction with CST [22]. Amongst the three natural variants of CST (Gly364Ser, Gly367Val, and Pro370Leu) in US population, Pro370Leu-CST showed the highest potency of inhibiting catecholamine secretion and desensitizing catecholamine secretion, followed by WT-CST and Gly364-Ser-CST [23,24,25].
The hypotensive effects of intravenous CST were first shown in rats in 1998 after activation of the sympathetic outflow by electrical stimulation. CST effects were shown to be blocked by a histamine H1 receptor antagonist but not by blockade of the α- and β-adrenergic receptors [26]. In 2010, hand vein studies in humans also revealed the hypotensive effects of CST [27]. In 2005, the anti-hypertensive effects of CST were exhibited in a monogenetic model of rodent hypertension (CgA knockout mice) with excess plasma catecholamines [28]. Besides excess catecholamines, reactive oxygen species (ROS) were also implicated in the development of hypertension in CgA-KO mice [29]. The antihypertensive effects of CST were later determined in mice (polygenic model of hypertension) and spontaneously hypertensive rats (polygenic model of hypertension) in 2012 and 2014, respectively [30,31]. CST was also demonstrated to improve dampened baroreflex sensitivity [32] and dysregulated heart rate variability in CgA-KO mice [33]. Central effects of CST on regulation of arterial blood pressure include excitation of both the bulbospinal (glutamatergic) neurons in the rostral ventrolateral medulla (RVLM) and GABAergic neurons in the caudal ventrolateral medulla (CVLM). Loss and gain of function studies in CST knockout mice confirmed that CST is necessary and sufficient to regulate blood pressure [34]. The effects of natural variants of CST on blood pressure and autonomic nervous system have been described previously in a review article [35]. Several studies in Langendorff perfused heart preparations implicate CST as a cardioprotective peptide inhibiting both inotropy and lusitropy under basal and stimulated conditions [36,37,38]. CST was also indicated to provide comparable cardioprotection in isolated heart by pre-conditioning [39] and post-conditioning [40]. Plasma CST is diminished very early during development of hypertension [41], and the processing of CgA to CST is decreased in hypertensive patients [42]. Plasma CST is also low in black patients suffering from end-stage renal disease [43]. CST increases insulin sensitivity by decreasing infiltration of macrophages in the liver and the consequent inflammation [44], inhibiting hepatic glucose production [44], decreasing endoplasmic reticulum stress [45], and inducing glycogen production [46]. Although it is known that neurotransmitters released from parasympathetic (e.g., acetylcholine) and sympathetic nerve ending (e.g., norepinephrine) can bind with the acetylcholine receptors [47,48,49,50,51] and adrenergic receptors [52,53,54,55,56], respectively present on innate immune cells, the homeostatic regulation between these two systems is complex. CST, being an intrinsic regulator of neuroendocrine secretory system, can have both direct and indirect effects on the host-pathogen interactions. The goal of this review is to summarize the developing concept of the effect of CST as an antimicrobial and cell penetrating peptide.
2. Homology and non-synonymous single nucleotide polymorphisms of CST in mammals
2.1. Homology of CST in mammals
Sequence alignment of CST in 53 mammalian species belonging to 8 orders revealed >80% homology in 52 species except in Platypus (lowest in the mammalian phylogenetic tree) where the homology with the Primates (highest in the mammalian phylogenetic tree) was >58% (Figure 1), indicating that CST is highly conserved in mammals. The homology of individual amino acids is summarized in Figure 2.
2.2. Single nucleotide polymorphisms (SNPs) in the CST domain of mammals
Four non-synonymous SNPs have been identified in CST domain of CgA: Gly364Ser (US, Indian, and Japanese populations) [24,57,58], Gly367Val (only in Indian populations) [57], Pro370Leu (US and Indian populations) [24], and Arg374Gln (US populations only) [24]. Pro370Leu-CST has the highest potency of inhibiting catecholamine secretion and desensitizing catecholamine secretion, followed by WT-CST and Gly364-Ser-CST [24]. The Gly364Ser variant was demonstrated to cause profound changes in parasympathetic and sympathetic activity including a ~2.4-fold increase in cardiac parasympathetic index and a ~26% decrease in cardiac sympathetic index in comparison to wild-type individuals [59]. This variant protects men from developing cardiovascular diseases compared to women [59].
3. Catestatin and innate immunity
The first indication for the role of CST in innate immunity came from a study in rats where intravenous administration of CST was shown to reduce pressor responses by electrical stimulation [26]. The hypotensive effect of CST was revealed to be mediated at least in part by profuse histamine release (by ~21-fold) and action at the H1 receptor [26]. The in vivo studies were later confirmed in peritoneal and pleural mast cells where CST caused dose-dependent release of histamine utilizing signaling pathways established for wasp venom peptide mastoparan and other amphiphilic cationic neuropeptides (the peptidergic pathway) [60]. This pathway is in sharp contrast to the nicotinic-cholinergic pathway used by CST to induce catecholamine secretion from chromaffin cells [5]. Subsequent studies uncover the following: (i) release of immunoreactive CST-containing peptides from human stimulated polymorphonuclear neutrophils [61]; (ii) detection of CST in mouse peritoneal macrophages by Western blots [39]; (iii) detection of CST in human monocytes and monocyte-derived macrophages by Western blots [62]; (iv) blockade of lipopolysaccharide (LPS)-induced increase in expression of tumor necrosis factor alpha (TNF-α) [62]; (v) decreased expression of proinflammatory cytokines by CST in plasma and heart [39]; (vi) inhibition of infiltration of macrophages in obese liver [44]; (vii) degranulation of primary mast cells from human peripheral blood [63] and (viii) low plasma CST in fatal COVID-19 patients [64]. These findings implicate CST as an immunomodulatory peptide. Since, receptor-ligand interactions are an essential driver of host-immune response [65], it is important to examine if CST can bind with a receptor on immune cells and regulate their polarization and function in host defense.
4. Antibacterial effects of CST
4.1. Inhibition of bacterial growth by CST
Metz-Boutigue’s group was the first to demonstrate the antibacterial activity of CST. Her group used bCST344-358 (coining the term cateslytin to describe this antibacterial effect) to reveal inhibition of growth of the Gram-positive and Gram-negative bacteria [61]. The minimal inhibitory concentrations (MICs) of CST (bCgA344-358, hCgA352-372, Gly364Ser-CST and Pro370Leu-CST) for Gram-positive bacteria (Micrococcus luteus, Bacillus megaterium, Group A Streptococcus, S. aureus ATCC 25923, S. aureus ATCC 49775, S. aureus S1 MRSA, S. aureus S1 MSSA, and S. aureus DmprF) range from 0.8 µM to >100 µM (Figure 3) [61,66]. The minimal concentration with 100% inhibition (MIC100) for Gram-positive bacteria range from 2 µM to >100 µM of 2 µM. The MIC of CST was higher (8 µM to 50 µM) for Gram-negative bacteria (Escherichia coli D22, E. coli 029, and Pseudomonas aeruginosa) compared to Gram-positive bacteria (Figure 3). Likewise, the MIC100 of CST was higher (15 µM to 150 µM) for Gram-negative bacteria compared to Gram-positive bacteria (Figure 3). The higher MIC and MIC100 values of CST for Gram-negative bacteria are consistent with the presence of extra outer membrane containing LPS [67,68]. Besides the extra-thick cell membrane, Gram-negative bacteria release exotoxins like tetanus [69,70] and cholera toxins [71] that make worse prognosis.
4.2. Interaction of CST with the bacterial wall
While Gram-positive bacteria contain only the cytoplasmic membrane that surrounds the cell, Gram-negative bacteria contain an additional LPS-containing thick outer membrane [67,68]. Furthermore, the peptidoglycan layer on the outer side of the cytoplasmic membrane is much thicker in Gram-positive bacteria compared to Gram-negative bacteria [72,73]. In addition, the LPS in Gram-positive bacteria are lipoteichoic acids that are embedded in the cytoplasmic membrane [74]. In contrast, in Gram-negative bacteria, the LPS forms the major lipid component of the outer leaflet of the outer membrane [74]. Although the membranes in both groups possess phosphate groups and are negatively charged, the Gram-negative bacteria in general have a higher content of the zwitterionic phospholipid is phosphatidylethanolamine than Gram-positive bacteria [74,75,76,77]. The predominant anionic lipids in bacterial membranes are phosphatidylglycerol and cardiolipin [78,79,80]. Metz-Boutigue’s group has shown that cateslytin (bCgA344-358) is unstructured in solution but is converted to an antiparallel β-structure and forms aggregates at the surface of negatively charged bacterial membranes [81]. As for catecholamine secretion [14], arginine residues were found to be crucial for binding to negatively charged lipids [81,82]. They proposed that the phase boundary defects caused by zones of different rigidity and thickness lead to permeability induction and peptide crossing through the bacterial membrane [81]. That CST penetrates through the bacterial wall was shown by measuring the optical density of the released β-galactosidase from ML-35p [66]. Electron microscopical studies of E. coli ML-35p confirmed that CST rapidly disrupts the E. coli membrane with visible membrane blebbing compared to untreated cells within 10 minutes [66].
4.3. CST as a potential therapy for bacterial diseases
The antimicrobial peptides (AMPs) derived from CgA display antimicrobial activities by lytic effects at micromolar range against Gram-positive bacteria, filamentous fungi, and yeasts. Interestingly, Catestatin-derived peptides can kill superbugs and more particularly S. aureus [83]. Considering the actions of CST on E. coli, it could be useful as a therapeutic target for the Gram-negative bacteria cause many serious infections, including Cholera [84], E. coli infection [85], Plague [86], Campylobacter [87], Legionnaire’s disease [88], Salmonella [89], Klebsiella [90], Pseudomonas [91], Tularemia [92], and Typhoid fever [93] and microbes associated with drug resistance [94,95,96,97], CST might be used as a therapeutic target for the above diseases.
5. Antifungal and antiyeast effects of CST
5.1. Inhibition of growth of fungus and yeast by CST
Fungal infections are common on the surface of skin, nails, or mucous membranes (superficial or mucocutaneous), underneath skin (subcutaneous), or in the lungs, brain, or heart (deep infection). Deep fungal infections include Histoplasmosis [98,99], Coccidioidomycosis (Valley fever) [100,101], Blastomycosis [102,103], Aspergillosis [104,105], Candidal urinary tract infection [106,107], invasive candidiasis [108,109], Pneumocystis pneumonia [110,111], Mucormycosis [112,113], and Cryptococcosis [114,115]. It is becoming increasingly evident that resistance to antifungal therapy is on the rise [116,117], which calls for the development alternative therapy for these infections. Host-defense peptides are emerging as new promising candidates to counteract antifungal resistance [118]. To this end, Metz-Boutigue’s group tested the effects of CST on the growth of fungus and yeasts. They found MIC values of CST or its human variants ranging from 0.2 µM to 75 µM against a host of fungal species (Neurospora crassa, Aspergillus fumigatus, A. niger, Nectria haematococca, Fusarium culmorum, F. oxysporum, Trichophyton mentagrophytes, and T. rubrum) [61,66] (Figure 3). The MIC100 values of CST or its human variants against the above fungal species ranged from 0.8 µM to 100 µM [61,66] (Figure 4). CST and its human variants also displayed similar inhibitory effects on the growth of yeasts with MIC ranged from 1.2 µM to >240 µM (Figure 4) [61,66]. The MIC100 of CST and its variants against the above yeasts ranged from 6 µM to 75 µM [61,66] (Figure 5). Similar to the effects of retro-inverso (RI)-CST on catecholamine secretion [30], D-CST (L-amino acids were replaced by D-amino acids) exhibited comparable inhibitory effects on the growth of yeast compared to L-CST with MIC ranged from 2 µM to 9.6 µM [119]. D-CST was also uncovered to be resistant to proteolytic digestion [83,119,120]. Like L-CST, D-CST can also be used to develop therapies for drug-resistant microbial infection [121].
5.2. Mechanisms underlying the antifungal and antiyeast activities of CST
Metz-Boutigue’s group used confocal laser microscopy to analyze the interaction of the synthetic rhodamine-labeled cateslytin (bCgA344-358R) with fungal (A. fumigatus) and yeast (C. albicans) membranes [61]. Rhodaminated cateslytin (1 µM) was detected in the inner compartment after 2 min of incubation, implicating rapid and efficient penetration through the cell wall [61]. Using time-lapse videomicroscopy of fungal growth, they have shown that rhodaminated cateslytin blocked the growth and development of nascent fungus [61]. Penetration of rhodaminated cateslytin takes place at both ends of the small fungi (three cells and expressing a slow growth rate) as compared to larger fungi with a higher growth rate where penetration takes place at one end [61]. Sequence homology of the well-known cell-permeable peptide penetratin with CST representing 7 mammalian orders (Primates, Rodentia, Artiodactyla, Perissodactyla, Carnivora, Cetacea and Monotremata) revealed 63.64% to 75% similarity, which should qualify CST as a cell-permeable peptide (Figure 6).
6. CST regulation of gut microbiota
6.1. Microbiomes in colonic mucosa versus feces
Recent studies have identified a larger role of gut microbiota in gut-immune homeostasis and in intestinal pathology. The human intestinal microbiota is dominated by five phyla: Bacillota (aka Firmicutes), Bacteroidota, Actinomycetota (aka Actinobacteria), Pseudomonadota (aka Proteobacteri), and Verrucomicrobiota [122]. In contrast, the intestinal microbiota of mice is dominated by four phyla: Bacillota, Bacteroidota, Deferribacterota, and Pseudomonadota [123], where the phylum Deferribacterota is in high abundance and the phyla Actinomycetota and Verrucomicrobiota are in low abundance compared to humans. In human adults, more than 80% of the species belong to just two phyla: Gram-negative Bacteriodota and Gram-positive Bacillota (aka Firmicutes). In mouse colonic mucosa samples, 19 phyla were identified [124]. The high-abundant (≥1%) phyla included Bacillota, Bacteroidota, Deferribacterota, and Pseudomonadota [124] (Figure 7). The low-abundant (<1%) phyla included the following: Actinomycetota (aka Actinobacteria), Parcubacteria (aka OD1), Saccharibacteria (aka TM7), Omnitrophota (aka OP3), Acidobacteriota, Armatimonadota, Chlamydiae, Chlorobiota, Cyanobacteria, Fibrobacterota, Mycoplasmatota (aka Tenericutes), Lentisphaerota, Planctomycetota, Spirochaetes, and Verrucomicrobiota) [124] (Figure 7). Although CST failed to alter bacterial populations in the four high-abundant phyla, it altered colonic mucosa-associated bacterial community composition at lower taxonomic levels. Thus, CST showed a positive association with Orders including Bacteroidales, Clostridiales, and YS2; Families including Chitinophagaceae, Clostridiaceae, Coriobacteriaceae, Pseudomonadaceae, Rikenellaceae, and Ruminococcaceae; and Genera Bifidobacterium and Stenotrophomonas [124]. Fecal samples identified 10 phyla including the high-abundant (≥1%) Bacillota, Bacteroidota, Pseudomonadota, and Deferribacterota and low-abundant (<1%) Actinomycetota, Cyanobacteria, Fibrobacterota, Saccharibacteria, Mycoplasmotota and Verrucomicrobiota [124]. The relative abundance of Bacteroidota was observed to increase significantly after CST treatment. In contrast, CST was learned to cause a marked decrease in Bacillota population. Like mucosal samples, fecal samples disclosed positive associations with the Class Alpharoteobacteria; Orders including Bacteroidales, RF32, and YS2; and genera Prevotella, Bacteroides, Ovatus, Parabacteroidesdistarosis, Parabacteroides, and Dorea (Figure 7). Bacteroides and Parabacteroides species, representing ~25% of the colonic microbiota, transform simple and complex sugars into volatile short-chain fatty acids (SCFAs) such as acetate, butyrate, and propionate [125,126,127], which are absorbed in colon as a nutrient [128,129,130]. Besides colonic nutrients, SCFAs are well established for their roles in accelerating gut transit time via release of serotonin [131,132,133,134]. SCFAs also release glucagon-like peptide 1 (GLP1) from the enteroendocrine L-cells [135,136,137,138] and improve insulin sensitivity [139,140,141]. Bacteroides thetaiotaomicron produces significant amounts of glycosylhydrolases, which prevent obesity [142]. Other Bacteroides species are also reported to prevent obesity and increase insulin sensitivity [143,144,145,146]. Furthermore, Bacteroides fragilis produces zwitterionic polysaccharide, which activates CD4+ T cells to produce interleukin 10 (IL-10). IL-10 plays crucial roles in preventing abscess formation and other unchecked inflammatory responses [142,147,148]. Negative association of CST was reported for the Class Clostrida, Families Bacteroidaceae and Ruminococcaceae, and Genera Adlercreutzia and Allobaculum [124]. Interestingly, the functional correlation between different CST mutants across species and their respective microbiota remains elusive. It is important to decipher the effect of different CST mutant on microbial diversity between species.
6.2. Microbiomes in CST knockout (CST-KO) mice and inflammation
CST knockout (CST-KO) mice were generated in 2018 and are: insulin-resistant on a normal chow diet [44], hyperadrenergic [39], hypertensive [39], and with a leaky gut [149]. The microbiome in CST-KO mice was unearthed to be quite different in composition than its WT littermates [149]. Microbial richness, assessed by the number of amplicon sequence variants (ASVs) and ACE (abundance-based Coverage Estimator) index revealed a significant decrease in CST-KO compared to WT mice [150]. The most prominent observation was the higher ratio of Bacillota to Bacteroidota, which was opposite to the decreased ratio observed in WT mice intrarectally infused with CST [124] (Figure 7). The ratio of Bacilotta to Bacteroidota plays crucial roles in regulation of obesity and metabolic syndrome [151,152]. Thus, the ratio of Bacilotta to Bacteroidota in adults increases as BMI increased [153]. Considering these reports, it remains to be seen if this change in microbiota can promote host defense against enteric pathogens like Salmonella. Furthermore, Verrucomicrobiota population was very low in CST-KO mice, which indicates low levels of Akkermansia species. Since Akkermansia muciniphila modulates obesity by regulating metabolism and energy homeostasis to improve insulin sensitivity and glucose homeostasis [154], low Verrucomicrobiota population contributes to the insulin resistance reported for CST-KO mice [44].
6.3. Alteration of diversity and composition of the microbiota in the CST-KO after supplementation with CST.
Decreased ASVs and ACE indices in CST-KO mice were restored after supplementation with CST for 15 days [150]. Like richness scores, supplementation of CST-KO mice with CST increased the diversity index as assessed by Shanon’s H and inverted Simpson’s index [150]. At the phylum level, CST was testified to decrease Bacillota phylum and increase Bacteroidota, Patescibacteria, Desulfobacterota, and Proteobacteria in both CST-KO and WT mice [150]. Low bacteria levels in Verrucomicrobiota phylum in CST-KO mice were markedly increased in CST-supplemented CST-KO mice. CST was also described to decrease abundance of Staphylococcus and Turicibacter in both WT and CST-KO mice. In contrast, CST increased Alistipes, Akkermansia, and Roseburia genera only in CST-KO mice [150]. A. muciniphila, a mucin-degrading bacterium, is a member of Verrucomicrobiota, which use mucin as a carbon, nitrogen, and energy source [155,156]. Murine studies show a causative role for A. muciniphila in lowering body fat mass, improving glucose homeostasis, decreasing adipose tissue inflammation, and increasing gut integrity [157,158,159]. Human studies show a negative correlation between A. muciniphila abundance and being overweight, being obese, having untreated type 2 diabetes mellitus, and having hypertension [160,161,162,163,164,165,166].
6.4. Restoration of microbial dysbiosis in CST-KO mice after fecal microbial transplant (FMT) from WT donor mice
FMT, first tested in 1958 to modify the human microbiome and consequently ameliorate fulminant pseudomembranous enterocolitis [167], has become established as an effective therapeutic modality in the treatment of antibiotic-refractory recurrent Clostridium difficile colitis with a success rate of up to 95% [168,169,170,171,172] . Subsequently, FMT was tested for the treatment of constipation, irritable bowel syndrome, and inflammatory bowel disease [173,174,175,176,177]. As discussed above, CST-KO mice are associated with an altered gut microbiota composition and richness. WT mice that received FMT from the CST-KO mice were shown to encompass a reduction of Clostridia and Akkermansia [178], which are linked to metabolic disorders and insulin resistance [179,180]. Furthermore, WTFMT-CST-KO mice exhibit a marked increase in the Proteobacteria population, which are associated with active inflammatory bowel disease (IBD) states [181,182]. Of note, CST-KO mice are insulin-resistant on normal chow diet [44]. In contrast, CST-KO that received FMT from the WT mice (CST-KOFMT-WT) showed an increase in richness, a notable reduction of Staphylococcus, and an increase in the butyrate-producing Intestinimonas [178] (Figure 7). I. butyriciproducens produces butyrate from not only sugars but also lysine and even glycated lysine [183]. Butyrate, taken up directly by colonocytes, serves as a direct source of energy and directly contributes to a healthy gut. In addition, butyrate as a signaling molecule affects many factors such as satiety, secretion of hormones, and glucose metabolism [184,185,186]. Of note, reduced levels of butyrate are strongly associated with IBD and metabolic disorders [187,188]. Further, butyrate has shown promise in restoring gut barrier integrity [189] and modulating regulatory T cell function by inhibiting histone deacetylase [190,191,192] though the former role is understudied. Butyrate modulation of certain serine proteases [193,194] would be a promising therapeutic target in future studies.
7. Conclusions
Alignment of CST sequences from 53 mammalian species belonging to eight orders revealed that CST sequence is highly conserved (>90% in 90% species) in mammals: Five (23.8%) amino acids (M3, L5, F7, F14, and G18) are 100% conserved; nine (42.8%) amino acids (S2, K4, S6, R8, R10, R15, P17, Q20 and L21) are 90-96% conserved; and three (14.28%) amino acids (A9, A11, and G16), are >80% conserved. The least conserved sequences are G13 (>66%) and P19 (>58%), where human variants of CST were reported for G13 (G13S) and P19 (P19L), indicating that natural selection pressures still exist on those two amino acids [195,196,197,198]. Existing literature (expression of CST in innate immune cells, inhibition of macrophage infiltration in tissues, decreased expression of pro-inflammatory cytokines by CST, and low plasma CST in fatal COVID-19 patients) implicate CST as an immunomodulatory peptide. Prominent effects of CST in low micromolar range on inhibition of growth of Gram-positive and Gram-negative bacteria, fungi, and yeast establish CST as an antimicrobial peptide. Penetration of CST (pI 12.03-12.48) in bacteria, fungus, yeast, and neutrophils, coupled with 70-75% homology with cell penetrating peptide Penetratin (pI 12.62) rightfully qualify CST as a member of the cell permeable peptide. Increased ratio of Bacilotta to Bacteroidota, together with low levels of Verrucomicrobiota (e.g., Akkermansia spp) in CST-KO mice, not only explains insulin resistance in CST-KO mice but also implicates that CST is necessary for the maintenance of insulin sensitivity. Decreased ratio of Bacilotta to Bacteroidota coupled with increased abundance of Verrucomicrobiota after supplementation of CST-KO mice with CST confirm that CST is necessary and sufficient to increase insulin sensitivity by modulating gut microbiota. Decreased population of Akkermansia and increased population of Proteobacteria in WTFMT-CST-KO coupled with increased population of butyrate producing Intestimonas in CST-KOFMT-WT further substantiates regulation of obesity and insulin resistance by CST [44] via regulation of gut microbial population [150,178].
Author Contributions
S.K.M. conceived the idea and wrote major portion of the manuscript. S.M. made all the Figures. S.J., S.M., S.D., S.C., and S.K.M. performed literature search, interpreted data, and wrote the Manuscript. All authors approved the submitted manuscript.
Funding
This research was supported by grants from the National Institutes of Health (1 R21 AG072487-01 and 1 R21 AG080246-01 to S.K.M). S.J. is supported by AFTD Holloway Postdoctoral Fellowship (Award #2020-02).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
S.K.M. is a co-inventor of a patent on CST regulation of obesity.
References
- Winkler, H.; Fischer-Colbrie, R. , The chromogranins A and B: the first 25 years and future perspectives. Neuroscience 1992, 49, 497–528. [Google Scholar] [CrossRef] [PubMed]
- Iacangelo, A.L.; Eiden, L.E. , Chromogranin A: current status as a precursor for bioactive peptides and a granulogenic/sorting factor in the regulated secretory pathway. Regul. Pept. 1995, 58, 65–88. [Google Scholar] [CrossRef]
- Montero-Hadjadje, M.; Vaingankar, S.; Elias, S.; Tostivint, H.; Mahata, S.K.; Anouar, Y. , Chromogranins A and B and secretogranin II: evolutionary and functional aspects. Acta Physiol. (Oxf.) 2008, 192, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Bartolomucci, A.; Possenti, R.; Mahata, S.K.; Fischer-Colbrie, R.; Loh, Y.P.; Salton, S.R. , The extended granin family: structure, function, and biomedical implications. Endocr. Rev. 2011, 32, 755–797. [Google Scholar] [CrossRef] [PubMed]
- Mahata, S.K.; O’Connor, D.T.; Mahata, M.; Yoo, S.H.; Taupenot, L.; Wu, H.; Gill, B.M.; Parmer, R.J. , Novel autocrine feedback control of catecholamine release. A discrete chromogranin A fragment is a noncompetitive nicotinic cholinergic antagonist. J. Clin. Invest. 1997, 100, 1623–1633. [Google Scholar] [CrossRef] [PubMed]
- Mahata, S.K.; Corti, A. , Chromogranin A and its fragments in cardiovascular, immunometabolic, and cancer regulation. Ann. N. Y. Acad. Sci. 2019, 1455, 34–58. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.V.; Taupenot, L.; Mahata, S.K.; Mahata, M.; Wu, H.; Yasothornsrikul, S.; Toneff, T.; Caporale, C.; Jiang, Q.; Parmer, R.J.; Hook, V.Y.; O’Connor, D.T. , Formation of the catecholamine release-inhibitory peptide catestatin from chromogranin A. Determination of proteolytic cleavage sites in hormone storage granules. J. Biol. Chem. 2000, 275, 22905–22915. [Google Scholar] [CrossRef] [PubMed]
- Parmer, R.J.; Mahata, M.; Gong, Y.; Mahata, S., K.; Jiang, Q.; O’Connor, D.T.; Xi, X.-P.; Miles, L.A. , Processing of chromogranin A by plasmin provides a novel mechanism for regulating catecholamine secretion. J. Clin. Invest. 2000, 106, 907–915. [Google Scholar] [CrossRef]
- Jiang, Q.; Taupenot, L.; Mahata, S.K.; Mahata, M.; O’Connor, D.T.; Miles, L.A.; Parmer, R.J. , Proteolytic cleavage of chromogranin A (CgA) by plasmin: selective liberation of a specific bioactive CgA fragment that regulates catecholamine release. J. Biol. Chem. 2001, 276, 25022–25029. [Google Scholar] [CrossRef]
- Lee, J.C.; Taylor, C.V.; Gaucher, S.P.; Toneff, T.; Taupenot, L.; Yasothornsrikul, S.; Mahata, S.K.; Sei, C.; Parmer, R.J.; Neveu, J.M.; Lane, W.S.; Gibson, B.W.; O’Connor, D.T.; Hook, V.Y. , Primary sequence characterization of catestatin intermediates and peptides defines proteolytic cleavage sites utilized for converting chromogranin a into active catestatin secreted from neuroendocrine chromaffin cells. Biochemistry 2003, 42, 6938–6946. [Google Scholar] [CrossRef]
- Biswas, N.; Vaingankar, S.M.; Mahata, M.; Das, M.; Gayen, J.R.; Taupenot, L.; Torpey, J.W.; O’Connor, D.T.; Mahata, S.K. , Proteolytic cleavage of human chromogranin a containing naturally occurring catestatin variants: differential processing at catestatin region by plasmin. Endocrinology 2008, 149, 749–757. [Google Scholar] [CrossRef]
- Biswas, N.; Rodriguez-Flores, J.L.; Courel, M.; Gayen, J.R.; Vaingankar, S.M.; Mahata, M.; Torpey, J.W.; Taupenot, L.; O’Connor, D.T.; Mahata, S.K. , Cathepsin L Co-Localizes with Chromogranin a in Chromaffin Vesicles to Generate Active Peptides. Endocrinology 2009, 150, 3547–3557. [Google Scholar] [CrossRef]
- Taupenot, L.; Mahata, S.K.; Mahata, M.; Parmer, R.J.; O’Connor, D.T. , Interaction of the catecholamine release-inhibitory peptide catestatin (human chromogranin A(352-372)) with the chromaffin cell surface and Torpedo electroplax: implications for nicotinic cholinergic antagonism. Regul. Pept. 2000, 95, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Mahata, S.K.; Mahata, M.; Wakade, A.R.; O’Connor, D.T. , Primary structure and function of the catecholamine release inhibitory peptide catestatin (chromogranin A344-364): Identification of amino acid residues crucial for activity. Mol. Endocrinol. 2000, 14, 1525–1535. [Google Scholar]
- Preece, N.E.; Nguyen, M.; Mahata, M.; Mahata, S.K.; Mahapatra, N.R.; Tsigelny, I.; O’Connor, D.T. , Conformational preferences and activities of peptides from the catecholamine release-inhibitory (catestatin) region of chromogranin A. Regul. Pept. 2004, 118, 75–87. [Google Scholar] [CrossRef]
- Mahata, S.K. , Catestatin - The catecholamine release inhibitory peptide: A structural and functional overview. Curr Med Chem - Immun Endoc & Metab Agents 2004, 4, 221–234. [Google Scholar]
- Mahapatra, N.R.; Mahata, M.; Mahata, S.K.; O’Connor, D.T. , The chromogranin A fragment catestatin: specificity, potency and mechanism to inhibit exocytotic secretion of multiple catecholamine storage vesicle co-transmitters. J. Hypertens. 2006, 24, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Mahata, S.K.; Mahata, M.; Parmer, R.J.; O’Connor, D.T. , Desensitization of catecholamine release: The novel catecholamine release-inhibitory peptide catestatin (chromogranin A344-364) acts at the receptor to prevent nicotinic cholinergic tolerance. J. Biol. Chem. 1999, 274, 2920–2928. [Google Scholar] [CrossRef] [PubMed]
- Tsigelny, I.; Mahata, S.K.; Taupenot, L.; Preece, N.E.; Mahata, M.; Khan, I.; Parmer, R.J.; O’Connor, D.T. , Mechanism of action of chromogranin A on catecholamine release: molecular modeling of the catestatin region reveals a b-strand/loop/b-strand structure secured by hydrophobic interactions and predictive of activity. Regul. Pept. 1998, 77, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Mahata, S.K.; Mahapatra, N.R.; Mahata, M.; Wang, T.C.; Kennedy, B.P.; Ziegler, M.G.; O’Connor, D.T. , Catecholamine secretory vesicle stimulus-transcription coupling in vivo. Demonstration by a novel transgenic promoter/photoprotein reporter and inhibition of secretion and transcription by the chromogranin A fragment catestatin. J. Biol. Chem. 2003, 278, 32058–32067. [Google Scholar] [CrossRef]
- Herrero, C.J.; Ales, E.; Pintado, A.J.; Lopez, M.G.; Garcia-Palomero, E.; Mahata, S.K.; O’Connor, D.T.; Garcia, A.G.; Montiel, C. , Modulatory mechanism of the endogenous peptide catestatin on neuronal nicotinic acetylcholine receptors and exocytosis. J. Neurosci. 2002, 22, 377–388. [Google Scholar] [CrossRef]
- Sahu, B.S.; Mohan, J.; Sahu, G.; Singh, P.K.; Sonawane, P.J.; Sasi, B.K.; Allu, P.K.; Maji, S.K.; Bera, A.K.; Senapati, S.; Mahapatra, N.R. , Molecular interactions of the physiological anti-hypertensive peptide catestatin with the neuronal nicotinic acetylcholine receptor. J. Cell Sci. 2012, 125, 2323–2337. [Google Scholar] [CrossRef]
- Mahata, S.K.; Mahata, M.; Fung, M.M.; O’Connor, D.T. , Catestatin: a multifunctional peptide from chromogranin A. Regul. Pept. 2010, 162, 33–43. [Google Scholar] [CrossRef]
- Wen, G.; Mahata, S.K.; Cadman, P.; Mahata, M.; Ghosh, S.; Mahapatra, N.R.; Rao, F.; Stridsberg, M.; Smith, D.W.; Mahboubi, P.; Schork, N.J.; O’Connor, D.T.; Hamilton, B.A. , Both rare and common polymorphisms contribute functional variation at CHGA, a regulator of catecholamine physiology. Am. J. Hum. Genet. 2004, 74, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Mahata, S.K.; Mahata, M.; Wen, G.; Wong, W.B.; Mahapatra, N.R.; Hamilton, B.A.; O’Connor, D.T. , The catecholamine release-inhibitory “catestatin” fragment of chromogranin a: naturally occurring human variants with different potencies for multiple chromaffin cell nicotinic cholinergic responses. Mol. Pharmacol. 2004, 66, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.P.; Mahata, S.K.; O’Connor, D.T.; Ziegler, M.G. , Mechanism of cardiovascular actions of the chromogranin A fragment catestatin in vivo. Peptides 1998, 19, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Fung, M.M.; Salem, R.M.; Mehtani, P.; Thomas, B.; Lu, C.F.; Perez, B.; Rao, F.; Stridsberg, M.; Ziegler, M.G.; Mahata, S.K.; O’Connor, D.T. , Direct vasoactive effects of the chromogranin A (CHGA) peptide catestatin in humans in vivo. Clin. Exp. Hypertens. 2010, 32, 278–287. [Google Scholar] [CrossRef]
- Mahapatra, N.R.; O’Connor, D.T.; Vaingankar, S.M.; Hikim, A.P.; Mahata, M.; Ray, S.; Staite, E.; Wu, H.; Gu, Y.; Dalton, N.; Kennedy, B.P.; Ziegler, M.G.; Ross, J.; Mahata, S.K. , Hypertension from targeted ablation of chromogranin A can be rescued by the human ortholog. J. Clin. Invest. 2005, 115, 1942–1952. [Google Scholar] [CrossRef]
- Gayen, J.R.; Zhang, K.; Ramachandrarao, S.P.; Mahata, M.; Chen, Y.; Kim, H.-S.; Naviaux, R.K.; Sharma, K.; Mahata, S.K.; O’Connor, D.T. , Role of reactive oxygen species in hyperadrenergic hypertension: Biochemical, physiological, and pharmacological evidence from targeted ablation of the chromogranin A gene. Circ. Cardiovasc. Genet. 2010, 3, 414–425. [Google Scholar] [CrossRef]
- Biswas, N.; Gayen, J.; Mahata, M.; Su, Y.; Mahata, S.K.; O’Connor, D.T. , Novel peptide isomer strategy for stable inhibition of catecholamine release: application to hypertension. Hypertension 2012, 60, 1552–1559. [Google Scholar] [CrossRef]
- Avolio, E.; Mahata, S.K.; Mantuano, E.; Mele, M.; Alo, R.; Facciolo, R.M.; Talani, G.; Canonaco, M. , Antihypertensive and neuroprotective effects of catestatin in spontaneously hypertensive rats: Interaction with GABAergic transmission in amygdala and brainstem. Neuroscience 2014, 270, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Gayen, J.R.; Gu, Y.; O’Connor, D.T.; Mahata, S.K. , Global disturbances in autonomic function yield cardiovascular instability and hypertension in the chromogranin A null mouse. Endocrinology 2009, 150, 5027–5035. [Google Scholar] [CrossRef]
- Dev, N.B.; Gayen, J.R.; O’Connor, D.T.; Mahata, S.K. , Chromogranin A and the autonomic system: Decomposition of heart rate variability by time and frequency domains, along with non-linear characteristics during chromogranin A ablation, with “rescue” by its catestatin. Endocrinology 2010, 151, 2760–2768. [Google Scholar] [CrossRef] [PubMed]
- Gaede, A.H.; Pilowsky, P.M. , Catestatin in rat RVLM is sympathoexcitatory, increases barosensitivity, and attenuates chemosensitivity and the somatosympathetic reflex. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R1538–1545. [Google Scholar] [CrossRef]
- Mahata, S.K.; Kiranmayi, M.; Mahapatra, N.R. , Catestatin: A Master Regulator of Cardiovascular Functions. Curr. Med. Chem. 2018, 25, 1352–1374. [Google Scholar] [CrossRef] [PubMed]
- Angelone, T.; Quintieri, A.M.; Brar, B.K.; Limchaiyawat, P.T.; Tota, B.; Mahata, S.K.; Cerra, M.C. , The antihypertensive chromogranin a peptide catestatin acts as a novel endocrine/paracrine modulator of cardiac inotropism and lusitropism. Endocrinology 2008, 149, 4780–4793. [Google Scholar] [CrossRef] [PubMed]
- Imbrogno, S.; Garofalo, F.; Cerra, M.C.; Mahata, S.K.; Tota, B. , The catecholamine release-inhibitory peptide catestatin (chromogranin A344-364) modulates myocardial function in fish. J. Exp. Biol. 2010, 213, 3636–3643. [Google Scholar] [CrossRef]
- Mazza, R.; Gattuso, A.; Mannarino, C.; Brar, B.K.; Barbieri, S.F.; Tota, B.; Mahata, S.K. , Catestatin (chromogranin A344-364) is a novel cardiosuppressive agent: inhibition of isoproterenol and endothelin signaling in the frog heart. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H113–122. [Google Scholar] [CrossRef]
- Ying, W.; Tang, K.; Avolio, E.; Schilling, J.M.; Pasqua, T.; Liu, M.A.; Cheng, H.; Gao, H.; Zhang, J.; Mahata, S.; Ko, M.S.; Bandyopadhyay, G.; Das, S.; Roth, D.M.; Sahoo, D.; Webster, N.J.G.; Sheikh, F.; Ghosh, G.; Patel, H.H.; Ghosh, P.; van den Bogaart, G.; Mahata, S.K. , Immunosuppression of Macrophages Underlies the Cardioprotective Effects of CST (Catestatin). Hypertension 2021, 77, 1670–1682. [Google Scholar] [CrossRef]
- Penna, C.; Alloatti, G.; Gallo, M.P.; Cerra, M.C.; Levi, R.; Tullio, F.; Bassino, E.; Dolgetta, S.; Mahata, S.K.; Tota, B.; Pagliaro, P. , Catestatin improves post-ischemic left ventricular function and decreases ischemia/reperfusion injury in heart. Cell. Mol. Neurobiol. 2010, 30, 1171–1179. [Google Scholar] [CrossRef]
- O’Connor, D.T.; Kailasam, M.T.; Kennedy, B.P.; Ziegler, M.G.; Yanaihara, N.; Parmer, R.J. , Early decline in the catecholamine release-inhibitory peptide catestatin in humans at genetic risk of hypertension. J. Hypertens. 2002, 20, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.T.; Zhu, G.; Rao, F.; Taupenot, L.; Fung, M.M.; Das, M.; Mahata, S.K.; Mahata, M.; Wang, L.; Zhang, K.; Greenwood, T.A.; Shih, P.A.; Cockburn, M.G.; Ziegler, M.G.; Stridsberg, M.; Martin, N.G.; Whitfield, J.B. , Heritability and genome-wide linkage in US and australian twins identify novel genomic regions controlling chromogranin a: implications for secretion and blood pressure. Circulation 2008, 118, 247–257. [Google Scholar] [CrossRef]
- Salem, R.M.; Cadman, P.E.; Chen, Y.; Rao, F.; Wen, G.; Hamilton, B.A.; Rana, B.K.; Smith, D.W.; Stridsberg, M.; Ward, H.J.; Mahata, M.; Mahata, S.K.; Bowden, D.W.; Hicks, P.J.; Freedman, B.I.; Schork, N.J.; O’Connor, D.T. , Chromogranin A polymorphisms are associated with hypertensive renal disease. J. Am. Soc. Nephrol. 2008, 19, 600–614. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.; Mahata, S.; Bandyopadhyay, G.K.; Zhou, Z.; Wollam, J.; Vu, J.; Mayoral, R.; Chi, N.W.; Webster, N.J.G.; Corti, A.; Mahata, S.K. , Catestatin Inhibits Obesity-Induced Macrophage Infiltration and Inflammation in the Liver and Suppresses Hepatic Glucose Production, Leading to Improved Insulin Sensitivity. Diabetes 2018, 67, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Bandyopadhyay, G.K.; Ray, I.; Bandyopadhyay, K.; Chowdhury, N.; De, R.K.; Mahata, S.K. , Catestatin improves insulin sensitivity by attenuating endoplasmic reticulum stress: In vivo and in silico validation. Comput. Struct. Biotechnol. J. 2020, 18, 464–481. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, G.; Tang, K.; Webster, N.J.G.; van den Bogaart, G.; Mahata, S.K. , Catestatin induces glycogenesis by stimulating the phosphoinositide 3-kinase-AKT pathway. Acta Physiol. (Oxf.) 2022, e13775. [Google Scholar] [CrossRef]
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, H.; Ulloa, L.; Al-Abed, Y.; Czura, C.J.; Tracey, K.J. , Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef]
- Mashimo, M.; Fujii, T.; Ono, S.; Moriwaki, Y.; Misawa, H.; Kawashima, K. , Minireview: Divergent roles of alpha7 nicotinic acetylcholine receptors expressed on antigen-presenting cells and CD4(+) T cells in the regulation of T cell differentiation. Int. Immunopharmacol. 2020, 82, 106306. [Google Scholar] [CrossRef]
- Nakata, Y.; Miura, K.; Yamasaki, N.; Ogata, S.; Miura, S.; Hosomi, N.; Kaminuma, O. , Expression and Function of Nicotinic Acetylcholine Receptors in Induced Regulatory T Cells. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Rueda Ruzafa, L.; Cedillo, J.L.; Hone, A.J. , Nicotinic Acetylcholine Receptor Involvement in Inflammatory Bowel Disease and Interactions with Gut Microbiota. Int. J. Environ. Res. Public Health 2021, 18. [Google Scholar] [CrossRef]
- Hone, A.J.; McIntosh, J.M. , Nicotinic acetylcholine receptors: Therapeutic targets for novel ligands to treat pain and inflammation. Pharmacol. Res. 2023, 190, 106715. [Google Scholar] [CrossRef] [PubMed]
- Araujo, L.P.; Maricato, J.T.; Guereschi, M.G.; Takenaka, M.C.; Nascimento, V.M.; de Melo, F.M.; Quintana, F.J.; Brum, P.C.; Basso, A.S. , The Sympathetic Nervous System Mitigates CNS Autoimmunity via beta2-Adrenergic Receptor Signaling in Immune Cells. Cell. Rep. 2019, 28, 3120–3130 e3125. [Google Scholar] [CrossRef] [PubMed]
- Emeny, R.T.; Gao, D.; Lawrence, D.A. , Beta1-adrenergic receptors on immune cells impair innate defenses against Listeria. J. Immunol. 2007, 178, 4876–4884. [Google Scholar] [CrossRef] [PubMed]
- Lorton, D.; Bellinger, D.L. , Molecular mechanisms underlying beta-adrenergic receptor-mediated cross-talk between sympathetic neurons and immune cells. Int. J. Mol. Sci. 2015, 16, 5635–5665. [Google Scholar] [CrossRef]
- Grisanti, L.A.; Perez, D.M.; Porter, J.E. , Modulation of immune cell function by alpha(1)-adrenergic receptor activation. Curr. Top. Membr. 2011, 67, 113–138. [Google Scholar]
- Sharma, D.; Farrar, J.D. , Adrenergic regulation of immune cell function and inflammation. Semin. Immunopathol. 2020, 42, 709–717. [Google Scholar] [CrossRef]
- Sahu, B.S.; Obbineni, J.M.; Sahu, G.; Allu, P.K.; Subramanian, L.; Sonawane, P.J.; Singh, P.K.; Sasi, B.K.; Senapati, S.; Maji, S.K.; Bera, A.K.; Gomathi, B.S.; Mullasari, A.S.; Mahapatra, N.R. , Functional genetic variants of the catecholamine-release-inhibitory peptide catestatin in an Indian population: allele-specific effects on metabolic traits. J. Biol. Chem. 2012, 287, 43840–43852. [Google Scholar] [CrossRef]
- Choi, Y.; Miura, M.; Nakata, Y.; Sugasawa, T.; Nissato, S.; Otsuki, T.; Sugawara, J.; Iemitsu, M.; Kawakami, Y.; Shimano, H.; Iijima, Y.; Tanaka, K.; Kuno, S.; Allu, P.K.; Mahapatra, N.R.; Maeda, S.; Takekoshi, K. , A common genetic variant of the chromogranin A-derived peptide catestatin is associated with atherogenesis and hypertension in a Japanese population. Endocr. J. 2015, 62, 797–804. [Google Scholar] [CrossRef]
- Rao, F.; Wen, G.; Gayen, J.R.; Das, M.; Vaingankar, S.M.; Rana, B.K.; Mahata, M.; Kennedy, B.P.; Salem, R.M.; Stridsberg, M.; Abel, K.; Smith, D.W.; Eskin, E.; Schork, N.J.; Hamilton, B.A.; Ziegler, M.G.; Mahata, S.K.; O’Connor, D.T. , Catecholamine release-inhibitory peptide catestatin (chromogranin A(352-372)): naturally occurring amino acid variant Gly364Ser causes profound changes in human autonomic activity and alters risk for hypertension. Circulation 2007, 115, 2271–2281. [Google Scholar] [CrossRef]
- Kruger, P.G.; Mahata, S.K.; Helle, K.B. , Catestatin (CgA344-364) stimulates rat mast cell release of histamine in a manner comparable to mastoparan and other cationic charged neuropeptides. Regul. Pept. 2003, 114, 29–35. [Google Scholar] [CrossRef]
- Briolat, J.; Wu, S.D.; Mahata, S.K.; Gonthier, B.; Bagnard, D.; Chasserot-Golaz, S.; Helle, K.B.; Aunis, D.; Metz-Boutigue, M.H. , New antimicrobial activity for the catecholamine release-inhibitory peptide from chromogranin A. Cell. Mol. Life Sci. 2005, 62, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Ozawa, N.; Mori, Y.; Takahashi, Y.; Watanabe-Kominato, K.; Shirai, R.; Watanabe, R.; Sato, K.; Matsuyama, T.A.; Ishibashi-Ueda, H.; Koba, S.; Kobayashi, Y.; Hirano, T.; Watanabe, T. , Catestatin Prevents Macrophage-Driven Atherosclerosis but Not Arterial Injury-Induced Neointimal Hyperplasia. Thromb. Haemost. 2018, 118, 182–194. [Google Scholar] [CrossRef]
- Aung, G.; Niyonsaba, F.; Ushio, H.; Kajiwara, N.; Saito, H.; Ikeda, S.; Ogawa, H.; Okumura, K. , Catestatin, a neuroendocrine antimicrobial peptide, induces human mast cell migration, degranulation and production of cytokines and chemokines. Immunology 2011, 132, 527–539. [Google Scholar] [CrossRef]
- Kljakovic-Gaspic, T.; Tokic, D.; Martinovic, D.; Kumric, M.; Supe-Domic, D.; Stojanovic Stipic, S.; Delic, N.; Vrdoljak, J.; Vilovic, M.; Ticinovic Kurir, T.; Bozic, J. , Prognostic Value of Catestatin in Severe COVID-19: An ICU-Based Study. J. Clin. Med. 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Jati, S.; Kundu, S.; Chakraborty, A.; Mahata, S.K.; Nizet, V.; Sen, M. , Wnt5A Signaling Promotes Defense Against Bacterial Pathogens by Activating a Host Autophagy Circuit. Front. Immunol. 2018, 9, 679. [Google Scholar] [CrossRef] [PubMed]
- Radek, K.A.; Lopez-Garcia, B.; Hupe, M.; Niesman, I.R.; Elias, P.M.; Taupenot, L.; Mahata, S.K.; O’Connor, D.T.; Gallo, R.L. , The neuroendocrine peptide catestatin is a cutaneous antimicrobial and induced in the skin after injury. J. Invest. Dermatol. 2008, 128, 1525–1534. [Google Scholar] [CrossRef]
- Scheffers, D.J.; Pinho, M.G. , Bacterial cell wall synthesis: new insights from localization studies. Microbiol. Mol. Biol. Rev. 2005, 69, 585–607. [Google Scholar] [CrossRef]
- Brown, A.R.; Gordon, R.A.; Hyland, S.N.; Siegrist, M.S.; Grimes, C.L. , Chemical Biology Tools for Examining the Bacterial Cell Wall. Cell. Chem. Biol. 2020, 27, 1052–1062. [Google Scholar] [CrossRef]
- Lopez-Siles, M.; Corral-Lugo, A.; McConnell, M.J. , Vaccines for multidrug resistant Gram negative bacteria: lessons from the past for guiding future success. FEMS Microbiol. Rev. 2021, 45. [Google Scholar] [CrossRef]
- Saier, M.H., Jr. , Protein secretion and membrane insertion systems in gram-negative bacteria. J. Membr. Biol. 2006, 214, 75–90. [Google Scholar] [CrossRef]
- Welch, R.A. , Pore-forming cytolysins of gram-negative bacteria. Mol. Microbiol. 1991, 5, 521–528. [Google Scholar] [CrossRef]
- Vollmer, W.; Seligman, S.J. , Architecture of peptidoglycan: more data and more models. Trends Microbiol. 2010, 18, 59–66. [Google Scholar] [CrossRef]
- Rojas, E.R.; Billings, G.; Odermatt, P.D.; Auer, G.K.; Zhu, L.; Miguel, A.; Chang, F.; Weibel, D.B.; Theriot, J.A.; Huang, K.C. , The outer membrane is an essential load-bearing element in Gram-negative bacteria. Nature 2018, 559, 617–621. [Google Scholar] [CrossRef]
- Percy, M.G.; Grundling, A. , Lipoteichoic acid synthesis and function in gram-positive bacteria. Annu. Rev. Microbiol. 2014, 68, 81–100. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. , Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef]
- Hancock, R.E.; Diamond, G. , The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef]
- Lee, T.H.; Hirst, D.J.; Aguilar, M.I. , New insights into the molecular mechanisms of biomembrane structural changes and interactions by optical biosensor technology. Biochim. Biophys. Acta 2015, 1848, 1868–1885. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M.; Rotem, S.; Mor, A.; Berno, B.; Epand, R.F. , Bacterial membranes as predictors of antimicrobial potency. J. Am. Chem. Soc. 2008, 130, 14346–14352. [Google Scholar] [CrossRef]
- Wadhwani, P.; Epand, R.F.; Heidenreich, N.; Burck, J.; Ulrich, A.S.; Epand, R.M. , Membrane-active peptides and the clustering of anionic lipids. Biophys. J. 2012, 103, 265–274. [Google Scholar] [CrossRef]
- Bogdanov, M.; Pyrshev, K.; Yesylevskyy, S.; Ryabichko, S.; Boiko, V.; Ivanchenko, P.; Kiyamova, R.; Guan, Z.; Ramseyer, C.; Dowhan, W. , Phospholipid distribution in the cytoplasmic membrane of Gram-negative bacteria is highly asymmetric, dynamic, and cell shape-dependent. Sci. Adv. 2020, 6, eaaz6333. [Google Scholar] [CrossRef]
- Jean-Francois, F.; Castano, S.; Desbat, B.; Odaert, B.; Roux, M.; Metz-Boutigue, M.H.; Dufourc, E.J. , Aggregation of cateslytin beta-sheets on negatively charged lipids promotes rigid membrane domains. A new mode of action for antimicrobial peptides? Biochemistry 2008, 47, 6394–6402. [Google Scholar] [CrossRef]
- Epand, R.M.; Epand, R.F. , Lipid domains in bacterial membranes and the action of antimicrobial agents. Biochim. Biophys. Acta 2009, 1788, 289–294. [Google Scholar] [CrossRef]
- Scavello, F.; Mutschler, A.; Helle, S.; Schneider, F.; Chasserot-Golaz, S.; Strub, J.M.; Cianferani, S.; Haikel, Y.; Metz-Boutigue, M.H. , Catestatin in innate immunity and Cateslytin-derived peptides against superbugs. Sci. Rep. 2021, 11, 15615. [Google Scholar] [CrossRef]
- Costa, T.R.; Felisberto-Rodrigues, C.; Meir, A.; Prevost, M.S.; Redzej, A.; Trokter, M.; Waksman, G. , Secretion systems in Gram-negative bacteria: structural and mechanistic insights. Nat. Rev. Microbiol. 2015, 13, 343–359. [Google Scholar] [CrossRef]
- Bonten, M.; Johnson, J.R.; van den Biggelaar, A.H.J.; Georgalis, L.; Geurtsen, J.; de Palacios, P.I.; Gravenstein, S.; Verstraeten, T.; Hermans, P.; Poolman, J.T. , Epidemiology of Escherichia coli Bacteremia: A Systematic Literature Review. Clin. Infect. Dis. 2021, 72, 1211–1219. [Google Scholar] [CrossRef]
- Pechous, R.D.; Sivaraman, V.; Stasulli, N.M.; Goldman, W.E. , Pneumonic Plague: The Darker Side of Yersinia pestis. Trends Microbiol. 2016, 24, 190–197. [Google Scholar] [CrossRef]
- Kim, S.H.; Chelliah, R.; Ramakrishnan, S.R.; Perumal, A.S.; Bang, W.S.; Rubab, M.; Daliri, E.B.; Barathikannan, K.; Elahi, F.; Park, E.; Jo, H.Y.; Hwang, S.B.; Oh, D.H. , Review on Stress Tolerance in Campylobacter jejuni. Front Cell. Infect. Microbiol. 2020, 10, 596570. [Google Scholar] [CrossRef]
- Cunha, B.A.; Burillo, A.; Bouza, E. , Legionnaires’ disease. Lancet 2016, 387, 376–385. [Google Scholar] [CrossRef]
- Coburn, B.; Grassl, G.A.; Finlay, B.B. , Salmonella, the host and disease: a brief review. Immunol. Cell Biol. 2007, 85, 112–118. [Google Scholar] [CrossRef]
- Lee, G.C.; Burgess, D.S. , Treatment of Klebsiella pneumoniae carbapenemase (KPC) infections: a review of published case series and case reports. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 32. [Google Scholar] [CrossRef]
- Capatina, D.; Feier, B.; Hosu, O.; Tertis, M.; Cristea, C. , Analytical methods for the characterization and diagnosis of infection with Pseudomonas aeruginosa: A critical review. Anal. Chim. Acta 2022, 1204, 339696. [Google Scholar] [CrossRef]
- Hennebique, A.; Boisset, S.; Maurin, M. , Tularemia as a waterborne disease: a review. Emerg. Microbes Infect. 2019, 8, 1027–1042. [Google Scholar] [CrossRef] [PubMed]
- Butler, T. , Treatment of typhoid fever in the 21st century: promises and shortcomings. Clin. Microbiol. Infect. 2011, 17, 959–963. [Google Scholar] [CrossRef]
- Mazzariol, A.; Bazaj, A.; Cornaglia, G. , Multi-drug-resistant Gram-negative bacteria causing urinary tract infections: a review. J. Chemother. 2017, 29, 2–9. [Google Scholar] [CrossRef]
- Blair, J.M.; Richmond, G.E.; Piddock, L.J. , Multidrug efflux pumps in Gram-negative bacteria and their role in antibiotic resistance. Future Microbiol. 2014, 9, 1165–1177. [Google Scholar] [CrossRef]
- Shorr, A.F. , Review of studies of the impact on Gram-negative bacterial resistance on outcomes in the intensive care unit. Crit. Care Med. 2009, 37, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Vasoo, S.; Barreto, J.N.; Tosh, P.K. , Emerging issues in gram-negative bacterial resistance: an update for the practicing clinician. Mayo Clin. Proc. 2015, 90, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Wheat, L.J. , Histoplasmosis: a review for clinicians from non-endemic areas. Mycoses 2006, 49, 274–282. [Google Scholar] [CrossRef]
- Sizemore, T.C. , Rheumatologic manifestations of histoplasmosis: a review. Rheumatol. Int. 2013, 33, 2963–2965. [Google Scholar] [CrossRef]
- DiCaudo, D.J. , Coccidioidomycosis: a review and update. J. Am. Acad. Dermatol. 2006, 55, 929–942, quiz 943-925. [Google Scholar] [CrossRef] [PubMed]
- Jude, C.M.; Nayak, N.B.; Patel, M.K.; Deshmukh, M.; Batra, P. , Pulmonary coccidioidomycosis: pictorial review of chest radiographic and CT findings. Radiographics 2014, 34, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Linder, K.A.; Kauffman, C.A.; Miceli, M.H. , Blastomycosis: A Review of Mycological and Clinical Aspects. J. Fungi (Basel) 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Bariola, J.R.; Perry, P.; Pappas, P.G.; Proia, L.; Shealey, W.; Wright, P.W.; Sizemore, J.M.; Robinson, M.; Bradsher, R.W., Jr. , Blastomycosis of the central nervous system: a multicenter review of diagnosis and treatment in the modern era. Clin. Infect. Dis. 2010, 50, 797–804. [Google Scholar] [CrossRef]
- Mengoli, C.; Cruciani, M.; Barnes, R.A.; Loeffler, J.; Donnelly, J.P. , Use of PCR for diagnosis of invasive aspergillosis: systematic review and meta-analysis. Lancet Infect. Dis. 2009, 9, 89–96. [Google Scholar] [CrossRef]
- Mahdavinia, M.; Grammer, L.C. , Management of allergic bronchopulmonary aspergillosis: a review and update. Ther. Adv. Respir. Dis. 2012, 6, 173–187. [Google Scholar] [CrossRef]
- Fisher, J.F. , Candida urinary tract infections--epidemiology, pathogenesis, diagnosis, and treatment: executive summary. Clin. Infect. Dis. 2011, 52 Suppl 6, S429–432. [Google Scholar] [CrossRef]
- Griffith, N.; Danziger, L. , Candida auris Urinary Tract Infections and Possible Treatment. Antibiotics (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Ben-Ami, R. , Treatment of Invasive Candidiasis: A Narrative Review. J. Fungi (Basel) 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- McCarty, T.P.; Pappas, P.G. , Invasive Candidiasis. Infect. Dis. Clin. North Am. 2016, 30, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Carmona, E.M.; Limper, A.H. , Update on the diagnosis and treatment of Pneumocystis pneumonia. Ther. Adv. Respir. Dis. 2011, 5, 41–59. [Google Scholar] [CrossRef]
- de Boer, M.G.; de Fijter, J.W.; Kroon, F.P. , Outbreaks and clustering of Pneumocystis pneumonia in kidney transplant recipients: a systematic review. Med. Mycol. 2011, 49, 673–680. [Google Scholar]
- Jeong, W.; Keighley, C.; Wolfe, R.; Lee, W.L.; Slavin, M.A.; Kong, D.C.M.; Chen, S.C. , The epidemiology and clinical manifestations of mucormycosis: a systematic review and meta-analysis of case reports. Clin. Microbiol. Infect. 2019, 25, 26–34. [Google Scholar] [CrossRef]
- Nair, A.G.; Dave, T.V. , Transcutaneous retrobulbar injection of amphotericin B in rhino-orbital-cerebral mucormycosis: a review. Orbit 2022, 41, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Setianingrum, F.; Rautemaa-Richardson, R.; Denning, D.W. , Pulmonary cryptococcosis: A review of pathobiology and clinical aspects. Med. Mycol. 2019, 57, 133–150. [Google Scholar] [CrossRef]
- Montoya, M.C.; Magwene, P.M.; Perfect, J.R. , Associations between Cryptococcus Genotypes, Phenotypes, and Clinical Parameters of Human Disease: A Review. J. Fungi (Basel) 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Bermas, A.; Geddes-McAlister, J. , Combatting the evolution of antifungal resistance in Cryptococcus neoformans. Mol. Microbiol. 2020, 114, 721–734. [Google Scholar] [CrossRef]
- Wall, G.; Lopez-Ribot, J.L. , Current Antimycotics, New Prospects, and Future Approaches to Antifungal Therapy. Antibiotics (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Buda De Cesare, G.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. , Antimicrobial Peptides: a New Frontier in Antifungal Therapy. mBio 2020, 11. [Google Scholar] [CrossRef]
- Zaet, A.; Dartevelle, P.; Daouad, F.; Ehlinger, C.; Quiles, F.; Francius, G.; Boehler, C.; Bergthold, C.; Frisch, B.; Prevost, G.; Lavalle, P.; Schneider, F.; Haikel, Y.; Metz-Boutigue, M.H.; Marban, C. , D-Cateslytin, a new antimicrobial peptide with therapeutic potential. Sci. Rep. 2017, 7, 15199. [Google Scholar] [CrossRef] [PubMed]
- Aslam, R.; Marban, C.; Corazzol, C.; Jehl, F.; Delalande, F.; Van Dorsselaer, A.; Prevost, G.; Haikel, Y.; Taddei, C.; Schneider, F.; Metz-Boutigue, M.H. , Cateslytin, a chromogranin A derived peptide is active against Staphylococcus aureus and resistant to degradation by its proteases. PLoS One 2013, 8, e68993. [Google Scholar] [CrossRef]
- Mancino, D.; Kharouf, N.; Scavello, F.; Helle, S.; Salloum-Yared, F.; Mutschler, A.; Mathieu, E.; Lavalle, P.; Metz-Boutigue, M.H.; Haikel, Y. , The Catestatin-Derived Peptides Are New Actors to Fight the Development of Oral Candidosis. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Salazar, N.; Gueimonde, M.; de Los Reyes-Gavilan, C.G. , Shaping the Metabolism of Intestinal Bacteroides Population through Diet to Improve Human Health. Front. Microbiol. 2017, 8, 376. [Google Scholar] [CrossRef]
- Rabbi, M.F.; Eissa, N.; Munyaka, P.M.; Kermarrec, L.; Elgazzar, O.; Khafipour, E.; Bernstein, C.N.; Ghia, J.E. , Reactivation of Intestinal Inflammation Is Suppressed by Catestatin in a Murine Model of Colitis via M1 Macrophages and Not the Gut Microbiota. Front. Immunol. 2017, 8, 985. [Google Scholar] [CrossRef] [PubMed]
- Rabbi, M.F.; Munyaka, P.M.; Eissa, N.; Metz-Boutigue, M.H.; Khafipour, E.; Ghia, J.E. , Human Catestatin Alters Gut Microbiota Composition in Mice. Front. Microbiol. 2016, 7, 2151. [Google Scholar] [CrossRef]
- Horvath, T.D.; Ihekweazu, F.D.; Haidacher, S.J.; Ruan, W.; Engevik, K.A.; Fultz, R.; Hoch, K.M.; Luna, R.A.; Oezguen, N.; Spinler, J.K.; Haag, A.M.; Versalovic, J.; Engevik, M.A. , Bacteroides ovatus colonization influences the abundance of intestinal short chain fatty acids and neurotransmitters. iScience 2022, 25, 104158. [Google Scholar] [CrossRef]
- Fernandez-Julia, P.J.; Munoz-Munoz, J.; van Sinderen, D. , A comprehensive review on the impact of beta-glucan metabolism by Bacteroides and Bifidobacterium species as members of the gut microbiota. Int. J. Biol. Macromol. 2021, 181, 877–889. [Google Scholar] [CrossRef]
- Bornet, E.; Westermann, A.J. , The ambivalent role of Bacteroides in enteric infections. Trends Microbiol. 2022, 30, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. , Colonic health: fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Friedel, D.; Levine, G.M. , Effect of short-chain fatty acids on colonic function and structure. JPEN J. Parenter. Enteral Nutr. 1992, 16, 1–4. [Google Scholar] [CrossRef]
- Cummings, J.H.; Macfarlane, G.T. , Role of intestinal bacteria in nutrient metabolism. JPEN J. Parenter. Enteral Nutr. 1997, 21, 357–365. [Google Scholar] [CrossRef]
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F., 3rd; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C. , Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 2015, 29, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Pan, J.; Liu, Y.; Wang, H.; Zhou, W.; Wang, X. , Intestinal Crosstalk between Microbiota and Serotonin and its Impact on Gut Motility. Curr. Pharm. Biotechnol. 2018, 19, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Grider, J.R.; Piland, B.E. , The peristaltic reflex induced by short-chain fatty acids is mediated by sequential release of 5-HT and neuronal CGRP but not BDNF. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G429–437. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, S.; Tatewaki, M.; Yamada, T.; Fujimiya, M.; Mantyh, C.; Voss, M.; Eubanks, S.; Harris, M.; Pappas, T.N.; Takahashi, T. , Short-chain fatty acids stimulate colonic transit via intraluminal 5-HT release in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R1269–1276. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. , Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef]
- Christiansen, C.B.; Gabe, M.B.N.; Svendsen, B.; Dragsted, L.O.; Rosenkilde, M.M.; Holst, J.J. , The impact of short-chain fatty acids on GLP-1 and PYY secretion from the isolated perfused rat colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G53–G65. [Google Scholar] [CrossRef]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. , The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. (Lond.) 2015, 39, 424–429. [Google Scholar] [CrossRef]
- Ma, Y.; Lee, E.; Yoshikawa, H.; Noda, T.; Miyamoto, J.; Kimura, I.; Hatano, R.; Miki, T. , Phloretin suppresses carbohydrate-induced GLP-1 secretion via inhibiting short chain fatty acid release from gut microbiome. Biochem. Biophys. Res. Commun. 2022, 621, 176–182. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. , Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Muller, M.; Hernandez, M.A.G.; Goossens, G.H.; Reijnders, D.; Holst, J.J.; Jocken, J.W.E.; van Eijk, H.; Canfora, E.E.; Blaak, E.E. , Circulating but not faecal short-chain fatty acids are related to insulin sensitivity, lipolysis and GLP-1 concentrations in humans. Sci. Rep. 2019, 9, 12515. [Google Scholar] [CrossRef]
- Hernandez, M.A.G.; Canfora, E.E.; Jocken, J.W.E.; Blaak, E.E. , The Short-Chain Fatty Acid Acetate in Body Weight Control and Insulin Sensitivity. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Wexler, H.M. , Bacteroides: the good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef]
- Yang, J.Y.; Lee, Y.S.; Kim, Y.; Lee, S.H.; Ryu, S.; Fukuda, S.; Hase, K.; Yang, C.S.; Lim, H.S.; Kim, M.S.; Kim, H.M.; Ahn, S.H.; Kwon, B.E.; Ko, H.J.; Kweon, M.N. , Gut commensal Bacteroides acidifaciens prevents obesity and improves insulin sensitivity in mice. Mucosal Immunol. 2017, 10, 104–116. [Google Scholar] [CrossRef]
- Yoshida, N.; Yamashita, T.; Osone, T.; Hosooka, T.; Shinohara, M.; Kitahama, S.; Sasaki, K.; Sasaki, D.; Yoneshiro, T.; Suzuki, T.; Emoto, T.; Saito, Y.; Ozawa, G.; Hirota, Y.; Kitaura, Y.; Shimomura, Y.; Okamatsu-Ogura, Y.; Saito, M.; Kondo, A.; Kajimura, S.; Inagaki, T.; Ogawa, W.; Yamada, T.; Hirata, K.I. , Bacteroides spp. promotes branched-chain amino acid catabolism in brown fat and inhibits obesity. iScience 2021, 24, 103342. [Google Scholar] [CrossRef]
- Nadal, I.; Santacruz, A.; Marcos, A.; Warnberg, J.; Garagorri, J.M.; Moreno, L.A.; Martin-Matillas, M.; Campoy, C.; Marti, A.; Moleres, A.; Delgado, M.; Veiga, O.L.; Garcia-Fuentes, M.; Redondo, C.G.; Sanz, Y. , Shifts in clostridia, bacteroides and immunoglobulin-coating fecal bacteria associated with weight loss in obese adolescents. Int. J. Obes. (Lond.) 2009, 33, 758–767. [Google Scholar] [CrossRef]
- Gauffin Cano, P.; Santacruz, A.; Moya, A.; Sanz, Y. , Bacteroides uniformis CECT 7771 ameliorates metabolic and immunological dysfunction in mice with high-fat-diet induced obesity. PLoS One 2012, 7, e41079. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Kasper, D.L. , The love-hate relationship between bacterial polysaccharides and the host immune system. Nat. Rev. Immunol. 2006, 6, 849–858. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. , Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 12204–12209. [Google Scholar] [CrossRef]
- Muntjewerff, E.M.; Tang, K.; Lutter, L.; Christoffersson, G.; Nicolasen, M.J.T.; Gao, H.; Katkar, G.D.; Das, S.; Ter Beest, M.; Ying, W.; Ghosh, P.; El Aidy, S.; Oldenburg, B.; van den Bogaart, G.; Mahata, S.K. , Chromogranin A regulates gut permeability via the antagonistic actions of its proteolytic peptides. Acta Physiol. (Oxf.) 2021, 232, e13655. [Google Scholar] [CrossRef]
- Gonzalez-Davila, P.; Schwalbe, M.; Danewalia, A.; Dalile, B.; Verbeke, K.; Mahata, S.K.; El Aidy, S. , Catestatin selects for colonization of antimicrobial-resistant gut bacterial communities. ISME J. 2022, 16, 1873–1882. [Google Scholar] [CrossRef]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. , Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef]
- Indiani, C.; Rizzardi, K.F.; Castelo, P.M.; Ferraz, L.F.C.; Darrieux, M.; Parisotto, T.M. , Childhood Obesity and Firmicutes/Bacteroidetes Ratio in the Gut Microbiota: A Systematic Review. Child. Obes. 2018, 14, 501–509. [Google Scholar] [CrossRef]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; Sineok, L.; Lushchak, O.; Vaiserman, A. , Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef] [PubMed]
- Corb Aron, R.A.; Abid, A.; Vesa, C.M.; Nechifor, A.C.; Behl, T.; Ghitea, T.C.; Munteanu, M.A.; Fratila, O.; Andronie-Cioara, F.L.; Toma, M.M.; Bungau, S. , Recognizing the Benefits of Pre-/Probiotics in Metabolic Syndrome and Type 2 Diabetes Mellitus Considering the Influence of Akkermansia muciniphila as a Key Gut Bacterium. Microorganisms 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. , Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Uchiyama, K.; Takagi, T. , A next-generation beneficial microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr. 2018, 63, 33–35. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; de Vos, W.M.; Cani, P.D. , Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. U. S. A. 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- Shin, N.R.; Lee, J.C.; Lee, H.Y.; Kim, M.S.; Whon, T.W.; Lee, M.S.; Bae, J.W. , An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef]
- Anhe, F.F.; Roy, D.; Pilon, G.; Dudonne, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; Marette, A. , A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef]
- Brahe, L.K.; Le Chatelier, E.; Prifti, E.; Pons, N.; Kennedy, S.; Hansen, T.; Pedersen, O.; Astrup, A.; Ehrlich, S.D.; Larsen, L.H. , Specific gut microbiota features and metabolic markers in postmenopausal women with obesity. Nutr. Diabetes 2015, 5, e159. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. , Human gut microbiota changes reveal the progression of glucose intolerance. PLoS One 2013, 8, e71108. [Google Scholar] [CrossRef]
- Yassour, M.; Lim, M.Y.; Yun, H.S.; Tickle, T.L.; Sung, J.; Song, Y.M.; Lee, K.; Franzosa, E.A.; Morgan, X.C.; Gevers, D.; Lander, E.S.; Xavier, R.J.; Birren, B.W.; Ko, G.; Huttenhower, C. , Sub-clinical detection of gut microbial biomarkers of obesity and type 2 diabetes. Genome Med. 2016, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; Zhang, W.; Weldon, R.; Auguste, K.; Yang, L.; Liu, X.; Chen, L.; Yang, X.; Zhu, B.; Cai, J. , Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; Xia, H.; Liu, Z.; Cui, B.; Liang, P.; Xi, L.; Jin, J.; Ying, X.; Wang, X.; Zhao, X.; Li, W.; Jia, H.; Lan, Z.; Li, F.; Wang, R.; Sun, Y.; Yang, M.; Shen, Y.; Jie, Z.; Li, J.; Chen, X.; Zhong, H.; Xie, H.; Zhang, Y.; Gu, W.; Deng, X.; Shen, B.; Xu, X.; Yang, H.; Xu, G.; Bi, Y.; Lai, S.; Wang, J.; Qi, L.; Madsen, L.; Wang, J.; Ning, G.; Kristiansen, K.; Wang, W. , Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; Consortium, M.I.-O.; Dumas, M.E.; Rizkalla, S.W.; Dore, J.; Cani, P.D.; Clement, K. , Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; de Barsy, M.; Loumaye, A.; Hermans, M.P.; Thissen, J.P.; de Vos, W.M.; Cani, P.D. , Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Eiseman, B.; Silen, W.; Bascom, G.S.; Kauvar, A.J. , Fecal enema as an adjunct in the treatment of pseudomembranous enterocolitis. Surgery 1958, 44, 854–859. [Google Scholar]
- Bhutiani, N.; Schucht, J.E.; Miller, K.R.; McClave, S.A. , Technical Aspects of Fecal Microbial Transplantation (FMT). Curr. Gastroenterol. Rep. 2018, 20, 30. [Google Scholar] [CrossRef]
- Brandt, L.J.; Aroniadis, O.C.; Mellow, M.; Kanatzar, A.; Kelly, C.; Park, T.; Stollman, N.; Rohlke, F.; Surawicz, C. , Long-term follow-up of colonoscopic fecal microbiota transplant for recurrent Clostridium difficile infection. Am. J. Gastroenterol. 2012, 107, 1079–1087. [Google Scholar] [CrossRef]
- Mattila, E.; Uusitalo-Seppala, R.; Wuorela, M.; Lehtola, L.; Nurmi, H.; Ristikankare, M.; Moilanen, V.; Salminen, K.; Seppala, M.; Mattila, P.S.; Anttila, V.J.; Arkkila, P. , Fecal transplantation, through colonoscopy, is effective therapy for recurrent Clostridium difficile infection. Gastroenterology 2012, 142, 490–496. [Google Scholar] [CrossRef]
- Friedman-Korn, T.; Livovsky, D.M.; Maharshak, N.; Aviv Cohen, N.; Paz, K.; Bar-Gil Shitrit, A.; Goldin, E.; Koslowsky, B. , Fecal Transplantation for Treatment of Clostridium Difficile Infection in Elderly and Debilitated Patients. Dig. Dis. Sci. 2018, 63, 198–203. [Google Scholar] [CrossRef]
- Cohen, N.A.; Maharshak, N. , Novel Indications for Fecal Microbial Transplantation: Update and Review of the Literature. Dig. Dis. Sci. 2017, 62, 1131–1145. [Google Scholar] [CrossRef]
- Borody, T.J.; George, L.; Andrews, P.; Brandl, S.; Noonan, S.; Cole, P.; Hyland, L.; Morgan, A.; Maysey, J.; Moore-Jones, D. , Bowel-flora alteration: a potential cure for inflammatory bowel disease and irritable bowel syndrome? Med. J. Aust. 1989, 150, 604. [Google Scholar] [CrossRef]
- Borody, T.J.; Cole, P.; Noonan, S.; Morgan, A.; Lenne, J.; Hyland, L.; Brandl, S.; Borody, E.G.; George, L.L. , Recurrence of duodenal ulcer and Campylobacter pylori infection after eradication. Med. J. Aust. 1989, 151, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Winslet, M.C.; Andrews, H.; Allan, R.N.; Keighley, M.R. , Fecal diversion in the management of Crohn’s disease of the colon. Dis. Colon Rectum 1993, 36, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Grehan, M.J.; Borody, T.J.; Leis, S.M.; Campbell, J.; Mitchell, H.; Wettstein, A. , Durable alteration of the colonic microbiota by the administration of donor fecal flora. J. Clin. Gastroenterol. 2010, 44, 551–561. [Google Scholar] [CrossRef]
- Bakken, J.S.; Borody, T.; Brandt, L.J.; Brill, J.V.; Demarco, D.C.; Franzos, M.A.; Kelly, C.; Khoruts, A.; Louie, T.; Martinelli, L.P.; Moore, T.A.; Russell, G.; Surawicz, C.; Fecal Microbiota Transplantation, W. , Treating Clostridium difficile infection with fecal microbiota transplantation. Clin. Gastroenterol. Hepatol. 2011, 9, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Davila, P.; Schwalbe, M.; Danewalia, A.; Wardenaar, R.; Dalile, B.; Verbeke, K.; Mahata, S.K.; El Aidy, S. , Gut microbiota transplantation drives the adoptive transfer of colonic genotype-phenotype characteristics between mice lacking catestatin and their wild type counterparts. Gut Microbes 2022, 14, 2081476. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, M.; Everard, A.; Gomez-Valades, A.G.; Matamoros, S.; Ramirez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. , Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci. Rep. 2015, 5, 16643. [Google Scholar] [CrossRef]
- Zhou, Q.; Pang, G.; Zhang, Z.; Yuan, H.; Chen, C.; Zhang, N.; Yang, Z.; Sun, L. , Association Between Gut Akkermansia and Metabolic Syndrome is Dose-Dependent and Affected by Microbial Interactions: A Cross-Sectional Study. Diabetes Metab. Syndr. Obes. 2021, 14, 2177–2188. [Google Scholar] [CrossRef]
- Earley, H.; Lennon, G.; Balfe, A.; Coffey, J.C.; Winter, D.C.; O’Connell, P.R. , The abundance of Akkermansia muciniphila and its relationship with sulphated colonic mucins in health and ulcerative colitis. Sci. Rep. 2019, 9, 15683. [Google Scholar] [CrossRef] [PubMed]
- Glassner, K.L.; Abraham, B.P.; Quigley, E.M.M. , The microbiome and inflammatory bowel disease. J. Allergy Clin. Immunol. 2020, 145, 16–27. [Google Scholar] [CrossRef]
- Bui, T.P.; Ritari, J.; Boeren, S.; de Waard, P.; Plugge, C.M.; de Vos, W.M. , Production of butyrate from lysine and the Amadori product fructoselysine by a human gut commensal. Nat. Commun. 2015, 6, 10062. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. , The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Bolognini, D.; Tobin, A.B.; Milligan, G.; Moss, C.E. , The Pharmacology and Function of Receptors for Short-Chain Fatty Acids. Mol. Pharmacol. 2016, 89, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.J.; Preston, T. , Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- De Preter, V.; Geboes, K.P.; Bulteel, V.; Vandermeulen, G.; Suenaert, P.; Rutgeerts, P.; Verbeke, K. , Kinetics of butyrate metabolism in the normal colon and in ulcerative colitis: the effects of substrate concentration and carnitine on the beta-oxidation pathway. Aliment. Pharmacol. Ther. 2011, 34, 526–532. [Google Scholar] [CrossRef]
- Huda-Faujan, N.; Abdulamir, A.S.; Fatimah, A.B.; Anas, O.M.; Shuhaimi, M.; Yazid, A.M.; Loong, Y.Y. , The impact of the level of the intestinal short chain Fatty acids in inflammatory bowel disease patients versus healthy subjects. Open Biochem. J. 2010, 4, 53–58. [Google Scholar] [CrossRef]
- Wang, H.B.; Wang, P.Y.; Wang, X.; Wan, Y.L.; Liu, Y.C. , Butyrate enhances intestinal epithelial barrier function via up-regulation of tight junction protein Claudin-1 transcription. Dig. Dis. Sci. 2012, 57, 3126–3135. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; Rudensky, A.Y. , Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; Takahashi, M.; Fukuda, N.N.; Murakami, S.; Miyauchi, E.; Hino, S.; Atarashi, K.; Onawa, S.; Fujimura, Y.; Lockett, T.; Clarke, J.M.; Topping, D.L.; Tomita, M.; Hori, S.; Ohara, O.; Morita, T.; Koseki, H.; Kikuchi, J.; Honda, K.; Hase, K.; Ohno, H. , Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Chen, L.; Sun, M.; Wu, W.; Yang, W.; Huang, X.; Xiao, Y.; Ma, C.; Xu, L.; Yao, S.; Liu, Z.; Cong, Y. , Microbiota Metabolite Butyrate Differentially Regulates Th1 and Th17 Cells’ Differentiation and Function in Induction of Colitis. Inflamm. Bowel. Dis. 2019, 25, 1450–1461. [Google Scholar] [CrossRef]
- Fregeau, C.J.; Helgason, C.D.; Bleackley, R.C. , Two cytotoxic cell proteinase genes are differentially sensitive to sodium butyrate. Nucleic Acids Res. 1992, 20, 3113–3119. [Google Scholar] [CrossRef]
- Tsuda, H.; Ochiai, K.; Suzuki, N.; Otsuka, K. , Butyrate, a bacterial metabolite, induces apoptosis and autophagic cell death in gingival epithelial cells. J. Periodontal Res. 2010, 45, 626–634. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Copeland, B.R.; Mustoe, A.M.; Goldstein, D.B. , Natural Selection Shapes Codon Usage in the Human Genome. Am. J. Hum. Genet. 2020, 107, 83–95. [Google Scholar] [CrossRef]
- McGarrah, R.W.; White, P.J. , Branched-chain amino acids in cardiovascular disease. Nat. Rev. Cardiol. 2023, 20, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Rees, J.S.; Castellano, S.; Andres, A.M. , The Genomics of Human Local Adaptation. Trends Genet. 2020, 36, 415–428. [Google Scholar] [CrossRef]
- Brunner, J.S.; Finley, L.W.S. , Metabolic determinants of tumour initiation. Nat. Rev. Endocrinol. 2023, 19, 134–150. [Google Scholar] [CrossRef]
Figure 1.
Homology of CST sequence in 53 mammalian species belonging to 8 orders. CST sequences were aligned using the MUSCLE method provided by SnapGene software from the following mammalian species: human (Homo sapiens: NM_001275), bonobo (Pan paniscus: XP_008956465.1), chimpanzee (Pan troglodytes: PNI97600.1), western low-land gorilla (Gorilla gorilla gorilla: XM_019009788.2), Sumatran orangutan (Pongo abelii: XM_002825045.3), silvery gibbon (Hylobates moloch: XP_031990963.1), black snub-nosed monkey (Rhinopithecus bieti: XM_017857899.1), crab-eating macaque (Macaca fascicularis: XP_045252830.1), golden snub-nosed monkey (Rhinopithecus roxellana: XM_010384506.1), green monkey (Chlorocebus sabaeus: XM_007987644.2), night monkey (Aotus nancymaae: XM_012455409.1), old world monkey (Colobus angolensis palliates: XM_011949380.1), olive baboon (Papio Anubis: XM_031667888.1), pig-tailed macaque (Macaca nemestrina: XM_011717182.1), red colobus (Piliocolobus tephrosceles: XM_023205512.3), Sooty mangabey (Cercocebus atys: XM_012083744.1), rhesus monkey (Macaca mulatta: NM_001278450.1), tufted capuchin (Sapajus apella: XM_032287580.1), black rat (Rattus rattus: XM_032908276.1), brown rat (Rattus norvegicus: XM_032908276.1), golden hamster (Mesocricetus auratus: XM_005068386.4), grasshopper mouse (Onychomys torridus: XM_036206345.1), house mouse (Mus musculus: NM_007693.2), multimammate mouse (Mastomys coucha: XM_031357132.1), Ryuku mouse (Mus caroli: XM_021179357.1), Shrew mouse (Mus pahari: XM_021202342.2), European rabbit (Oryctolagus cuniculus: XM_051826432.1), alpaca (Vicugna pacos: XP_031534667.1), Arabian camel (Camelus dromedarius: XM_031454226.1), cattle (Bos taurus: NM_181005.2), goat (Capra hircus: XM_018066172.1), pig (Sus scrofa: NP_001157477.2), sheep (Ovis aries: XP_004018008.3), donkey (Equus asinus: XP_014687627.1), horse (Equus caballus: NP_001075283.2), southern white rhinoceros (Ceratotherium simum: XP_004434274.1), California sea lion (Zalophus californianus: XP_027424506.2), cat (Felis catus: XP_023111743.1), cheetah (Acinonyx jubatus: XP_026922275.1), dog (Canis lupus: XP_038528993.1), giant panda (Ailuropoda melanoleuca: XP_019660005.1), grizzly bear (Ursus arctos horribilis: XP_048075839.1), harbor seal (Phoca vitulina: XP_032261715.1), leopard (Panthera pardus: XP_019317643.2), mongooses (Suricata suricatta: XP_029807749.1), monk seal (Neomonachus schauinslandi: XP_021535325.1), Northern fur seal (Callorhinus ursinus: XP_025726236.1), walrus (Odobenus rosmarus divergens: XP_004394547.1), weddell-seal (Leptonychotes weddellii: XP_030873380.1), dolphin (Lipotes vexillifer: XP_007454783.1), killer whale (Orcinus orca: XP_004262400.1), sperm whale (Physeter catodon: XP_023986851.1), and platypus (Ornithorhynchus anatinua: XP_039767777.1). Yellow shows an amino acid match between species.
Figure 1.
Homology of CST sequence in 53 mammalian species belonging to 8 orders. CST sequences were aligned using the MUSCLE method provided by SnapGene software from the following mammalian species: human (Homo sapiens: NM_001275), bonobo (Pan paniscus: XP_008956465.1), chimpanzee (Pan troglodytes: PNI97600.1), western low-land gorilla (Gorilla gorilla gorilla: XM_019009788.2), Sumatran orangutan (Pongo abelii: XM_002825045.3), silvery gibbon (Hylobates moloch: XP_031990963.1), black snub-nosed monkey (Rhinopithecus bieti: XM_017857899.1), crab-eating macaque (Macaca fascicularis: XP_045252830.1), golden snub-nosed monkey (Rhinopithecus roxellana: XM_010384506.1), green monkey (Chlorocebus sabaeus: XM_007987644.2), night monkey (Aotus nancymaae: XM_012455409.1), old world monkey (Colobus angolensis palliates: XM_011949380.1), olive baboon (Papio Anubis: XM_031667888.1), pig-tailed macaque (Macaca nemestrina: XM_011717182.1), red colobus (Piliocolobus tephrosceles: XM_023205512.3), Sooty mangabey (Cercocebus atys: XM_012083744.1), rhesus monkey (Macaca mulatta: NM_001278450.1), tufted capuchin (Sapajus apella: XM_032287580.1), black rat (Rattus rattus: XM_032908276.1), brown rat (Rattus norvegicus: XM_032908276.1), golden hamster (Mesocricetus auratus: XM_005068386.4), grasshopper mouse (Onychomys torridus: XM_036206345.1), house mouse (Mus musculus: NM_007693.2), multimammate mouse (Mastomys coucha: XM_031357132.1), Ryuku mouse (Mus caroli: XM_021179357.1), Shrew mouse (Mus pahari: XM_021202342.2), European rabbit (Oryctolagus cuniculus: XM_051826432.1), alpaca (Vicugna pacos: XP_031534667.1), Arabian camel (Camelus dromedarius: XM_031454226.1), cattle (Bos taurus: NM_181005.2), goat (Capra hircus: XM_018066172.1), pig (Sus scrofa: NP_001157477.2), sheep (Ovis aries: XP_004018008.3), donkey (Equus asinus: XP_014687627.1), horse (Equus caballus: NP_001075283.2), southern white rhinoceros (Ceratotherium simum: XP_004434274.1), California sea lion (Zalophus californianus: XP_027424506.2), cat (Felis catus: XP_023111743.1), cheetah (Acinonyx jubatus: XP_026922275.1), dog (Canis lupus: XP_038528993.1), giant panda (Ailuropoda melanoleuca: XP_019660005.1), grizzly bear (Ursus arctos horribilis: XP_048075839.1), harbor seal (Phoca vitulina: XP_032261715.1), leopard (Panthera pardus: XP_019317643.2), mongooses (Suricata suricatta: XP_029807749.1), monk seal (Neomonachus schauinslandi: XP_021535325.1), Northern fur seal (Callorhinus ursinus: XP_025726236.1), walrus (Odobenus rosmarus divergens: XP_004394547.1), weddell-seal (Leptonychotes weddellii: XP_030873380.1), dolphin (Lipotes vexillifer: XP_007454783.1), killer whale (Orcinus orca: XP_004262400.1), sperm whale (Physeter catodon: XP_023986851.1), and platypus (Ornithorhynchus anatinua: XP_039767777.1). Yellow shows an amino acid match between species.
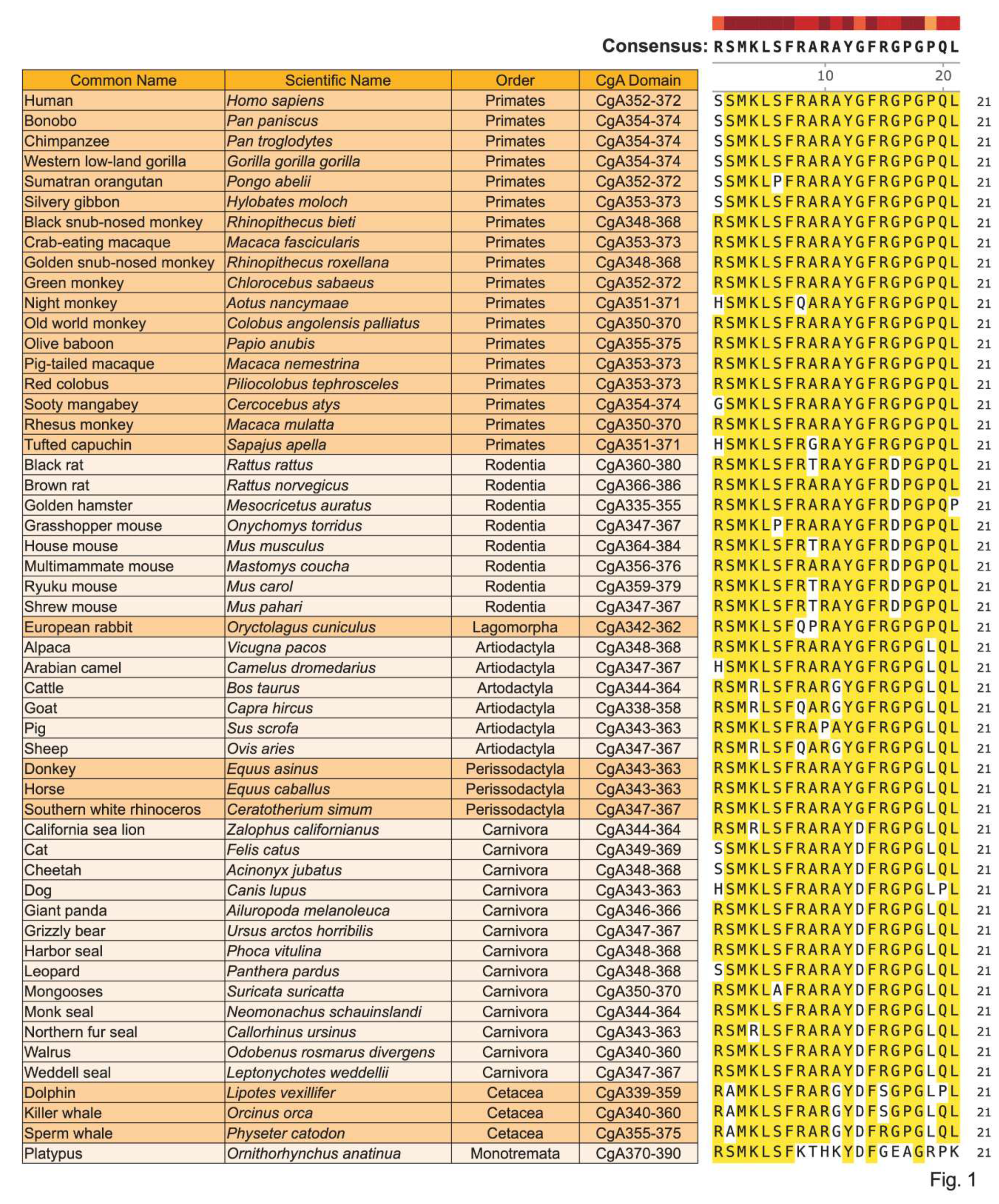
Figure 2.
Homology of the individual amino acid in catestatin sequence in 53 mammalian species belonging to 7 orders.
Figure 2.
Homology of the individual amino acid in catestatin sequence in 53 mammalian species belonging to 7 orders.
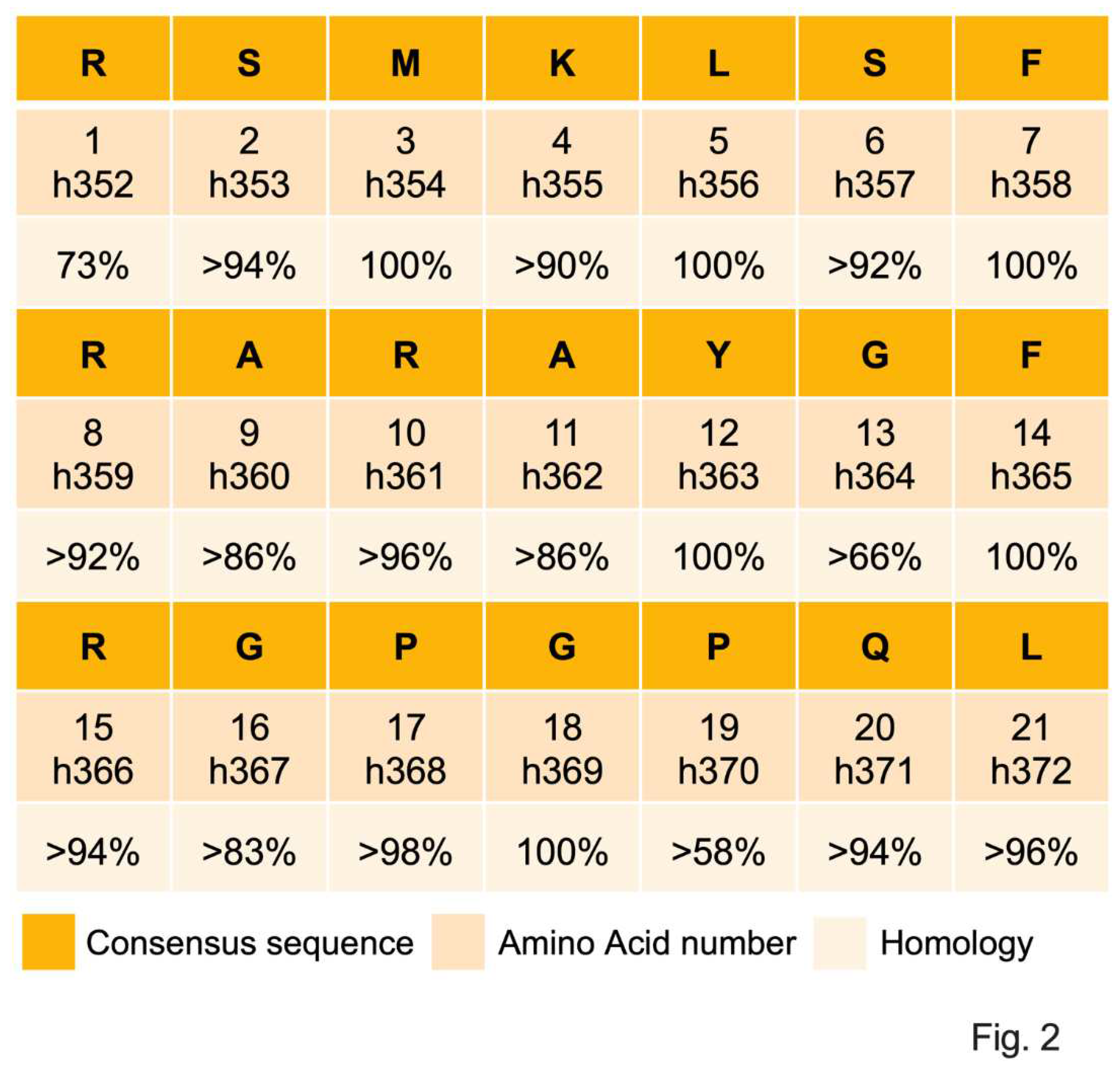
Figure 3.
Effects of wild-type (WT)-CST and natural human variants of CST (Gly364Ser and Pro370Leu) on the growth of Gram-positive and Gram-negative bacteria showing minimal inhibitory concentration (MIC) and lethal concentration (MIC100) of CST.
Figure 3.
Effects of wild-type (WT)-CST and natural human variants of CST (Gly364Ser and Pro370Leu) on the growth of Gram-positive and Gram-negative bacteria showing minimal inhibitory concentration (MIC) and lethal concentration (MIC100) of CST.
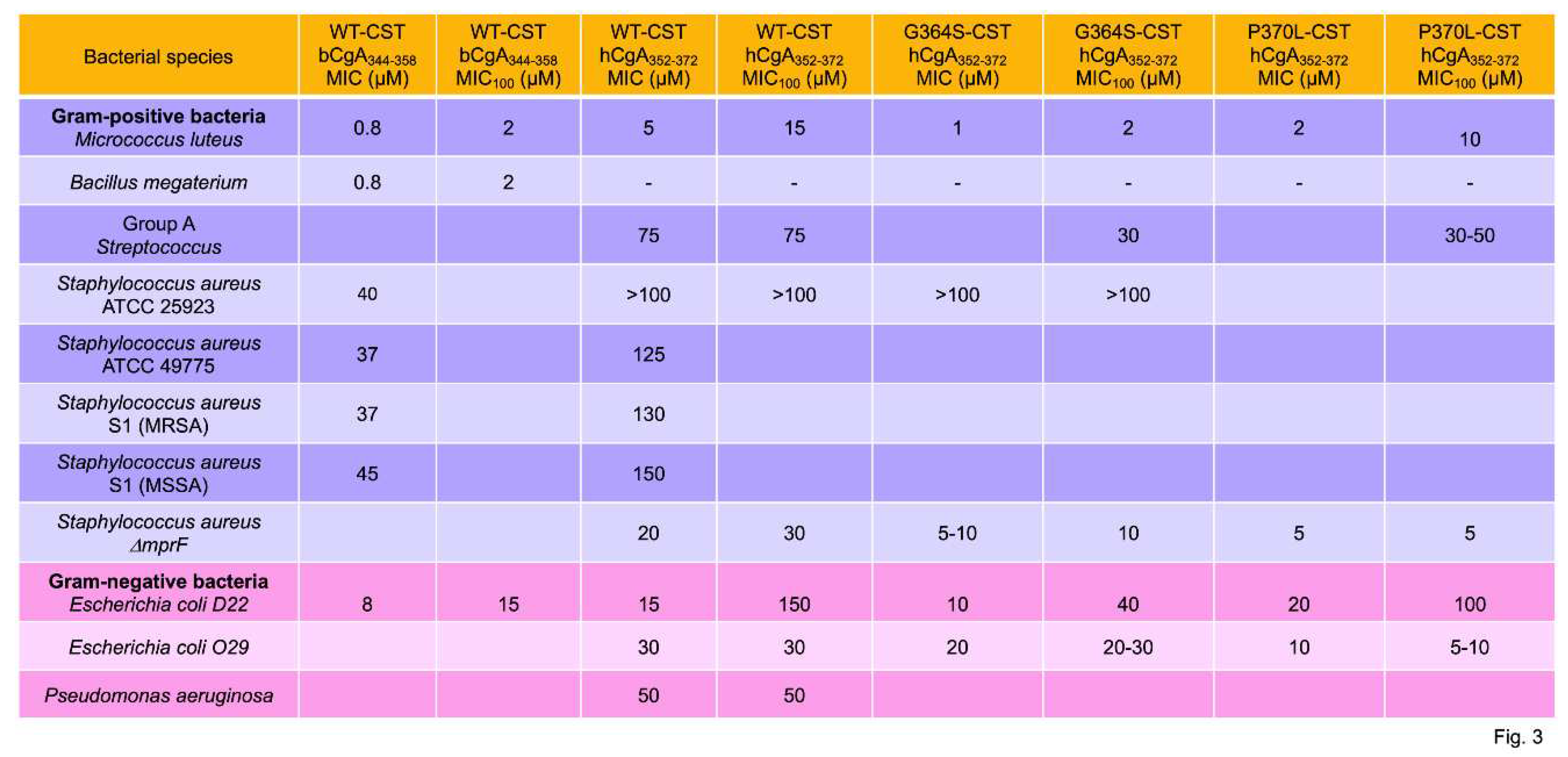
Figure 4.
Effects of WT-CST and natural human variants of CST (Gly364Ser and Pro370Leu) on the growth of fungal species showing MIC and MIC100 of CST.
Figure 4.
Effects of WT-CST and natural human variants of CST (Gly364Ser and Pro370Leu) on the growth of fungal species showing MIC and MIC100 of CST.
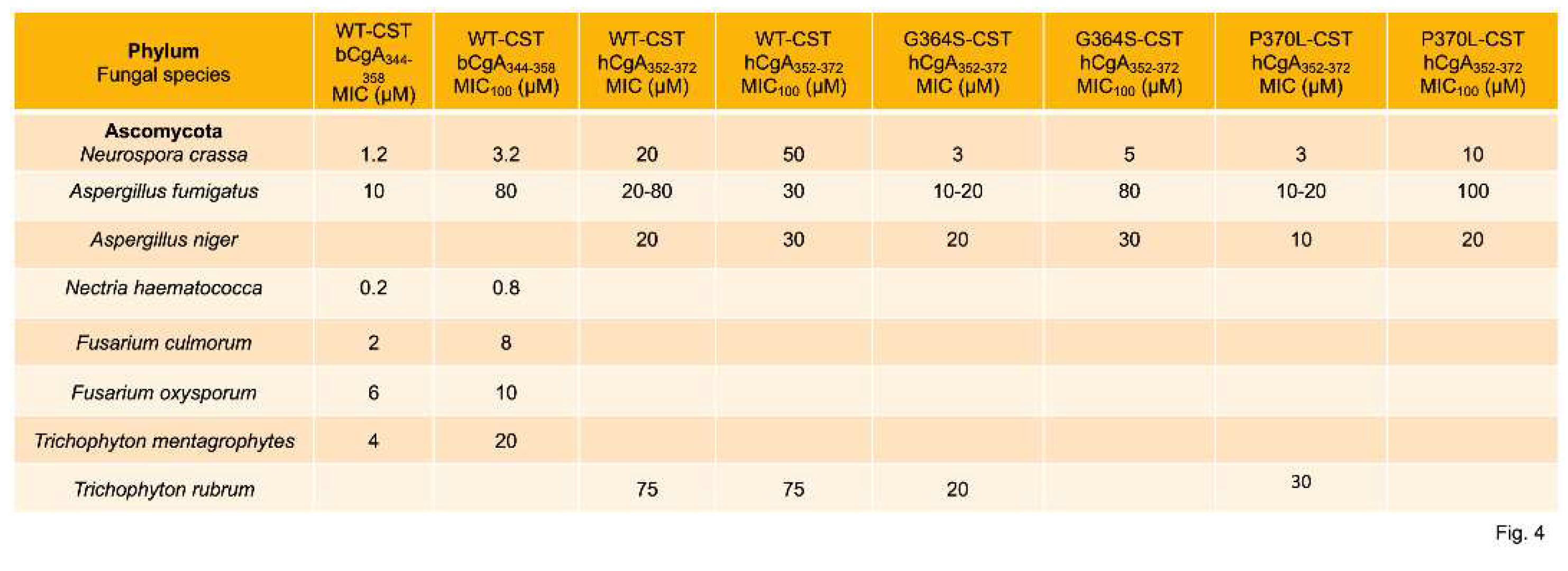
Figure 5.
Effects of WT-CST and natural human variants of CST (Gly364Ser and Pro370Leu) on the growth of yeast species showing MIC and MIC100 of CST.
Figure 5.
Effects of WT-CST and natural human variants of CST (Gly364Ser and Pro370Leu) on the growth of yeast species showing MIC and MIC100 of CST.
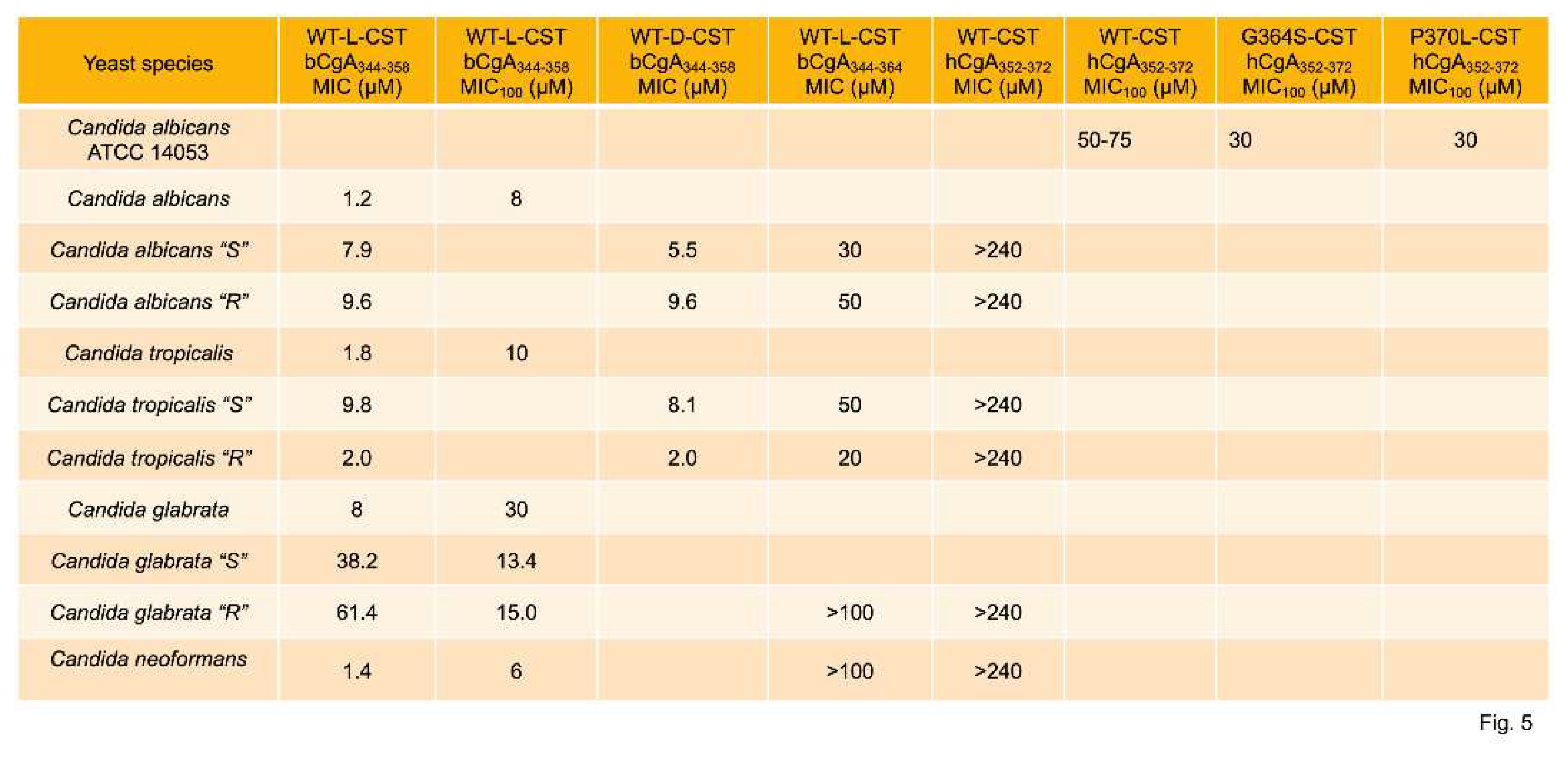
Figure 6.
Homology between cell permeable peptide penetratin and CST in 7 mammalian orders.
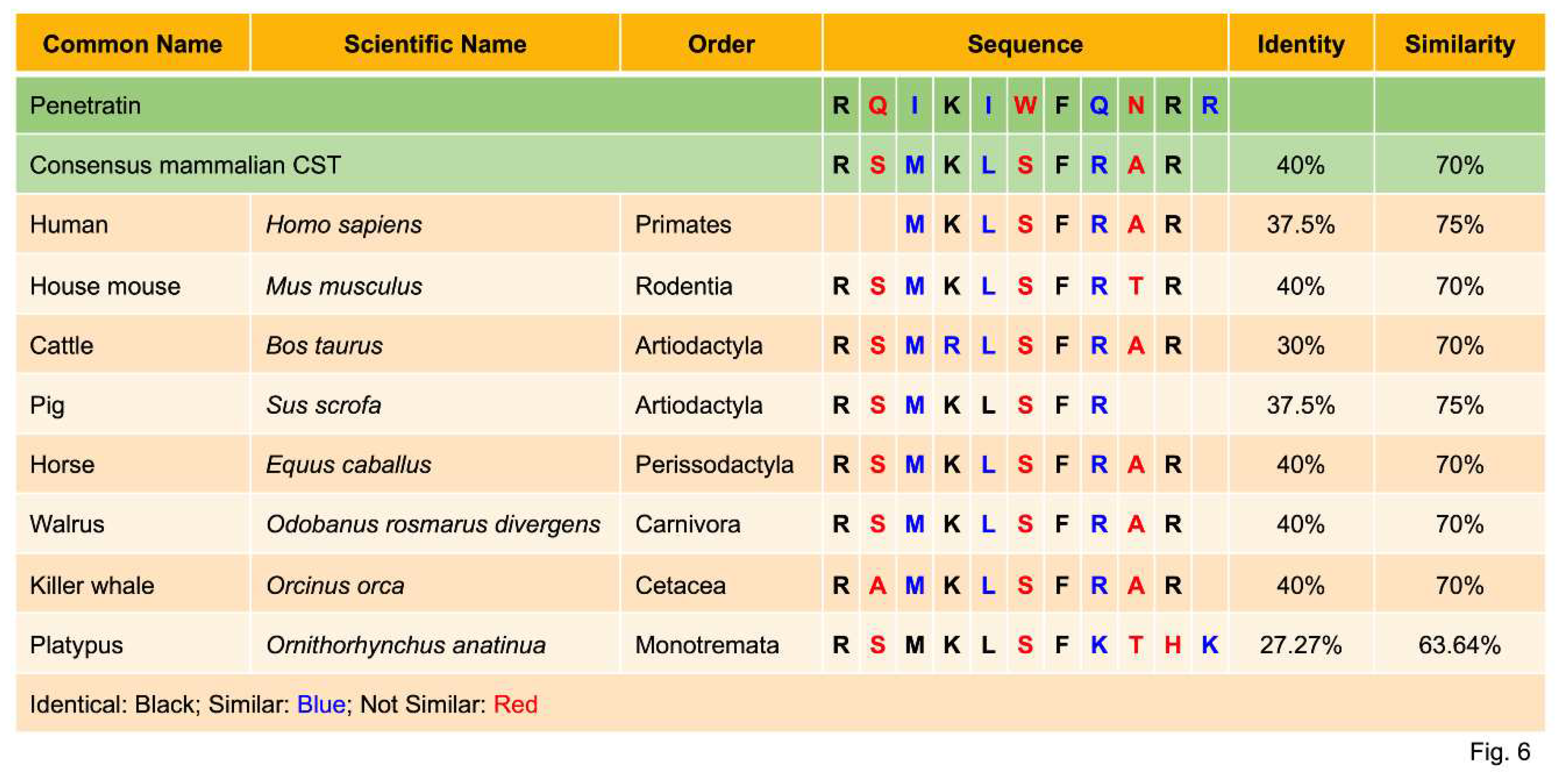
Figure 7.
Abundance of bacterial species in mucosal and fecal samples in WT and CST-KO mice in presence or absence of CST as well as after fecal microbial transplant. Green arrows indicate CST effects; red arrows indicate FMT effects.
Figure 7.
Abundance of bacterial species in mucosal and fecal samples in WT and CST-KO mice in presence or absence of CST as well as after fecal microbial transplant. Green arrows indicate CST effects; red arrows indicate FMT effects.
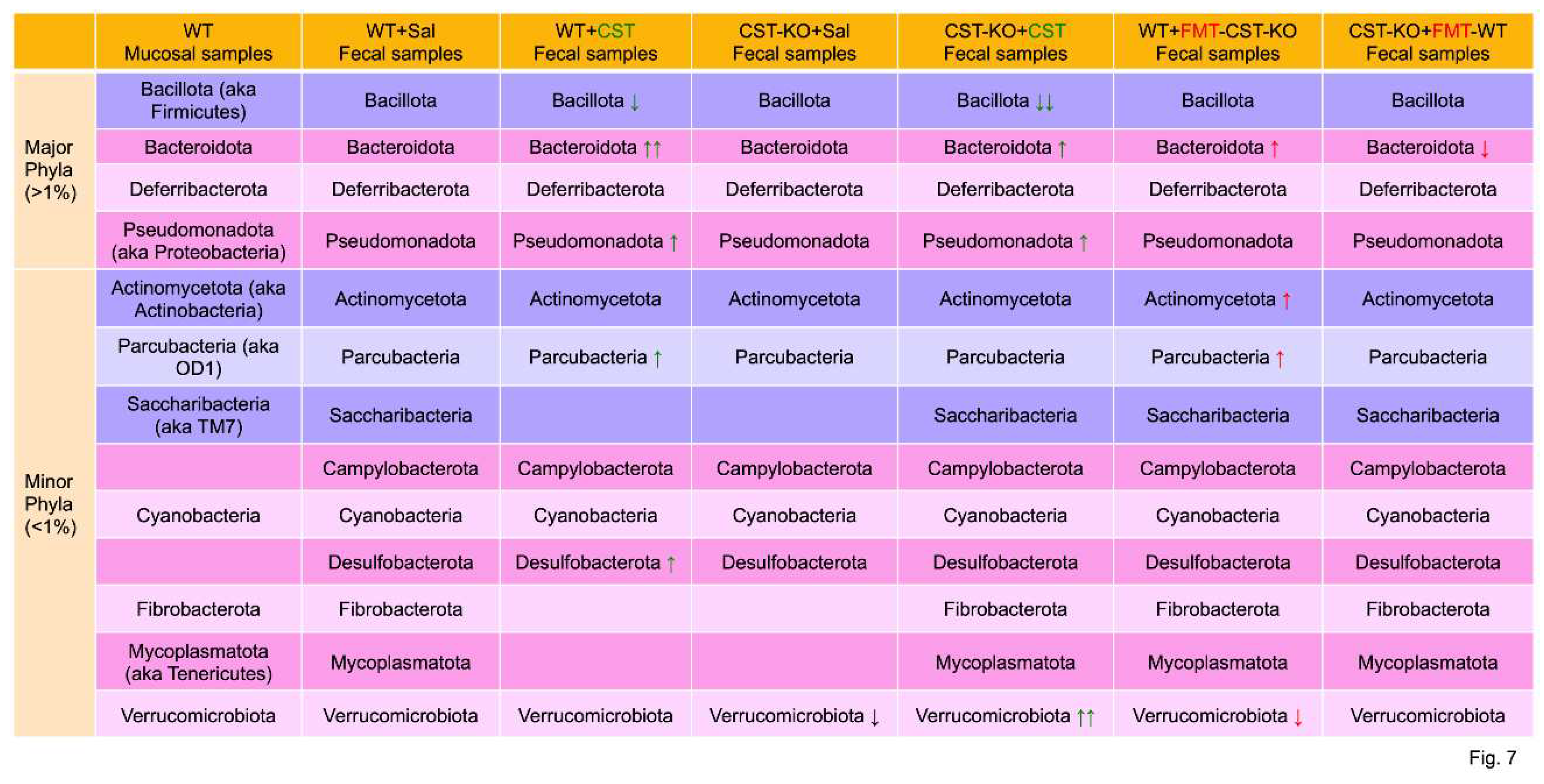
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



