
Submitted:
31 March 2023
Posted:
06 April 2023
You are already at the latest version
Abstract
A key component of efforts to identify the biological and drug-specific aspects that contribute to therapeutic failure or unexpected exposure-associated toxicity is the study of drug-intestinal barrier interactions. While methods supporting such assessments are widely described for human therapeutics, there is relatively little information available for similar evaluation in support of veterinary pharmaceuticals. There is, therefore, a critical need to develop novel approaches for evaluating drug-gut interactions in veterinary medicine. Three-dimensional (3D) organoids can address these difficulties in a reasonably affordable system that circumvents the need for more invasive in vivo assays in live animals. However, a first step in the development of such systems is understanding organoid interactions in a 2D monolayer. Given the importance of orally administered medications for meeting the therapeutic need of companion animals, we demonstrate growth conditions under which canine colonoid-derived intestinal epithelial cells survive, mature, and differentiate into confluent cell systems with high monolayer integrity. We further examine the applicability of this canine colonoid-derived 2D model to assess the permeability of three structurally diverse, passively absorbed β-blockers (e.g., propranolol, metoprolol, and atenolol). Both absorptive and secretive apparent permeability (Papp) of these drugs at two different pH conditions were evaluated in canine colonoid-derived monolayers and were compared with that of Caco-2 cells. This proof-of-concept study provides promising preliminary results with regard to the utility of canine-derived organoid monolayers for species-specific assessments of therapeutic drug passive permeability.
Keywords:
canine
; 3D organoid
; permeability
; Caco-2
; colon
1. Introduction
Oral dosing is the most common route of administration for human therapeutics, accounting for over 90% of all drug formulations[1]. For systemically active pharmaceuticals, orally delivered dosage forms must be absorbed by the gastrointestinal (GI) tract to exert their clinical effects at the target site. Disintegration, dissolution, and permeation are three key processes involved in the GI absorption of solid dosage forms[2,3]. Solubilized molecules must pass through the single layer of enterocytes lining the gut lumen to be absorbed into the portal circulation. Thus, the intestinal epithelium serves both as an absorptive surface and an absorption barrier to the systemic entry of therapeutic drugs. Barrier functions of the enterocytes include metabolizing enzymes within the cells, membrane efflux transporters, and tight junctions (TJs); the latter consisting of extracellular folds of the transmembrane proteins and multiprotein junctional complexes that form pore-like structures constraining movement across the intercellular spaces[4] (Figure 1).
Movement of a drug (rate of flux) in a diffusion cell system from a donor compartment to a receiver compartment can be used to obtain a mathematical estimate of the apparent permeability (Papp) (Figure 2). The Papp estimate, which is a function of both passive and active transport mechanisms, can be derived using a variety of in vitro systems[5]. Excluding the possibility of drug metabolism within the enterocyte, drug absorption can be evaluated within monolayer cells systems such as those associated with the parallel artificial membrane permeability assay (PAMPA), which typically reflects passive permeability only [6], the human colon adenocarcinoma (Caco-2) cell line [7,8,9,10], and more recently intestinal organoids [11].
Mathematically, the in vitro Papp can be used to estimate in vivo effective permeability (Peff) by incorporating information on the fraction of drug molecules existing in their neutral form at a given pH, the surface area for absorption, the fraction of unbound drug in the unstirred boundary layer and the drug permeability across the unstirred boundary layer[12]. Determination of a drug Papp early within the drug product development process is critical for predicting possible challenges with oral bioavailability and setting up corresponding formulation strategies that can circumvent these issues. Cell culture systems, e.g., Caco-2, are frequently employed to determine oral drug permeability and are currently considered the gold standard for estimating in vitro intestinal permeability and oral absorption of candidate therapeutics[13]. The Caco-2 cell line forms monolayers that both morphologically and functionally resemble the small intestinal (absorptive) enterocytes[14].
One of the reasons for the extensive use of the Caco-2 assay lies in the versatility of the cell line, allowing for the study of passive diffusion processes, active drug transport, and pre-systemic drug metabolism[15]. Caco-2 cells spontaneously differentiate into mature small intestinal enterocytes that express morphological (polarized columnar epithelium) and functional features such as intercellular TJs, efflux [e.g., P-glycoprotein (P-gp) and member of the Multidrug Resistance-Associated Protein (MRP) family] and influx [e.g., the Organic Anion Transporting Protein (OATP)] transporters, and the presence of enzymes such as the cytochrome P450 (CYP) isoenzymes[15,16,17]. While Caco-2 cells are widely used in the pharmaceutical industry and are accepted by regulatory agencies to predict human intestinal drug permeability, caution should be exercised when extrapolating data from these in vitro models to in vivo physiology[18,19]. More specifically, in addition to issues outlined above, there have been discrepancies in the expression of influx transporters in Caco-2 cells between studies conducted in different laboratories[20,21,22,23,24]. Additionally, one disadvantage of Caco-2 cells is that they only represent one cell type from the epithelial layer of the small intestine[25]. Specifically, the absence of goblet cells, which are responsible for mucus production, makes it impossible to evaluate mucus-drug interactions [26]. The presence of mucus from goblet cells represents a physiological barrier that drugs cross to enter the intestinal enterocytes. It should also be noted that Caco-2 cultures lack expression of numerous key nuclear receptors normally found in the intestine, including the pregnane X receptor (PXR), steroid X receptor (SXR), and constitutive androstane receptor (CAR)[27]. As a result, Caco-2 cultures are unable to simulate the induction of drug transporters and enzymes by certain drugs (e.g., rifampin) that interact with these receptors[28].
Research in biomedical science, disease modeling, and personalized medicine has advanced since Sato et al. (2009) first reported the in vitro culture of intestinal organoids[19,29,30,31,32,33,34,35,36,37,38,39]. Three-dimensional (3D) organoids are derived from leucine-rich repeat containing G protein-coupled receptor 5 (Lgr5)-positive adult stem cells [35,38,39]. Their 3D structure enables organoids to morphologically, physiologically, and structurally mimic endogenous epithelia. Therefore, these systems provide an opportunity to evaluate the transport and intestinal metabolism of administered orally therapeutic drugs [19]. Although human adult-stem cell derived intestinal organoids are now used as intestinal permeability models[40,41], canine organoids present a promising alternative to human organoids due to ethical constraints related to research on human stem cells[42] and the lack of availability of large human organoid bioarchives.
Canine-specific permeability tools are also needed for assessing the unique absorption challenges associated with the canine GI tract. In this regard, although Caco-2 cells are commonly used to evaluate human drug permeability, their predictive performances in modeling Papp in dogs remain to be demonstrated. The use of canine intestinal organoids in permeability studies may be more accurate in predicting canine, not human, intestinal permeability and metabolism of small pharmacological molecules as compared to that derived using Caco-2 or Madin-Darby Canine Kidney (MDCK) cells, with a potential to avoid the ethical and financial constraints associated with the use of live animal models. At last, the possibility of re-using organoids preserved in a biorepository further supports the 3R “Replacement, Reduction, and Refinement” initiative.
The potential utility of organoid systems is gaining recognition within pharmaceutical sciences, with examples being applied to exploring drug screening, cancer therapies, gene therapy, and a host of other kinds of diseases [43,44,45]. Examples of cultured organoid systems include human kidney tubuloids[46], dog prostate cancer organoids[47], and human bladder cancer organoids[29,48,49]. Although still early in its development, with many questions and challenges yet to be addressed, there are recent efforts to improve the ability of the 3D organoid to recapitulate in vivo organ physiology through the development of the organ-on-a-chip technology[50]. Microfluidic organ-on-chips [50] provide a constant flow of media through the cell culture, while various other physiologically important parameters, such as oxygen saturation and shear stress, can be manipulated and mimicked more closely for physiological representation. However, this technology is still relatively new and bears a number of limitations, such as significant batch-to-batch variability causing a wide range of results for the same parameters [51,52]. Also, due to the complexity of the microfluidic system, experiments involving organs-on-a-chip require additional technical skill sets, thereby limiting its broader use in preclinical research [51,52].
Whether or not influx and efflux transporters are involved in the transmembrane movements of a drug candidate, it is essential that a model system does not introduce a bias by failure to adequately control passive transport. Accordingly, the first step in characterizing the function of cell monolayers is to assess their passive permeability. Furthermore, predictive performances of the novel canine colonoid model need to be compared with those of the current gold standard Caco-2 cell monolayer to identify possible interspecies differences in drug passive permeability. To meet this objective, the study presented herein used model drugs with well-characterized transcellular absorption (propranolol and metoprolol) and paracellular diffusion (atenolol) attributes. Since this was a proof-of-concept study, this choice was motivated based on the known in vivo and in vitro permeability of these drugs. Metoprolol and propranolol are human BCS Class I (high solubility, high permeability) drugs [53], while atenolol is a BCS Class III drug (low permeability, high solubility) [54].
In this study, we thoroughly validated the integrity of canine monolayers and their functionality by transepithelial electrical resistance (TEER) measurement and FITC-Dextran assay. We also report expression data for key intestinal epithelial cell markers, tight and adherens junction proteins, transporters and cytochrome P450 (CYP) enzymes. At last, both the absorptive (apical to basolateral, AP→BL) and secretive (basolateral to apical, BL→AP) Papp estimates of these model drugs were evaluated in Caco-2 cell and canine colonoid-derived monolayers under various pH conditions intended to reflect the in vivo intestinal physiology. Our working hypothesis was that the passive permeability of the two transcellularly absorbed molecules would be similar across the two cell systems, but that differences may be observed with atenolol [55]. Noteworthily, this first proof-of concept study focused on the number of conditions being explored rather than replicates per condition, with the option of expanding the number of replicates for studying a particular condition as needed.
2. Materials and Methods
2.1. Materials
Caco-2 cells (passage #47) and FITC-dextran were purchased from Millipore Sigma (Sigma-Aldrich Inc, St. Louis, MO). Propranolol hydrochloride, metoprolol tartrate, and atenolol were obtained from Tocris Bioscience (Bio-Techne Corporation, Minneapolis, MN). Corning® Transwell™ 24-well plates with permeable polyester membrane inserts (6.5mm diameter and 0.4μm pore size), GIBCO™ TrypLE™ express enzyme (1X, no phenol red), trypan blue solution (0.4%), Corning™ Matrigel® growth factor reduced (GFR) basement membrane matrix (phenol red-free, LDEV-free) for organoid culture, Fetal bovine serum (FBS), and GIBCO Hank's balanced salt solution (HBSS) were purchased from Thermo Fisher Scientific (Bedford, MA).
2.2. Caco-2 cell culture and maintenance
Caco-2 cells (passage 29) were cultured and maintained according to the methods previously described by Volpe et al.[7,56]. After rapid thawing in a 37°C water bath, Caco-2 cells were transferred to a centrifuge tube containing 2.0 mL Dulbecco's modified Eagle medium/Ham's F-12 (GIBCO™ Advanced DMEM/F-12, Thermo Fisher Scientific) supplemented with 10% FBS (CCCM) and were centrifuged at 100 x g for 5 min at 4°C. The cell pellet was resuspended with 10 mL of CCCM (Caco-2 cell culture medium) and cultured in a cell culture flask at 37°C in a humidified O2/CO2 incubator (Panasonic, PHC Corporation of North America, IL) in the presence of air supplemented with 5% CO2. After 24 hours, CCCM media was supplemented with 100 U/mL penicillin and 100 µg/mL streptomycin (Pen-Strep). During subculture, CCCM media with Pen-Strep was replaced three times per week until cells attained 80% confluence.
For subculture (passaging), Caco-2 cells at approximately 80% confluence were rinsed initially to remove all traces of trypsin inhibitor and then treated with 0.25% (w/v) trypsin and 0.53 mM EDTA solution (Thermo Fisher Scientific) at 37°C for 10 min. The cell culture flask was observed frequently under an inverted microscope (DMi1, Leica Microsystems, Germany) to monitor trypsinization progress. Over-trypsinization was avoided as it can damage cells and induce clumping. After approximately 70%-80% of the cells had been detached from the surface, 3 mL of pre-warmed CCCM media was added to the cell culture flask, and cells were separated from clumps using repeated pipetting. Caco-2 cells were sub-cultured in new flasks with a medium at a ratio of 1:6 to 1:10.
2.3. Caco-2 two-dimensional (2D) monolayer preparation and maintenance
Once Caco-2 cells were at least 80% confluent, epithelial cells were passaged as described in the previous section. The harvested Caco-2 cell suspension was then filtered through a sterile cell strainer (40 µm nylon mesh, Thermo Fisher Scientific) to obtain a single-cell suspension. Caco-2 cell viability and concentration were determined by trypan blue exclusion test[57,58]. Caco-2 single cells (passages 50-55) were seeded at a density of 75,000 cells/cm2 in the apical chamber of the insert and cultured in CCCM media. The culture medium in both apical and basolateral chambers was changed every other day. In the transport studies, the monolayers were utilized 18 to 20 days after plating. Transepithelial electrical resistance (TEER) was measured every other day (described later in Section 2.6).
2.4. Maintenance of 3D colonoids
The collection and analysis of canine colon biopsy samples were approved by the Iowa State University (ISU) Institutional Animal Care and Use Committee (IACUC) (IACUC-22-050). All methods were performed in accordance with the relevant guidelines and regulations of IACUC as required by U.S. federal regulations. The study is reported in accordance with ARRIVE guidelines (https://arriveguidelines.org). The tissue used for generating the canine colonoids was obtained from a healthy male, 18-month-old dog and colonoids were cultured and maintained according to methods previously described by our laboratory [32,35,36]. In brief, colonic crypts containing primary adult intestinal stem cells (ISC) were isolated using the cold EDTA chelation method[35]. Complete medium with ISC growth factors (CMGF+) containing advanced DMEM/F12 (GIBCO) supplemented with 1:100 GlutaMAX™ supplement (Fisher), 200 mM HEPES (Fisher), and 100 μg/mL Primocin® antibiotics (InvivoGen), 1X B-27™ (Fisher), 1X N-2 supplement (GIBCO™), 1mM N-acetylcysteine (Sigma-Aldrich), 50 ng/mL EGF (PeproTech), 100 ng/mL Noggin (PeproTech), 500 ng/mL R-spondin-1 (PeproTech), 100 ng/mL Wnt 3a (PeproTech), 10 nM gastrin (Sigma-Aldrich), 10 mM nicotinamide (Sigma-Aldrich), 500 nM A83-01 (TGFβ type I receptor inhibitor; Tocris), 10 μM SB202190 (P38 inhibitor; Sigma-Aldrich), and 8% fetal bovine serum (Atlanta Biologicals), supplemented with 10 μM Rho-associated kinase inhibitor (ROCKi) Y-27632 (StemGent) and 2.5 μM glycogen synthase kinase 3β (GSK3β) inhibitor CHIR99021 (StemGent) was used as the culture medium. The isolated ISCs were grown in Matrigel™ in CMGF+ media to progress differentiation[35]. Though CMGF+ medium with ROCKi and GSK3 inhibitor was used for the first two days of ISC culture to boost ISC survival and prevent dissociation-induced apoptosis (anoikis), CMGF+ medium without ROCKi and GSK3 inhibitor was utilized after the first two days of culture to drive differentiation of the canine colonoids[35]. Organoids were fully differentiated (Figure 3A) after 6–8 days in differentiation media (CMGF+ without ROCKi and GSK3 inhibitor), exhibiting a luminal compartment, crypt epithelium, and villi-like structures, as well as the exfoliation of denuded epithelia into the lumen[35]. The CMGF+ culture medium was changed every other day, and organoids were passaged once a week when they were potentially “mature”.
2.5. Preparation of 2D canine colonic monolayer and maintenance
After seven days of culture (Figure 3A), the 3D colonoids were harvested from Matrigel® using Corning™ cell recovery solution. Matrigel® plugs were dissociated by repeated pipetting after adding 500 μL of pre-chilled cell recovery solution into each well, and the plate was incubated for 30 min at 4°C to dissolve Matrigel®. The mixture was then transferred to a 15 mL tube and centrifuged at 100 x g for 5 min at 4°C. The organoid pellet was resuspended in 1 mL TrypLE Express and incubated at 37°C for 10 min with shaking. To stop TrypLE action, 6 mL of DMEM/F12 was added to the mixture, before centrifugation at 100 x g for 5 min at 4oC. The supernatant was removed, and the pellet was resuspended with CMGF+ media. The harvested colonoid cell suspension was filtered through a sterile cell strainer (40 µm nylon mesh, Thermo Fisher Scientific) to obtain a single-cell suspension. The viability of dissociated cells was assessed by trypan blue exclusion test [58,59], and cells were counted manually using a hemocytometer (Hausser Scientific). The Transwell™ inserts were pre-coated with Matrigel® (100 μg/mL; Corning®) and rat tail collagen I (30 μg/mL; Fisher Scientific), to promote adhesion, growth, and differentiation, in CMGF+ media at 37°C for 1 h. Dissociated colonoid cells (passages #15-20) were seeded at a density of 100,000 cells/cm2 in pre-coated inserts and allowed to grow and differentiate in the presence of CMGF+ media for 10 to 12 days before the experiments (Figure 3B). The CMGF+ culture medium was changed every other day. The structural integrity of the cell monolayer was monitored every other day for up to two weeks by phase-contrast microscopy. Colonoid-derived monolayers and colonoids were fixed and processed for histology and/or TEM, as described in prior studies [35,36,60].
2.6. RNA Extraction and quantitative real-time polymerase chain reaction (qPCR) analyses
After adding TRIzol RNA isolation reagent (ThermoFisher Scientific) to each monolayer (when they attain steady Transepithelial Electrical Resistance (TEER) value, i.e., Day 11 for colonoid-derived monolayers and Day 19 for Caco-2-derived monolayers) and pipetting up and down, cellular contents were removed for total RNA extraction using the TRIzol method as described by the manufacturer. After extraction, total RNA samples were purified using the RNeasy MinElute Cleanup Kit (QIAGEN) following the manufacturer's instructions. Purified RNA samples were quantified and assessed for quality using a Bioanalyzer 2100 instrument (Agilent Technologies, Palo Alto, CA, USA). A total of 0.5 µg of purified RNA was used to synthesize first-strand cDNA according to the manufacturer’s instructions using SuperScript III first-strand synthesis kit (Invitrogen).
qPCR for expression of intestinal epithelial cell markers, tight and adherens junction proteins, transporters and cytochrome P450 (CYP) enzymes (primers listed in Table 1) was carried out using the synthesized complementary DNA (cDNA), which contains only the coding DNA sequences, using PowerUp SYBR Green Master Mix following the manufacturer’s protocol using QuantStudio™ 3 (Applied Biosystems by Thermo Fisher). Thermocycling conditions were as follows: 50°C for 2 min and then 95°C for 2 min, followed by 35 cycles of 95 °C for 15 s, 55 °C for 15 s, and 72°C for 1 min. The expression of each gene of interest was normalized using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and quantified using the delta-delta Ct method [38]. The gene expression results were displayed as normalized Ct (dCt) to GAPDH. A canine liver sample was used for RNA extraction and cDNA synthesis for use as positive control for Oatp2b1 expression studies.
2.7. Assessment of monolayer integrity by TEER measurement and Fluorescein Isothiocyanate (FITC)-Dextran leakage assay
TEER is a quantitative technique that measures the electrical resistance of the monolayers, reflecting the ionic conductance of the paracellular pathway in each monolayer[61] and is indicative of the integrity of the cell monolayer. A Millipore Millicell® ERS-2 epithelial volt-ohm meter was used to measure TEER values every day. TEER measurements of individual monolayers (Rsample) were normalized by comparison to a cell-free insert (Rblank) and multiplied by the area of the insert (0.33 cm²) as follows (Eq. 1):
TEER (Ω.cm2) = [Rsample − Rblank] ×0.33 cm2
TEER measurements were performed in all experimental replicates for both Caco-2 cell and canine colonoid monolayers. Prior to incubation with the model drugs, only monolayers with steady state TEER values[36] above 500 Ω×cm2 and 4000 Ω×cm2 were utilized for Caco-2 cells and colonoid-derived monolayers, respectively[9,62,63].
In addition to TEER values the canine colonoid monolayer integrity was evaluated with the commonly used zero permeability compound 4kDa FITC-Dextran[61]. Since FITC-dextran does not impact the permeability of metoprolol or the integrity of the monolayer, FITC-dextran was combined with metoprolol to simultaneously confirm TJ structural integrity of the canine colonoid monolayer [64]. FITC-dextran concentrations over time were measured using a SpectraMax® M2e (Molecular Devices, San Jose, CA) microplate reader at excitation and emission wavelengths of 490 and 520 nm, respectively[13]. Briefly, the donor (AP) side (pH 6.8) contained 200 µg/ml FITC-dextran with or without metoprolol (0.4 mg/mL). The receiver (BL) side contained the HBSS transport buffer at a pH of 7.4. The percentage of relative fluorescence units (%RFU) was measured for the donor and receiver sides of the wells at 120 min, as previously described[13].
2.8. Preparation of transport buffer and drug solutions
Stock solutions of the test drugs (metoprolol, atenolol, and propranolol) were prepared in distilled water. Working solutions were prepared in the appropriate transport buffer for use at the pH (6.8 or 7.4) of the in vivo condition being replicated. All assays were performed in triplicates. Hank's balanced salt solution (HBSS) buffer with Ca2+ and Mg2+ was used as a transport buffer (GIBCO™). Inclusion of Ca2+ and Mg2+ in the buffer prevents monolayer detachment from the filter. The pH of the transport buffer was adjusted to pH 6.8 or 7.4 with 1N HCl or 1N NaOH before use. Final drug concentrations were 0.04 mg/mL and 0.4 mg/mL metoprolol, 0.2 and 2.0 mg/mL atenolol, and 1.0 mg/mL propranolol. The physicochemical characteristics of the three tested drugs are presented in Table 2, based on the information available in PubChem [65].
2.9. Bidirectional transport experiments
All transport experiments were conducted at room temperature in accordance with the conditions outlined in earlier reports during sampling and incubation [66,67,68]. The culture media was aspirated from both AP (0.2 mL) and BL (0.7 mL) chambers. For both the canine colonoid and the Caco-2 monolayer, the BL chambers with HBSS buffer at pH 7.4 (0.7 mL) and the AP chambers with appropriate HBSS buffer at pH 6.8 or 7.4 (0.2 mL) (canine) or pH 6.8 (0.2 mL) (Caco-2 cells) were rinsed and equilibrated at room temperature for 30 min. Two to three wells per drug/pH/direction were used for data generation.
2.9.1. Apical-to-basolateral [AP→BL] permeability
After Caco-2 and canine colonoid monolayers were equilibrated in the transport buffer for 30 min, the buffer was aspirated from the AP (0.2 mL) and BL (0.7 mL) chambers. The media of the AP chamber was replaced with 0.2 mL of the drug solution prepared in an appropriate transport buffer. HBSS buffer with pH 7.4 was added to the BL chambers. At pre-defined timepoints (15, 30, 45, 60, 90, and 120 min), samples of 100 µL buffer were collected from the receiver chamber (BL) and replaced with the same volume (i.e., 100 µL) of transport buffer at pH 7.4. For the AP side, drug concentrations of the donor solution were measured prior to introduction into the well and at the end of the study (120 min). In so doing, no buffer replacement was needed. All study samples were labeled and stored at -80°C until further analysis.
2.9.2. Basolateral-to-apical [BL→AP] permeability
The same procedures as described above for studying AP→BL permeability were followed when studying BL→AP, with the only difference being that the donor chamber was now the BL side, and the receiver chamber was now the AP side. At pre-determined time intervals (15, 30, 45, 60, 90, and 120 min), 100 µL of buffer was removed from the receiver chamber (AP) and replaced with the same volume (i.e., 100 µL) of transport buffer with the specified pH. The collected samples were stored at -80°C until further analysis by mass spectrometry (details below).
2.10. Sample preparation for mass spectrometer analysis
All the experimental samples in HBSS buffer were prepared by transferring 40 µL of buffer into a 1.5 mL polypropylene tube. A volume of 160 µL of acetonitrile was added to each sample, and all samples were vortexed for 30 seconds. Samples were diluted in water based on the expected concentration in the donor or receiver wells. Samples containing atenolol were diluted into a linear range of 0.5-30 µg/mL. Specimens containing metoprolol were diluted into a linear range of 0.2-40 µg/mL, and samples containing propranolol were diluted into a linear range of 1-300 µg/mL. A calibration curve and three quality control (QC) sample replicates were prepared in pH 6.8 HBSS buffer and pH 7.4 HBSS buffer for each analyte. Calibration curves and QCs were prepared and diluted in the same manner as the samples. All calibration curves had a R2 value of 0.99 and all QC samples showed a bias of less than 10% (Supplemental Figure 1 and Supplemental Table 1).
2.10.1. Preparation of standards and solutions
Metoprolol acid-d5, atenolol, atenolol-d7, and propranolol HCl analytical standards were purchased from Toronto Research Chemicals (Toronto, ON, CA). Metoprolol was purchased from Sigma-Aldrich (St. Louis, MO, USA). Propranolol-d7 was purchased from Cerilliant® (Round Rock, TX, USA) as a 100 ng/µL solution in methanol. All stock solutions were prepared at 1 mg/mL in methanol or 50/50 (v/v) methanol/water except for propranolol-d7, which was purchased in solution at a concentration of 100 ng/µL.
2.10.2. Analytical method
A Vanquish™ Flex LC pump interfaced with a TSQ Altis mass spectrometer (MS) (Thermo Scientific™, San Jose, CA, USA) was used to analyze all analytes. The column used for all analyses was a Hypersil GOLD™ aQ Vanquish 50 x 2.1, 1.9 µm (Thermo Scientific™). Mobile phase A was water plus 0.1% formic acid, and mobile phase B was acetonitrile plus 0.1% formic acid for all analyses. The column compartment temperature was 40°C for all analyses. The chromatographic gradient was as follows: start 0% organic, linear ramp from 0.5 to 2 min up to 80% organic (100% organic for propranolol), hold at 80% organic (100% organic for propranolol) for 0.4 min, return to starting conditions for 0.01 min, and hold at starting conditions for 0.59 min. The solvent flow rate was 0.4 mL/min. A 2 µL injection volume was used.
The following MS method parameters were used for the analysis of all analytes. The resolution of Q1 and Q3 was 0.7 FWHM. The CID gas was set to 1.5 mTorr. The chromatographic peak width was 2 sec, and the cycle time was 0.2 sec. The source conditions were as follows: positive mode spray voltage 2000 V, sheath gas 60 Arb, auxiliary gas 22 Arb, sweep gas 1 Arb, ion transfer tube temperature 350°C, and vaporizer temperature 350°C. The total run time of the method was 3 min.
2.11. Papp Calculations
The uncorrected Papp estimates for the three drugs of interest (Papp-uncorrected, cm/sec) were calculated as follows (Eq. 2)[7]:
where VR is the volume in the receiver chamber (mL), is the surface area of the filter (), C0 is the initial drug concentration (µM) in the donor chamber, and is the maximum slope of the line defining the cumulative drug concentration (mg/mL) vs. time (min) in the receiver compartment. Drug concentrations in the donor and receiver chambers at designated timepoints were measured by mass spectrometry (see details above in Section 2.9.2). The estimated concentrations in the receiver compartment were corrected for drug removed during prior samples. The efflux ratio was calculated as Papp,BL-AP/Papp,AP-BL, where a ratio greater than 2 indicates active efflux[69,70].
These Papp value estimates were further corrected for the percent recovery (Rec (%)) in each well[71,72] as follows (Eq. 3):
The corrected Papp estimates reported herein were therefore calculated as follows (Eq. 4):
The slope of the relationship between sampling time and the corrected Papp values was calculated as the least squares estimate of the linear regression line defining the drug transfer rate from the first quantifiable drug concentration in the donor cell to 120 min.
2.12. Statistics
Since only two wells were tested per monolayer type and assay condition in this exploratory study, there was insufficient power to assess statistical significance of Papp differences between model systems. Therefore, descriptive statistics (arithmetic mean, SD, SEM, CV%) were used to summarize Papp estimates for the various experimental conditions in this preliminary study. Comparative expression of transporters and CYP enzymes expression in Caco-2-derived and canine colonoid-derived monolayers was performed with GraphPad Prism 9 (version 9.4.1) (https://graphpad.com/) using a one-way ANOVA with Šídák’s adjustment for multiple-comparisons, or a two-tailed Student’s t-test, as appropriate. P-values < 0.05 were considered as statistically significant for all analyses.
3. Results
3.1. Assessment of monolayer integrity
TEER values were recorded for both colonoids and Caco-2 cells to determine the integrity and confluence of the monolayer, as reported in Figure 4 and Figure 5. Before initiating the permeability studies, the wells containing the 2D canine colonic monolayer were maintained for 11 days. The colonic monolayer TEER values began to plateau on Day 10-12 at an average of 4198.3 ± 142.4 Ω.cm2 (mean ± SD) (Supplemental Table 2A). Caco-2 cell monolayers were monitored for a total of 23 days. The TEER values began to plateau around Day 18-21, with an average of 501.4 ± 15.6 Ω.cm2 (mean ± SD) (Supplemental Table 2B).
In the colonoid monolayer, FITC-Dextran was combined with metoprolol to assess monolayer integrity and measure the permeability of metoprolol simultaneously. For FITC-dextran without metoprolol, the percentage of relative fluorescence was estimated at 93.7% RFU and < 0.02% RFU in the donor and receiver side, respectively. In comparison, these estimates were 94.6% Relative Fluorescence (RFU) (donor side) and < 0.02% RFU (receiver side) for FITC-dextran with metoprolol (Table 3). This suggests that the FITC-dextran transport was less than 0.02% both with and without metoprolol, thus confirming the integrity of the colonoid monolayer.[38]
3.2. Drug-specific Papp estimates
The estimated Papp values per drug, direction, pH, and system are provided in Table 4 (see Supplemental Table 3 for estimated drug concentration).
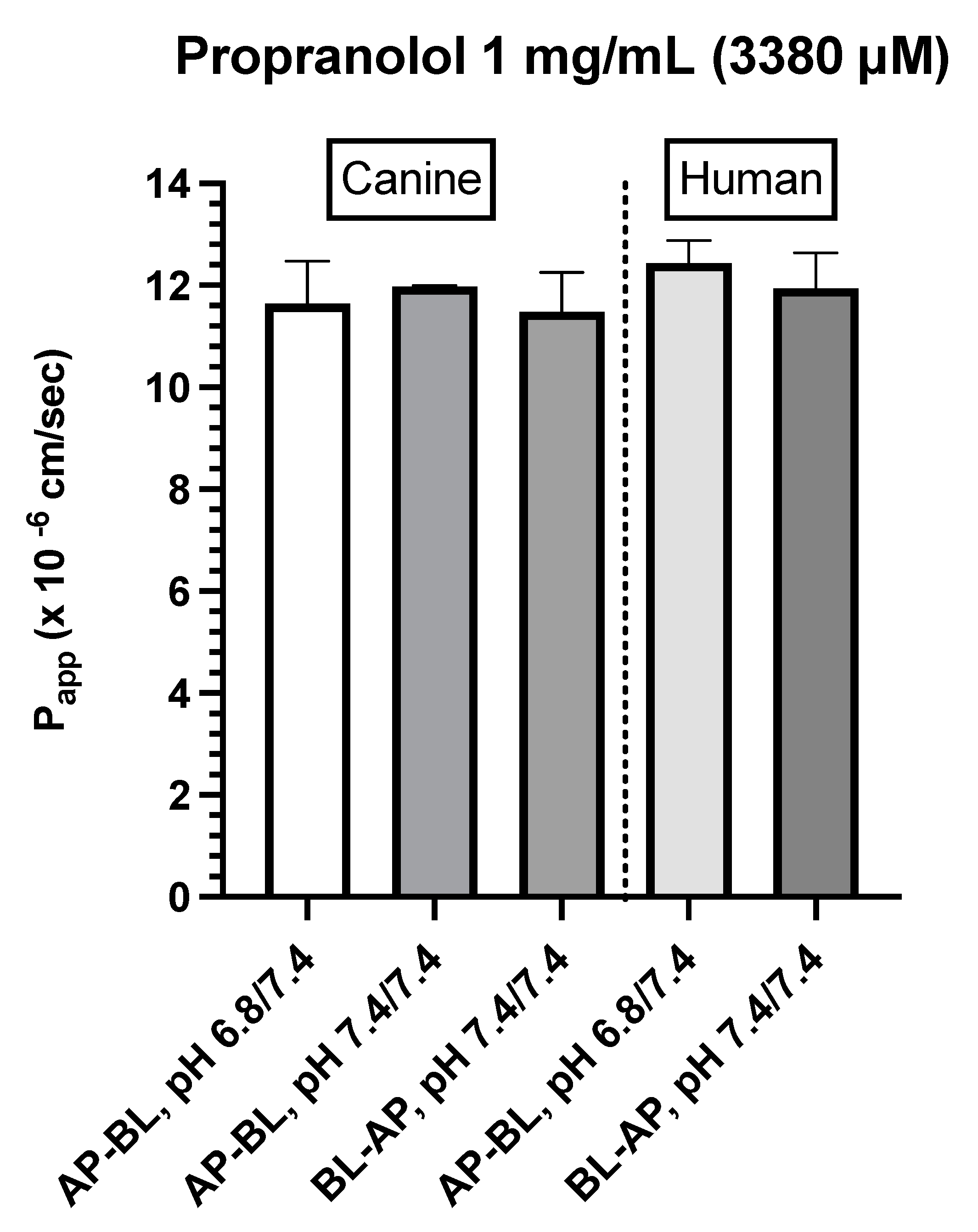
Propranolol (1 mg/mL)
Figure 6.
A. Comparison of propranolol Papp estimates in canine colonoid vs. human Caco-2 monolayers as a function of transport direction (AP-BL vs. BL-AP) and apical pH (6.8 vs. 7.4).
Figure 6.
A. Comparison of propranolol Papp estimates in canine colonoid vs. human Caco-2 monolayers as a function of transport direction (AP-BL vs. BL-AP) and apical pH (6.8 vs. 7.4).
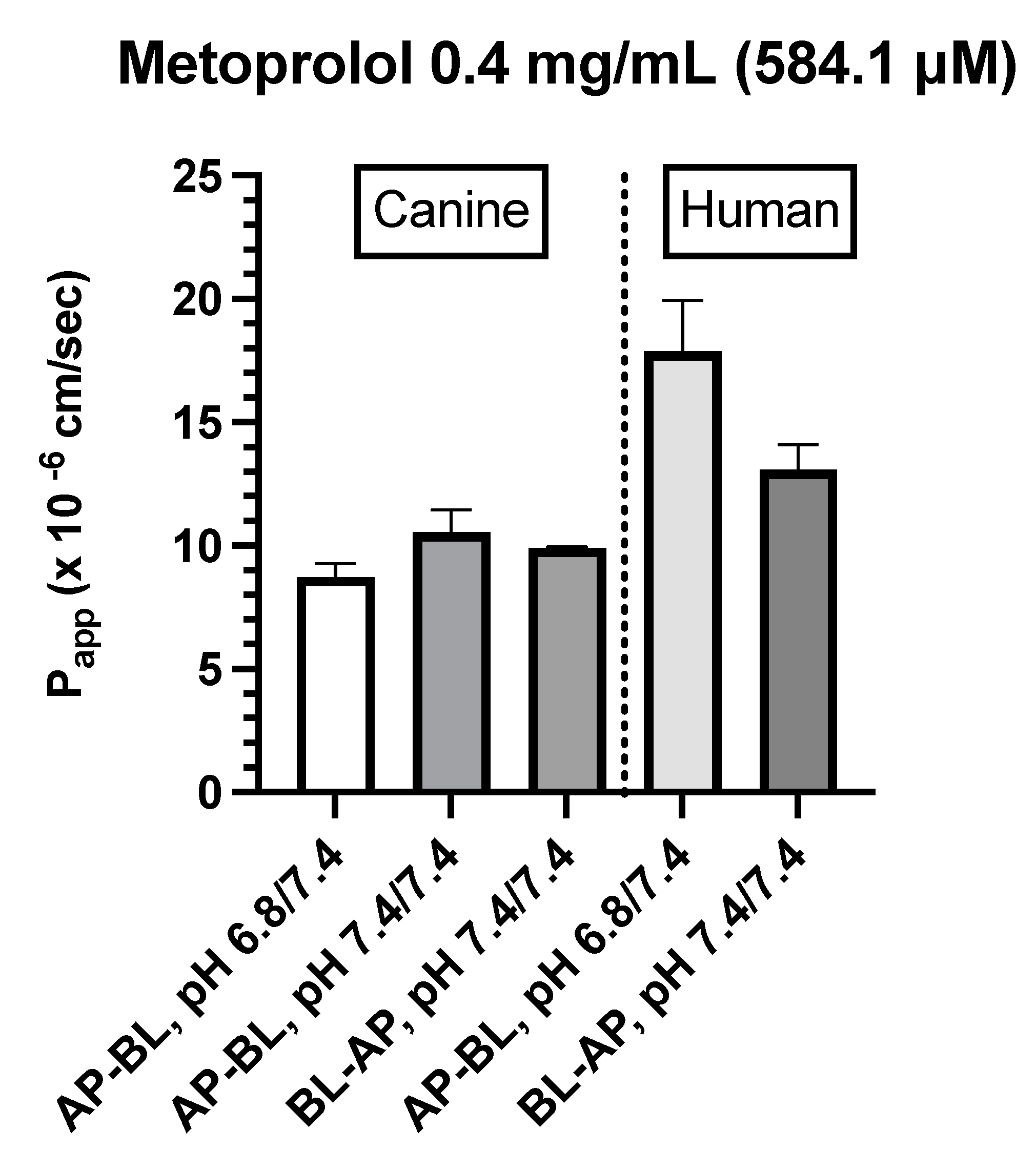
Metoprolol (0.04 and 0.4 mg/mL)
- Canine Colonoids: The AP→BL Papp values were lower in the medium containing 0.4 vs. 0.04 mg/mL metoprolol. Moreover, while at the 0.04 mg/mL concentration, the movement from AP→BL was slightly greater than that from BL→AP. The transport from AP→BL and BL→AP tended to be similar when evaluated at the 0.4 mg/mL concentration. Upon considering the data generated across the two metoprolol concentrations, the pH of the apical chamber did not consistently influence the magnitude of the Papp estimate (Figure 6B,C & Table 4).
- Caco-2: Unlike canine colonoids, Papp values were not markedly influenced by metoprolol concentration and at both concentrations, the movement from AP→BL and BL→AP were comparable (Figure 6B,C & Table 4). The concentration-associated differences in Papp values seen with the colonoid were not observed with the Caco-2 monolayer. Moreover, although the two cell-line monolayers exhibited similar AP→BL Papp values in the presence of 0.04 mg/mL metoprolol, the AP→BL Papp values for the Caco-2 monolayer tended to be higher than that of the canine colonoid when the donor concentration was increased to 0.4 mg/mL. Although the movement from AP→BL was somewhat greater than that seen in the BL→AP direction in the colonoid (0.04 mg/mL metoprolol but not at the 0.4 mg/mL donor concentration), that difference was not seen with the 0.04 mg/ml concentration or was only minimally appreciated at the 0.4 mg/mL concentration when the Caco-2 monolayer was used. When considering the variability across observations and the small number of wells tested, statistical inferences should not be linked to these outcomes (Figure 6C,D & Table 4). Thus, unlike propranolol, differences in the behavior of metoprolol were seen when comparing the two cell-line systems. However, these preliminary findings should be interpreted cautiously in light of our limited sample size and background variability in our system.
Figure 6.
B. Comparison of metoprolol (0.04 mg/mL) Papp estimates in canine colonoid vs. human Caco-2 monolayers as a function of transport direction (AP-BL vs. BL-AP) and apical pH (6.8 vs. 7.4).
Figure 6.
B. Comparison of metoprolol (0.04 mg/mL) Papp estimates in canine colonoid vs. human Caco-2 monolayers as a function of transport direction (AP-BL vs. BL-AP) and apical pH (6.8 vs. 7.4).
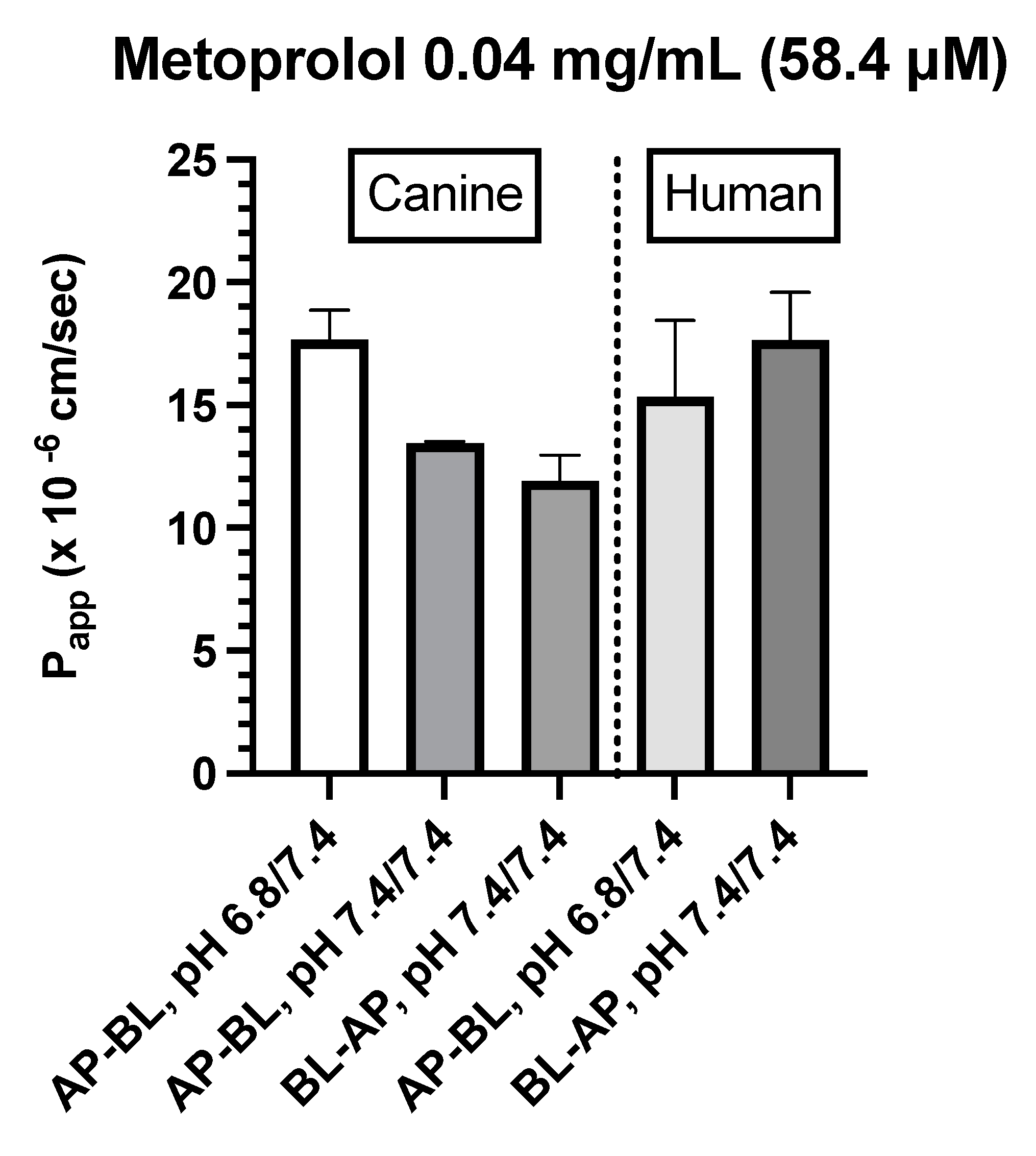
Figure 6.
C. Comparison of metoprolol (0.4 mg/mL) Papp estimates in canine colonoid vs. human Caco-2 monolayers as a function of transport direction (AP-BL vs. BL-AP) and apical pH (6.8 vs. 7.4).
Figure 6.
C. Comparison of metoprolol (0.4 mg/mL) Papp estimates in canine colonoid vs. human Caco-2 monolayers as a function of transport direction (AP-BL vs. BL-AP) and apical pH (6.8 vs. 7.4).
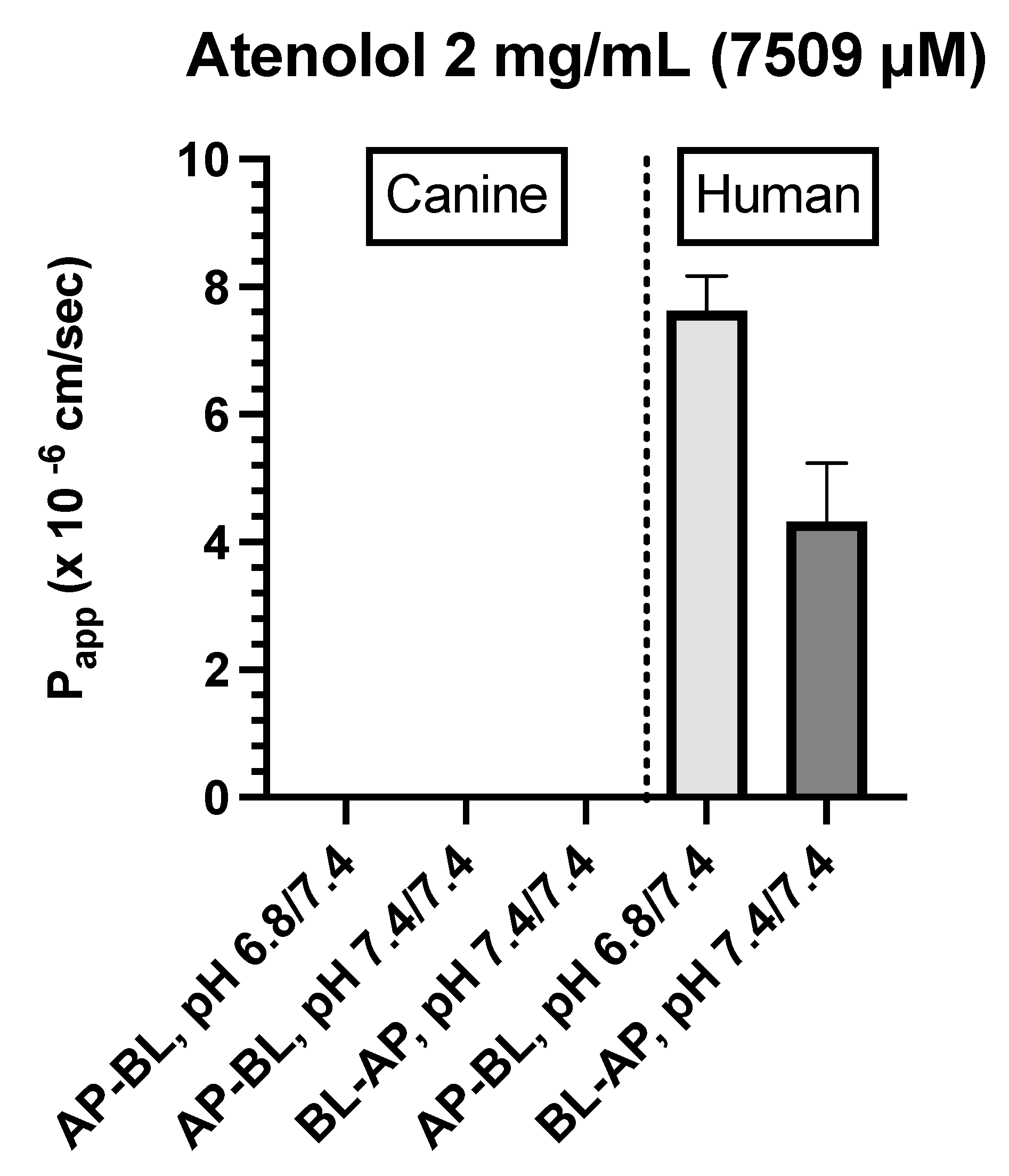
Atenolol (0.2 mg/mL)
For atenolol (0.2 mg/mL), the transport was not quantifiable in either direction for the canine colonoid and the Caco-2 monolayer (Table 4). However, because values were below the analytical limit of quantification, we increased the donor concentration 10-fold (i.e., 2.0 mg/mL) to ensure that we were able to determine if some paracellular movement did in fact occur across either of the two monolayers.
Atenolol (2.0 mg/mL)
- Caco-2: Incubation of Caco-2 cells with 2.0 mg/mL atenolol resulted in measurable concentrations in the receiver compartment for both AP→BL and BL→AP directions, with higher Papp estimates reported after incubation in the apical chamber (Figure 6D & Table 4). These preliminary findings suggest a somewhat greater ability for atenolol to undergo paracellular transport across the Caco-2 monolayer as compared to that of the dog colonoid. However, at the lower atenolol concentration, neither system was associated with quantifiable movement from donor to receiver compartment (irrespective of direction).
Figure 6.
D. Comparison of atenolol (2 mg/mL) Papp estimates in canine colonoid vs. human Caco-2 monolayers as a function of transport direction (AP-BL vs. BL-AP) and apical pH (6.8 vs. 7.4).
Figure 6.
D. Comparison of atenolol (2 mg/mL) Papp estimates in canine colonoid vs. human Caco-2 monolayers as a function of transport direction (AP-BL vs. BL-AP) and apical pH (6.8 vs. 7.4).
3.3. Gene Expression Analyses
The integrity and differentiation of colonoid-derived monolayers were confirmed by measuring the expression of tight and adherens junction proteins, including CDH1, OCLN, and TJP1, as well as intestinal epithelial cell differentiation markers like MUC2 (for goblet cells), NEUROG3 (for enteroendocrine cells) and ALP (for absorptive epithelium) (Figure 7; Supplemental Table 4). Additionally, stem cell markers such as OLFM4, HOPX, PROM1, and SOX9 were detected in colonoid-derived monolayers (Figure 7; Supplemental Table 4. See caption of Figure 7 for definition of abbreviations). The electron microscopic images of colonoids also clearly showed the presence of goblet cells containing mucin granules (Figure 8). The expression of key intestinal epithelial transporter molecules, such as Mdr1 and Oatp2b1, and important CYP enzymes expressed in the intestine, such as Cyp3a12, Cyp2b11, and Cyp2c21, was measured in canine colonoid-derived monolayers and compared to expression of their orthologs in human Caco-2-derived monolayers. In canine colonoid-derived monolayers, the expression of Mdr1, Cyp3a12, and Cyp2c21 was increased relative to their orthologs in Caco-2-derived monolayers, as expected (Figure 9 and Figure 10; Supplemental Table 4). We also used qPCR to evaluate transporter and CYP metabolic enzyme expression in other canine cell lines utilized for drug uptake study, such MDCK cells, which demonstrated low expression of these markers (data not shown). Our findings were consistent with those of prior research [73]; hence it was not included in the study.
Submucosa (SM); Muscularis mucosa (MM); Lamina propria (LP); Rows of goblet cells (GC) are oriented toward the gland lumen (L); mucus-filled goblet cell (GC); microvilli (MV); zonula adherens (ZA); desmosome or macula adherens (MA).
Gene expression results are displayed as normalized Ct (dCt) to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (see Supplemental Table 4 for Ct values). Values are expressed as the arithmetic mean of the data and one standard deviation of N = 3 monolayers. Between-groups statistics were performed with GraphPad Prism 9 (version 9.4.1) (https://graphpad.com/) using one-way ANOVA with Šídák’s adjustment for multiple-comparisons (for OATP2B1), or a two-tailed Student’s t-test (for MDR1). P-values < 0.05 were considered as statistically significant for all analyses. **** P < 0.0001. ns = no significance difference.
4. Discussion
Identifying the permeability characteristics of an orally administered drug is one of the critical steps in the prediction of GI drug absorption. This information helps identify factors that can influence oral bioavailability and guide formulators in their efforts to optimize the fraction of the administered dose that is absorbed. While in vitro tools for exploring drug permeability and enterocyte drug metabolism are available for human therapeutics, there is no corresponding tool available that faithfully models the canine GI tract. Therefore, we studied the feasibility of using canine colonoid-derived 2D-monolayers for an apparent permeability assessment, focusing first on the passive diffusion of drugs across the canine enterocyte membrane.
Generally, we expect that passive transcellular permeability characteristics are translatable across biological membranes. This assumption is supported by the use of the MDCK cells and Lewis lung carcinoma-porcine kidney cells (LLC-PK1) for the evaluation of passive absorption mechanisms [75]. However, differences between cell lines may exist when assessing paracellular absorption [55,76]. Therefore, we explored the feasibility of the canine colonoid system using three well-characterized β-blockers with transcellular or paracellular absorption and determined the validity of our assumptions by comparing canine colonoid Papp values to those obtained when the same compounds were tested using the Caco-2 cell system. If differences in Papp estimates between systems were identified, then the use of Caco-2 data to predict canine drug intestinal passive permeability would need to be further examined.
Consistent with our expectations, we observed comparable absorption (AP→BL) and secretion (BL→AP) characteristics across the two systems for propranolol. In contrast, system dissimilarities were observed for metoprolol. For example, while the concentration of metoprolol influenced colonoid Papp values across both AP→BL and BL→AP directions, such differences were not seen when using the Caco-2 monolayer. Furthermore, Papp values in both directions were higher for the Caco-2 monolayer when the donor concentration was 0.4 mg/mL as compared to that of the colonoid. In contrast, when the donor concentration was 0.04 mg/mL, AP→BL (apical pH = 6.8), Papp values were comparable. Only movement in the BL→AP direction was greater for the Caco-2 vs canine colonoid monolayer systems.
Although we assumed that transcellular permeability would be similar across species and tissues, one possible reason for the observed differences between Caco-2 and colonoids could be unanticipated regional differences across the enterocytes lining the various intestinal regions and differential expression of transporters and CYP enzymes. This possibility was explored by Incecayir et al.[77] and Zur et al.[78]. Incecayir et al. estimated metoprolol Peff values using an in situ single-pass intestinal perfusion system (SPIPS) in mice and/or rats[77]. They observed that the metoprolol intestinal permeability of both species higher in the distal ileum vs. the jejunum. Recognizing that Peff values factor surface area into its estimate, this outcome likely reflects segmental surface area differences or dissimilarity in intestinal mucus boundary layers. However, in contrast to the aforementioned rodent study, using a SPIPS study design in human subjects, Dahlgren et al. detected no statistically significant regional differences in metoprolol Peff in the colon vs. the ileum [79]. Altogether, these results are not consistent with a possible influence of cellular configuration of the Caco-2 cell vs. canine colonoid.
With regard to potential transporter involvement, Incecayir et al. confirmed that metoprolol is not a substrate for P-gp transport, excluding the notion that observations were due to differences in efflux transporter expression [77]. Moreover, if P-gp transporters were in fact involved, it would have negatively influenced movement from AP→BL (decrease in Papp values) but positively increased Papp values in the BL→AP direction. This is not consistent with what was observed experimentally, either at a donor concentration of 0.04 mg/mL or for the BL→AP Papp values (colonoid vs. Caco-2 results) at the donor metoprolol concentration of 0.4 mg/mL. Factoring this point along with our correction for drug loss during our study, we can reasonably assume that expression of efflux transporters (or losses associated with enterocyte drug metabolism) cannot explain the differences seen between the two systems.
Another point considered in the SPIPS studies by Incecayir et al.[77] and Zur et al.[78] was the potential influence of pH on metoprolol permeability. Although higher pH tended to be associated with higher Peff values, this difference was not statistically significant in the rat (N=6). In contrast, in the rat SPIPS study conducted by Zur et al. markedly higher Peff values were observed as perfusate pH was raised from 6.5 to 7.5 (N=6 per experiment) [78]. In addition, both Incecayir et al.[77] and Zur et al.[78] reported a significant pH-associated change in permeability using either the Caco-2 monolayer or octanol/buffer partition coefficient (and PAMPA membrane)[78]. In all cases, the permeability of metoprolol decreased as the donor pH decreased. The investigators attributed this pH effect to metoprolol being a basic secondary amine that serves as its only ionizable center. As a result, the fraction unionized of metoprolol is negligible at low pH (i.e., at pH values less than the corresponding pKa), and gradually increases as the pH rises. Incecayir et al. suggested that the small effect of pH on in vivo absorption vs. in vitro permeability may have been attributable to the in vivo presence of a mucous layer which retains the microclimate pH, regardless of the luminal pH [77]. Interestingly, however, we observed the opposite effect of pH on our 0.04 mg/mL metoprolol Papp values where there tended to be a decrease rather than increase as pH was raised from 6.8 to 7.4. Therefore, again, the reasons for our observations with metoprolol are not readily apparent.
Finally, unlike that of the Caco-2 monolayer or colonoid permeability studies conducted at a donor metoprolol concentration of 0.04 mg/mL, FITC-dextran was included when the canine colonoid donor concentrations were 0.4 mg/mL. While this fluorescent probe has not been associated with changes in drug transcellular permeability, we do not have the data to exclude that possibility in our current investigation. Therefore, an impact of this design difference between permeability study conditions cannot be excluded. The permeability findings on Caco-2 monolayers from Day 14 with low TEER value[54] indicate that the transport of hydrophilic marker (FITC-dextran) and hydrophilic drug (atenolol) was significantly increased, whereas the transport of metoprolol remained unchanged. The findings indicate that a hydrophilic drug such as atenolol can cross the leaky intercellular connection via the paracellular channel. This assessment is consistent with the observations of Yang et al. (2007), where palmitoylcarnitine (PCC), a compound that opens tight junctions, increased the transport of sotalol (a hydrophilic drug) and of FITC-dextran, produced only a small increase in metoprolol transport, but significantly decreased the TEER, the latter indicating a loosening of the tight junction[54].
Unlike the unanswered questions associated with the metoprolol study results, reasons for the observations with atenolol may be found in the investigation by Dahlgren et al. [79]. Using a Single-Pass Intestinal Perfusion (SPIP) study design in human subjects, atenolol exhibited >10-fold lower Peff in the colon as compared to that of the ileum and >350-fold lower Peff in the colon as compared to that in the jejunum[79]. This translated to statistically significantly different drug exposure in vivo values (expressed as area under the concentration time-curve, AUC) when doses were administered to the colon vs. ileum. Extrapolating their observations to our study results and recognizing that the Peff values were influenced by regional differences in absorptive surface area[80], we cannot exclude the possibility that differences observed between atenolol Papp in colonoids vs. Caco-2 cells may reflect differences in the TJ expression between these two systems, since they represent different intestinal segments showing both enterocyte and colonocyte features[7,8]. Under normal culture conditions, Caco-2 cells can spontaneously undergo morphological and biochemical enterocytic differentiation[7]. The cells become polarized, forming a cell monolayer with apical brush boundary microvilli, tight intercellular junctions, villin expression, and dome formation. When cells approach confluence, the number of proteins characteristic of the colon decreases while the number of proteins characteristic of the enterocytes increases[7]. Consistent with this interpretation is the results of the TEER values we obtained to determine the integrity of the intercellular junctional complex.
The TEER of the monolayers used to assess pre-assay integrity of the monolayers in the present study is comparable to previous reports in Caco-2 cells (Figure 5). In the present study, TEER values greater than 500 Ω×cm2 were required in order for Caco-2 cell monolayers to be considered appropriate for use in the transmembrane transport studies [62]. Studies have identified that TEER values in the range of 300-600 Ω×cm2 imply the establishment of robust TJs between cells (which are essential for maintaining good monolayer integrity) [9,62,63]. However, TEER of the Caco-2 cell line is generally higher than that of the in vivo human intestine. Therefore, the passive paracellular pathway in the Caco-2 cells is generally lower than what would be observed in vivo [10].
Based on their relative permeability to small ions, epithelia can be classified as "leaky" or "tight," as proposed by Machen et al. (1972) [81] and Fromter and Diamond (1972) [82], and this designation is still in use at present [83]. "Leaky" epithelia have higher paracellular small ion permeability than transcellular permeability and low transepithelial resistance, while "tight" epithelia have similar or better transcellular small ion permeability than paracellular small ions with higher transepithelial resistance [81,82]. A “leaky” epithelium [83] has a TEER of less than 100 Ω×cm2, indicating greater paracellular permeability, whereas a “tight” epithelium [83] has a TEER of about 2000 Ω×cm2, indicating lower paracellular permeability [61,84]. We observed that monolayers obtained from the canine colonoids have eight times higher TEER values than that of the Caco-2 monolayer, reflecting tighter intercellular junctional complexes than monolayers derived from Caco-2 cells (Figure 4 and Figure 5). That observation is consistent with the higher atenolol Papp values observed in Caco-2 monolayer as compared with that of the canine colonoids when the 2 mg/mL atenolol concentrations was used. Regarding the difference in Papp estimates obtained with 2 mg/mL vs. 0.2 mg/mL atenolol concentrations, we cannot determine if some movement did in fact occur across the Caco-2 monolayer when testing 0.2 mg/mL atenolol, because all receiver compartment concentrations were below the analytical limit of quantification. Higher P-gp (Mdr1) expression in colonoid-derived monolayers relative to MDR1 in Caco-2 cell monolayers is also a potential explanation for the absence of quantifiable atenolol concentrations in colonoid-derived receiver compartments as atenolol is a substrate for P-gp. This was evidenced in studies showing that co-administration of P-gp inhibitors such as verapamil and zosuquidar decreased the efflux ratio of atenolol[85,86] and that co-administration of another P-gp inhibitor, cyclosporine (a non-specific inhibitor of both efflux and influx transporters) increased the absorption rate of atenolol[85,86]. On a final note, we recognize that the Papp values estimated in our study tended to be lower than those reported by others [7,8,10,54,87]. This underscores the importance of comparing permeability results obtained with both systems from the same laboratory. In that regard, variations in permeability results have been observed between laboratories and different Caco-2 culture batches [7,24]. Possible reasons for this observation are the differences in culture and transport protocols between the different research groups [7,42].
Intestinal stem cells (ISCs) can differentiate into progenitor cells that develop into a diverse range of cell lineages. While a variety of secretory cells (including enteroendocrine cells, goblet cells, and tuft cells), and M cells within Peyer's patches are all present in the normal canine large intestine [88], tuft cells are only infrequently observed [89]. Therefore, it is important to confirm that the monolayer used in our studies was in fact colonic enterocytes.
In the present study, while the expression of mucin 2 (MUC2), intestinal alkaline phosphatase (ALP) and neurogenin 3 (NEUROG3) in the canine colonoid-derived monolayers validate the differentiation of intestinal epithelial cells (IECs) into different cell subtypes, the expression of olfactomedin 4 (OLFM4), HOP homeobox (HOPX), prominin 1 (PROM1), SRY-box transcription factor 9 (SOX9), and leucine-rich repeat-containing G protein-coupled receptor 5 (LGR5) genes suggests the presence of intestinal epithelial stem cells[35,38]. PROM1 is a marker for stem cells and early progenitors in the intestine [90]. Both microvilli and brush border enzyme ALP expression also indicate the presence of differentiated enterocytes[35] OLFM4 is also highly expressed in the crypt base columnar cells in the colon [91]. In our previous research, monolayers generated from canine colonoids were also comprehensively studied for differentiated cell lineages displaying both Ki67 expression and the expression of LGR5, an important marker for adult intestinal stem cells. These monolayers showed NEUROG3 and Chromogranin A (CgA) expression further confirming the presence of differentiated neuroendocrine cells. Moreover, monolayers contained epithelia covered with mucus-like substances, indicating mucus production[36]. The presence of differentiated goblet cells containing mucus visualized on transmission electron microscopy (TEM) and Alcian blue staining in the present study confirm findings from our prior research[36].
Continuation and expansion of this preliminary work is based upon the importance of using species-specific monolayers to assess factors such as active influx and efflux transporters, as well as intracellular metabolism on intestinal drug permeability. Moreover, this information should be generated across the various intestinal segments. A first step in supporting the use of such systems is their ability to adequately reflect the passive transport properties (transcellular and paracellular) of the enterocyte membrane.
Despite the limited number of replicates per condition, our study succeeded in providing promising results regarding the utility of canine-derived organoid monolayers for species-relevant assessments of drug passive permeability processes. It also highlighted potential sources of error and the challenges remaining to be addressed. Furthermore, can influence the reliable extrapolation of passive permeability estimates from the Caco-2 cell line to that associated with the canine colon. Clearly, these observed differences warrant future investigations.
5. Conclusion
The potential for the usage of canine colonoid-derived monolayers as a model system for drug permeability assessment is described herein. Despite multiple limitations associated with the use of single-cell monolayers, these systems provide important information on potential barriers to the movement of dissolved-drug from the gut lumen to the portal system. Additional studies with organoids derived from other canine intestinal segments and the testing of a broader array of drugs from the BCS classification are needed to conclusively determine the utility of canine-derived intestinal organoid monolayers for predicting the absorption of therapeutic drugs in dogs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
DKS: data acquisition, analysis and interpretation of data and manuscript draft and critical revision of the manuscript. MNM: concept and design of the study, analysis and interpretation of data, and critical revision of the manuscript. KD: data acquisition and revising manuscript. VG, CZ, TA and Cl-S: data acquisition and revising manuscript. AEJ: concept and design of the study, interpretation of data, and critical revision of the manuscript. LEB and DS: LC-MS data acquisition and technical support. DAV: inputs in study design, analysis and interpretation of data, and revising the manuscript. KA, and JPM: concept and design of the study, interpretation of data, and critical revision of manuscript, funding.
Data Availability Statements
All data generated or analyzed during this study are included in this published article [and its supplementary information files].
Acknowledgments
The authors acknowledge the Department of Veterinary Clinical Sciences (VCS) Core Lab for GraphPad Prism 9 and BioRender program support.
Conflicts of Interest
K. Allenspach, J. Mochel and A.E. Jergens are founders of a biotechnology start-up company, 3D Health Solutions, Inc.
References
- Alqahtani, M.S.; Kazi, M.; Alsenaidy, M.A.; Ahmad, M.Z. Advances in Oral Drug Delivery. Front Pharmacol 2021, 12, 62. [CrossRef]
- Sugano, K.; Kansy, M.; Artursson, P.; Avdeef, A.; Bendels, S.; Di, L.; Ecker, G.F.; Faller, B.; Fischer, H.; Gerebtzoff, G.; et al. Coexistence of Passive and Carrier-Mediated Processes in Drug Transport. Nat Rev Drug Discov 2010, 9, 597–614.
- Roos, C. The Impact of Regional Permeability, Nanoparticles, and Absorption-Modifying Excipients. 2018.
- Furuse, M. Molecular Basis of the Core Structure of Tight Junctions. Cold Spring Harb Perspect Biol 2010, 2. [CrossRef]
- Fleth-James, J. Suitability of in Vitro, in Silico and in Vivo Methods to Predict Intestinal Absorption in Drug Development. 2018. [CrossRef]
- Teksin, Z.S.; Seo, P.R.; Polli, J.E. Comparison of Drug Permeabilities and BCS Classification: Three Lipid-Component PAMPA System Method versus Caco-2 Monolayers. AAPS J 2010, 12, 238. [CrossRef]
- Volpe, D.A. Variability in Caco-2 and MDCK Cell-Based Intestinal Permeability Assays. J Pharm Sci 2008, 97, 712–725. [CrossRef]
- Volpe, D.A. Drug-Permeability and Transporter Assays in Caco-2 and MDCK Cell Lines. Future Med Chem 2011, 3, 2063–2077. [CrossRef]
- Volpe, D.A. Advances in Cell-Based Permeability Assays to Screen Drugs for Intestinal Absorption. [CrossRef]
- Volpe, D.A. Application of Method Suitability for Drug Permeability Classification. AAPS Journal 2010, 12, 670–678. [CrossRef]
- Hill, D.R.; Huang, S.; Tsai, Y.H.; Spence, J.R.; Young, V.B. Real-Time Measurement of Epithelial Barrier Permeability in Human Intestinal Organoids. J Vis Exp 2017, 2017. [CrossRef]
- Pade, D.; Jamei, M.; Rostami-Hodjegan, A.; Turner, D.B. Application of the MechPeff Model to Predict Passive Effective Intestinal Permeability in the Different Regions of the Rodent Small Intestine and Colon. Biopharm Drug Dispos 2017, 38, 94–114. [CrossRef]
- Macedo, M.H.; Martinez, E.; Barrias, C.C.; Sarmento, B. Development of an Improved 3D in Vitro Intestinal Model to Perform Permeability Studies of Paracellular Compounds. Front Bioeng Biotechnol 2020, 0, 1076. [CrossRef]
- Lea, T. Caco-2 Cell Line. The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models 2015, 103–111. [CrossRef]
- Tavelin, S.; Gråsjö, J.; Taipalensuu, J.; Ocklind, G.; Artursson, P. Applications of Epithelial Cell Culture in Studies of Drug Transport. Methods Mol Biol 2002, 188, 233–272. [CrossRef]
- Sai, Y.; Kaneko, Y.; Ito, S.; Mitsuoka, K.; Kato, Y.; Tamai, I.; Artursson, P.; Tsuji, A. Predominant Contribution of Organic Anion Transporting Polypeptide OATP-B (OATP2B1) to Apical Uptake of Estrone-3-Sulfate by Human Intestinal Caco-2 Cells. Drug Metab Dispos 2006, 34, 1423–1431. [CrossRef]
- Balimane, P. v.; Chong, S. Cell Culture-Based Models for Intestinal Permeability: A Critique. Drug Discov Today 2005, 10, 335–343. [CrossRef]
- Ozawa, T.; Takayama, K.; Okamoto, R.; Negoro, R.; Sakurai, F.; Tachibana, M.; Kawabata, K.; Mizuguchi, H. Generation of Enterocyte-like Cells from Human Induced Pluripotent Stem Cells for Drug Absorption and Metabolism Studies in Human Small Intestine. Scientific Reports 2015 5:1 2015, 5, 1–11. [CrossRef]
- Mochel, J.P.; Jergens, A.E.; Kingsbury, D.; Kim, H.J.; Martín, M.G.; Allenspach, K. Intestinal Stem Cells to Advance Drug Development, Precision, and Regenerative Medicine: A Paradigm Shift in Translational Research. AAPS Journal 2018, 20, 1–9.
- Maubon, N.; Le Vee, M.; Fossati, L.; Audry, M.; Le Ferrec, E.; Bolze, S.; Fardel, O. Analysis of Drug Transporter Expression in Human Intestinal Caco-2 Cells by Real-Time PCR. Fundam Clin Pharmacol 2007, 21, 659–663. [CrossRef]
- Neuhoff, S.; Ungell, A.L.; Zamora, I.; Artursson, P. PH-Dependent Passive and Active Transport of Acidic Drugs across Caco-2 Cell Monolayers. European Journal of Pharmaceutical Sciences 2005, 25, 211–220. [CrossRef]
- Yamashita, T.; Inui, T.; Yokota, J.; Kawakami, K.; Morinaga, G.; Takatani, M.; Hirayama, D.; Nomoto, R.; Ito, K.; Cui, Y.; et al. Monolayer Platform Using Human Biopsy-Derived Duodenal Organoids for Pharmaceutical Research. Mol Ther Methods Clin Dev 2021, 22, 263–278. [CrossRef]
- Ito, K.; Suzuki, H.; Horie, T.; Sugiyama, Y. Apical/Basolateral Surface Expression of Drug Transporters and Its Role in Vectorial Drug Transport. Pharmaceutical Research 2005, 22, 1559–1577. [CrossRef]
- Hayeshi, R.; Hilgendorf, C.; Artursson, P.; Augustijns, P.; Brodin, B.; Dehertogh, P.; Fisher, K.; Fossati, L.; Hovenkamp, E.; Korjamo, T.; et al. Comparison of Drug Transporter Gene Expression and Functionality in Caco-2 Cells from 10 Different Laboratories. European Journal of Pharmaceutical Sciences 2008, 35, 383–396. [CrossRef]
- Natoli, M.; Leoni, B.D.; D’Agnano, I.; Zucco, F.; Felsani, A. Good Caco-2 Cell Culture Practices. Toxicology in Vitro 2012, 26, 1243–1246. [CrossRef]
- Hoffmann, P.; Burmester, M.; Langeheine, M.; Brehm, R.; Empl, M.T.; Seeger, B.; Breves, G. Caco-2/HT29-MTX Co-Cultured Cells as a Model for Studying Physiological Properties and Toxin-Induced Effects on Intestinal Cells. PLoS One 2021, 16, e0257824. [CrossRef]
- Sun, H.; Chow, E.C.Y.; Liu, S.; Du, Y.; Pang, K.S. The Caco-2 Cell Monolayer: Usefulness and Limitations. Expert Opin Drug Metab Toxicol 2008, 4, 395–411. [CrossRef]
- Thummel, K.E.; Brimer, C.; Yasuda, K.; Thottassery, J.; Senn, T.; Lin, Y.; Ishizuka, H.; Kharasch, E.; Schuetz, J.; Schuetz, E. Transcriptional Control of Intestinal Cytochrome P-4503A by 1α,25-Dihydroxy Vitamin D3. Mol Pharmacol 2001, 60, 1399–1406. [CrossRef]
- Minkler, S.; Lucien, F.; Kimber, M.J.; Sahoo, D.K.; Bourgois-Mochel, A.; Musser, M.; Johannes, C.; Frank, I.; Cheville, J.; Allenspach, K.; et al. Emerging Roles of Urine-Derived Components for the Management of Bladder Cancer: One Man’s Trash Is Another Man’s Treasure. Cancers 2021, 13, 422. [CrossRef]
- Shanahan, M.T.; Kanke, M.; Oyesola, O.O.; Hung, Y.H.; Koch-Laskowski, K.; Singh, A.P.; Peck, B.C.E.; Biraud, M.; Sheahan, B.; Cortes, J.E.; et al. Multiomic Analysis Defines the First MicroRNA Atlas across All Small Intestinal Epithelial Lineages and Reveals Novel Markers of Almost All Major Cell Types. Am J Physiol Gastrointest Liver Physiol 2021, 321, G668–G681. [CrossRef]
- Kopper, J.J.; Iennarella-Servantez, C.; Jergens, A.E.; Sahoo, D.K.; Guillot, E.; Bourgois-Mochel, A.; Martinez, M.N.; Allenspach, K.; Mochel, J.P. Harnessing the Biology of Canine Intestinal Organoids to Heighten Understanding of Inflammatory Bowel Disease Pathogenesis and Accelerate Drug Discovery: A One Health Approach. Frontiers in Toxicology 2021, 0, 52. [CrossRef]
- Gabriel, V.; Zdyrski, C.; Sahoo, D.K.; Dao, K.; Bourgois-Mochel, A.; Kopper, J.; Zeng, X.-L.; Estes, M.K.; Mochel, J.P.; Allenspach, K. Standardization and Maintenance of 3D Canine Hepatic and Intestinal Organoid Cultures for Use in Biomedical Research. J Vis Exp 2022. [CrossRef]
- Zdyrski, C.; Iennarella-Servantez, C.A.; Sahoo, D.K.; Ward, J.; Long, E.; Gabriel, V.; Minkler, S.; Mao, S.; Bourgois-Mochel, A.; Jergens, A.; et al. Su124 HOMOLOGY DIRECTED REPAIR IN CANINE DUODENAL ENTEROIDS TO MIMIC THE WILD-TYPE P-GLYCOPROTEIN MUTATION. Gastroenterology 2021, 160, S-625-S-626. [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 Stem Cells Build Crypt-Villus Structures in Vitro without a Mesenchymal Niche. Nature 2009, 459, 262–265. [CrossRef]
- Chandra, L.; Borcherding, D.C.; Kingsbury, D.; Atherly, T.; Ambrosini, Y.M.; Bourgois-Mochel, A.; Yuan, W.; Kimber, M.; Qi, Y.; Wang, Q.; et al. Derivation of Adult Canine Intestinal Organoids for Translational Research in Gastroenterology. BMC Biology 2019, 17, 1–21. [CrossRef]
- Ambrosini, Y.M.; Park, Y.; Jergens, A.E.; Shin, W.; Min, S.; Atherly, T.; Borcherding, D.C.; Jang, J.; Allenspach, K.; Mochel, J.P.; et al. Recapitulation of the Accessible Interface of Biopsy-Derived Canine Intestinal Organoids to Study Epithelial-Luminal Interactions. PLoS One 2020, 15, e0231423. [CrossRef]
- Gabriel, V.; Zdyrski, C.; Sahoo, D.K.; Dao, K.; Bourgois-Mochel, A.; Atherly, T.; Martinez, M.N.; Volpe, D.A.; Kopper, J.; Allenspach, K.; et al. Canine Intestinal Organoids in a Dual-Chamber Permeable Support System. JoVE (Journal of Visualized Experiments) 2022, e63612. [CrossRef]
- Sahoo, D.K.; Borcherding, D.C.; Chandra, L.; Jergens, A.E.; Atherly, T.; Bourgois-Mochel, A.; Ellinwood, N.M.; Snella, E.; Severin, A.J.; Martin, M.; et al. Differential Transcriptomic Profiles Following Stimulation with Lipopolysaccharide in Intestinal Organoids from Dogs with Inflammatory Bowel Disease and Intestinal Mast Cell Tumor. Cancers (Basel) 2022, 14, 3525. [CrossRef]
- Bedos, L.; Wickham, H.; Gabriel, V.; Zdyrski, C.; Allbaugh, R.A.; Sahoo, D.K.; Sebbag, L.; Mochel, J.P.; Allenspach, K. Culture and Characterization of Canine and Feline Corneal Epithelial Organoids: A New Tool for the Study and Treatment of Corneal Diseases. Front Vet Sci 2022, 9. [CrossRef]
- Kodama, N.; Iwao, T.; Katano, T.; Ohta, K.; Yuasa, H.; Matsunaga, T. Characteristic Analysis of Intestinal Transport in Enterocyte-Like Cells Differentiated from Human Induced Pluripotent Stem Cells. DRUG METABOLISM AND DISPOSITION Drug Metab Dispos 2016, 44, 1662–1667. [CrossRef]
- Akazawa, T.; Yoshida, S.; Ohnishi, S.; Kanazu, T.; Kawai, M.; Takahashi, K. Application of Intestinal Epithelial Cells Differentiated from Human Induced Pluripotent Stem Cells for Studies of Prodrug Hydrolysis and Drug Absorption in the Small Intestine. Drug Metab Dispos 2018, 46, 1497–1506. [CrossRef]
- Lo, B.; Parham, L. Ethical Issues in Stem Cell Research. Endocr Rev 2009, 30, 204–213. [CrossRef]
- Ergün, S.; Wörsdörfer, P. Organoids, Assembloids and Embryoids: New Avenues for Developmental Biology, Disease Modeling, Drug Testing and Toxicity Assessment without Animal Experimentation. Organoids 2022, Vol. 1, Pages 37-40 2022, 1, 37–40. [CrossRef]
- Liebau, S.; Parvin, B.; Ergün, S.; Richiardone, E.; Van Den Bossche, V.; Corbet, C. Metabolic Studies in Organoids: Current Applications, Opportunities and Challenges. Organoids 2022, 1, 85–105. [CrossRef]
- Burgess, T.; Barker, N.; Torresi, J.; Munro, M.J.; Tan, S.T.; Gray, C. Applications for Colon Organoid Models in Cancer Research. Organoids 2023, 2, 37–49. [CrossRef]
- Schutgens, F.; Rookmaaker, M.B.; Margaritis, T.; Rios, A.; Ammerlaan, C.; Jansen, J.; Gijzen, L.; Vormann, M.; Vonk, A.; Viveen, M.; et al. Tubuloids Derived from Human Adult Kidney and Urine for Personalized Disease Modeling. Nat Biotechnol 2019, 37, 303–313. [CrossRef]
- Usui, T.; Sakurai, M.; Nishikawa, S.; Umata, K.; Nemoto, Y.; Haraguchi, T.; Itamoto, K.; Mizuno, T.; Noguchi, S.; Mori, T.; et al. Establishment of a Dog Primary Prostate Cancer Organoid Using the Urine Cancer Stem Cells. Cancer Sci 2017, 108, 2383–2392. [CrossRef]
- Sun, G.; Ding, B.; Wan, M.; Chen, L.; Jackson, J.; Atala, A. Formation and Optimization of Three-Dimensional Organoids Generated from Urine-Derived Stem Cells for Renal Function in Vitro. Stem Cell Res Ther 2020, 11, 1–12. [CrossRef]
- Zhang, Y.; McNeill, E.; Tian, H.; Soker, S.; Andersson, K.E.; Yoo, J.J.; Atala, A. Urine Derived Cells Are a Potential Source for Urological Tissue Reconstruction. Journal of Urology 2008, 180, 2226–2233. [CrossRef]
- Yu, F.; Hunziker, W.; Choudhury, D. Engineering Microfluidic Organoid-on-a-Chip Platforms. Micromachines (Basel) 2019, 10. [CrossRef]
- Danku, A.E.; Dulf, E.H.; Braicu, C.; Jurj, A.; Berindan-Neagoe, I. Organ-On-A-Chip: A Survey of Technical Results and Problems. Front Bioeng Biotechnol 2022, 10, 94. [CrossRef]
- van Berlo, D.; Nguyen, V.V.T.; Gkouzioti, V.; Leineweber, K.; Verhaar, M.C.; van Balkom, B.W.M. Stem Cells, Organoids, and Organ-on-a-Chip Models for Personalized in Vitro Drug Testing. Curr Opin Toxicol 2021, 28, 7–14. [CrossRef]
- Menon-Andersen, D.; Florian, J.; Madabushi, R. CLINICAL PHARMACOLOGY REVIEW NDA Number 205410 Submission Type; Code. 2013.
- Yang, Y.; Faustino, P.J.; Volpe, D.A.; Ellison, C.D.; Lyon, R.C.; Yu, L.X. Biopharmaceutics Classification of Selected β-Blockers: Solubility and Permeability Class Membership. Mol Pharm 2007, 4, 608–614. [CrossRef]
- Martinez, M.N.; Mochel, J.P.; Neuhoff, S.; Pade, D. Comparison of Canine and Human Physiological Factors: Understanding Interspecies Differences That Impact Drug Pharmacokinetics. AAPS J 2021, 23. [CrossRef]
- Volpe, D.A.; Faustino, P.J.; Ciavarella, A.B.; Asafu-Adjaye, E.B.; Ellison, C.D.; Yu, L.X.; Hussain, A.S. Classification of Drug Permeability with a Caco-2 Cell Monolayer Assay. Ceased 2008, 24, 39–47. [CrossRef]
- Sahoo, D.K.; Roy, A.; Bhanja, S.; Chainy, G.B.N. Hypothyroidism Impairs Antioxidant Defence System and Testicular Physiology during Development and Maturation. Gen Comp Endocrinol 2008, 156. [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr Protoc Immunol 2015, 111, A3.B.1–A3.B.3. [CrossRef]
- Sahoo, D.K.; Roy, A.; Chainy, G.B.N. Protective Effects of Vitamin E and Curcumin on L-Thyroxine-Induced Rat Testicular Oxidative Stress. Chem Biol Interact 2008, 176. [CrossRef]
- Gabriel, V.; Zdyrski, C.; Sahoo, D.K.; Dao, K.; Bourgois-Mochel, A.; Atherly, T.; Martinez, M.N.; Volpe, D.A.; Kopper, J.; Allenspach, K.; et al. Canine Intestinal Organoids in a Dual-Chamber Permeable Support System. J Vis Exp 2022, 2022. [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J Lab Autom 2015, 20, 107–126.
- Wang, Y.; Bai, X.; Hu, B.; Xing, M.; Cao, Q.; Ji, A.; Song, S. Transport Mechanisms of Polymannuronic Acid and Polyguluronic Acid Across Caco-2 Cell Monolayers. Pharmaceutics 2020, 12. [CrossRef]
- Stockdale, T.P.; Challinor, V.L.; Lehmann, R.P.; de Voss, J.J.; Blanchfield, J.T. Caco-2 Monolayer Permeability and Stability of Chamaelirium Luteum (False Unicorn) Open-Chain Steroidal Saponins. 2019. [CrossRef]
- Frost, T.S.; Jiang, L.; Lynch, R.M.; Zohar, Y. Permeability of Epithelial/Endothelial Barriers in Transwells and Microfluidic Bilayer Devices. Micromachines 2019, 10, 533. [CrossRef]
- PubChem Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 22 December 2022).
- Falavigna, M.; Stein, P.C.; Flaten, G.E.; di Cagno, M.P. Impact of Mucin on Drug Diffusion: Development of a Straightforward In Vitro Method for the Determination of Drug Diffusivity in the Presence of Mucin. Pharmaceutics 2020, 12. [CrossRef]
- Kulkarni, U.D.; Mahalingam, R.; Li, X.; Pather, I.; Jasti, B. Effect of Experimental Temperature on the Permeation of Model Diffusants Across Porcine Buccal Mucosa. AAPS PharmSciTech 2011, 12, 579. [CrossRef]
- Flaten, G.E.; Dhanikula, A.B.; Luthman, K.; Brandl, M. Drug Permeability across a Phospholipid Vesicle Based Barrier: A Novel Approach for Studying Passive Diffusion. Eur J Pharm Sci 2006, 27, 80–90. [CrossRef]
- Caco-2 Permeability Assay Available online: https://www.cyprotex.com/admepk/in-vitro-permeability/caco-2-permeability (accessed on 13 March 2022).
- In Vitro Drug Interaction Studies-Cytochrome P450 Enzyme-and Transporter-Mediated Drug Interactions Guidance for Industry. 2020.
- Katneni, K.; Pham, T.; Saunders, J.; Chen, G.; Patil, R.; White, K.L.; Abla, N.; Chiu, F.C.K.; Shackleford, D.M.; Charman, S.A. Using Human Plasma as an Assay Medium in Caco-2 Studies Improves Mass Balance for Lipophilic Compounds. [CrossRef]
- Heinlein, A.; Metzger, M.; Walles, H.; Buettner, A. Transport of Hop Aroma Compounds across Caco-2 Monolayers. Food Funct 2014, 5, 2719. [CrossRef]
- Quan, Y.; Jin, Y.; Faria, T.N.; Tilford, C.A.; He, A.; Wall, D.A.; Smith, R.L.; Vig, B.S. Expression Profile of Drug and Nutrient Absorption Related Genes in Madin-Darby Canine Kidney (MDCK) Cells Grown under Differentiation Conditions. Pharmaceutics 2012, 4, 314–333. [CrossRef]
- Almeida, L.D.; Quaglio, A.E.V.; De Almeida Costa, C.A.R.; Di Stasi, L.C. Intestinal Anti-Inflammatory Activity of Ground Cherry (Physalis Angulata L.) Standardized CO2 Phytopharmaceutical Preparation. World J Gastroenterol 2017, 23, 4369–4380. [CrossRef]
- Youhanna, S.; Lauschke, V.M. The Past, Present and Future of Intestinal In Vitro Cell Systems for Drug Absorption Studies. J Pharm Sci 2021, 110, 50–65. [CrossRef]
- He, Y.L.; Murby, S.; Warhurst, G.; Gifford, L.; Walker, D.; Ayrton, J.; Eastmond, R.; Rowland, M. Species Differences in Size Discrimination in the Paracellular Pathway Reflected by Oral Bioavailability of Poly(Ethylene Glycol) and D-Peptides. J Pharm Sci 1998, 87, 626–633. [CrossRef]
- Incecayir, T.; Tsume, Y.; Amidon, G.L. Comparison of the Permeability of Metoprolol and Labetalol in Rat, Mouse, and Caco-2 Cells: Use as a Reference Standard for BCS Classification. Mol Pharm 2013, 10, 958–966. [CrossRef]
- Zur, M.; Gasparini, M.; Wolk, O.; Amidon, G.L.; Dahan, A. The Low/High BCS Permeability Class Boundary: Physicochemical Comparison of Metoprolol and Labetalol. Mol Pharm 2014, 11, 1707–1714. [CrossRef]
- Dahlgren, D.; Roos, C.; Sjögren, E.; Lennernäs, H. Direct In Vivo Human Intestinal Permeability (Peff) Determined with Different Clinical Perfusion and Intubation Methods. J Pharm Sci 2015, 104, 2702–2726. [CrossRef]
- Dahlgren, D.; Roos, C.; Johansson, P.; Lundqvist, A.; Tannergren, C.; Abrahamsson, B.; Sjögren, E.; Lennernäs, H. Regional Intestinal Permeability in Dogs: Biopharmaceutical Aspects for Development of Oral Modified-Release Dosage Forms. Mol Pharm 2016, 13, 3022–3033. [CrossRef]
- Machen, T.E.; Erlij, D.; Wooding, F.B.P. PERMEABLE JUNCTIONAL COMPLEXES : The Movement of Lanthanum across Rabbit Gallbladder and Intestine. J Cell Biol 1972, 54, 302. [CrossRef]
- Frömter, E.; Diamond, J. Route of Passive Ion Permeation in Epithelia. Nat New Biol 1972, 235, 9–13. [CrossRef]
- Monaco, A.; Ovryn, B.; Axis, J.; Amsler, K. The Epithelial Cell Leak Pathway. Int J Mol Sci 2021, 22. [CrossRef]
- Amidon, G.; Lee, P.; Topp, E. Transport Processes in Pharmaceutical Systems; 1999;
- Saaby, L.; Helms, H.C.C.; Brodin, B. IPEC-J2 MDR1, a Novel High-Resistance Cell Line with Functional Expression of Human P-Glycoprotein (ABCB1) for Drug Screening Studies. Mol Pharm 2016, 13, 640–652. [CrossRef]
- Dahlgren, D.; Lennernäs, H. Intestinal Permeability and Drug Absorption: Predictive Experimental, Computational and In Vivo Approaches. Pharmaceutics 2019, 11. [CrossRef]
- Li, J.; Volpe, D.A.; Wang, Y.; Zhang, W.; Bode, C.; Owen, A.; Hidalgo, I.J. Use of Transporter Knockdown Caco-2 Cells to Investigate the In Vitro Efflux of Statin Drugs. Drug Metabolism and Disposition 2011, 39, 1196–1202. [CrossRef]
- Meneses, A.M.C.; Schneeberger, K.; Kruitwagen, H.S.; Penning, L.C.; van Steenbeek, F.G.; Burgener, I.A.; Spee, B. Intestinal Organoids—Current and Future Applications. Veterinary Sciences 2016, 3, 31. [CrossRef]
- Reineking, W.; Schauerte, I.E.; Junginger, J.; Hewicker-Trautwein, M. Sox9, Hopx, and Survivin and Tuft Cell Marker DCLK1 Expression in Normal Canine Intestine and in Intestinal Adenoma and Adenocarcinoma. Vet Pathol 2022, 59, 415–426. [CrossRef]
- Karim, B.O.; Rhee, K.J.; Liu, G.; Yun, K.; Brant, S.R. Prom1 Function in Development, Intestinal Inflammation, and Intestinal Tumorigenesis. Front Oncol 2014, 4, 323. [CrossRef]
- Wang, X.Y.; Chen, S.H.; Zhang, Y.N.; Xu, C.F. Olfactomedin-4 in Digestive Diseases: A Mini-Review. World J Gastroenterol 2018, 24, 1881. [CrossRef]
Figure 1.
Summary diagram of the transport of a therapeutic drug through the intestine facilitated by transcellular transport, carrier-mediated (CM) and passive diffusion pathways such as passive lipoidal diffusion, CM influx, CM efflux, paracellular diffusion, endocytosis, and transcytosis. The figure was produced with BioRender.com.
Figure 1.
Summary diagram of the transport of a therapeutic drug through the intestine facilitated by transcellular transport, carrier-mediated (CM) and passive diffusion pathways such as passive lipoidal diffusion, CM influx, CM efflux, paracellular diffusion, endocytosis, and transcytosis. The figure was produced with BioRender.com.
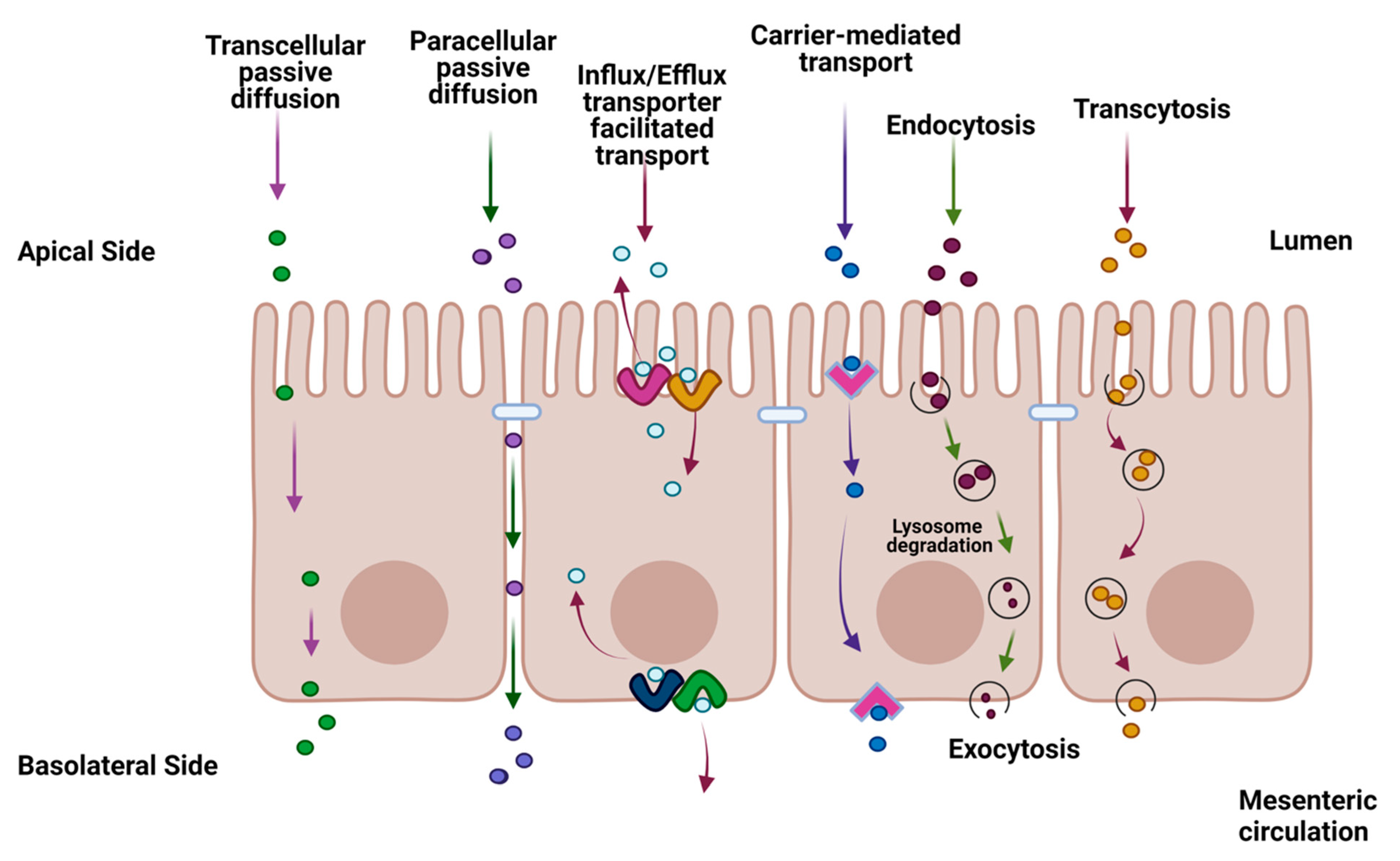
Figure 2.
Representative diagram of a dual-chamber culture apparatus showing a cell monolayer grown on filter membrane with apical and basolateral chambers and validation of monolayer integrity by transepithelial electrical resistance (TEER) measurement. Absorption is measured by adding drug to the apical chamber and measuring its appearance in the basolateral chamber over time. Conversely, secretion (efflux) is measured by adding drug to the basolateral chamber and measuring its appearance in the apical chamber over time. The figure was produced with BioRender.com.
Figure 2.
Representative diagram of a dual-chamber culture apparatus showing a cell monolayer grown on filter membrane with apical and basolateral chambers and validation of monolayer integrity by transepithelial electrical resistance (TEER) measurement. Absorption is measured by adding drug to the apical chamber and measuring its appearance in the basolateral chamber over time. Conversely, secretion (efflux) is measured by adding drug to the basolateral chamber and measuring its appearance in the apical chamber over time. The figure was produced with BioRender.com.
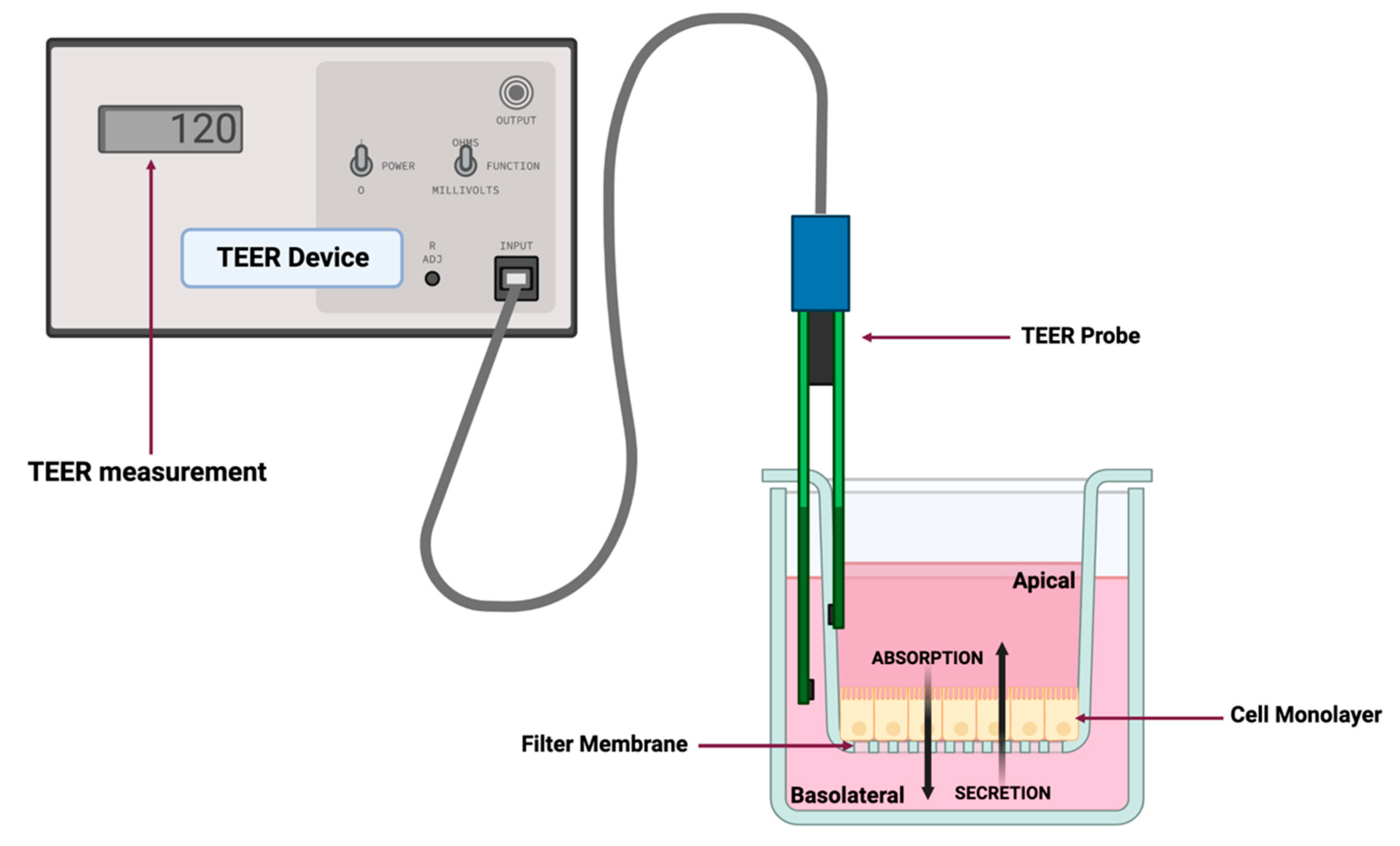
Figure 3.
Formation of the canine colonoid-derived monolayer. (A) Representative phase-contrast micrograph of a differentiated colonoid on Day 5 used for monolayer preparation. Scale Bar, 500 μm. (B) Representative phase-contrast micrographs of colonoid-derived monolayers on Days 3, 9, and 11 and representative micrographs of eosin-stained colonoid-derived monolayers on Day 11. Scale Bar, as indicated.
Figure 3.
Formation of the canine colonoid-derived monolayer. (A) Representative phase-contrast micrograph of a differentiated colonoid on Day 5 used for monolayer preparation. Scale Bar, 500 μm. (B) Representative phase-contrast micrographs of colonoid-derived monolayers on Days 3, 9, and 11 and representative micrographs of eosin-stained colonoid-derived monolayers on Day 11. Scale Bar, as indicated.

Figure 4.
Evaluation of canine colonoid-derived monolayer integrity. Continuous TEER analysis over 15 days was used to assess the integrity of each colonoid monolayer. Monolayers were prepared for propranolol, atenolol, and metoprolol studies. Values are expressed as the arithmetic mean of the data and one standard deviation of N = 18 monolayers.
Figure 4.
Evaluation of canine colonoid-derived monolayer integrity. Continuous TEER analysis over 15 days was used to assess the integrity of each colonoid monolayer. Monolayers were prepared for propranolol, atenolol, and metoprolol studies. Values are expressed as the arithmetic mean of the data and one standard deviation of N = 18 monolayers.
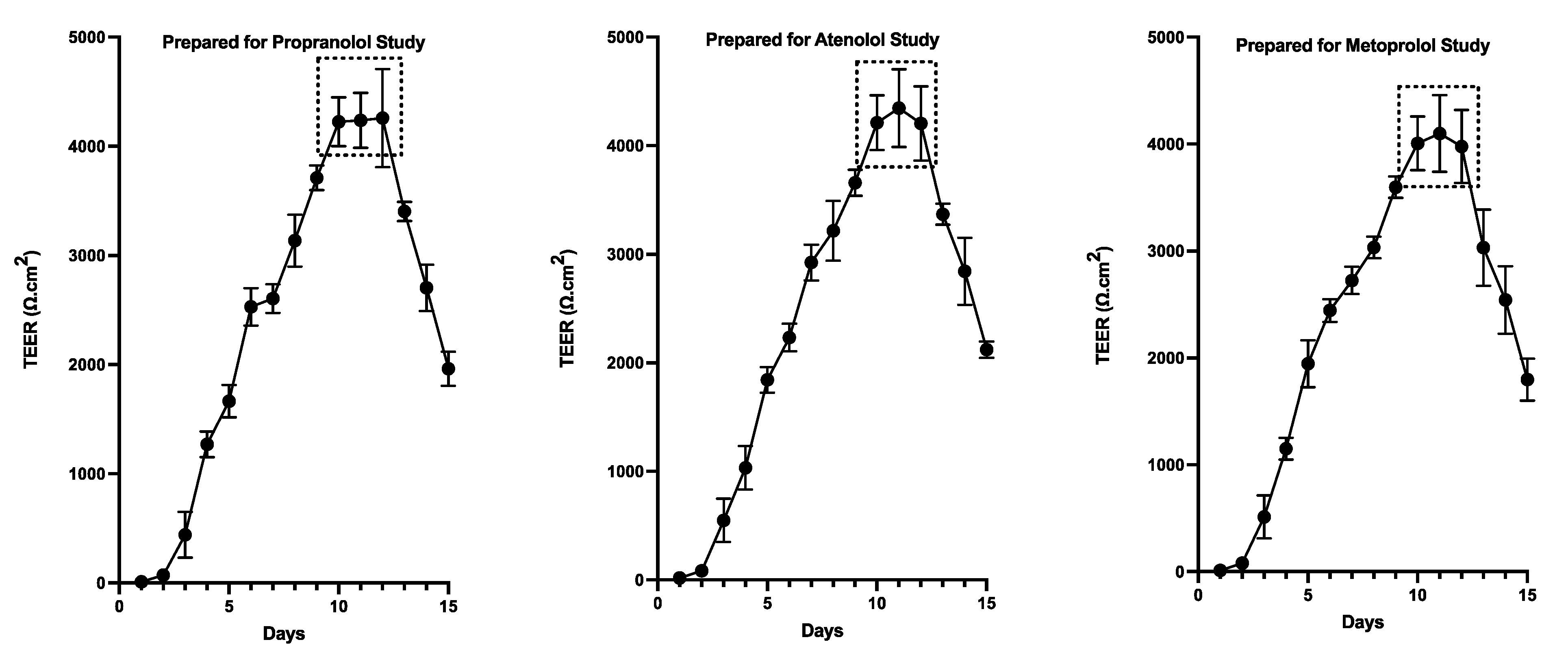
Figure 5.
Evaluation of Caco-2 cell monolayer integrity. Continuous TEER analysis of the monolayer over 23 days was used to assess the permeability of propranolol, atenolol, and metoprolol. Values are expressed as the arithmetic mean of the data and one standard deviation of N = 12 monolayers.
Figure 5.
Evaluation of Caco-2 cell monolayer integrity. Continuous TEER analysis of the monolayer over 23 days was used to assess the permeability of propranolol, atenolol, and metoprolol. Values are expressed as the arithmetic mean of the data and one standard deviation of N = 12 monolayers.
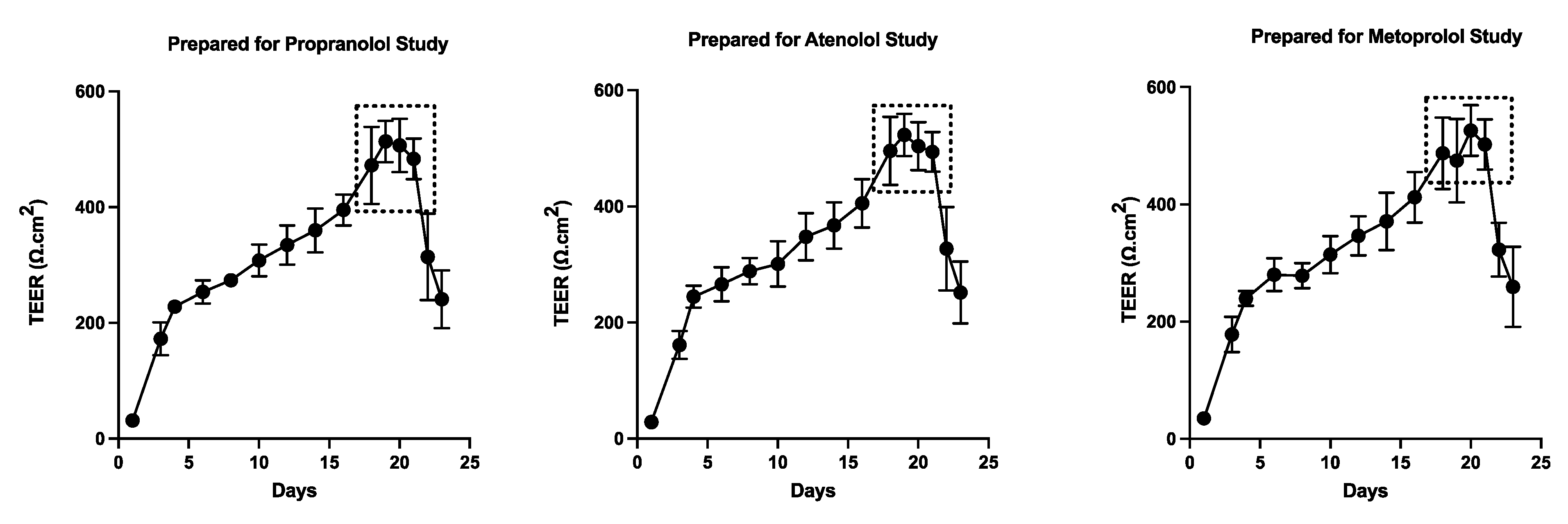
Figure 7.
Expression of tight and adherens junction proteins, intestinal epithelial cell differentiation and stem cell markers in canine colonoid-derived monolayers. The gene expression results are displayed as normalized Ct (dCt) to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (see Supplemental Table 4 for Ct values). Values are expressed as the arithmetic mean of the data and one standard deviation of N = 3 monolayers. Cadherin 1 (CDH1); occludin (OCLN); tight junction protein 1 (TJP1); mucin 2 (MUC2); neurogenin 3 (NEUROG3); intestinal alkaline phosphatase (ALP); olfactomedin 4 (OLFM4); HOP homeobox (HOPX); prominin 1 (PROM1); SRY-box transcription factor 9 (SOX9); leucine-rich repeat-containing G protein-coupled receptor 5 (LGR5).
Figure 7.
Expression of tight and adherens junction proteins, intestinal epithelial cell differentiation and stem cell markers in canine colonoid-derived monolayers. The gene expression results are displayed as normalized Ct (dCt) to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (see Supplemental Table 4 for Ct values). Values are expressed as the arithmetic mean of the data and one standard deviation of N = 3 monolayers. Cadherin 1 (CDH1); occludin (OCLN); tight junction protein 1 (TJP1); mucin 2 (MUC2); neurogenin 3 (NEUROG3); intestinal alkaline phosphatase (ALP); olfactomedin 4 (OLFM4); HOP homeobox (HOPX); prominin 1 (PROM1); SRY-box transcription factor 9 (SOX9); leucine-rich repeat-containing G protein-coupled receptor 5 (LGR5).
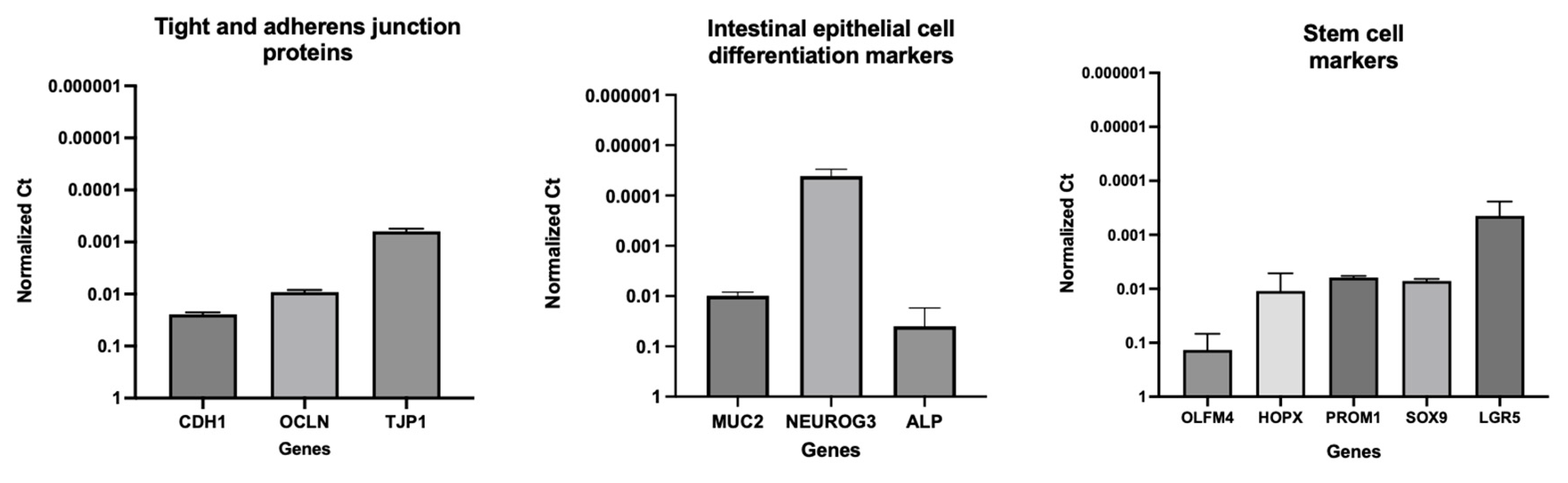
Figure 8.
Visualization of goblet cells containing mucus in canine colon tissue and colonoids. (A) Histological images of hematoxylin and eosin (H&E) staining of colon tissue. (B) Alcian blue staining of colon tissue reveal goblet cells containing mucus. (C) The H&E staining of colonoids at 5 days after passage. (D) Colonoids at 5 days after passage with mucus production are shown by Alcian blue staining. (E) A representative transmission electron microscopy (TEM) image of colonoid shows a goblet cell with mucin granules. (F) A TEM image of the intercellular junctional complex in canine colonoid. The microvilli of the canine colonoid are visible at higher magnifications using TEM than with a standard optical light microscope [74]. (G) A zoom-in that shows a high-power magnification of the red dashed area in “F”.
Figure 8.
Visualization of goblet cells containing mucus in canine colon tissue and colonoids. (A) Histological images of hematoxylin and eosin (H&E) staining of colon tissue. (B) Alcian blue staining of colon tissue reveal goblet cells containing mucus. (C) The H&E staining of colonoids at 5 days after passage. (D) Colonoids at 5 days after passage with mucus production are shown by Alcian blue staining. (E) A representative transmission electron microscopy (TEM) image of colonoid shows a goblet cell with mucin granules. (F) A TEM image of the intercellular junctional complex in canine colonoid. The microvilli of the canine colonoid are visible at higher magnifications using TEM than with a standard optical light microscope [74]. (G) A zoom-in that shows a high-power magnification of the red dashed area in “F”.
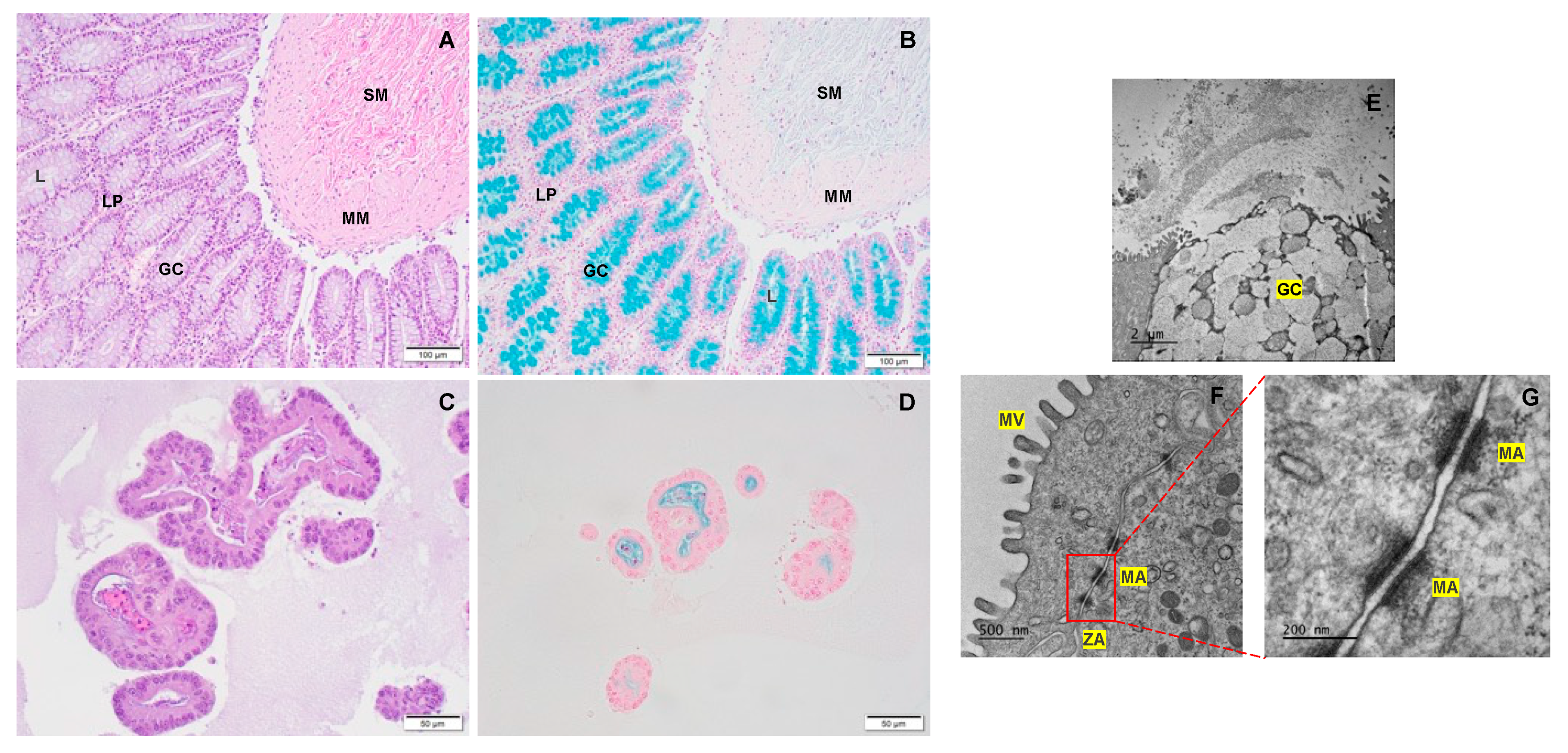
Figure 9.
Comparative expression of transporters in Caco-2-derived and canine colonoid-derived monolayers. Organic anion transporting polypeptide (OATP2B1, human; Oatp2b1, dog); multidrug resistance p-glycoprotein (MDR1, human; Mdr1, dog).
Figure 9.
Comparative expression of transporters in Caco-2-derived and canine colonoid-derived monolayers. Organic anion transporting polypeptide (OATP2B1, human; Oatp2b1, dog); multidrug resistance p-glycoprotein (MDR1, human; Mdr1, dog).
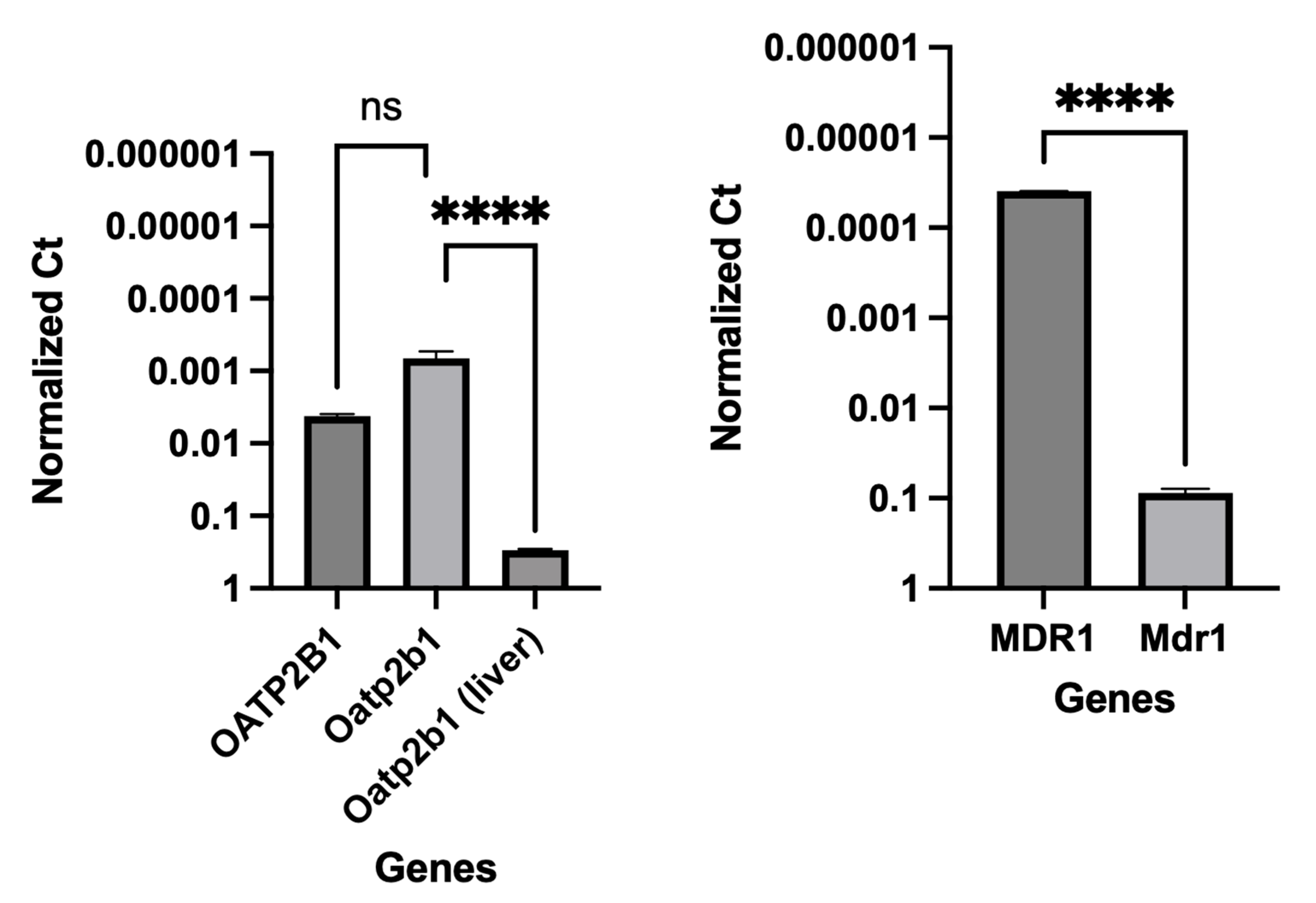
Figure 10.
Comparative expression of CYP enzymes in Caco-2-derived and their orthologs in canine colonoid-derived monolayers. Gene expression results are displayed as normalized Ct (dCt) to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (see Supplemental Table 4 for Ct values). Values are expressed as the arithmetic mean of the data and one standard deviation of N = 3 monolayers. CYP enzymes in human Caco-2 (CYP3A4, CYP2B6, CYP2C9 and CYP2C19); CYP enzymes in canine colonoids (Cyp3a12, Cyp2b11 and Cyp2c21). Group comparisons were performed with GraphPad Prism 9 using a two-tailed Student’s t-test. P-values < 0.05 (*) were considered as statistically significant for all analyses. **** P < 0.0001. ns = no significance difference.
Figure 10.
Comparative expression of CYP enzymes in Caco-2-derived and their orthologs in canine colonoid-derived monolayers. Gene expression results are displayed as normalized Ct (dCt) to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (see Supplemental Table 4 for Ct values). Values are expressed as the arithmetic mean of the data and one standard deviation of N = 3 monolayers. CYP enzymes in human Caco-2 (CYP3A4, CYP2B6, CYP2C9 and CYP2C19); CYP enzymes in canine colonoids (Cyp3a12, Cyp2b11 and Cyp2c21). Group comparisons were performed with GraphPad Prism 9 using a two-tailed Student’s t-test. P-values < 0.05 (*) were considered as statistically significant for all analyses. **** P < 0.0001. ns = no significance difference.
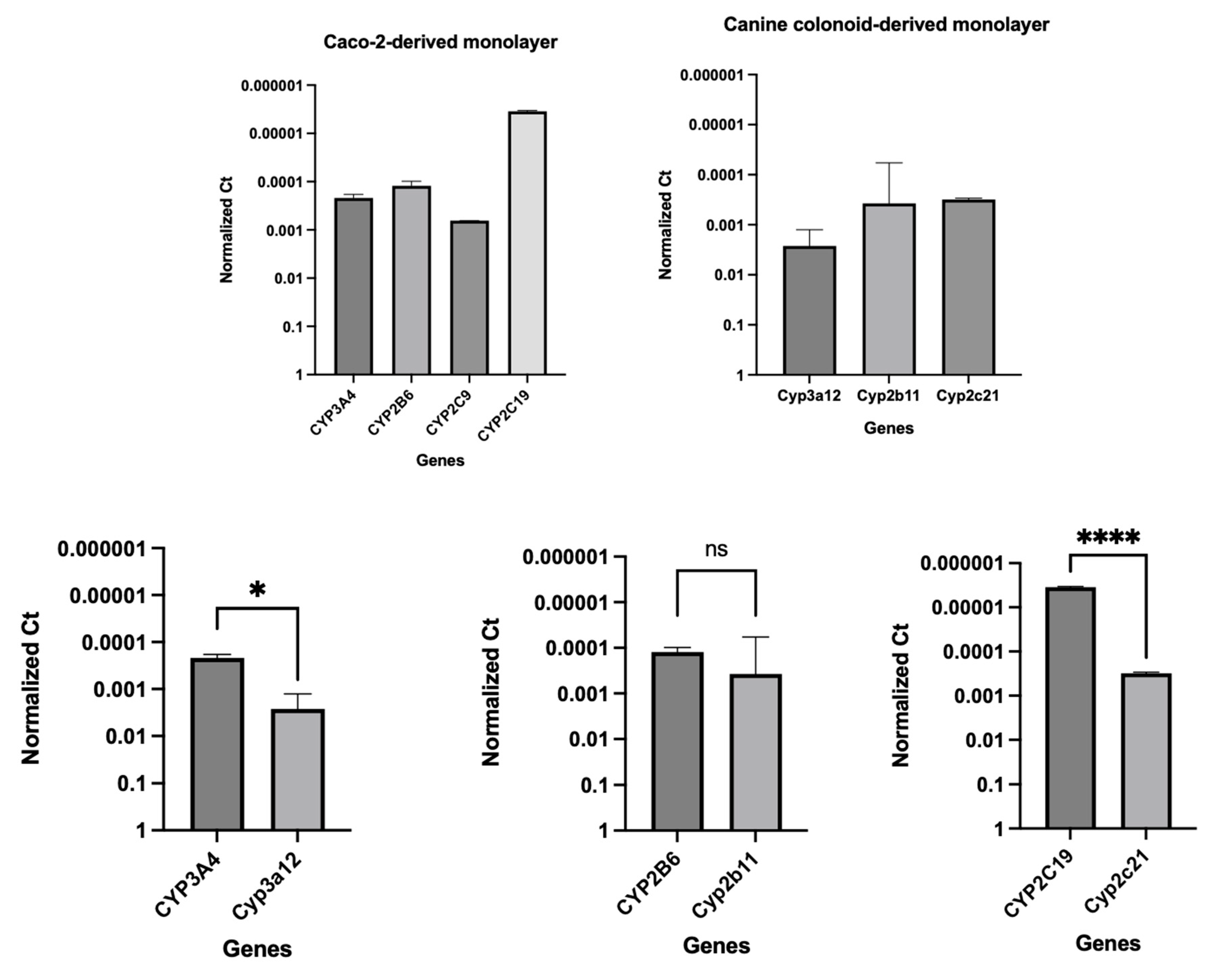

Table 1.
Primers used for real-time PCR expression of tight and adherens junction proteins, intestinal epithelial cell markers, transporters, and cytochrome P450 (CYP) enzymes.
Table 1.
Primers used for real-time PCR expression of tight and adherens junction proteins, intestinal epithelial cell markers, transporters, and cytochrome P450 (CYP) enzymes.
| Species | Category | Gene Full Name | Symbol | Forward Primer Sequence (5'-3') | Reverse Primer Sequence (5'-3') |
|---|---|---|---|---|---|
| Canis lupus familiaris (dog) | Housekeeping gene | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | TCAACGGATTTGGCCGTATTGG | TGAAGGGGTCATTGATGGCG |
| Tight and adherens junction proteins | Cadherin 1 | CDH1 | GACCCAGTAACTAACGACG | CTTCATTCACATCTTCCACG | |
| Occludin | OCLN | CACTACTGTGTGGTGGATCC | CCTTGTCCCACAATATATTCG | ||
| Tight junction protein 1 | TJP1 | GAGGGTGATCAAATTCTCAGG | CTGATTCTACAATGCGACG | ||
| Intestinal epithelial cell differentiation markers | Mucin 2 | MUC2 | CCTGTGCCCCATATTCTGC | GAGATGTTGGAATGGATGCC | |
| Neurogenin 3 | NEUROG3 | GAATGCACAACCTCAACTCG | GTAGAGGCTGTGGTCCGC | ||
| Intestinal alkaline phosphatase | ALP | CGTAGTAAACCGCAACTGG | GGAAACATGTACTTTCGGC | ||
| Stem cell markers | Olfactomedin 4 | OLFM4 | GTATCATGAATGTCAGCAAGC | CTGTAATATTCCAGAATTCTTCC | |
| HOP homeobox | HOPX | GACCAGGTGGAGATTCTGG | GCCAGACGCTGCTTAAACC | ||
| Prominin 1 | PROM1 | GATTATTATTTGTGCTGTCC | GAGACTGTAAAGTATTTCCTC | ||
| SRY-box transcription factor 9 | SOX9 | GTCATCTCCAACATAGAGACC | CTGCTTGGACATCCACACG | ||
| Leucine-rich repeat-containing G protein-coupled receptor 5 | LGR5 | GCTAGATCTGTCTTACAACC | GTTCCAGGCTAAATTCAGC | ||
| Transporters | Organic anion transporting polypeptide | Oatp2b1 | GATGACTTTGCCCACAACAGC | CAGCAGCAGAGATGAGGAAGC | |
| Multidrug resistance p-glycoprotein | Mdr1 | GTAGCTGAAGAAGTCTTAGCAGC | GCGGCACCAATAGAAATGTTGGC | ||
| Cytochrome P450 (CYP) enzymes | Cytochrome P-450 3a12 | Cyp3a12 | GATCATGAACATGAAACTTGC | CTTTTCAGGTTGAATAATCCC | |
| Cytochrome P450 2b11 | Cyp2b11 | CTGAGGGAGTCCTCCAGGACCC | CACATAGAACAAGTTCATCAGG | ||
| Cytochrome P450 2C21 | Cyp2c21 (Cyp2c18) | CAAGCACCTCCTGGATACAGC | CTTCGTGTTCTTTTATTTTTTCC | ||
| Homo sapiens (human) | Housekeeping gene | Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | TGCACCACCAACTGCTTAGC | GGCATGGACTGTGGTCATGAG |
| Transporters | Organic anion transporting polypeptide 2B1/ solute carrier organic anion transporter family member 2B1 | OATP2B1 (SLCO2B1) | CAAACCTGACTGTGATCCAG | GAGCAGGTTGGCGTATGAGG | |
| ATP binding cassette subfamily B member 1 | ABCB1 (MDR1) | CAGTAGCTGAAGAGGTCTTGGC | CTGTAATAGCTTTCTTTATCCC | ||
| Cytochrome P450 (CYP) enzymes | Cytochrome P450 family 3 subfamily A member 4 | CYP3A4 | GAGATGGTCCCTATCATTGCC | GATGTTCACTCCAAATGATGTGC | |
| Cytochrome P450 family 2 subfamily B member 6 | CYP2B6 | GAAACCGCTGGAAGGTGCTTCG | CTCCTCTATCAGACACTGAGC | ||
| Cytochrome P450 family 2 subfamily C member 9 | CYP2C9 | GAAGGAGATCCGGCGTTTCTCC | CTTGGTTTTTCTCAACTCCTCC | ||
| Cytochrome P450 family 2 subfamily C member 19 | CYP2C19 | GATCTGCTCCATTATTTTCC | GTTTTTAAGTAATTTGTTATGG |
Table 2.
Physicochemical characteristics of metoprolol, atenolol, and propranolol.
| Drug | Molecular Formula (MF) | Molecular Weight (MW) | Aqueous solubility (25°C) | Log P | Dissociation Constants (Basic pKa) |
|---|---|---|---|---|---|
| Metoprolol | C15H25NO3 | 267.36 g/mol | >1000 mg/mL | 2.15 | 9.56 |
| Atenolol | C14H22N2O3 | 266.34 g/mol | 13.3 mg/mL | 0.16 | 9.58 |
| Propranolol | C16H21NO2 | 259.339 g/mol | 0.0617 mg/L | 3.48 | 9.53 |
Table 3.
Transport of FITC–dextran (expressed in percentage) across the colonoid-derived monolayer. Values are expressed as the arithmetic mean of the data and one standard deviation of 3 monolayers.
Table 3.
Transport of FITC–dextran (expressed in percentage) across the colonoid-derived monolayer. Values are expressed as the arithmetic mean of the data and one standard deviation of 3 monolayers.
| Compound | Time (min) | Donor (%) | Receiver (%) |
|---|---|---|---|
| FITC-Dextran (200 µg/mL) | 0 | 100 ± 0.0 | 0.020 ± 0.001 |
| 15 | 0.021 ± 0.003 | ||
| 30 | 0.024 ± 0.004 | ||
| 45 | 0.022 ± 0.002 | ||
| 60 | 0.024 ± 0.003 | ||
| 90 | 0.023 ± 0.004 | ||
| 120 | 93.7 ± 7.8 | 0.022 ± 0.002 | |
| Metoprolol (0.4 mg/mL) + | 0 | 100 ± 0.0 | 0.021 ± 0.001 |
| FITC-Dextran (200 µg/mL) | 15 | 0.023 ± 0.001 | |
| 30 | 0.020 ± 0.00 | ||
| 45 | 0.020 ± 0.00 | ||
| 60 | 0.022 ± 0.001 | ||
| 90 | 0.023 ± 0.00 | ||
| 120 | 94.6 ± 1.4 | 0.020 ± 0.001 |
Table 4.
Adjusted Papp value estimates as a function of drug/concentration, direction, experimental pH and the cell culture system. BLQ = below the analytical limit of quantification. *Metoprolol (0.4 mg/mL) ± FITC-dextran (200 µg/mL) testing was exclusively performed in canine colonoid monolayers. Values presented herein were aggregated from both Metoprolol (0.4 mg/mL) ± FITC-dextran (200 µg/mL) experiments. Transport of metoprolol, atenolol and propranolol across the Caco-2-derived (human) and canine colonoid-derived (dog) monolayers. BL→AP and AP→BL transports of the drugs were studied in two different pH conditions i.e., 6.8 and 7.4 (AP site). Two different monolayers (N = 2) were used for each system for specific directions and pH conditions (for dog: AP→BL, pH 6.8/7.4 or BL→AP, pH 7.4/7.4 or AP→BL, pH 7.4/7.4 and for human: AP→BL, pH 6.8/7.4 or BL→AP, pH 7.4/7.4). In total, 20 Caco-2 monolayers and 34 colonoid monolayers were used throughout the study.
Table 4.
Adjusted Papp value estimates as a function of drug/concentration, direction, experimental pH and the cell culture system. BLQ = below the analytical limit of quantification. *Metoprolol (0.4 mg/mL) ± FITC-dextran (200 µg/mL) testing was exclusively performed in canine colonoid monolayers. Values presented herein were aggregated from both Metoprolol (0.4 mg/mL) ± FITC-dextran (200 µg/mL) experiments. Transport of metoprolol, atenolol and propranolol across the Caco-2-derived (human) and canine colonoid-derived (dog) monolayers. BL→AP and AP→BL transports of the drugs were studied in two different pH conditions i.e., 6.8 and 7.4 (AP site). Two different monolayers (N = 2) were used for each system for specific directions and pH conditions (for dog: AP→BL, pH 6.8/7.4 or BL→AP, pH 7.4/7.4 or AP→BL, pH 7.4/7.4 and for human: AP→BL, pH 6.8/7.4 or BL→AP, pH 7.4/7.4). In total, 20 Caco-2 monolayers and 34 colonoid monolayers were used throughout the study.
| Drug | Species | Direction | Well | Papp × 10-6 | Avg | SD | %CV |
|---|---|---|---|---|---|---|---|
| Metoprolol (0.4 mg/mL or 584.1 μM) |
Human | AP→BL pH 6.8/7.4 | 1 | 19.34 | |||
| 2 | 16.45 | 17.89 | 2.05 | 11.43 | |||
| BL→AP pH 7.4/7.4 | 1 | 13.79 | |||||
| 2 | 12.37 | 13.08 | 1.01 | 7.72 | |||
| Dog* | AP→BL pH 6.8/7.4 | 1 | 8.33 | ||||
| 2 | 9.10 | 8.72 | 0.55 | 6.32 | |||
| AP→BL pH 7.4/7.4 | 1 | 11.19 | |||||
| 2 | 9.90 | 10.54 | 0.91 | 8.66 | |||
| BL→AP pH 7.4/7.4 | 1 | 9.88 | |||||
| 2 | 9.94 | 9.91 | 0.05 | 0.46 | |||
| Metoprolol (0.04 mg/mL or 58.4 μM) |
Human | AP→BL pH 6.8/7.4 | 1 | 17.54 | |||
| 2 | 13.13 | 15.33 | 3.12 | 20.38 | |||
| BL→AP pH 7.4/7.4 | 1 | 16.30 | |||||
| 2 | 19.03 | 17.67 | 1.93 | 10.92 | |||
| Dog | AP→BL pH 6.8/7.4 | 1 | 16.83 | ||||
| 2 | 18.52 | 17.68 | 1.19 | 6.75 | |||
| AP→BL pH 7.4/7.4 | 1 | 13.44 | |||||
| 2 | 13.50 | 13.47 | 0.04 | 0.32 | |||
| BL→AP pH 7.4/7.4 | 1 | 11.19 | |||||
| 2 | 12.67 | 11.93 | 1.04 | 8.76 | |||
| Atenolol (0.2 mg/mL or 750.9 μM) |
Human | AP→BL pH 6.8/7.4 | 1 | BLQ | |||
| 2 | BLQ | ||||||
| BL→AP pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| Dog | AP→BL pH 6.8/7.4 | 1 | BLQ | ||||
| 2 | BLQ | ||||||
| AP→BL pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| BL→AP pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| Atenolol (2 mg/mL or 7,509 μM) |
Human | AP→BL pH 6.8/7.4 | 1 | 7.25 | |||
| 2 | 8.01 | 7.63 | 0.54 | 7.09 | |||
| BL→AP pH 7.4/7.4 | 1 | 3.69 | |||||
| 2 | 4.97 | 4.33 | 0.90 | 20.89 | |||
| Dog | AP→BL pH 6.8/7.4 | 1 | BLQ | ||||
| 2 | BLQ | ||||||
| AP→BL pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| BL→AP pH 7.4/7.4 | 1 | BLQ | |||||
| 2 | BLQ | ||||||
| Propranolol (1 mg/mL or 3,380 μM) |
Human | AP→BL pH 6.8/7.4 | 1 | 12.12 | |||
| 2 | 12.75 | 12.44 | 0.45 | 3.59 | |||
| BL→AP pH 7.4/7.4 | 1 | 11.45 | |||||
| 2 | 12.43 | 11.94 | 0.70 | 5.84 | |||
| Dog | AP→BL pH 6.8/7.4 | 1 | 12.23 | ||||
| 2 | 11.05 | 11.64 | 0.83 | 7.15 | |||
| AP→BL pH 7.4/7.4 | 1 | 11.99 | |||||
| 2 | 11.97 | 11.98 | 0.01 | 0.09 | |||
| BL→AP pH 7.4/7.4 | 1 | 10.94 | |||||
| 2 | 12.03 | 11.49 | 0.77 | 6.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



