
Submitted:
30 March 2023
Posted:
11 April 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Background: Chronic neurologic diseases are common sequelae of COVID. They severely impact the quality of life and increase the burden on healthcare systems. The long COVID neurological symptoms are due to the robust replication of SARS-CoV-2 in the nasal neuroepithelial cells, leading to neuroinvasion and inflammation of the central nerve system (CNS). Currently used medications and vaccines do not inhibit the robust SARS-CoV-2 replication in the nasal epithelial cells. EGCG-palmitate (EC16), a multifunctional compound, has the potential to become a novel intranasal-delivered drug for minimizing post-COVID neurologic symptoms. Method: EC16-containing formulations were developed and tested in vitro against human β coronavirus OC43 (CoV-OC43) using a TCID50 assay following three test protocols differing in exposure sequence. Results: EC16 formulations in normal saline, phosphate buffered saline, and cell culture medium were found to effectively inhibit human β-coronavirus infection (>99.99%) after a 30-min contact. A single 10-min application to cells after infection (i.e., without direct contact with the virus) resulted in >99% inhibition of viral replication. Conclusion: With its antiviral, antioxidant, anti-inflammatory, and neuroprotective properties, EC16 in nasal formulations could be further developed for clinical applications to COVID-19 patients for minimizing long COVID neurological symptoms.
Keywords:
COVID-19
; Long COVID
; EC16
; EGCG-palmitate
; Formulations
1. Introduction
Following recovery from the acute infection stage of the SARS-CoV-2 virus (COVID-19), survivors can experience a wide range of persistent post-acute sequelae referred to as long COVID (PASC). It has been estimated that 50% of COVID-19 survivors developed a broad array of pulmonary and extrapulmonary clinical manifestations, including nervous system and neurocognitive disorders, which include headache, persistent loss of smell and/or taste, memory loss, brain fog (difficulty concentrating, sense of confusion or disorientation), dizziness, anxiety, depression, earache, hearing loss, and/or ringing in ears (tinnitus) [1]. According to the US National Research Action Plan on Long COVID, 2022, up to 23.7 million Americans suffer from long COVID, and approximately one million workers may be out of the workforce each day due to these symptoms, leading to a $50 billion annual loss of salary.
The post-COVID chronic neurologic symptoms have been shown to be due to the robust replication of SARS-CoV-2 in the nasal neuroepithelial cells, leading to neuroinvasion and inflammation of the central nervous system (CNS) [2].
Aerosol transmission of SARS-CoV-2 is a significant route for viral entry to infect humans [3]. Similar to other respiratory viruses, the nasal epithelia are the major initial site of entry for SARS-CoV-2 prior to spreading to upper respiratory tissues and invasion of the CNS. Specifically, multiciliated cells in the nasal respiratory epithelium serve as a reservoir for SARS-CoV-2 replication [3,4]. Newly published evidence confirms that “respiratory viruses, including SARSCoV-2, bypass the defensive mucus/mucin layer of the airway by entering and exiting epithelial cells via their protruding motile cilia and microvilli” [5].
Currently used medications and vaccines do not target (at least directly) the neuroinvasion of SARS-CoV-2, and these methods do not inhibit the robust SARS-CoV-2 replication in the nasal epithelial cells. Therefore, a significant gap exists in treatment/prevention strategies that needs to be filled by methods to rapidly inhibit SARS-CoV-2 replication in the nasal cavity and block viral invasion of the CNS, in order to minimize neurologic damage. A known agent with antiviral, anti-inflammatory, antioxidant, and neuroprotective properties and able to rapidly inhibit viral replication in nasal epithelia would be a candidate for such a method.
One of the new drug candidate agents is epigallocatechin-3-gallate-palmitate (EGCG-palmitate or EC16), a stable lipid-soluble form of EGCG with broad virucidal, antiviral, anti-inflammatory, antioxidant, and neuroprotective properties that has significant potential for minimizing long COVID [6-11]. We reported previously that EC16 is able to effectively inhibit influenza virus, norovirus, and herpes simplex virus [12-15]. Particularly, results from our clinical trial on herpes labialis indicate that a lipid-soluble EGCG topical formulation possesses significant antiviral efficacy [13]. We also tested the antiviral activity of EGCG against SARS-CoV-2, with promising results [16]. Another advantage of using EC16 as a new drug candidate is that it is an FDA categorized generally recognized as a safe (GRAS) compound (GRAS Notice 772) [17] and an EPA-approved safe inert.
The long-term goal of our study is to develop intranasally applied EC16-containing new drugs to minimize long COVID symptoms either through prevention or therapeutic approaches. The current study aimed to test the proof-of-concept that EC16 in nasal formulations is able to effectively inhibit human coronavirus in vitro.
2. Results
2.1. Formulation F18-Containing EC16
Dilution of F18 into aqueous buffers resulted in fast formation of a flocculate precipitate that rapidly rose to a cream. Microscopic imaging of the vortexed samples revealed large aggregates of material (Figure 1), together with barely visible particles. Separation of the larger cream particles by centrifugation and quantitation of the polyphenol content showed that 103+11% of the polyphenol was recovered in the liquid phase, and 9.0+8.1% in the cream phase, as determined by a Folin-Ciocalteu assay. Given that there was remaining liquid (containing polyphenol) in the cream phase, essentially all the reactive polyphenol was present in the liquid suspension, and little (if any) in the bulk cream aggregate material. The composition of the cream was not further evaluated, and vortexed suspensions (i.e., including cream material) were used for subsequent virus tests.
2.2 Size Distribution of Small Particles
After brief low-speed centrifugation to remove larger particles and aggregates (cream), the particle size distribution of 10-fold dilutions of F18 in saline was evaluated by nanoparticle tracking analysis (NTA). The particles showed high polydispersity, with sizes ranging from <100 nm to about 1 μm (Figure 2). The median size (average of 3 independent preparations) was 210+42 nm (SD), and the cutoff size for 90% of the particles was <547+190 nm (n=3). The mean initial particle density was 3.07+0.06 x107/ml.
2.3. Dose Effect Results
Incubation of virus with dilutions of F18 demonstrated a dose-dependent increase in contact inhibition antiviral activity of the formulation, as represented by an increase in the log10 reduction in viral titer (Figure 3). A plateau in viral titer reduction was evident at higher doses. Non-linear regression with a fit to a hyperbola gave a good fit to the data (r2 =0.97, D’Agostino and Pearson omnibus K2 test of residual normality p=0.76). The Bmax was 4.21+0.12 (SE)) for the plateau in log10 viral titer reduction, equivalent to a 99.994% reduction. The Kd (equivalent to the concentration giving a reduction of 50% of Bmax) was 0.025+0.005 mM. Using these values, the concentration of EC16 giving a reduction in titer 90% of Bmax (99.984% reduction) was 0.225 mM.
2.4. Time Effect Results
To determine the time effect of EC16, the F18 stock was diluted to 1.25 mM EC16 with serum-free EMEM and incubated with OC43 virus for different time periods before neutralization by immediate dilution with serum-free EMEM. Figure 4 shows the results from three independent tests. The antiviral activity showed a rapid increase in log10 titer reduction up to a plateau. Non-linear regression with a fit to a hyperbola gave a good fit to the data (r2 =0.97, D’Agostino and Pearson omnibus K2 test of residual normality p=0.31). The Bmax for the plateau in log10 viral titer reduction was 4.77+0.23 (SE)), equivalent to a 99.998% reduction, and consistent with the fold reduction observed in the dose response testing.
The Kd (equivalent to the time giving a reduction of 50% of Bmax) was 5.86+1.26 min. Using these values, a 90% Bmax reduction in titer was predicted at 42.9 min. The 30 min exposure used for dose testing was predicted to give a 3.994 log10 reduction in titer, or 83.7% Bmax, and consistent with the values observed for dose testing.
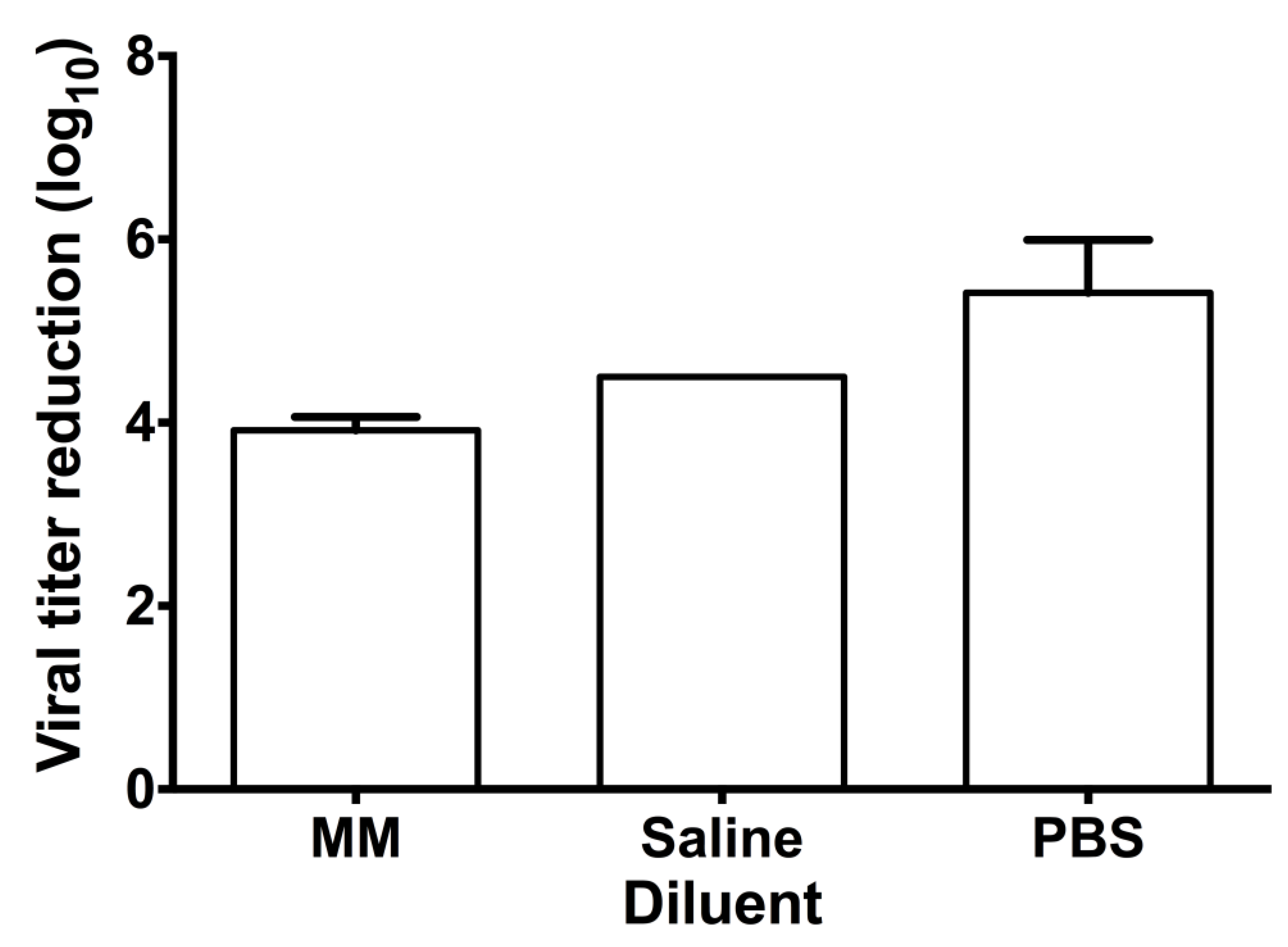
Figure 5.
Contact inhibition antiviral activity of F18 EC16 suspensions in three diluents. The glycerol-based F18 EC16 nasal formulation stock (1%) was diluted 10 X with serum-free EMEM (MM), normal saline, or phosphate buffered saline (PBS) to 1.25 mM. The working suspensions were incubated with OC43 virus for 30 min at a 1:9 ratio (virus : formulation) prior to serial 10x dilution and subjected to TCID50 assay. Results are means obtained from 3 independent experiments with standard deviation (the saline test result values were identical).
Figure 5.
Contact inhibition antiviral activity of F18 EC16 suspensions in three diluents. The glycerol-based F18 EC16 nasal formulation stock (1%) was diluted 10 X with serum-free EMEM (MM), normal saline, or phosphate buffered saline (PBS) to 1.25 mM. The working suspensions were incubated with OC43 virus for 30 min at a 1:9 ratio (virus : formulation) prior to serial 10x dilution and subjected to TCID50 assay. Results are means obtained from 3 independent experiments with standard deviation (the saline test result values were identical).
2.5. Contact Inhibition Test of F18 in Different Diluents
The effect of the formulation diluent on direct contract log10 reduction was tested using a 30 min exposure of OC43 virus (titer log 9.75/ml) to 1.25 mM EC16. There was a significant difference between the three diluents tested (saline, PBS and MM) (one -way ANOVA, (F2,6)=14.53; p=0.005). Post hoc testing (Tukey’s multiple comparisons test) showed no significant difference between MM and saline (p=0.18), but PBS (mean log10 5.42 + 0.58 SD) gave a significantly higher reduction (p<0.04) in comparison to MM or saline (mean log10 3.92 + 0.14 and 4.50 + 0.00 respectively). That is, F18 diluted in PBS gave a 99.9996% reduction in viral titer.
2.6. Contact Inhibition Test of F18m-Containing EC16m
The F18m formulation was diluted with serum-free EMEM to a working concentration of 1.40 mM EC16m. This formulation was incubated with virus (titer log 7.5-7.75/ml) for 30 min and then 10x serial dilutions were prepared and remaining viral titer determined. A 30 min incubation with 1.40 mM EC16m in serum-free EMEM resulted in a log10 3.00 + 0.43 (99.9%, n=3) reduction of viral infectivity The reduction in titer with F18m at this dose was somewhat modest, and significantly less than that seen with the higher 1.25 mM EC16 dose tested with F18 (t-test, assumption of equal variance, p=0.040).
2.7. Pre-Infection Tests
To mimic a nasal application, the F18m formulation at 50 μM was added to the testing wells with a monolayer of cells for just 10 min before removal and exposure of the cells to virus (note: these tests used a lower viral concentration of log10 6.25/ml). The result indicated that 10 min pre-infection incubation led to a log 1.81 + 0.38 (98.45%; n=4) reduction in viral infectivity.
2.8. Post-Infection Tests
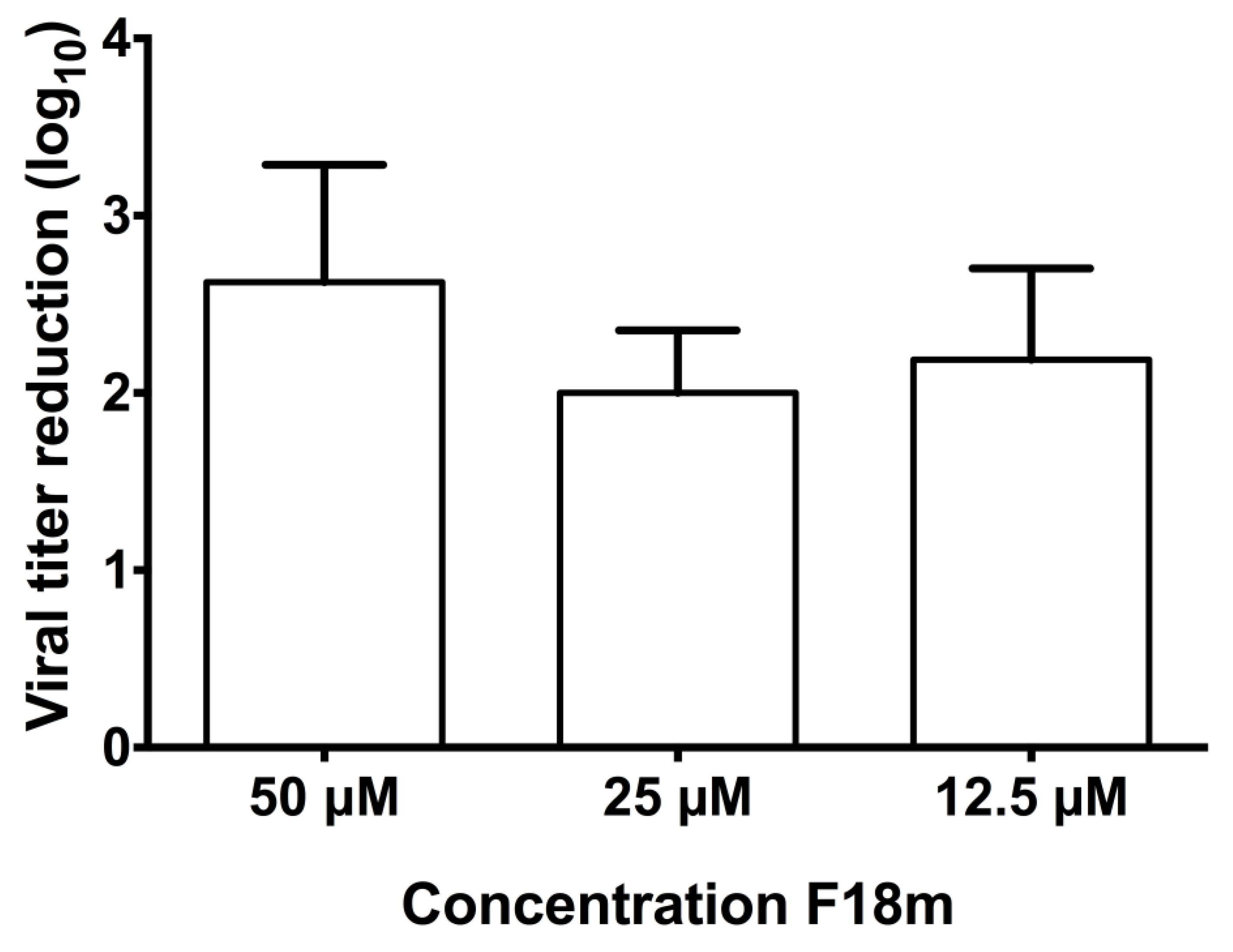
To determine the post-infection dose effect of EC16m in serum-free EMEM, the F18m stock was diluted with serum-free EMEM to 50, 25 and 12.5 μM, and antiviral activity was determined using a 10 min exposure to the diluted agent before changing to media. There was no significant difference in the reduction in viral titer (one-way repeated measures ANOVA, Geisser-Greenhouse epsilon 0.511; F(1.021,3.064)=2.548; p=0.21 between treatment groups (Figure 6, n=4). A dose of just 12.5 μM F18m gave a log10 titer reduction of 2.19+0.52, and 25 μM F18m gave a log10 titer reduction of 2.00+0.36 (99.0%), about the same as 50% of the Bmax determined for this dose of F18. A 50 μM dose gave a log10 2.63+0.66 (99.77%) reduction.
3. Discussion
The current study aimed to determine the feasibility of using EC16 as a nasal delivered drug to provide antiviral activity in terms of contact inactivation and pre- and post-infection inhibition of viral replication. The human nasal cavity is made of the respiratory epithelium (RE), which consists of ciliated cells, basal cells, brush cells, and secretory cells (3, 19), and the olfactory epithelium (OE), consisting of olfactory sensory neurons, sustentacular cells, microvillar cells, globose basal cells, and horizontal basal cells (19). The non-neuronal cells express ACE2 and TMPRSS2, and the olfactory sensory neurons express neuropilin-1, which facilitate SARS-CoV-2 infection [18]. Entry of SARS-CoV-2 into the nasal cavity results in infection and initial replication in the RE during the early stage of COVID-19, mainly in the ciliated cells that are rich in ACE2 and TMPRSS2 [19]. The rapid accumulation of SARS-CoV-2 in RE could cause concomitant infection in the OE. Indeed, recent clinical and animal studies demonstrate that SARS-CoV-2 infection of the olfactory sensory neurons and their support cells in the OE results in local inflammation and apoptosis, which could be the mechanisms leading to OE destruction, anosmia, and other neuronal dysfunctions in the CNS [18,20]. Thus, active SARS-CoV-2 replication in RE, OE, and the olfactory bulb appears to be the cause of acute anosmia, and persistent presence of the virus in the RE and OE cells could be associated with chronic neurologic symptoms [18]. Neutralizing antibodies, either injected in the nasal cavity or acquired via vaccination, are not effective at reducing the SARS-CoV-2 viral load in the nasal cavity due to robust viral replication in the nasal turbinates [21]. In addition, asymptomatic patients have a nasal viral load comparable to symptomatic patients, suggesting both symptomatic and asymptomatic patients are at risk for anosmia [22]. Therefore, inactivation and clearance of viral particles in the nasal cavity could effectively minimize the risks for post COVID neurologic symptoms.
Recent studies indicate that nose-to-brain (NTB) drug delivery using nanoparticles of lipid-soluble drug or nanocarrier is a promising method to increase the drug bioavailability with rapid action [23-25]. Specifically, NTB drug delivery technology has the potential to treat neurologic disorders by decreasing reactive oxygen species using natural antioxidants [26]. If the EC16 formulation not only performs the beneficial activity in the nasal epithelia, but also provide the multiple effects in the central nerve system (CNS), it would be a first-in-class drug to prevent and minimize SARS-CoV-2 associated neurologic symptoms, including long COVID.
As of today, the US FDA have approved a number of nasal delivered drugs to treat different symptoms such as Spravato (esketamine) nasal spray for depression (fast-track), Astepro (azelastine hydrochloride nasal spray, 0.15%) for seasonal and perennial allergic rhinitis, Narcan (naloxone hydrochloride) nasal spray for opioid overdose, and Ryaltris (mometasone furoate monohydrate) nasal spray for seasonal allergic rhinitis, etc. However, to the best of our knowledge, there is no intranasally administered drug for use against respiratory viral infection or post-infection symptoms.
The antiviral activity of EGCG has been widely reported. In addition, EGCG has been shown to provide neuroprotective effects to nerve cells. Results from preclinical and clinical trials on Fragile X syndrome patients demonstrate that 5-7 mg/kg/day EGCG combined with cognitive training significantly improved cognitive function in 3 months, without adverse effect [27]. The neuroprotective effects of EGCG include reducing Aβ and tau toxicity, and inhibition of apoptosis, suggesting a potential to prevent/treat neurodegenerative diseases such as Alzheimer's disease [28]. Collectively, CNS exposure to EGCG is safe and beneficial, and olfactory function could be protected.
EC16 is a lipid-soluble compound mixture derived from EGCG by esterification with palmitate. Our previous studies showed that EC16 is able to enter epithelial cells and is hydrolyzed in the cytoplasm, releasing free EGCG [29,30]. Since the bioavailability of EGCG is very low [33,34], nasal delivery could be a preferred route to administrate lipid-soluble EC16 for neuroprotection. In comparison to water-soluble EGCG, EC16 is significantly more potent against influenza virus, herpes simplex virus, and norovirus [12,15,31]. Other advantages of EC16 over EGCG are that EC16 is more stable and long-lasting [13,15,32]. In addition, the US FDA approved the use of tea polyphenol palmitates (contains 50% EC16) as a GRAS green tea extract (FDA GRAS Notice 772). EC16 undergoes hydrolysis after consumption, consistent with our findings that EC16 is hydrolyzed to free EGCG after entering cells [29,30]. These studies concluded that the intended use of EC16 meets the GRAS requirement [17]. Furthermore, we have tested EC16 in different animal and human epithelial cells (MDCK, CRFK, RAW.264.7, FRhk-4, human keratinocytes) and in a herpes labialis clinical trial (12-16). The results indicate that EC16 has the potential to protect nasal epithelial cells from SARS-CoV-2 infection as well as exerting anti-inflammatory, antioxidant, and neuroprotective effects.
Our previous in vivo studies demonstrate that the neuroinvasion of SARS-CoV-2 occurs closely following the peaking of viral replication in the nasal epithelia [2]. Thus, the formulations must possess rapid antiviral actions to inactivate the coronavirus in the nasal epithelial cells and be able to deliver the multiple benefits of EC16 into the brain, bypassing the blood-brain barrier (BBB) and digestive system. The nasal delivery method would overcome the well-documented poor bioavailability of EGCG, which has a serum maximum concentration in the sub-micromolar range (0.57 μM), well below the effective concentration for beneficial effects [33,34]. If EC16 is applied to the nasal epithelial cells, the long chain fatty acyl group would allow EC16 to attach to the cell membrane for prolonged effect against SARS-CoV-2 and its variants.
Our goal is therefore to develop an intranasally applied drug (in the form of a spray and/or irrigation) based on EC16 to prevent and treat respiratory viral infection and post-infection symptoms. To achieve this goal, the primary property of interest is antiviral activity, and the secondary property is anti-inflammatory, antioxidant, and neuroprotection in both nasal neuroepithelia and the brain. The current study aimed to test the feasibility of formulating candidate nasal formulations containing EC16 with antiviral properties for COVID and long COVID use.
Due to the lipid-soluble nature of EC16, the solubility of EC16 is very low in aqueous solutions. By using our patent-pending technology, we were able to bring EC16 into aqueous suspension as particles ranging in size from nano- to micro-meters. Among 62 formulations tested thus far, the F18 glycerol-based stock formulation showed the most promising results for antiviral activity with the simplest composition. Dilution of F18 into aqueous buffer systems resulted in rapid formation of a flocculate precipitate, with little, if any, polyphenol in the flocculate material, which was comprised of large aggregates of various-sized particles (Figure 5). Analysis of particle size distribution in the saline liquid phase showed a broad polydisperse suspension ranging <100 nm to about 1 μm (Figure 2).
The F18 formulation diluted in EMEM showed potent antiviral activity when virus was directly exposed to it, with a maximum inhibitory effect of log10 4.21+0.12 (SE) (99.996%) at saturating concentrations in the 1 mM range (Figure 3). Based on a regression analysis and determination of the curve constants, a concentration of 0.225 mM was predicted to give a log10 3.79 reduction (90% of maximum). Thus, experiments conducted with 1.25 mM were firmly in the saturated region. The antiviral results of F18 diluted in serum-free EMEM indicated that at 50 μM (approximately 40 ng/ml), a 30 min incubation with the virus reduced the infectivity by 99.90%. A concentration of 8 μM was predicted to give a 90% reduction (the EC90; log10 1 reduction). This would be significantly more potent than the antiviral activity of EGCG against SARS-CoV-2 ([10], one-hour incubation EC90 = 69 μM).
At a saturating 1.25 mM concentration of ECV16, a 5-min direct contact with the virus reduced the viral infectivity by >99% (Figure 4). These results suggested that the rapid and potent inactivation of viral infectivity is associated with a completely different mechanism of action in comparison to antiviral drugs in use, such as the COVID drug Remdesivir (a nucleoside analog to inhibit RNA polymerization), which has no known contact inhibition of coronavirus.
For our ongoing animal study and a future human study, the F18 formulation was tested by dilution in PBS and saline, which showed comparable direct virus log10 inhibition efficacy results (Figure 5), although dilution in PBS gave significantly greater reduction (log10 5.41 versus log10 4.21 (average saline and MM), equivalent to 16.2-fold better), suggesting that the phosphate content may enhance the antiviral activity, presumably by modification of the surface charge of particles. Of note, the difference is based on an already very high level (log 3.92-4.5 vs. log 5.42).
EC16 is a mixture of EGCG-palmitates, with the majority being EGCG-mono-palmitate (EC16m), followed by EGCG-di-palmitates, and EGCG-tri-palmitates. Therefore, the single molecule EC16m would most likely be the new drug form. In this report, a series of initial tests for EC16m was performed with the F18m formulation of EC16m in serum-free EMEM dilutions. After a 30 min incubation with 1.41 mM (0.1%) EC16m the infectivity of the OC43 virus was reduced by 99.9%, similar to that seen with EC16 (99.996%, Figure 3) at a moderately higher dose. The measured log10 reduction value was influenced by the titer of the virus used in the tests, with a lower titer resulting in a lower proportionate log10 reduction. Since the F18m tests used a lower titer virus preparation (log 7.75 in comparison to > log 9.0 in EC16 contact inhibition tests), EC16m had similar activity to EC16.
A human study indicated that when symptoms appear during initial SARS-CoV-2 infection, the nasal cavity viral load is less than log10 8 and peaks (day 2 to 6) at > log10 9 [35]. Therefore, the tests conducted in this study have been at clinically relevant viral loads. Here, a 10-min pre-infection incubation of cells with 50 μM (approximately 35 ng/ml) EC16m gave a 98.45% inhibition of subsequent viral replication in the cells. On the other hand, without direct contact with the virus, a 10 min incubation of infected MRC-5 cells with 50 μM EC16m (approximately 35 ng/ml) reduced the viral replication by 99.77% (Figure 6). Even at just 12.5 μM, EC16m was able to inhibit viral replication by >99% after 10 min post-infection treatment with the cells before removal. These initial results from a base EC16m formulation suggested that EC16m entered the cells and blocked the viral replication effectively, because the inhibition was the result of a single application and viral titer was observed over a 4 to 7-day incubation period. Repeated applications of EC16m may produce a higher inhibitory effect on viral replication.
According to the characteristics of SARS-CoV-2 replication in the nasal cavity, investigators studied if saline irrigation (gavage) in the nasal cavity would assist in infection recovery. Indeed, a randomized clinical trial using isotonic saline pressured irrigation, supplemented with either sodium bicarbonate or povidone-iodine, significantly reduced hospitalization rate during the early stage of COVID-19 pandemic [36]. Saline, either isotonic or hypertonic, has been used for respiratory conditions as an economical and effective alternative to medications [37,38]. Thus, a novel saline-based formulation containing EC16 (EC16m for pharmaceutical use) would be a potential nasal application for COVID and long COVID prevention and intervention.
A multi-mechanism of action has been reported for the inhibitory effects of EGCG against influenza virus [39]. Taken together, the results presented here showed that F18 base formulations containing EC16 or +EC16m similarly demonstrated antiviral activities to either rapidly inactivate human coronavirus by direct contact or inhibit viral entry and replication without direct contact with the virus.
4. Materials and Methods
4.1. Virus and Cell Line
OC43 human coronavirus (ATCC VR-1558) and MCR-5 human respiratory fibroblast cells (ATCC CCL-171) were purchased from ATCC.
4.2. EC16 and Other Supplies
Epigallocatechin-3-Gallate-Palmitates (EC16); a mixture of mono- di- and tri-palmitates) and Epigallocatechin-3-Gallate-mono-palmitate (EC16m), was provided by Camellix, LLC (Evans, GA). Eagle’s Minimal Essential Medium (EMEM) was purchased from ATCC (30-2003). Fetal bovine serum (FBS) was from Neuromics (Edina, MN). Trypsin (0.25%)-EDTA was provided by Fisher Scientific. Penicillin, streptomycin, and amphotericin B solution (100x) was ordered from Corning (Glendale, AR). Plasticwares were purchased from Southern Labware (Cumming, GA).
4.3. EC16 Formulations
EC16 and EC16m were initially dispersed as stable glycerol-based stocks (F18 and F18m respectively) at 1% w/v using a proprietary method. Working formulations were made by a 10 X dilution with serum-free EMEM (MM, for cell-based studies), normal saline solution (for future clinical studies), or phosphate buffer saline (PBS, for animal studies). These 0.1% EC16 or EC16m formulations were equal to approximately 1.25 mM (EC16) and 1.40 mM (EC16m), respectively, from which they were diluted to lower concentrations for the experiments.
4.4. Quantitation of EC16 Polyphenol in Mixtures
After dilution of F18 in aqueous buffers the flocculent material was separated by centrifugation (700 rpm for 3 min). The liquid under the cream was removed and the cream reconstituted by vigorous vortexing in the same volume of liquid. The distribution of polyphenols in the fractions was determined by the Folin-Ciocalteu reaction as described with minor modifications [40], using EC16 dissolved in ethanol as the standard.
4.5. Evaluation of Particle Size Distribution
ZetaView nanoparticle tracking analysis was performed according to a method described previously [41]. The particle size distribution and concentration were measured using the Zetaview x20 (Particle Metrix, Meerbusch, Germany) and corresponding software. The measuring range for particle diameter is 10-2000 nm. The samples were diluted by the same volume of 1×PBS and then loaded into the cell. The instrument collected particle information from 11 different positions across the cell, with two cycles of readings respectively. The standard operating procedure was set to a temperature of 23°C, a sensitivity of 70, a frame rate of 30 frames per second, and a shutter speed of 100. The post-acquisition parameters were set to a minimum brightness of 20, a maximum area of 1000, a minimum area of 10, and the tracelength of 15 (41).
4.6. Antiviral Activity Tests
Infection of cells by OC43 virus, and viral titer: MRC-5 cells were cultured in EMEM Medium supplemented with 10% FBS and 1% penicillin, streptomycin, and amphotericin B. The viral infection assay and viral titering were performed in 96 well cell culture plates when the cells had reached 90% confluency. A 10-fold series dilution of OC43 virus in serum-free EMEM was loaded into wells in quadruplicates per dilution. After a one-hour absorption, the viral dilutions were removed and 100 µl serum-free EMEM was added, followed by incubation at 33 °C with 5% CO2 for >4 days to allow a CPE (cytopathic effect) to become visible. Viral titer was calculated by a TCID50 protocol and software [42]. A minimum of three independent experiments were performed and results recorded. To compare EC16 activity to an antiviral in clinical use, exposure of cells to 50 µM remdesivir was used.
4.7. Dose and Time Tests of Direct Contact with Virus
EC16 nasal formulations were used with different concentrations or incubation times in direct contact with OC43 virus in serum free EMEM. To determine the dose effect of EC16, in three independent tests the F18 stock formulation was diluted with serum-free EMEM to a series of concentrations from 1.25 mM down to 0.05 mM, and incubated with OC43 (titer log 9.25/ml diluted in serum-free EMEM) for 30 min. To determine the time of exposure effects, a 1.25 mM dilution of F18 in serum-free EMEM was mixed with virus for 5, 15, 30 or 60 min, followed by rapid serial dilution with serum-free EMEM and loaded onto 96-well plates containing 90% confluent MRC-5 cells for one-hour viral absorption, followed by media change. The infectivity rate was determined by TCID50 method after 4-7 days of incubation.
4.8. Pre-Infection Test
EC16m nasal formulations were incubated with MRC-5 cells in 96 well plates for 10 min, and then removed. A series dilution of OC43 in serum-free EMEM was added to the cells and incubated for 1 h. The viral dilutions were then replaced with serum-free EMEM, and TCID50 infection rate was determined after 4-7 days of incubation.
4.9. Post-Infection Test
To test if EC16m nasal formulations possess a post-infection effect, MRC-5 cells in 96 well cell culture plate were infected for one hour with the virus in series dilutions before removal. Then, 100 µl of either EC16 formulations or Remdesivir solutions of various concentrations were applied onto the cells for 10 min before being replaced by serum-free EMEM. The TCID50 values were determined after incubation for 4-7 days.
4.10. Microphotography of the Formulations
EC16 particles in the stock and in the three F18 diluted suspension formulations (MM, saline, and PBS) were photographed with a Zeiss Axio Imager M.2 microscope at 40x magnification under DIC illumination, and the images were recorded.
4.11. Statistical Analysis
The primary statistical tests were parametric one-way ANOVA (with appropriate adjunct tests for homogeneity of variance, etc.) and non-linear regression using a fit to a parabolic curve. Alpha was 0.05. GraphPad Prism version 6.0 software (www.graphpad.com) was used for most analyses. Reported errors are given as standard deviation (SD) unless otherwise noted.
5. Conclusions
We report here, for the first time according to the best of our knowledge, that epigallocatechin-3-gallate-palmitate(s) is a candidate for intranasally applied formulations to minimize COVID-associated neurologic symptoms through its strong and rapid antiviral and other beneficial properties. Our ongoing studies are testing the broad-spectrum of antiviral activity of EC16 on human α-coronavirus (229E) in vitro; and SARS-CoV-2 variants in vitro and in vivo using K18-hACE2 mice. In addition, saline-based formulations with significantly higher potency are being explored. These ongoing studies have shown promising initial results (data not shown). For new drug development, the next phase studies should include formulation optimization and finalization (chemistry, compatibility, stability, validation, etc.), and initial efficacy and toxicity with in vivo and ex vivo models, in order to collect data for pre-Investigational New Drug (IND) studies (including chemistry-manufacturing-control, pharmacology-toxicity, etc.). The advantage of the formulation is that all ingredients are known nontoxic molecules, and saline is regularly used in nasal irrigation/spray solutions. In conclusion, we have developed a simple nasal formulation to incorporate lipid-soluble EC16 in saline, without surfactant, to rapidly inactivate human β-coronavirus OC43 by contact or inhibit viral replication by a single 10-min application on infected cells. These results suggest EC16 nasal applications are potential prophylactic and therapeutic methods to minimize respiratory virus associated, such as COVID and post-COVID, symptoms, pending additional studies.
Author Contributions
Conceptualization, D.D., L.H.L., T.C., M.K., S.S., Y.L., and S.H.; methodology, N.F., D.D., and S.H..; validation, D.D., L.H.L., T.C., and S.H.; formal analysis, S.H., D.D.; investigation, N.F., D.D., W.G., L.X., A.X., H.Y., J.C., and S.H.; resources, S.H.; data curation, N.F., and D.D.; writing—original draft preparation, S.H.; writing—review and editing, N.F., D.D., W.G., L.X., A.X., L.H.L, T.C., M.K., S.S., Y.L., J.C., and S.H.; visualization, N.F., and S.H.; supervision, D.D., and S.H.; project administration, D.D., and S.H.; funding acquisition, D.D., and S.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the National Institute of Deafness and Other Communication Disorders (NIDCD) to Camellix, LLC (1R41DC020678-01), Augusta University-Georgia State University Seed Award to SH and MK, and the Office of Innovation Commercialization, Augusta University Research Institute. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Groff, D.; Sun, A.; Ssentongo, A.E.; Ba, D.M.; Parsons, N.; Poudel, G.R.; Lekoubou, A.; Oh, J.S.; Ericson, J.E.; Ssentongo, P.; et al. Short-term and Long-term Rates of Postacute Sequelae of SARS-CoV-2 Infection: A Systematic Review. JAMA Netw Open 2021, 4, e2128568. [Google Scholar] [CrossRef]
- Kumari, P.; Rothan, H.A.; Natekar, J.P.; Stone, S.; Pathak, H.; Strate, P.G.; Arora, K.; Brinton, M.A.; Kumar, M. Neuroinvasion and Encephalitis Following Intranasal Inoculation of SARS-CoV-2 in K18-hACE2 Mice. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat Med 2020, 26, 681–687. [Google Scholar] [CrossRef]
- Gallo, O.; Locatello, L.G.; Mazzoni, A.; Novelli, L.; Annunziato, F. The central role of the nasal microenvironment in the transmission, modulation, and clinical progression of SARS-CoV-2 infection. Mucosal Immunol 2021, 14, 305–316. [Google Scholar] [CrossRef]
- Wu, C.T.; Lidsky, P.V.; Xiao, Y.; Cheng, R.; Lee, I.T.; Nakayama, T.; Jiang, S.; He, W.; Demeter, J.; Knight, M.G.; et al. SARS-CoV-2 replication in airway epithelia requires motile cilia and microvillar reprogramming. Cell 2023, 186, 112–130 e120. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, S.; Dinda, M. Therapeutic potential of green tea catechin, (-)-epigallocatechin-3-O-gallate (EGCG) in SARS-CoV-2 infection: Major interactions with host/virus proteases. Phytomed Plus 2023, 3, 100402. [Google Scholar] [CrossRef]
- Goncalves, P.B.; Sodero, A.C.R.; Cordeiro, Y. Green Tea Epigallocatechin-3-gallate (EGCG) Targeting Protein Misfolding in Drug Discovery for Neurodegenerative Diseases. Biomolecules 2021, 11. [Google Scholar] [CrossRef]
- Hsu, S. Compounds Derived from Epigallocatechin-3-Gallate (EGCG) as a Novel Approach to the Prevention of Viral Infections. Inflamm Allergy Drug Targets 2015, 14, 13–18. [Google Scholar] [CrossRef]
- Kaihatsu, K.; Yamabe, M.; Ebara, Y. Antiviral Mechanism of Action of Epigallocatechin-3-O-gallate and Its Fatty Acid Esters. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Mokra, D.; Joskova, M.; Mokry, J. Therapeutic Effects of Green Tea Polyphenol (‒)-Epigallocatechin-3-Gallate (EGCG) in Relation to Molecular Pathways Controlling Inflammation, Oxidative Stress, and Apoptosis. Int J Mol Sci 2022, 24. [Google Scholar] [CrossRef]
- Payne, A.; Nahashon, S.; Taka, E.; Adinew, G.M.; Soliman, K.F.A. Epigallocatechin-3-Gallate (EGCG): New Therapeutic Perspectives for Neuroprotection, Aging, and Neuroinflammation for the Modern Age. Biomolecules 2022, 12. [Google Scholar] [CrossRef]
- de Oliveira, A.; Adams, S.D.; Lee, L.H.; Murray, S.R.; Hsu, S.D.; Hammond, J.R.; Dickinson, D.; Chen, P.; Chu, T.C. Inhibition of herpes simplex virus type 1 with the modified green tea polyphenol palmitoyl-epigallocatechin gallate. Food Chem Toxicol 2013, 52, 207–215. [Google Scholar] [CrossRef]
- Dickinson, D.P.; Xayaraj, S.; Dickinson, S.; Shao, X.; Hsu, S. Effect of Novel Formulations using Lipophilic Epigallocatechin-3-Gallate against Influenza Virus Infection. Microbiology & Infectious Diseases. 2018, 14.
- Zhao, M.; Zheng, R.; Jiang, J.; Dickinson, D.; Fu, B.; Chu, T.C.; Lee, L.H.; Pearl, H.; Hsu, S. Topical lipophilic epigallocatechin-3-gallate on herpes labialis: a phase II clinical trial of AverTeaX formula. Oral Surg Oral Med Oral Pathol Oral Radiol 2015, 120, 717–724. [Google Scholar] [CrossRef]
- Zhong, J.; Dickinson, D.; Hsu, S. Effects of Epigallocatechin-3-Gallate-Palmitate (EC16) on In Vitro Norovirus Infection. Microbiol Infect Dis 2021, 5. [Google Scholar] [CrossRef]
- Hurst, B.L.; Dickinson, D.; Hsu, S. Epigallocatechin-3-Gallate (EGCG) Inhibits SARS-CoV-2 Infection in Primate Epithelial Cells: (A Short Communication). Microbiol Infect Dis 2021, 5. [Google Scholar] [CrossRef]
- 772, U.G.N. GRAS Notice for Oil-Soluble Green Tea Extract (Green Tea Catechin Palmitate). 2018.
- de Melo, G.D.; Lazarini, F.; Levallois, S.; Hautefort, C.; Michel, V.; Larrous, F.; Verillaud, B.; Aparicio, C.; Wagner, S.; Gheusi, G.; et al. COVID-19-related anosmia is associated with viral persistence and inflammation in human olfactory epithelium and brain infection in hamsters. Sci Transl Med 2021, 13. [Google Scholar] [CrossRef]
- Ahn, J.H.; Kim, J.; Hong, S.P.; Choi, S.Y.; Yang, M.J.; Ju, Y.S.; Kim, Y.T.; Kim, H.M.; Rahman, M.D.T.; Chung, M.K.; et al. Nasal ciliated cells are primary targets for SARS-CoV-2 replication in the early stage of COVID-19. J Clin Invest 2021, 131. [Google Scholar] [CrossRef]
- Bryche, B.; St Albin, A.; Murri, S.; Lacote, S.; Pulido, C.; Ar Gouilh, M.; Lesellier, S.; Servat, A.; Wasniewski, M.; Picard-Meyer, E.; et al. Massive transient damage of the olfactory epithelium associated with infection of sustentacular cells by SARS-CoV-2 in golden Syrian hamsters. Brain Behav Immun 2020, 89, 579–586. [Google Scholar] [CrossRef]
- Zhou, D.; Chan, J.F.; Zhou, B.; Zhou, R.; Li, S.; Shan, S.; Liu, L.; Zhang, A.J.; Chen, S.J.; Chan, C.C.; et al. Robust SARS-CoV-2 infection in nasal turbinates after treatment with systemic neutralizing antibodies. Cell Host Microbe 2021, 29, 551–563 e555. [Google Scholar] [CrossRef]
- Ra, S.H.; Lim, J.S.; Kim, G.U.; Kim, M.J.; Jung, J.; Kim, S.H. Upper respiratory viral load in asymptomatic individuals and mildly symptomatic patients with SARS-CoV-2 infection. Thorax 2021, 76, 61–63. [Google Scholar] [CrossRef]
- Giunchedi, P.; Gavini, E.; Bonferoni, M.C. Nose-to-Brain Delivery. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Khan, A.R.; Yang, X.; Fu, M.; Zhai, G. Recent progress of drug nanoformulations targeting to brain. J Control Release 2018, 291, 37–64. [Google Scholar] [CrossRef]
- Maaz, A.; Blagbrough, I.S.; De Bank, P.A. In Vitro Evaluation of Nasal Aerosol Depositions: An Insight for Direct Nose to Brain Drug Delivery. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef]
- Bonferoni, M.C.; Rassu, G.; Gavini, E.; Sorrenti, M.; Catenacci, L.; Giunchedi, P. Nose-to-Brain Delivery of Antioxidants as a Potential Tool for the Therapy of Neurological Diseases. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- de la Torre, R.; de Sola, S.; Farre, M.; Xicota, L.; Cuenca-Royo, A.; Rodriguez, J.; Leon, A.; Langohr, K.; Gomis-Gonzalez, M.; Hernandez, G.; et al. A phase 1, randomized double-blind, placebo controlled trial to evaluate safety and efficacy of epigallocatechin-3-gallate and cognitive training in adults with Fragile X syndrome. Clin Nutr 2020, 39, 378–387. [Google Scholar] [CrossRef]
- Singh, N.A.; Mandal, A.K.; Khan, Z.A. Potential neuroprotective properties of epigallocatechin-3-gallate (EGCG). Nutr J 2016, 15, 60. [Google Scholar] [CrossRef]
- Chen, P.; Dickinson, D.; Hsu, S. Lipid-soluble Green Tea Polyphenols: Stabilized for Effective Formulation. In Handbook of Green Tea and Health Research, McKinley, H., Jamieson, M., Eds.; Nova Science Publishers, Inc.: New York, 2009; pp. 45–61. [Google Scholar]
- Hsu, S.; Dickinson, D. Green tea and skin protection: Mechanism of action and practical applications. Household and Personal Care TODAY 2009, n 2, 33–36. [Google Scholar]
- Mori, S.; Miyake, S.; Kobe, T.; Nakaya, T.; Fuller, S.D.; Kato, N.; Kaihatsu, K. Enhanced anti-influenza A virus activity of (-)-epigallocatechin-3-O-gallate fatty acid monoester derivatives: effect of alkyl chain length. Bioorg Med Chem Lett 2008, 18, 4249–4252. [Google Scholar] [CrossRef]
- Wei, Y.; Chen, P.; Ling, T.; Wang, Y.; Dong, R.; Zhang, C.; Zhang, L.; Han, M.; Wang, D.; Wan, X.; et al. Certain (-)-epigallocatechin-3-gallate (EGCG) auto-oxidation products (EAOPs) retain the cytotoxic activities of EGCG. Food Chem 2016, 204, 218–226. [Google Scholar] [CrossRef]
- Cai, Z.Y.; Li, X.M.; Liang, J.P.; Xiang, L.P.; Wang, K.R.; Shi, Y.L.; Yang, R.; Shi, M.; Ye, J.H.; Lu, J.L.; et al. Bioavailability of Tea Catechins and Its Improvement. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Yang, C.S.; Chen, L.; Lee, M.J.; Balentine, D.; Kuo, M.C.; Schantz, S.P. Blood and urine levels of tea catechins after ingestion of different amounts of green tea by human volunteers. Cancer Epidemiol Biomarkers Prev 1998, 7, 351–354. [Google Scholar]
- Savela, E.S.; Viloria Winnett, A.; Romano, A.E.; Porter, M.K.; Shelby, N.; Akana, R.; Ji, J.; Cooper, M.M.; Schlenker, N.W.; Reyes, J.A.; et al. Quantitative SARS-CoV-2 Viral-Load Curves in Paired Saliva Samples and Nasal Swabs Inform Appropriate Respiratory Sampling Site and Analytical Test Sensitivity Required for Earliest Viral Detection. J Clin Microbiol 2022, 60, e0178521. [Google Scholar] [CrossRef]
- Baxter, A.L.; Schwartz, K.R.; Johnson, R.W.; Kuchinski, A.M.; Swartout, K.M.; Srinivasa Rao, A.S.R.; Gibson, R.W.; Cherian, E.; Giller, T.; Boomer, H.; et al. Rapid initiation of nasal saline irrigation to reduce severity in high-risk COVID+ outpatients. Ear Nose Throat J 2022, 1455613221123737. [Google Scholar] [CrossRef]
- Huijghebaert, S.; Hoste, L.; Vanham, G. Essentials in saline pharmacology for nasal or respiratory hygiene in times of COVID-19. Eur J Clin Pharmacol 2021, 77, 1275–1293. [Google Scholar] [CrossRef]
- Panta, P.; Chatti, K.; Andhavarapu, A. Do saline water gargling and nasal irrigation confer protection against COVID-19? Explore (NY) 2021, 17, 127–129. [Google Scholar] [CrossRef]
- Xu, J.; Xu, Z.; Zheng, W. A Review of the Antiviral Role of Green Tea Catechins. Molecules 2017, 22. [Google Scholar] [CrossRef]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Thorough study of reactivity of various compound classes toward the Folin-Ciocalteu reagent. J Agric Food Chem 2010, 58, 8139–8144. [Google Scholar] [CrossRef]
- Helwa, I.; Cai, J.; Drewry, M.D.; Zimmerman, A.; Dinkins, M.B.; Khaled, M.L.; Seremwe, M.; Dismuke, W.M.; Bieberich, E.; Stamer, W.D.; et al. A Comparative Study of Serum Exosome Isolation Using Differential Ultracentrifugation and Three Commercial Reagents. PLoS One 2017, 12, e0170628. [Google Scholar] [CrossRef]
- REED, L.J.; MUENCH, H. A SIMPLE METHOD OF ESTIMATING FIFTY PER CENT ENDPOINTS12. American Journal of Epidemiology 1938, 27, 493–497. [Google Scholar] [CrossRef]
Figure 1.
Light microscopy imaging of F18 stock and dilutions. The F18 glycerol stock, and 1:10 dilutions in MM, PBS and saline were examined by transmitted light microscopy at 40x magnification using DIC illumination. (A) F18 stock; (B) F18 MM; (C) F18 Saline; (D) F18 PBS. .
Figure 1.
Light microscopy imaging of F18 stock and dilutions. The F18 glycerol stock, and 1:10 dilutions in MM, PBS and saline were examined by transmitted light microscopy at 40x magnification using DIC illumination. (A) F18 stock; (B) F18 MM; (C) F18 Saline; (D) F18 PBS. .
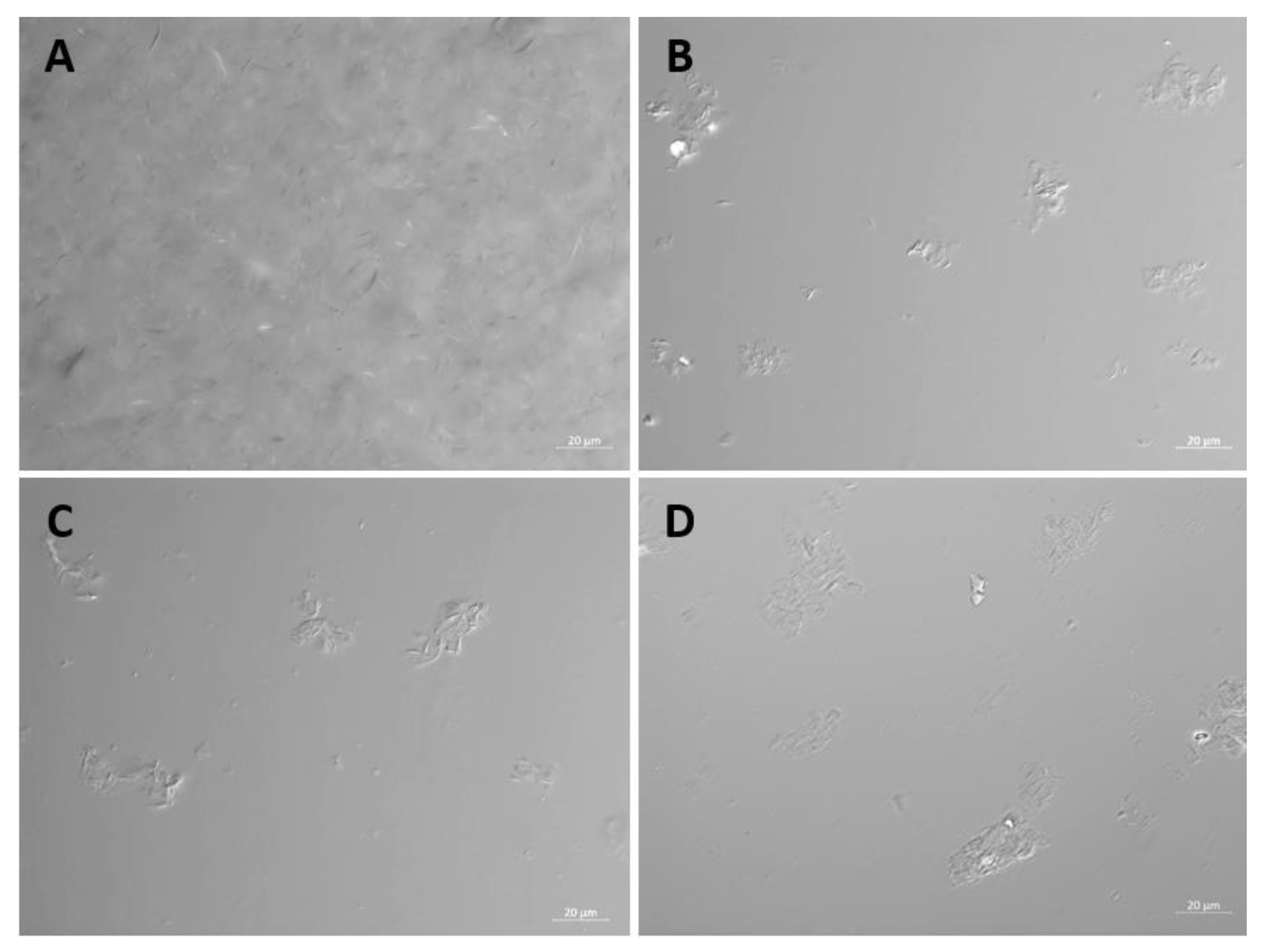
Figure 2.
Size distribution of particles in saline-diluted F18 determined by NTA. The size distribution profile of one representative sample is shown.
Figure 2.
Size distribution of particles in saline-diluted F18 determined by NTA. The size distribution profile of one representative sample is shown.
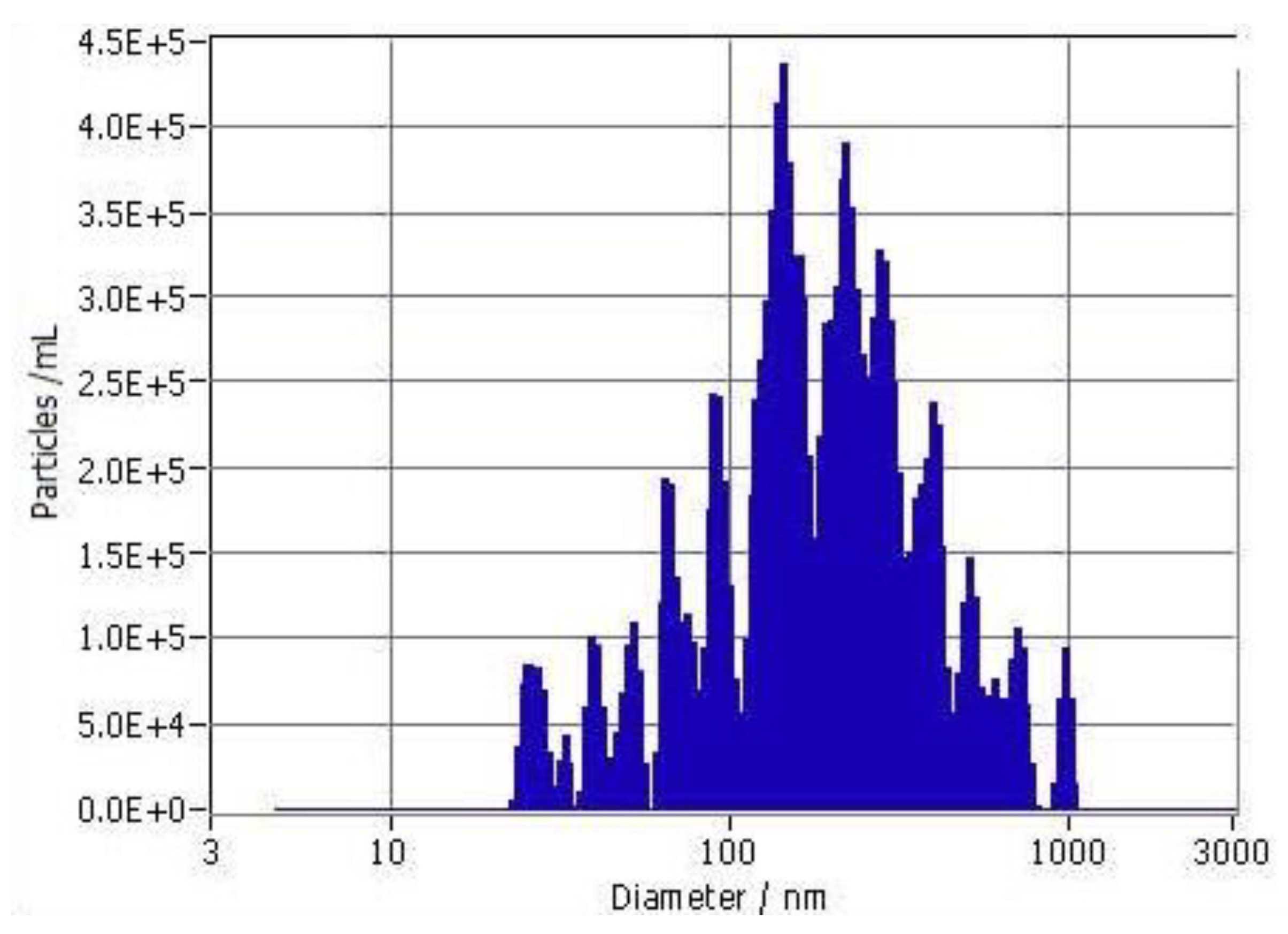
Figure 3.
Dose response of contact inhibition antiviral activity of F18 EC16 nasal formulation. The formulations were diluted from an F18 glycerol stock of EC16 (1%) into serum-free EMEM to a concentration of 0.05, 0.125, 0.25, 0.625, and 1.25 mM prior to incubation with OC43 virus at a 1:9 ratio (virus to formulation) for 30 min, followed by a series of 10 X dilutions and TCID50 assay. The contact inhibition antiviral activity was calculated and expressed as log10 reduction of the original titer (log10 9.25/ml). Results are the mean from three independent experiments (bars show standard deviation).
Figure 3.
Dose response of contact inhibition antiviral activity of F18 EC16 nasal formulation. The formulations were diluted from an F18 glycerol stock of EC16 (1%) into serum-free EMEM to a concentration of 0.05, 0.125, 0.25, 0.625, and 1.25 mM prior to incubation with OC43 virus at a 1:9 ratio (virus to formulation) for 30 min, followed by a series of 10 X dilutions and TCID50 assay. The contact inhibition antiviral activity was calculated and expressed as log10 reduction of the original titer (log10 9.25/ml). Results are the mean from three independent experiments (bars show standard deviation).
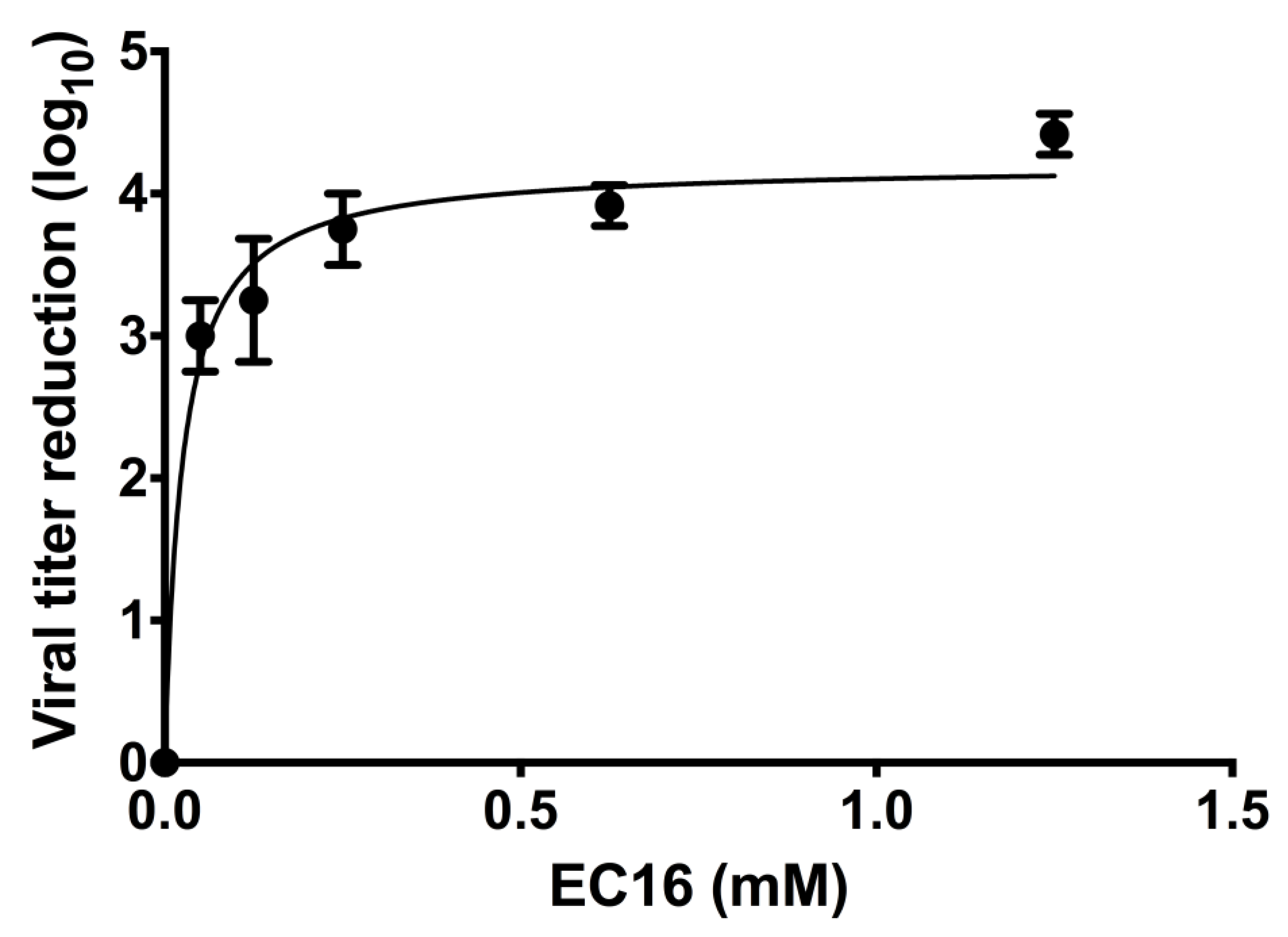
Figure 4.
Time response of contact inhibition antiviral activity of F18 EC16 nasal formulation. The formulation was diluted from an F18 glycerol stock of EC16 (1%) in serum-free EMEM, to a concentration of 1.25 mM EC16. This working formulation was incubated with OC43 virus (initial titer log 9.0/ml in serum-free EMEM) at a 1:9 ratio (virus to formulation) for 5, 15, 30, and 60 min before 10x serial dilutions and subjected to TCID50 assay. The antiviral activity was calculated and expressed as log10 reduction with standard deviation. Results are from three independent experiments.
Figure 4.
Time response of contact inhibition antiviral activity of F18 EC16 nasal formulation. The formulation was diluted from an F18 glycerol stock of EC16 (1%) in serum-free EMEM, to a concentration of 1.25 mM EC16. This working formulation was incubated with OC43 virus (initial titer log 9.0/ml in serum-free EMEM) at a 1:9 ratio (virus to formulation) for 5, 15, 30, and 60 min before 10x serial dilutions and subjected to TCID50 assay. The antiviral activity was calculated and expressed as log10 reduction with standard deviation. Results are from three independent experiments.
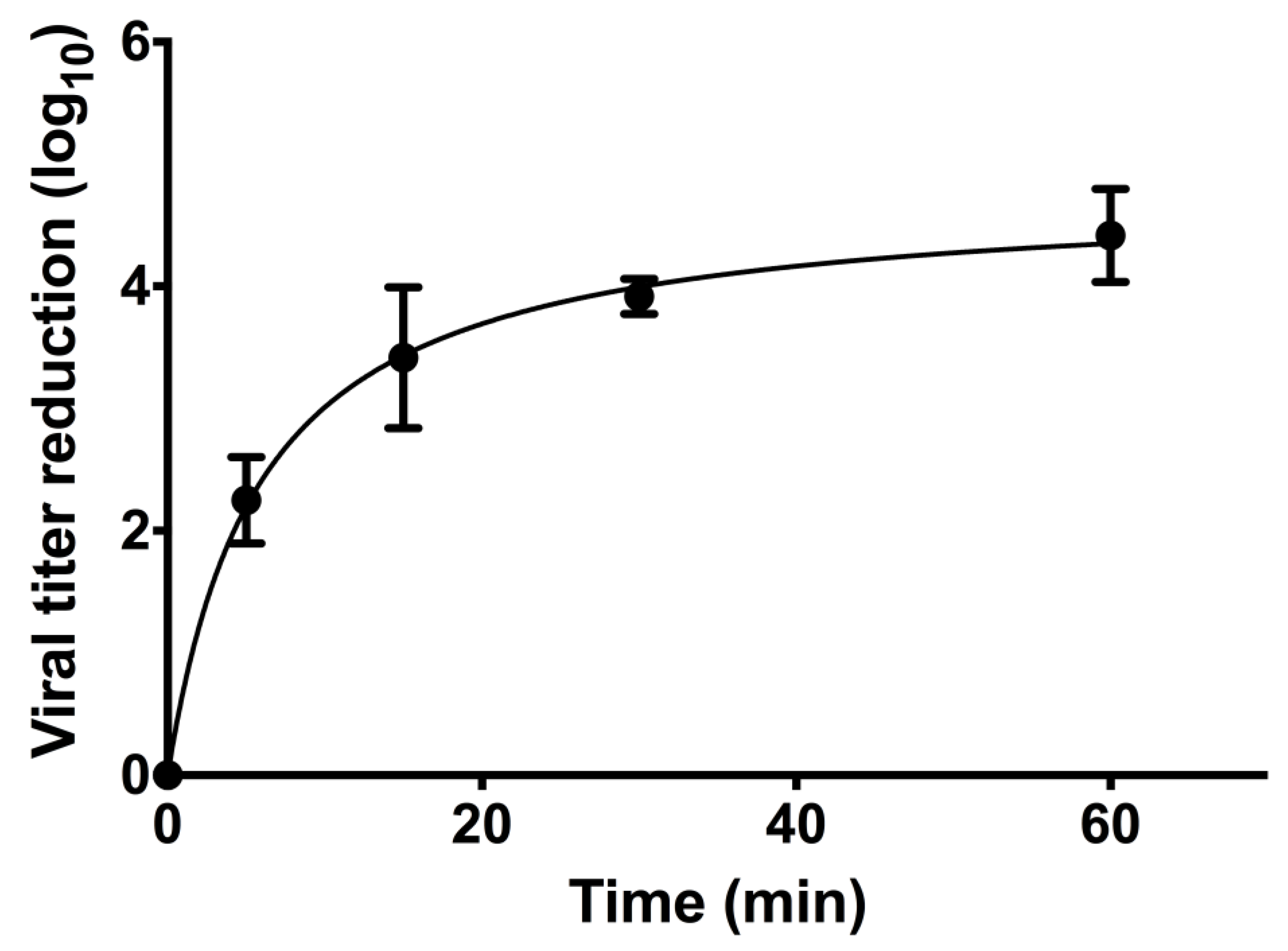
Figure 6.
Initial antiviral tests using EC16m to determine the antiviral activity on post-infection with OC43 virus. Results were obtained from four independent experiments (bars show SD).
Figure 6.
Initial antiviral tests using EC16m to determine the antiviral activity on post-infection with OC43 virus. Results were obtained from four independent experiments (bars show SD).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



