
Submitted:
11 April 2023
Posted:
12 April 2023
You are already at the latest version
Abstract
The Lin28B gene is involved in the initiation of puberty, but its regulatory mechanisms remain unclear. Therefore, in this study, we aimed to study the regulatory mechanism of the Lin28B promoter by cloning the Lin28B proximal promoter for bioinformatics analysis. Next, a series of deletion vectors were constructed based on the bioinformatics analysis results for dual-fluorescein activity detection. The transcriptional regulation mechanism of the Lin28B promoter region was analyzed by detecting mutations in transcription factor-binding sites and overexpression of transcription factors. The dual-luciferase assay showed that the Lin28B promoter region -837 to -338 bp had the highest transcriptional activity, and the transcriptional activity of the Lin28B transcriptional regulatory region decreased significantly after Egr1 and SP1 mutations. Overexpression of the Egr1 transcription factor significantly enhanced the transcription of Lin28B, and the results indicated that Egr1 and SP1 play an important role in regulating Lin28B. These results provide a theoretical basis for further research on the transcriptional regulation of sheep Lin28B during puberty initiation.
Keywords:
Dolang sheep
; Lin28B
; promoter
; transcription factor
1. Introduction
Puberty in female animals is the period when estrus appears for the first time and ovulation occurs. It plays an important role in animal growth and development. In addition, this is an important symbol for animals to acquire reproductive ability[1]. In the early stages, the hypothalamic tissue of Dolang sheep was used to screen the gene Lin28B related to the onset of puberty through transcriptome sequencing technology and high expression was detected in the hypothalamus, pituitary, ovary, and fallopian tube through qPCR and western blot technology. The Lin28B gene has a certain relationship with the initiation of puberty in Dolang sheep [2].
The Lin28B gene was first cloned from human hepatocellular carcinoma cells. It is overexpressed in hepatocellular carcinoma and is highly similar to the heterochronous gene Lin28. These two may originate from the duplication of the same ancestral gene. Its mRNA has a long 3'-UTR with the complementary site of Let7 microRNA, which may be the natural target gene of Let7. Lin28B contains a cold shock domain (CSD) and a pair of CCHC zinc finger domains[3]. Related studies have shown that Lin28B is a related gene that regulates puberty[4,5]. Sangiao-Alvarellos et al. (2013) found that Lin28B mRNA was abundantly expressed in the hypothalamus of neonatal male and female rats and that Lin28B expression decreased sharply during the transition from infancy to puberty[6]. Grieco et al. (2013) found that Lin28B expression decreases in the ovaries of female mice from infancy to puberty[7].
The promoter is usually located upstream of the 5′end of the structural gene, and is a nucleotide sequence that RNA polymerase recognizes and specifically binds to the template DNA to ensure the accuracy of transcription initiation[8] . Studies have shown that gene regulation at the transcriptional level is through the combination of TFBS DNA sequences, making the interaction between promoters and TFs[9]. At present, there is no related research on the promoter region of sheep Lin28B at home and abroad. In order to clarify the significance of the promoter region to the transcriptional regulation of the Lin28B, this study constructed a promoter deletion fragment vector to explore the core promoter region. To further elucidate the transcription regulation mechanism of Lin28B, this study predicted and verified the related transcription factors SP1 and Egr1, thus providing a theoretical basis for the regulation mechanism of sheep Lin28B on puberty initiation.
2. Materials and Methods
2.1. Ethics Statement
All animal experiments were approved by the Animal Protection and Use Committee of the Tarim University of Science and Technology College (Approval No. 2020.050).
2.2. Sample Collection
Fifteen female Dolang sheep maintained at the Tarim University experimental station were used as models in this study. Blood (5 mL) was collected from the jugular vein of each sheep, treated with EDTA-K anticoagulant (5:1 ratio of blood to EDTA-K (V/V)), and stored in a refrigerator at -20 ℃ for later use. Genomic DNA was extracted from Dolang sheep blood using a DNA extraction kit (Tiangen, Beijing, China) according to the manufacturer’s instructions. Its purity was detected using 1.5% agarose gel electrophoresis, and its content was detected using an ultraviolet spectrophotometer. Extracted DNA was stored at -20 ºC.
2.3. Lin28B promoter sequence analysis and vector construction
According to the promoter sequence of the sheep, the Lin28B gene in the Ensemble database, using the 2837 bp upstream and 162 bp downstream of the transcription start site as templates, six specific primers for deletion fragments were designed, namely L1 (-2837 to + 162 ), L2 (-2337 to +162), L3 (-1837 to +162), L4 (-1337 to +162), L5 (-837 to +162) and L6 (-337 to +162), and also designed for high transcriptional activity. The primer for the fragment was L7 (-837 to -338). The restriction endonuclease sites NheI and XhoI (Takara, Beijing, China) were added to the upstream and downstream primers, respectively, and protective bases were added before the enzyme cutting sites. Primer sequences are listed in Table 1. Using the conventional PCR method with the sheep Lin28B gene promoter in the Ensemble database as a template, deletion fragments of different lengths of the Duolang sheep Lin28B gene promoter were amplified. PCR system (25 μL ): DNA 1.0 µL, upstream and downstream primers (10 µmol/L) each 1.0 µL, 2× EasyTaq PCR SuperMix (+dye) 12.5 µL, sterile deionized water (ddH2O) 9.5 µL. PCR conditions: pre-denaturation at 94 ºC for 5 min; denaturation at 94 ºC for 30 s, annealing at 60 ºC for 30 s, extension at 72 ºC for 2 min, 40 cycles; extension at 72 ºC for 10 min.
2.4. Vector construction and transfection
The agarose gel was subjected to gel electrophoresis and recovered. The recovered target fragment and pGL3-Basic (Promega) were double-digested overnight with NheI and XhoI. The target fragment and carrier were recovered by the solution recovery method, the concentration was measured by NanoDrop 2000, and the connection was carried out at 4 ℃ for over 12 h. Afterward, the ligation product was transformed into DH5α, spread on an LB solid petri dish, cultured at 37 ºC for 12 h, then picked as a single colony with a regular shape and appropriate size for propagation, and the plasmid was extracted. The correct plasmid, identified by enzyme digestion, was sent to the Shanghai Sangon Bioengineering Company for sequencing. Plasmids with correct sequencing were amplified in large quantities and stored at -20 ºC for later use.
2.5. Cell Transfection and Dual-Luciferase Detection of Deletion Fragment Luciferase Reporter Gene Vector
The day before transfection, seed 1 × 10 4 293T cells (provided by the Chinese Academy of Sciences Cell Bank) in a 96-well plate were shaken gently to prevent cells from accumulating in the center of the well and causing massive cell death. They were cultured in a 37 ºC constant temperature incubator for 24 h. When the confluence of the cells reached 70%–80%, the cells were transfected according to the instructions of Lipofectamine™ 3000 transfection reagent (Invitrogen, USA) with at least three biological repeats. The 96-well plate was taken out of the incubator 36 h after transfection, and the dual-luciferase activity was detected according to the instructions of the Dual-Glo® Luciferase Assay System kit (Promega, USA).
2.6. Transcription factor prediction of highly transcriptionally active fragments
Using the combination of the online software JASPAR and Animal TFDB 3.0 to predict the L7 (-837bp to -338 bp) fragment of the Lin28B promoter, the possible binding sites of transcription factors related to the regulation of puberty were obtained with higher scores.
2.7. Construction of Point Mutation Vectors for Transcription Factors SP1 and Egr1
The assay was performed according to the manufacturer’s instructions for the Express Point Mutation Kit (Tiangen, Beijing, China). Primer design instructions, using Primer Premier software (version 6.0) to design transcription factor EGR1 primers ETF1 and ETR1, transcription factor SP1 primers, transcription factor point mutation primers (Table 2) for STF2 and STR2, and construct mutation vectors ET1 and ST2, respectively.
According to the instructions of Lipofectamine™ 3000 transfection reagent, were co-transfected with pGL3-basic, plasmid L7, mutant plasmids ET1, ST2, respectively, and pRL -TK (Promega, USA) and at least three biological replicates were set up for each treatment. The 96-well plate was removed from the incubator 36 h after transfection, and dual-luciferase activity was detected according to the instructions of the Dual-Glo Luciferase Assay System kit.
2.8. Construction of transcription factor Egr1 overexpression vector and dual luciferase detection
The Egr1 overexpression vector was constructed by Suzhou GENEWIZ Co., Ltd. The Egr1 overexpression vector, pEGFP-N1 and plasmid L7 were co-transfected into 293T cells according to the instructions of Lipofectamine™ 3000 transfection reagent, and at least three biological replicates were set up for each treatment. The 96-well plate was removed from the incubator 36 h after transfection, and dual-luciferase activity was detected according to the instructions of the Dual-Glo Luciferase Assay System kit.
2.9. Data Analysis
Relative activity value of dual luciferases = F value/R value. The data were analyzed using SPSS and expressed as the standard error of the mean. At least three biological replicates were set up for each treatment group, and significance analysis was performed using a one-way ANOVA. *P<0.05 and **P<0.01 indicated significant and extremely significant differences, respectively.
3. Results
3.1. Construction of the Deletion Vector of Lin28B Gene Promoter Fragment in Dolang Sheep
To determine the core region of the Lin28B promoter, we gradually reduced the length of the promoter using a deletion mutation and measured its relative activity. As shown in Figure 1, using the sheep Lin28B promoter fragment as a template, a series of deletion primers were used to clone Lin 28 gene promoter deletion fragments of different lengths, L 1 (-2837 to +16 2 ), L 2 (-2337 to +162 ), L 3 (- 1837 to +162 ), L 4 (- 1337 to +162 ), L 5 (-837 to +162 ), L 6 (-337 to +162 ) and L 7(-837 to-338 ). The missing fragment of the Lin28B promoter was connected to a luciferase reporter gene vector to obtain the corresponding recombinant vector.
3.2. Detection of dual-luciferase activity in the deletion vector of Lin28B gene promoter fragment in Dolang sheep
The successfully constructed promoter deletion fragment vector was transfected into 293T cells. After incubation for 36 h, the promoter activity of each fragment was detected (Figure 2). The results showed that the L1 and L6 promoters had the lowest activity, whereas the L2 and L5 promoters had the highest activity. Furthermore, the results found that from the deletion of -887bp to -338bp, promoter activity dropped suddenly and decreased approximately once. The above results indicate that the -887 to -338 bp region may play a very important role in maintaining the transcriptional activity of Lin28B, and further experiments are needed to prove this. It is the core region of the promoter.
3.3. Dual luciferase activity detection of fragment vectors with high transcriptional activity
The promoter deletion fragment vectors L5, L6, and L7 were transfected into 293T cells, and the relative activity of the dual luciferase was detected (Figure 3). The promoter activity of L5 was approximately the same as the sum of the promoter activities of L6 and L7, and the promoter activity of L5 and L7 was significantly higher than that of L6. The above test results showed that the L7 (-887 to -338) fragment is the core promoter region, which plays an important role in the transcriptional regulation of Lin28B in Dolang sheep.
3.4. Prediction of transcription factor binding sites in Lin28B promoter region of Dolang sheep
The online software JASPAR and Animal TFDB 3.0 were used to predict the transcription factor binding site of the Lin28B promoter L7 (-837bp to -338 bp) fragment and the transcription factors related to the transcriptional regulation of puberty with higher scores were selected. The results are summarized in Table 3. Two transcription factors were mainly predicted, namely transcription factors SP1 and Egr1. This study selected the transcription factors SP1 and Egr1 related to transcriptional regulation during puberty. These transcription factors may play a role in transcription by binding to the promoters.
3.5. Verification of the point mutation of the promoter transcription factor in the core region of the Lin28B promoter of Dolang sheep
According to the software to predict the transcripts of the Lin28B promoter core region, the transcription factors SP1 and Egr1 were selected to verify the correct existence of the two transcription factor binding sites in the -837 to -338bp promoter core region, according to the sequence of the core region and the prediction results. For the sequence of the transcription factor binding site, we conducted site-directed mutagenesis analysis on the transcription factor binding sequence to verify the mutation of 2-3 bases. Using the L7 fragment as a plate, a mutation kit was used to construct a mutation vector for the two L7 fragment transcription factors. The mutant vectors were transfected into 293T cells, and the L7 fragment was used as a control. The promoter activity of the mutant and normal groups was detected using a double luciferase assay, and significant differences were compared. The results showed that the promoter activity of EGR1 and SP1 mutations compared with the non-mutated fragment, the difference was extremely significant (P<0.01), and the difference between the L7 fragment and the EGRI mutant vector was more significant (Figure 4), indicating that these two sites may regulate the promoter activity by combining transcription factors SP1 and Egr1. Egr1 has a greater influence on the activity of the Lin28B promoter than SP1; thus, the EGR1 transcription factor was selected to construct an overexpression vector to further verify the role of transcription factor binding sites.
3.6. Effect of Egr1 overexpression on the transcriptional activity of Lin28B promoter
In site-directed mutation verification of the transcription factors, mutation of the two transcription factors led to a significant decrease in Lin28B promoter activity. Therefore, we preliminarily determined that these two transcription factors affect the transcriptional regulation of the Lin28B promoter. To further verify the possible existence of two transcription factors in the Lin28B gene promoter for the factor binding site, based on point mutations, an overexpression vector of the transcription factor Egr1 was constructed to detect whether there was a significant difference between the promoter activity of the overexpression group and the promoter activity of the normal L7 fragment group after the transcription factor was overexpressed. The cells were co-transfected with the Egr1 overexpression vector and the L7 fragment, and the control group was co-transfected with the blank vectors pEGFP-N1 and the L7 fragment. The results showed that the fluorescence activity ratio of the L7 fragment co-transfected with the Egr1 overexpression vector was significantly higher (P<0.01) than that of the control group co-transfected with the pEGFP-N1 and L7 fragments (Figure 5), indicating that overexpression of Egr1 has an impact on the transcriptional activity of the L7 fragment. It has been speculated that an Egr1 transcription factor binding site in the Lin28B promoter region can affect the transcriptional activity of the Lin28B promoter, which may play a certain role in the transcriptional regulation of the Lin28B promoter.
4. Discussion
The identification of promoters is key to the regulation of gene transcription[10]. Luciferase analysis is a common method for studying promoter core and regulatory regions[11,12]. The dual-luciferase reporter assay used the firefly luciferase (pGL3) to kidney luciferase (pRL-TK) ratio to determine gene expression. This method can reduce experimental errors caused by factors such as cell activity, transcription efficiency, and cleavage efficiency, and the test results are reliable and accurate[13]. In this study, a promoter deletion fragment vector was constructed and transfected into cells. Double luciferase detection revealed that the L1 (-2837 to +162) and L6 (-337 to +162) vectors had the least activity, whereas the L2 (-2337 to +162) and L5 (-837 to +162) had the most active promoters. There was a significant difference in the promoter activity of L7 (-837 to -338bp) and L5 (-837 to +162 bp), and the core promoter region was generally located in the upstream 500 bp[14]; thus, we infer that L7 (-837 to -338bp) is the core promoter region of the gene.
A gene promoter is a complex region containing many transcription factor-binding sites determining the transcription start site and frequency[15]. Transcription factors (TFs) can specifically recognize the active region of the gene promoter, thereby activating or inhibiting the transcriptional activity of the target gene, regulating the transcription level of gene mRNA, and affecting the protein’s function [16]. Transcriptional regulation of genes is affected by the activity of different transcription sites in the promoter region. In this study, JASPAR and Animal TFDB 3.0 were used to predict the transcription factor-binding site of the Lin28B promoter L7 (-837 bp to -338 bp) fragment, and the intersection of higher scores was selected to obtain the two transcription factors Egr1 and SP1. The Erg1 transcription factor was point-mutated in this study, and an overexpression vector was constructed. Double-luciferase detection results showed that the Egr1 transcription factor plays an important role in the transcriptional regulation of Lin28B. According to relevant studies, Egr1 plays an important role in the reproductive system, and Egr1 can maintain sufficient LHb expression in the female pituitary [17,18]. During proestrus in rats, GnRH stimulates the expression of Egr1 and simultaneously binds Egr1 to two conserved cis-acting elements of the LH proximal promoter [19-21]. Egr1 knockout mice showed reduced fertility, blocked GnRH-induced pituitary LH production, and impaired ovarian LH receptor expression. Egr1 regulates the expression of many important factors involved in folliculogenesis and ovulation in the ovary [17,22,23]. Therefore, this study suggests that the Egr1 transcription factor plays an important role in the transcriptional regulation of Lin28B during puberty in sheep. In this study, we constructed a point mutation vector for the transcription factor SP1. After testing, it was found that the SP1 transcription factor was found to affect the transcriptional regulation of Lin28B. Studies have reported that SP1 interacts with various epigenetic factors, actively regulates gene transcription, and promotes gene expression [24]. Further studies have shown that SP1 interacts with epigenetic factors that negatively regulate gene transcription to inhibit gene expression [25,26]. During primordial follicle formation in the ovary, SP1 is present in oocytes and somatic cells. Knockdown of SP1 expression, especially in pregranulosa cells, significantly inhibits oocyte apoptosis and primordial follicle formation, suggesting that somatically expressed SP1 plays a role in primordial folliculogenesis clarifying that SP1 regulates pregranulosa cells during mammalian ovarian development to control the establishment of ovarian reserves [27]. Therefore, our results suggest that SP1 plays a role in the transcriptional regulation of Lin28B during puberty in sheep.
5. Conclusions
In this study, by constructing a promoter fragment deletion vector, we found that -887 to -338bp was a region with high transcriptional activity. To further understand the transcriptional regulatory mechanism of Lin28B, we constructed a transcription factor point mutation vector and an Egr1 overexpression vector. Double luciferase assay revealed that the transcription factors SP1 and Egr1 can significantly affect the activity of the promoter. Therefore, this study suggests that the transcription factors SP1 and Egr1 play a role in the transcriptional regulation of Lin28B during sheep puberty.
Supplementary Materials
Not applicable
Author Contributions
Conceptualization, Z.S., F.X. and M.C.; data curation, Z.S., Y.Z., Z.Z. and C.W.; investigation, Y.Z., Z.Z., C.W. and J.L.; supervision, Y.Z., Z.Z., C.W. and J.L.; visualization, Z.S., Y.Z. and Z.Z.; writing—original draft, Z.S., F.X. and M.C.; writing—review and editing, Z.S., F.X., M.C and Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The study was made possible thanks to funding from the National Natural Science Foundation of China (Grant/Award Numbers: 31960655)
Institutional Review Board Statement
This study was conducted following specifications of the Ethics Committee of Tarim University of Science and Technology (NO. 2020.050).
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable
Acknowledgments
The study was made possible thanks to funding from the National Natural Science Foundation of China (Grant/Award Numbers: 31960655)
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sui, Z.; Zhang, Z.; Zhang, Y.; Zhang, J.; Li, Q.; Xing, F. Analysis of Methylation and mRNA Expression of Lin28B Gene Promoter Region in the Hypothalamus of Dolang Sheep During Pubertal Initiation. DNA Cell Biol 2023, 42, 130–139. [Google Scholar] [CrossRef]
- Xing, F.; Zhang, C.; Kong, Z. Cloning and expression of lin-28 homolog B gene in the onset of puberty in Duolang sheep. Asian-Australas J Anim Sci 2019, 32, 23–30. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, Y.; Ito, H.; Watanabe, A.; Ge, X.; Kodama, T.; Aburatani, H. Identification and characterization of lin-28 homolog B (LIN28B) in human hepatocellular carcinoma. Gene 2006, 384, 51–61. [Google Scholar] [CrossRef]
- Ong, K.K.; Elks, C.E.; Li, S.; Zhao, J.H.; Luan, J.a.; Andersen, L.B.; Bingham, S.A.; Brage, S.; Smith, G.D.; Ekelund, U. Genetic variation in LIN28B is associated with the timing of puberty. Nature Genetics 2009, 41, 729–733. [Google Scholar] [CrossRef]
- Ong, K.K.; Elks, C.E.; Wills, A.K.; Wong, A.; Wareham, N.J.; Loos, R.J.F.; Kuh, D.; Hardy, R. Associations between the Pubertal Timing-Related Variant in LIN28B and BMI Vary Across the Life Course. J. Clin. Endocrinol. Metab. 2011, 96, E125–E129. [Google Scholar] [CrossRef]
- Sangiao-Alvarellos, S.; Manfredi-Lozano, M.; Ruiz-Pino, F.; Navarro, V.M.; Sanchez-Garrido, M.A.; Leon, S.; Dieguez, C.; Cordido, F.; Matagne, V.; Dissen, G.A.; et al. Changes in Hypothalamic Expression of the Lin28/let-7 System and Related MicroRNAs During Postnatal Maturation and After Experimental Manipulations of Puberty. Endocrinology 2013, 154, 942–955. [Google Scholar] [CrossRef]
- Grieco, A.; Rzeczkowska, P.; Alm, C.; Palmert, M.R. Investigation of peripubertal expression of Lin28a and Lin28b in C57BL/6 female mice. Mol. Cell. Endocrinol. 2013, 365, 241–248. [Google Scholar] [CrossRef]
- Lemon, B.; Tjian, R. Orchestrated response: a symphony of transcription factors for gene control. Genes Dev 2000, 14, 2551–2569. [Google Scholar] [CrossRef]
- Dong, W.H.; Yin, X.M.; Sun, L.; Wang, J.; Sun, S.Y.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Age-associated methylation change of TAP1 promoter in piglet. Gene 2015, 573, 70–74. [Google Scholar] [CrossRef]
- Haberle, V.; Lenhard, B. Promoter architectures and developmental gene regulation. Semin. Cell Dev. Biol. 2016, 57, 11–23. [Google Scholar] [CrossRef]
- Wang, J.M.; Lang, B.; Zhu, H.Y.; Du, H.T.; Tian, Y.M.; Su, Y.H. Cloning and transcriptional activity analysis of the porcine cofilin 2 gene promoter. Gene 2014, 547, 280–287. [Google Scholar] [CrossRef]
- Annicotte, J.; Schoonjans, K.; Haby, C.; Auwerx, J. An E-box in pGL3 reporter vectors precludes their use for the study of sterol regulatory element-binding proteins. Biotechniques 2001, 31, 993–994. [Google Scholar]
- Wei, X.F.; Zhu, Y.C.; Du, J.; Ma, X.J.; Zhao, X.; Ma, Y.Y.; Han, S.; Ma, Y. Analysis of ANGPTL8 promoter activity and screening of related transcription factors in bovine. Gene 2021, 784, 6. [Google Scholar] [CrossRef]
- Xiang, G.M.; Huang, L.; Zhang, X.L.; Wang, N.; Wang, H.; Mu, Y.L.; Li, K.; Liu, Z.G. Molecular Characteristics and Promoter Analysis of Porcine COL1A1. Genes 2022, 13, 14. [Google Scholar] [CrossRef]
- Inukai, S.; Kock, K.H.; Bulyk, M.L. Transcription factor-DNA binding: beyond binding site motifs. Curr. Opin. Genet. Dev. 2017, 43, 110–119. [Google Scholar] [CrossRef]
- Yang, L.; Orenstein, Y.; Jolma, A.; Yin, Y.M.; Taipale, J.; Shamir, R.; Rohs, R. Transcription factor family-specific DNA shape readout revealed by quantitative specificity models. Mol. Syst. Biol. 2017, 13, 14. [Google Scholar] [CrossRef]
- Lee, S.L.; Sadovsky, Y.; Swirnoff, A.H.; Polish, J.A.; Goda, P.; Gavrilina, G.; Milbrandt, J. Luteinizing hormone deficiency and female infertility in mice lacking the transcription factor NGFI-A (Egr-1). Science 1996, 273, 1219–1221. [Google Scholar] [CrossRef]
- Topilko, P.; Schneider-Maunoury, S.; Levi, G.; Trembleau, A.; Gourdji, D.; Driancourt, M.A.; Rao, C.V.; Charnay, P. Multiple pituitary and ovarian defects in Krox-24 (NGFI-A, Egr-1)-targeted mice. Mol Endocrinol 1998, 12, 107–122. [Google Scholar] [CrossRef]
- Slade, J.P.; Carter, D.A. Cyclical expression of egr-1/NGFI-A in the rat anterior pituitary: a molecular signal for ovulation? J Neuroendocrinol 2000, 12, 671–676. [Google Scholar] [CrossRef]
- Wolfe, M.W.; Call, G.B. Early growth response protein 1 binds to the luteinizing hormone-β promoter and mediates gonadotropin-releasing hormone-stimulated gene expression. Molecular Endocrinology 1999, 13, 752–763. [Google Scholar]
- Tremblay, J.J.; Drouin, J. Egr-1 is a downstream effector of GnRH and synergizes by direct interaction with Ptx1 and SF-1 to enhance luteinizing hormone β gene transcription. Molecular and Cellular Biology 1999, 19, 2567–2576. [Google Scholar] [CrossRef] [PubMed]
- Fortin, J.; Lamba, P.; Wang, Y.; Bernard, D.J. Conservation of mechanisms mediating gonadotrophin-releasing hormone 1 stimulation of human luteinizing hormone beta subunit transcription. Mol. Hum. Reprod. 2009, 15, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Tourtellotte, W.G.; Nagarajan, R.; Bartke, A.; Milbrandt, J. Functional compensation by Egr4 in Egr1-dependent luteinizing hormone regulation and Leydig cell steroidogenesis. Molecular and Cellular Biology 2000, 20, 5261–5268. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Kimura, A.; Nagai, R.; Horikoshi, M. Regulation of interaction of the acetyltransferase region of p300 and the DNA-binding domain of Sp1 on and through DNA binding. Genes to Cells 2000, 5, 29–41. [Google Scholar] [CrossRef]
- He, S.; Sun, J.-M.; Li, L.; Davie, J.R. Differential intranuclear organization of transcription factors Sp1 and Sp3. Molecular Biology of the Cell 2005, 16, 4073–4083. [Google Scholar] [CrossRef]
- Li, L.; Davie, J.R. The role of Sp1 and Sp3 in normal and cancer cell biology. Ann. Anat. 2010, 192, 275–283. [Google Scholar] [CrossRef]
- Cai, H.; Liu, B.Y.; Wang, H.R.; Sun, G.H.; Feng, L.Z.; Chen, Z.Q.; Zhou, J.Q.; Zhang, J.W.; Zhang, T.; He, M.N.; et al. SP1 governs primordial folliculogenesis by regulating pregranulosa cell development in mice. J. Mol. Cell Biol. 2020, 12, 230–244. [Google Scholar] [CrossRef]
Figure 1.
Figure 1. Dolang sheep Lin28B promoter fragment deletion vector electrophoresis map. Note: M, marker 5000; L1(-2837 to +162), L2(-2337 to +162), L3(-1837 to +162), L4(-1337 to +162), L5(-837 to +162), L6(-337 to +162), and L7(-837 to -338).
Figure 1.
Figure 1. Dolang sheep Lin28B promoter fragment deletion vector electrophoresis map. Note: M, marker 5000; L1(-2837 to +162), L2(-2337 to +162), L3(-1837 to +162), L4(-1337 to +162), L5(-837 to +162), L6(-337 to +162), and L7(-837 to -338).
Figure 2.
The activity analysis of different Dolang sheep Lin28B promoter lengths. Note: The left side is a schematic diagram of the length of each deletion fragment, and the right side is the promoter activity of the corresponding length; different lowercase letters indicate significant differences (P<0.05).
Figure 2.
The activity analysis of different Dolang sheep Lin28B promoter lengths. Note: The left side is a schematic diagram of the length of each deletion fragment, and the right side is the promoter activity of the corresponding length; different lowercase letters indicate significant differences (P<0.05).
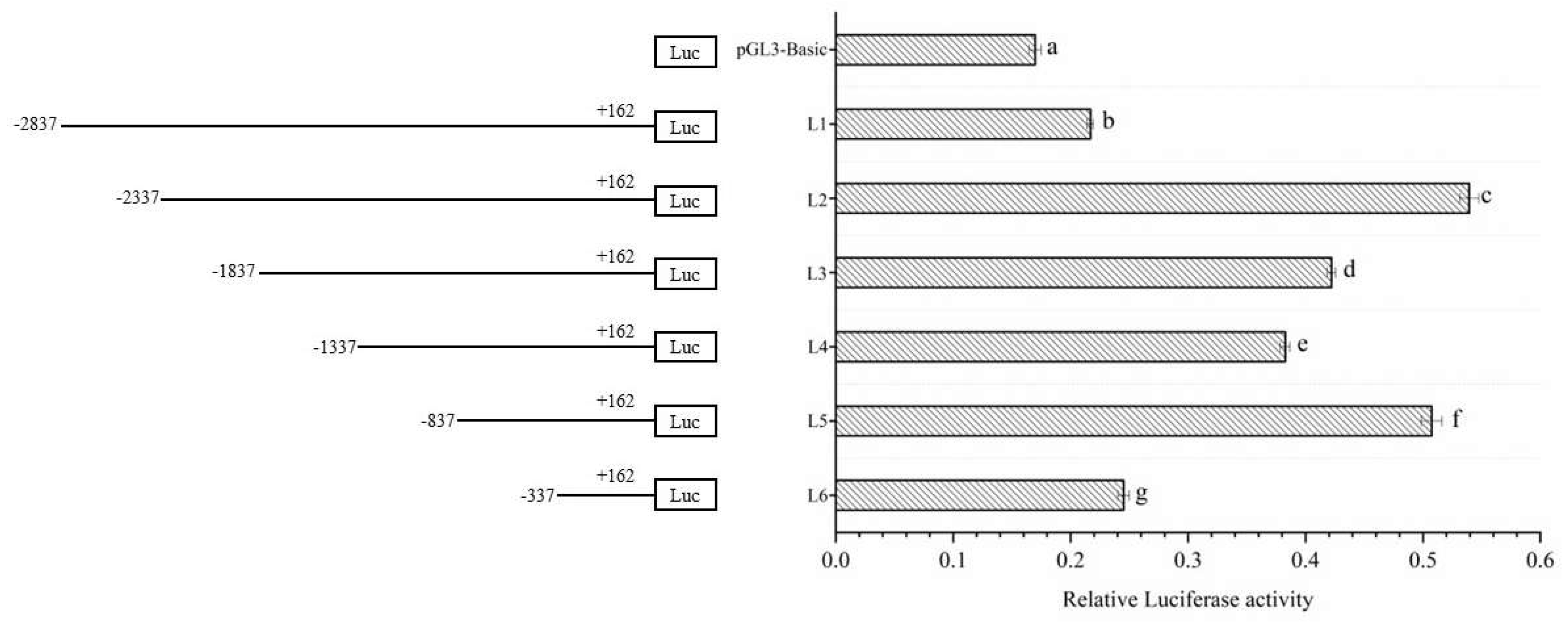
Figure 3.
The activity analysis of the second time different lengths of Dolang sheep Lin28B promoter. Note: The left side is a schematic diagram of the length of each deletion fragment, and the right side is the promoter activity of the corresponding length; different lowercase letters indicate significant differences (P<0.05).
Figure 3.
The activity analysis of the second time different lengths of Dolang sheep Lin28B promoter. Note: The left side is a schematic diagram of the length of each deletion fragment, and the right side is the promoter activity of the corresponding length; different lowercase letters indicate significant differences (P<0.05).
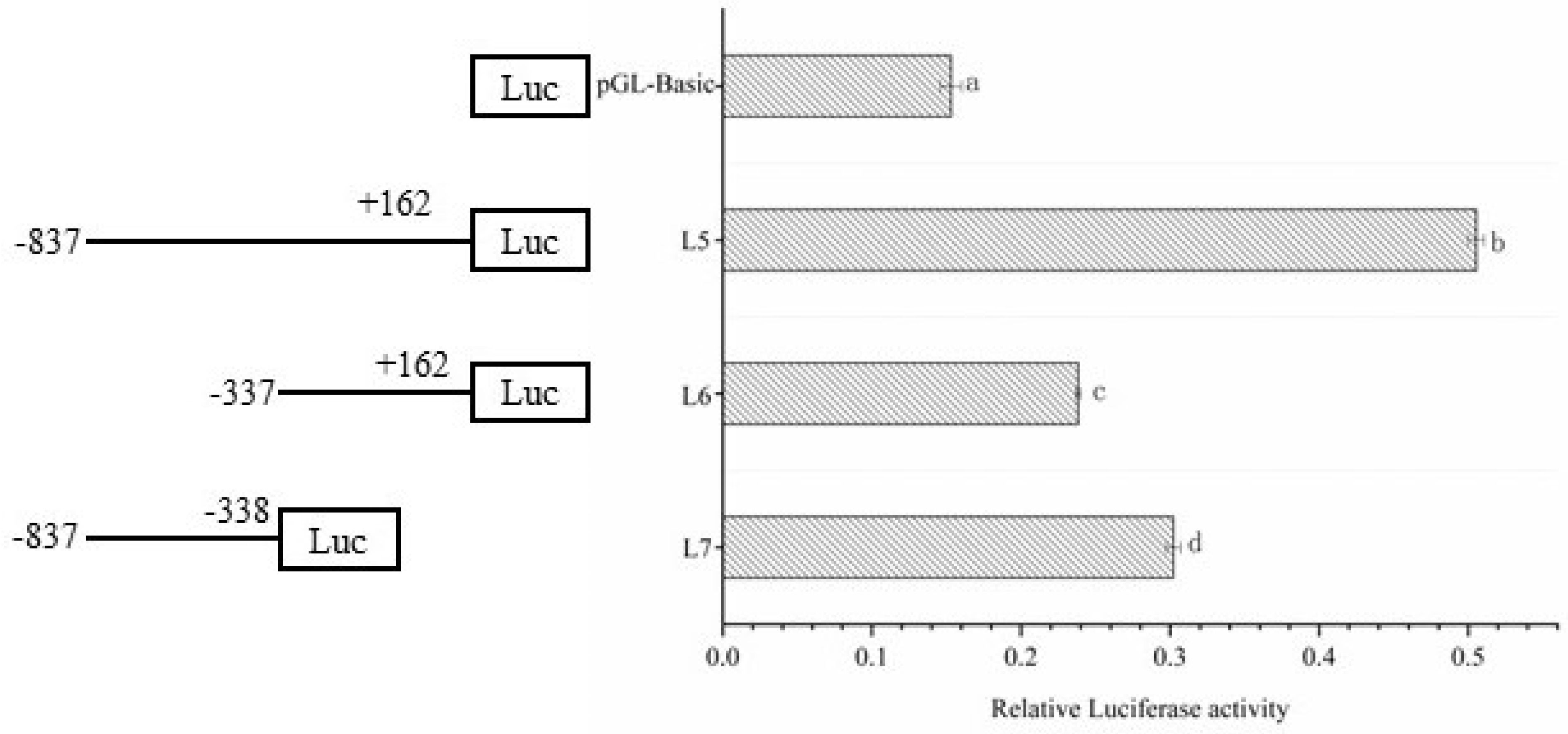
Figure 4.
The point mutation analysis of Egr1 and SP1.
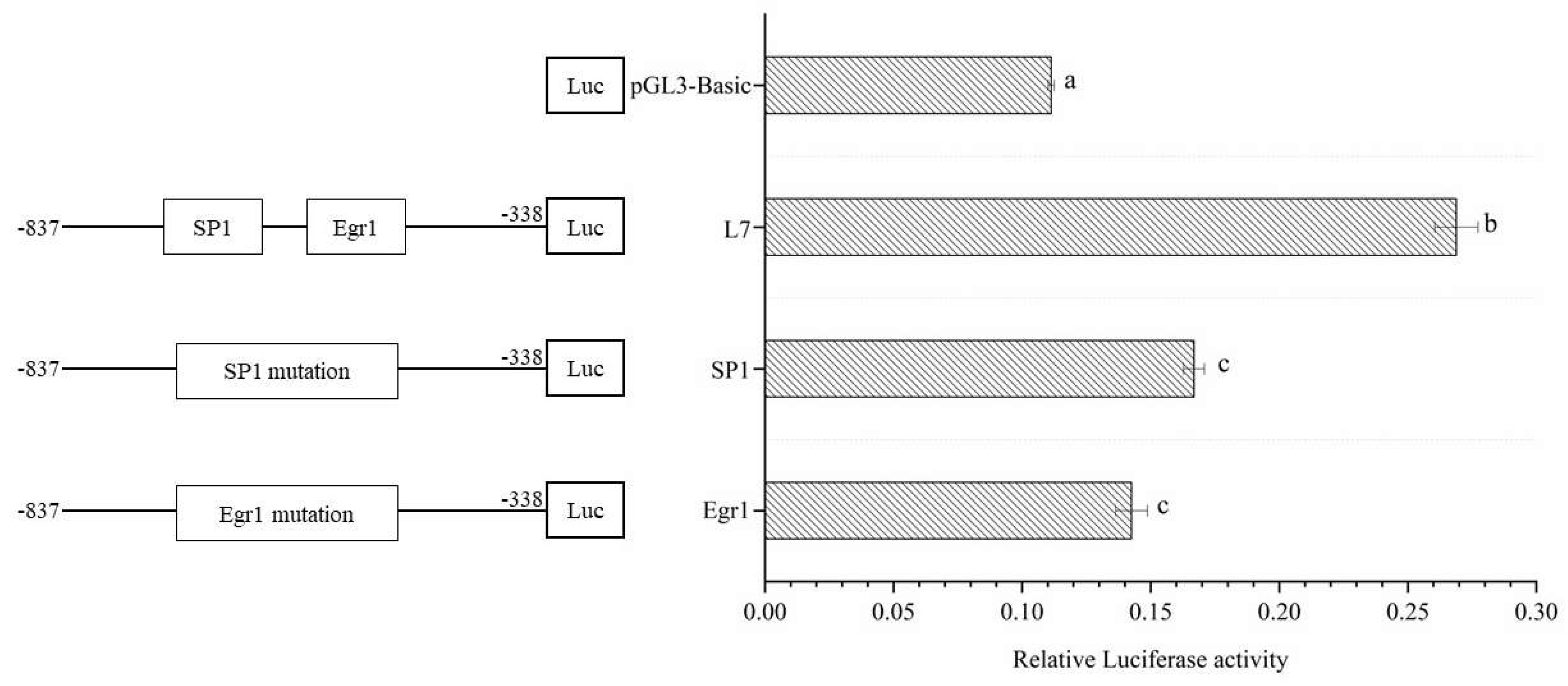
Figure 5.
Promoter activity analysis of Egr1 overexpression.
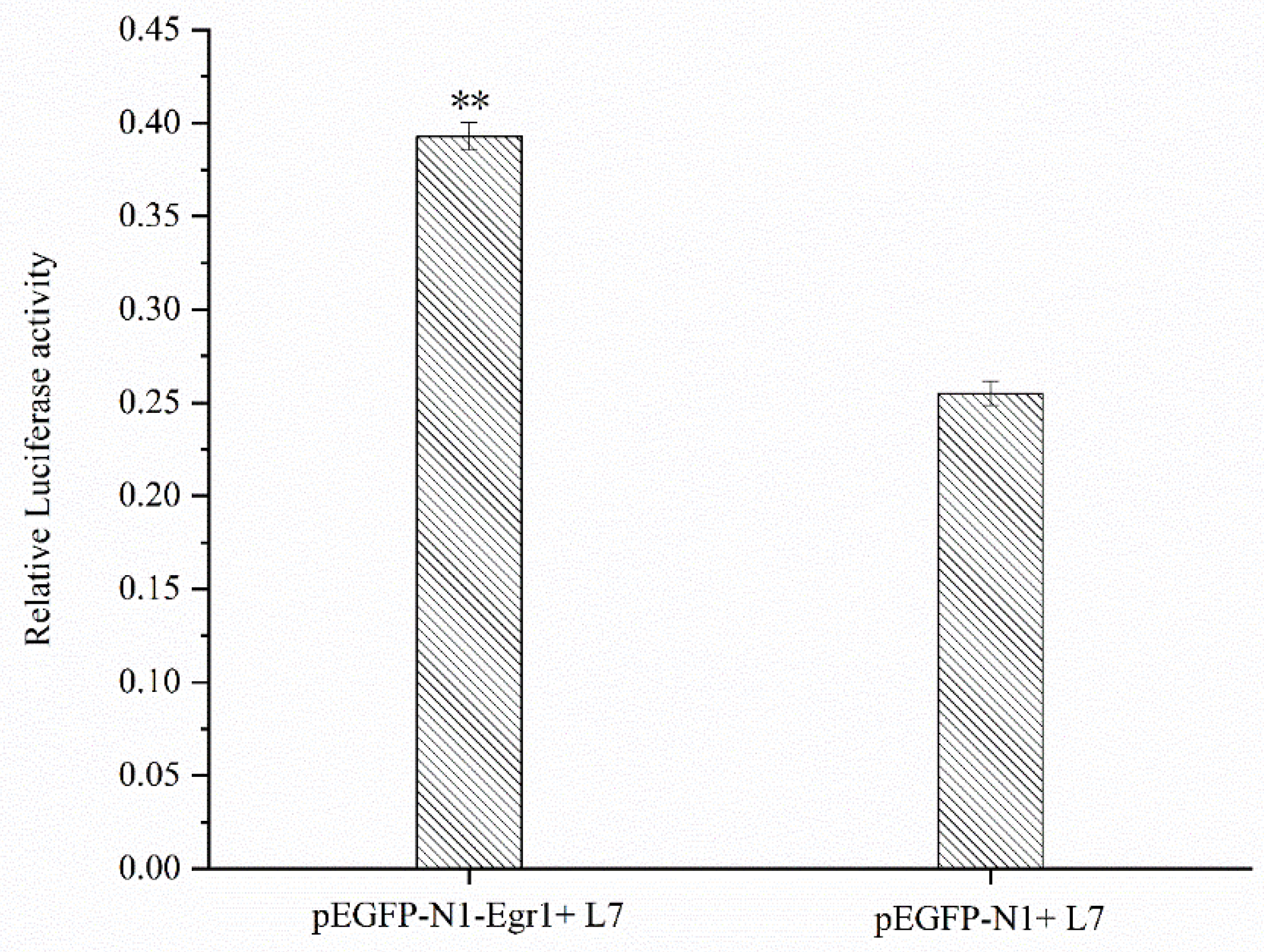

Table 1.
Primers used for fragment deletion of sheep Lin28B gene promoter constructs.
| Primer name | Primer sequence (5’-3’) | Product length (bp) |
|---|---|---|
| L1 | F: CTAgctagcGATAACCAACGGGCATTTA R: CCGctcgagCTGGCAAGAGGAAGAGATAAC |
3012 |
| L2 | F: CTAgctagcCTCCCTCTCCTCTCCCCTCCT CTCCTTTCTTTTACTCTCCACA R: CCGctcgagCTGGCAAGAGGAAGAGATAAC |
2512 |
| L3 | F: CTAgctagcTTTATGATAATTTTACATAATGA CATGTCCTCA R: CCGctcgagCTGGCAAGAGGAAGAGATAAC |
2012 |
| L4 | F: CTAgctagcTATACATCCATTTATTTC AGATCTGAACTAATTAATTGTCCAT R: CCGctcgagCTGGCAAGAGGAAGAGATAAC |
1512 |
| L5 | F: CTAgctagcAGGGACGGTAGGAGCCT AATCCGTTATT R: CCGctcgagCTGGCAAGAGGAAGAGATAAC |
1012 |
| L6 | F: CTAgctagcCAGGGCACAATCAGGTACT TGTGT R: CCGctcgagCTGGCAAGAGGAAGAGATAAC |
512 |
| L7 | F: CTAgctagcAGGGACGGTAGGAGCCTAAT CCGTTATT R: CCGctcgagAAATATTTCTCAAATTTAAAAT AAAATCCTACCGGAAAAATCGCT |
512 |
Table 2.
Information on site-directed mutation primers of porcine BMP7 promoter.
| Primer name | Primer sequence(5’-3’) |
|---|---|
| ETF1 | GCCTGCCTGGCGCTCCCTTCTCCCTCCCTGCCCATACATA |
| ETR1 | R: TATGTATGGGCAGGGAGGGAGAAGGGAGCG CCAGGCAGGC |
| STF2 | F: GGCCCCTCTTTTTCACCTCGCTCCCCAGCCTCGCCCGCTG |
| STR2 | R: CAGCGGGCGAGGCTGGGGAGCGAGGTGAAAAAGAGGGGCC |
Table 3.
Prediction of transcription factors in the regulatory region of the Lin28B promoter in Dolang sheep.
Table 3.
Prediction of transcription factors in the regulatory region of the Lin28B promoter in Dolang sheep.
| Transcription Factors | Start | Stop | Strand | Matched sequence(5’ - 3’) |
|---|---|---|---|---|
| SP1 | 142 | 151 | + | CCCCTCCCCC |
| SP1 | 219 | 228 | + | CCCCTCCCCC |
| Egr1 | 218 | 231 | + | TCCCCTCCCCCTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



