Submitted:
14 April 2023
Posted:
14 April 2023
You are already at the latest version
Abstract
Mitochondrial alternative oxidase 1a (AOX1a) plays an extremely important role in critical node of seed viability. However, the regulatory mechanism is still poorly understood. The study aims to identify regulatory mechanisms by comparing between OsAOX1a-RNAi and wild type (WT) rice seed during artificial aging treatment. Gain weight and P50 significantly decreased in OsAOX1a-RNAi rice seed, indicating that there might be impaired in seed development and storability. Compared to WT seeds in the 100%, 90%, 80%, and 70% germination, respectively, NADH and succinate-dependent O2 consumption, the activity mitochondrial malate dehydro-genase and ATP contents were decreased in OsAOX1a-RNAi seeds in the 100%, 90%, 80%, and 70% germination, respectively, indicating that mitochondrial status in the OsAOX1a-RNAi seeds after imbibition was weaken than the WT. In addition to, the reduced abundance of complex I N and P module subunits might showed that the capacity of mitochondrial electron transfer chain was significantly inhibited in the OsAOX1a-RNAi seed at critical node of seed viability. Above results might indicated that the ATP production was impaired in OsAOX1a-RNAi seeds during ageing. Therefore, we conclude that the activities of mitochondrial metabolism and alternative pathways were severely inhibited in OsAOX1a-RNAi seeds at critical node of viability, which was leading to accelerate the collapse of seed viability. The precise regulatory mechanism of the alternative pathway at the critical node of viability still needs to be further analyzed.
Keywords:
alternative oxidase
; mitochondria
; critical node
; rice
1. Introduction
Mitochondrial redox homeostasis is a key factor affecting seed viability. During the imbibition process of seeds with high viability, the number, inner membrane and cristae of mitochondria were significantly improved, and proteins and genes involved in energy and material metabolism, enzyme activity of TCA cycle, electron transport chain and oxidative phosphorylation were significantly improved [1,2]. However, in the process of imbibition, aging seeds inhibit the repair of mitochondrial structure and function, the total number of mitochondria, the outer membrane and inner membrane are difficult to distinguish and the integrity is low, the number of matrix and cristae is less than that of high viability seeds, and the shape is irregular. The proportion of mitochondria increased and the motility decreased, and the physiological functions such as electron transport chain, oxidative phosphorylation and marker enzymes were also lower than normal seeds, and mitochondrial cytochrome c was released into the cytoplasm, inducing programmed cell death [3,4,5].
With the decrease of seed viability, the activities of SOD, APX, MDHAR and GR of mitochondrial antioxidant system decreased significantly, and the total amount of ascorbic acid and glutathione decreased, and the ratio of reduced and oxidized forms also decreased significantly, resulting in the accumulation of ROS [6,7]. The accumulation of ROS will oxidize lysine, arginine, and proline in the protein side chain, resulting in protein carbonylation modification, inducing protein peptide chain breakage, and changing the oxidation site to carbonyl [8]. Carbonylation leads to changes in protein structure and loss of function, which are easily degraded by hydrolases, leading to physiological and metabolic disorders of cells and tissues, aging or death [9]. Previous studies revealed that when rice seed viability drops to a critical node (CN, 85% germination), ROS accumulate and attack lipids, and 4-HNE and other carbonyl small molecules accumulate, resulting in mitochondrial ATP synthase, malate dehydrogenase, and succinate dehydrogenase, etc. The protein undergoes carbonylation modification, which inhibits the ATP supply required for seed germination [10,11].
Mitochondrial redox homeostasis is closely related to the mitochondrial electron transport chain (mETC). Plant mETC mainly includes cytochrome pathway and alternate oxidase pathway. The cytochrome pathway is the main pathway of mitochondrial electron transfer, electrons are transferred from reduced substrates (NADH and succinate) to complex I or complex II, and then to ubiquinone, complex III, cytochrome c (Cyt c), complex IV, and finally transfer electrons to O2 to generate H2O, while protons are pumped from the mitochondrial matrix to the mitochondrial intermembrane space, forming proton kinetic potential on both sides of the inner membrane, driving the synthesis of ATP [12,13]. Under stress, the cytochrome pathway is inhibited, the oxidative capacity of NADH is reduced, the transfer of electrons to ubiquinone is inhibited, the proportion of oxidized ubiquinone is increased, and the transfer of electrons to O2 is promoted to generate O2-, resulting in oxidative damage; at the same time induces the alternative oxidase pathway, which facilitates electron transfer to alternative oxidase (AOX), and the pumping of protons from the mETC membrane arm to generate ATP [14,15,16]. Kühn et al. [17] to reveal the molecular mechanism by which the redox homeostasis of aox1a mutants regulates the cytochrome pathway and the alternate oxidase pathway under stress in Arabidopsis.
Different plants contain different numbers of AOX genes. For example, the rice genome contains four AOX genes, OsAOX1a, OsAOX1c, OsAOX1d (previously named OsAOX1b), and OsAOX1e [18]. AOX1a plays an important role in response to stress in Arabidopsis and rice. The previously reports revealed that the redox homeostasis of critical node of rice seed viability was significantly altered, inhibiting the cytochrome pathway, and the expression of OsAOX1a was significantly up-regulated than that of unaged seeds, and the expression of the core subunit of the membrane arm of mETC complex I significantly up-regulated, improved electron and proton transfer ability, and maintained ATP generation [5,19].
Previous work confirmed that the cytochrome pathway, ROS accumulation, and mitochondrial protein carbonylation were inhibited during the critical node of rice seed viability, but AOX1a was induced to maintain mitochondrial redox homeostasis and enhance the function of ATP generation. The molecular mechanism of regulation is still unclear. This study will explore the regulatory mechanism of rice seed conservation longevity from the perspective of mitochondrial activity regulation using wild-type and OsAOX1a-RNAi rice seeds during ageing.
2. Results
2.1. Pattern of survival curves between WT and OsAOX1a-RNAi seeds after AA treatment
Comparing the OsAOX1a-RNAi and wild-type rice seeds harvested in the same, while OsAOX1a low expression seeds have longer grain length smaller (Figure 1a) Compared seed width, length, and 100-grain weight with WT, the OsAOX1a-RNAi seeds decreased by 1.9%, 2.2% and 8.2%, respectively (Figure 1b–d). These results showed that the accumulation of dry matter in OsAOX1a-RNAi seeds decreased compared with WT seed.
The initial germination percentage of the WT and OsAOX1a-RNAi seeds were 98.5% and 96.5%, respectively. Under artificially aging treatment with 40 °C and 75% relative humidity for 30 d, the survival curves showed an inverse S-shaped curve with plateau, CN and rapid decline, were significantly different (Figure 2). Compared to WT seeds, OsAOX1a-RNAi seeds showed a similar plateau that the GP reached 90.0%, 80.0% and 70.0% after 6 d, 9 d and 12 d of AA treatment (Figure 2a). However, the vigor index of OsAOX1a-RNAi seeds during AA treatment were always significantly lower than that of WT (Figure 2b). Furthermore, after CN (12 days of aging), the germination rate of OsAOX1a-RNAi seeds began to decline rapidly comparing to WT seeds. A half-viability period (P50) was estimated to be 13.6 d and of 14.8 d for OsAOX1a-RNAi and WT seeds, respectively. These results indicated that the storability of OsAOX1a-RNAi seeds were significant decrease compared to WT seeds.
2.2. Assessment of mitochondria status during AA treatments
To gain deeper insight into the mitochondria status between OsAOX1a-RNAi and WT seeds during AA treatments, crude mitochondria were extracted from seeds imbibed for 48 hours at 100%, 90%, 80%, and 70% germination, respectively. The mitochondrial electron transport capacity was evaluated by monitoring NADH and succinate-dependent O2 consumption (Table 1). The respiratory rate of mitochondrial oxygen consumption decreased significantly with the decrease of seed viability in both materials. Compared to the WT seeds in the 100%, 90%, 80%, and 70% germination, the NADH-dependent O2 consumption of the OsAOX1a-RNAi seeds were decrease by 62%, 61%, 70% and 53%, respectively; the succinate-dependent O2 consumption were decrease by 21%, 28%, 54% and 19%, respectively. In addition to, the abundances of AOX1 and cyt c were further studied which played extremely important roles in mitochondrial electron transport (Figure 3). Compared to the WT seeds, the abundances of AOX1 and cyt c in the OsAOX1a-RNAi seeds were always lower than that in the WT during AA treatment. These results suggested that the capacity for electron transport were inhibited in the mitochondria of the OsAOX1a-RNAi seeds after imbibition, indicating decrease in the ATP supply.
The mitochondrial malate dehydrogenase (MDH) activity and ATP content was further measured between OsAOX1a-RNAi and WT seeds in the 100%, 90%, 80% and 70% germination, respectively. The MDH activities and ATP contents decreased significantly with the decrease of seed viability in both materials (Figure 4). Compared to the WT seeds in the 100%, 90%, 80%, and 70% germination, the MDH activities of the OsAOX1a-RNAi seeds were decrease by 14.4%, 14.3%, 13.18% and 24.8% respectively, while ATP contents were decrease by 47.2%, 60.5%, 53.8% and 51.8%, respectively. Therefore, above results indicated that mitochondrial status in the OsAOX1a-RNAi seeds after imbibition was weaken than the WT.
2.3. The abundance of complex I subunits during AA treatments
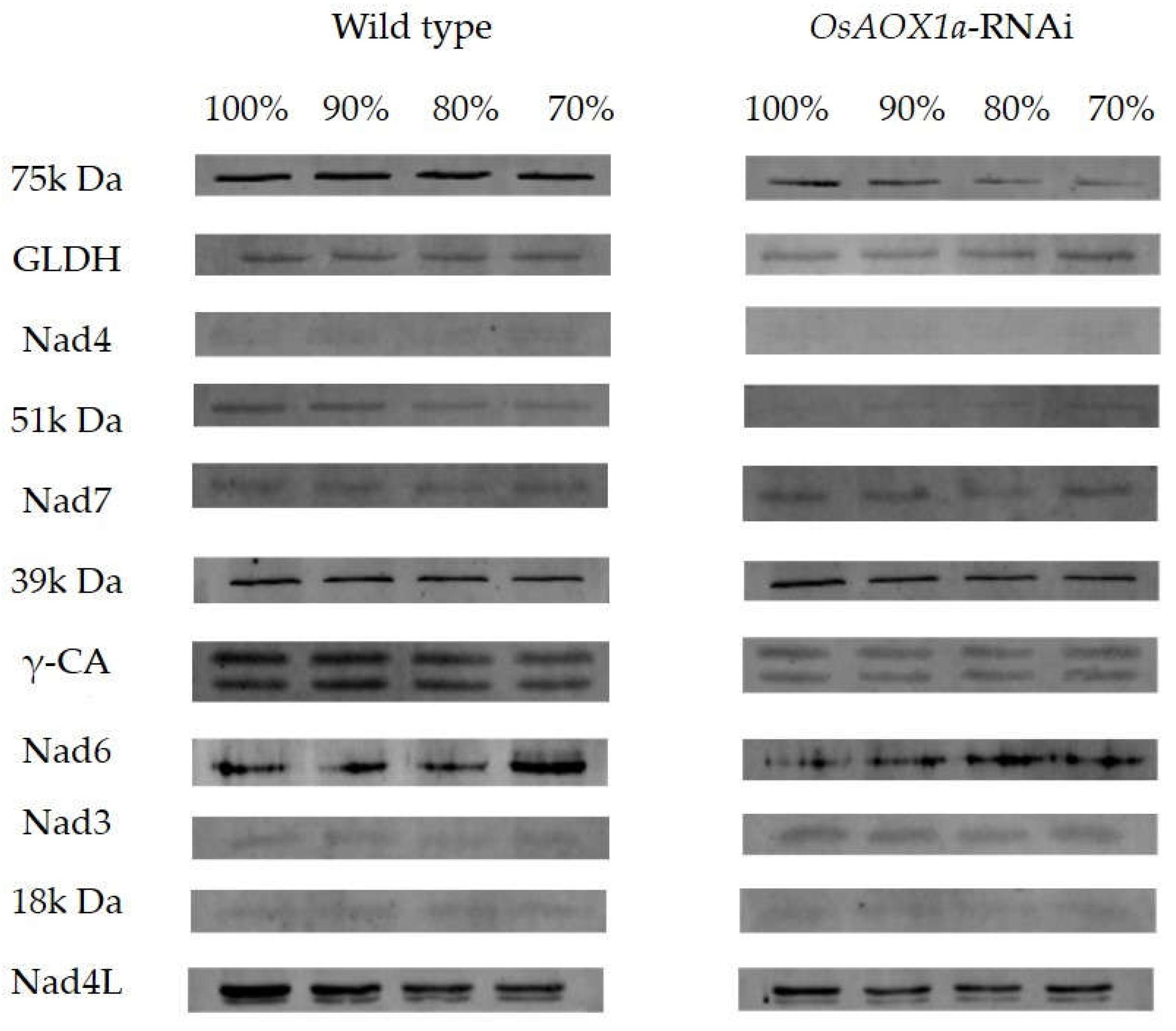
Mitochondrial electron transfer chain Complex I is upstream of the alternative oxidase pathway which were classified into five function sections: N module, Q module, P module, CA domain, GLDH domain. The abundances of Complex I subunits were investigated between OsAOX1a-RNAi and WT seeds in the 100%, 90%, 80% and 70% germination, respectively (Figure 5).
The abundance of 75 kDa and 51 kDa subunit, the core subunits in the N module, in the OsAOX1a-RNAi seeds were always lower than that in the WT during AA treatment. In addition to, the abundance of 75 kDa subunit in the WT seeds has no significantly changed during AA treatment, however, that in the OsAOX1a-RNAi seeds was gradually down-regulated. The abundance of 51 kDa subunit was down-regulated during AA treatment, however, that in the OsAOX1a-RNAi seeds was too low to be resolved, these results indicating that the capacity of electron transfer in OsAOX1a-RNAi was inhibited, especially in the CN of seed viability.
The abundance of Nad3, Nad4, Nad4L and Nad6 subunit, the core subunits in the P module, showed different patterns between OsAOX1a-RNAi and WT seeds during AA treatments. The abundance of Nad4 subunit were too low to be resolved in both materials. The abundance of Nad3 and Nad6 subunit in the OsAOX1a-RNAi seeds were always higher than that in the WT during AA treatment, respectively. In addition to, the abundance of Nad3 has no significantly changed, while the abundance of Nad6 was gradually down-regulated in both materials during AA treatment. The abundance of Nad4L subunit in the OsAOX1a-RNAi seeds were always lower than that in the WT during AA treatment. In addition to, the abundance of Nad4L subunit in the WT seeds was gradually down-regulated during AA treatment, however, that in the OsAOX1a-RNAi seeds has no significantly changed.
The abundance of 39 kDa and Nad7 subunit, the core subunits in the Q module, in the OsAOX1a-RNAi seeds were always higher than that in the WT during AA treatment. The subunits of GLDH and γ-carbonic anhydrase (γ-CA) are non-conserved subunit of complex I. The abundances of GLDH subunit were gradually up-regulated in both materials during AA treatment. The abundance of γ-CA subunit in the OsAOX1a-RNAi seeds were always lower than that in the WT during AA treatment.
Figure 5.
Verification abundance of mitochondrial complex I subunits in crude mitochondria from the wild type and OsAOX1a-RNAi seeds with germination percentage of ~90, ~80, ~70% and the control group after 48 h imbibition by westernblotting.
Figure 5.
Verification abundance of mitochondrial complex I subunits in crude mitochondria from the wild type and OsAOX1a-RNAi seeds with germination percentage of ~90, ~80, ~70% and the control group after 48 h imbibition by westernblotting.
3. Discussion
3.1. Low AOX1a impaired seed development and storability
AOX1a plays an extremely important role in maintaining growth and promoting plant survival under adverse conditions. Many studies have shown that the expression of AOX1a is up-regulated during seed germination, stress response, flowering and pollination, which plays a key role in plant growth and development [16,20,21]. The expression level of AOX1a under suitable environment had no significant effect on plant vegetative growth, but under low temperature condition, the expression level of AOX1a had no significant effect on plant vegetative growth. The growth of OsAOX1a-RNAi Arabidopsis thaliana leaves was slower and weaker under the same treatment [22]. The ripening, respiration, and ethylene production were severely impaired in OsAOX1a-RNAi tomato [23]. In this study, the OsAOX1a-RNAi seeds showed decrease in seed width, length, and 100-grain weight compared with the wild type seeds (Figure 1). The dry matter accumulation of seeds was positively correlated with the expression level of AOX1a. The down-regulation of AOX1a impaired the regulation of cell life activities at the seed development. Although germination percentage aged at 3 d, 6 d and 9 d of OsAOX1a-RNAi rice seeds is similar with WT, the vigor index of OsAOX1a-RNAi seeds during AA treatment were always significantly lower than that of WT (Figure 2). In addition to, the P50 of OsAOX1a-RNAi rice seeds is significantly longer than that of WT. These results indicated the down-regulation of AOX1a is reduced the seed storability. It can be concluded that AOX1a is involved in the regulation of seed development and storability.
3.2. Low AOX1a impaired mitochondrial activity during seed imbibition
AOX1a is key component in the mitochondrial alternative chain, which was inactivated that could lead to a different response [24]. In this study, compared to WT, the NADH and succinate-dependent O2 consumption were reduced in OsAOX1a-RNAi seeds during AA treatment, whereas the abundances of AOX1 and cyt c were inhibited expression (Table 1 and Figure 3). These results consistent with that the respiratory capacity of aox1a mitochondria was significantly restricted by impairing in the alternative-terminal oxidase, the cytochrome oxidase, or both [17,25]. Compared to our previous study, the capacity of O2 consumption of the pure mitochondria were higher than the crude mitochondria in present study, which were isolated from the imbibed rice seed after AA treatment. In addition to, the MDH activity and ATP content were reduced in OsAOX1a-RNAi seeds during AA treatment compared to the WT (Figure 4). These results consistent with that the activity of the malate-oxaloacetate shuttle in the aox1a mutant was lower than that of WT [26]. MDH catalyzes the interconversion of malate and oxaloacetate in mitochondria [27,28]. MDH activity were reduction in rice seed during AA treatment [5]. These results indicated that ATP production were severely inhibited in OsAOX1a-RNAi seeds at critical node of viability. ATP supply was core factor for seed germination. The insufficient of ATP in OsAOX1a-RNAi seeds at critical node of viability, might lead to rapid loss of seed viability. Above results indicated that mitochondrial metabolism might be inhibited in the OsAOX1a-RNAi seeds after imbibition.
3.3. Low AOX1a altered mitochondrial electron transfer chain complex I during seed imbibition
In our previous study, the alternative pathway of ETC was significantly induced, whereas the COX pathway was inhibited at critical node of seed viability in rice [5]. Complex I is the entry site for electrons into the ETC by oxidizing NADH to reduce uniquinone with translocation of H+ across the inner membrane, which contained three core modules: N (NADH oxidation site) and Q (ubiquinone reduction site) modules, and P (proton transfer site) module [29,30].
When Complex I activity is inhibited, the subunits of NADH dehydrogenase II, belonged P module, and AOX were upregulated under stress conditions, which formed an alternative pathway for maintaining cellular respiration and energy supply [31,32]. Our previous study has shown that the P module was significantly induced, whereas the N module was inhibited at critical node of seed viability in rice [19]. In this study, we also found that the core subunits of N module were down-regulated, whereas the subunits of P module were up-regulated in OsAOX1a-RNAi and WT at critical node of seed viability (Figure 5). However, the abundance of complex I subunits of the WT was higher than that of the OsAOX1a-RNAi, which showed that the capacity of ETC of the WT was significantly higher than that of OsAOX1a-RNAi at critical node of seed viability, indicating that the capacity of ATP production in the WT was higher than that in the OsAOX1a-RNAi seed. This might be the reason that the loss of viability of the OsAOX1a-RNAi seeds was faster than that of the WT after critical node of viability.
4. Materials and Methods
4.1. Materials and treatments
Rice seed (Oryza sativa L. japonica nipponbare) was used in all experiments. The homologous sequence of the OsAOX1a cDNA fragment was used for a double-stranded RNA interference (RNAi) trial. The construct AOX1a-RNAi was generated by the fragment and its inverted repeat fragment inserted downstream of the CaMV 35S promoter at the BamHI, SacI, Kpn I, and Spe I restriction sites of the modified pTCK303 vector. The primers were shown in Table 2. Transgenic plants were generated by Agrobacterium tumefaciens (strain EHA105)-mediated transformation, which showed positive in real-time PCR testing by HygR gene.
The seeds were sealed in aluminum foil bag at 40 °C for 7 days to break the dormancy. The seed germination percentage and vigor index were carried out at 28 °C for 7 d in the dark (ISTA, 1995). AA treatment was performed at 40 °C and 75% relative humidity for 21 days [5]. The aging curve were characterized by seeds withdrawn from the treatment every 3 days for each genotype. And the time for seed germination to decrease to 50% (P50) was calculated using Avrami equation with OriginPro software [33].
4.2. Crude mitochondria purification
For mitochondrial purification, rice seeds were imbibition at 28 °C for 48 h in the dark. 1200 embryos were grinded in buffer, centrifuge at 2000× g for 5 min, take the supernatant, grind The buffer was resuspended and the pellet was centrifuged at 2000× g for 5 min. The supernatant was collected and centrifuged at 12000× g for 15 min. Discard the supernatant, wash with elution buffer to collect all the precipitates, centrifuge again at 2000× g for 5 min, take the supernatant, and centrifuge at 12000× g for 15 min to obtain crudely extracted mitochondria. Each step of the experiment was performed at 4 °C, and the purification mitochondria were stored at −80 °C.
4.3. Mitochondrial respiration rate assay
Mitochondrial respiration rate was measured using an oxygen electrode (Hansatech, UK). Crude mitochondria (1 mg mitochondrial protein) was added to 1 mL of reaction solution containing 0.3 M sucrose, 10 mM TES-KOH (pH 7.5), 5 mM KH2PO4, 10 mM NaCl, 2 mM MgSO4 and 0.1% (w/v) BSA, and was always kept at 20 °C. NADH (10 mM), succinic acid (10 mM) and ADP (0.8 mM) were added according to Logan et al. [34].
4.4. ATP content determination
ATP content was determined using ATP Content Assay Kit (Solarbio, Beijing, China) according to the instructions of the manufacturer. Mitochondria pellets were resuspended in 1 mL extraction buffer and centrifuged at 10,000× g for 10 min. 500 μL chloroform were added in supernatant, and then centrifuged at 10,000× g for 3 min. The supernatant was collected for detection ATP.
4.5. Mitochondrial malate dehydrogenase activity
Mitochondrial malate dehydrogenase activity was based on the reversible reaction of NAD+ oxidizing L-malic acid to oxaloacetate, which was determined by monitoring the increase in absorbance at 340 nm and 25 °C. and 0.5 mg of mitochondrial protein was added to the reaction solution (0.1 M potassium phosphate, pH 7.5, 0.2 mM NADH, 5 mM oxaloacetic acid).
4.6. Western blot t analysis
Equal amounts of mitochondrial protein (10 μg per lane) were loaded onto 12% SDS-PAGE gels, transferred to PVDF, using the Mini Trans-Blot cell electrophoresis apparatus (Bio-Rad, USA). Blots were incubated with the following antibodies: Nad4, Nad4L, Nad6, Nad7 (Beijing Protein Innovation, China); 51 kDa, 75 kDa, 18 kDa, 39 kDa (Phytoab, USA); cyt c, AOX1, Nad3, GLDH, γ-CA (Agrisera, Sweden). A secondary antibody was anti-rabbit IgG (Agrisera, Sweden). LI-COR Odyssey dual-color infrared fluorescence imaging system (LI-COR Biosciences, USA) was used for imaging observation.
4.7. Statistical Analyses
High-confidence results were obtained by using moderated t-test (Limma) with rank products based on well-defined null hypotheses. Differences at the level of p < 0.05, 0.01, and 0.001 were considered as significant, which were respectively labeled as ∗, ∗∗, and ∗∗∗, respectively.
5. Conclusions
In conclusion, seed development and storability was impaired in OsAOX1a-RNAi seeds. In addition to, the activities of mitochondrial metabolism and alternative pathways were severely inhibited in OsAOX1a-RNAi seeds at critical node of viability, which could not supply effectively ATP after imbibition, which was leading to accelerate the collapse of seed viability. This study further explored the biological mechanism of the critical nodes of seed viability, and provided a scientific basis for constructing the technology of seed safety conservation.
Author Contributions
G.Y. and X.L. designed the research. J.J., S.L., J.H., X.X. and Y.Z. performed experiments, G.Y., J.J. and G.S. analysed the results. G.Y. and X.L. wrote the paper, X.X., Y.L. helped with editing the paper before submission. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Key Research and Development Program of China (2021YFD1200501)
Data Availability Statement
The data presented in this study are available on reasonable request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carrie, C.; Murcha, M.W.; Giraud, E.; Ng, S.; Zhang, M.F.; Narsai, R.; Whelan, J. How do plants make mitochondria? Planta 2013, 237, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Law, S.R.; Narsai, R.; Taylor, N.L.; Delannoy, E.; Carrie, C.; Giraud, E.; Millar, A.H.; Small, I.; Whelan, J. Nucleotide and RNA metabolism prime translational initiation in the earliest events of mitochondrial biogenesis during Arabidopsis germination. Plant Physiol. 2012, 158, 1610–1627. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Tian, Q.; Yin, G.; Chen, X.; Zhang, J.; Ng, S.; Lu, X. Reduced mitochondrial and ascorbate-glutathione activity after artificial ageing in soybean seed. J Plant Physiol. 2014, 171, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Xue, H.; Pritchard, H.W.; Wang, X. Reactive oxygen species-provoked mitochondria-dependent cell death during ageing of elm (Ulmus pumila L.) seeds. Plant J. 2015, 81, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Whelan, J.; Wu, S.; Zhou, J.; Chen, B.; Chen, X.; Zhang, J.; He, J.; Xin, X.; Lu, X. Comprehensive mitochondrial metabolic shift during the critical node of Seed Ageing in Rice. PLoS One 2016, 11, e0148013. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Xue, H.; Pritchard, H.W.; Wang, X. Changes in the mitochondrial protein profile due to ROS eruption during ageing of elm (Ulmus pumila L.) seeds. Plant Physiol Biochem. 2017, 114, 72–87. [Google Scholar] [CrossRef]
- Sun, H.; Li, L.; Wang, X.; Wu, S.; Wang, X. Ascorbate-glutathione cycle of mitochondria in osmoprimed soybean cotyledons in response to imbibitional chilling injury. J Plant Physiol. 2011, 168, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R.; Levine, R.L. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino Acids 2003, 25, 207–218. [Google Scholar] [CrossRef]
- Ramallo Guevara, C.; Philipp, O.; Hamann, A.; Werner, A.; Osiewacz, H.D.; Rexroth, S.; Rögner, M.; Poetsch, A. Global protein oxidation profiling suggests efficient mitochondrial proteome homeostasis during aging. Mol Cell Proteomics. 2016, 15, 1692–1709. [Google Scholar] [CrossRef]
- Yin, G.; Xin, X.; Fu, S.; An, M.; Wu, S.; Chen, X.; Zhang, J.; He, J.; Whelan, J.; Lu, X. Proteomic and carbonylation profile analysis at the critical node of Seed Ageing in Oryza sativa. Sci Rep. 2017, 7, 40611. [Google Scholar] [CrossRef]
- Fu, S.; Yin, G.; Xin, X.; Wu, S.; Wei, X.; Lu, X. Levels of crotonaldehyde and 4-hydroxy-(E)-2-nonenal and expression of genes encoding carbonyl-Scavenging enzyme at critical node during rice seed aging. Rice Sci. 2018, 25, 152–160. [Google Scholar]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu Rev Plant Biol. 2011, 62, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Moller, I.M. Plant mitochondria and oxidative stress: electron transport, NADPH Turnover, and metabolism of reactive oxygen species. Annu Rev Plant Physiol Plant Mol Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [PubMed]
- Fromm, S.; Senkler, J.; Zabaleta, E.; Peterhänsel, C.; Braun, H.P. The carbonic anhydrase domain of plant mitochondrial complex I. Physiol Plant. 2016, 157, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Senkler, J.; Senkler, M.; Braun, H.P. Structure and function of complex I in animals and plants—A comparative view. Physiol Plant 2017, 161, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Selinski, J.; Scheibe, R.; Day, D.A.; Whelan, J. Alternative oxidase is positive for plant performance. Trends Plant Sci. 2018, 23, 588–597. [Google Scholar] [CrossRef]
- Kühn, K.; Yin, G.; Duncan, O.; Law, S.R.; Kubiszewski-Jakubiak, S.; Kaur, P.; Meyer, E.; Wang, Y.; Small, C.C.; Giraud, E.; Narsai, R.; Whelan, J. Decreasing electron flux through the cytochrome and/or alternative respiratory pathways triggers common and distinct cellular responses dependent on growth conditions. Plant Physiol. 2015, 167, 228–250. [Google Scholar] [CrossRef]
- Costa, J.H.; McDonald, A.E.; Arnholdt-Schmitt, B.; Fernandes de Melo, D. A classification scheme for alternative oxidases reveals the taxonomic distribution and evolutionary history of the enzyme in angiosperms. Mitochondrion 2014, 19 Pt B, 172–183. [Google Scholar] [CrossRef]
- Chen, B.; Yin, G.; Whelan, J.; Zhang, Z.; Xin, X.; He, J.; Chen, X.; Zhang, J.; Zhou, Y.; Lu, X. Composition of mitochondrial Complex I during the critical node of seed aging in Oryza sativa. J Plant Physiol. 2019, 236, 7–14. [Google Scholar] [CrossRef]
- Garmash, E.V.; Velegzhaninov, I.O.; Ermolina, K.V.; Rybak, A.V.; Malyshev, R.V. Altered levels of AOX1a expression result in changes in metabolic pathways in Arabidopsis thaliana plants acclimated to low dose rates of ultraviolet B radiation. Plant Sci. 2020, 291, 110332. [Google Scholar] [CrossRef]
- Clifton, R.; Millar, A.H.; Whelan, J. Alternative oxidases in Arabidopsis: a comparative analysis of differential expression in the gene family provides new insights into function of non-phosphorylating bypasses. Biochimica et biophysica Acta 2006, 1757, 730–741. [Google Scholar] [CrossRef]
- Fiorani, F.; Umbach, A.L.; Siedow, J.N. The alternative oxidase of plant mitochondria is involved in the acclimation of shoot growth at low temperature. A study of Arabidopsis AOX1a transgenic plants. Plant Physiol. 2005, 139, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Yuan, S.; Zhang, D.W.; Lv, X.; Lin, H.H. The role of alternative oxidase in tomato fruit ripening and its regulatory interaction with ethylene. J Exp Bot. 2012, 63, 5705–5716. [Google Scholar] [CrossRef]
- Selinski, J.; Scheibe, R.; Day, D.A.; Whelan, J. Alternative oxidase is ppositive for plant performance. Trends in Plant Sci. 2018, 23, 588–597. [Google Scholar] [CrossRef]
- Giraud, E.; Ho, L.H.; Clifton, R.; Carroll, A.; Estavillo, G.; Tan, Y.F.; Howell, K.A.; Ivanova, A.; Pogson, B.J.; Millar, A.H.; Whelan, J. The absence of alternative oxidase1a in Arabidopsis results in acute sensitivity to combined light and drought stress. Plant Physiol. 2008, 147, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Gandin, A.; Duffes, C.; Day, D.A.; Cousins, A.B. The absence of alternative oxidase AOX1A results in altered response of photosynthetic carbon assimilation to increasing CO2 in Arabidopsis thaliana. Plant Cell Physiol. 2012, 53, 1627–1637. [Google Scholar] [CrossRef] [PubMed]
- Imsande, J.; Pittig, J.; Palmer, R.G.; Wimmer, C.; Gietl, C. Independent spontaneous mitochondrial malate dehydrogenase null mutants in soybean are the result of deletions. The Journal of heredity 2001, 92, 333–338. [Google Scholar] [CrossRef]
- Day, D.A.; Millar, A.H.; Whelan, J. Plant mitochondria: from genome to function. In Advances in Photosynthesis and Respiration; Govindjee Kluwer, Ed.; Academic Press: Dordrecht, 2004; pp. 2–7. [Google Scholar]
- Hunte, C.; Zickermann, V.; Brandt, U. Functional modules and structural basis of conformational coupling in mitochondrial complex I. Science 2010, 329, 448–451. [Google Scholar] [CrossRef]
- Braun, H.P.; Binder, S.; Brennicke, A.; Eubel, H.; Fernie, A.R.; Finkemeier, I.; Klodmann, J.; König, A.C.; Kühn, K.; Meyer, E.; Obata, T.; Schwarzländer, M.; Takenaka, M.; Zehrmann, A. The life of plant mitochondrial complex I. Mitochondrion 2014, 19 Pt B, 295–313. [Google Scholar] [CrossRef]
- Sweetman, C.; Waterman, C.D.; Rainbird, B.M.; Smith, P.M.C.; Jenkins, C.D.; Day, D.A.; Soole, K.L. AtNDB2 is the main external NADH dehydrogenase in mitochondria and is important for tolerance to environmental stress. Plant Physiol. 2019, 181, 774–788. [Google Scholar] [CrossRef]
- Sweetman, C.; Miller, T.K.; Booth, N.J.; Shavrukov, Y.; Jenkins, C.L.D.; Soole, K.L.; Day, D.A. Identification of alternative mitochondrial electron transport pathway components in chickpea indicates a differential response to salinity stress between cultivars. Int J Mol Sci. 2020, 21, 3844. [Google Scholar] [CrossRef] [PubMed]
- Walters, C.; Wheeler, L.; Grotenhuis, J. Longevity of seeds stored in a genebank: Species characteristics. Seed Sci Res. 2005, 15, 1–20. [Google Scholar] [CrossRef]
- Logan, D.C.; Millar, A.H.; Sweetlove, L.J.; Hill, S.A.; Leaver, C.J. Mitochondrial biogenesis during germination in maize embryos. Plant physiol. 2001, 125, 662–672. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparison of seed width and length (a) between WT and OsAOX1a-RNAi rice, with with statistics for width (b), length (c) and 100-grain weight (d). Data represent the mean ± standard deviation of 3 independent experiments. * represent that the significant change was at the level of p < 0.05.
Figure 1.
Comparison of seed width and length (a) between WT and OsAOX1a-RNAi rice, with with statistics for width (b), length (c) and 100-grain weight (d). Data represent the mean ± standard deviation of 3 independent experiments. * represent that the significant change was at the level of p < 0.05.
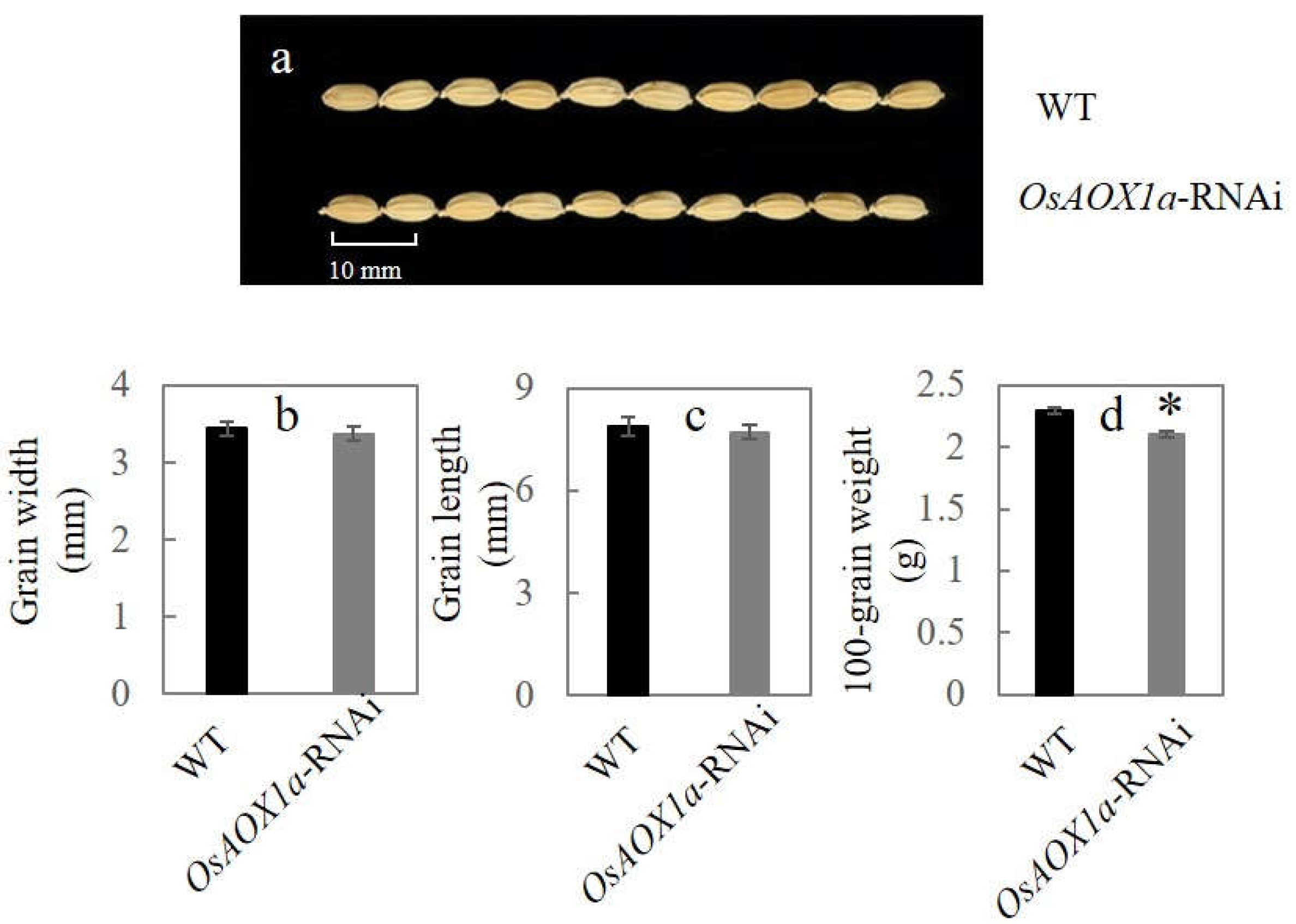
Figure 2.
Seed viability curves and P50 (a) and the vigor index (b) of the wild type and OsAOX1a-RNAi rice following artificial aging treatment. Data represent the mean ± standard deviation of 3 independent experiments. *, **, *** represent that the significant change was at the level of p < 0.05, 0.01, 0.001, respectively.
Figure 2.
Seed viability curves and P50 (a) and the vigor index (b) of the wild type and OsAOX1a-RNAi rice following artificial aging treatment. Data represent the mean ± standard deviation of 3 independent experiments. *, **, *** represent that the significant change was at the level of p < 0.05, 0.01, 0.001, respectively.
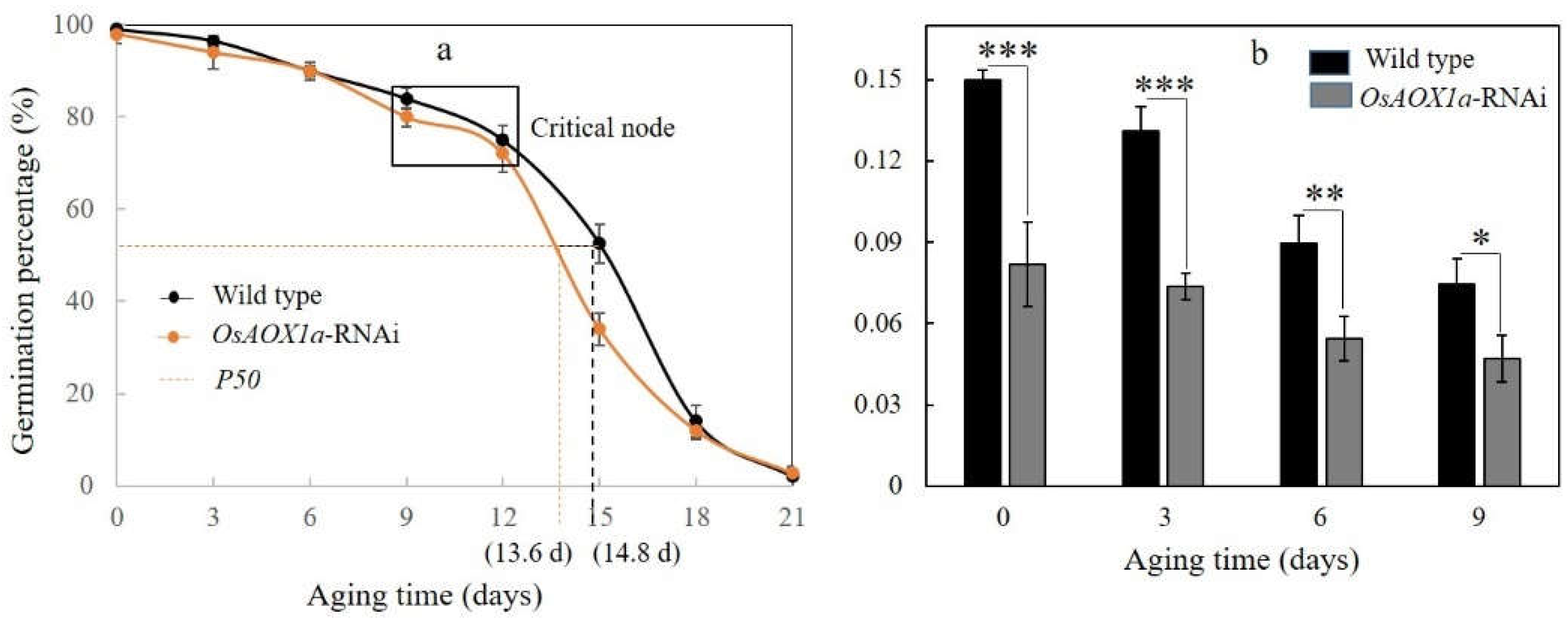
Figure 3.
Verification abundance of AOX1 and cyt c in crude mitochondria from the wild type and OsAOX1a-RNAi seeds with germination percentage of ~90, ~80, ~70% and the control group after 48 h imbibition by westernblotting.
Figure 3.
Verification abundance of AOX1 and cyt c in crude mitochondria from the wild type and OsAOX1a-RNAi seeds with germination percentage of ~90, ~80, ~70% and the control group after 48 h imbibition by westernblotting.

Figure 4.
Malate dehydrogenase (MDH, a) activity and ATP content (b) in crude mitochondria from the wild type and OsAOX1a-RNAi seeds with germination percentage of ~90, ~80, ~70% and the control group after 48 h imbibition. Data represent the mean ± standard deviation of 3 independent experiments. *, **, *** represent that the significant change was at the level of p < 0.05, 0.01, 0.001, respectively.
Figure 4.
Malate dehydrogenase (MDH, a) activity and ATP content (b) in crude mitochondria from the wild type and OsAOX1a-RNAi seeds with germination percentage of ~90, ~80, ~70% and the control group after 48 h imbibition. Data represent the mean ± standard deviation of 3 independent experiments. *, **, *** represent that the significant change was at the level of p < 0.05, 0.01, 0.001, respectively.
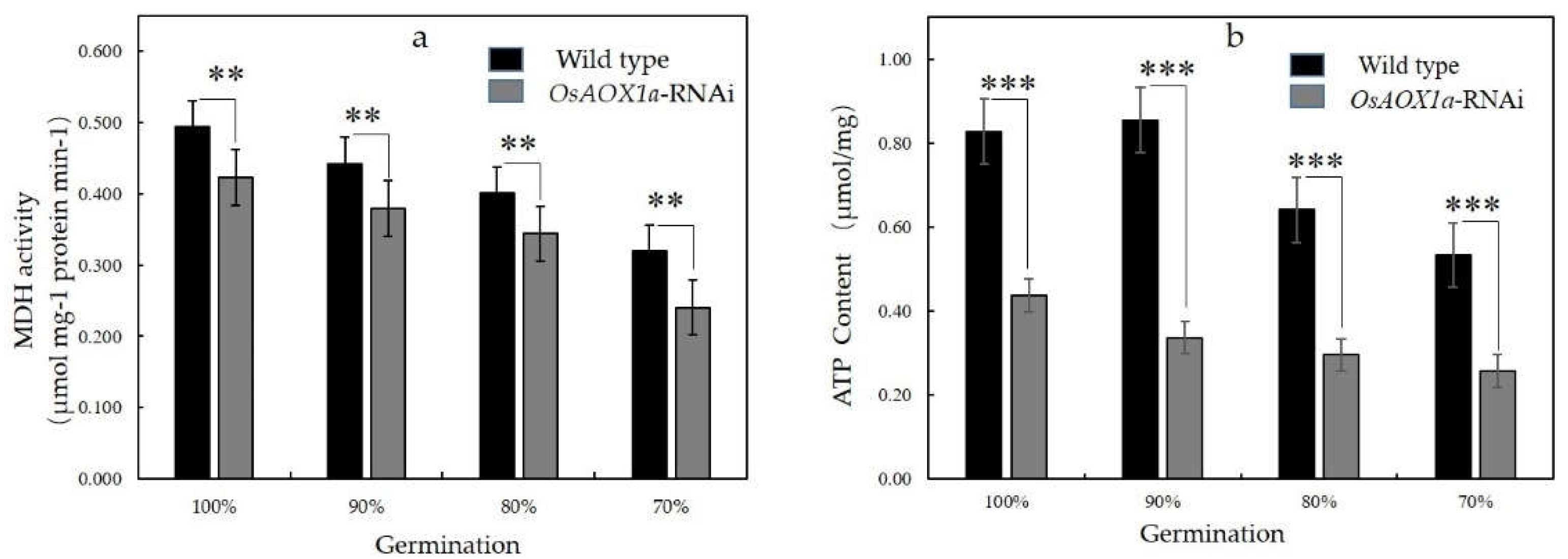

Table 1.
Rate of oxygen uptake (nmolO2·min−1mg−1 protein) by mitochondria from the wild type and OsAOX1a-RNAi seeds with germination percentage of ~90, ~80, ~70% and the control group after 48 h imbibition. O2 consumption rates are presented as means ± SD (n = 3).
Table 1.
Rate of oxygen uptake (nmolO2·min−1mg−1 protein) by mitochondria from the wild type and OsAOX1a-RNAi seeds with germination percentage of ~90, ~80, ~70% and the control group after 48 h imbibition. O2 consumption rates are presented as means ± SD (n = 3).
| Materials | Substrate | O2 consumption rate (nmolO2·min−1mg−1 protein) | |||
|---|---|---|---|---|---|
| 100% | 90% | 80% | 70% | ||
| Wild type | NADH | 44.4 ± 3.2 | 38.1 ± 2.2 | 18.9 ± 0.6 | 11.4 ± 2.6 |
| NADH + ADP | 97.8 ± 3.2 | 65.5 ± 2.9 | 43.3 ± 3.0 | 22.7 ± 2.6 | |
| Succinate | 24.5 ± 2.0 | 18.8 ± 0.8 | 11.5 ± 1.0 | 7.9 ± 0.7 | |
| Succinate + ADP | 40.5 ± 1.9 | 25.8 ± 1.8 | 14.2 ± 1.7 | 9.8 ± 2.6 | |
| OsAOX1a-RNAi | NADH | 17.3 ± 1.9 | 11.8 ± 1.2 | 8.3 ± 0.7 | 6.6 ± 0.4 |
| NADH + ADP | 36.7 ± 0.9 | 25.2 ± 1.1 | 12.9 ± 0.7 | 10.5 ± 1.0 | |
| Succinate | 7.9 ± 0.2 | 6.0 ± 0.3 | 4.5 ± 0.5 | 4.1 ± 0.2 | |
| Succinate + ADP | 32.0 ± 1.2 | 18.5 ± 0.7 | 8.8 ± 0.6 | 7.9 ± 0.5 | |
Table 2.
Primers used in real-time PCR.
| name | Primer (5'→3') | restriction site |
|---|---|---|
| OsAOX1a-F | CGGGTACCACTAGTGTGCGGCGTACGAAAAACAG | Kpn I + Spe I |
| OsAOX1a-R | GGGGATCCGAGCTCTGCCGAGGATTTGCATCACT | BamH I + Sac I |
| HygF | CTATTTCTTTGCCCTCGGAC | |
| HygR | AAGCCTGAACTCACCGCGAC | |
| OsUBQ5-F | ACCACTTCGACCGCCACTACT | |
| OsUBQ5-R | ACGCCTAAGCCTGCTGGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



