
Submitted:
14 April 2023
Posted:
17 April 2023
You are already at the latest version
Abstract
Drought stress inducing pollen sterility can reduce crop yield worldwide. The regulatory crosstalk associated with the effects of drought on pollen formation at the cellular level have not been explored in detail so far.
In this study, we performed morphological and cytoembryological analysis of anther perturbations and examined pollen development in two spring barley genotypes that differ in earliness and drought tolerance. The Syrian breeding line CamB (drought-tolerant) and the European cultivar Lubuski (drought-sensitive) were used as experimental materials to analyze the drought-induced changes in yield performance, chlorophyll fluorescence kinetics, the pollen grain micromorphology and ultrastructure during critical stages of plant development. In addition, fluctuations in HvGAMYB expression were studied, as this transcription factor is closely associated with the anthers development.
In the experiments, the studied plants were affected by drought, as was confirmed by the analyses of yield performance and chlorophyll fluorescence kinetics. However, contrary to our expectations, the pollen development of plants grown under specific conditions was not severely affected. The results also suggest that growth modification, as well as the perturbation in light distribution, can affect the HvGAMYB expression. This study showed that the duration of the vegetation period can influence plant drought responses and, as a consequence, the processes associated with pollen development as every growth modification changes the dynamics of drought effects as well as the duration of plant exposition to drought.
Keywords:
Drought
; Gibberellins
; HvGAMYB
; Light Deprivation
; Pollen Development
1. Introduction
Pollen development is an important process in the life cycle of plants, since only normally formed pollen can allow proper fertilization and the formation of full-fledged seeds. However, under unfavorable conditions, the anther development is disturbed, which leads to the formation of sterile pollen [1]. Elevated temperatures and frequent episodes of drought driven by climate change affect developmental and physiological processes in plants, and ultimately crop yield and quality [2]. Drought stress also inhibits the growth of plants by influencing their various biochemical functions such as photosynthesis, chlorophyll synthesis, nutrient metabolism, ion uptake and translocation, respiration, and carbohydrate metabolism [3,4]. Moreover, it affects pollen development by disturbing the internal homeostasis of plant cells, possibly by altering the intracellular levels of sugars, hormones, and reactive oxygen species [5,6]. However, the extent of damages, the recovery capacity, and the impact on the yield depend on the developmental stage at which the crop is exposed to this abiotic stress [7]. Although the impact of stress on plant yield and development has been extensively studied [8,9], the interactions of stress responses with plant phenology have often been neglected.
Pollen development within anthers has been well-studied [10,11]. Due to its role in anther and pollen development, the growth regulator gibberellic acid (GA) has been studied in several works. GA is vital to anther development as well as pollen viability [12], and its signaling predominantly occurs in tapetal cells [13]. Transcription factor families such as MYB, AP2/ERF, NAC, and MDAS regulate flower development. GAMYB encodes R2R3-MYB transcription factor, which belongs to the MYB transcription factor family playing an important role in plant growth and development, including defense, cell differentiation, secondary metabolism, and responses to biotic and abiotic stresses [14,15]. GAMYB appears to be the key transcription factor in the GA signaling pathway [16]. The crosstalk between gibberellin and photoperiod pathways as well as potential gene integrators has been studied in different plant species [17,18,19]. Although studies have analyzed different MYB genes due to their involvement in the regulation of abiotic stress response [20,21], the role of HvGAMYB in drought responses in the context of plant phenology remains elusive.
In order to survive in drought conditions, plants develop a few strategies to cope with limited water availability. These strategies include: (i) drought avoidance [22], (ii) drought tolerance [23], (iii) drought resistance [24], (iv) drought abandonment [25], and (v) drought-prone activation of biochemical/physiological traits [26]. In addition to these, (vi) drought escape (DE) is an important strategy adopted by plants to cope with impending unfavorable environmental conditions. However, classifying plants based on their drought response mechanism is not entirely accurate since the timing of the onset of the mechanism varies from one plant to another, such as prior to or after the occurrence of a drought event. This implies that plants use a combination of DE and other strategies to survive (and reproduce) under drought. Although DE is considered characteristic of “ephemeral native plants,” major cereals such as wheat and barley may exhibit a mechanism similar to DE, which is referred to as “earliness” or “early flowering” [27].
Both light and darkness act as environmental stimuli regulating plant growth and development from seedling emergence to senescence. Darkness induces the secretion of phytohormones such as GA and ethylene in germinating seeds [28]. Under darkness, GA accumulates and binds to its receptor gibberellin-insensitive dwarf 1 as well as its targets, DELLAs (GAI and RGA), which are the negative regulators of GA signal transduction [29]. Detailed studies on gene expression patterns have revealed various discrepancies between dark-induced and developmentally controlled processes [30,31,32].
As increasing evidence suggests that stress responses depend on the developmental stage of the plant [33], we decided to study specific developmental stages to understand the extent of the negative influence of stress on plant development. Because barley (Hordeum vulgare L.) is usually used as a model plant in studies on abiotic stress responses of plants [34] and precision developmental scales are currently available for barley [35], we designed the experiment with two spring barley genotypes that differ in phenology and resistance to abiotic stresses [36,37]. Experiments were designed from a multi-perspective approach to enrich the knowledge about better adaptations of plants to stressful growth conditions.
The aims of our study were to (i) investigate the effects of drought on spikes fertility, (ii) explore drought-induced alterations in pollen grain ultrastructure, and (iii) characterize the expression pattern of HvGAMYB under light deprivation and drought conditions in phenologically differentiated barley plants.
2. Material and Methods
2.1. Plant material and growth conditions
In this study, two barley genotypes were used –Lubuski is an old Polish cultivar derived from a Heines-Haisa/Skrzeszowicki hybrid and Cam/B1/CI08887//CI05761 (hereafter referred as CamB) is the Syrian breeding line supplied to Dr. A. Górny (IPG PAS) by Drs S. Grando and S. Ceccarelli from ICARDA in Aleppo. The two cultivars were selected as they show different phenology and are adapted to contrasting environments (Lubuski, late-heading, Poland-grown cultivar; CamB, early-heading, adapted to arid condition) [36,38].
2.2. Growth conditions, treatments and experimental designs
All experiments were conducted in growth chambers under fully controlled conditions (IPG PAS phytotrons). Five seeds from each accession were sown in plastic pots (40 cm×26 cm×26 cm) filled with field-soil and peat (3:1, w/w). Plants were cultivated in optimal conditions: 22 °C/18 °C day/night, air humidity of 50–60%, and a photoperiod of 16/8 h of light/dark (an exception: dark condition). All treatments were replicated three times. In the study the barley external development stages scale [35], in which the later stages from Zadoks scale [39] have been replaced by the last flag elongation (LFE) stages to provide a clear system for developmental staging associated with reproductive development. Samples were collected at LFE1 (flag leaf emerged completely and unrolling, ligule may be visible; last flag sheath extended 0.5–5 cm - development point 1) and LFE4 (spike has completed its upward movement and was entirely localized within the last flag sheath - development point 2) stages.
2.3. Drought treatment
Plants were irrigated until the onset of flag leaf emergence (Z37), then they were exposed to two watering treatments: (i) well-watering treatment (soil maintained at ~70% of field capacity (FC)); and (ii) severe drought stress (at 20% FC) following methodology implemented by Kuczyńska et al. [40]. To maintain the targeted control and drought conditions, soil moisture in each pot was controlled gravimetrically by weighing and, additionally, volumetrically (if necessary) using the FOM/mts device [36,41].
2.4. GA3 and TRinexapac application
10 mM GA3 (100 mg/l) solution was sprayed directly on the leaves at the tillering stage (Z21). Bioactive GA3 (Sigma-Aldrich) solution was prepared by dissolving powder in distilled water. For each plant, 1ml GA3 solution was applied. The GA3 treatment option was applied following the method descripted by Boden et al. [42]. Trinexapac-ethyl was used as a commercial product Moddus 250 EC (Syngenta, USA). The spraying was applied at the tillering stage (Z21). The treatment option was implemented following methods descripted by Grijalva-Contreras et al. [43].
2.5. Light/Dark Cycle Treatment
In this study four types of treatments were applied: C – control condition; D – drought condition; D+GA - drought condition combined with GA3 application; D+TR - drought condition combined with TRinexapac application, in two experiments - experiment 1 (hereafter referred as Exp 1), and experiment 2 (hereafter referred as Exp 2). In Exp 2, additional stress application – light deprivation was implemented. The dark conditions (started at the beginning of abiotic stress – Z37) were achieved by carefully covering the barley plants in a dark growth chamber. The dark treatment was carried out for 7 days and covered the beginning of the flowering period. After the dark treatment, the plants were exposed to the normal light regime.
2.6. Phenology and phenotypic evaluation
In this study, four developmental stages (tillering, flag leaf, flowering, heading) appearances were noticed when for at least 51% of all plants grown under given experimental variants the developmental phase was observed and presented as a number of days after sowing (DAS). 14 yield related traits were also investigated. Name of these traits with abbreviations and description of measurements methods is given in Table 1.
2.7. Anther morphology evaluation
Anther measurements were performed using stereomicroscope (Motic SMZ-161) following the protocol descripted by Browne et al. [44]. Anthers were photographed at a fixed magnification using a digital camera system (Moticam CMOS BTU8). The images were measured using Motic Advanced 3.2 software (China). Anther length (mm) was determined following removal of the anther from the primary floret of the largest spikelet. Anther width (mm) was the width of the anther at its broadest point. The probes were collected at LFE3, just before flowering (LFE3 - flag leaf opening and awns clearly visible; last flag sheath extended over 10 cm; rachis starts elongation, moving the spike upwards towards the last flag sheath).
2.8. Pollen viability and morphology evaluation
Pollen viability and fertility were evaluated using KI/I2 (method 1 – abbreviated as Pv1) and TTC (2,3,5-Triphenyl Tetrazolium Chloride) (method 2 – abbreviated as Pv2) staining methods as described by Wang et al. [45] and Ma et al. [46], respectively, with minor modifications. Only mature pollen was used for the evaluations (LFE4). Anthers from different plants were used for each replicate. Pollen was extracted according to the protocol of Impe et al. [47]. First, fresh pollen was collected from the studied plants on the same day at 09:00. Then, pollen from each line was divided into two samples; one sample was incubated in a 1.5-ml centrifuge tube containing 0.1% TTC at 37°C for 1 h, and the other was incubated in a 1.5-ml centrifuge tube containing 1% KI/I2 (potassium iodide/iodine) stain at room temperature for 5 min. The stained pollen was examined using a light microscope at a magnification of 400× (Motic BA410-E) and photographed using the Moticam digital camera. The images were analyzed using the Motic Advanced 3.2 software (Motic China Group Co., China). The pollen grains that had a round shape and were stained black with KI/I2 were classified as viable or alive, whereas those that were stained yellow or bright red were classified as sterile or dead. The pollen grains that stained red or pink with TTC were classified as viable (by the response to the presence of enzymatic activity), whereas those that appeared gray or colorless were considered sterile. In the viability assays, all the treatments (C, D, D+GA and D+TR) were conducted in triplicate in each of the three slides of all genotypes. At least 100–450 pollen were counted per single observation. Ca. 2000 pollen were counted for each genotype in this study. Pollen viability for each genotype was then expressed as the percentage of the total number of live pollen grains to the total number of grains observed per field.
The pollen grain area, perimeter, width (the widest point) and length were evaluated. The morphological evaluations were based on 100 randomly selected pollen grains observed in each genotype in each treatments.
2.9. Chlorophyll fluorescence measurements
Chlorophyll content and fluorescence were measured on both control and stressed plants. Data were collected at two development points— LFE1 and LFE3, always at the same time of the day (09:00). Measurements were taken using a FluorPen FP 100-MAX (Photon Systems Instruments, Drasov, Czech Republic). Fluorescence transients for chlorophyll-a were recorded from the center of the completely spent leaf (second from the top). The leaves were dark-adapted for 30 min, before starting the measurements using leaf clips provided by the manufacturer. Then the leaves were exposed to a pulse of saturating light at an intensity of 3,000 μmol m-2 s-1 and all studied parameters were measured. In each block, leaves from three plants of each line were measured. Nine replicates were performed for each cultivar and treatment (three leaves from three plants/treatment). The parameters analyzed in this study are presented in Table 2.
2.10. Relative Water Content
The RWC was calculated according to the method of Barrs and Weatherley [48] using the formula RWC (%) = (FW–DW)/ (TW–DW) × 100, where: FW is the fresh weight of the detached second leaf; TW is the turgid weight of the second leaf, which was incubated in distilled water for 24 h in darkness after detachment; and DW is the dry weight of the second leaf after it was dried in a dryer at 60°C for 48 h. Nine replicates (three replicates from three plants/treatment) were determined for each combination of genotype and treatment. The samples were collected at LFE4.
2.11. Microscopic observations
For cytological studies, anthers from spikelets of the middle zone of spike (three per each experimental treatment) were collected. Isolated anthers were fixed in mixture of equal volume of 2.0% glutaraldehyde and 2.0% paraformaldehyde in 0.05 M cacodylic buffer. The dates of sample collection were showed in Supplementary Table S1.
2.12. Light microscopy observation
The probes were collected at LFE4. For observations in light microscope (LM) commercially available acrylic resin Technovit 7100 (Heraeus Kulzer Gmbh, Wahrheim, Germany) was used. Fixed samples were dehydrated in ethanol series (from 5% to 100%) and embedded in Technovit resin. Sections of 4 µm thick were stained with 0.05 toluidine blue in 1% sodium borate. Micrographs were taken using a light microscope (Axioscope A1, Zeiss, Germany) with an attached AxioCam MRc5 digital camera (Carl Zeiss GmbH, Jena, Germany). On average, eight anthers from each experimental treatment were examined.
2.13. Electron microscopy observation
The probes were collected at LFE4. For observations in transmission electron microscope (TEM) the fixed anthers were rinsed three times in 0.05 M cacodylic buffer (pH 6.8), post-fixed for two hours in 1% osmium tetroxide in room temperature, and counter-stained with 2% aqueous uranyl acetate (pH 5.0). Samples were dehydrated in a graded acetone series (from 5% to 100%) and embedded in low-viscosity Spurr’s resin (Polysciences). The sections of 90 nm in thickness were counter-stained for twenty minutes in lead citrate and observed with JEOL JEM 1200 EXII (Jeol, Tokyo, Japan) transmission electron microscope at acceleration of 80 kV. At least five anthers from each experimental treatment were examined. Moreover, from the same samples, semithin sections were collected and observed microscopically (Axioscope A1, Zeiss, Germany) and documented with an attached AxioCam MRc5 digital camera (Carl Zeiss GmbH, Jena, Germany).
2.14. RNA Extraction, Reverse Transcription, and Real-Time PCR
Anthers were collected at two developmental stages (LFE1 and LFE4). Four biological replicates were collected for each stage, comprising approximately 100 anthers from four individual spikes. After dissection, the anthers were immediately frozen in liquid nitrogen and stored at −80°C until RNA extraction was performed. The RNA was extracted using the RNeasy Mini Kit (QIAGEN, Germany) according to the manufacturer’s protocol with on-column DNase treatment (QIAGEN, Germany). Additionally, all isolated RNA samples were treated with TURBO DNase (Thermo Fisher Scientific, Lithuania) according to the manufacturer’s instructions to exclude trace contamination of samples with genomic DNA. The purity of all RNA samples was assessed via OD260/280 and OD260/230 absorbance ratios, whereas their structural integrity was evaluated using denaturing agarose gel electrophoresis. All RNA samples were adjusted to the same concentration (100 ng/μl). The quantitative real-time PCR (RT-qPCR) analysis performed in this study met the MIQE criteria [49]. Single-stranded cDNA was synthesized from 1 μg of total RNA using the iTaq Universal SYBR Green One-Step Kit according to the manufacturer’s instructions. To analyze the specific expression of each reference/target gene, RT-qPCR was performed using the CFX Connect Real-time PCR Detection System (Bio-Rad). Each 10 μl mixture for PCR contained 1 μl of a diluted RNA and 5 µM of each primer. To confirm the specificity of amplification and the absence of primer dimers, each run was completed with melting curve analysis (melting curve 63°C to 95°C, increasing by 0.5°C for 0.05 s). Moreover, each pooled RT-qPCR product underwent sequencing process (AMU, Poznań, Poland). Data were normalized using three stable reference genes (UBI - GenBank ID: M60175.1, ACT1 - GenBank ID: AY145451.1, UPL - GenBank ID: XM_045123725.1) and the stability of reference genes in the experimental setup was confirmed using a tool (Bio-Rad) that supports the geNorm algorithm. The RT-qPCR data for the genes and the endogenous controls were obtained from the means of three independent amplification reactions performed on four plants harvested at the same phenological stage (biological replicates). In each RT-qPCR run, extracts from the negative controls as well as the NTC controls were applied. Gene expression data were analyzed using the Bio-Rad CFX Manager (Bio-Rad) software-CFX Maestro v2.0. Relative changes in the gene expression were calculated using the comparative 2-ΔΔCt method and were normalized to the appropriate reference genes [50]. HvGAMYB primers were designed using the Primer3 tool (https://primer3.org/). The complete list of primers and probes used is presented in Supplementary Table S2.
2.15. Statistical analysis
Analysis of variance for observed quantitative traits was performed in the model containing fixed effects of genotypes (CamB, Lubuski - G), treatment (T), development point (DP) (for chlorophyll fluorescence parameters), G × T interaction, G × DP interaction (for chlorophyll fluorescence parameters) and T × DP interaction (for chlorophyll fluorescence parameters). Significant effects were selected at p < 0.001 (approximate threshold resulting from application of the Bonferroni correction with respect to multiple testing for all traits). In case of physiological parameters, due to non-normal distributions of observed variables, analysis of variance was performed on the data transformed by optimal Box-Cox transformation [51]. All statistical computations and visualizations were made in Genstat 22 [52].
3. Results
3.1. Differences in plant phenology, chlorophyll fluorescence kinetics and yield reduction in drought
3.1.1. Phenology
The results of the analysis of variance (ANOVA) showed significant differences (at p < 0.001) between the studied genotypes with respect to time of reaching two phenological stages—flag leaf stage (Z38) and flowering (Z65) - in both types of experiments (Supplementary Table S3). Tillering process was initiated much earlier in the late-heading Lubuski genotype, whereas both flag leaf stage and flowering were reached earlier in CamB compared to Lubuski. In general, the applied treatments caused the delay of plant development with two exceptions (a similar number of DAS was recorded in heading for both CamB and Lubuski plants subjected to C and D+GA conditions in Exp 2) (Figure 1).
3.1.2. Yield performance
The results of ANOVA revealed significant effects of the applied treatments on almost all the investigated yield-related traits (except for NSSm and NSSl) in Exp 1 (Supplementary Table S3). In Exp 2, significant effects of treatment were observed only for four investigated traits (Tn, WGSm, NSSl, TGW). In Exp 1, significant genotypic effects were recorded for traits linked to main spike (LSm, NSSm, NGSm, WGSm) and lateral spike (LSl, NSSl) morphology as well as for PTn (Supplementary Table S3). In the second experiment, the studied genotypes showed significant differences in all the same yield-related traits with one exception (PTn). In addition, in Exp 2, a significant effect of genotype was also noticed for FSl. The mean values of the nine studied traits recorded in both types of experiments are presented in Supplementary Figure S1. The means of most of the studied traits were significantly reduced under drought conditions in Exp 1 (except for Tn and PTn). Greater mean values of PTn were recorded for CamB in D condition (Exp 1) compared to the mean values of this trait recorded for Lubuski. In Exp 2, greater mean values (compared to C condition) of traits associated with the morphology of main spike (NSSm, NGSm, WGSm) as well as lateral spike (NSSl, NGSl, WGSl) were recorded in D condition for CamB.
3.1.3. Chlorophyll fluorescence kinetics and RWC
The results of ANOVA from Exp 1 showed significant effects of genotype for ABS_RC, TRo_RC, ETo_RC, Dlo_RC, and Pi_Abs. Significant effects of DP on all chlorophyll fluorescence parameters were noted in both Exp 1 (except for ETo_RC) and Exp 2 (Supplementary Table S3). In Exp 2, significant effects of the applied treatment were observed for almost all the studied traits (except for Φ_Eo). Significant effects of G × T interaction were recorded for ETo_RC, Ψ_o, and Φ_Eo in Exp 1 and for ABS_RC, TRo_RC, Dlo_RC, Φ_Do, and Pi_Abs in Exp 2. The mean values recorded for chlorophyll fluorescence parameters in both types of experiments are presented in Supplementary Figure S2. The results of ANOVA from both Exp 1 and Exp 2 showed no significant effects of genotypes on relative water content (RWC); this is contrary to the highly significant effects of the applied treatments on this trait observed in both types of experiments (Supplementary Table S3). For both the genotypes, higher mean RWC values were observed in C condition compared to the values recorded in stress conditions in Exp 1 and Exp 2 (Supplementary Figure S3). In Exp 2, higher mean RWC values were observed for CamB in D condition compared to the values recorded for Lubuski in this condition.
3.2. Spike fertility and anther morphology
The results of ANOVA from Exp 1 showed significant effects of the treatments on FSm and FSl and the significant effect of G × T interaction on FSm (Supplementary Table S3). In Exp 2, significant effects of genotype and G × T interaction on FSl were recorded. No significant effects of treatments were observed in this experiment for spike fertility-related traits.
Lubuski showed higher mean FSm values in C condition compared to the Syrian genotype in Exp 1, but no significant differences between the studied genotypes were noticed for this trait in C condition in Exp 2 (Figure 2). In D condition, a rapid increase of FSm values was observed in CamB in both types of experiments. The application of growth regulator (GA) in D condition resulted in decreases in the mean values of FSm compared to the condition where drought was applied alone for both studied genotypes (but no significant differences in mean FSm values between D and D+GA condition were recorded for Lubuski in Exp 2). For Lubuski, lower mean FSm values were recorded in C condition in Exp 1 compared to C condition in Exp 2. For CamB, higher mean values of FSl were observed under D condition in both types of experiments compared to the late-heading genotype. For the Syrian genotype, higher mean FSl values were noticed under all types of treatments (except for D+TR) in Exp 2 compared to Lubuski.
The ANOVA results revealed the significant effects of treatment on anther length and width. Significant effects of G × T interaction were recorded only for anther width—in C condition, a much higher anther width was observed for CamB (Figure 2L).
3.3. Pollen micromorphology and ultrastructure
3.3.1. Pollen grain morphology and viability
The ANOVA results showed significant effects of treatment and G × T interaction on all studied traits associated with pollen morphology in Exp 1 and significant effects of genotype, treatment, and G × T interaction on these traits in Exp 2. The late-heading genotype showed higher mean pollen grain area values in C condition compared to CamB in Exp 1. On the other hand, in Exp 2, higher mean values of this trait were recorded in C condition for the Syrian genotype (Figure 3). No significant differences between genotypes were noticed in D condition in Exp 1, whereas in Exp 2 higher mean values of this trait were observed in D condition for CamB compared to Lubuski. No significant differences in mean pollen grain area values were recorded between C and D conditions for CamB in Exp 1. The D+GA condition negatively affected the pollen grain area of Lubuski in both types of experiments. Similarly, the mean values of pollen grain perimeter, width, and length, decreased in this condition for Lubuski in both types of experiments.
The ANOVA results showed significant effects of treatments on pollen viability in both types of experiments (Supplementary Table S3). In Exp 1, higher mean values of pollen viability (estimated using method 1—Pv1) were recorded in C and D conditions for Lubuski compared to CamB (Supplementary Figure S4). On the other hand, the Syrian genotype showed higher mean values for this trait in D condition compared to C condition. Similar mean values of this trait were observed for D+GA condition in both studied genotypes. The mean values of Pv1 recorded for CamB subjected to D+GA condition in Exp 2 were also much higher compared to D condition in this type of experiment. Pollen viability evaluation by method 2 (Pv2) revealed that in C condition the studied genotypes were characterized by similar levels of pollen viability in Exp 1. For CamB, higher mean values of this trait were recorded in D condition (Exp 1 and Exp 2) compared to the values observed for the late-heading genotype. For Lubuski, much higher mean Pv2 values were recorded in D+TR condition in Exp 1.
3.3.2. Pollen cytological observations
The pollen grains collected from plants in the flowering stage (Z65) analyzed by light microscopy (LM) revealed presence of bi-nucleate or tri-nucleate stages of pollen development and varying degree of cytoplasm vacuolization (Figure 4, Figure 5, Figure 6 and Figure 7).
Transmission electron microscopy (TEM) observations revealed differences in cell’s organelle composition and structures between the studied genotypes grown under different conditions. TEM images of pollen grains collected from the Lubuski plant grown under C condition in Exp 1 showed a normal pollen structure; in the cytoplasm, mitochondria with well-developed cristae, amyloplasts with starch grains, and endoplasmic reticulum (ER) (both smooth and rough) were observed (Figure 8). Trinuclear microspores with visible ER structures and numerous other organelles were observed under this condition in Exp 2. The structure of pollen grains collected from Lubuski plant grown under D condition in Exp 1 showed differentiation in organization: electron-dense areas with starch-rich amyloplasts and numerous mitochondria with swollen cristae (membrane) structures were localized in the organelle-free cytosol. The ultrastructural observations of the pollen grains collected from Lubuski plants subjected to drought in Exp 2 revealed the presence of starch grains, small vacuoles, and numerous vesicles. Under D+GA condition in Exp 1, numerous small vacuoles as well as mitochondria with swollen cristae (membrane) structures were observed for this genotype, whereas under this condition in Exp 2 no alterations in the organelles structure were noticed. Diluted cytoplasmic areas with few organelles and visible ER structures were observed in pollen grains obtained from Lubuski plants grown under D+TR condition in Exp 1. Under the same condition in Exp 2, the pollen grains showed no alterations in cell organization but the presence of numerous vesicles and starch grains in the cytoplasm.
The ultrastructural observations of pollen grains collected from the Syrian genotype grown under optimal (C) water condition in Exp 1 revealed large vacuoles and mitochondria with well-developed crista and amyloplasts (Figure 9). A similar cell organization, but much more amyloplasts with starch grains, was observed in the pollen grains from the Syrian genotype plants grown in this condition in Exp 2. In D condition in Exp 1, there were diluted cytoplasm areas with numerous small vesicles and vacuoles in the pollen grains. In the same water condition in Exp 2, CamB pollen grains showed well-developed ER structure and starch-rich amyloplasts. In the cytoplasm of pollen grains collected from the Syrian genotype grown under D+GA condition in Exp 1, numerous mitochondria and small vacuoles were observed, whereas in the same condition in Exp 2 the pollen grains showed diluted cytoplasm areas with only a few amyloplasts. In the drought condition with the application of the second growth regulator (D+TR), pollen grains collected from CamB plants grown in Exp 1 had diluted cytoplasm areas, mitochondria with swollen cristae (membrane) structures, numerous vesicles and starch grain. Under D+TR condition in Exp 2 trinuclear pollen grains were noticed with well-developed amyloplasts and mitochondria with swollen cristae (membrane) structures.
4. HvGAMYB transcript level
The results of ANOVA showed significant G × T effects on HvGAMYB expression level in the second point of measurement in Exp 1. No significant effects of the variance source on the relative expression level of the studied gene were recorded in Exp 2. In Exp 1, differences in the relative level of expression between the genotypes were noticed in D condition—for CamB, a much higher relative level of HvGAMYB expression was observed in LFE1. During the second developmental point, decreases in the expression of the studied transcription factor were recorded for both genotypes almost in all applied conditions (except for a rapid increase in gene expression recorded for the late-heading genotype in D+TR condition and a stable level of expression recorded for Lubuski in D+GA condition) (Figure 10).
5. Discussion
Of all the stages in the reproduction process, anther and pollen development is the most sensitive to drought [53]. It has been shown that severe drought, which is already fatal for male development, can only limit female organ development, indicating that these organs are insensitive to drought [54]. Therefore, it is crucial to understand the mechanisms and processes associated with drought-related male sterility to ensure food security. The present study examined morphological and cytoembryological changes in the anther and pollen developmental stages in two spring barley genotypes that differ in earliness and drought tolerance. To the best of our knowledge, this study is the first to explore the ultrastructural pollen alterations in spring barley plants exhibiting contrasting growth habits under drought conditions.
5.1. Differences between studied genotypes
Plant genotypes with different growth habits adapt to the stress environment through different ways [55] and show significant differences in their responses to water scarcity [56]. In particular, barley landraces from regions with challenging climatic conditions near the origin of crop domestication are characterized by valuable drought tolerance traits [57]. In this regard, flowering time is one of the most important agronomic traits that affect both grain yield and quality. The Syrian breeding line CamB, studied here, exhibited extraordinary development by achieving reproductive success as early as possible. Its early-heading plants showed flag leaf phase even before the onset of tillering. This phenomenon was also noticed in our previous studies on the genotypes of Syrian origin [36,37,58,59]. Lubuski is an old cultivar adapted to East European conditions, where water availability in the past was different than it is now. In this environment, plants follow a stable growth pattern: during the tillering process, biomass develops, the number of stems increases, and then stem elongation begins. After the simultaneous development of many stems, flowering is triggered by internal and external factors. CamB plants adapted to arid conditions show a different growth strategy: tillering is postponed, and plants use all available resources (water and nutrition) to promote rapid development of the main stem with the spike. After the initiation of flowering and the development of seeds, which guarantee reproductive success, the early-heading genotype starts to develop more tillers. The adjusted developmental pattern may be beneficial under stressful environments where water conditions are unfavorable almost every day [60] — in this case, the plants have a short life cycle which allow their successful adaptation to different abiotic stresses. As frequent episodes of drought driven by climate change are a real threat to agriculture and food security, DE seems to be a promising strategy that can be adopted by plants, which enables to improve crop yield and performance under unfavorable water conditions.
The studied genotypes also varied in spike morphology as this trait is strictly linked to the plant growth pattern [61]. Interestingly, for CamB, similar mean values of NGSm were recorded under C and D conditions (Exp 1), but the mean WGSm values were lower in D, which suggests that this genotype did not reduce the number of seeds under stress condition but the quality of seeds was probably negatively affected by unfavorable conditions. These findings highlight the ability of arid-tolerant plants to adapt to the water condition, which is inconsistent with the general fact that drought limits the number of seeds in barley [62]. However, considering early heading as a drought resistance strategy, it is important to consider the yield quality from an economic point of view.
In many studies, chlorophyll fluorescence has long been used as a convenient and sensitive stress responses indicator of different plant species (e.g., [63,64,65]), including barley [37,66]. In the present study, the ANOVA results showed significant effects of genotype and development points on chlorophyll fluorescence parameters linked to specific energy fluxes per reaction center (RC) (ABS_RC, TRo_RC, Dlo_RC in Exp 1 and ABS_RC, TRo_RC, ETo_RC in Exp 2), which suggests the differences in the drought response strategy of the studied plants and also highlights the negative impact of prolonged stress on plant performance. According to Rapacz et al. [67], drought reduces the number of active RCs, while further steps of photosynthetic electron transfer are less affected, which could explain the findings of the present study. In addition, changes in, for example, ABS_RC recorded here may suggest that the early-heading genotype reacts differently to the initial phase of drought stress, but after a while, alterations in RCs also occur in this plant. Our study confirmed that the chlorophyll fluorescence induction (OJIP) parameters are very sensitive to drought effects compared to changes in RWC in leaves. These parameters reflects the balance between water supply to the leaf tissue and transpiration rate, and thus are considered as an important indicator of plants’ water status [68]. The results of chlorophyll fluorescence analysis followed by OJIP testing corresponded well with the RWC measurements. Although the mean values of this trait recorded in our study decreased in stress conditions, no significant differences were noticed between the stress treatments for both genotypes, which contrasts with the OJIP test showing significant effects of genotype, treatment, and G × T interaction for some parameters.
All these findings suggest that the genotypes studied here are characterized by different growth patterns and drought response strategies (in terms of both yield and chlorophyll fluorescence kinetics), thus constituting a promising model to explore the differences in pollen development under drought conditions.
5.2. Spike fertility and anther morphology
In the present study, significant effects of treatment were observed for the traits associated with spike fertility (FSm and FSl), only in Exp 1. In the experiment with temporary light deprivation, no significant effects of different water conditions were recorded, which suggests that the dark stress affected traits associated with spike fertility and contributed to the limitation of drought effects on spike morphology. Like several previous studies [69,70] demonstrating the negative impact of light distortion on the vegetative and generative cycle of plant development and other different aspects of plant biology [71,72], the present study also revealed that temporary light deficiency is a stronger stressor for seed development than drought occurrence. Interestingly, the drought condition — applied in both Exp 1 and Exp 2 — contributed positively to FSm with increased values recorded for CamB. This can be explained by the fact that this early-heading plant adapted an extraordinary survival strategy that guarantees its wide adaptation in many morphological, biochemical, and physiological aspects to unfavorable conditions. In Exp 2, relatively higher mean FSl values were recorded in C, D, and D+GA conditions for CamB, which can be related to the development of secondary tillers and highlight the effective adaptation of CamB to drought. It is consistent with the nature of tiller development, which is considered a plastic process that is strongly dependent on environmental factors capable of promoting, or repressing, lateral shoot development through a complex network of hormonal and regulatory signals [73].
5.3. Pollen viability and micromorphology
Pollen viability highly depends on the ability of plants to accumulate enough reserve metabolites, especially carbohydrates [74]. Due to drought-induced disruption in the enzymatic processes associated with starch synthesis [75], pollen viability decreases rapidly in plants subjected to water scarcity. Our study only partially confirmed this phenomenon because no differences between C and D conditions were observed for Lubuski — evaluation of pollen viability using the method 1 showed decreases in Pv1 only when additional plant growth regulator were applied. This suggests that starch accumulation in Lubuski seeds was not negatively affected by drought but when there was some disruption in plant development, a rapid decline in pollen viability was noticed. Moreover, better pollen viability (Pv1) recorded for CamB in the D condition than in the C condition also showed the adaptation of this genotype to unfavorable water condition as pollen grains are extremely vulnerable to moisture losses [76]. TTC reacts with dehydrogenase in pollen of plants growing in optimal conditions and appears red, which is considered the most common [77] and indicates enzyme activity in the pollen grains collected from plants grown under different abiotic stresses. Evaluation of pollen viability using method 2 showed the negative impact of all types of drought conditions on Pv2 but still relatively high mean values of this trait were recorded for CamB, suggesting that the Syrian genotype maintained relatively high pollen viability in drought.
The effects of genotype on pollen grain micromorphology were obtained only in Exp 2, which demonstrated the impact of light deprivation on the reproductive phase and, as a consequence, the variations in pollen morphology of plants varied in terms of earliness. Moreover, CamB pollen grain size and shape were stable even under combination of drought and application of artificial growth regulator, which is contrary to our expectations as any perturbations in plant development subjected to drought may additionally damage the cell structures. For Lubuski, almost all traits associated with pollen grain morphology were negatively affected by GA application in drought condition which clearly shows that the pollen grain of the late-heading genotype was vulnerable to the negative impact of growth acceleration. The pollen alterations also had an impact on spike fertility, as in this type of condition, a rapid decrease in mean FSm values was noticed for the late-heading genotype in Exp 1.
5.4. Pollen cytological observations - LM
For all four types of treatments, trinuclear microspores were observed in pollen grains, which indicates that the development of the Syrian genotype’s pollen was not negatively affected under drought conditions or this genotype was able to activate some repair processes. Interestingly, strongly vacuolated microspores were noticed in pollen from plants grown in C and D conditions. It has been shown that vacuolization always occurs during pollen development [78], which suggests that the Syrian genotype is adapted to arid conditions to an extent that under drought conditions the cell structure of pollen grains was almost similar to the pollen collected from plants grown under well-watered condition. Contrary to the observations for CamB, for Lubuski extremely vacuolated, binuclear microspores were observed in pollen for D condition. In a partially dehydrated state, pollen exhibits reduced metabolic activity, lives longer and can better tolerate further desiccation during dispersal [76,79]. Such bicellular pollen consists of a vegetative cell and a sperm precursor cell - the generative cell. Pollen dehydration imposes developmental arrest on maturing pollen, and shifts in the timing of developmental arrest might prevent or enable pollen cell cycle progression to the tricellular condition before dispersal [80]. All together suggest that a delay occurred in pollen development in Lubuski plant as in LFE4 trinuclear pollen should be present [35] or some perturbations have occurred in cell division process causing rapid dehydration of the mother plant in drought condition. The vacuolization process observed in our study was expected as plants under abiotic stress can modulate their development and growth by altering morphological and cellular mechanisms, and cells’ responses to stress might involve changes in the distribution and sorting of specific proteins and molecules. Hence, as vacuoles serve physical and metabolic functions and are essential for cellular responses to general cell homeostasis [81], as well as to abiotic and biotic stresses [82], changes in the number or structure of these organelles are expected in drought condition. On the other hand – as we already mentioned - vacuolization is a common process in pollen development, and microscopic observation of vacuoles did not provide enough information on the effect of drought on these organelles. It seems that the artificial regulation of development (GA) did not affect the reaction of CamB to drought, whereas in this condition (D+GA, Exp 1) pollen grains from Lubuski plants showed chromatin condensation, which may suggest the intensification of cellular processes associated with metabolism remodeling as chromatin rearrangements occur in plant cells subjected to drought [83,84]. Interestingly, in Exp 2 pollen development in samples collected from Lubuski plants did not seem to be affected by any applied stress conditions, whereas in samples obtained from CamB plants grown under the same stress conditions processes associated with chromatin condensation can be visible.
5.5. Pollen ultrastructure observations - TEM
The effects of the applied treatment on the pollen cellular architecture of the two studied genotypes were studied in detail by TEM. The observations indicated that under adverse conditions, the destruction of organelle structures eventually led to a decrease or even loss of cell physiological function [85]. Similar to previous studies reporting the presence of several sterile pollen grains with diluted cytoplasm and reduced starch under water-deficient conditions [54], in our study (Exp 1) pollen grains from both the studied genotypes showed diluted cytoplasm with accumulation of numerous, small vesicles. The functions of these vesicles under drought condition still remain elusive but the appearance of these organelles could be affected by metabolism remodeling triggered by stress as the transport of cargo molecules between compartments is mainly carried out by vesicle shuttles (i.e., transport vesicles) [86]. The findings for previously analyzed parameters indicated that CamB reacted in a similar way to both C and D condition, but ultrastructural observations showed that changes associated with drought-induced reaction can be triggered at the cellular level. Interestingly, as suggested by TEM micrograph observations, pollen grains obtained from CamB plants subjected to D+GA condition did not seem to be affected by drought. This phenomenon was probably linked to the acceleration of plant development resulting in a faster onset of second developmental stages (LFE4) in plants grown under this condition compared to plants grown under D condition. Consequently, D+GA plants were exposed only for a shorter time to stress conditions compared to CamB plants grown under D condition. On the other hand, under D condition with application of the second growth regulator (TR), CamB was subjected for a prolonged time to stress (but the pollen development process was completed) and the TEM observations revealed numerous signs of cellular damage: diluted cytoplasm, accumulation of vesicles and presence of mitochondria with swollen cristae (membrane) structures. The study conducted by Grigorova et al. [87] on wheat revealed that under drought conditions mitochondria were swollen and vacuolized, which can be observed on TEM micrographs of pollen grains collected from CamB plants subjected to D+TR condition. Some studies suggest that developmental alterations can be irreversible [88,89] and highlight the negative effects of growth modification applied under drought in plants adapted to arid condition. Although the extent of mitochondrial damage in plants grown under drought is hard to specify [90], it is probably related to the species and variety of plants [91]. These types of mitochondrial alterations were also observed in pollen grains from Lubuski plants grown under D and D+GA conditions in Exp 1 in our study. Interestingly, no swollen mitochondrial membranes were noticed on TEM micrographs for CamB plants grown under D conditions in this type of experiment. It seems that cellular organization was not that much affected by drought as the organelles of pollen grains collected from Lubuski plants subjected to drought in Exp 1. Diluted cytoplasm structures were observed in the micrographs of pollen grains taken from CamB plants grown under D+GA condition in Exp 2, contrary to Exp 1, where similar cellular alterations were noticed for D+TR conditions. This was probably associated with the changes in plant development triggered by growth regulators — in Exp 2 acceleration in plant development caused that the CamB plants were subjected to light deprivation conditions from a very beginning stress duration, whereas the plants grown under C, D, and D+TR conditions modified their growth which allowed the completion of pollen grain development when dark stress was over. Although, the presence of dense systems of ER on the TEM micrographs of CamB plants grown under D condition in Exp 2 may suggest same perturbations in pollen development as abiotic stress can lead to misfolding of proteins and their accumulation, causing an ER stress situation [92,93]. This may be linked to the modification of plant development by light deprivation stress. In Exp 1 for the late-heading genotype, drought-induced alterations were observed in organelle structures—mitochondria with swollen membrane structures in D and D+GA conditions and diluted cytoplasm areas with a small number of cell organelles. The presence of dense systems of ER was noticed on the micrographs of Lubuski plants grown under D+TR condition in Exp 1, which showed that some processes related to cellular metabolism modification were triggered. This can be explained by prolonged exposure of plants to drought but did not provide enough information about the disruption of the pollen development process. Surprisingly, no such cellular changes were observed on the TEM micrographs of Lubuski plants grown under the same condition in Exp 2. This was presumably associated with the late growth habit of Lubuski and the delay of development process, especially under drought — the plants limited the speed of development and extended the period between light deprivation and the flowering process. Moreover, stress exposure of mother plants did not affect the development of spikes in the secondary tillering process, which indicated that Lubuski plants managed to obtain enough nutrition to complete pollen development after drought.
5.6. HvGAMYB expression fluctuation
The present study confirmed the expression of HvGAMYB in the anther tissues of plants subjected to different water conditions. In the study conducted by Murray et al. [94], a progressive decrease in anther size was associated with the increase in GAMYB levels, particularly a decrease in anther length. These findings are in line with the results of the present study, in which for CamB plants with a higher level of HvGAMYB expression and lower mean values of anther width (trait linked to anther morphology) were recorded compared to Lubuski plants under the same stress treatment. In our study, HvGAMYB was expressed at a relatively high level in anthers collected from the CamB plants subjected to drought in LFE1. Transgenic barley lines with a fourfold higher level of endogenous GAMYB protein in their anthers were reported to be male sterile [94,95]. In our study, the relatively high expression level of this transcription factor in anthers collected from CamB plants cannot be linked to pollen — even partially — sterility as in D condition the traits associated with pollen morphology as well as pollen viability did not seem to be affected by drought. Interestingly, under D conditions with the application of growth regulator, the relative expression levels of HvGAMYB were lower compared to those recorded under drought condition without spraying interactions, which highlighted the complex nature of the expression fluctuations of some transcription factors in terms of plant growth modifications under drought condition. Surprisingly, HvGAMYB expression was relatively high in tissues collected from Lubuski plants grown under D+TR in LFE4. This condition should have resulted in plant development delay—this phenomenon can be linked to the involvement of this gene in pollen and anther development process [94] in spike tissues developing during secondary tillering. The results of this study clearly showed that plant development perturbations affected the circadian rhythm functions and the expression of transcription factors, which can be directly linked to the development of generative organs and pollen. Light deprivation applied in Exp 2 did not result in significant differences in HvGAMYB expression—contrary to the treatment applied in Exp 1. As HvGAMYB is part of the GA-dependent flowering pathway [14], this finding confirmed the existence of a close association between gibberellin and photoperiod pathways [19] and showed the potential role of HvGAMYB as a flowering pathway integrator.
6. Conclusions
This study clearly showed that the duration of the vegetation period has an influence on plant’s responses to drought condition as every modification (application of growth regulators or light deprivation) can change the dynamics of drought effects as well as the duration of plant’s exposure to drought. As a consequence, plants exhibited different drought reactions which also had an impact on pollen development. When the mother plant experiences stress conditions, the signal will reach pollen grains and affect pollen preparation and the timing of pollen dispersal [96,97]. The results of our study suggest that this conclusion is partially correct—in some cases, the exposure of mother plant to drought did not seem to severely affect pollen grain development in spikes that elude (by growth delay or secondary tillering process) the unfavorable conditions and their consequences, even in plants not adapted to arid condition. In our study, CamB and Lubuski mother plants were affected by drought which was confirmed by the evaluation of yield performance and chlorophyll fluorescence kinetics. However, the pollen development of plants grown under specific condition (e.g., Lubuski in Exp 2) was not much affected, contrary to our expectations. Even though no damage in cellular components was noticed in pollen grains for the late-heading genotype in TEM observations, the evaluation of pollen viability indicated low pollen quality after dispersal. An important indicator of proper pollen development may also be the two proposed traits associated with spike fertility, which showed clearly a negative impact of drought on the generative process, and consequently, seed development. According to phenology observations for Exp 2, the timing between the flag leaf stage (where stresses begin) and flowering was extremely short for CamB plants, due to which this genotype completed the flowering process under light deprivation treatment. At the same time, the late-heading genotype delayed generative development and, probably, recovered to some extent, which can be observed in TEM and LM micrographs. The results of this study suggest that growth modification, as well as perturbations in light distribution, can affect the HvGAMYB expression. As this transcription factors should be expressed in a tailored pattern, the changes in the expression can have a significant impact on many development-linked processes. As we previously stated [36,37], growth pattern modification may be an effective strategy that allows plants to cope with unfavorable water conditions—this study showed that the duration of the vegetative period is an important factor for proper pollen development, which significantly affects the biological success of plants.
Supplementary Materials
Supplementary Table S1: The dates of sample collection for microscopic observation conducted in both experiments (Exp 1 and Exp 2. Treatments: C – control condition; D – drought condition; D+GA - drought condition combined with GA3 application; D+TR - drought condition combined with Trinexapac application. NoD – number of days, the first day is the day of droughts beginning. Supplementary Table S2: DNA sequences of the gene specific primers. Supplementary Table S3: Results of analysis of variance for observed traits (P values for testing significance of variation sources). Supplementary Figure S1: Mean values of yield-related traits (with standard errors). Supplementary Figure S2: Mean values (with standard errors) recorded for chlorophyll fluorescence parameters in both type of experiments at two development points (LFE1 and LFE3. Supplementary Figure S3: Means values (with standard errors) of RWC recorded for two contrasting genotypes in different water regimes in both type of experiments. Supplementary Figure S4: Mean values (with standard errors) of pollen viability evaluation traits (two different methods: A – KI/I2 – Pv1; B – TTC - Pv2) recorded for two contrasting genotypes in different water regimes in both type of experiments.
Funding
The research was supported by National Science Centre, Poland, project SONATA 12 no. 2016/23/D/NZ9/00042
Author Contribution
P.O., conceptualization, methodology, investigation, writing—original draft preparation, funding acquisition, project administration, data visualization. M.K.W., methodology, investigation, writing—review and editing. A.K., supervision, methodology, investigation, writing—review and editing. P.K., statistical data analysis, validation, data visualization, writing—review and editing. M.K., investigation, writing—review. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflict of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Dong, B.; Zheng, X.; Liu, H.; Able, J.A.; Yang, H.; Zhao, H.; Zhang, M.; Qiao, Y.; Wang, Y.; Liu, M. Effects of Drought Stress on Pollen Sterility, Grain Yield, Abscisic Acid and Protective Enzymes in Two Winter Wheat Cultivars. Front. Plant Sci. 2017, 8, 1008. [Google Scholar] [CrossRef] [PubMed]
- Calleja-Cabrera, J.; Boter, M.; Oñate-Sánchez, L.; Pernas, M. Root Growth Adaptation to Climate Change in Crops. Front. Plant Sci. 2020, 11, 544. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Gopi, R.; Sankar, B.; Gomathinayagam, M.; Panneerselvam, R. Differential Responses in Water Use Efficiency in Two Varieties of Catharanthus Roseus under Drought Stress. Comptes. Rendus. Biol. 2008, 331, 42–47. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Santiago, J.P.; Sharkey, T.D. Pollen development at high temperature and role of carbon and nitrogen metabolites. Plant Cell Environ. 2019, 42, 2759–2775. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Jiang, M.; Guo, C. Crop Pollen Development under Drought: From the Phenotype to the Mechanism. Int. J. Mol. Sci. 2019, 20, 1550. [Google Scholar] [CrossRef] [PubMed]
- Alqudah, A.M.; Samarah, N.H.; Mullen, R.E. Drought stress effect on crop pollination, seed set, yield and quality. 2011, pp. 193-213. In: Lichtfouse E. (ed), Alternative farming systems, biotechnology, drought stress and ecological fertilisation, Sustainable Agriculture Reviews 6. Springer Science+Business Media B.V. ISBN 978-94-007-0186-1. [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, L. Specification of tapetum and microsporocyte cells within the anther. Curr. Opin. Plant Biol. 2014, 17, 49–55. [Google Scholar] [CrossRef]
- Gómez, J.F.; Talle, B.; Wilson, Z.A. Anther and pollen development: A conserved developmental pathway. J. Integr. Plant Biol. 2015, 57, 876–91. [Google Scholar] [CrossRef]
- Kwon, C.T.; Kim, S.H.; Kim, D.; Paek, N.C. The rice floral repressor Early flowering1 affects spikelet fertility by modulating gibberellin signaling. Rice, 2015, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Plackett. A.R.; Thomas, S.G.; Wilson, Z.A.; Hedden, P. Gibberellin control of stamen development: a fertile field. Trends Plant Sci. 2011, 16, 568–78. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, H.; Aya, K.; Ueguchi-Tanaka, M.; Shimada, Y.; Nakazono, M.; Watanabe, R.; Nishizawa, N.; Gomi, K.; Shimada, A.; Kitano, H.; et al. GAMYB controls different sets of genes and is differentially regulated by microRNA in aleurone cells and anthers. Plant J, 2006, 47, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, Y.; Zhu, J.K. Thriving under stress: how plants balance growth and the stress response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Aya, K.; Ueguchi-Tanaka, M.; Kondo, M.; Hamada, K.; Yano, K.; Nishimura, M.; Matsuoka, M. Gibberellin modulates anther development in rice via the transcriptional regulation of GAMYB. Plant Cell. 2009, 21, 1453–72. [Google Scholar] [CrossRef] [PubMed]
- Osnato, M.; Castillejo, C.; Matías-Hernández, L.; Pelaz, S. TEMPRANILLO genes link photoperiod and gibberellin pathways to control flowering in Arabidopsis. Nat Commun. 2012, 3, 808. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Pan, J.; Li, Y.; Lou, D.; Hu, Y.; Yu, D. The DELLA-CONSTANS Transcription Factor Cascade Integrates Gibberellic Acid and Photoperiod Signaling to Regulate Flowering. Plant Physiol. 2016, 172, 479–88. [Google Scholar] [CrossRef]
- Quiroz, S.; Yustis, J.C.; Chávez-Hernández, E.C.; Martínez, T.; Sanchez, M.d.l.P.; Garay-Arroyo, A.; Álvarez-Buylla, E.R.; García-Ponce, B. Beyond the Genetic Pathways, Flowering Regulation Complexity in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 5716. [Google Scholar] [CrossRef]
- Roy, S. Function of MYB domain transcription factors in abiotic stress and epigenetic control of stress response in plant genome. Plant Signal Behav. 2016, 11, e1117723. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.; Wang, Y.; Liu, Z.; Gao, C. Comprehensive Analysis of MYB Gene Family and Their Expressions Under Abiotic Stresses and Hormone Treatments in Tamarix hispida. Front. Plant Sci. 2018, 9, 1303. [Google Scholar] [CrossRef]
- Jackson, R.B.; Sperry, J.S.; Dawson, T.E. Root water uptake and transport: using physiological predictions in global predictions. Trends in Plant Sci. 2000, 5, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.M. Osmoregulation and water stress in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 299–319. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Simontacchi, M.; Tambussi, E.; Beltrano, J.; Montaldi, E.; Puntarulo, S. Drought and watering-dependent oxidative stress: effect on antioxidant content in Triticum aestivum L. leaves. J. Exp. Bot. 1999, 50, 375–383. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought - from genes to the whole plant. Funct Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Maherali, H.; Caruso, C.M.; Sherrard, M.E.; Latta, R.G. Adaptive value and costs of physiological plasticity to soil moisture limitation in recombinant inbred lines of Avena barbata. Am. Nat. 2010, 175, 211–224. [Google Scholar] [CrossRef]
- Dolferus, R. To grow or not to grow: a stressful decision for plants. Plant Sci. 2014, 229, 247–261. [Google Scholar] [CrossRef]
- Willige, B.C.; Ogiso-tanaka, E.; Zourelidou, M.; Schwechheimer, C. (2012). WAG2 represses apical hook opening downstream from gibberellin and PHYTOCHROME INTERACTING FACTOR 5. Development 2012, 139, 4020–4028. [Google Scholar] [CrossRef]
- de Lucas, M.; Davière, J.M.; Rodríguez-Falcón, M.; Pontin, M.; Iglesias-Pedraz, J.M.; Lorrain, S.; Fankhauser, C.; Blázquez, M.A.; Titarenko, E.; Prat, S. A molecular framework for light and gibberellin control of cell elongation. Nature 2008, 451, 480–4. [Google Scholar] [CrossRef]
- van der Graaff, E.; Schwacke, R.; Schneider, A.; Desimone, M.; Flugge, U.I.; Kunze, R. Transcription analysis of Arabidopsis membrane transporters and hormone pathways during developmental and induced leaf senescence. Plant Physiol. 2006, 141, 776–792. [Google Scholar] [CrossRef]
- Guo, Y.; Gan, S.S. Convergence and divergence in gene expression profiles induced by leaf senescence and 27 senescence-promoting hormonal, pathological and environmental stress treatments. Plant Cell Environ. 2012, 35, 644–655. [Google Scholar] [CrossRef]
- Roberts, I.N.; Veliz, C.G.; Criado, M.V.; Signorini, A.; Simonetti, E.; Caputo, C. Identification and expression analysis of 11 subtilase genes during natural and induced senescence of barley plants. J. Plant Physiol. 2017, 211, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Rankenberg, T.; Geldhof, B.; van Veen, H.; Holsteens, K.; Van de Poel, B.; Sasidharan, R. Age-dependent abiotic stress resilience in plants. Trends Plant Sci. 2021, 26, 692–705. [Google Scholar] [CrossRef] [PubMed]
- Gürel, F.; Öztürk, Z.N.; Uçarlı, C.; Rosellini, D. (2016) Barley Genes as Tools to Confer Abiotic Stress Tolerance in Crops. Front. Plant Sci. 2016, 7, 1137. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.F.; Wilson, Z.A. Non-destructive staging of barley reproductive development for molecular analysis based upon external morphology. J. Exp. Bot. 2012, 63, 4085–94. [Google Scholar] [CrossRef] [PubMed]
- Ogrodowicz, P.; Adamski, T.; Mikołajczak, K.; Kuczyńska, A.; Surma, M.; Krajewski, P.; Sawikowska, A.; Górny, A.G.; Gudyś, K.; Szarejko, I.; et al. QTLs for earliness and yield-forming traits in the Lubuski × CamB barley RIL population under various water regimes. J. Appl. Genet. 2017, 58, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Ogrodowicz, P.; Kuczyńska, A.; Krajewski, P.; Kempa, M. The effects of heading time on yield performance and HvGAMYB expression in spring barley subjected to drought. J. Appl. Genet. 2023, 64, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Ogrodowicz, P.; Mikołajczak, K.; Kempa, M.; Mokrzycka, M.; Krajewski, P.; Kuczyńska, A. Genome-wide association study of agronomical and root-related traits in spring barley collection grown under field conditions. Front. Plant Sci. 2023, 14, 1077631. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Kuczyńska, A.; Cardenia, V.; Ogrodowicz, P.; Kempa, M.; Rodriguez-Estrada, M.T.; Mikołajczak, K. Effects of multiple abiotic stresses on lipids and sterols profile in barley leaves (Hordeum vulgare L.). Plant Physiol. Biochem. 2019, 141, 215–224. [Google Scholar] [CrossRef]
- Mikołajczak, K.; Kuczyńska, A.; Krajewski, P.; Sawikowska, A.; Surma, M.; Ogrodowicz, P.; Adamski, T.; Krystkowiak, K.; Górny, A.G.; Kempa, M.; et al. Quantitative trait loci for plant height in Maresi × CamB barley population and their associations with yield-related traits under different water regimes. J. Appl. Genet. 2017, 58, 23–35. [Google Scholar] [CrossRef]
- Boden, S.A.; Weiss, D.; Ross, J.J.; Davies, N.W.; Trevaskis, B.; Chandler, P.M.; Swain, S.M. EARLY FLOWERING3 regulates flowering in spring barley by mediating gibberellin production and FLOWERING LOCUS T expression. Plant Cell 2014, 26, 1557–1569. [Google Scholar] [CrossRef]
- Grijalva-Contreras, R.L.; Macías-Duarte, R.; Martínez-Díaz, G.; Robles-Contreras, F.; Nuñez-Ramírez, F. Effects of trinexapac-ethyl on different wheat varieties under desert conditions of Mexico. Agric. Sci. 2012, 3, 658–662. [Google Scholar] [CrossRef]
- Browne, R.G.; Iacuone, S.; Li, S.F.; Dolferus, R.; Parish, R.W. Anther Morphological Development and Stage Determination in Triticum aestivum. Front. Plant Sci. 2018, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Huang, H.J.; Ren, S.T.; Li, J.J.; Sun, Y.; Sun, D.Y. Zhang, S.Q. The rice wall-associated receptor-like kinase gene OsDEES1 plays a role in female gametophyte development. Plant Physiol. 2012, 160, 696–707. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, J.; Dong, J.; Yu, J.; Huang, S.; Lin, H.; Hu, S.; Wang, J. Optimized qualitative and quantitative methods for barley viability testing using triphenyl tetrazolium chloride staining. Cereal Chem. 2019, 96, 421–428. [Google Scholar] [CrossRef]
- Impe, D.; Reitz, J.; Köpnick, C.; Rolletschek, H.; Börner, A.; Senula, A.; Nagel, M. Assessment of pollen viability for wheat. Front. Plant Sci. 2020, 10, 1588. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A Re-Examination of the Relative Turgidity Techniques for Estimating Water Deficits in Leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–22. [Google Scholar] [CrossRef]
- Dawidziuk, A.; Koczyk, G.; Popiel, D.; Kaczmarek, J.; Buśko, M. Molecular diagnostics on the toxigenic potential of Fusarium spp. Plan pathogens. J. Appl. Microbio. 2014, 16, 1607–1620. [Google Scholar] [CrossRef] [PubMed]
- Box, G.E.P.; Cox, D.R. An analysis of transformations. J. R. Stat. Soc. Series B. 1964, 26, 211–252. [Google Scholar] [CrossRef]
- VSN International. Genstat for Windows 22nd Edition. VSN International 2022, Hemel Hempstead, UK. Web page: Genstat.co.uk.
- Guo, C.; Yao, L.; You, C.; Wang, S.; Cui, J.; Ge, X.; Ma, H. MID1 plays an important role in response to drought stress during reproductive development. Plant J. 2016, 88, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Saini, H.S. Effects of water stress on male gametophyte development in plants. Sex. Plant Reprod. 1997, 10, 67–73. [Google Scholar] [CrossRef]
- Bakhtiari, M.; Formenti, L.; Caggia, V.; Glauser, G.; Rasmann, S. Variable effects on growth and defense traits for plant ecotypic differentiation and phenotypic plasticity along elevation gradients. Ecol. Evol. 2019, 9, 3740–3755. [Google Scholar] [CrossRef] [PubMed]
- Zare, M.; Azizi, M.H.; Bazrafshan, F. Effect of drought stress on some agronomic traits in ten barley (Hordeum vulgar) cultivars. TJEAS 2011, 1, 57–62. [Google Scholar]
- Ellis, R.P.; Forster, B.P.; Robinson, D.; Handley, L.L.; Gordon, D.C.; Russell, J.R.; Powell, W. Wild barley: A source of genes for crop improvement in the 21st century? J. Exp. Bot. 2000, 51, 9–17. [Google Scholar] [CrossRef]
- Mikołajczak, K.; Ogrodowicz, P.; Gudyś, K.; Krystkowiak, K.; Sawikowska, A.; Frohmberg, W.; Górny, A.; Kędziora, A.; Jankowiak, J.; Józefczyk, D.; et al. Quantitative Trait Loci for Yield and Yield-Related Traits in Spring Barley Populations Derived from Crosses between European and Syrian Cultivars. PLoS ONE 2016, 11(5), e0155938. [Google Scholar] [CrossRef]
- Ogrodowicz, P.; Kuczyńska, A.; Mikołajczak, K.; Adamski, T.; Surma, M.; Krajewski, P.; Ćwiek-Kupczyńska, H.; Kempa, Michał.; Rokicki, Michał.; Jasińska, D. Mapping of quantitative trait loci for traits linked to fusarium head blight in barley. PLoS ONE 2020, 15(2), e0222375. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, Y.; Ma, Y.; Yang, X.; Yin, X.; Zhang, Y.; Xiao, Y.; Liu, W.; Li, Y.; Li, S.; et al. The Strawberry Transcription Factor Farav1 Positively Regulates Anthocyanin Accumulation by Activation of Famyb10 and Anthocyanin Pathway Genes. Plant Biotechnol. J. 2020, 18, 2267–2279. [Google Scholar] [CrossRef]
- Gol, L.; Haraldsson, E.B.; von Korff, M. Ppd-H1 integrates drought stress signals to control spike development and flowering time in barley. J. Exp. Bot. 2021, 72, 122–136. [Google Scholar] [CrossRef]
- Oguz, M.C.; Aycan, M.; Oguz, E.; Poyraz, I.; Yildiz, M. Drought Stress Tolerance in Plants: Interplay of Molecular, Biochemical and Physiological Responses in Important Development Stages. Physiologia 2022, 2, 180–197. [Google Scholar] [CrossRef]
- Goltsev, V.; Chernev, P.; Zaharieva, I.; Lambrev, P.; Strasser, R.J. Kinetics of delayed chlorophyll a fluorescence registered in milliseconds time range. Photosynth. Res. 2005, 84, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant 2016, 38, 1–11. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef] [PubMed]
- Daszkowska-Golec, A.; Collin, A.; Sitko, K.; Janiak, A.; Kalaji, H.M.; Szarejko, I. Genetic and Physiological Dissection of Photosynthesis in Barley Exposed to Drought Stress. Int. J. Mol. Sci. 2019, 20, 6341. [Google Scholar] [CrossRef] [PubMed]
- Rapacz, M.; Wójcik-Jagła, M.; Fiust, A.; Kalaji, H.M. Kościelniak, J. Genome-Wide Associations of Chlorophyll Fluorescence OJIP Transient Parameters Connected With Soil Drought Response in Barley. Front. Plant Sci. 2019, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Soltys-Kalina, D.; Plich, J.; Strzelczyk-Żyta, D.; Śliwka, J.; Marczewski, W. The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of 'Katahdin'-derived potato cultivars. Breed. Sci. 2016, 66, 328–331. [Google Scholar] [CrossRef]
- Longstaff, B.J.; Loneragan, N.R.; O’Donohue, M.J.; Dennison, W.J. Effects of light deprivation on the survival and recovery of the seagrass Halophila ovalis (R.Br.) Hook. J. Exp. Mar. Biol. Ecol. 1999, 234, 1–27. [Google Scholar] [CrossRef]
- Lu, D.; Liu, B.; Ren, M.; Wu, C.; Ma, J.; Shen, Y. Light Deficiency Inhibits Growth by Affecting Photosynthesis Efficiency as well as JA and Ethylene Signaling in Endangered Plant Magnolia sinostellata. Plants 2021, 10, 2261. [Google Scholar] [CrossRef]
- Liu, H.; Wang, X.; Ren, K.; Li, K.; Wei, M.; Wang, W.; Sheng, X. Light Deprivation-Induced Inhibition of Chloroplast Biogenesis Does Not Arrest Embryo Morphogenesis But Strongly Reduces the Accumulation of Storage Reserves during Embryo Maturation in Arabidopsis. Front. Plant Sci. 2017, 8, 1287. [Google Scholar] [CrossRef]
- Paluch-Lubawa, E.; Stolarska, E.; Sobieszczuk-Nowicka, E. Dark-Induced Barley Leaf Senescence – A Crop System for Studying Senescence and Autophagy Mechanisms. Front. Plant Sci. 2021, 12, 635619. [Google Scholar] [CrossRef]
- Kebrom, T.H.; Chandler, P.M.; Swain, S.M.; King, R.W.; Richards, R.A.; Spielmeyer, W. Inhibition of tiller bud outgrowth in the tin mutant of wheat is associated with precocious internode development. Plant Physiol. 2012, 160, 308–18. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, S.V.K. Heat stress during flowering in cereals – effects and adaptation strategies. New Phytol. 2020, 226, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; Nepi, M.; Pacini, E. Water status and associated processes mark critical stages in pollen development and functioning. Ann. Bot. 2012, 109, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Riano, T.; Dafni, A. A new procedure to asses pollen viability. Sex. Plant Reprod. 2000, 12, 241–244. [Google Scholar] [CrossRef]
- Pacini, E.; Jacquard, C.; Clément, C. Pollen vacuoles and their significance. Planta 2011, 234, 217–227. [Google Scholar] [CrossRef]
- Hoekstra, F.A.; Golovina, E.A.; Buitink, J. Mechanisms of plant desiccation tolerance. Trends in Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef]
- Williams, J.H.; Taylor, M.L.; O'Meara, B.C. Repeated evolution of tricellular (and bicellular) pollen. Am. J. Bot. 2014, 101, 559–71. [Google Scholar] [CrossRef]
- Martinoia, E.; Maeshima, M.; Neuhaus, H.E. Vacuolar transporters and their essential role in plant metabolism. J. Exp. Bot. 2007, 58, 83–102. [Google Scholar] [CrossRef]
- Signorelli, S.; Tarkowski, Ł.P.; Van den Ende, W.; Bassham, D.C. Linking Autophagy to Abiotic and Biotic Stress Responses. Trends Plant Sci. 2019, 24, 413–430. [Google Scholar] [CrossRef]
- Han, S.K.; Wagner, D. Role of chromatin in water stress responses in plants. J. Exp. Bot. 2014, 65, 2785–2799. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; Sasaki, T.; Ueda, M.; Sako, K.; Seki, M. Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Front. Plant Sci. 2015, 6, 114. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Boutté, Y.; Vernhettes, S.; Satiat-Jeunemaitre, B. Involvement of the cytoskeleton in the secretory pathway and plasma membrane organisation of higher plant cells. Cell Biol. Int. 2007, 31, 649–54. [Google Scholar] [CrossRef] [PubMed]
- Grigorova, B.; Vassileva, V.; Klimchuk, D.; Vaseva, I.; Demirevska, K.; Feller, U. (2012) Drought, high temperature, and their combination affect ultrastructure of chloroplasts and mitochondria in wheat (Triticum aestivum L.) leaves. J. Plant Interact. 2012, 7, 204–213. [Google Scholar] [CrossRef]
- Liu, J.X.; Bennett, J. Reversible and irreversible drought-induced changes in the anther proteome of rice (Oryza sativa L.) genotypes IR64 and Moroberekan. Mol. Plant. 2011, 4, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Kong, F.; Zhang, S.; Meng, X.; Wang, Y.; Meng, Q. A tomato chloroplast-targeted DnaJ protein protects Rubisco activity under heat stress. J. Exp. Bot. 2015, 66, 3027–3040. [Google Scholar] [CrossRef]
- Tang, H.; Zhu, H. Specific Changes in Morphology and Dynamics of Plant Mitochondria under Abiotic Stress. Horticulturae 2023, 9, 11. [Google Scholar] [CrossRef]
- Vassileva, V.; Simova-Stoilova, L.; Demirevska, K.; Feller, U. Variety-specific response of wheat (Triticum aestivum L.) leaf mitochondria to drought stress. J. Plant Res. 2009, 122, 445–454. [Google Scholar] [CrossRef]
- Wan, S.; Jiang, L. Endoplasmic reticulum (ER) stress and the unfolded protein response (UPR) in plants. Protoplasma 2016, 253, 753–764. [Google Scholar] [CrossRef]
- Wang, X.; Xu, M.; Gao, C.; Zeng, Y.; Cui, Y.; Shen, W.; Jiang, L. The roles of endomembrane trafficking in plant abiotic stress responses. J. Integr. Plant Biol. 2020, 62, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Murray, F.; Kalla, R.; Jacobsen, J.; Gubler, F. A role of HvGAMYB in anther development. Plant J. 2003, 33, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Duca, M.; Port, A.; Orozco-Cardenas, M.; Lovatt, C. Gibberellin-Induced Gene Expression Associated with Cytoplasmic Male Sterility in Sunflower. Biotechnol. Biotechnol. Equip. 2008, 22, 691–698. [Google Scholar] [CrossRef]
- Begcy, K.; Dresselhaus, T. Epigenetic responses to abiotic stresses during reproductive development in cereals. Plant Reprod. 2018, 31, 343–355. [Google Scholar] [CrossRef]
- Pacini, E.; Dolferus, R. Pollen Developmental Arrest: Maintaining Pollen Fertility in a World With a Changing Climate. Front. Plant Sci. 2019, 10, 679. [Google Scholar] [CrossRef]
Figure 1.
Phenological differentiation between the studied genotypes. Photos showing plants sowed at the same time (0 DAS) and subjected to different water regimes: CamB plants grown under C condition (A), D condition (B), D+GA condition (C), and D+TR condition (D) and Lubuski plants grown under C condition (E), D condition (F), D+GA condition (G), and D+TR condition (H). The development pattern of the studied genotypes representing different times (DAS) of tillering (I), flag leaf (J), flowering (K), and heading (L) stages. DAS—days after sowing.
Figure 1.
Phenological differentiation between the studied genotypes. Photos showing plants sowed at the same time (0 DAS) and subjected to different water regimes: CamB plants grown under C condition (A), D condition (B), D+GA condition (C), and D+TR condition (D) and Lubuski plants grown under C condition (E), D condition (F), D+GA condition (G), and D+TR condition (H). The development pattern of the studied genotypes representing different times (DAS) of tillering (I), flag leaf (J), flowering (K), and heading (L) stages. DAS—days after sowing.
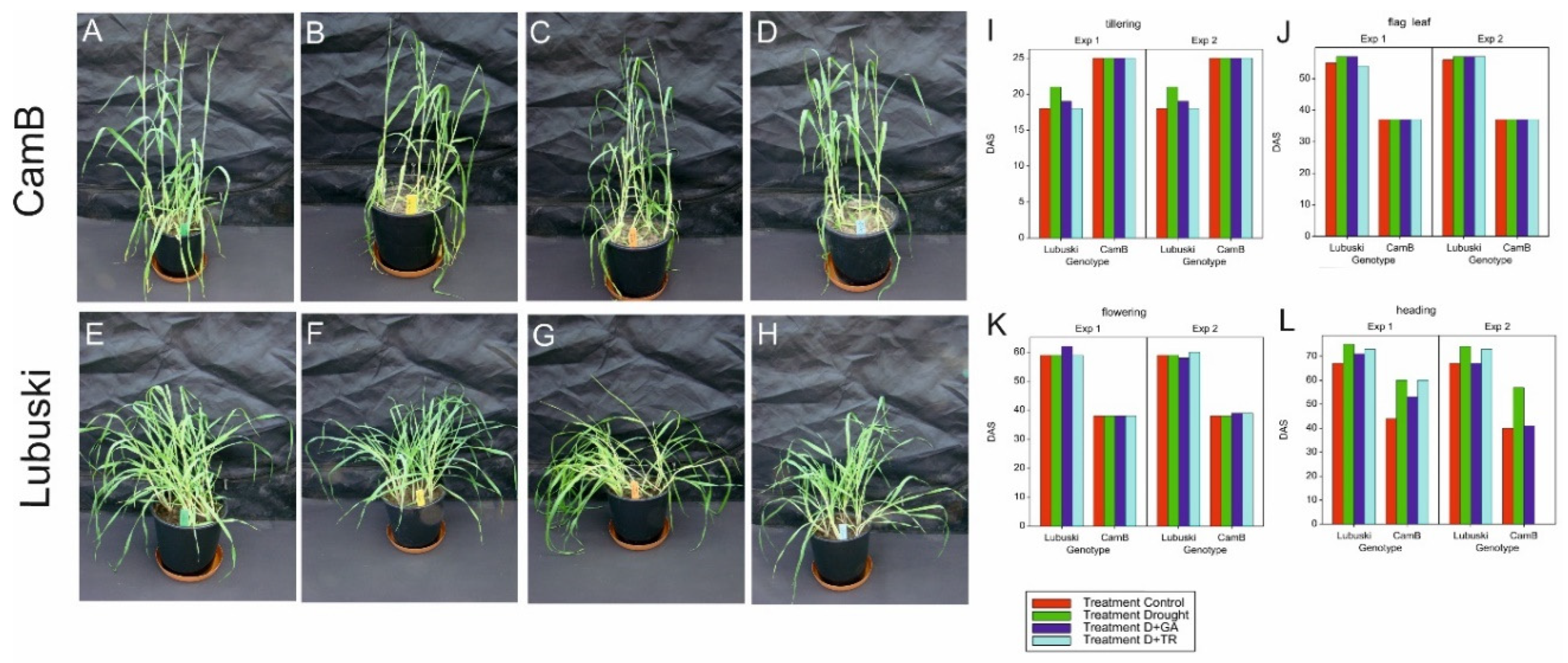
Figure 2.
Photos showing the differences in the morphology of spikes and anthers among the studied plants subjected to different water regimes in Exp 1: spikes and anthers of CamB plants grown under C condition (A), D condition (B), D+GA condition (C), and D+TR condition (D) and spikes and anthers of Lubuski plants grown under C condition (E), D condition (F), D+GA condition (G), and D+TR condition (H). Mean values (with standard errors) of FSm (I), FSl (J), anther length (K), and width (L) recorded for the studied genotypes under four different applied treatments.
Figure 2.
Photos showing the differences in the morphology of spikes and anthers among the studied plants subjected to different water regimes in Exp 1: spikes and anthers of CamB plants grown under C condition (A), D condition (B), D+GA condition (C), and D+TR condition (D) and spikes and anthers of Lubuski plants grown under C condition (E), D condition (F), D+GA condition (G), and D+TR condition (H). Mean values (with standard errors) of FSm (I), FSl (J), anther length (K), and width (L) recorded for the studied genotypes under four different applied treatments.
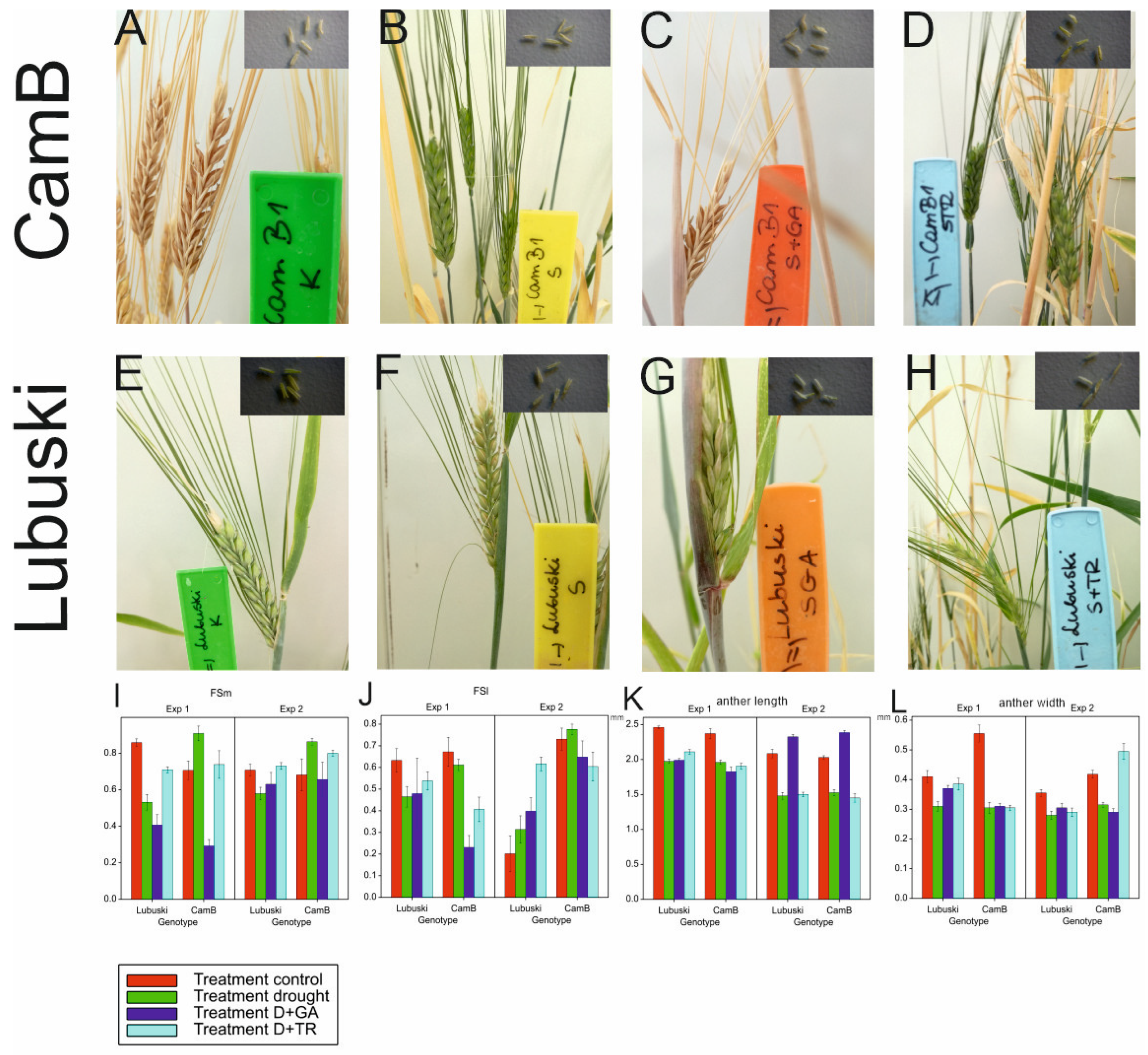
Figure 3.
Images presenting the LM analysis results obtained using two different methods of pollen viability evaluation (Pv1 and Pv2): pollen grains collected from CamB plants grown under four types of water regime in Exp 1 (A) and Exp 2 (B), pollen grains collected from Lubuski plants grown under four types of water regime in Exp 1 (C) and Exp 2 (D). Mean values (with standard errors) of pollen morphology-related traits: pollen grain area (E), perimeter (F), width (G), and length (H).
Figure 3.
Images presenting the LM analysis results obtained using two different methods of pollen viability evaluation (Pv1 and Pv2): pollen grains collected from CamB plants grown under four types of water regime in Exp 1 (A) and Exp 2 (B), pollen grains collected from Lubuski plants grown under four types of water regime in Exp 1 (C) and Exp 2 (D). Mean values (with standard errors) of pollen morphology-related traits: pollen grain area (E), perimeter (F), width (G), and length (H).
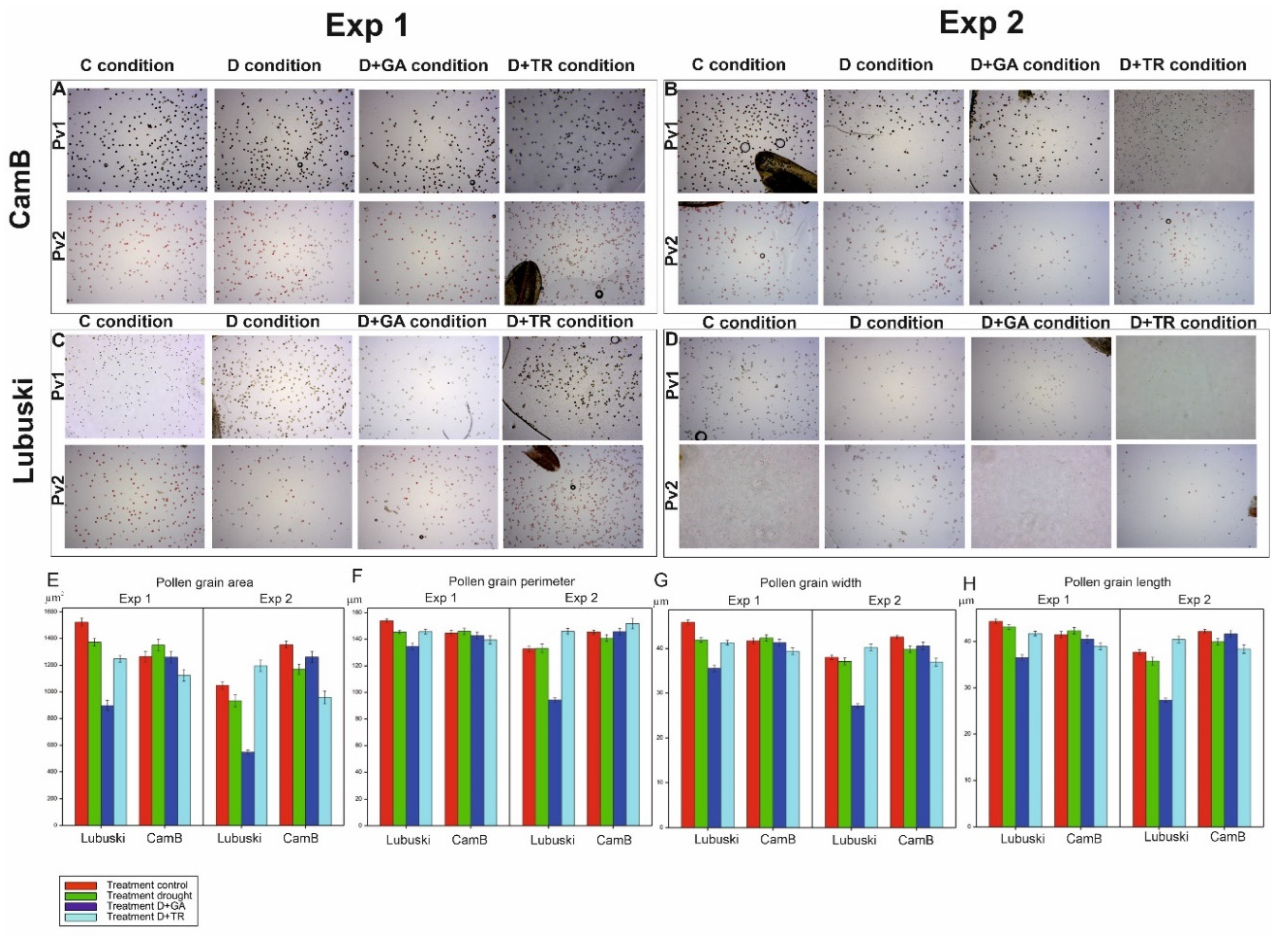
Figure 4.
Micrographs showing the pollen grains collected from the late-heading genotype grown under four different water conditions in Exp 1. Trinuclear microspores can be seen in the image taken from the tissues of Lubuski plant grown under C condition. In the vegetative cell, the nucleus with well-developed nucleolus and condensed cytoplasm can be observed (A). Binuclear, vacuolated microspores can be seen in the image taken from the tissues of Lubuski plant grown under D condition (B). Trinuclear microspores are visible in the image taken from the tissues of Lubuski plant grown under D+GA condition. The intensity of toluidine blue staining indicated chromatin condensation in the generative and vegetative nuclei (C). Trinuclear microspores can be observed in the image taken from the tissues of Lubuski plant grown under D+TR condition (D). VM — vacuolated microspores, E — epidermis, En — endothecium, Msp — microspore, and T — tapetum.
Figure 4.
Micrographs showing the pollen grains collected from the late-heading genotype grown under four different water conditions in Exp 1. Trinuclear microspores can be seen in the image taken from the tissues of Lubuski plant grown under C condition. In the vegetative cell, the nucleus with well-developed nucleolus and condensed cytoplasm can be observed (A). Binuclear, vacuolated microspores can be seen in the image taken from the tissues of Lubuski plant grown under D condition (B). Trinuclear microspores are visible in the image taken from the tissues of Lubuski plant grown under D+GA condition. The intensity of toluidine blue staining indicated chromatin condensation in the generative and vegetative nuclei (C). Trinuclear microspores can be observed in the image taken from the tissues of Lubuski plant grown under D+TR condition (D). VM — vacuolated microspores, E — epidermis, En — endothecium, Msp — microspore, and T — tapetum.
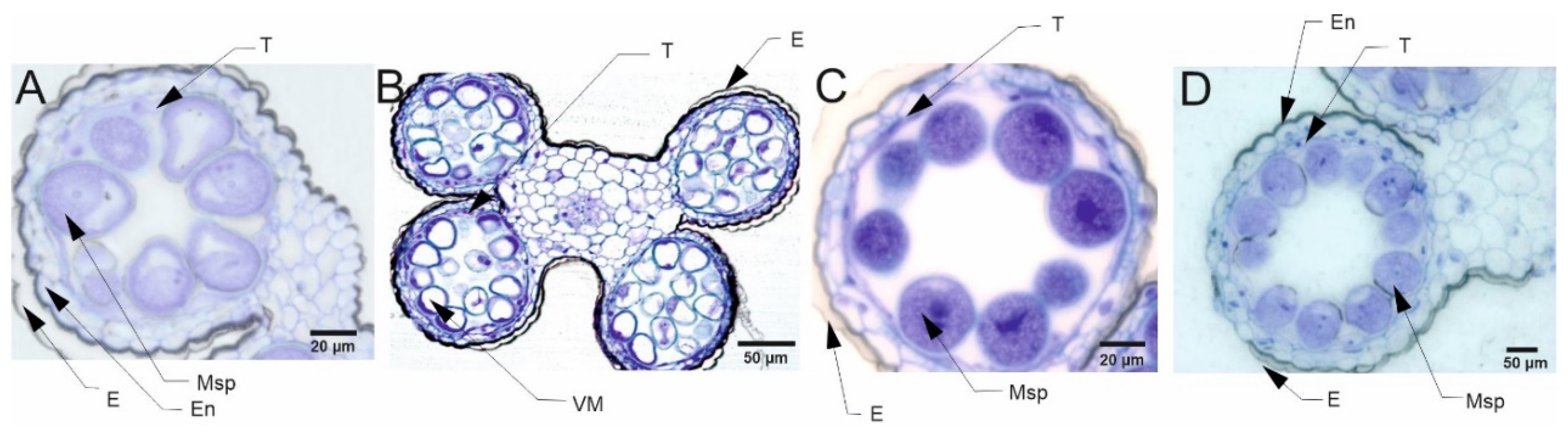
Figure 5.
Micrographs showing the pollen grains collected from the early-heading genotype grown under four different water conditions in Exp 1. Trinuclear, strongly vacuolated microspores can be seen in the image taken from the tissues of CamB plant grown under C condition (A). Trinuclear, partially vacuolated microspores are visible in the image taken from the tissues of CamB plant grown under D condition (B). Trinuclear microspores with weak development of vacuoles can be seen in the image taken from the tissues of CamB plant grown under D+GA condition (C). Trinuclear microspores are visible in the image taken from the tissues of CamB plant grown under D+TR condition (D). VM — vacuolated microspores, E — epidermis, En — endothecium, MI — middle layer, Msp — microspore, T — tapetum, and Sm — stomium.
Figure 5.
Micrographs showing the pollen grains collected from the early-heading genotype grown under four different water conditions in Exp 1. Trinuclear, strongly vacuolated microspores can be seen in the image taken from the tissues of CamB plant grown under C condition (A). Trinuclear, partially vacuolated microspores are visible in the image taken from the tissues of CamB plant grown under D condition (B). Trinuclear microspores with weak development of vacuoles can be seen in the image taken from the tissues of CamB plant grown under D+GA condition (C). Trinuclear microspores are visible in the image taken from the tissues of CamB plant grown under D+TR condition (D). VM — vacuolated microspores, E — epidermis, En — endothecium, MI — middle layer, Msp — microspore, T — tapetum, and Sm — stomium.
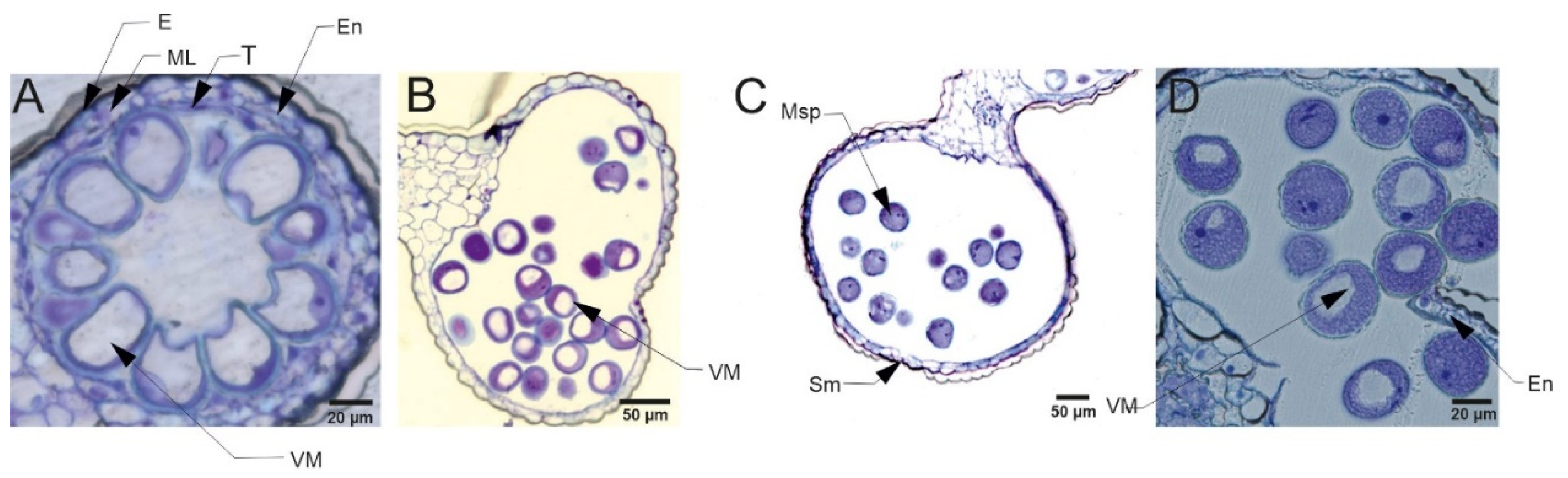
Figure 6.
Micrographs showing the pollen grains collected from the late-heading genotype grown under four different water conditions in Exp 2. Vacuolated microspores with well-developed middle layer can be seen in the image taken from the tissues of Lubuski plant grown under C condition (A). Vacuolated microspores can be seen in the image taken from the tissues of Lubuski plant grown under D condition (B). Trinuclear microspores are visible in the image taken from the tissues of Lubuski plant grown under D+GA condition (C). Binuclear microspores can be observed in the image taken from the tissues of Lubuski plant grown under D+TR condition (D). VM — vacuolated microspores, E — epidermis, En — endothecium, MI — middle layer, Msp — microspore, and T — tapetum.
Figure 6.
Micrographs showing the pollen grains collected from the late-heading genotype grown under four different water conditions in Exp 2. Vacuolated microspores with well-developed middle layer can be seen in the image taken from the tissues of Lubuski plant grown under C condition (A). Vacuolated microspores can be seen in the image taken from the tissues of Lubuski plant grown under D condition (B). Trinuclear microspores are visible in the image taken from the tissues of Lubuski plant grown under D+GA condition (C). Binuclear microspores can be observed in the image taken from the tissues of Lubuski plant grown under D+TR condition (D). VM — vacuolated microspores, E — epidermis, En — endothecium, MI — middle layer, Msp — microspore, and T — tapetum.
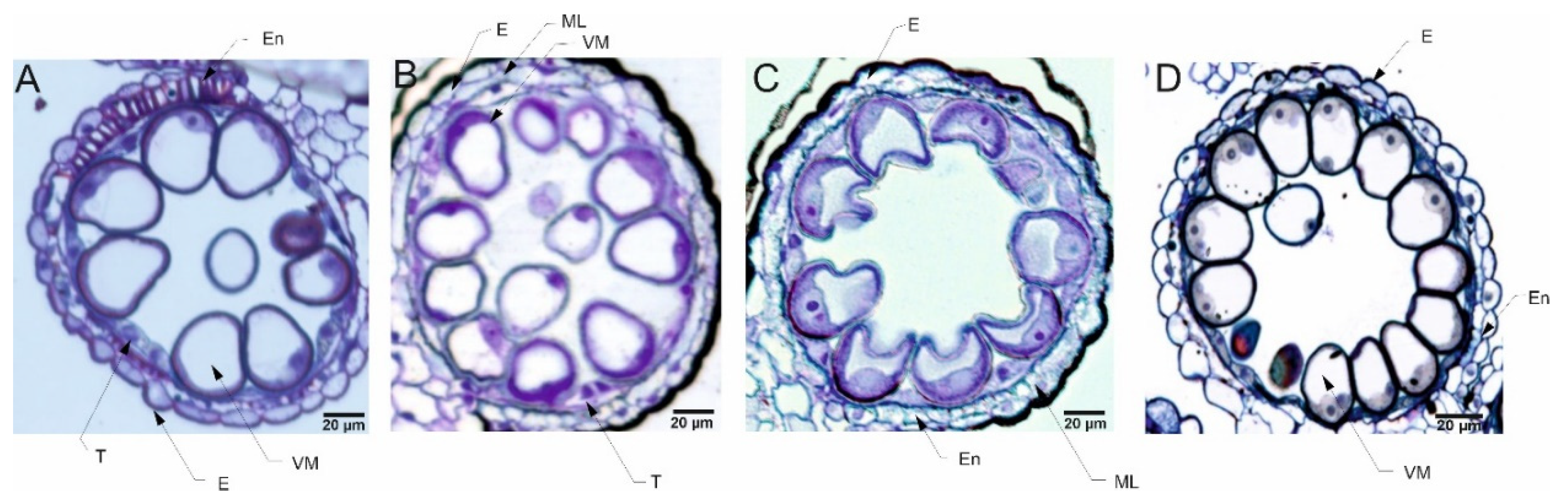
Figure 7.
Micrographs showing the pollen grains collected from the early-heading genotype grown under four different water conditions in Exp 2. Partially vacuolated microspores with well-developed middle layer can be seen in the image taken from the tissues of CamB plant grown under C condition (A). Strongly vacuolated microspores are visible in the image taken from the tissues of CamB plant grown under D condition (B). Trinuclear microspores are visible in the image taken from the tissues of CamB plant grown under D+GA condition. The intensity of toluidine blue staining indicated chromatin condensation in the generative and vegetative nuclei (C). Trinuclear microspores are visible in the image taken from the tissues of CamB plant grown under D+TR condition (D). VM — vacuolated microspores, E — epidermis, En — endothecium, MI — middle layer, Msp — microspore, and T — tapetum.
Figure 7.
Micrographs showing the pollen grains collected from the early-heading genotype grown under four different water conditions in Exp 2. Partially vacuolated microspores with well-developed middle layer can be seen in the image taken from the tissues of CamB plant grown under C condition (A). Strongly vacuolated microspores are visible in the image taken from the tissues of CamB plant grown under D condition (B). Trinuclear microspores are visible in the image taken from the tissues of CamB plant grown under D+GA condition. The intensity of toluidine blue staining indicated chromatin condensation in the generative and vegetative nuclei (C). Trinuclear microspores are visible in the image taken from the tissues of CamB plant grown under D+TR condition (D). VM — vacuolated microspores, E — epidermis, En — endothecium, MI — middle layer, Msp — microspore, and T — tapetum.
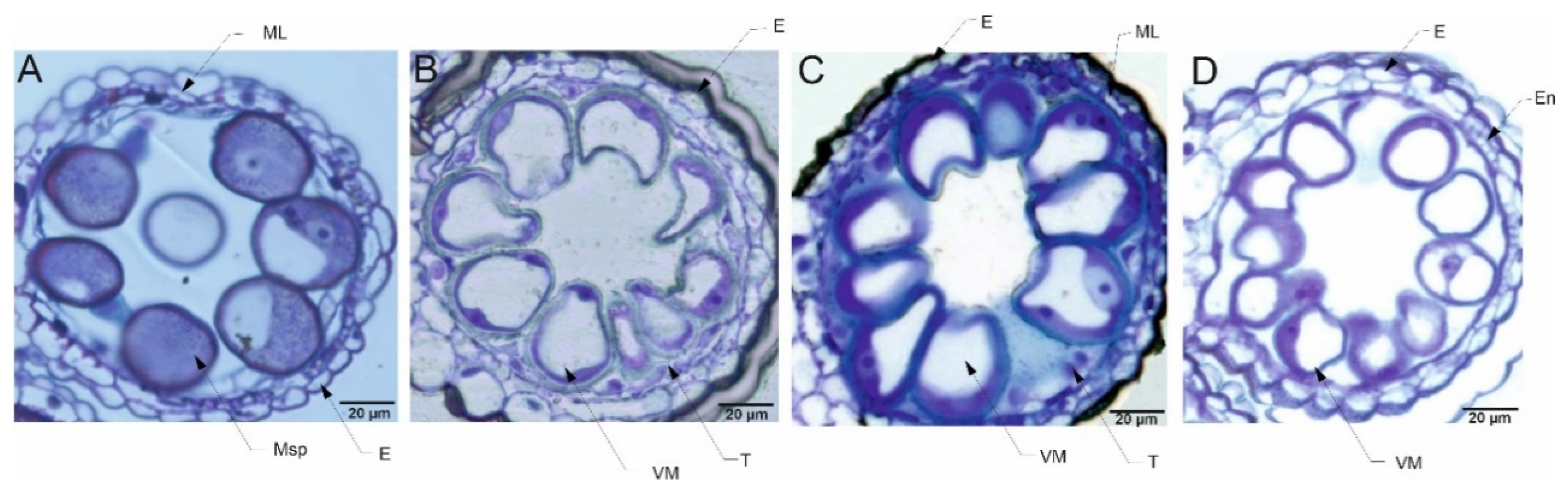
Figure 8.
TEM micrographs of pollen grains collected from Lubuski plants grown under control (A), drought (B), drought+GA (C), and drought+TR (D) conditions in Exp 1 and under control (E), drought (F), drought+GA (G), and drought+TR (H) conditions in Exp 2. ER — endoplasmic reticulum, M — mitochondria with swollen cristae (membrane) structures, A — amyloplast, Vi — vesicle, and V — vacuole. Bars = 2 µm for (A), (E); 1 µm for (G); 500 nm for (B), (F), (H); 200 nm for (C), (D).
Figure 8.
TEM micrographs of pollen grains collected from Lubuski plants grown under control (A), drought (B), drought+GA (C), and drought+TR (D) conditions in Exp 1 and under control (E), drought (F), drought+GA (G), and drought+TR (H) conditions in Exp 2. ER — endoplasmic reticulum, M — mitochondria with swollen cristae (membrane) structures, A — amyloplast, Vi — vesicle, and V — vacuole. Bars = 2 µm for (A), (E); 1 µm for (G); 500 nm for (B), (F), (H); 200 nm for (C), (D).
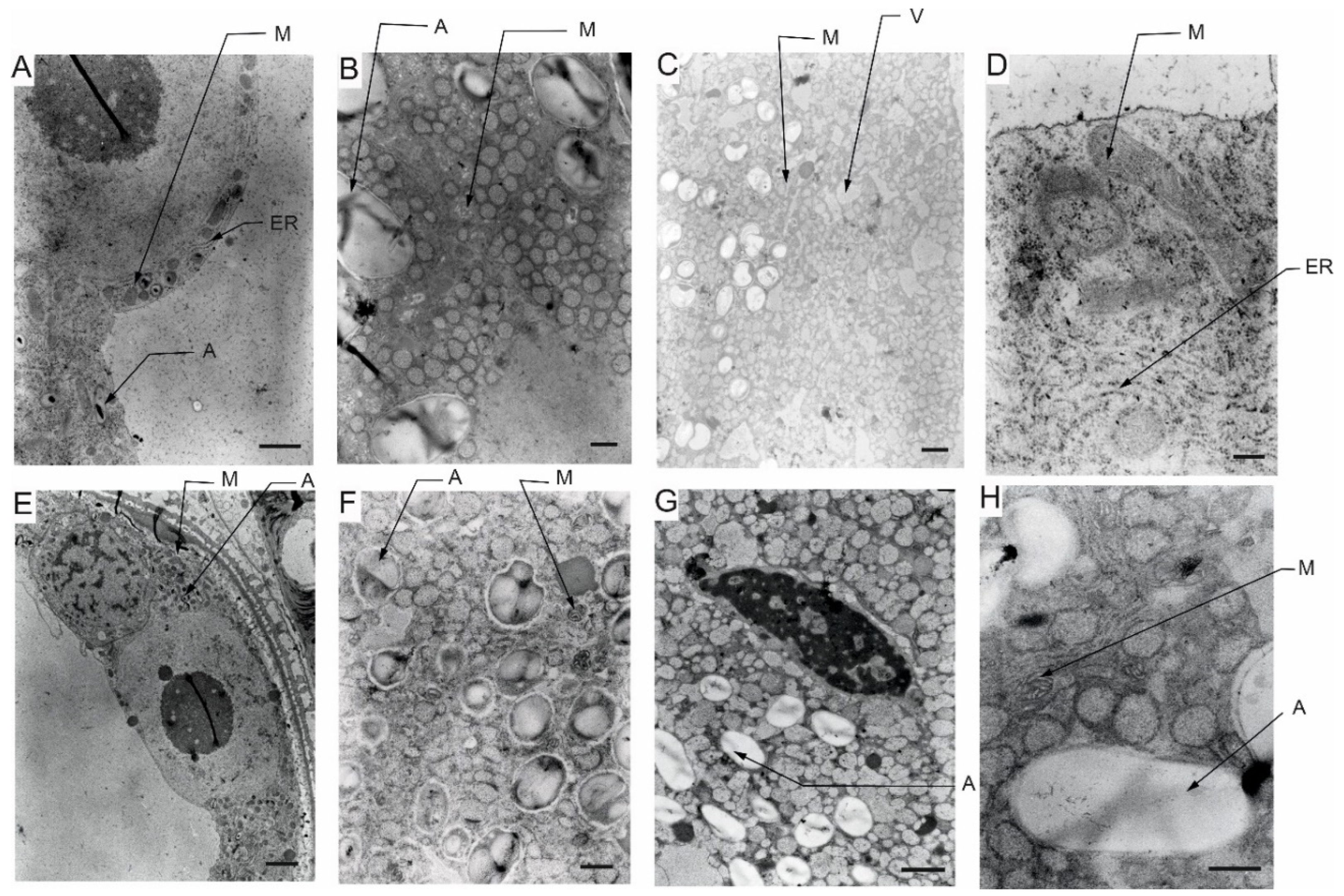
Figure 9.
TEM micrographs of pollen grains collected from CamB plants grown under control (A), drought (B), drought+GA (C), and drought+TR (D) conditions in Exp 1 and under control (E), drought (F), drought+GA (G), and drought+TR (H) conditions in Exp 2. ER — endoplasmic reticulum, M — mitochondria with swollen cristae (membrane) structures, A — amyloplast, Vi — vesicle, and V — vacuole. Bars = 2 µm for (A), (B), (E), (G); 1 µm for (F), (H); 500 nm for (C); 200 nm for (D). .
Figure 9.
TEM micrographs of pollen grains collected from CamB plants grown under control (A), drought (B), drought+GA (C), and drought+TR (D) conditions in Exp 1 and under control (E), drought (F), drought+GA (G), and drought+TR (H) conditions in Exp 2. ER — endoplasmic reticulum, M — mitochondria with swollen cristae (membrane) structures, A — amyloplast, Vi — vesicle, and V — vacuole. Bars = 2 µm for (A), (B), (E), (G); 1 µm for (F), (H); 500 nm for (C); 200 nm for (D). .
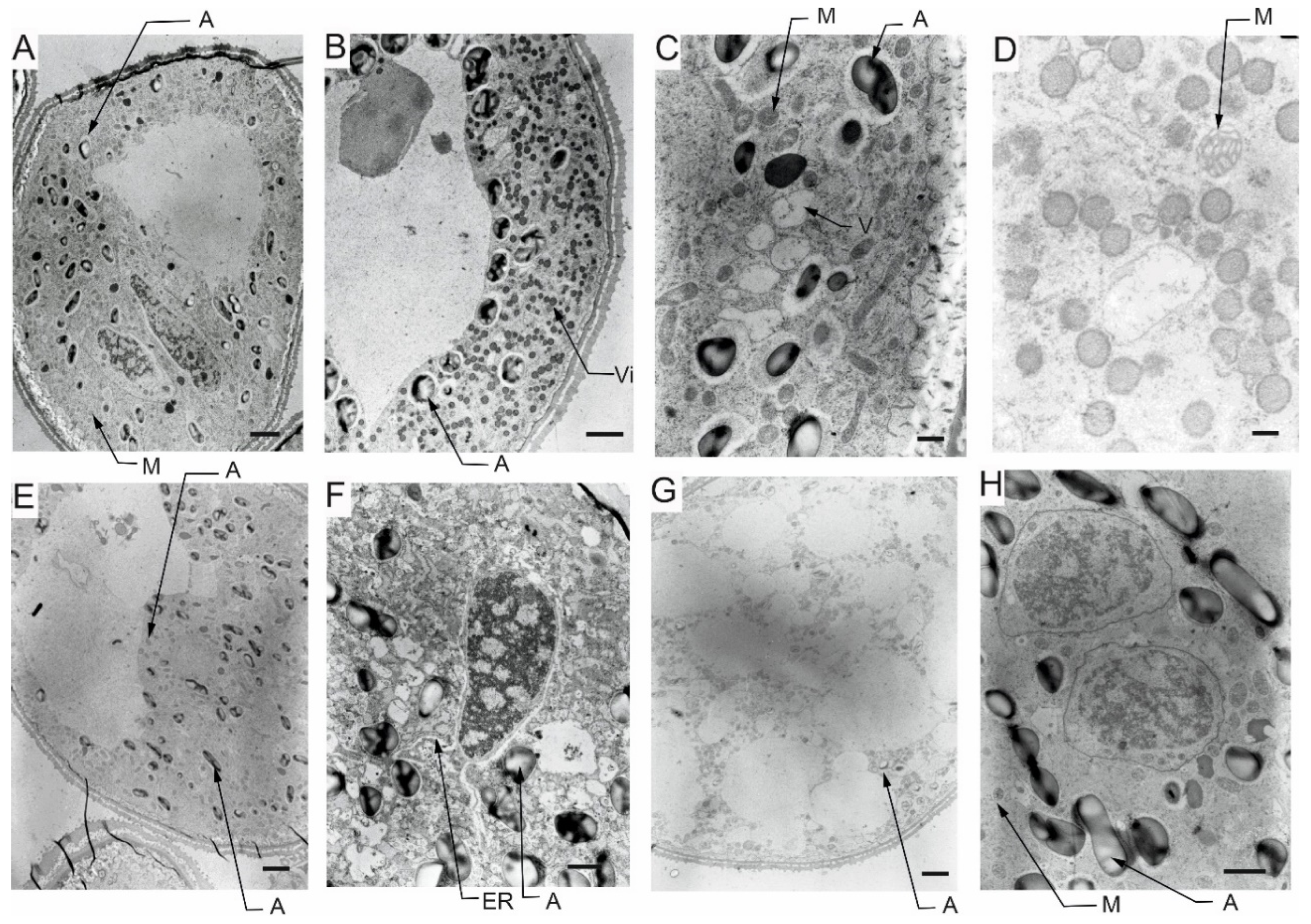
Figure 10.
Relative levels of expression (with standard errors) of the HvGAMYB gene in two different development points (1—LFE1 and 2—LFE4) in four different water regimes applied in Exp 1 (A) and Exp 2 (B).
Figure 10.
Relative levels of expression (with standard errors) of the HvGAMYB gene in two different development points (1—LFE1 and 2—LFE4) in four different water regimes applied in Exp 1 (A) and Exp 2 (B).
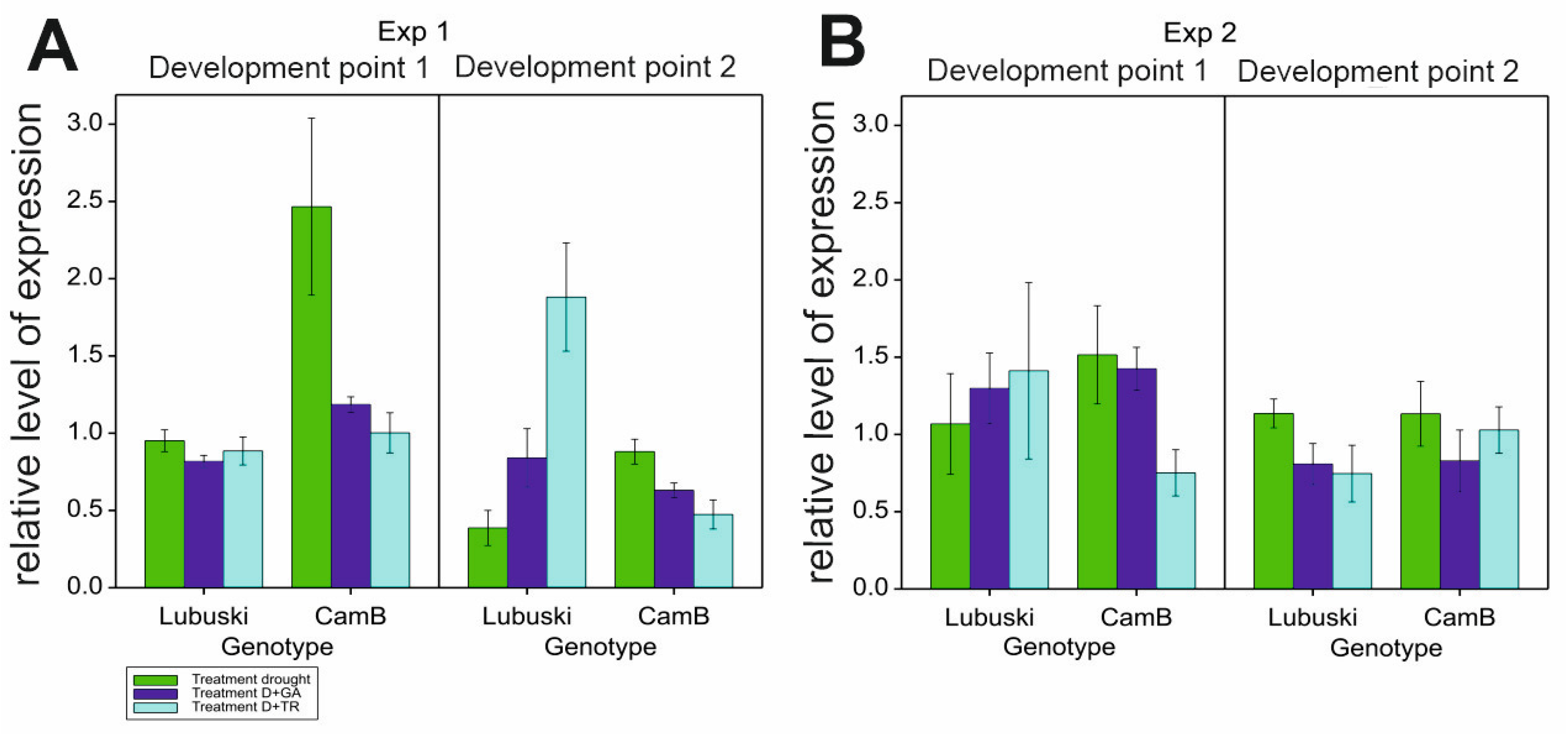

Table 1.
List of phenotypic traits with description, abbreviations and measured units.
| Trait (unit), (abbrev.) | Trait description |
| Total number of tillers, (Tn) | Number of tillers with fertile and non-fertile (without grains) spikes |
| Number of productive tillers, (PTn) | Number of tillers with fertile spikes |
| Length of main spike (cm), (LSm) | Length of main spike from 10 randomly selected spikes in a pot (without awns) |
| Number of spikelets per main spike, (NSSm) | Number of spikelets in spike of main stem-average e for 10 main spikes in a pot |
| Number of grains per main spike, (NGSm) | Number of grains collected from one spike of main stem - average for 10 main spikes in a pot |
| Weight of grains per main spike, (WGSm) | Weight of grain collected from one spike of the main stem - average for 10 main spikes in a pot |
| Length of lateral spike (cm), (LSl) | Length of spike from lateral stem - average for 10 lateral spikes in a pot (without awns) |
| Number of spikelets per lateral spike, (NSSl) | Number of spikelets per spike of lateral stem - average for 10 lateral spikes in a pot |
| Number of grains per lateral spike, (NGSl) | Number of grains collected from spike of lateral stem - average for 10 lateral spikes in a pot |
| Weight of grains per main spike, (WGSl) | Weight of grain collected from one spike of the lateral stem - average for 10 lateral spikes in a pot |
| Grain yield (g), (GY) | Average weight of grains collected from one plant, calculated as average of measurements of grain weight for 10 plants. |
| Thousand grain weight (g), (TGW) | Average weight of 1000 grains, calculated as average of 1000 * average weight of one grain for 20 spikes in a pot |
| Fertility of the main spike, (FSm) | NGSm/NSSm ratio |
| Fertility of the lateral spike, (FSl) | NGSl/NSSl ratio |
Table 2.
Chlorophyll fluorescence induction (OJIP) parameters calculated in the study (with abbreviations).
Table 2.
Chlorophyll fluorescence induction (OJIP) parameters calculated in the study (with abbreviations).
| Trait | Abbrev. |
| Quantify the PSII behavior were the absorbed energy flux | ABS_RC |
| Trapped energy flux | TRo_RC |
| Electron transport flux | Eto_RC |
| Dissipated energy flux | DIo_RC |
| Maximum quantum yield of primary photochemistry | Fv_Fm |
| Probability/efficiency that a trapped exciton moves an electron into the electron transport chain beyond QA | Ψ_o |
| Quantum yield of electron transport | Φ_Eo |
| Probability that the energy of an absorbed photon is dissipated as heat | Φ_Do |
| Performance index | Pi _Abs |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



