
Submitted:
18 April 2023
Posted:
19 April 2023
You are already at the latest version
Abstract
Grape production worldwide is increasingly threatened by grapevine trunk diseases (GTDs). No grapevine cultivar is known to be entirely resistant to GTDs, but susceptibility varies greatly. To quantify these differences, four Hungarian grape germplasm collections containing 305 different cultivars were surveyed to determine the ratios of GTDs based on symptom expression and mortality rate. The cultivars of monophyletic Vitis vinifera L. origin were amongst the most sensitive ones, and their sensitivity was significantly (p<0.01) higher than that of the interspecific (hybrid) cultivars assessed, which are defined by the presence of Vitis species other than V. vinifera (e.g. V. labrusca L., V. rupestris Scheele, V. amurensis Rupr.) in their pedigree. We conclude that ancestral diversity of the grape confers higher degrees of resilience against GTDs.
Keywords:
interspecific cultivars
; Vitis vinifera
; Vitis amurensis
; Vitis rupestris
; Vitis labrusca
; grape germplasm collection
; GTDs
1. Introduction
Grapevine trunk diseases (GTDs) are amongst the most important diseases of grapevines with estimated losses of 1.5 billions of USD worldwide while the average GTD incidences were reported to be between 10 % (Spain) to 22 % (Italy) in European vineyards [1,2,3,4]. Moreover, the increases in disease incidence has been recognized in several grape growing countries such as Spain, Italy, Canada [5,6,7,8]. GTD fungal pathogens colonize the woody part of the plant, producing different toxins and enzymes, and resulting leaf symptoms (tiger stripes) stunted growth, reduced quantity and quality of grape, and dieback of the plant [2]. GTDs are complex diseases, including esca, eutypa dieback, black foot, botryosphaeria and Petri diseases, and are affected by several biotic and abiotic factors [2,4,9,10,11,12]. More than 100 fungal species have been recognized as GTD pathogens, characterized by different taxonomic status, disease cycle, fungicide sensitivity and host range [13]. Moreover, infections usually do not manifest rapidly and can linger on for years. Factors and circumstances that turn the latent infection into an active one giving rise to mild (e.g., foliar symptoms) or serious symptoms (partial or whole plant dieback) are not fully understood. Wounds, environmental stress (frost, drought, flood), and increased age of the vineyards appear to correlate with increased disease incidence of GTDs [4,14,15,16,17]. Chronic symptom expression does not necessarily lead to significant yield or quality loss of the fruit, or plant loss within a few years [4,18]; conversely, apoplexy of the trunk leads to plant loss and results in irreversible economic loss in the plantation. Replenishing vineyards with young, healthy vines is challenging and often unsuccessful.
Tolerant plant cultivars are widely in use as they are one of the most effective means to control plant disease, providing economic and environmentally friendly plant protection technology while reducing pesticide usage and dependency [19]. Disease-resistant cultivars would also provide solutions when effective protection by chemical pesticides is not available, like in the case of GTDs [20,21,22].
Due to the susceptibility of traditional European grape varieties to different pathogens, an interspecific hybrid breeding program was started in France in the early fourties of the 19th century , by crossing Vitis vinifera varieties from France with American species, which resulted in more resistant, high-quality hybrids that exhibit partial resistance toward the fungal pathogens [23]. The hybrid offspring were subsequently used in the resistance breeding programs in Hungary as 'Seibel' and 'Seyve-Villard' varieties [24]. However, with the propagating plants, the phylloxera (Daktulosphaira vitifoliae Fitch) insect pest was also introduced, which resulted in dramatic loss of plants in the European vineyards. It also has become common practice to graft American rootstock, resistant to phylloxera, to preserve susceptible cultivated European varieties, and grape breeding programs were initiated to control phylloxera, powdery mildew (Erysiphe necator Schwein.) and downy mildew (Plasmopara viticola (Berk. et Curt.) Berl. et De Toni) [23,25,26,27,28].
Vitis amurensis Rupr., native to China, has several beneficial properties, such as cold resistance and resistance against several phytopathogens causing diseases, like grape crown gall (Allorhizobium vitis), white rot (Coniella diplodiella (Speg.) Petr. et Syd.), downy mildew and anthracnose of grapes (Elsinoe ampelina Shear). Therefore, it is often used as rootstock or in breeding interspecific hybrids [29,30,31,32,33,34,35,36]. Introduction of American and Asian grape species to the breeding programs increases genetic diversity, and compensates the bottleneck effect (when the size of a population is severely reduced), that developed historically as the consequence of the domestication of V. vinifera [37,38].
There are no V. vinifera cultivars known to be completely resistant to GTD pathogens, however considerable differences in sensitivity have been recognized during in planta tests and in field surveys (Table 1). There were differences observed between the tolerance to different GTD pathogen fungi in one cultivar, which may due to the various climate conditions and/or grape-producing technologies. In the case of eutypa dieback, Dubos [39] categorized Aligote, Grolleau, Merlot, Semillion and Sylvaner cultivars as resistant, and later Carter [40] reported possible resistance against Eutypa lata (Pers.) Tul. & C. Tul in some French cultivars. Borgo et al. [41] and Murolo-Romanazzi [42] classified the degree of GTD expression for six and 86 varieties, distinguishing between red and white grape varieties. Sosnowski et al. [43] ranked 118 varieties based on plant death and foliar symptoms. These and other studies have shown that, among internationally recognized and cultivated varieties, Cabernet Sauvignon, Cabernet Franc and Sauvignon Blanc are particularly susceptible to GTDs, while the Merlot is much more resilient.
Both GTD chronic symptom expression and apoplexy combined with subsequent loss of plants were monitored in four Hungarian grape germplasm collections containing a total of 305 different cultivars. Disease incidence (DI) was calculated to compare (i) the degree of GTD sensitivity of the most important international and national grape cultivars (ii) the severity of GTD symptoms in cultivars with monophyletic V. vinifera origin and interspecific (hybrid) cultivars with various American or Asian Vitis species in their pedigree. These data may provide important information for extended and future grape breeding programmes.
2. Results
Four grapevine germplasm collections with 537 cultivars were surveyed. GTD symptoms were categorized as new symptoms during the annual vegetative period (leaf stripes with white or brown rot, and dieback) (Figure 1 a-d) or as dead and missing (removed) plants from previous dieback events in past years (Figure 1 e-f).
The total disease incidence (DI %) was over 25 % in each of the survey sites (Table 2), therefore, the conditions for a meaningful survey of symptom expression rates were considered adequate for further analysis. The average ratio of dead plants and total disease incidence (i.e., all symptoms) was similar in each germplasm collection. Altogether, these results, with previous records of dieback symptoms of currently dead and removed (dead) plants validated the connection between missing plants and previous dieback.
The total DI of the most important cultivars with only V. vinifera ancestors were compared (Figure 2). Sauvignon Blanc and Cabernet Sauvignon were the most susceptible cultivars, while Merlot and Syrah were the less susceptible ones (Figure 2, Table 1). There were both white and red grapes among the most and the less sensitive cultivars within the analyzed genuine V. vinifera cultivars. The susceptibility of Furmint, one of the most important Hungarian white cultivars was similar to that of Veltliner Gruen and Muscat Lunel, while another indigenous white cultivar, Juhfark, was less susceptible, more similar to that of Blauburger and Pinot Blanc. The indigenous table grape, Csaba Gyoengye, was less susceptible than Furmint, showing similar DI to those of Welschriesling, Cabernet Franc and Muscat Ottonel. Blaufraenkish, a grapevine variety with regional importance was amongst the less susceptible cultivars, like e.g., Pinot Blanc and Pinot Noir.
Severity of disease expression categories were defined to separate cultivars. When a cultivar has a tendency to not demonstrate GTD symptoms in situ, it is defined as unsusceptible. When only annual developed (usually mild) GTD symptoms are displayed, the cultivar is listed as resilient. Sensitive cultivars demonstrated tendency of developing dieback symptoms eventually resulting in plant loss in parallel with other GTD symptoms in other individuals, while exclusively plant loss of infected specimen was detected in vulnerable cultivars that are highly sensitive. The majority of the cultivars with only V. vinifera ancestors in their pedigree were categorized as highly sensitive or sensitive to GTDs with exclusively plant loss or high plant demise concurrent with non-lethal symptoms (Figure 3). The level of resistence to GTD pathogens was generally better or much better in the case of interspecific hybrid Vitis cultivars, with a considerably higher ratio of unsusceptible or resilient cultivars, than encountered amongst monophyletic V. vinifera ones (Figure 3).
The tendency of the cultivars with different origins for plant loss, was compared in binomial test. The ratio of the monophyletic V. vinifera cultivars was lower in the less sensitive groups (unsusceptible and resilient), than expected based that of all tested cultivars (Figure 4a). This indicates, that monophyletic V. vinifera cultivars have a higher tendency to display serious GTD symptoms including plant loss, than the average of all examined cultivars (overall samples). On the contrary, the ratio of cultivars without plant loss (less sensitive groups) was significantly higher for the group of the interspecific hybrids. Similarly, when the hybrids with American (V. labrusca, V. riparia or V. rupestris) or Asian (V. amurensis) ancestors were split and compared separately, the ratio of the cultivars in both groups were higher in the less susceptible categories compared to all cultivars studied (Figure 4b).
The susceptibility of the cultivars with different species ancestry (i.e., exclusively V. vinifera or interspecific hybrids) was compared regarding the cultivar specimen mortality from GTDs as part of the total GTD disease incidence (i.e., all symptoms). Plant death as a consequence of GTD expression was more likely in cultivars with monophyletic V. vinifera origin, than in the interspecific Vitis cultivars. Separating the group of the interspecific cultivars into cultivars with Asian and American origins, the ratio of dead plants within the displayed GTD symptoms was meaningfully lower exclusively for cultivars with V. amurensis ancestry , than the ones with monophyletic V. vinifera cultivars (Figure 5). Thus, the calculated difference was not significant for the group of cultivars with V. labrusca, V. riparia or V. rupestris (American species) in their pedigree.
3. Discussion
There are differences in sensitivity to GTDs displayed by the V. vinifera cultivars, however, no completely resistant has been identified. The physiological and genetic background of these differences in sensitivity or resistance against GTD-causing pathogens is not understood [53,54]. In accordance with previous results, Sauvignon Blanc and Cabernet Sauvignon showed the highest DI in the surveyed Hungarian germplasm collections, all four with their own climate and soil characteristics, while Furmint, Chardonnay and Cabernet Franc were found less GTD susceptible [12,42,43,46,55]. Blaufraenkisch (also referred to as Limberger), again confirmed by our current results, constistently one of the lowest DI [42,43,46,55], while Merlot and Pinot Noir usually were also found less susceptible to most of GTDs in general [12,42,46,55,56].
Comparing the sensitivity of different grapevine cultivars to esca, significant differences were found between those with red and those with white berries, and their respective xylem vessel diameter and density [46]. The average vessel diameter of the white cultivars was larger with higher densities, compared to the red grapevines. A similar trend was observed for overall disease incidence, where the mean disease incidence was higher for white-berry cultivars than for red-berry cultivars. Foliar symptom symptoms are hypothesized to result from fungal toxins translocated to leaves from primary infection sites [53,57,58]. Higher rates of leaf symptoms were explained by the larger vessel diameters, since it provides space for more intensive xylem cavitation, which can assist toxin translocation to the green plant parts [46]. Moreover, Pouzoulet et al. [59] stated, that the esca pathogens may escape compartmentalization more efficiently when the vessels are wider, and the more gel and tyloses in the vessels, the more substrate is provided for wood pathogens [59].
No GTDs symptom expression was detected in the Hungarian germplasm collections on the extant Merlot cultivars, whose outstanding tolerance was reported in several previous studies in other countries [21,39,42,44,50,60,61]. The lignin content of Merlot was found to be significantly higher than in Cabernet Sauvignon, a cultivar to be considerably more susceptible to GTDs [41,43,46,50]. Other cultivars identified as less sensitive to GTD had in general smaller vessel diameter and higher lignin content than the most sensitive grapevine varieties [62,63]. The results of Rolshausen et al. [62] highlighted the potential importance of lignin in the E. lata-grapevine interaction. The costumary defence response of grapevines to infection is compartmentalization, where the plant attempts to contain the invading agent by depositing suberin and lignin which impedes the spread of the pathogens throughout the xylem. A higher lignin content was detected in the infected grape tissues, which indicates that lignin deposition is initiated in response to the fungal infection [62].
GTDs are complex diseases, resulting serious economic loss by reduced grape productivity, and characterized by remarkable differences in the disease severity and manifestation [2]. Infection with GTD fungal pathogens may result in latency, accidental or repeated annual disease expression, and serious partial or whole plant dieback [64]. The most serious disease symptom is plant loss resulting irreversible economic damage. Previously, only foliar or chronic and dead cordon or apoplexic (partial and whole plant) individual disease expressions were differentiated amongst GTD symptoms [43,46,55]. This traditional categorization or subsequent merging different symptom manifestations and calculating disease incidence indicates only the susceptibility of a cultivar, and does not take into account the severity of the infection and the plant’s responses. Cultivars that are able to survive infection for a longer period of time - specimens of which are more likely to express the milder foliar symptoms and partial dieback rather than whole plant apoplexy and death - are considered more resistant to the fungal GTD pathogens in our present survey and analysis.
The survey and analysis of four Hungarian germplasm collections concluded that the interspecific hybrid cultivars, in particular the ones with Asian V. amurensis ancestry are generally less susceptible to GTDs, expressing no or milder symptoms, than monophyletic cultivars with only V. vinifera ancestors. In these hybrid cultivars with some level of East-Asian ancestry, infection by GTD fungal pathogens resulted in less plant losses, which is the most serious, and irreversible consequence of GTD infection. One of the possible backgrounds of this lower sensitivity (or higher resistance) may concur with the xylem vessel diameter, as V. amurensis had the smallest vessel diameter amongst different grape species [65,66]. By contrast, the vessel diameter of the American species V. labrusca was reported to be rather large [65]. In a more recent study, there was no substantial difference in xylem vessel diameter recorded between V. vinifera and American interspecific hybrid called Noiret, with V. labrusca ancestry [63].
Since most of the GTD pathogens are wound-colonizing fungi, frost cracks of the wood parts of the plant could facilitate the prevalence of the GTD disease complex in grapevine [67,68]. Compared to V. vinifera and V. labrusca species, V. amurensis is extraordinarily cold-resistant and can survive long and cold winters as a result of its relatively low respiratory intensity, a lower level of active metabolism and a longer dormancy period [32]. V. amurensis is cultivated as a cold-resistant grape in the colder regions of China [32,69,70,71]. Wang et al. [72] identified 17 genes possibly involved in this increased cold hardiness. Accumulation of several amino acids (valine, isoleucine and proline) was reported to be higher in V. amurensis than in V. vinifera cultivars, the level of which was subject to abiotic stress [73]. This property together with the accumulation of other bioactive compounds (polyphenols, tannin, and the stilbene phytoalexin resveratrol) can protect plants from long-term cold damage [32,74].
The induction of stilbene biosynthesis was found correlated with basal immunity against the downy mildew and eutypa dieback [48,75]. American Vitis species are also employed in breeding more cold-hardy cultivars [76]. Increased stilbene biosynthesis has relevance in increased resistance to different fungal diseases [77], and may have importance in GTD tolerance, as grapevine rootstock transformed with grapevine stilbene synthase gene expressed from a pathogen-inducible promoter showed increased resistance against E. lata [48].
V. amurensis is not only cold-tolerant, but also resistant to white rot, grape anthracnose and grape bitter rot (Greeneria uvicola (Berk. & M.A. Curtis) Punith) fungal diseases, and has a high resistance to downy mildew, caused by the Oomycete P. viticola [29,31,32,33,74,78,79,80]. The resistance of grapevine against the bacterial trunk pathogen A. vitis was introgressed from V. amurensis upon interspecific breeding [30]. Hybrids with V. amurensis ancestry were unambiguously less sensitive to GTD pathogens in our survey, illustrated by the considerably higher ratio of resilient and tolerant hybrid cultivars to Botryosphaeria dieback (BD) and esca diseases.
Pretorius and Høj [81] assumed that the product of a single gene or its pyramid (stacking multiple genes into a single genotype to combine desirable traits) is effective only against a narrowly related group of pathogens within the GTD complex. These authors differentiated the tolerance toward various GTD pathogens in numerous monophyletic American Vitis cultivars and hybrids. The resistance loci Rda1 and Rda2 originating from Vitis cinerea (Engelm.) Engelm. ex Millard B9, a native American grape, and the interspecific Horizon cultivar, respectively, largely prevented the development of Phomopsis dieback symptoms [82]. Concord, an interspecific cultivar with parental varieties Catawba and V. labrusca showed a reduced sensitivity to Neofusicoccum parvum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips, in inoculation assays. On the other hand, the American Vitis spp. were found to be more susceptible against Eutypa dieback, than V. vinifera [44]. Co-evolution of V. vinifera and E. lata in a natural habitat, could have increased the resistance of the plants prior to domestication [44].
One of the main goals of breeding programs nowadays is to pyramid extant, independent biotic and abiotic resistance genes from different lineages of American or Asian grapes and to attain additive accumulation of broad resistance against or tolerance to phytopathogens into one parent which can be crossed with European V. vinifera [84]. The domestication bottleneck effect, the result of thousands of years of vegetative propagation without meiosis and recombination, and the continuous incrossings of the high-quality cultivar resulted in low genetic diversity across domesticated V. vinifera grapes [37,38]. Engaging American and Asian Vitis species in breeding has the potential to enhance biotic and abiotic vine stress tolerance lost over the course of domestication [29,30,31,32,33,34,35,36,69,70,71], which is relevant to GTD symptom expression and disease severity in grape cultivars.
4. Materials and Methods
4.1. Survey sites and cultivars
The survey was conducted in 2022 involving four Hungarian germplasm collections (Figure 6) containing high number of cultivars with worldwide, Central-European, or Carpathian basin significance, and valuable parental lines for further breeding. The climatic and edaphic conditions differ considerably at the four locations, despite their geographical closeness (ranging from 60 to 330 km in distance). Pallag (University of Debrecen, Institutes for Agricultural Research and Educational Farm, Horticultural Experimental Plant of Pallag) and Kecskemét (Hungarian University of Agriculture and Life Sciences, Research Institute for Viticulture and Oenology) are in the eastern part of Hungary which has a continental climate with a relatively low annual precipitation (500-700 mm) [85]. These lowland sites in the Carpathian basin were established on phylloxera immune sandy soils, thus the plants growing at these locations are not grafted (Pallag) or in part growing on their own root (Kecskemét) [86].
Badacsonytomaj (Hungarian University of Agriculture and Life Sciences, Research Institute for Viticulture and Oenology) and Pécs (University of Pécs, Research Institute for Viticulture and Oenology) are in occidental part of the country, where the influence of the westerly winds associated with the more moderate oceanic climate is more pronounced. Both of these sites have mountain slope relief with terrace cultivation and a sub-Mediterranean climate with annual precipitation between 600-800 mm [85,88]. The soil type in Badacsonytomaj is volcanic erubase and eroded loess slope sediment and the region is heavily affected by the humidifying and moderating effect of the water body of the Lake Balaton [89]. The soil type in Pécs is Brown earth (Ramann’s brown forest soil) overlying carbonate-rich red sandstone.
The germplasm collections are considered to be free from bacterial phytopathogens A. vitis, and Rhizobium radiobacter. Vineyard parts potentially affected by Flavescence dorée (Ca. Phytoplasma vitis) were consistently excluded from our survey. BD and esca symptoms were predominant at the surveyed sites but Eutypa-like symptoms [13] were encountered in few instances. The GTDs were visually diagnosed by the typical tiger-strip foliar symptoms (Figure 1a,d) while white and/or brown rot was detected on cross sections or debarked woody parts (Figure 1c) of the plants. The ensemble of BD, esca and Eutypa-like symptoms were counted as GTD symptoms. The new apoplectic symptoms (dead young shoots with leaves, Figure 1b) were considered as annual GTD symptoms. If there were no fresh sprouts in the vine specimen, the plant was considered as dead (Figure 1 1e-f). All evaluated cultivars were surveyed in over 10-years-old plants, therefore the chronic/milder (non-lethal) symptoms were evaluable [90,91].

Table 6.
Characteristics of germplasm collection locations.
| Badacsonytomaj | Kecskemét | Pallag | Pécs | |
|---|---|---|---|---|
| Soil | erubase soil | sand | acidic sand | Brown earth |
| Relief | mountain slope (top-valley row direction, terrace cultivation) | lowland | lowland | mountain slope (terrace cultivation) |
| Cultivation type | grafted | own rooted | own rooted | grafted |
| Climate | submediterranean with dry, warm summer | continental | continental | submediterranean with dry, warm summer |
| Relative climate sector1 | IIIc | Ib | Ia | IIIb |
| Average temperature fluctuation (°C) | 21-22 | 23-24.5 | 23-24 | 21-22 |
| Annual precipitation (mm) | 600-800 | 500-550 | 550-700 | 600-800 |
| Annual sunshine duration (h) | 1950-2050 | 2000-2150 | 1900-2050 | 2000-2100 |
1 Relative climate sector as taken from [85].
Many of the surveyed cultivars had non-V. vinifera ancestry. The different Vitis spp. in the pedigree of a cultivar was certified based on the data of the Vitis International Variety Catalogue (VIVC) [51]. The cultivars were grouped for the further analysis based on ancestry from different Vitis sp. (Table 7).
4.2. Data analysis
4.2.1. Susceptibility analysis
The total disease incidence (DI% - the ratio of plants showed fresh leaf symptoms and dieback, and whole plant apoplexy in previous years) was evaluated in the the cultivars of the surveyed germplasm collections. Since the overall disease incidence was over 25 % in every site and the spatial distribution of the symptom expressing plants was homogenous in all the vineyards, similar probabilities of infection were assumed for each cultivar. Given these conditions, the same the cultivars in the different surveyed sites could be considered as replicates in statistical analysis.
4.2.2. Sensitivity categories and analysis
The cultivars were categorized based on a new method to determine the disease expression severity (i.e., the severity of visible symptoms). Four categories were established to differentiate between (1) no symptom expression, (2) exclusively new (annual) symptoms, (3) both new symptoms and previous dieback resulting plant loss, and (4) exclusively previous dieback events all resulting in plant loss.
Four GTD sensitivity groups were created to catagorize the studied cultivars, based on the type (annual foliar symptoms and dieback, or apoplexy), and the frequency of the different symptoms. Highly sensitive (HS), where all symptomatic plants of the cultivar are dead; sensitive (S), where both dead plant (resulting from apoplexy of the trunk), and fresh GTD leaf and dieback symptoms are detected. The cultivar was considered resilient (R), if only foliar symptoms were present, while neither apoplexy nor annual GTD leaf and dieback symptoms were detected in unsusceptible (U) cultivars (Table 8).
To reveal potential differences in pathogen sensitivity amongst differences in the ancestry groups, the four original groups were re-appreciated, where the two more sensitive (HS and S) and the two less sensitive (R and U) categories were merged. The ratio of the lineage groups within each of these two re-defined sensitivity categories was compared to the theoretically expected distributions with the binominal test.
The tendency of GTD to kill the host plant was determined in parallel by calculating the proportion of individual plant losses within the total disease incidence of the lineage groups and compare the lineage groups in pairs. (1) Monophyletic European V. vinifera (Vv) cultivars against the interspecific (I) ones and hybrids with American (V. rupestris, V. riparia, V. labrusca - Ao) and Asian (V. amurensis – Va) species co-origin.
4.3. Statistical analysis and software background
The data sets did not fulfill the assumptions of parametric tests (i.e., normality and homogeneity of variances), that were analyzed with Q-Q plots and Levene test. During the analysis nonparametric Kruskal-Wallis test was used for comparison, which was backed up with Mann-Whitney U test as pairwise comparison with Statsoft Statistica 10 software.
The ratio of the sensitivity groups in different ancestral groups was compared with the binominal test executed using the online calculator of Stat Trek [92]. The Sankey diagram was generated by the Sankeymatic online diagram builder (https://sankeymatic.com).
5. Conclusions
Regarding the order in V. vinifera cultivar susceptibility based on total disease incidence, earlier data from the literature in other grape producing countries were confirmed and the main cultivars of the Carpathian basin were inserted in this ranking, where Juhfark proved to be more tolerant and Furmint more susceptible. Merlot did not show GTD symptoms in any of the Hungarian germplasm collections.
Interspecific Vitis cultivars had a lower tendency to plant loss following infection with GTD fungal pathogens Hybrid varieties with Asian V. amurensis ancestry have outstanding tolerance in our experimental set of more, than 300 cultivars. Engaging American and Asian Vitis species in breeding programs to enhance tolerance and resistence against GTDs has a great potential.
Author Contributions
Conceptualization, Erzsébet Sándor; Data curation, András Csótó; Funding acquisition, Levente Karaffa; Investigation, András Csótó, Nóra Laurinyecz, Zóra Annamária Nagy, Csaba Németh, Erzsébet Krisztina Németh, Anna Csikász-Krizsics, Nándor Rakonczás and Erzsébet Sándor; Methodology, András Csótó, Antal Nagy and Erzsébet Sándor; Project administration, András Csótó; Resources, Erzsébet Fekete and Levente Karaffa; Supervision, Erzsébet Sándor; Visualization, András Csótó, Antal Nagy and Florence Fontaine; Writing – original draft, András Csótó, Antal Nagy, Nóra Laurinyecz, Zóra Annamária Nagy, Csaba Németh, Erzsébet Krisztina Németh, Anna Csikász-Krizsics, Nándor Rakonczás, Florence Fontaine, Erzsébet Fekete, Michel Flipphi, Levente Karaffa and Erzsébet Sándor; Writing – review & editing, András Csótó, Antal Nagy, Florence Fontaine, Michel Flipphi, Levente Karaffa and Erzsébet Sándor.
Funding
This research was supported by the Hungarian National Research, Development, and Innovation Fund (grants: NN 128867 to L.K. and K 138489 to E.F.).
Data Availability Statement
The datasets used in the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hofstetter, V.; Buyck, B.; Croll, D.; Viret, O.; Couloux, A.; Gindro, K. What if esca disease of grapevine were not a fungal disease? Fungal Divers. 2012, 54, 51–67. [Google Scholar] [CrossRef]
- De la Fuente, M.; Gramaje, D.; Armengol, J.; Fontaine, F.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.-F. ;. Grapevine Trunk Diseases. A Review; OIV Publications: Paris, France, 2016; ISBN 979-10-91799-60-7. https://hal.science/hal-01604038/. (accessed on 28 03 2023); ISBN 979-10-91799-60-7. [Google Scholar]
- Pollastro, S.; Dongiovanni, C.; Abbatecola, A.; Faretra, F. Observations on the fungi associated with esca and on spatial distribution of esca symptomatic plants in Apulian (Italy) vineyards. Phytopathol. Mediterr. 2000, 39, 206 210. Available online: http://www.jstor.org/stable/26456547 accessed on 28 March 2023.
- Kovács, C.; Balling, P.; Bihari, Z.; Nagy, A.; Sándor, E. Incidence of grapevine trunk diseases is influenced by soil, topology and vineyard age, but not by Diplodia seriata infection rate in the Tokaj Wine Region, Hungary. Phytoparasitica 2017, 45, 21–32. [Google Scholar] [CrossRef]
- Martín, M.T.; Cobos, R. Identification of fungi associated with grapevine decline in Castilla y León (Spain). Phytopathol. Mediterr. 2007, 46, 18–25, http://www.jstor.org/stable/26463267. [Google Scholar]
- Úrbez-Torres, J.R.; Haag, P.; Bowen, P.A.; O’Gorman, D.T. Grapevine Trunk Diseases in British Columbia: Incidence and characterization of the fungal pathogens associated with esca and Petri diseases of grapevine. Plant Dis. 2014, 98, 469 482. [Google Scholar] [CrossRef] [PubMed]
- Surico, G.; Marchi, G.; Braccini, P.; Mugnai, L. Epidemiology of esca in some vineyards in Tuscany (Italy). Phytopathol. Mediterr. 2000, 39, 190–205. [Google Scholar] [CrossRef]
- Calzarano, F.; Di Marco, Wood, S. Discoloration and decay in grapevines with esca proper and their relationship with foliar symptoms. Phytopathol. Mediterr. 2007, 46, 96–101. [Google Scholar] [CrossRef]
- Lehoczky, J. Black dead arm disease of grapevine caused by Bortyosphaeria stevensii infection. Acta Phytopathol. Hun. 1974, 9, 319–327. [Google Scholar]
- Marchi, G.; Peduto, F.; Mugnai, L.; Di Marco, S.; Calzarano, F.; Surico, G. Some observations on the relationship of manifest and hidden esca to rainfall. Phytopathol. Mediterr. 2006, 45, 117–126, http://www.jstor.org/stable/26463242. (accessed on 28 03 2023). [Google Scholar] [CrossRef]
- Lecomte, P.; Darrieutort, G.; Dewasme, C.; Blancard, D.; Goutouly, J.P.; Rey, P.; Guérin-Dubrana, L. Impact of biotic and abiotic factors on the development of Esca decline disease. Integrated protection and production in viticultureIOBC/ wprs Bulletin 2009, 2011, 67. 171–180, https://iobc-wprs.org/product/impact-of-biotic-and-abiotic-factors-on-the-development-of-esca-decline-disease/. (accessed on 28 03 2023). [Google Scholar]
- Jakab, M.K.; Werner, J.; Csikászné Krizsics, A. Az évjáratok hatása a faszöveti betegségek tüneteinek jelentkezésére különböző szőlőfajtákon.G. for Agric. 2016, 20, 39-43. Available online: https://journal.uni-mate.hu/index.php/gfa/issue/view/433/138 (accessed on 1 April 2023).
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M.R. Managing Grapevine Trunk diseases With Respect to Etiology and Epidemiology: Current Strategies and Future Prospects. Plant Dis. 2018, 102, 1, 1239, 31. [Google Scholar] [CrossRef]
- Surico, G.; Mugnai, L.; Marchi, G. Older and more recent observations on esca: a critical overview. Phytopathol. Mediterr. 2006, 45, 68–86. [Google Scholar] [CrossRef]
- Dubos, B. 2002 Le syndrome de l'Esca. In Editions Féret, Maladies cryptogamiques de lavigne, 2° ed., Bordeaux: 127-136. cited in: Dewasme, C.; Mary, S.; Darrieutort, G.; Roby, J.P.;Gambetta, G.A. (2022). Long-Term Esca Monitoring Reveals Disease Impacts on Fruit Yield and Wine Quality. Plant Dis., 106(12), 3076–3082. [CrossRef]
- Fussler, L.; Kobès, N.; Bertrand, F.; Maumy, M.; Grosman, M.; Savary, S. A characterization of grapevine trunk diseases in France from data generated by the National Grapevine Wood Disease Survey. Phytopathology 2008, 98, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Songy, A.; Fernandez, O.; Clément, C.; Larignon, P.; Fontaine, F. Grapevine trunk diseases under thermal and water stresses. Planta 2019, 249, 1655–1679. [Google Scholar] [CrossRef] [PubMed]
- Dewasme, C.; Mary, S.; Darrieutort, G.; Roby, J.-P.; Gambetta, G.A. Long-Term Esca Monitoring Reveals Disease Impacts on Fruit Yield and Wine Quality. Plant Disease 2022, 106(12), 3076–3082. [Google Scholar] [CrossRef] [PubMed]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine Trunk Diseases: A Review of Fifteen Years of Trials for Their Control with Chemicals and Biocontrol Agents. Plant Dis. 2018, 102(7), 1189–1217. [Google Scholar] [CrossRef] [PubMed]
- Sosnowski, M.; Ayres, M.; Wicks, T.; McCarthy, M. In search of resistance to grapevine trunk diseases. Wine Vitic. J. 2013, 28(4), 55–58. Available online: https://winetitles.com.au/wvj/articles/wine-viticulture-journal-volume-28-no-4-2013/in-search-of-resistance-to-grapevine-trunk-diseases/.
- Guan, X.; Essakhi, S.; Laloue, H.; Nick, P.; Bertsch, C.; Chong, J. Mining new resources for grape resistance against Botryosphaeriaceae: a focus on Vitis vinifera subsp. sylvestris. Plant Pathol. 2016, 65, 273–284. [Google Scholar] [CrossRef]
- Martínez-Diz, M.P.; Díaz-Losada, E.; Barajas, E.; Ruano-Rosa, D.; Andrés-Sodupe, M.; Gramaje, D. Screening of Spanish Vitis vinifera germplasm for resistance to Phaeomoniella chlamydospora. Sci. Hortic. 2019, 246, 104–109. [Google Scholar] [CrossRef]
- Reynolds, A.G. 2015 Grapevine breeding in France – a historical perspective. In: Woodhead Publishing Series in Food Science, Tecgnology and Nutrition, Grapevine Breeding Programs for the Wine Industry, Reynolds, A.G. Ed.; Woodhead Publishing, 2015; 65-76. [CrossRef]
- Kozma, P. A szőlő és termesztése I. Akadémiai kiadó: Budapest, Hungary; 2000; Volume 91, p. 319. ISBN 9630577208. [Google Scholar]
- Buonassisi, D.; Colombo, M.; Migliaro, D.; Dolzani, C.; Peressotti, E.; Mizzotti, C.; Velasco, R.; Masiero, S.; Perazzolli, M.; Vezzulli, S. Breeding for grapevine downy mildew resistance: a review of “omics” approaches. Euphytica 2017, 213(5), 103. [Google Scholar] [CrossRef]
- Armijo, G.; Schlechter, R.; Agurto, M.; Munoz, D.; Nunez, C.; Arce-Johnson, P. Grapevine pathogenic microorganisms: understanding infection strategies and host response scenarios. Front. Plant Sci. 2016, 7, 382. [Google Scholar] [CrossRef]
- Merdinoglu, D.; Schneider, C.; Prado, E.; Wiedemann-Merdinoglu, S.; Mestre, P. Breeding for durable resistance to downy and powdery mildew in grapevine. OENO One 2018, 52, 203–209. [Google Scholar] [CrossRef]
- Villano, C.; Aversano, R. Towards grapevine (Vitis vinifera L.) mildews resistance: molecular defence mechanisms and new breeding technologies. Italus Hortus 2020, 27, 1–17. [Google Scholar] [CrossRef]
- Blasi, P.; Blanc, S.; Wiedemann-Merdinoglu, S.; Prado, E.; Rühl, E.H.; Mestre, P.; Merdinoglu, D. Construction of a reference linkage map of Vitis amurensis and genetic mapping of Rpv8, a locus conferring resistance to grapevine downy mildew. Theor. Appl. Genet. 2011, 123(1), 43–53. [CrossRef]
- Kuczmog, A.; Galambos, A.; Horváth, S.; Mátai, A.; Kozma, P.; Szegedi, E.; Putnoky, P. Mapping of crown gall resistance locus Rcg1 in grapevine. Theor. Appl. Genet. 2012, 125(7), 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Schwander, F.; Eibach, R.; Fechter, I.; Hausmann, L.; Zyprian, E.; Topfer, R. Rpv10: a new locus from the Asian Vitis gene pool for pyramiding downy mildew resistance loci in grapevine. Theor. Appl. Genet. 2012, 124, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, H. Review: Research progress in amur grape, Vitis amurensis Rupr. Can. J. Plant. Sci. 2013, 93(4), 565–575. [Google Scholar] [CrossRef]
- Venuti, S.; Copetti, D.; Foria, S.; Falginella, L.; Hoffmann, S.; Bellin, D.; Di Gaspero, G. Historical introgression of the downy mildew resistance gene Rpv12 from the Asian species Vitis amurensis into grapevine varieties. PLoS ONE 2013, 8, e61228. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Wu, W.; Lai, G.; Li, R.; Peng, Y.; Yang, B.; Wang, B.; Yin. ; L.; Qu, J.; Song, S.; Lu, J. Identifying Plasmopara viticola resistance loci in grapevine (Vitis amurensis) via genotyping-bysequencing-based QTL mapping. Plant Physiol. Biochem. 2020, 154, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kozma, P.; Dula, T. Inheritance of resistance to downy mildew and powdery mildew of hybrid family Muscadinia x V. vinifera x V. amurensis x Franco-American hybrid. Acta Hortic., 2003, 603, 457–63. [Google Scholar] [CrossRef]
- Foria, S.; Magris, G.; Jurman, I.; Schwope, R.; De Candido, M.; De Luca, M.; Ivanišević, D.; Morgante, M.; Di Gaspero, G. Extent of wild–to–crop interspecific introgression in grapevine (Vitis vinifera) as a consequence of resistance breeding and implications for the crop species definition. Hort. Res. 2022, 9, uhab010. [Google Scholar] [CrossRef]
- Myles, S.; Boyko, A.R.; Owens, C.L.; Brown, P.J.; Grassi, F.; Aradhya, M.K.; Prins, B.; Reynolds, A.; Chia, J.M.; Ware, D.; Bustamante, C.; Buckler, E.S. Genetic structure and domestication history of the grape. P. Natl. A. Sci. 2011, 108(9), 3530–3535. [Google Scholar] [CrossRef]
- Grassi, F.; De Lorenzis, G. Back to the Origins: Background and Perspectives of Grapevine Domestication. Int. J. Mol. Sci. 2021, 22, 4518. [Google Scholar] [CrossRef]
- Dubos, B. Mise au point sur les maladies de dépéréssiment dans la vignoble francais. Le Progrés Agriculture et Viticulture 1987, 104, 135–140, cited in: Sosnowski, M.R.; Ayres, M.R.; McCarthy, M.G; Scott, E.S. Winegrape cultivars (Vitis vinifera) vary in susceptibility to the grapevine trunk pathogens Eutypa lata and Diplodia seriata. Aust. J. Grape Wine. R. 2022 28, 166-174. [Google Scholar] [CrossRef]
- Carter, M.V. . The status of Eutypa lata as pathogen. Monograph- Phytopathological Paper N0.32 1991 International Mycological Institute, Surrey, UK. https://www.cabdirect.org/cabdirect/abstract/19912304635.
- Borgo, M.; Pegoraro, G.; Sartori, E. usceptibility of grape varieties to esca disease. In BIO Web Conf.; Proceedings of the 39th World Congress of Vine and Wine, Bento Goncalves, Brazil, 24-28. 10.2016. [CrossRef]
- Murolo, S.; Romanazzi, G. Efects of grapevine cultivar, rootstock and clone on esca disease. Australas. Plant Pathol. 2014, 43, 215–221. [Google Scholar] [CrossRef]
- Sosnowski, M.; Ayres, M.; McCarthy, M. Pests and diseases: investigating the potential for resistance to grapevine trunk diseases. Wine Vitic. J. 2016, 31, 41. [Google Scholar] [CrossRef]
- Travadon, R.; Rolshausen, P.E.; Gubler, W.D.; Cadle-Davidson, L.; Baumgartner, K. Susceptibility of cultivated and wild Vitis spp. to wood infection by fungal trunk pathogens. Plant Dis. 2013, 97, 1529–1536. [Google Scholar] [CrossRef] [PubMed]
- Billones-Baaijens, R.; Jones, E.E.; Ridgway, H.J.; Jaspers, M.V. Susceptiblity of common rootstock and scion varieties of grapevines to Botryosphaeriaceae species. Australas. Plant Path. 2014, 43, 25–31. [Google Scholar] [CrossRef]
- Foglia, R.; Landi, L.; Romanazzi, G. Analyses of Xylem Vessel Size on Grapevine Cultivars and Relationship with Incidence of Esca Disease, a Threat to Grape Quality. Appl. Sci. 2022, 12, 1177. [Google Scholar] [CrossRef]
- Feliciano, A.; Eskalen, A.; Gubler, W.D. Diferential susceptibility of three grapevine cultivars to Phaeomoniella chlamydospora in California. Phytopathol. Mediterr. 2004, 43, 66–69. [Google Scholar]
- Cardot, C.; Mappa, G.; La Camera, S.; Gaillard, C.; Vriet, C.; Lecomte, P.; Ferrari, G.; Coutos-Thévenot, P. Comparison of the molecular responses of tolerant, susceptible and highly susceptible grapevine cultivars during interaction with the pathogenic fungus Eutypa lata. Front. Plant Sci. 2019, 10, 991. [Google Scholar] [CrossRef]
- Péros, J.P.; Berger, G. A rapid method to assess the aggressiveness of Eutypa lata isolates and the susceptibility of grapevine cultivars to Eutypa Dieback. Agronomie 1994, 14, 515–523. [Google Scholar] [CrossRef]
- Quaglia, M.; Covarelli, L.; Zazzerini, A. Epidemiological survey on esca disease in Umbria, central Italy. Phytopathol. Mediterr. 2009, 48, 84–91. [Google Scholar] [CrossRef]
- Maul, E.; .Töpfer, R. (2023) Vitis International Variety Catalogue - www.vivc.de –. (accessed on 2 April 2023).
- Sankeymatic online diagram builder. Avaliable online: https://sankeymatic.com (accessed on (2023.01.07.).
- Claverie, M.; Notaro, M.; Fontaine, F.; Wéry, J. Current knowledge on grapevine trunk diseases with complex etiology: A systemic approach. Phytopathol. Med. 2020, 59, 29–53. [Google Scholar] [CrossRef]
- Moret, F.; Lemaître-Guillier, C.; Grosjean, C.; Clément, G.; Coelho, C.; Negrel, J.; Jacquens, L.; Morvan, G.; Mouille, G.; Trouvelot, S.; Fontaine, F.; Adrian, M. Clone-dependent expression of esca disease revealed by leaf metabolite analysis. Front. Plant Sci. 2019, 9, 1960. [Google Scholar] [CrossRef] [PubMed]
- Serra, S.; Ligios, V.; Schianchi, N.; Prota, V.A.; Deidda, A.; Scanu, B. Incidence of grapevine trunk diseases on four cultivars in Sardinia, Southern Italy. Vitis 2021, 60, 35–42. [Google Scholar] [CrossRef]
- Bruez, E.; Lecomte, P.; Grosman, J.; Doublet, B.; Bertsch, C.; Fontaine, F.; Ugaglia, A.; Teissedre, P.-L.; Da Costa, J.-P.; GuerinDubra, L. Overview of grapevine trunk diseases in France in the 2000s. Phytopathol. Mediterr. 2013, 52, 262–275. [Google Scholar]
- Andolfi, A.; Mugnai, L.; Luque, J.; Surico, G.; Cimmino, A.; Evidente, A. Phytotoxins produced by fungi associated with grapevine trunk diseases. Toxins. 2011, 3, 1569–1605. [Google Scholar] [CrossRef] [PubMed]
- Bortolami, G.; Gambetta, G.A.; Cassan, C.; Dayer, S.; Farolfi, E.; Ferrer, N.; Gibon, Y.; Jolivet, J.; Lecomte, P.; Delmas, C.E.L. Grapevines under drought do not express esca leaf symptoms. Proc. Natl. Acad. Sci. USA 2021, 118, e2112825118. [Google Scholar] [CrossRef] [PubMed]
- Pouzoulet, J.; Scudiero, E.; Schiavon, M.; Rolshausen, P.E. Xylem vessel diameter affects the compartmentalization of the vascular pathogen Phaeomoniella chlamydospora in grapevine. Front. Plant. Sci. 2017; 8, 1442. [Google Scholar] [CrossRef]
- Pouzoulet, C.A.; Pivovaroff, A.L.; Santiago, L.S.; Rolshausen, P.E. Can vessel dimension explain tolerance toward fungal vascular wilt diseases in woody plants? Lessons from Dutch elm disease and esca disease in grapevine. Front. Plant. Sci. 2014, 5, 253. [Google Scholar] [CrossRef] [PubMed]
- Csótó, A.; Balling, P.; Nagy, A.; Sándor, E. The Role of Cultivar Susceptibility and Vineyard Age in GTD: Examples from theCarpathian Basin. Acta Agrar. Debr. 2020, 2, 57–63. [Google Scholar] [CrossRef]
- Rolshausen, P.E.; Greve, L.C.; Labavitch, J.M.; Mahoney, N.E.; Molyneux, R.J.; Gubler, W.D. Pathogenesis of Eutypa lata in grapevine: identification of virulence factors and biochemical characterization of cordon dieback. Biochemistry and Cell Biology. Phytopathology 2007, 98, 222–229. [Google Scholar] [CrossRef]
- Smith, M.S.; Centinari, M. Young grapevines exhibit interspecific differences in hydraulic response to freeze stress but not in recovery. Planta 2019, 250, 495–505. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (black measles) and brown wood-streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.L.; Rodriguez-Zaccaro, F.D.; Lee, T.F.; Valdovinos, J.; Toschi, H.S.; Martinez, J.A.; Pratt, R.B. Grapevine xylem development, architecture, and function. In Functional and Ecological Xylem Anatomy; Hacke, U., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 133–162. [Google Scholar]
- Guo, X.W.; Fu, W.H.; Wang, G.J. Studies on cold hardiness of grapevine roots. Vitis 1987, 26, 161–171. [Google Scholar] [CrossRef]
- DeKrey, D.H.; Klodd, A.E.; Clark, M.D.; Blanchette, R.A. Grapevine trunk diseases of cold-hardy varieties grown in Northern Midwest vineyards coincide with canker fungi and winter injury. PLoS One 2022, 17, e0269555. [Google Scholar] [CrossRef] [PubMed]
- Csótó, A.; Balling, P.; Rakonczás, N.; Kovács, C.; Nagy, A.; Sándor, E. The effect of extreme weather conditions on the incidence and spreading of grapevine trunk diseases. In: 16th Congress of the Mediterranean Phytopathological Union (2022) pp. 60-60. , 1 p. https://cyprusconferences.org/mpu2022/wp-content/uploads/2022/04/MPU-2022_ABSTRACTS-ALL-13_04_2022-2.pdf.
- Zhao, Y.; Wang, Z.X.; Yang, Y.M.; Liu, H.S.; Shi, G.L.; Ai, J. Analysis of the cold tolerance and physiological response differences of amur grape (Vitis amurensis) germplasms during overwintering. Sci. Hortic. 2020, 259, 108760. [Google Scholar] [CrossRef]
- Xin, H.P.; Zhu, W.; Wang, L.; Xiang, Y.; Fang, L.; Li, J.; Sun, X.; Wang, N.; Londo, J.P.; Li, S. Genome wide transcriptional profile analysis of Vitis amurensis and Vitis vinifera in response to cold stress. PLoS One 2013, 8, e58740. [Google Scholar] [CrossRef]
- Zhang, J.X.; Wu, X.C.; Niu, R.X.; Liu, Y.; Liu, N.; Xu, W.; Wang, Y. Cold-resistance evaluation in 25 wild grape species. Vitis 2012, 51, 153–160. [Google Scholar] [CrossRef]
- Wang, Y.; Xin, H.; Fan, P.; Zhang, J.; Liu, Y.; Dong, Y.; Wang, Z.; Yang, Y.; Zhang, Q.; Ming, R.; Zhong, G.Y.; Li, S.; Liang, Z. The genome of Shanputao (Vitis amurensis) provides a new insight into cold tolerance of grapevine. T. P. J. 2021, 105, 1495–1506. [Google Scholar] [CrossRef]
- Chai, F.; Liu, W.; Xiang, Y.; Meng, X.; Sun, X.; Cheng, C.; Liu, G.; Duan, L.; Xin, H.; Li, S. Comparative metabolic profiling of Vitis amurensis and Vitis vinifera during cold acclimation. Hortic. Res. 2019, 6, 8. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Liu, S.Y.; Sun, X.; Fang, Y. Oenological potential and health benefits of Chinese non-Vitis vinifera species: An opportunity to the revalorization and to breed new varieties. Food Res. Int. 2020, 137, 109443. [Google Scholar] [CrossRef]
- Ciaffi, M.; Paolacci, A.R.; Paolocci, M.; Alicandri, E.; Bigini, V.; Badiani, M.; Muganu, M. Transcriptional regulation of stilbene synthases in grapevine germplasm differentially susceptible to downy mildew. BMC Plant Biol. 2019, 19, 404. [Google Scholar] [CrossRef]
- Clark, M.D. Development of cold climate grapes in the Upper Midwestern US: The Pioneering Work of Elmer Swenson. In Plant Breeding Reviews; Goldman, I., Ed.; Wiley: Hoboken, NJ, USA, 2020; Volume 43, pp. 31–60. [Google Scholar] [CrossRef]
- Viret, O.; Spring, J.-L.; Gindro, K. Stilbenes: biomarkers of grapevine resistance to fungal diseases OENO One 2018, 52, 235-240. https://doi.org/10.20870/oeno-one.2018.52.3.2033. 52,. [CrossRef]
- Hong, L.; Huan, L.; Yinshan, G.; Satoru, K.; Yuhui, Z.; Guangli, S.; Xiuwu, G. QTLs and candidate genes for downy mildew resistance conferred by interspecific grape (V. vinifera L.×V. amurensis Rupr.) crossing. Sci. Hortic. 2019, 244, 200–207. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Y.; Yin, L.; Lu, J. The mode of host resistance to Plasmopara viticola infection of grapevines. Phytopathology 2012, 102, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Xinlong, L.; Jiao, W.; Ling, Y.; Yali, Z.; Junjie, Q.; Jiang, L. Comparative transcriptome analysis reveals defense-related genes and pathways against downy mildew in Vitis amurensis grapevine. Plant Physiol. Bioch. 2015, 95, 1–14. [Google Scholar] [CrossRef]
- Pretorius, I.S.; Høj, P.B. Grape and wine biotechnology: challenges, opportunities and potential benefits. Aust. J. Grape Wine R. 2005, 11, 83–108. [Google Scholar] [CrossRef]
- Barba, P.; Lillis, J.; Luce,R. S.; Travadon, R.; Osier, M.; Baumgartner, K.; Wilcox, W.F.; Reisch, B.I.; Cadle-Davidson, L. Two dominant loci determine resistance to Phomopsis cane lesions in F1 families of hybrid grapevines. Theor. Appl. Genet. 2018, 131, 1173–1189. [Google Scholar] [CrossRef]
- Travadon, R.; Baumgartner, K.; Rolshausen, P.E.; Gubler, W.D.; Sosnowski, M.R.; Lecomte, P.; Halleen, F.; Peros, J.P. Genetic structure of the fungal grapevine pathogen Eutypa lata from four continents. Plant Pathol. 2012, 61, 85–95. [Google Scholar] [CrossRef]
- Töpfer, R.; Hausmann, L.; Harst, M.; Maul, E.; Zyprian, E.; Eibach, R. New horizons for grapevine breeding. In Methods in Temperate Fruit Breeding; Flachowsky, H., Hanke, M.V., Eds.; Vegetable and cereal science and biotechnology; Global Science Books Ltd., UK: Kagawa, Japan, 2011; pp. 79–100, http://www.globalsciencebooks.info/Online/GSBOnline/images/2011/FVCSB_5(SI1)/FVCSB_5(SI1)79-100o.pdf. (accessed on 07 04 2023); ISBN 978-4-903313-75-7. [Google Scholar]
- Bartholy, J.; Weidinger, T. 2010 Magyarország éghajlati képe. In: Pannon Enciklopédia - Magyarország földje. Ed.: Karátson, D. Urbis, Budapest, Hungary, 2010; pp. 240-241. ISBN: 978-963-9706-68-2.
- Rakonczás, N. Production data of wine grape gene bank (Vitis spp.) of University of Debrecen, East Hungary. I. J. H. S. 2019, 25, 32–36. [Google Scholar] [CrossRef]
- KSH: Borvidékek. Available online: https://www.ksh.hu/docs/hun/teruleti/egyeb_egysegek/borvidekek.pdf (accessed on 8 April 2023).
- Patocskai, Z.; Vidéki, R.; Szépligeti, M.; Bidlo, A.; Kovács; G. Talajviszonyok a Szent György-hegyen. Talajvédelem special issue, 2008, 639-644. http://publicatio.bibl.u-szeged.hu/5647/7/talajtani%20vandorgyules2008.pdf.
- Fehér, O.; Füleky, G.; Madarász, B.; Kertész, A. Hét vulkáni kőzeten kialakult talajszelvény morfológiai és diagnosztikai jellemzői a hazai genetikai talajosztályozás és a WRB (World Reference Base for Soil Resources, 1998) szerint (Morphological and diagnostic properties of seven volcanic soil profiles according to the Hungarian Soil Classification and the World Reference Base for Soil Resources (WRB, 1998). ) Agrokémia és Talajtan 2011, 60, 131–148. [Google Scholar] [CrossRef]
- Díaz, G.A.; Latorre, B.A. Efficacy of paste and liquid fungicide formulations to protect pruning wounds against pathogens associated with grapevine trunk diseases in Chile. Crop Prot. 2013, 2013 46, 106–112. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Leavitt, G.M.; Guerrero, J.C.; Guevara, J.; Gubler, W.D. Identification and Pathogenicity of Lasiodiplodia theobromae and Diplodia seriata, the Causal Agents of Bot Canker Disease of Grapevines in Mexico. Plant Dis. 2008, 92, 519–529. [Google Scholar] [CrossRef]
- Berman, H.B. , "Statistics and Probability", Available online:. Available online: https://stattrek.com/. (accessed on 7 April 2023).
Figure 1.
GTDs symptoms: a, d: leaf stripes; b: partial dieback; c: esca symptoms with white rot and leaf stripe; e: dead plant from previous dieback (indicated by arrow), and new (annual) symptomatic plants (middle and right side); f: dead plant from previous vintage.
Figure 1.
GTDs symptoms: a, d: leaf stripes; b: partial dieback; c: esca symptoms with white rot and leaf stripe; e: dead plant from previous dieback (indicated by arrow), and new (annual) symptomatic plants (middle and right side); f: dead plant from previous vintage.

Figure 2.
Total disease incidence (DI) of grapevine trunk diseases of the most important international and national grape cultivars, surveyed in 3 or 4 Hungarian germplasm collections. The capital letters between brackets indicate the berry skin color: (N): noir, (B): blanc, (G): gris as defined in the VIVC database [51]. Small letters show significant differences based on Mann-Whitney U-test (p<0.05).
Figure 2.
Total disease incidence (DI) of grapevine trunk diseases of the most important international and national grape cultivars, surveyed in 3 or 4 Hungarian germplasm collections. The capital letters between brackets indicate the berry skin color: (N): noir, (B): blanc, (G): gris as defined in the VIVC database [51]. Small letters show significant differences based on Mann-Whitney U-test (p<0.05).
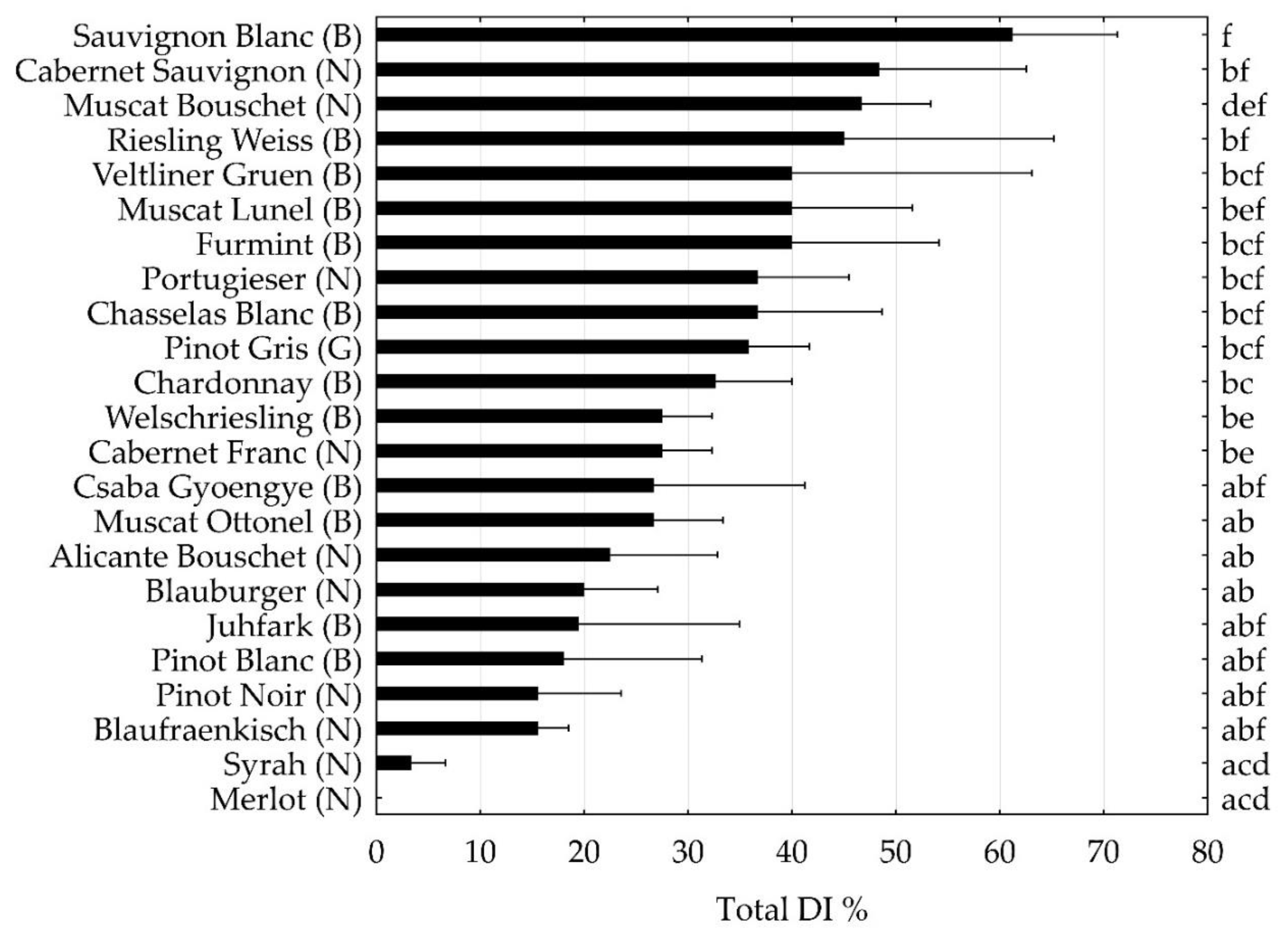
Figure 3.
Distribution of the studied cultivars regarding their Vitis pure or mixed ancestry, and the GTD pathogens sensitivity groups. Diagram created by SankeyMATIC [52].
Figure 3.
Distribution of the studied cultivars regarding their Vitis pure or mixed ancestry, and the GTD pathogens sensitivity groups. Diagram created by SankeyMATIC [52].
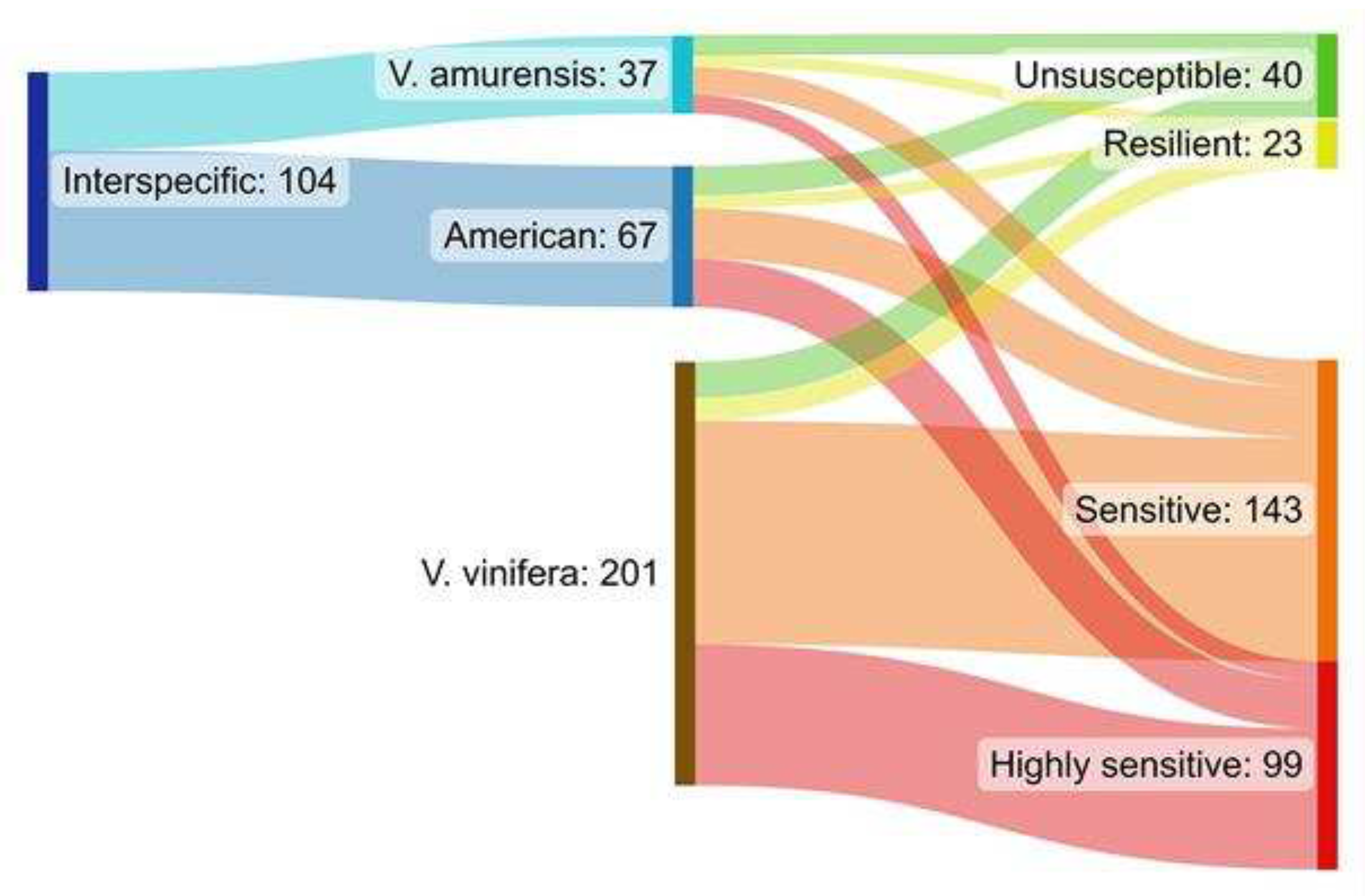
Figure 4.
The ratio of the cultivars categorized in the less GTD pathogens sensitive group within monophyletic V. vinifea cultivars (Vv), and those in all interspecific cultivars combined (I), or with the interspecific cultivars with those with Asian (V. amurensis) (Va) or American species (V. labrusca, V. riparia or V. rupestris) (Va) ancestors treated separately. Results of the binomial probability test, indicating the difference between the examined group and the averages of all cultivars: * = p<0.05; ** = p<0.01.
Figure 4.
The ratio of the cultivars categorized in the less GTD pathogens sensitive group within monophyletic V. vinifea cultivars (Vv), and those in all interspecific cultivars combined (I), or with the interspecific cultivars with those with Asian (V. amurensis) (Va) or American species (V. labrusca, V. riparia or V. rupestris) (Va) ancestors treated separately. Results of the binomial probability test, indicating the difference between the examined group and the averages of all cultivars: * = p<0.05; ** = p<0.01.

Figure 5.
Proportion of plant loss within all recorded GTD symptoms (a) comparing the average of these cultivars with that in exclusively V. vinifera (Vv) ancestors, and with that in all the interspecific hybrids (I) and (b) the same comparison with Vv but now with hybrids with V. amurensis (Va) in their pedigree or those with American(Ao) (V. labrusca, V. riparia or V. rupestris) ancestry, separatedly. Small letters indicate significant differences between datasets based on Mann-Whitney U-test (p<0.01).
Figure 5.
Proportion of plant loss within all recorded GTD symptoms (a) comparing the average of these cultivars with that in exclusively V. vinifera (Vv) ancestors, and with that in all the interspecific hybrids (I) and (b) the same comparison with Vv but now with hybrids with V. amurensis (Va) in their pedigree or those with American(Ao) (V. labrusca, V. riparia or V. rupestris) ancestry, separatedly. Small letters indicate significant differences between datasets based on Mann-Whitney U-test (p<0.01).
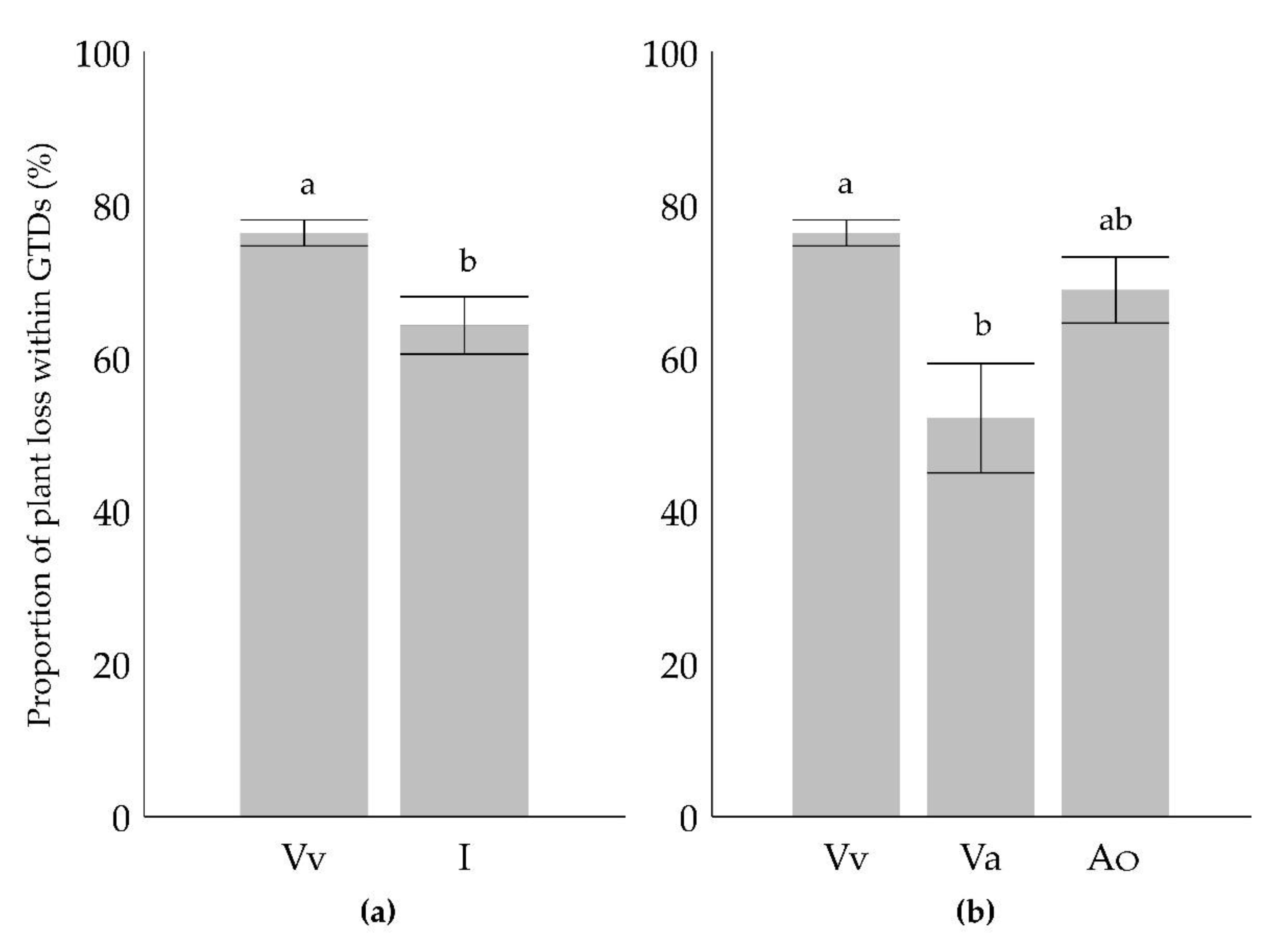
Figure 6.
The location of the surveyed germplasm collections in Hungary. Different colors indicate the different Wine Regions [87].
Figure 6.
The location of the surveyed germplasm collections in Hungary. Different colors indicate the different Wine Regions [87].
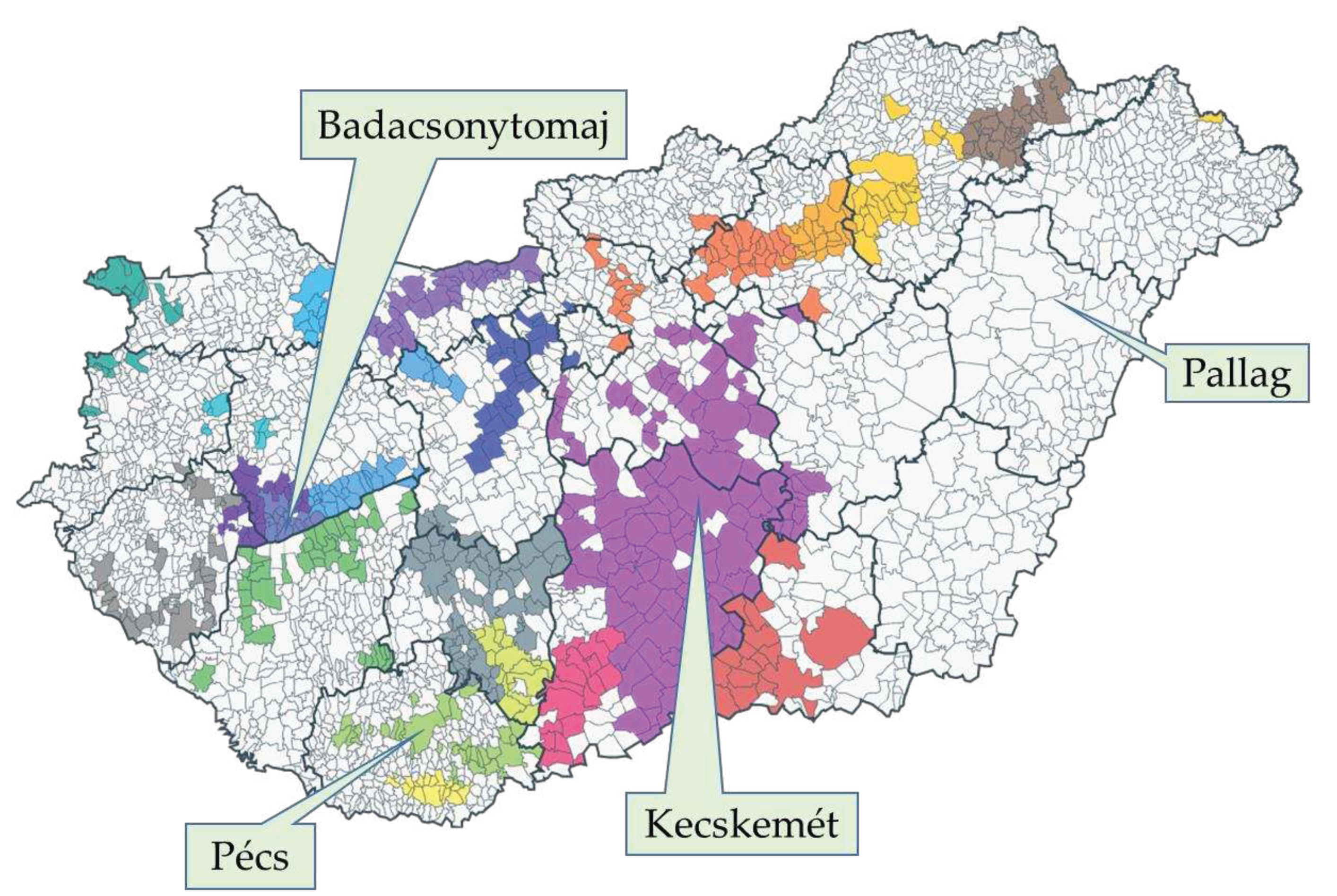
Table 1.
The tolerance of V. vinifera cultivars to different grapevine trunk diseases. Adopted from Songy et al. [17].
Table 1.
The tolerance of V. vinifera cultivars to different grapevine trunk diseases. Adopted from Songy et al. [17].
| Cultivars1 | GTDs | Inoculation test/Disease incidence survey3 | References | ||
|---|---|---|---|---|---|
| Tolerance | Disease2 | ||||
| White | Chardonnay | high | BD, Eutypa | Test | [44] |
| medium | BD | Survey | [43] | ||
| medium | Esca | Survey | [41] | ||
| Pinot Gris | high | BD, Eutypa | Survey | [43] | |
| medium | Esca | Survey | [10] | ||
| Riesling | high | Eutypa | Survey | [43] | |
| medium | BD | Test | [45] | ||
| medium/low | Esca | Test | [44] | ||
| Sauvignon Blanc | high | BD | Test | [45] | |
| medium | Eutypa | Test and Survey | [43] | ||
| low | BD | Test and Survey | [43] | ||
| low | Esca | Survey | [42,46] | ||
| Semillon | high | BD, Eutypa | Test and Survey | [43] | |
| low | Esca | Survey | [10] | ||
| Thompson seedless | high | Esca | Test | [44,47] | |
| medium/low | Eutypa | Test | [44] | ||
| low | BD, Eutypa | Test | [44,47] | ||
| Ugni Blanc | medium/high | BD | Survey | [43] | |
| low | Eutypa | Test | [48] | ||
| low | Esca, Eutypa | Test | [49] | ||
| Survey | [41,43] | ||||
| Welshriesling | high | BD, Eutypa | Test and Survey | [43] | |
| Survey | [41] | ||||
| low | Esca | Survey | [7] | ||
| Red | Cabernet Franc | medium/high | Eutypa | Test | [44] |
| medium | BD | Test | [44] | ||
| low | Esca | Test and Survey | [43] | ||
| Cabernet Sauvignon | high | BD | Test | [45] | |
| low | Eutypa | Test | [48] | ||
| low | Esca, Eutypa | Survey | [41,46,50 | ||
| BD | Survey | [43] | |||
| Grenache | high | Esca, Eutypa | Survey | [43] | |
| high | Esca | Test | [47] | ||
| BD | medium/high | Survey | [43] | ||
| Merlot | high | Eutypa | Test | [44,48] | |
| medium/high | BD | Test | [44] | ||
| medium | Esca | Survey | [42,50] | ||
| Pinot Noir | high | Esca | Survey | [41] | |
| Eutypa, Esca | Test and Survey | [43] | |||
| medium | BD | Test and Survey | [43] | ||
| Sangiovese | high | BD, Esca, Eutypa | Test and Survey | [43] | |
| medium | Esca | Survey | [41] | ||
| Syrah | high | Esca | Survey | [41] | |
| low | BD, Eutypa | Test | [21,44] | ||
| Test and Survey | [43] | ||||
| Hybrid(V. labrusca hybrid) | Concord (Vitis labrusca hybrid) |
high | BD, Esca, Eutypa | Test | [44] |
1Cultivar primer names from VIVC database [51]. 2BD: Botryosphaeria dieback; Eutypa: Eutypa dieback. 3Test: Inoculation of cuttings; Survey: in field survey of disease incidence.
Table 2.
Total disease incidence (DI) of grapevine trunk diseases (GTDs) (mean±SE) and the ratio of dead plants (mean±SE) within GTD symptoms in the different germplasm collections. Small letters show significant differences based on Mann-Whitney U-test (p<0.05).
Table 2.
Total disease incidence (DI) of grapevine trunk diseases (GTDs) (mean±SE) and the ratio of dead plants (mean±SE) within GTD symptoms in the different germplasm collections. Small letters show significant differences based on Mann-Whitney U-test (p<0.05).
| Location | No. Cultivars | GTDs | |
|---|---|---|---|
| Total DI % (±SE) | Ratio of dead plants (% ±SE) | ||
| Badacsonytomaj | 90 | 44.58(±2.62) c | 74.63(±3.14) a |
| Kecskemét | 130 | 28.05(±2.19) a | 76.49(±3.11) a |
| Pallag | 166 | 37.05(±2.16) b | 73.78(±3.10) a |
| Pécs | 151 | 28.41(±1.92) a | 69.94(±3.23) a |
| Total | 537 | 33.70(±1.13) | 73.56(±1.59) |
Table 7.
Categories of cultivars with multiple Vitis species ancestry.
| Ancestors in parent or grandparent level | Categorization I. | Categorization II. |
|---|---|---|
| Vitis vinifera | Vitis vinifera (Vv) | Vitis vinifera (Vv) |
| Occurrence of American species 1 | Interspecific (I) | American origin (Ao) |
| Occurrence of Vitis amurensis | Interspecific (I) | Vitis amurensis origin (Va) |
1 V. labrusca, V. riparia or V. rupestris.
Table 8.
Categorization of cultivars according to the observed sensitivity toward grapevine trunk diseases (GTDs).
Table 8.
Categorization of cultivars according to the observed sensitivity toward grapevine trunk diseases (GTDs).
| Sensitivity categories | GTDs symptoms | ||
|---|---|---|---|
| Two groups | Four groups | apoplexy (dead plant) | leaf symptoms and fresh dieback |
| More sensitive | Highly sensitive (HS) | exclusively | - |
| Sensitive (S) | present | present | |
| Less sensitive | Resilient (R) | - | exclusively |
| Unsusceptible (U) | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



