
Submitted:
19 April 2023
Posted:
20 April 2023
You are already at the latest version
Abstract
The present study examined mechanisms underlying memory deficits in patients with Parkinson disease (PD)and their associations with structural metrics. Nineteen PD and 22 matched controls underwent two memory experiments. In experiment 1 (delayed memory task), subjects were asked to remember an array of colored rectangles with varying memory set sizes [Low-Load (2 items), Low-Load with Distractor, & High-Load (5 items)]. After a 7s delay period, they reported whether the orientation of any relevant figures had changed (test period). In experiment 2 (working memory task), memory arrays were presented in varying set sizes (2 to 6 items) with no distractors that were followed by a 2s delay period and subsequent test period. Brain MRI data were acquired to assess structural differences (volumes and cortical thickness) in brain areas related to attention, working memory storage, and episodic memory. Compared to controls, PD patients had lower memory capacity scores in all memory load conditions for experiment 1 (p <0.021) whereas there were no group differences in any memory load conditions for experiment 2 (p>0.06). In addition, PD patients had lower thickness in the left superior temporal gyrus (p=0.02). Lower thickness values in the left superior temporal gyrus were significant predictors of lower delayed memory performance in Low-Load and Low-Load with Distractor conditions (ps’<0.044) and working memory performance of memory load conditions of 4 and 5 items (p’s<0.012). The present findings suggest that PD patients may have intact working memory storage capacity but impaired attentional filtering and memory consolidation that may lead to lower delayed memory scores. Lower delayed memory in PD may partly be associated with lower cortical thickness in the left superior temporal gyrus.
Keywords:
visuospatial working memory
; delayed memory
; Parkinson’s disease
; attentional filtering
; lower memory storage
1. Introduction
Parkinson’s disease (PD) is a progressive neurodegenerative disorder with predominant loss of dopaminergic neurons in subtantia nigra pars compacta and subsequent depletion of dopamine levels in the basal ganglia. Prominent characteristics of PD include motor symptoms such as tremor, rigidity and bradykinesia. While the motor symptoms dominate clinical pictures in PD, many patients with PD experience a wide range of non-motor symptoms. These may include autonomic disturbances (e.g., constipation and bladder control problems) and sensory complaints (e.g., numbness, burning or tingling sensation) but also include psychiatric (e.g., depression and anxiety) or non-psychiatric cognitive dysfunctions such as executive functions, attention, and memory (Gabrieli et al., 1996; Zgaljardic et al., 2003; Owen, 2004).
Given that the basal ganglia have extensive interconnections with the prefrontal cortex (Jellinger, 2001; Lewis et al., 2003), cognitive symptoms in PD are often ascribed to compromised information flow through this fronto-striatal pathway (Lewis et al., 2003). In fact, the pattern of cognitive deficits in PD appears to be similar to that observed in frontal lobe patients such as difficulties with planning, selective attention, and set shifting (Morris et al., 1988, Owen, 1995; West et al., 1998). Converging evidence, however, suggests that PD may experience various cognitive dysfunctions beyond frontal lobe-related executive dysfunctions.
Working memory deficits in PD
Working memory is a capacity limited system which temporarily maintains information in a highly active and accessible state but also manipulates that information for performing tasks (Baddeley, 2003; Conway et al., 2005; Cowan et al., 2005). Theoretically, poor performance on working memory tasks can be due to reduced storage capacity per se, inability to effectively process information, or both (Cowan et al., 2005). Many studies have shown poor performance on various working memory tasks in PD patients and reported lower working memory capacity. One possible cause, however, may be a reduced ability to filter out irrelevant information so that they unnessarily usurp capacity-limited working memory space.
Episodic memory deficits in PD
Although PD present difficulties in various cognitive domains, learning/memory problems, particularly deficits in episodic memory (Bäckman et al. 2001) are one of the most common and devastrating cognitive symptoms in PD (Weintraub et al 2015, Yarnall 2014). Episodic memoryis a system that involves consciously retrieving information that was acquired in a particular time and space (Tulving 1984) and a key function of medial temporal lobe memory areas, especially hippocampus (Eichenbaum and Lipton 2008; Sugar and Moser 2019). Episodic memory problems have been reported as earliest neurobehavioral deficits observed in dementia patients (Bäckman et al. 2001). The prevalence of dementia in PD ranges between 24% and 50% and up to 80% PD patients may eventually develop dementia (Aarsland and Kurz 2010; Anang et al. 2014). Previous studies reported more than 20% of newly diagnosed PD patients had lower performance on episodic memory tasks (Breen and Drutyte 2013).
In the present study, two visual array comparison experiments were conducted to examine the mechanisms underlying potential memory deficits in PD patients. In both experiments, participants viewed an array of colored rectangles. In some trials, the presented array contained task-irrelevant items that should be ignored. After a short (2s) or relatively long (7s) delay, subjects reported whether the orientation of any relevant figures had changed. Structural differences were assessed by volumes or cortical thickness in brain areas related to attention (e.g., superior frontal gyrus, superior parietal lobe, and intraparietal sulcus for the dorsal attentional system and inferior and middle frontal gyri, inferior parietal lobe, and superior temporal gyrus for ventral attentional system), working memory storage capacity (e.g., intraparietal sulcus), and episodic memory (e.g., medial temporal lobe structures). Our central hypotheses were: 1) compared to controls, PD patients will show lower performance in both working memory and delayed memory tasks; 2) The memory performance in PD will be lower with distractors than without; 3) There will be significant structural differences in brain areas related to attention, working memory storage capacity, or episodic memory; 4) There will be positive associations between MRI structural metrics in ROIs and memory metrics.
2. Materials and Methods
2.1. Participants
Fourty one subjects (19 patients with Parkinson’s disease (PD) and 22 age- and education-matched neurologically normal subjects) were recruited for the study. All subjects except for one patient and one control subject reported having normal color vision and normal or corrected-to-normal acuity. One patient and one control who were identical twins were partially color-blind, but they were able to tell the difference between red and green rectangles used in this study. All subjects had Mini-Mental Status Examination (MMSE) scores >=26. Depression was evaluated by means of Geriatric Depression Scale-long form (GDS; Sheikh and Yesavage, 1986). Three patients and one control subject were taking antidepressants (e.g., Fluoxetine, Bupropion, or Sertraline) at the time of the study. The general pattern of results was essentially the same with or without these three patients, so their data have been retained.
PD patients were free from other neurological disorders. Fifteen patients were receiving the dopamine precursor Levodopa as treatment. On patient was not taking any anti-parkinsonian medication. Another patient was taking only Pramipexole (a dopamine agonist). The remaining two patients were receiving Ropinirole in conjunction with Trihexyphenidyl (an anticholinergic agent) or Azilect (an MAO inhibitor). On the morning of the experiment, PD patients skipped their initial dose of anti-parkinsonian medication. The mean withdrawal period of 11 h (at least 9 h) would not be enough to achieve complete clearance. Rather, it was intended to enhance differences between experimental groups while minimizing the burden imposed on patients. The severity of the disease was reassessed just before the start of the experiment using the Hoehn and Yahr scale (1967). Control subjects reported neither a history of neurological problems nor any significant current psychiatric disorder. All participants gave their informed consent according to procedures approved by the ethics board at the University of Missouri-Columbia (Approval number: 1170557).
2.2. Stimuli and Procedures for Neuropsychological Experiments
Stimulus arrays were presented within a 4 x 7.3° rectangular region centered at fixation on a dark background. Arrays consisted of either two or five colored rectangles. Item positions were randomized across trials. Both red and green rectangles subtended .65 x 1.15° of visual angle, with orientations selected randomly from a set of four possible values (vertical, horizontal, left-tilting 45°, and right-tilting 45°).
In experiment 1 (Figure 1), each trial began with a 2 s get-ready signal (3 yellow crosses). Next there was an instructional cue, which was followed by a 1 s long memory array, consisting of either two red or two red and three green rectangles. The instructional cue indicated whether subjects should ignore the green rectangles as distracters (“X”: Low Load+ Distracter) or remember them as part of target memory array with no distracters (“o”: Low Load, or “O”: High Load).
After a 7-s long retention interval, subjects were presented with a single probe stimulus for 2 s and asked to press a specified button on either the left- or right-hand key pad to report whether the orientation of the tested rectangle changed (same or different). Following an inter-trial interval of 2, 4 or 6 s, the next trial commenced, starting with the yellow get-ready signal. Accuracy was emphasized over speed, and subjects were allowed to correct their response before the next trial began. The experiment 2 consists of twelve blocks of 12 trials (total 144 trials). Between blocks, participants were allowed to take as long a break as they wanted.
In experiment 2, subjects performed a change detection task to estimate their working memory capacity. Subjects sat upright in a comfortable chair and viewed the stimuli at a distance of ~70 cm. In this version of the task, there were neither precues nor green distracters, only relevant items. The number of to-be-remembered red rectangles varied from 2 to a maximum of 6, which slightly exceeds the typical capacity of an older adult. Each trial began with a 2 s get-ready signal followed by a 1 s long memory array. After a brief (200 ms) pattern mask and a 2 s delay period, the test stimulus was presented until a response was made. Following a 3 s inter-trial interval, the next trial commenced. Accuracy was emphasized over speed, and subjects were allowed to correct their response before the next trial began. This version of the task was structured as five blocks of 32 trials (total 160 trials).
2.3. MRI image acquisition and image processing
Images were acquired on the 3-Tesla Siemens scanner at the University of Missouri’s Brain Imaging Center. Technical parameters for the structural scans were as follows: T1-weighted MPRAGE images: repetition time (TR) =1920 ms, echo time (TE) =2.92 ms, flip angle=9º, field of view (FOV) = 256 mm, matrix: 256 x 256, 176 slices in the sagittal plane, voxel size = 1x1x1 mm, slice thickness = 1 mm with acquisition time of 8 min, 13 s. T2-weighted images: TR = 3200 ms, TE = 402 ms, FOV = 256 mm, matrix = 258 x 256, slice thickness = 1 mm.
2.3.1. Brain regions of interest
Brain regions that previously had reported associated with attention including attentional filtering process (superior frontal gyrus, superior parietal lobe, and intraparietal sulcus for the dorsal attentional system and inferior and middle frontal gyri, inferior parietal lobe, and superior temporal gyrus for ventral attentional system; Figure 3a), working memory storage (retention; intraparietal sulcus) or episodic memory [medial temporal lobe (hippocampus, and entorhinal and parahippocampal cortices; Figure 3b) were selected as regions-of-interest (ROIs). The ROIs were defined for each subject using Freesurfer. The segmentation quality then was confirmed visually by a reviewer blinded to group assignment.
2.3.2. Hippocampal volumes and cortical thickness
Volumetric segmentation and cortical parcellation for thickness calculation were performed with the Freesurfer image analysis suite (http://surfer.nmr.mgh.harvard.edu/). The processing included motion correction, removal of non-brain tissue using a hybrid watershed/surface deformation procedure (Ségonne et al. 2004), automated Talairach transformation, and segmentation of the deep gray matter volumetric structures and parcellation of cortical gay matter structures (Fischl et al. 2002; Fischl et al. 2004).
2.4. Statistical Analysis
Group comparisons of demographic data were conducted using one-way analysis of variance (ANOVA) or χ2 test. Group comparisons of neuropsychological and MRI structural ROI metrics were conducted using multivariate analysis of covariance (MANCOVA) in order to account for potential inter-correlations among outcome variables. For MANCOVA, age and education were used as covariates. To compare the performance differences between Low-Load and Low-Load with Distractor conditions, within-subjects analysis of covariance (ANCOVA) was performed using age and education as covariates. When comparing the hippocampal volume, total intracranial volume (TIV) additionally was used as a covariate. The primary neuropsychological metrics were K scores that were derived from hit rate (proportion of correct responses when a change was present) and false alarm rate (proportion of incorrect responses on no-change trials): K = N * (H-FA), where N is the number of relevant, to-be-stored items, H is the hit rate and FA is the false alarm rate as suggested by Cowan (2001). Association analyses of MRI structural (volume and cortical thickness) with K scores were conducted for controls and PD patients separately using Pearson partial correlation analyses with adjustment for age and education. Following the association analyses, regression analyses were conducted for the variables that demonstrated significant associations between MRI structural and neuropsychological metrics after controlling for age and education. In order to determine structural metrics that could explain the variances of group differences in memory metrics, a stepwise regression analysis was conducted for controls and PD separately. For the stepwise regression analysis, structural metrics that showed significant associations with memory metrics were included in addition to age and education. Statistical significance was defined as α=0.05. The association analyses were corrected for multiple comparisons using the Holm-Bonferroni stepdown method (Holm 1979) to control the familywise error rate (FWER) at p=0.05. We report uncorrected raw p values but indicate significant results with FWER-correction. SAS 9.4 was used for all statistical analyses.
3. Results
3.1. Demographics
There were no significant group differences in age, gender, education, MMSE, and depression scores (p’s >0.073).

Table 1.
Demographics.
| Controls (N=22) |
PD Patients (N=19) |
p-values | ||
|---|---|---|---|---|
| a. Demographics | ||||
| Age (y) | 69.05 ± 5.58 | 66.16 ± 8.81 | 0.211 | |
| Gender (m/f) | 12/10 | 14/5 | 0.205 | |
| Education (years) | 14.77 ± 3.19 | 16.63 ± 3.25 | 0.073 | |
| MMSE | 29.40 ± 0.99 | 29.00 ± 1.29 | 0.284 | |
| Hoehn & Yahr Scale | 0 | 2.03 ± 0.77 | ||
| Disease duration (years) | 0 | 6.65 ± 4.76 | ||
| GDS | 2.78 | 4.11 ± 3.11 | 0.147 | |
Note. Descriptive data for participants’ demographics. MMSE: Mini-Mental-State-Exam, GDS: Geriatric Depression Scale.
3.2. Group comparison of memory metrics
In Experiment 1, there were significant group differences of memory K scores in overall memory conditions [F(3,35)=3.04, p=0.042) and in each individual memory condition [F(1,37)=5.79, p=0.021, R2=0.152 for Low-Load; [F(1,37)=8.33, p=0.007, R2=0.206 for Low-Load with Distractor; F(1,37)=5.89, p=0.020, R2=0.164 for High-Load; Figure 4a). The K socre in the Low-Load with distractor condition was significantly lower compared to that in Low-Load condition for PD (t=-2.81, p=0.008) but not for controls (t=-1.29, p=0.206).
In Experiment 2, there were no significant group differences in overall working memory condicitons (F(5,33)=0.84, p=0.534) and in each individual conditions with different memory set sizes (F’s >0.49, p’s >0.060, R2<0.199; Figure 4b).
3.3. Group comparison of MRI structural metrics
There were no overall significant group differences in ROIs related to dorsal and ventral attentional systems (p’s>0.121). When considering attention-related individual subregions, there was a significantly lower thickness in the left superior temporal gyrus for PD compared to controls [F(3,37)=5.92, p=0.020, R2=0.270]. There were no significant group differences in bilateral intraparietal sulci (p’s>0.064), entorhinal and parahippocampal cortices (p’s>0.390). There also were no significant group differences in bilateral hippocampal volumes (p’s >0.114).
3.4. Associations of MRI structural metrics with memory metrics
Within controls, higher thickness in the superior frontal gyrus was associated with higher memory K scores in the Low-Load condition of experiment 1 (R=0.466, p=0.038). There were no signficant correlations between structural metrics in other ROIs and memory K scores in experiment 1 (p’s>0.083). For working memory conditions in experiment 2, higher thickness in the left inferior frontal-opercular gyrus and lower thickness in the left superior temporal gyrus were associated with higher K scores in memory load condition of 2 itmes (p’s <0.048). Higher thickness in bilateral superior frontal gyri were associated with higher memory K scores in memory load condition of 6 itmes (R’s >0.460, p’s<0.041). None of the associations, however, remained significant after FWER-correction. There were no signficant correlations between structural metrics in other ROIs and memory K scores in experiment 1 and 2 (p’s>0.052; see Supplementary Materials 1 for details).
Subsequent regression analyses confirmed that higher thickness in the left superior frontal gyrus was a significant predictor of higher K scores in the Low-Load condition of experiment 1 after controlling for age and education (ß=2.0199, t=2.23, p=0.038, R2=0.273). Higher thickness in the left inferior frontal-opercular gyrus and lower thickness in the left superior temporal gyrus were significant predictors of memory load of 2 items in experiment 2 (ß=1.2427, t=2.93, p=0.009 for inferior frontal gyrus and ß=-1.1137, t=-2.76, p=0.014 for superior temporal gyrus, total R2=0.486).
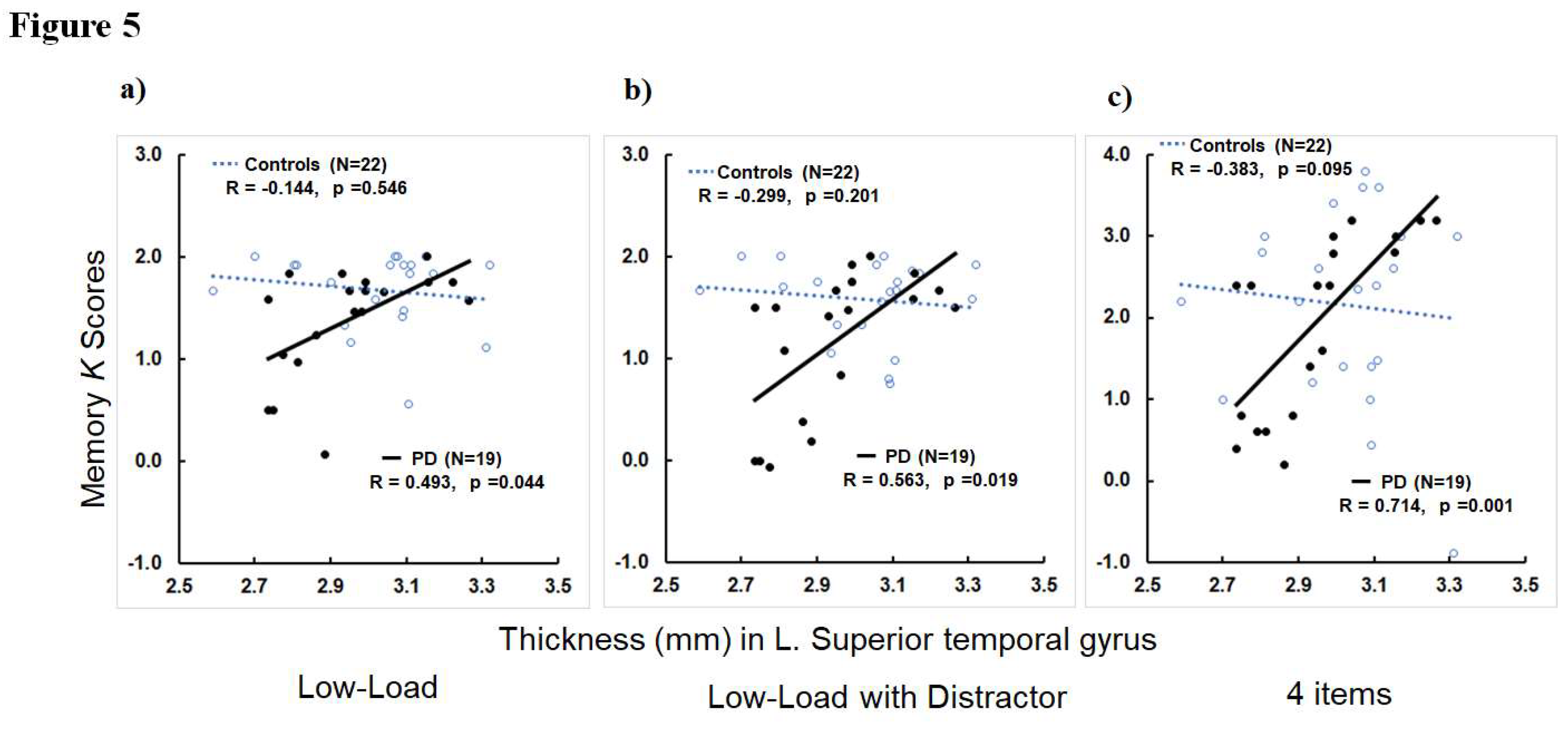
Within PD patients, greater thickness in the left superior temporal gyrus was associated with higher K scores in the Low-Load and Low-Load with Distractor conditions of experiment 1 (R=0.563, p=0.019 for Low-Load and R=0.493, p=0.044 for Low-Load with Distractor; Figure 5a-b). Higher thicknessin in the right intraparietal sulcus was associated with higher K scores in the Low-Load condition (R=0.490, p=0.046). For memory conditions in experiment 2, greater thickness in the left superior temporal gyrus was associated with higher K scores in all memory load conditions of 2 to 6 items (R’s>0.575, p’s<0.016; Figure 5c for 4 items). The correlations between left superior temporal thickness and K scores in 3 and 4 items conditions remained significant after FWER-correction. Greater thickness in the right superior temporal gyrus also was associated with higher K scores in memory load condition of 2 itmes (R=0.514, p=0.035). Higher thickness in the left inferior frontal-opercular gyrus was associated with higher K scores in memory load conditions of 2, 3, 4, and 6 items (R’s>0.489, p’s<0.046). The association between left inferior frontal-opercular gyrus and memory load of 2 items remained significant after FWER-correction. Higher thickness in the right inferior frontal-opercular gyrus also was associated with higher K scores in memory load condition of 6 items (R=0.681, p=0.003) that remained significant after FWER-correction. Higher thickness in the left inferior frontal-triangular gyrus was associated with higher K scores in memory load conditions of 2 and 3 items (R’s>0.489, p’s<0.046). Higher thickness in bilateral supramarginal gyrus was associated with higher K scores of memory load condition of 6 items (R’s>0.570, p’s<0.017). The association between left supramarginal thickness and memory load of 6 items remained significant after FWER-correction. Greater left intraparietal sulcus thickness was associated with higher K scores of the memory load condition of 3 items (R=0.568, p=0.017). Higher thickness in the right superior frontal gyrus was associated with higher K scores of memory load conditions of 2 and 3 items (R’s>0.550, p’s<0.022). Greater thickness in the right superior parietal gyrus was associated with higher K scores in memory set load conditions of 2 and 3 items (R’2>0.568, p’s<0.017). Higher thickness in the right intraparietal sulcus was associated with higher K scores of memory load conditions of 3 and 6 items (R’s>0.569, p’s<0.017). The correlations of right superior frontal and parietal gyri and intraparietal sulcus thickness with K scores in memory condition of 3 items remained significant after FWER-correction. For hippocampal volume metrics, there were no significant correlattions of hippocampal volumes with any memory metrics in both experiments 1 and 2 ( p’s >0.235; see Supplementary Materials 1 for details).
Subsequent regression analyses revealed that higher thickness in the left superior temporal gyrus was a significant predictor of higher K scores in the Low-Load with Distractor condition of experiment 1 (ß=2.5189, t=2.64, p=0.019, R2=0.397) and memory load conditions of 4 and 5 items in experiment 2 (ß=4.1656, t=2.71, p=0.017, R2=0.655 for 4 items and ß=4.4272, t=2.86, p=0.012, R2=0.580 for 5 items). Higher thickness in the left supramarginal gyrus was associated with higher K scores in the memory load condition of 6 items (ß=6.6444, t=2.63, p=0.025, R2=0.891).
3.5. Stepwise regression analysis to determine factors predicting memory metrics
The stepwise regression analyses were conducted using structural metrics that showed significant associations with memory metrics. In controls, left inferior frontal-opercular and superior temporal gyri and bilateral superior frontal gyri thickness values were used for stepwise regression analyses in addition to age and education as predictors for K scores of Low-Load in experiment 1 and memory load conditions of 2 and 6 items in experiment 2 (see Supplementary Materials 1 for association analysis results). For PD, thickness values in left inferior frontal-triangular and right superior frontal and parietal gyri, and bilateral inferior frontal-opercular, supramarginal, superior temporal gyri and bilateral intraparietal sulci were used as predictors analyses in addition to age and education.
In controls, higher thickness in the left superior frontal gyrus was a significant predictor of higher K scores in the Low-Load condition of experiment 1 (ß=2.0199, p=0.038, partial R2=0.202). Higher thickness in the left inferior frontal-opercular gyrus and lower thickness in the left superior temporal gyrus were significant predictors of memory load of 2 items in experiment 2 (ß=1.2427, p=0.009, partial R2=0.260 for inferior frontal gyrus and ß=-1.1137, t=-2.76, p=0.014 for superior temporal gyrus, partial R2=0.229). Higher thickness in the right superior frontal gyrus was a significant predictor of higher K scores of memory load condition of 6 items (ß=7.3819, p=0.021, partial R2=0.218).
In PD, higher thickness in the left superior temporal gyrus was a significant predictor for higher K scores of Low-Load (ß=1.6034, p=0.044, partial R2=0.203) and Low-Load with Distractor (ß=2.5189, p=0.019, partial R2=0.279) conditions in experiment 1. Higher thickness in the left inferior frontal-opercular gyrus was a significant predictor for higher K scores of memory condition of 2 items in expediment 2 (ß=3.2703, p=0.006, partial R2=0.365). Higher thickness in the right superior parietal thickness was asignificant predictor for higher K scores of memory condition of 3 items (ß=4.8596, p=0.007, partial R2=0.315). Higher thickness in the left superior temporal gyrus a significant predictor for higher K scores of memory load conditions of 4 items (ß=4.3709, p=0.001, partial R2=0.360) and 5 items (ß=4.4272, p=0.012, partial R2=0.228). Higher thickness in the right inferior frontal-opercular and supramarginal gyri were significant predictors for higher K scores of memory condition of 6 items (ß=4.7980, p=0.019, partial R2=0.242 for inferior frontal and ß=6.2697, p=0.019, partial R2=0.093 for supramarginal).
4. Discussion
The present study examined mechanisms underlying memory deficits in PD patients and their associations with structural metrics. Compared to controls, PD patients had lower delayed memory whereas there were no group differences in working memory performance for any memory load conditions. In addition, PD patients had lower thickness in the left superior temporal gyrus, aregion related ventral attentional system. To the contrary, there were no structural differences in ROIs related to dorsal attentional system, working memory storage, and episodic memory. Lower thickness values in the left superior temporal gyrus were significant predictors of lower delayed memory performance in Low-Load and Low-Load with Distractor conditions and working memory performance of memory load conditions of 4 and 5 items. The present findings suggest that PD patients may have intact working memory storage capacity but impaired attentional filtering and memory consolidation that may lead to lower delayed memory scores. Lower delayed memory in PD may partly be associated with lower cortical thickness in the left superior temporal gyrus. Future studies should be warranted to elucidate early brain structural or functional changes related to poorer delayed memory in PD.
4.1. Lower dealyed memory due to reduced memory consolidation
In the present study, 2 different delay periods (2s vs. 7s) were utilized between memory and test arrays to mimic working memory and episodic memory formation processes. Interestingly, PD patients’ memory K scores were comparable to those of controls when their memory was tested 2s after the memory array offset whereas their memory performance became significantly worse with a 7s delay period. This result suggests that PD patients may have intact working memory storage capacity compared to neurologically normal controls. PD patients’ ability to encode and retrieve visual information may also be intact compared to those in controls. This result is inconsistent with a previous finding demonstrating that PD patients had lower K scores and CDA (contralateral delay activity) amplitudes, EEG correlates reflecting items held in working memory, in memory load conditions even with no distractors (Lee et al. 2010). In that study, however, a bilateral display was utilized to measure CDA, which required additional filtering by the participant even in conditions with no distracters. So, it is possible that patients’ lower K scores and CDA amplitudes in that study could be due to impaired filtering rather than diminished working memory storage capacity. The current finding supports this interpretation.
Instead, PD patients seemed to have difficulty to continue holding information in memory over a prolonged period of time where memory consolidation may increasingly gain importance to form stable episodic memory. Memory consolidation is a process by which a temporary and unstable memory trace is transformed into a more stable and long-lasting memory (Squire et al. 2015) and may serve as a critical component for successful episodic memory formation process. Note that studies that reported impaired episodic memory performance in PD typically tested memory performance after a 20 min delay period (Weintraub et al. 2015; Yarnall et al. 2014). The current finding, however, suggests that impaired episodic memory formation process may be detected as early as within a 7s delay period. Thus, the curent finding of lower delayed memory performance in PD without difference in working memory performance suggests intact working memory storage but impaired memory consolidation process in PD.
Alzheimer’s disease (AD) is the most common age-related neurodegenerative disorder, comprising about 50-70% of dementia cases (Brookmeyer et al. 2011) and the major and early AD-related behavioral deficits entail learning/memory problems (Bäckman et al. 2005), particularly episodic memory (Bäckman et al. 2001). Given that selective episodic memory decline may occur in AD-at-risk-populations (Bäckman et al. 2001; Lim et al. 2014), current finding of lower K scores in delayed memory conditions suggests that AD-related early neurobehavioral differences may occur in PD despite still intact working memory performance.
4.2. Lower delayed memory due to impaired attentional filtering
Both controls and patients had some difficulty ignoring distracters. They demonstrated lower K scores compared to Low-Load condition although the number of to-be-remembered items was same for both conditions with and without distractors. Interestingly, K score difference between these two memory conditions was significant only for PD patients, suggesting difficulty to ignore irrelevant information (attentional filtering difficulty) in PD. This finding is consistent with a previous study (Lee etal., 2010) reporting that PD patients had lower K scores but higher CDA (contralateral delay activity) amplitudes in memory load conditions with distractors suggesting lower memory performance for to-be-remembered task-relevant information probably due to unnessary storage for task-irrelevant items. The current finding also is in line with previous findings reporting a critical role of fronto-striatal pathway in controlling the access of incoming information into memory system (McNab and Klingberg, 2008; Cohen and Frank, 2009). Patients with impaired fronto-striatal pathway may especially be vulnerable to filtering deficits. The loss of dopaminergic input to the basal ganglia in Parkinson’s disease may lead to a diminished fronto-striatal functions and reduced ability to filter out distracters so that they do not unnecessarily usurp memory space. Indeed, PD patients do seem to be vulnerable to attention deficits in a general sense. Studies have shown that PD patients have more difficulty inhibiting automatic response to ignored salient but irrelevant stimuli such as flanking distracters (Praamstra & Plat, 2001; Verleger et al., 2010).
4.3. Structural correlates of lower delayed memory in PD
In the present study, we examined structural metrics (volumes and cortical thickness) in brain areas that were known to be related to attentional processes including attentional filtering, working memory storage, or episodic memory including memory consolidation processes. Consistent with comparable working memory storage capacity throughout different memory load conditions, there were no significant group differences in the intraparietal sulcus, an area known to be sensitive to working memory storage (Postle et al., 2006). Instead, PD patients had lower thickness in the left superior temporal gyrus. Moreover, lower thickness in the left superior temporal gyrus was a significant predictor of both lower delayed memory performance and working memory performance. The superior temporal gyrus is known to be part of ventral attention network that is generally associated with bottom-up detection of salient stimuli by shifting attention to unexpected information while a dorsal attention network employs top–down goal-directed attentional shifts to designated features of stimuli (Nani et al. 2019). Both ventral and dorsal attention netwkrs, however, seem to simultaneously influence and integrate each other in real life situations (Nani et al. 2019). The volume of the left superior temporal gyrus has been reported to sensitively reflect verbal working memory capacity and ability to comprehend spoken sentences (Leff et al. 2009). Since properly understanding spoken sentences may be involved in accessing to long-term storage of lexico-semantic representations of verbal information, the role of the left superior temporal gyrus in accessing to long-term storage especially for verbal information has been implicated (Leff et al., 2008; Schofield et al., 2009). The significant associations of the left superior temporal gyrus thickness with working memory and delayed memory capacity observed in the present study may extend previous findings suggesting that the role of the left superior temporal gyrus may be multimodal and generic to working memory capacity and transferring information to and accessing to long-term storage. Future studies are waranteed to replicated current findings and test this inriguing hypothesis.
It is intriguing to note that there were no volumetric or thickness differences in any of medial temporal ROIs or associations with memory metrics although PD patients demonstrated robust decline in delayed memory performance. It is possible that our sample size was too small reliably to detect structural differences in medial temporal areas. It also is possible that patients’ lower delayed memory observed in this study may partly be associated with early brain microstructural or functional changes that may occur before medial temporal morphological changes. For example, studies report that diffusion tensor imaging (DTI) metrics that measure random translational water motion may reflect early microstructural changes (Le Bihan et al. 2001). These DTI measures have been suggested to be more sensitive than volume or thickness measures in capturing AD-related early brain changes including changes in the medial temporal lobe (Douaud et al. 2013; Fellgiebel et al. 2006; Lancaster et al. 2016; Müller et al. 2007). Further studies utilizing brain imaging markers that sensitiely capture early brain microstructural or functional changes should be warranted to confirm current findings and to elucidate neural correlates of delayed memory in PD.
5. Conclusions
Lower attentional filtering and memory consolidation problems may contribute to lower delayed memory performance in PD that may partly be associated with lower cortical thickness in the left superior temporal gyrus.
Supplementary Materials
Supplementary Materials 1.
Funding
This work was supported by the Dong-A University research fund.
Institutional Review Board Statement
All participants gave their informed consent according to procedures approved by the ethics board at the University of Missouri-Columbia (protocol code: 1170557 on 2010.09.01).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data can be available upon requests.
Acknowledgments
This work was supported by the Dong-A University research fund.
Conflicts of Interest
The author declares no conflict of interest.
References
- Albin, R.L.; Young, A.B.; Penney, J.B. The functional anatomy of basal ganglia disorders. Trends Neurosci. 1989, 12, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, J. & Friston, K. (2000). Voxel-based morphometry - The methods. NeuroImage, 11, 805-821. [CrossRef]
- Atkinson, R. C. , & Shiffrin, R. M. (1968). Human memory: A proposed system and its control processes. In K. W. Spence & J. T. Spence (Eds.). The psychology of learning and motivation: Advances in research and theory (Vol. 2, pp. 89–195). New York: Academic Press. [CrossRef]
- Awh, E.; Barton, B.; Vogel, E.K. Visual Working Memory Represents a Fixed Number of Items Regardless of Complexity. Psychol. Sci. 2007, 18, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Awh & Vogel (2008). The bouncer in the brain. Nature Neuroscience, 11(1), 5-6. [CrossRef]
- Baddeley, A.D. , & Hitch, G.J. (1974).Working memory. In G. Bower (Ed.), The psychology of learning and motivation (Vol. VIII, pp. 47-90). New York: Academic Press.
- Baddeley, A. Working memory: looking back and looking forward. Nat. Rev. Neurosci. 2003, 4, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.M.B.; Waterman, C.; Scarpa, M.; Castiello, U. Covert visuospatial attentional mechanisms in Parkinson's disease. Brain 1995, 118, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Bradley, A.V.; Welch, J.L.; Dick, D.J. Visuospatial working memory in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 1989, 52, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.; Steur, E.N.J.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Buchel, C.; Friston, K.J. Modulation of connectivity in visual pathways by attention: cortical interactions evaluated with structural equation modelling and fMRI. Cereb. Cortex 1997, 7, 768–778. [Google Scholar] [CrossRef]
- Cameron, I. G. , Watanabe, M., Pari, G., & Munoz, D. P. (2010). Executive impairment in Parkinson's disease: response automaticity and task switching. Neuropsychologia, 48(7), 1948-57. [CrossRef]
- Case, R.; Kurland, D.; Goldberg, J. Operational efficiency and the growth of short-term memory span. J. Exp. Child Psychol. 1982, 33, 386–404. [Google Scholar] [CrossRef]
- Chang, C.; Crottaz-Herbette, S.; Menon, V. Temporal dynamics of basal ganglia response and connectivity during verbal working memory. NeuroImage 2007, 34, 1253–1269. [Google Scholar] [CrossRef]
- Cohen, J. Statistical power analysis for the behavioral sciences, 2nd ed.; Lawrence Erlbaum Associates: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Cohen, M.X.; Frank, M.J. Neurocomputational models of basal ganglia function in learning, memory and choice. Behav. Brain Res. 2009, 199, 141–156. [Google Scholar] [CrossRef]
- function in learning, memory and choice. Behav Brain Res, 199(1), 141-156. [CrossRef]
- Cools, R.; Gibbs, S.E.; Miyakawa, A.; Jagust, W.; D'Esposito, M. Working Memory Capacity Predicts Dopamine Synthesis Capacity in the Human Striatum. J. Neurosci. 2008, 28, 1208–1212. [Google Scholar] [CrossRef] [PubMed]
- Cools, R.; Barker, R.A.; Sahakian, B.J.; Robbins, T.W. Enhanced or Impaired Cognitive Function in Parkinson's Disease as a Function of Dopaminergic Medication and Task Demands. Cereb. Cortex 2001, 11, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Cools, R.; Ivry, R.B.; D'Esposito, M. The Human Striatum is Necessary for Responding to Changes in Stimulus Relevance. J. Cogn. Neurosci. 2006, 18, 1973–1983. [Google Scholar] [CrossRef] [PubMed]
- Cools, R.; Miyakawa, A.; Sheridan, M.; D'Esposito, M. Enhanced frontal function in Parkinson's disease. Brain 2009, 133, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J. A. , Sagar, H. J., Jordan, N., Harvey, N. S., & Sullivan, E. V. (1991). Cognitive impairment in early, untreated Parkinson's disease and its relationship to motor disability. Brain, 114 (5), 2095-2122. [CrossRef]
- Cowan, N. The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behav. Brain Sci. 2001, 24, 87–114. [Google Scholar] [CrossRef] [PubMed]
- Cowan, N. , Elliott, E. M., Saults, J. S., Morey, C. C., Mattox, S., Hismjatullina, A.
- Cowan, N.; Elliott, E.M.; Saults, J.S.; Morey, C.C.; Mattox, S.; Hismjatullina, A.; Conway, A.R. On the capacity of attention: Its estimation and its role in working memory and cognitive aptitudes. Cogn. Psychol. 2005, 51, 42–100. [Google Scholar] [CrossRef]
- Cowan, N.; Li, D.; Moffitt, A.; Becker, T.M.; Martin, E.A.; Saults, J.S.; Christ, S.E. A Neural Region of Abstract Working Memory. J. Cogn. Neurosci. 2011, 23, 2852–2863. [Google Scholar] [CrossRef]
- Conway, A.R.A.; Kane, M.J.; Bunting, M.F.; Hambrick, D.Z.; Wilhelm, O.; Engle, R.W. Working memory span tasks: A methodological review and user’s guide. Psychon. Bull. Rev. 2005, 12, 769–786. [Google Scholar] [CrossRef]
- Dalrymple-Alford, J.C.; Kalders, A.S.; Jones, R.D.; Watson, R.W. A central executive deficit in patients with Parkinson's disease. J. Neurol. Neurosurg. Psychiatry 1994, 57, 360–367. [Google Scholar] [CrossRef]
- Daneman, M.; Carpenter, P.A. Individual differences in working memory and reading. J. Verbal Learn. Verbal Behav. 1980, 19, 450–466. [Google Scholar] [CrossRef]
- Daneman, M.; Merikle, P.M. Working memory and language comprehension: A meta-analysis. Psychon. Bull. Rev. 1996, 3, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Eimer, M.; Kiss, M. An electrophysiological measure of access to representations in visual working memory. Psychophysiology 2009, 47, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Erdfelder, E.; Faul, F.; Buchner, A. GPOWER: A general power analysis program. Behav. Res. Methods Instrum. Comput. 1996, 28, 1–11. [Google Scholar] [CrossRef]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-Mental State”. A Practical Method for Grading the Cognitive State of Patients for the Clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.J. Dynamic Dopamine Modulation in the Basal Ganglia: A Neurocomputational Account of Cognitive Deficits in Medicated and Nonmedicated Parkinsonism. J. Cogn. Neurosci. 2005, 17, 51–72. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Frith, C.D.; Frackowiak, R.S.J. Time-dependent changes in effective connectivity measured with PET. Hum. Brain Mapp. 1993, 1, 69–79. [Google Scholar] [CrossRef]
- Friston, K.J. Functional and effective connectivity in neuroimaging: A synthesis. Hum. Brain Mapp. 1994, 2, 56–78. [Google Scholar] [CrossRef]
- Friston, K.; Buechel, C.; Fink, G.; Morris, J.; Rolls, E.; Dolan, R.; Friston, K.; Buechel, C.; Fink, G.; Morris, J.; et al. Psychophysiological and Modulatory Interactions in Neuroimaging. NeuroImage 1997, 6, 218–229. [Google Scholar] [CrossRef]
- Friston, K.J. , Buchel, C. (2000). Attentional modulation of effective connectivity from V2 to V5/MT in humans. Proc Natl Acad Sci U S A, 97, 7591–7596. [CrossRef]
- Friston, K.J., Harrison, L., Penny, W. (2003). Dynamic causal modelling. NeuroImage, 19, 1273–1302. [CrossRef]
- Gabrieli, J.D.E.; Singh, J.; Stebbins, G.T.; Goetz, C.G. Reduced working memory span in Parkinson's disease: Evidence for the role of frontostriatal system in working and strategic memory. Neuropsychology 1996, 10, 322–332. [Google Scholar] [CrossRef]
- Gerfen, C.R. Molecular effects of dopamine on striatal-projection pathways. Trends Neurosci. 2000, 23, S64–S70. [Google Scholar] [CrossRef]
- Geweke, J. Measurement of Linear Dependence and Feedback Between Multiple Time Series. J. Am. Stat. Assoc. 1982, 77, 304. [Google Scholar] [CrossRef]
- Gilbert, B. , Belleville, S., Bherer, L., & Chouinard, S. (2005). Study of verbal working memory in patients with Parkinson's disease. Neuropsychology, 19, 106-114. [CrossRef]
- Goebel, R. , Roebroeck, A., Kim, D.S., Formisano, E. (2004). Directed cortical interactions during dynamic sensory-motor mapping. In: Duncan, J. (Ed.), Attention and Performance XX. Oxford University Press, New York, pp. 439–462.
- Good, C.; Johnsrude, I.; Ashburner, J.; Henson, R.; Friston, K.; Frackowiak, R. A voxel-based morphometric study of ageing in 465 normal adult human brains. 5th IEEE EMBS International Summer School on Biomedical Imaging, 2002.. LOCATION OF CONFERENCE, FranceDATE OF CONFERENCE; p. 16 pp.
- Gruber, A.J.; Dayan, P.; Gutkin, B.S.; Solla, S.A. Dopamine modulation in the basal ganglia locks the gate to working memory. J. Comput. Neurosci. 2006, 20, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Gouttard, S.; Styner, M.; Joshi, S.; Smith, R.G.; Hazlett, H.C.; Gerig, G. Subcortical structure segmentation using probabilistic atlas priors. Medical Imaging. LOCATION OF CONFERENCE, COUNTRYDATE OF CONFERENCE; pp. 65122J–65122J-11.
- Harrison, A.; Jolicoeur, P.; Marois, R. "What" and "Where" in the Intraparietal Sulcus: An fMRI Study of Object Identity and Location in Visual Short-Term Memory. Cereb. Cortex 2010, 20, 2478–2485. [Google Scholar] [CrossRef] [PubMed]
- Cereb Cortex, 20(10): 2478–2485. [CrossRef]
- Harrison, L.; Penny, W.; Friston, K.; Harrison, L.; Penny, W.; Friston, K.; Harrison, L.; Penny, W.; Friston, K. Multivariate autoregressive modeling of fMRI time series. NeuroImage 2003, 19, 1477–1491. [Google Scholar] [CrossRef] [PubMed]
- Hochstadt, J.; Nakano, H.; Lieberman, P.; Friedman, J. The roles of sequencing and verbal working memory in sentence comprehension deficits in Parkinson’s disease. Brain Lang. 2006, 97, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Hoehn M, Yahr M. (1967). Parkinsonism: onset, progression and mortality. Neurology, 17: 427–42.
- Hsieh, S.; Hwang, W.-J.; Tsai, J.-J.; Tsai, C.-Y. Visuospatial Orienting of Attention in Parkinson's Disease. Percept. Mot. Ski. 1996, 82, 1307–1315. [Google Scholar] [CrossRef]
- Jellinger, K. A. (2001). The pathology of Parkinson’s disease. Advances in Neurology, 86, 55-72.
- Jolicœur, P.; Brisson, B.; Robitaille, N. Dissociation of the N2pc and sustained posterior contralateral negativity in a choice response task. Brain Res. 2008, 1215, 160–172. [Google Scholar] [CrossRef]
- Joshi, S.; Davis, B.; Jomier, M.; Gerig, G. Unbiased diffeomorphic atlas construction for computational anatomy. NeuroImage 2004, 23, S151–S160. [Google Scholar] [CrossRef]
- Just, M.A.; Carpenter, P.A. A capacity theory of comprehension: Individual differences in working memory. Psychol. Rev. 1992, 99, 122–149. [Google Scholar] [CrossRef]
- Kay, S.M. (1988). Modern Spectral Estimation: Theory and Application. Prentice Hall, Englewood Cliffs, NJ.
- Kennett, S.; van Velzen, J.; Eimer, M.; Driver, J. Disentangling gaze shifts from preparatory ERP effects during spatial attention. Psychophysiology 2006, 44, 69–78. [Google Scholar] [CrossRef]
- Kensinger, E. A. , Shearer, D. K., Locascio, J. J., Growdon, J. H., & Corkin, S. (2003). Working memory in mild Alzheimer's disease and early Parkinson's disease. Neuropsychology, 17(2), 230-239. [CrossRef]
- Lee, E. Y. , Cowan, N., Vogel, E. K., Rolan, T., Valle-Inclan, F., & Hackley, S. A. (2010). Visual working memory deficits in patients with Parkinson's disease are due to both reduced storage capacity and impaired ability to filter out irrelevant information. Brain, 133(9), 2677-2689. [CrossRef]
- Lewis, S.J.G.; Dove, A.; Robbins, T.W.; Barker, R.A.; Owen, A.M. Cognitive impairments in early Parkinson’s disease are accompanied by reductions in activity in frontostriatal neural circuitry. J. Neurosci. 2003, 23, 6351–6356. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S. J. , Slabosz, A., Robbins, T. W., Barker, R. A., & Owen, A. M. (2005). Dopaminergic basis for deficits in working memory but not attentional set-shifting in Parkinson's disease. Neuropsychologia, 43(6), 823-832. [CrossRef]
- Linden, D.E.; Bittner, R.A.; Muckli, L.; Waltz, J.A.; Kriegeskorte, N.; Goebel, R.; Singer, W.; Munk, M.H. Cortical capacity constraints for visual working memory: dissociation of fMRI load effects in a fronto-parietal network. NeuroImage 2003, 20, 1518–1530. [Google Scholar] [CrossRef]
- Lu, Z.-L.; Neuse, J.; Madigan, S.; Dosher, B.A. Fast decay of iconic memory in observers with mild cognitive impairments. 102, 1797. [Google Scholar] [CrossRef]
- Luck, S. J. , & Hillyard, S. A. (1994). Spatial filtering during visual search: evidence from human electrophysiology. J Exp Psychol Hum Percept Perform, 20(5), 1000-1014. [CrossRef]
- Luck, S.J.; Vogel, E.K. The capacity of visual working memory for features and conjunctions. Nature 1997, 390, 279–281. [Google Scholar] [CrossRef]
- Matsui, H.; Nishinaka, K.; Oda, M.; Niikawa, H.; Komatsu, K.; Kubori, T.; Udaka, F. Wisconsin Card Sorting Test in Parkinson's disease: diffusion tensor imaging. Acta Neurol. Scand. 2007, 116, 108–112. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Klingberg, T. Prefrontal cortex and basal ganglia control access to working memory. Nat. Neurosci. 2007, 11, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Miller, G. A. (1956). The magical number seven, and or minus two: Some limits on our capacity for processing information. Psychological Review, 63, 81–97. [CrossRef]
- Miyake, A. , & Shah, P. (Eds.). (1999). Models of Working Memory: Mechanisms of active maintenance and executive control. Cambridge, UK: Cambridge University Press.
- Morris, R.G.; Downes, J.J.; Sahakian, B.J.; Evenden, J.L.; Heald, A.; Robbins, T.W. Planning and spatial working memory in Parkinson's disease. J. Neurol. Neurosurg. Psychiatry 1988, 51, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.A.; Sherman, S.J.; Frank, M.J. A dopaminergic basis for working memory, learning and attentional shifting in Parkinsonism. Neuropsychologia 2008, 46, 3144–3156. [Google Scholar] [CrossRef] [PubMed]
- O'Reilly, R.C.; Frank, M.J. Making Working Memory Work: A Computational Model of Learning in the Prefrontal Cortex and Basal Ganglia. Neural Comput. 2006, 18, 283–328. [Google Scholar] [CrossRef]
- Owen, A.M.; Iddon, J.L.; Hodges, J.R.; A Summers, B.; Robbins, T.W. Spatial and non-spatial working memory at different stages of Parkinson's disease. Neuropsychologia 1997, 35, 519–532. [Google Scholar] [CrossRef]
- Owen, A.M. Cognitive Dysfunction in Parkinson’s Disease: The Role of Frontostriatal Circuitry. Neurosci. 2004, 10, 525–537. [Google Scholar] [CrossRef]
- Pashler, H. Familiarity and visual change detection. Percept. Psychophys. 1988, 44, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Patenaude, B.; Smith, S.M.; Kennedy, D.N.; Jenkinson, M. A Bayesian model of shape and appearance for subcortical brain segmentation. NeuroImage 2011, 56, 907–922. [Google Scholar] [CrossRef] [PubMed]
- Petit, L.; Courtney, S.M.; Ungerleider, L.G.; Haxby, J.V. Sustained Activity in the Medial Wall during Working Memory Delays. J. Neurosci. 1998, 18, 9429–9437. [Google Scholar] [CrossRef] [PubMed]
- Postle, B.R., Ferrarelli, F., Hamidi, M., Feredoes, E., Massimini, P. M., Alexander, A., and Tononi, G. (2006). Repetitive transcranial magnetic stimulation dissociates working memory manipulation from retention functions in prefrontal, but not posterior parietal cortex. Journal of Cognitive Neuroscience 2006, 18, 1712–1722. [PubMed]
- Praamstra, P. , & Plat, F. M. (2001). Failed suppression of direct visuomotor activation in Parkinson's disease. J Cogn Neurosci, 13(1), 31-43.
- Praamstra, P. , Stegeman, D. F., Cools, A. R., & Horstink, M. W. (1998). Reliance on external cues for movement initiation in Parkinson's disease. Evidence from movement-related potentials. Brain, 121, 167-177. [CrossRef]
- Roebroeck, A.; Formisano, E.; Goebel, R. Mapping directed influence over the brain using Granger causality and fMRI. NeuroImage 2005, 25, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Ruchkin, D.S.; Grafman, J.; Cameron, K.; Berndt, R.S. Working memory retention systems: A state of activated long-term memory. Behav. Brain Sci. 2003, 26, 709–728. [Google Scholar] [CrossRef] [PubMed]
- Salimi-Khorshidi, G.; Smith, S.M.; Nichols, T.E. Adjusting the effect of nonstationarity in cluster-based and TFCE inference. NeuroImage 2011, 54, 2006–2019. [Google Scholar] [CrossRef]
- Samuel, M.; O Ceballos-Baumann, A.; Blin, J.; Uema, T.; Boecker, H.; E Passingham, R.; Brooks, D.J. Evidence for lateral premotor and parietal overactivity in Parkinson's disease during sequential and bimanual movements. A PET study. Brain 1997, 120, 963–976. [Google Scholar] [CrossRef]
- Sen, S. , Flynn M., Du G., Tröster A., An H., Huang X. (2011). Manganese accumulation in the olfactory bulbs and other brain regions of "asymptomatic" welders. Toxicol Sci. 121(1):160-7. [CrossRef]
- Sheikh, J. I. & Yesavage, J. A. (1986).Geriatric Depression Scale (GDS): Recent evidence and development of a shorter version. Clinical Gerontology: A Guide to Assessment and Intervention. NY: The Haworth Press, 165-173.
- Skeel, R. L. , Crosson, B., Nadeau, S. E., Algina, J., Bauer, R. M., & Fennell, E. B. (2001). Basal ganglia dysfunction, working memory, and sentence comprehension in patients with Parkinson's disease. Neuropsychologia, 39(9), 962-971. [CrossRef]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. NeuroImage 2004, 23, S208–S219. [Google Scholar] [CrossRef]
- Smith, S.M. & Nichols, T.E. (2009).Threshold-freeclusterenhancement: Addressing problems of smoothing, threshold dependence and localisation in cluster inference. NeuroImage, 44, 83-98. [CrossRef]
- Sperling, G. The information available in brief visual presentations. Psychol. Monogr. Gen. Appl. 1960, 74, 1–29. [Google Scholar] [CrossRef]
- Surmeier, D.J.; Ding, J.; Day, M.; Wang, Z.; Shen, W. D1 and D2 dopamine-receptor modulation of striatal glutamatergic signaling in striatal medium spiny neurons. Trends Neurosci. 2007, 30, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Turner, M. L. , & Engle, R. W. (1989). Is working memory capacity task dependent? Journal of Memory and Language, 28, 127–154. [CrossRef]
- Unsworth, N.; Redick, T.S.; Heitz, R.P.; Broadway, J.M.; Engle, R.W. Complex working memory span tasks and higher-order cognition: A latent-variable analysis of the relationship between processing and storage. Memory 2009, 17, 635–654. [Google Scholar] [CrossRef] [PubMed]
- Verleger, R.; Hagenah, J.; Weiß, M.; Ewers, T.; Heberlein, I.; Pramstaller, P.P.; Siebner, H.R.; Klein, C. Responsiveness to distracting stimuli, though increased in Parkinson's disease, is decreased in asymptomatic PINK1 and Parkin mutation carriers. Neuropsychologia 2010, 48, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Vogel, E.K.; Woodman, G.F.; Luck, S.J. Pushing around the Locus of Selection: Evidence for the Flexible-selection Hypothesis. J. Cogn. Neurosci. 2005, 17, 1907–1922. [Google Scholar] [CrossRef]
- Vogel, E.K.; McCollough, A.W.; Machizawa, M.G. Neural measures reveal individual differences in controlling access to working memory. Nature 2005, 438, 500–503. [Google Scholar] [CrossRef]
- West, R. , Ergis, A. M., Winocur, G., & Saint-Cyr, J. (1998). The contribution of impaired working memory monitoring to performance of the self-ordered pointing task in normal aging and Parkinson’s disease. Neuropsychology, 12, 546-554. [CrossRef]
- Wiecki, T. V. , & Frank, M. J. (2010). Neurocomputational models of motor and cognitive deficits in Parkinson's disease. Prog Brain Res, 183, 275-297. [CrossRef]
- Woodward, T.S.; Bub, D.N.; A Hunter, M. Task switching deficits associated with Parkinson’s disease reflect depleted attentional resources. Neuropsychologia 2002, 40, 1948–1955. [Google Scholar] [CrossRef]
- with Parkinson’s disease reflect depleted attentional resources. Neuropsychologia, 40(12), 1948–1955.
- Woolrich, M.W.; Jbabdi, S.; Patenaude, B.; Chappell, M.; Makni, S.; Behrens, T.; Beckmann, C.F.; Jenkinson, M.; Smith, S.M. Bayesian analysis of neuroimaging data in FSL. NeuroImage 2009, 45, S173–S186. [Google Scholar] [CrossRef]
- Wu, T. , & Hallett, M. (2005). A functional MRI study of automatic movements in patients with Parkinson's disease, Brain, 128 (10), 2250-2259. [CrossRef]
- Zgaljardic, D.J.; Borod, J.C.; Foldi, N.S.; Mattis, P. A Review of the Cognitive and Behavioral Sequelae of Parkinson's Disease: Relationship to Frontostriatal Circuitry. Cogn. Behav. Neurol. 2003, 16, 193–210. [Google Scholar] [CrossRef]
- Aarsland D, Kurz MW. 2010. The epidemiology of dementia associated with parkinson's disease. Brain pathology. 20(3):633-639. [CrossRef]
- Anang JB, Gagnon J-F, Bertrand J-A, Romenets SR, Latreille V, Panisset M, Montplaisir J, Postuma RB. 2014. Predictors of dementia in parkinson disease: A prospective cohort study. Neurology. 83(14):1253-1260. [CrossRef]
- Bäckman L, Jones S, Berger A-K, Laukka EJ, Small BJ. 2005. Cognitive impairment in preclinical alzheimer's disease: A meta-analysis. Neuropsychology. 19(4):520. [CrossRef]
- Bäckman L, Small BJ, Fratiglioni L. 2001. Stability of the preclinical episodic memory deficit in alzheimer's disease. Brain : a journal of neurology. 124(1):96-102. [CrossRef]
- Breen KC, Drutyte G. 2013. Non-motor symptoms of parkinson’s disease: The patient’s perspective. Journal of neural transmission. 120:531-535. [CrossRef]
- Brookmeyer, R.; Evans, D.A.; Hebert, L.; Langa, K.M.; Heeringa, S.G.; Plassman, B.L.; Kukull, W.A. National estimates of the prevalence of Alzheimer's disease in the United States. Alzheimer's Dement. 2011, 7, 61–73. [Google Scholar] [CrossRef]
- Douaud G, Menke RA, Gass A, Monsch AU, Rao A, Whitcher B, Zamboni G, Matthews PM, Sollberger M, Smith S. 2013. Brain microstructure reveals early abnormalities more than two years prior to clinical progression from mild cognitive impairment to alzheimer's disease. Journal of Neuroscience. 33(5):2147-2155. [CrossRef]
- Eichenbaum, H.; Lipton, P.A. Towards a functional organization of the medial temporal lobe memory system: Role of the parahippocampal and medial entorhinal cortical areas. Hippocampus 2008, 18, 1314–1324. [Google Scholar] [CrossRef]
- Fellgiebel, A.; Dellani, P.R.; Greverus, D.; Scheurich, A.; Stoeter, P.; Müller, M.J. Predicting conversion to dementia in mild cognitive impairment by volumetric and diffusivity measurements of the hippocampus. Psychiatry Res. Neuroimaging 2006, 146, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Fischl, B.; Salat, D.H.; Busa, E.; Albert, M.; Dieterich, M.; Haselgrove, C.; van der Kouwe, A.; Killiany, R.; Kennedy, D.; Klaveness, S.; et al. Whole Brain Segmentation: Automated Labeling of Neuroanatomical Structures in the Human Brain. Neuron 2002, 33, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Fischl, B.; Salat, D.H.; van der Kouwe, A.J.; Makris, N.; Ségonne, F.; Quinn, B.T.; Dale, A.M. Sequence-independent segmentation of magnetic resonance images. NeuroImage 2004, 23 (Suppl. 1), S69–S84. [Google Scholar] [CrossRef] [PubMed]
- Holm, S. 1979. A simple sequentially rejective multiple test procedure. Scandinavian journal of statistics.65-70.
- Lancaster, M.A.; Seidenberg, M.; Smith, J.C.; Nielson, K.A.; Woodard, J.L.; Durgerian, S.; Rao, S.M. Diffusion Tensor Imaging Predictors of Episodic Memory Decline in Healthy Elders at Genetic Risk for Alzheimer’s Disease. J. Int. Neuropsychol. Soc. 2016, 22, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Le Bihan, D.; Mangin, J.; Poupon, C.; Clark, C.A.; Pappata, S.; Molko, N.; Chabriat, H. Diffusion tensor imaging: Concepts and applications. J. Magn. Reson. Imaging 2001, 13, 534–546. [Google Scholar] [CrossRef]
- Leff, A.P.; Schofield, T.M.; Crinion, J.T.; Seghier, M.L.; Grogan, A.; Green, D.W.; Price, C.J. The left superior temporal gyrus is a shared substrate for auditory short-term memory and speech comprehension: evidence from 210 patients with stroke. Brain 2009, 132, 3401–3410. [Google Scholar] [CrossRef]
- Lim, Y.Y.; Maruff, P.; Pietrzak, R.H.; Ames, D.; Ellis, K.A.; Harrington, K.; Lautenschlager, N.T.; Szoeke, C.; Martins, R.N.; Masters, C.L.; et al. Effect of amyloid on memory and non-memory decline from preclinical to clinical Alzheimer’s disease. Brain 2014, 137, 221–231. [Google Scholar] [CrossRef]
- Müller, M.J.; Greverus, D.; Weibrich, C.; Dellani, P.R.; Scheurich, A.; Stoeter, P.; Fellgiebel, A. Diagnostic utility of hippocampal size and mean diffusivity in amnestic MCI. Neurobiol. Aging 2007, 28, 398–403. [Google Scholar] [CrossRef]
- Nani, A.; Manuello, J.; Mancuso, L.; Liloia, D.; Costa, T.; Cauda, F. The Neural Correlates of Consciousness and Attention: Two Sister Processes of the Brain. Front. Neurosci. 2019, 13, 1169. [Google Scholar] [CrossRef]
- Ségonne F, Dale AM, Busa E, Glessner M, Salat D, Hahn HK, Fischl B. 2004. A hybrid approach to the skull stripping problem in mri. Neuroimage. 22(3):1060-1075. [CrossRef]
- Squire LR, Genzel L, Wixted JT, Morris RG. 2015. Memory consolidation. Cold Spring Harbor perspectives in biology. 7(8):a021766.
- Sugar, J.; Moser, M.-B. Episodic memory: Neuronal codes for what, where, and when. Hippocampus 2019, 29, 1190–1205. [Google Scholar] [CrossRef]
- Tulving, E. Précis of Elements of episodic memory. Behav. Brain Sci. 1984, 7, 223–238. [Google Scholar] [CrossRef]
- Weintraub D, Simuni T, Caspell-Garcia C, Coffey C, Lasch S, Siderowf A, Aarsland D, Barone P, Burn D, Chahine LM. 2015. Cognitive performance and neuropsychiatric symptoms in early, untreated parkinson's disease. Movement Disorders. 30(7):919-927. [CrossRef]
- Yarnall, A.J.; Breen, D.P.; Duncan, G.W.; Khoo, T.K.; Coleman, S.Y.; Firbank, M.J.; Nombela, C.; Winder-Rhodes, S.; Evans, J.R.; Rowe, J.B.; et al. Characterizing mild cognitive impairment in incident Parkinson disease: The ICICLE-PD Study. Neurology 2013, 82, 308–316. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Example of a typical trials of Experiment 1. ITI: Inter-Trial-Interval.
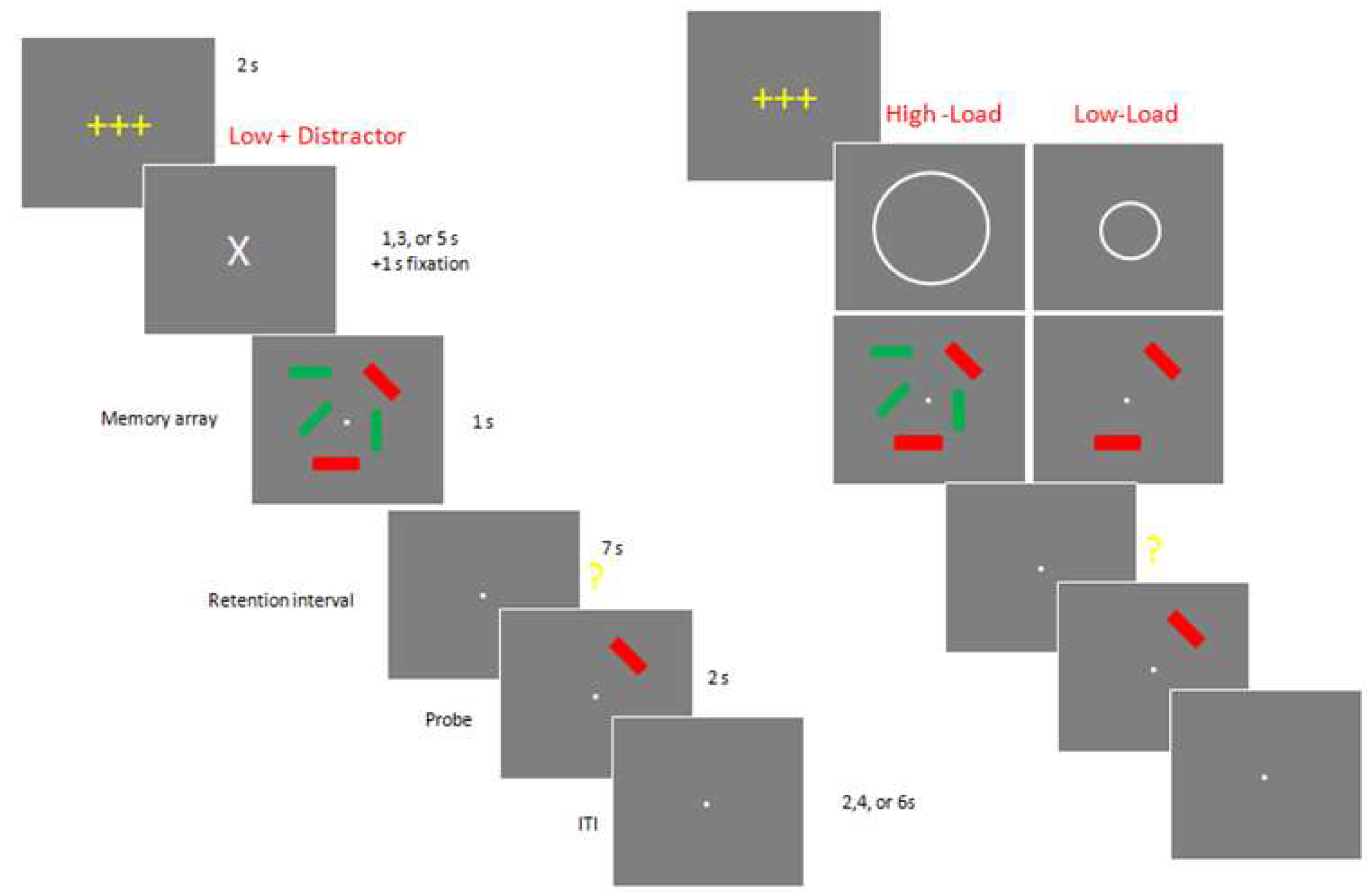
Figure 2.
Example of a typical trials of Experiment 2.

Figure 3.
Brain regions of interests.
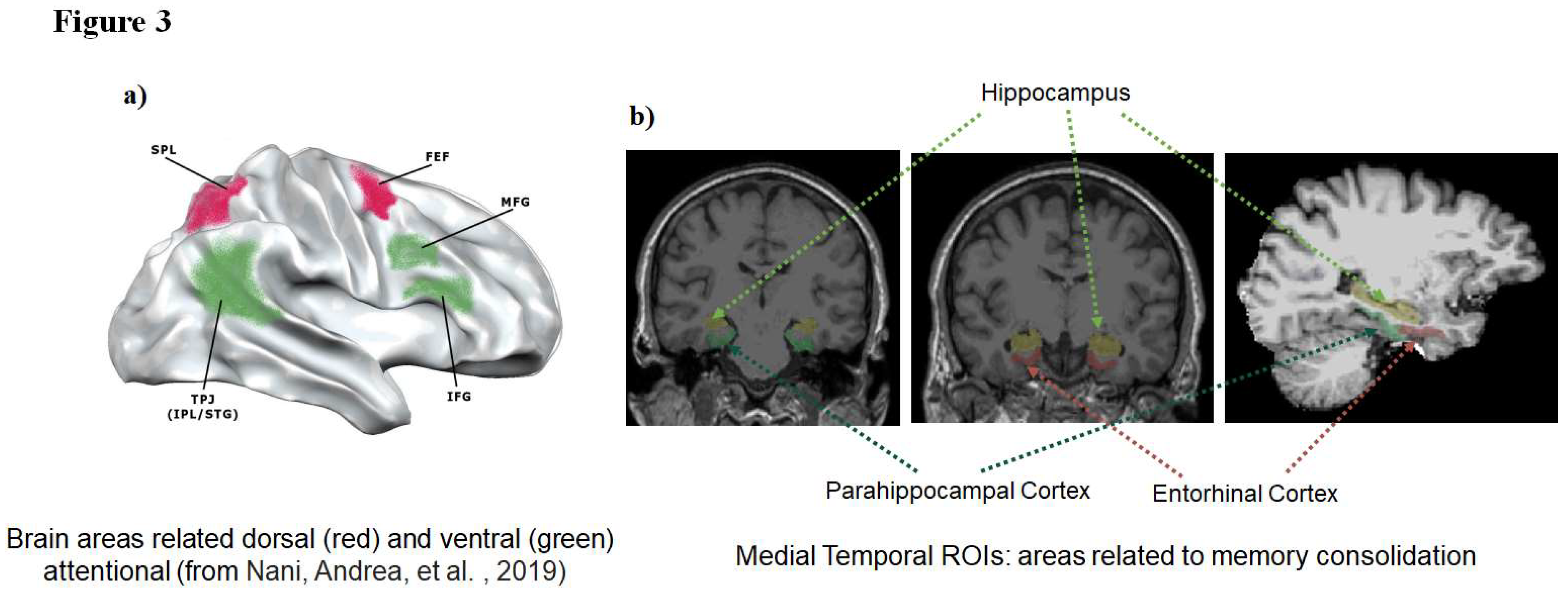
Figure 4.
Mean K scores for controls and PD patients in a) experiment 1 and b) experiment 2 depending on different memory array conditions: K = N * (H-FA), where N is the number of relevant, to-be-stored items, H is the hit rate and FA is the false alarm rate as suggested by Cowan (2001).
Figure 4.
Mean K scores for controls and PD patients in a) experiment 1 and b) experiment 2 depending on different memory array conditions: K = N * (H-FA), where N is the number of relevant, to-be-stored items, H is the hit rate and FA is the false alarm rate as suggested by Cowan (2001).
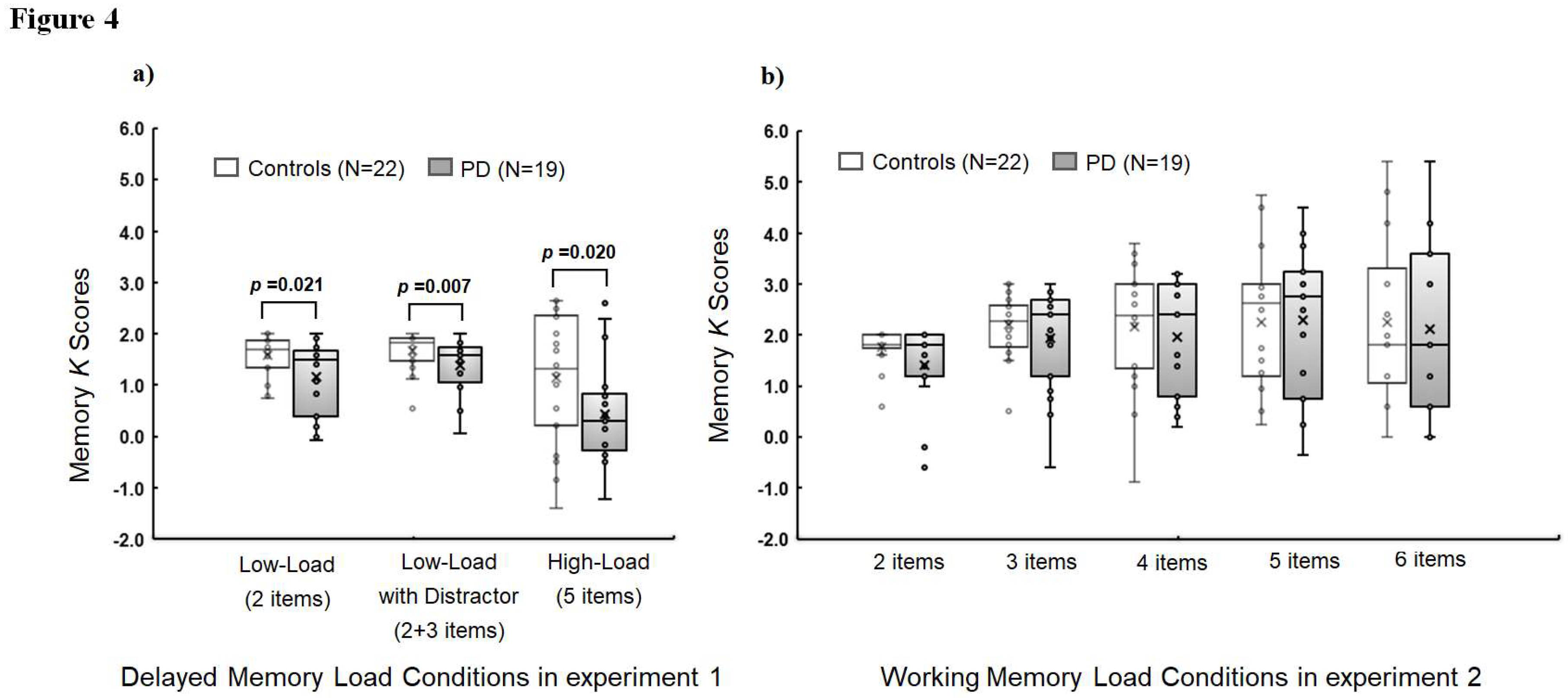
Figure 5.
Scatter plots show memory K scores (y-axis) in Low-Load (a), Low-Load with Distractor (b), and memory load of4 items (c) versus thickness values (mm) in the left superior temporal gyrus (x-axis) for controls and PD: K = N * (H-FA), where N is the number of relevant, to-be-stored items, H is the hit rate and FA is the false alarm rate as suggested by Cowan (2001).
Figure 5.
Scatter plots show memory K scores (y-axis) in Low-Load (a), Low-Load with Distractor (b), and memory load of4 items (c) versus thickness values (mm) in the left superior temporal gyrus (x-axis) for controls and PD: K = N * (H-FA), where N is the number of relevant, to-be-stored items, H is the hit rate and FA is the false alarm rate as suggested by Cowan (2001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



