
Submitted:
20 April 2023
Posted:
20 April 2023
You are already at the latest version
Abstract
Circular RNAs (circRNAs) served as covalently closed single-stranded RNAs have been proposed to influence plant development and stress resistance. Grapevine is the most economically valuable fruit crops cultivated worldwide and threaten by various abiotic stresses. Herein, we reported that a circRNA (Vv-circPTCD1) processed from the second exon of a pentatricopeptide repeat family gene PTCD1 was preferentially expressed in leaves and responded to salt and drought but not heat stress in grapevine. Additionally, the second exon sequence of PTCD1 was highly conserved but the biogenesis of Vv-circPTCD1 is species-dependent in plants. It was further found that the overexpressed Vv-circPTCD1 can slightly decreased abundance of the cognate host gene and the neighboring genes were barely affected in grapevine callus. Furthermore, we also successfully overexpressed the Vv-circPTCD1, and found that the Vv-circPTCD1 deteriorated the growth during heat, salt, and drought stresses in Arabidopsis. However, the biological effects on grapevine callus were not always consistent with that of Arabidopsis. Interestingly, we found that the transgenic plants of linear counterpart sequence also conferred the same phenotypes as that of circRNA during the three stress conditions, no matter what species. Those results imply that although the sequences are conserved, the biogenesis and functions of Vv-circPTCD1 are species-dependent. Our results indicate that the plant circRNA function investigation should be conducted in homologous species, which support valuable reference for further plant circRNA studies.
Keywords:
Grapevine
; Abiotic stress
; Non coding RNA
; CircRNA
; Pentatricopeptide repeat proteins
; Back-splicing
1. Introduction
Noncoding RNAs have increasingly crucial functions for growth and development and stress response in plants [1,2]. Circular RNAs (CircRNAs), which has not been found the coding ability in plants yet, are single strand non-coding RNA that are processed by backing-splicing, in which a downstream 5׳ splice site is linked by 3′,5′- phosphodiester bond to an upstream 3′ splice site in a reverse order crossing one exon or exons [3]. Owing to the development of high-throughput sequencing technology and associated analytical tools, thousands of circRNAs have been identified in various plants and were deposited in associated database [4,5]. At present, it has been reported that a total of 171,118 circRNAs from 21 plant species have been collected in PlantcircBase [6]. The biological functions of circRNA have been well investigated in mammals. Animal circRNAs have generally been suggested to function as miRNA sponges for decaying endogenous miRNA [7], as templates for translation [8,9], as mediator for regulating assembly and activation of AMPK complex under stress [10], as enhancer for cis-regulation of their host genes [11]. In plants, previous studies have focused on identification and annotation of putative circRNAs as well as function investigation. Until now, many studies have provided direct evidences that circRNAs shown important functions in plants by overexpression and gene editing technologies. Overexpression of circRNA derived from PSY1 (Phytoene Synthase 1) and PDS (Phytoene Desaturase) in tomato, respectively, causes color variation [12]. In Arabidopsis, circSEP3 from exon 6 of SEPALLATA3 can regulates alternative splicing of its host genes through R-loop formation with homological genomic region. Overexpression of this circRNA results in floral organ abnormality [13]. CRISPR-Cas9 strategy has been applied to remove Os06circ02797 processing by editing the flanking sequence in rice, resulted in a rapid growth phenotype and higher chlorophyll A/B content [14]. In Populus tomentosa, overexpression of Circ_0003418 derived from a RING-type E3 ligase gene XBAT32 deteriorates the callus thermotolerance [15]. Although some studies have been performed, functional investigation on circRNAs in plants is still very limited. Therefore, additional researches are further conducted.
Grapevine (Vitis spp.) is the most economically important fruit crops cultivated worldwide, for the purpose of producing table grape, dried fruits, and as the raw materials for juice and wine making [16]. The growth, development and flavor of berries are strongly influenced by various abiotic stresses, such as high temperature, water deficit, and salt during viticulture 17,18,19]. Previous findings suggested that high temperature influences grapevine photosynthesis, florescence and veraison [20,21,22]. In grapevine, the serine/arginine-rich splicing factors (SR) are generally phosphorylated on the condition of high temperature, which results in the disturbance of alternative splicing of pre-mRNA [23,24]. The mechanisms involved in grapevine tolerance to high temperature are poorly understood, although the heat shock factors (HSFs)-mediated regulatory networks have been well deciphered in other plants [25,26,27]. Drought is another important environment factor limiting grapevine growth and productivity, and studies have shown that the photosynthesis of vegetative organs, berries size and yield, and accumulation of secondary metabolites are greatly affected by the water deficit [28]. The abscisic acid (ABA) is induced and plays a key role for response to water deficit by regulating stomatal movements, as well as the signaling pathway has been well studied in plants [29]. Furthermore, the long non coding RNAs (LncRNA) and microRNAs (miRNA) are also serves as novel regulators of the plant response to drought stress in Arabidopsis [30], Brassica napus L [31], and grapevine [32]. Salt stress is another factor which is mainly due to the hyperosmotic conditions of soil solution with increase of Na+ and Cl-, causing the deficiency of the absorption of water and nutrients in plants [33]. In grapevine, previous studies found that many stress-induced genes, such as VvWRKY2, VvNAC17, and VvASMT1, perform functions for enhancing salt and osmatic stresses tolerance in ectopic overexpression [19,34,35]. Although the above findings greatly uncover the molecular mechanisms responding to abiotic stress in grapevine, the function of circRNAs also need to be further explored.
In grapevine, according to our previous study, a total of 8, 354 circRNAs have been identified by the whole transcriptome analysis, and a Vv-circATS1 spliced from glycerol-3-P acyltransferase was proved to improve cold tolerance in Arabidopsis [36]. In present study, a circRNA derived from the second exon of the pentatricopeptide repeat domain-containing protein 1 (PTCD1, VIT_211s0016g03830) was verified based on the grapevine circRNA dataset, named Vv-circPTCD1. Pentatricopeptide repeat (PPR) proteins is a large gene family in plants, which are characterized by 2 to 15 tandem repeats of 30-40 amino acid length motif. Some PPR proteins have been supposed to play roles in organellar RNA metabolism, organ development as well as stress response [37]. Overexpression of Vv-circPTCD1 deteriorated the tolerance to heat, salt, and PEG-mediated osmotic stresses in Arabidopsis. While, similarly, the linear fragment identical with the circRNA sequence also did. However, the Vv-circPTCD1 overexpressed callus mass of ‘Thompson Seedless’ were failure to display the phenotypes consistent with Arabidopsis. Those results implied that the functions of Vv-circPTCD1 potentially were sequence-dependent and not conserved among plants.
2. Results
2.1. Identification and verification of Vv-circPTCD1
Based on the previous published circRNA dataset of ‘Muscat Hamburg’ [36], a circRNA_4363, back-spliced from the second exon of a pentatricopeptide repeat (PPR) proteins family gene PTCD1, was identified and renamed Vv-circPTCD1 in this research. The PTCD1 encodes a protein with 636 amino acid residues length and contains the 9 tandem PPR domain (Figure 1A). In Arabidopsis genome, meanwhile, two orthologs (AT5G21222 and AT5G25630) were characterized and phylogenetic analysis showed that AT5G21222 and PTCD1 (VIT_211s0016g038030) were clustered into one clade with 96% confidence (1000 replicates) (Figure 1B). The Vv-circPTCD1 were further verified by cloning and sequencing as well as the back-splicing site was AG/GA (Figure 1C). Divergent and convergent primers were designed to amplify the target fragment form DNA and leaf cDNA, result showed that the fragment with the back-splicing site was only detected in cDNA templates (Figure 1D).
2.2. Nonconservative back-splicing of Vv-circPTCD1 in plants
To investigate whether the Vv-circPTCD1 was conserved in plants, we analyzed the physical gene structure of PTCD1 in plants, including monocots and dicots. The result showed that similar physical gene structure of PTCD1 orthologs was observed (Figure S1), as well as a CDS (coding sequence) back-spliced into Vv-circPTCD1 in grapevine was found and it was highly conserved by multiple sequence alignment among plants (Figure S2). It was reasonably hypothesized that this circRNA orthologous with Vv-circPTCD1 maybe also conserved in plants. Therefore, the divergent primer pair was tentatively designed to clone the orthologs in Arabidopsis and N. benthamiana, although the non-specific amplification was found in N. benthamiana, while the Vv-circPTCD1 orthologs were failure to cloned (Figure S3). Those results suggested that the back-splicing of Vv-circPTCD1 potentially was species-dependent.
2.3. The expression analyses of PTCD1 and Vv-circPTCD1 under abiotic stress
To investigate the potential functions in grapevine, we analyzed the tissue specificity and the expression patterns of Vv-CircPTCD1 and its corresponding host gene under salt, heat, and drought stress conditions, respectively. The Vv-CircPTCD1 was mainly expressed in young and mature leaves while scarcely expressed in roots, consistent with that of host gene (Figure 2A). However, the accumulation level of host gene was excessively higher than that of Vv-CircPTCD1 (more than sixty times) (Figure 2A). During salt stress, in addition to the host gene was mildly down-regulated during initial treatment, the Vv-CircPTCD1 was significantly down-regulated on the fifth day after treatment (Figure 2B, C). During drought treatment, the VvSnRK2I (VIT_207s0197g00080), a marker gene homologous with AtSRK2I/AtSnRK2.3 induced by ABA in Arabidopsis [42], was significantly induced when the soil water potential reduces to -0.8 and -1.0 MPa (Figure 2D). The expression level of PTCD1 was mildly up-regulation when the water potential was -0.4 MPa, while the expression level of PTCD1 was significantly reduced when the water potential was reduced to -0.8 and -1.0 MPa (Figure 2E). However, the expression pattern of Vv-CircPTCD1 was barely changed during drought treatment (Figure 2F). We further investigated the expression pattern during heat stress, the results showed that the heat shock factor genes VvHsfA7, as a marker of heat stress [20], were remarkable up-regulation (Figure 2G) and the expression level of PTCD1 was down-regulation with the prolongation of treatment (Figure 2H). Although the expression level of Vv-CircPTCD1 was down-regulation after 4 h treatment, however, the expression level was also slightly up-regulation after 2 h treatment (Figure 2I). Those results indicated that the Vv-CircPTCD1 expression was always fell behind that of PTCD1 and potentially play a role during abiotic stresses in grapevine.
2.4. Overexpression of Vv-CircPTCD1 in grapevine callus and Arabidopsis
To decipher the functions of Vv-CircPTCD1 in plants, the vector for Vv-CircPTCD1 overexpression (OE) was generated, meanwhile, corresponding linear fragment (Linear_PTCD1) identical with Vv-CircPTCD1 was also constructed as a control (Figure 3A). The overexpressed callus mass was verified by RT-PCR and RT-qPCR, respectively (Figure 3B, C, D, E). The expression level of host gene and flanking genes were further investigated in all OE callus mass. Among Vv-CircPTCD1 lines, the expression level of host gene was slightly down-regulated (Figure 3F), while the up-stream gene (VIT_211s0016g03850) and down-stream gene (VIT_211s0016g03820) were not significantly affected (Figure 3G, H). However, among Linear_PTCD1 OE lines, the expression level of host gene was discrepant among different lines, which is down-regulated in line 2 (Figure 3I). As well as the up-stream gene and down-stream gene were also slightly down-regulated (Figure 3J, K). In order to uncover whether the function of Vv-CircPTCD1 is conserved among plants, simultaneously, ectopic transformation was also performed in Arabidopsis (Figure S4).
2.5. Phenotype of OE lines under heat stress
To investigate whether Vv-CircPTCD1 plays a role in Arabidopsis, the four-weeks old seedlings were treated 12 h at 45℃ followed by exposure to 22 ℃ for 7 days to recovery. Results showed that, comparing with WT, the rosette leaves were significantly deteriorated in Vv-CircPTCD1 OE lines (Figure 4A,B). It was a wonder that the rosette leaves were also remarkably damaged in linear fragment OE lines, which keep an identical sequence with Vv-CircPTCD1 (Figure 4A, B). We also observed the phenotype of transgenic callus responding to heat stress, in which the callus was continuously incubated 15 d at 40℃. However, phenotypic differences were nonevent, no matter which OE callus mass (Figure 4C and Figure S5).
2.6. Phenotype of OE lines under salt stress
To further survey the function of Vv-CircPTCD1 in plants, the four days old seedlings of Arabidopsis were transplanted into normal and NaCl-containing medium to observe the variation. Results suggested that different OE lines and WT seedlings were normally and identically grow in NaCl-free medium (Figure 5A), while the survival rate of transgenic lines were lower on the NaCl-containing medium after five days compared to WT (Figure 5B, C). In grapevine callus, the OE callus mass transformed linear fragment adversely displayed albino phenotype, while the OE callus mass transformed Vv-CircPTCD1 was also did but much slighter compared to WT callus mass (Figure 5D). Those results also showed the Vv-CircPTCD1 potentially served as negative factor for salt stress and the linear fragment identical with the Vv-CircPTCD1 also plays the same roles in Arabidopsis and grapevine callus.
2.7. Phenotype of OE lines under drought stress
To comprehensively explore the function of Vv-CircPTCD1 in Arabidopsis, the four weeks old seedlings of Arabidopsis were stopped watering. Results showed that the transgenic lines were much more susceptible and withering (Figure 6A). The four days old seedlings were further transplanted into normal and PEG-containing medium, and found that different transgenic lines and WT seedlings were normally and identically grow in normal medium (Figure 6B), while the growth of rosette leaves were obviously inhibited in OE lines but the growth of root barely did (Figure 6C, D, E). Those results suggested that the Vv-CircPTCD1 and corresponding linear fragment also play negative roles during drought stress in Arabidopsis. In grapevine callus, oppositely, the OE callus mass of transgenic callus mass significantly displayed resistance to osmotic stress mediated by PEG-8000, while the WT has become brawn after 30 days incubation (Figure 6F). We further counted the growth index of callus mass, and found that the growth rate of transgenic callus was significantly higher than that of WT (Figure 6G and Figure S5). Those results implied that the function of Vv-CircPTCD1 potentially is species-dependent.
3. Discussion
CircRNAs are widespread in plants. Most circRNAs are generated by backing-splicing from the exon or exons of genic primary mRNA, for example 94.5% are derived from the coding genes in rice and the remaining were intergenic circRNAs [43]. In addition, the mitochondrion genome is also proved to generate circRNAs, called mitochondrion-encoded circular RNAs (mcircRNAs) [44]. Here, we identified a circRNA spliced from the second exon of PTCD1, a PPR family genes, in which it was spliced based on canonical GU/AG splicing signal [36] (Figure 1C). In general, hundreds of this superfamily proteins encoded by the PPR family genes located in the nuclear and organelle genome are found in plants and characterized by multiple tandem arrays of 35 residues PPR domain [45]. Previous finding proved that the PPR proteins, served as RNA binding protein, affect cytoplasmic male sterility, embryogenesis, seed development, and stress [46,47,48,49]. Interestingly, the physical gene structure of PTCD1 orthologs are similar, as well as the sequence of the CDS back-spliced into Vv-circPTCD1 are highly conserved among plants (Figure S1 and S2). However, the verification failure of corresponding circRNA in Arabidopsis and N. benthamiana imply that the circPTCD1 is not naturally conserved among plants. No matter what mammals and plants, a factually small proportion of circRNAs are supposed to be conserved, such as the overlap of 26% circRNAs in both human and mouse based on the syntenic region of genome [8]. In plants, only 8.7% circRNAs are genomic conservation in dicotyledonous plants, even that are less than half in Oryza genus [43]. Taken together, the Vv-circPTCD1 is not evolutionarily conserved, although the host genes are conserved. And this result partially supports the idea that circRNAs result largely from splicing errors [50,51].
We also successfully constructed the artificial vector containing a reverse complementary sequence pairs to generate the circRNA in Arabidopsis and grapevine callus according to the previous strategy, although the natural back-splicing of Vv-circPTCD1 is not conserved (Figure 3A) [36]. The intronic complementary sequences have been found in the flanking sequence of highly expressed circRNA in eukaryotes. In mammals, the exons flanked by the orientation-opposite Alu elements, a shorted interspersed elements in primates, are preferentially circularized [52]. In Populus tomentosa, the miniature inverted repeat transposable elements (MITEs) are enriched in circRNA flanking regions and served as cis-regulatory factor to significantly regulate the exon circularization [15]. Furthermore, previous finding supposed that the back-splicing of circRNA is also executed by the canonical spliceosome, a highly conserved precursor mRNA splicing mechanism in eukaryotes [53]. Additionally, the flanking intron sequence is crucial adjective for precise splicing and circularization of one exon and exons, which has been proved by the heterologous investigations [36,54]. Therefore, the inverted repeats and flanking sequence are commonly contributed to the fidelity and expression level of Vv-CircPTCD1 in Arabidopsis and grapevine callus (Figure 3B, C and Figure S4). Therefore, the artificial “stem-loop” structure mediated by the reverse complementary sequence greatly contributes to the back-splicing of Vv-CircPTCD1 in heterogeneous expression, in which the “stem-loop” brings the splice sites into close proximity to each other [55].
Additionally, previous investigations uncovered that circRNAs are incorporated into chromatin by R-loop structure interacting with the nuclear DNA to regulate the chromatin stabilization or alternative splicing [13,56]. The host gene and neighboring genes are potentially influenced thorough overexpressed or deleted circRNA. In rice, a neighboring gene is significantly up-regulated at circRNA mutant lines [14]. In this study, the Vv-CircPTCD1 has slightly negative effect on the host gene, and barely do on the neighboring genes after overexpression of Vv-CircPTCD1 (Figure 3F, G, H). In P. tomentosa, overexpression of Circ_0003418 significantly increased the expression abundance of its parental gene [15]. Therefore, the circRNA potentially have the ability to function through disturbing the expression of host or neighboring genes in plants.
The expression pattern of circRNAs is also determined by tissue and environment factors, and even genotype [15,57]. We found that the Vv-CircPTCD1 focused in this research were preferentially expressed in young and mature leaves, which is consistent with the expression pattern of corresponding host gene, while the expression abundance of Vv-CircPTCD1 greatly less than 2% that of host gene (Figure 2A). In addition to some circRNAs in animals (almost 10%) [8], the circRNAs barely hold a higher expression level with respect to the linear counterpart and are largely obeyed the expression pattern of host gene in plants, such as grapevine [36]. In this study, the expression level of PTCD1 is significantly inhibited during drought stress, while the expression of Vv-CircPTCD1 is slightly decreased (Figure 2D, E, F), indicating it plays a role in grapevine responding to drought stress. In addition, the expression level of Vv-CircPTCD1 is remarkably decreased during salt stress after 5 days, while the host gene never occurred (Figure 2B, C). Those results imply that the Vv-CircPTCD1 plays a role during abiotic stress, in which the expression of circRNA potentially is controlled on the condition of specific abiotic stress, at least during the processing or degradation pathway [58].
The hypothesis of “splicing error” is indeed uncontradictory with the biochemical activities and biological functions of circRNAs. According the previous idea, the biogenesis of circRNAs and the regulation of their abundance are managed by various mechanisms, such as cis- or trans-elements, export and turnover of circRNAs [3]. In this study, although the expression of Vv-CircPTCD1 is related to the salt and drought stress, the mechanism is also unclear. For investigation of the biological function, a problem with their circular conformation and sequence overlap with linear cognate mRNAs and incomplete cyclization is difficult to overcome [53]. The strategy to evade this problem is to transform the linear fragment identical with circRNA as a control. Generally, the linear fragment scarcely functions in plants [13,15,36]. In present research, the linear counterpart is also transformed as a control (Figure 3D, E and Figure S4). Interestingly, identical stress-induced phenotype is observed between Vv-CircPTCD1 and its linear counterpart OE lines (Figure 4, Figure 5 and Figure 6). It is putative that (1) the incompletely cyclized transcript plays roles identical with linear counterpart; (2) the Vv-CircPTCD1 functions through sequence-dependent style. We observed that only the phenotype induced by salt stress is conserved between Arabidopsis and grapevine callus (Figure 5), and that is distinctly inconsistent during the treatment of heat and PEG (Figure 3 and Figure 6). Coincidentally, although the cognate sequence generated Vv-CircPTCD1 are greatly conserved among multiple species (Figure S2), while the biogenesis is not conserved (Figure S3). Those results imply that the biological functions of circRNA are potentially species-dependent in plants.
4. Materials and methods
4.1. Plant materials and treatments
For drought treatment of grapevine, 1-year-old ‘Muscat Hamburg’ cuttings were planted in a mixture of loam and sand (1:1, v/v) in flowerpot in a greenhouse. Cuttings with four well-developed leaves were drought stress treated. The well-developed leaves were collected when the soil water potential was 0 MPa, -0.2 MPa, -0.4 MPa, -0.6 MPa, -0.8 MPa, and -1.0 MPa, respectively. For heat stress treatment of grapevine, the cuttings were treated at 45 ℃, and the leaves were collected at 0 h, 1 h, 2 h, and 4 h, respectively. For salt stress, the cuttings were planted in a chamber (25 ℃) and watered the 300 mM NaCl solution, and the mature leaves were sampled at 0 d, 1 d, 2 d, 3 d, and 5 d. All samples were quickly frozen in liquid nitrogen and then stored at -80 ℃. All treatments were carried out in three biological replicates.
Arabidopsis ecotype Col-0 was cultured in chamber under 16 h light/8 h dark conditions with a light intensity of 600 mmol m-2 s-1 at a temperature of 22°C. For heat stress treatment of Arabidopsis, 6 weeks-old WT and transgenic plants were exposed to heat stress at 45◦C for 12 h followed by exposure to 22◦C for 7 days to recovery. For drought stress treatment, 6-weeks-old WT and transgenic plants were stopped watering until the substrate is completely dry, and observed the phenotype. The PEG-8000 treatment (the water potential was -0.7 MPa) was performed according to the Verslues׳s descriptions [38]. And the 4 days old seedlings were transferred into the plate containing PEG-8000 and cultured in chamber for another 5 days. For salt treatment, the plates containing 1/2 MS salts supplemented with 300 mM NaCl were prepared and 4 days old seedlings were incubated. The survival rates were counted after 5 days.
4.2. Validation of circRNA and RT-qPCR
The total RNA of Arabidopsis and grapevine callus was extracted by TRIzol reagent (Sangon Biotech, China). And the total RNA of grapevine leaves was extracted by CTAB method. In brief, 200 mg sample are powered and transferred into RNase-free 2 mL tube. Added quickly 900 μL pre-heated (65 ℃) CTAB buffer (2% CTAB, 100 mM Tris-HCl, 20 mM EDTA, 2 M NaCl, 2% PVP-40, pH8.0) and 20 μL β-mercaptoethanol. Mixed the mixture robustly and then bathed at 65℃ for 15 min. Extracted lysate with 900 uL of chloroform: isoamylol (24:1, v/v) by vortexing for 30 s and centrifuged at 4 ℃ for 10 min at 12, 000 rpm. Repeated two times with chloroform: isoamylol extraction. Added 20 μL NaAC (pH5.2) and 60 μL pre-cooled (-20℃) absolute alcohol, and bathed 10 min in ice. And added volume chloroform: isoamylol for vortexing. After centrifuged at 4℃ for 10 min at 12,000 rpm, added 150 μL 10 M LiCl to 450 μL supernatant and mixed. The RNA was precipitated at 4℃ for 6 to 8 h, and harvested by centrifugation at 4℃ for 10 min at 12,000 rpm. Carefully removed the supernatant, and washed two times by 70 % pre-cooled (-20℃) alcohol. The pellet was dried and resuspended by 30 μL DEPC-treated ddH2O.
The total RNA was transcribed to cDNA using random primers by a FastKing RT Kit (TIANGEN, China) in accordance with the manufacturer’s instructions. To confirm the grapevine circRNAs predicted, divergent primer was designed by Primer 5 (Table S1). PCR was as follows: 94°C for 3 min; 40 cycles at 94°C for 30 s, 56°C for 15 s, and 72°C for 20 s; and then 1 cycle at 72°C for 5 min. For PCR, 2×Taq master mix (Vazyme, China) was used. The PCR products were separated by agarose gel electrophoresis and then purified. Sanger sequencing was further performed to verify the back-spliced junction sites. RT-qPCR (real-time quantitative PCR) analysis was conducted to evaluate the expression levels of circRNAs and linear counterparts using Talent qPCR PreMix (TIANGEN, China) with a qTOWER3 Real-Time PCR Detection System (Analytikjena, Germany). Gene expression levels were calculated by the 2-∆∆Ct method [39].
4.3. Vector construction
All expression vectors were constructed based on the pHB binary plasmids. The construction of circRNA expression vector was conducted based on previous published strategy [36]. For linear RNA expression vector, the linear fragment derived from the same sequence with circRNA was cloned into pHB vector which was digested by BamH І and Xba І. The constructed plasmids were individually transformed into Agrobacterium tumefaciens GV3101 strains and incubated at 28℃ for 2-3 d on the LB medium supplemented with 25 mg·L-1 rifampicin (Rif) and 50 mg·L-1 kanamycin (Kan). A single clone was incubated at 200 rpm and 28℃ for 8-10 h in LB liquid medium containing 25 mg·L-1 Rif and 50 mg·L-1 Kan. The presence of the corresponding plasmid was verified by PCR. The positive clones were added glycerol and stored at -80℃.
4.4. Arabidopsis transformation and verification
Arabidopsis transformation was performed according to floral dip method [40]. T1 seedling were selected by spraying of Glufosinate ammonium 10% solution (dilute 1000×) (Sangon Biotech, China). The positive transgenic ones were transferred into pots and confirmed by PCR detection using the gene primers. For circRNAs confirmation, the cDNA was used as templates for PCR and RT-qPCR reaction and the splicing site was verified by further sequenced.
4.5. Callus transformation of ‘Thompson Seedless’ and treatment
The embryogenic callus was induced from floral explants of ‘Thompson Seedless’ according to a previously published protocol [41]. The callus mass was monthly sub-cultured in MSTP medium (MS base salts, 20 g·L-1sucrose, 1 mg·L-1 TDZ, 2.2 mg·L-1 picloram, pH5.8) in dark condition at 26℃. For callus transformation, the A. tumefaciens containing vector was inoculated in 20 mL LB liquid medium and incubated until OD600 reached 1.0. After centrifugation for 5 min at 6000 rpm and 25℃, the medium was discarded. The pellet was resuspended and the OD600 was adjusted to 0.4 with MS medium (0.1 mM acetosyringone, pH5.8). The bacterial mixture was incubated with the embryogenic callus for 30 min at 40 rpm in 50 mL conical flask, and was then transferred onto filter paper to fully remove and evaporate excess liquid. Then, the callus was collected and transferred to solid MS medium (0.1 mM acetosyringone, 8g·L-1 agar, pH5.8) and co-cultivated in dark condition at 26 °C for 3 days. After 3 days, the callus was transferred to solid MSTP medium containing 50 mg·L-1 hygromycin, 200 mg·L-1 Timentin. Petri dishes were monthly sub-cultured in the dark for the induction of transgenic callus. The expression of circRNA in transgenic callus was confirmed by PCR and RT-qPCR using divergent primer pairs.
For heat stress treatment, the callus mass was sub-cultured into MSTP medium for one week at 26 ℃ and then further cultured at 40 ℃ for 15 days. The callus mass was sub-cultured into MSTP medium supplemented with 300 mM NaCl for salt stress at 26 ℃. For PEG-8000 treatment, the plates were prepared according to above description, of which the base salt was replaced by MSTP medium supplemented with 15 g·L-1 agar on solid layer but the liquid layer was the MSTP medium without sucrose and agar. Then the callus mass was sub-cultured into the PEG-containing plates and cultured at 26 ℃ for one month.
5. Conclusions
According to this result, Although the cognate counterpart mRNA is greatly conserved among plants, the biogenesis of circRNA generated from them potentially is not. Furthermore, the biological effects of circRNA are largely species-dependent.
Author Contributions
CM designed the experiments and revised the manuscript; YR performed the callus transformation, phenotype collection, and drafted the manuscript; JL and ZZ conducted the callus induction and sub-culture; YS, DF, ML, LZ, YX, and DG performed sample treatment, Arabidopsis transformation, and vector construction; JH, SS, and ZG contributed to the data collection and analyses.
Funding
This work was supported by the China Postdoctoral Science Foundation (2021M692102), the National Natural Science Foundation of China (Grant No. 32122076, 32202446), and the earmarked fund for CARS-29.
Data Availability Statement
No new datasets were generated or analyzed in this study.
Acknowledgments
Not applicable.
Conflicts of Interest
No conflicts of interest declared.
References
- Wang, Y.; Yang, M.; Wei, S.; Qin, F.; Zhao, H.; Suo, B. Identification of circular RNAs and their targets in leaves of Triticum aestivum L. under dehydration stress. Front Plant Sci 2016, 7, 2024. [CrossRef]
- Zhou, Y.; Wang, X.; Qi, K.; Bao, J.; Zhang, S.; Gu, C. Involvement of long non-coding RNAs in pear fruit senescence under high- and low-temperature conditions. Hortic Plant J 2022, 198, 112251. [CrossRef]
- Chen, L. L. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol 2020, 21, 475-490. [CrossRef]
- Chu, Q.; Zhang, X.; Zhu, X.; Liu, C.; Mao, L.; Ye, C.; Zhu, Q. H.; Fan, L. PlantcircBase: a database for plant circular RNAs. Mol Plant 2017, 10, 1126-1128. [CrossRef]
- Zhang, P.; Meng, X.; Chen, H.; Liu, Y.; Xue, J.; Zhou, Y.; Chen, M. PlantCircNet: a database for plant circRNA-miRNA-mRNA regulatory networks. Database (Oxford) 2017, 2017, bax089. [CrossRef]
- Chu, Q.; Bai, P.; Zhu, X.; Zhang, X.; Mao, L.; Zhu, Q. H.; Fan, L.; Ye, C. Y. Characteristics of plant circular RNAs. Brief Bioinform 2020, 21, 135-143. [CrossRef]
- Hansen, T. B.; Jensen, T. I.; Clausen, B. H.; Bramsen, J. B.; Finsen, B.; Damgaard, C. K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384-8. [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; Laneve, P.; Rajewsky, N.; Bozzoni, I. Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis. Mol Cell 2017, 66, 22-37. [CrossRef]
- Yang, Y.; Gao, X.; Zhang, M.; Yan, S.; Sun, C.; Xiao, F.; Huang, N.; Yang, X.; Zhao, K.; Zhou, H.; Huang, S.; Xie, B.; Zhang, N. Novel role of FBXW7 circular RNA in repressing glioma tumorigenesis. J Natl Cancer Inst 2018, 110, 304-315. [CrossRef]
- Li, Q.; Wang, Y.; Wu, S.; Zhou, Z.; Ding, X.; Shi, R.; Thorne, R. F.; Zhang, X. D.; Hu, W.; Wu, M. CircACC1 regulates assembly and activation of AMPK complex under metabolic stress. Cell Metab 2019, 30, 157-173. [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; Zhu, P.; Chang, Z.; Wu, Q.; Zhao, Y.; Jia, Y.; Xu, P.; Liu, H.; Shan, G. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol 2015, 22, 256-264. [CrossRef]
- Tan, J.; Zhou, Z.; Niu, Y.; Sun, X.; Deng, Z. Identification and functional characterization of tomato circRNAs derived from genes involved in fruit pigment accumulation. Sci Rep 2017, 7, 8594. [CrossRef]
- Conn, V. M.; Hugouvieux, V.; Nayak, A.; Conos, S. A.; Capovilla, G.; Cildir, G.; Jourdain, A.; Tergaonkar, V.; Schmid, M.; Zubieta, C.; Conn, S. J. A circRNA from SEPALLATA3 regulates splicing of its cognate mRNA through R-loop formation. Nat Plants 2017, 3, 17053. [CrossRef]
- Zhou, J.; Yuan, M.; Zhao, Y.; Quan, Q.; Yu, D.; Yang, H.; Tang, X.; Xin, X.; Cai, G.; Qian, Q.; Qi, Y.; Zhang, Y. Efficient deletion of multiple circle RNA loci by CRISPR-Cas9 reveals Os06circ02797 as a putative sponge for OsMIR408 in rice. Plant Biotechnol J 2021, 19, 1240-1252. [CrossRef]
- Song, Y.; Bu, C.; Chen, P.; Liu, P.; Zhang, D. Miniature inverted repeat transposable elements cis-regulate circular RNA expression and promote ethylene biosynthesis, reducing heat tolerance in Populus tomentosa. J Exp Bot 2021, 72, , 1978-1994. [CrossRef]
- Baroi, A. M.; Popitiu, M.; Fierascu, I.; Sardarescu, I. D.; Fierascu, R. C. Grapevine wastes: a rich source of antioxidants and other biologically active compounds. Antioxidants (Basel) 2022, 11, 393. [CrossRef]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine responses to heat stress and global warming. Plants (Basel) 2020, 9,1754. [CrossRef]
- Zhu, Z.; Quan, R.; Chen, G.; Yu, G.; Li, X.; Han, Z.; Xu, W.; Li, G.; Shi, J.; Li, B. An R2R3-MYB transcription factor VyMYB24, isolated from wild grape Vitis yanshanesis J. X. Chen., regulates the plant development and confers the tolerance to drought. Front Plant Sci 2022, 13, 966641. [CrossRef]
- Ju, Y. L.; Yue, X. F.; Min, Z.; Wang, X. H.; Fang, Y. L.; Zhang, J. X. VvNAC17, a novel stress-responsive grapevine (Vitis vinifera L.) NAC transcription factor, increases sensitivity to abscisic acid and enhances salinity, freezing, and drought tolerance in transgenic Arabidopsis. Plant Physiol Biochem 2020, 146, 98-111. [CrossRef]
- Zha, Q.; Xi, X. J.; He, Y. N.; Jiang, A. L. Transcriptomic analysis of the leaves of two grapevine cultivars under high-temperature stress. Sci Hortic 2020, 265, 109265. [CrossRef]
- Merrill, N. K.; García de Cortázar-Atauri, I.; Parker, A. K.; Walker, M. A.; Wolkovich, E. M. Exploring grapevine phenology and high temperatures response under controlled conditions. Front Env Sci 2020, 8, 516527. [CrossRef]
- Ryu, S.; Han, J. H.; Cho, J. G.; Jeong, J. H.; Lee, S. K.; Lee, H. J. High temperature at veraison inhibits anthocyanin biosynthesis in berry skins during ripening in 'Kyoho' grapevines. Plant Physiol Biochem 2020, 157, 219-228. [CrossRef]
- Jiang, J.; Liu, X.; Liu, C.; Liu, G.; Li, S.; Wang, L. Integrating omics and alternative splicing reveals insights into grape response to high temperature. Plant Physiol 2017, 173, 1502-1518. [CrossRef]
- Liu, G. T.; Jiang, J. F.; Liu, X. N.; Jiang, J. Z.; Sun, L.; Duan, W.; Li, R. M.; Wang, Y.; Lecourieux, D.; Liu, C. H.; Li, S. H.; Wang, L. J. New insights into the heat responses of grape leaves via combined phosphoproteomic and acetylproteomic analyses. Hortic Res 2019, 6, 100. [CrossRef]
- Haider, S.; Iqbal, J.; Naseer, S.; Yaseen, T.; Shaukat, M.; Bibi, H.; Ahmad, Y.; Daud, H.; Abbasi, N. L.; Mahmood, T. Molecular mechanisms of plant tolerance to heat stress: current landscape and future perspectives. Plant Cell Rep 2021, 40, 2247-2271. [CrossRef]
- Pillet, J.; Egert, A.; Pieri, P.; Lecourieux, F.; Kappel, C.; Charon, J.; Gomes, E.; Keller, F.; Delrot, S.; Lecourieux, D. VvGOLS1 and VvHsfA2 are involved in the heat stress responses in grapevine berries. Plant Cell Physiol 2012, 53, 1776-92. [CrossRef]
- Liu, G.; Chai, F.; Wang, Y.; Jiang, J.; Duan, W.; Wang, Y.; Wang, F.; Li, S.; Wang, L. Genome-wide identification and classification of HSF family in grape, and their transcriptional analysis under heat acclimation and heat stress. Hortic Plant J 2018, 4,133-143. [CrossRef]
- Gambetta, G. A.; Herrera, J. C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S. D. The physiology of drought stress in grapevine: towards an integrative definition of drought tolerance. J Exp Bot 2020, 71, 4658-4676. [CrossRef]
- Mao, H.; Jian, C.; Cheng, X.; Chen, B.; Mei, F.; Li, F.; Zhang, Y.; Li, S.; Du, L.; Li, T.; Hao, C.; Wang, X.; Zhang, X.; Kang, Z. The wheat ABA receptor gene TaPYL1-1B contributes to drought tolerance and grain yield by increasing water-use efficiency. Plant Biotechnol J 2022, 20, 846-861. [CrossRef]
- Qin, T.; Zhao, H.; Cui, P.; Albesher, N.; Xiong, L. A nucleus-localized long non-coding RNA enhances drought and salt stress tolerance. Plant Physiol 2017, 175, 1321-1336. [CrossRef]
- Li, J.; Duan, Y.; Sun, N.; Wang, L.; Feng, S.; Fang, Y.; Wang, Y. The miR169n-NF-YA8 regulation module involved in drought resistance in Brassica napus L. Plant Sci 2021, 313, 111062. [CrossRef]
- Pagliarani, C.; Vitali, M.; Ferrero, M.; Vitulo, N.; Incarbone, M.; Lovisolo, C.; Valle, G.; Schubert, A. The accumulation of miRNAs differentially modulated by drought stress is affected by grafting in grapevine. Plant Physiol 2017, 173, 2180-2195. [CrossRef]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J Integr Plant Biol 2018, 60, 796-804. [CrossRef]
- Yu, Y.; Ni, Y.; Qiao, T.; Ji, X.; Xu, J.; Li, B.; Sun, Q. Overexpression of VvASMT1 from grapevine enhanced salt and osmotic stress tolerance in Nicotiana benthamiana. PLoS One 2022, 17, e0269028. [CrossRef]
- Mzid, R.; Zorrig, W.; Ben Ayed, R.; Ben Hamed, K.; Ayadi, M.; Damak, Y.; Lauvergeat, V.; Hanana, M. The grapevine VvWRKY2 gene enhances salt and osmotic stress tolerance in transgenic Nicotiana tabacum. 3 Biotech 2018, 8, 277. [CrossRef]
- Gao, Z.; Li, J.; Luo, M.; Li, H.; Chen, Q.; Wang, L.; Song, S.; Zhao, L.; Xu, W.; Zhang, C.; Wang, S.; Ma, C. Characterization and cloning of Grape Circular RNAs Identified the Cold Resistance-Related Vv-circATS1. Plant Physiol 2019, 180,966-985. [CrossRef]
- Xing, H.; Fu, X.; Yang, C.; Tang, X.; Guo, L.; Li, C.; Xu, C.; Luo, K. Genome-wide investigation of pentatricopeptide repeat gene family in poplar and their expression analysis in response to biotic and abiotic stresses. Sci Rep 2018, 8, 2817. [CrossRef]
- Verslues, P. E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J. K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J 2006, 45, 523-39. [CrossRef]
- Livak, K. J.; Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-∆∆CT) Method. Methods 2001, 25, 402-408. [CrossRef]
- Clough, S. J.; Bent, A. F. Floral dip a simplified method for Agrobacterium -mediated transformation of Arabidopsis thaliana. Plant J 1998, 16, 9. [CrossRef]
- Gambino, G.; Ruffa, P.; Vallania, R.; Gribaudo, I. Somatic embryogenesis from whole flowers, anthers and ovaries of grapevine (Vitis spp.). Plant Cell Tiss Org 2007, 90, 79-83. [CrossRef]
- Fujita, Y.; Nakashima, K.; Yoshida, T.; Katagiri, T.; Kidokoro, S.; Kanamori, N.; Umezawa, T.; Fujita, M.; Maruyama, K.; Ishiyama, K.; Kobayashi, M.; Nakasone, S.; Yamada, K.; Ito, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Three SnRK2 protein kinases are the main positive regulators of abscisic acid signaling in response to water stress in Arabidopsis. Plant Cell Physiol 2009, 50, 2123-2132. [CrossRef]
- Chu, Q.; Ding, Y.; Xu, X.; Ye, C. Y.; Zhu, Q. H.; Guo, L.; Fan, L. Recent origination of circular RNAs in plants. New Phytol 2022, 233, 515-525. [CrossRef]
- Liao, X.; Li, X. J.; Zheng, G. T.; Chang, F. R.; Fang, L.; Yu, H.; Huang, J.; Zhang, Y. F. Mitochondrion-encoded circular RNAs are widespread and translatable in plants. Plant Physiol 2022, 51, 1-6. [CrossRef]
- Lurin, C.; Andres, C.; Aubourg, S.; Bellaoui, M.; Bitton, F.; Bruyere, C.; Caboche, M.; Debast, C.; Gualberto, J.; Hoffmann, B.; Lecharny, A.; Le Ret, M.; Martin-Magniette, M. L.; Mireau, H.; Peeters, N.; Renou, J. P.; Szurek, B.; Taconnat, L.; Small, I. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell 2004, 16, 2089-103. [CrossRef]
- Liu, Z.; Dong, F.; Wang, X.; Wang, T.; Su, R.; Hong, D.; Yang, G. A pentatricopeptide repeat protein restores nap cytoplasmic male sterility in Brassica napus. J Exp Bot 2017, 68, 4115-4123. [CrossRef]
- Sosso, D.; Canut, M.; Gendrot, G.; Dedieu, A.; Chambrier, P.; Barkan, A.; Consonni, G.; Rogowsky, P. M. PPR8522 encodes a chloroplast-targeted pentatricopeptide repeat protein necessary for maize embryogenesis and vegetative development. J Exp Bot 2012, 63, 5843-57. [CrossRef]
- Li, X.; Gu, W.; Sun, S.; Chen, Z.; Chen, J.; Song, W.; Zhao, H.; Lai, J. Defective Kernel 39 encodes a PPR protein required for seed development in maize. J Integr Plant Biol 2018, 60, 45-64. [CrossRef]
- Laluk, K.; Abuqamar, S.; Mengiste, T. The Arabidopsis mitochondria-localized pentatricopeptide repeat protein PGN functions in defense against necrotrophic fungi and abiotic stress tolerance. Plant Physiol 2011, 156, 2053-2068. [CrossRef]
- Xu, C.; Zhang, J. Mammalian circular RNAs result largely from splicing errors. Cell Rep 2021, 36, 09439. [CrossRef]
- Cocquerelle, C.; Mascrez, B.; Hétuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J 1993, 7, 6. [CrossRef]
- Zhang, X. O.; Wang, H. B.; Zhang, Y.; Lu, X.; Chen, L. L.; Yang, L., Complementary sequence-mediated exon circularization. Cell 2014, 159, 134-147. [CrossRef]
- Liu, C. X.; Chen, L. L. Circular RNAs: Characterization, cellular roles, and applications. Cell 2022, 185, 2016-2034. [CrossRef]
- Zhang, P.; Fan, Y.; Sun, X.; Chen, L.; Terzaghi, W.; Bucher, E.; Li, L.; Dai, M. A large-scale circular RNA profiling reveals universal molecular mechanisms responsive to drought stress in maize and Arabidopsis. Plant J 2019, 98, 697-713. [CrossRef]
- Liang, D.; Wilusz, J. E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev 2014, 28, 2233-2247. [CrossRef]
- Liu, Y.; Su, H.; Zhang, J.; Liu, Y.; Feng, C.; Han, F. Back-spliced RNA from retrotransposon binds to centromere and regulates centromeric chromatin loops in maize. PLoS Biol 2020, 18, e3000582. [CrossRef]
- Fan, J.; Quan, W.; Li, G. B.; Hu, X. H.; Wang, Q.; Wang, H.; Li, X. P.; Luo, X.; Feng, Q.; Hu, Z. J.; Feng, H.; Pu, M.; Zhao, J. Q.; Huang, Y. Y.; Li, Y.; Zhang, Y.; Wang, W. M. circRNAs Are Involved in the Rice-Magnaporthe oryzae Interaction. Plant Physiology 2020, 182, 272-286. [CrossRef]
- Liu, R.; Ma, Y.; Guo, T.; Li, G. Identification, biogenesis, function, and mechanism of action of circular RNAs in plants. Plant Commun 2022, 4, 100430. [CrossRef]
Figure 1.
Identification of Vv-circPTCD1 in grapevine. (A) The graphical representations of PTCD1 protein; (B) The phylogenetic analysis of PTCD1 (VIT_211s0016g038030) with orthologs in other species. (C) Verification of Vv-circPTCD1 processed in the second exon. (D) PCR amplification of divergent and convergent primers based on DNA and cDNA in grapevine. “►◄” indicated the convergent primer pairs, “◄►” indicated the divergent primer pairs.
Figure 1.
Identification of Vv-circPTCD1 in grapevine. (A) The graphical representations of PTCD1 protein; (B) The phylogenetic analysis of PTCD1 (VIT_211s0016g038030) with orthologs in other species. (C) Verification of Vv-circPTCD1 processed in the second exon. (D) PCR amplification of divergent and convergent primers based on DNA and cDNA in grapevine. “►◄” indicated the convergent primer pairs, “◄►” indicated the divergent primer pairs.
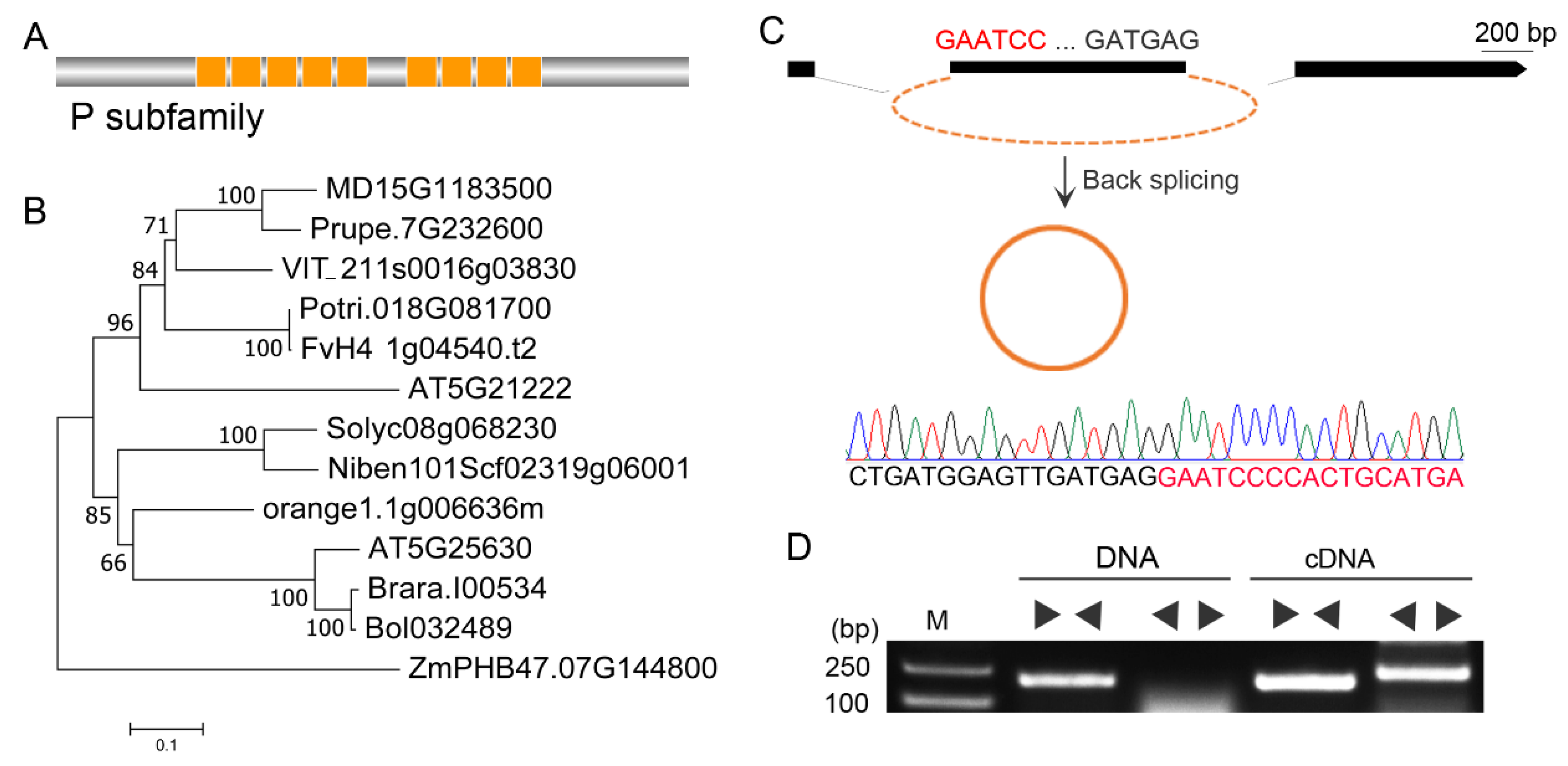
Figure 2.
The expression patterns of PTCD1 and Vv-CircPTCD1 under stress. (A) The tissue-specificity analyses of PTCD1 and Vv-CircPTCD1; (D, G) The expression pattern of marker genes VvSnRK2I and VvHsfA7 under drought and heat treatment; (B, E, H) The expression pattern of PTCD1 during salt, drought, and heat stress; (C, F, I) The expression pattern of Vv-CircPTCD1 during salt, drought, and heat stress.
Figure 2.
The expression patterns of PTCD1 and Vv-CircPTCD1 under stress. (A) The tissue-specificity analyses of PTCD1 and Vv-CircPTCD1; (D, G) The expression pattern of marker genes VvSnRK2I and VvHsfA7 under drought and heat treatment; (B, E, H) The expression pattern of PTCD1 during salt, drought, and heat stress; (C, F, I) The expression pattern of Vv-CircPTCD1 during salt, drought, and heat stress.
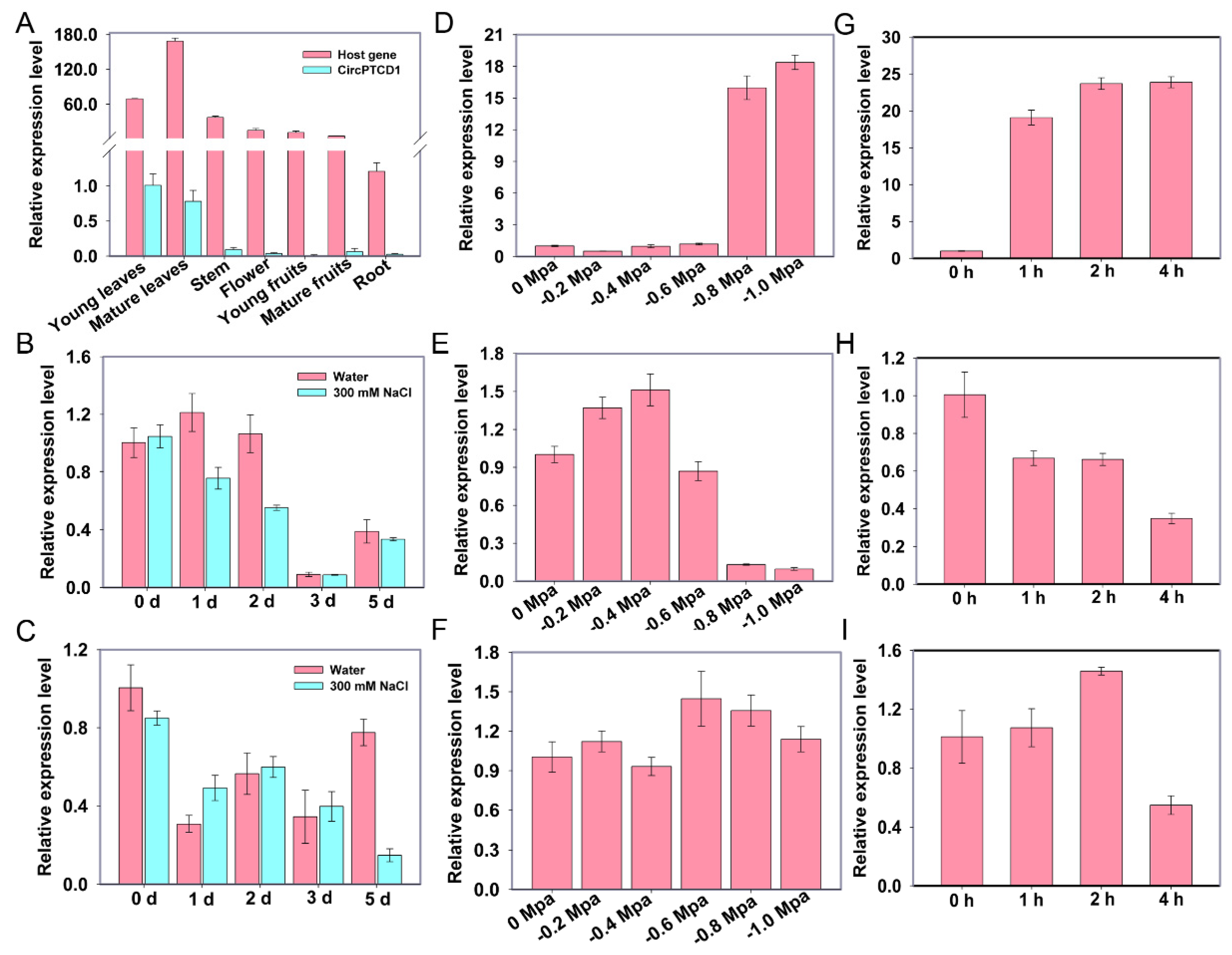
Figure 3.
The OE of Vv-CircPTCD1 and corresponding linear fragment (Linear_PTCD1) in callus of ‘Thompson Seedless’ (A) the schematic of vector construction for circRNA and linear fragment OE, the green line represents the sequence back-spliced into circRNA and the CaMV35S (35S) promoter was used to initiate transcription, Ter refers to transcription termination; (B, C) the PCR and RT-qPCR confirmation of Vv-CircPTCD1 OE in grapevine callus, three positive callus mass were identified, Hyg encode the hygromycin phosphotransferase responsible for resistance selection; (D, E) the PCR and RT-qPCR confirmation of Linear_PTCD1 OE in grapevine callus; (F, G, H) the expression level of host gene, upstream gene and downstream gene in Vv-CircPTCD1 OE lines; (I, J, K) the expression level of host gene, upstream gene and downstream gene in Linear_PTCD1 OE callus mass.
Figure 3.
The OE of Vv-CircPTCD1 and corresponding linear fragment (Linear_PTCD1) in callus of ‘Thompson Seedless’ (A) the schematic of vector construction for circRNA and linear fragment OE, the green line represents the sequence back-spliced into circRNA and the CaMV35S (35S) promoter was used to initiate transcription, Ter refers to transcription termination; (B, C) the PCR and RT-qPCR confirmation of Vv-CircPTCD1 OE in grapevine callus, three positive callus mass were identified, Hyg encode the hygromycin phosphotransferase responsible for resistance selection; (D, E) the PCR and RT-qPCR confirmation of Linear_PTCD1 OE in grapevine callus; (F, G, H) the expression level of host gene, upstream gene and downstream gene in Vv-CircPTCD1 OE lines; (I, J, K) the expression level of host gene, upstream gene and downstream gene in Linear_PTCD1 OE callus mass.
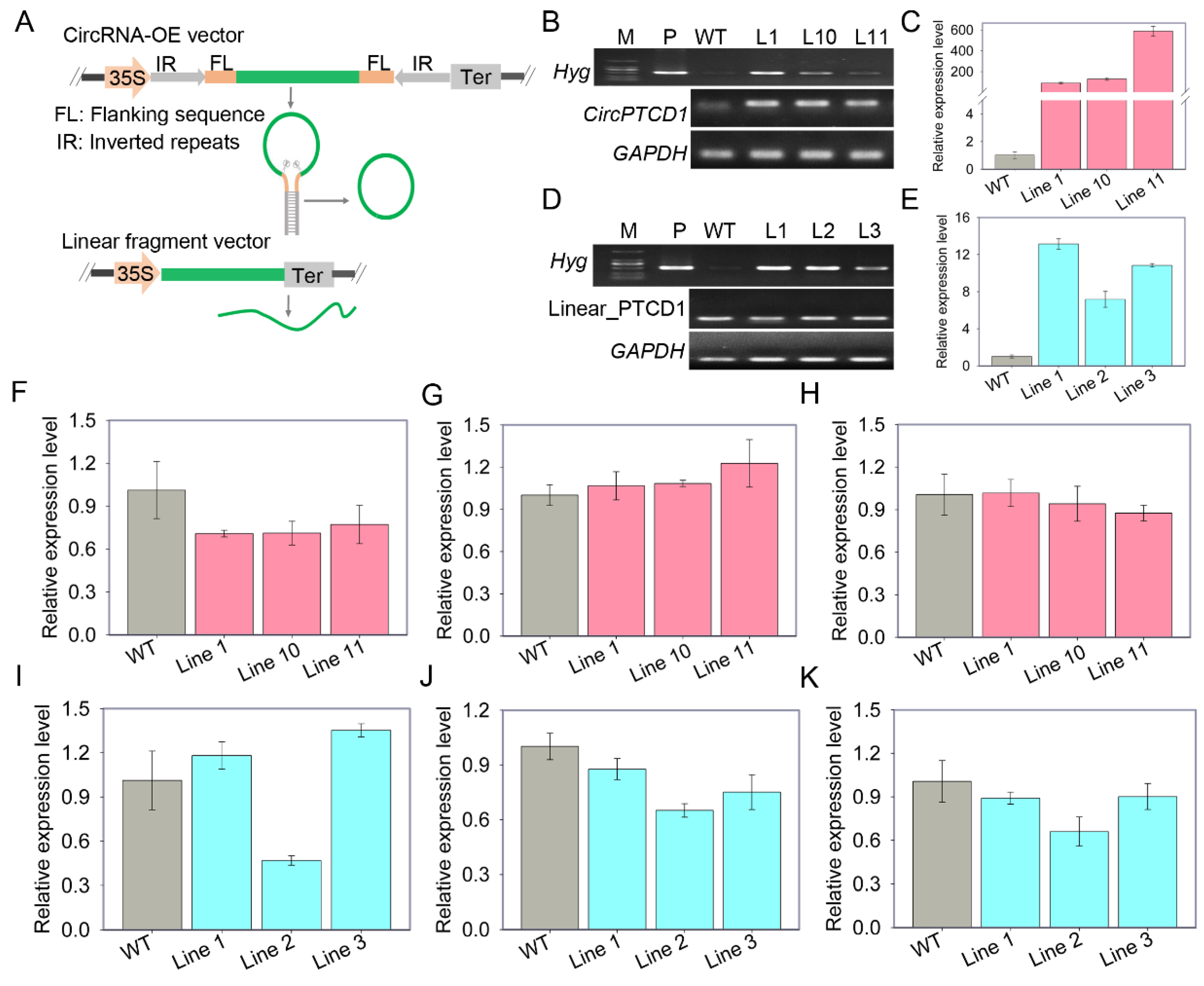
Figure 4.
The phenotype of OE lines in Arabidopsis and callus of grapevine under heat stress (A) Deteriorated phenotype of transgenic Arabidopsis; (B) damage rate of rosette leaves in overexpressed Arabidopsis; (C) the phenotype of overexpressed callus under heat stress. Differences between mean values of groups were compared using t-test (“**”: p <0.01).
Figure 4.
The phenotype of OE lines in Arabidopsis and callus of grapevine under heat stress (A) Deteriorated phenotype of transgenic Arabidopsis; (B) damage rate of rosette leaves in overexpressed Arabidopsis; (C) the phenotype of overexpressed callus under heat stress. Differences between mean values of groups were compared using t-test (“**”: p <0.01).
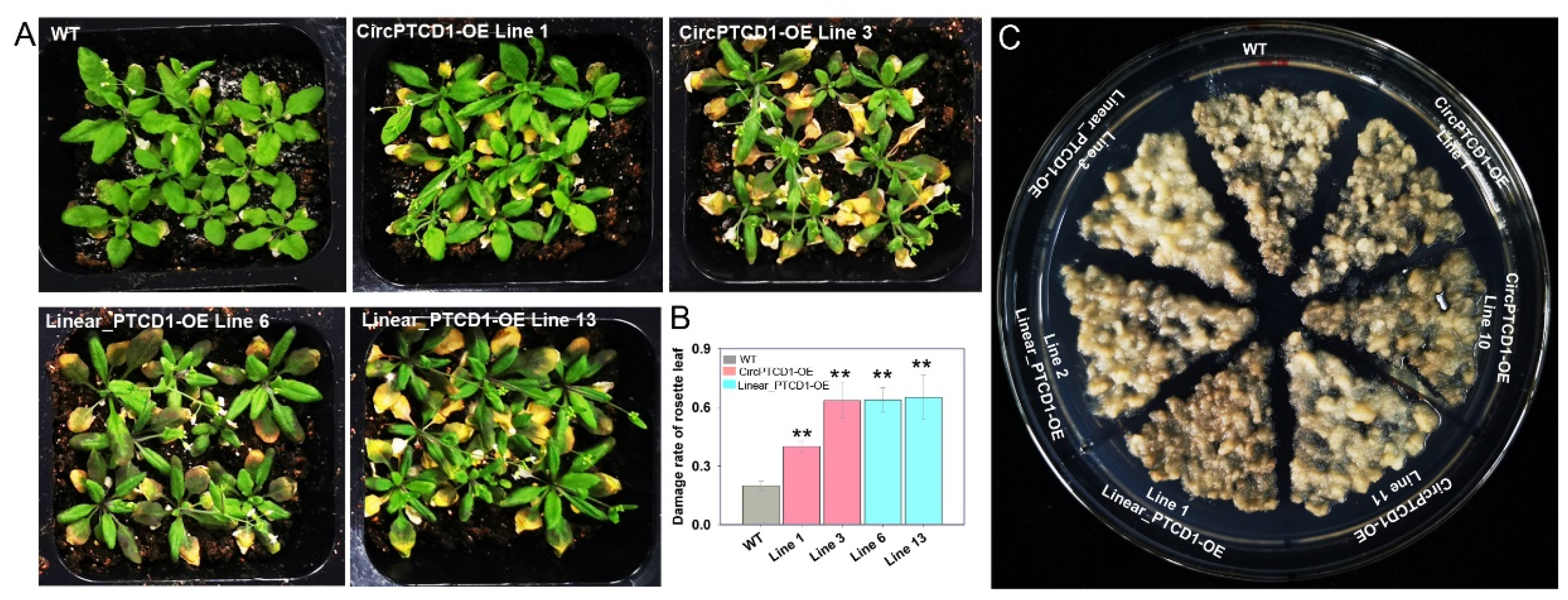
Figure 5.
The phenotype of OE lines in Arabidopsis and callus of grapevine under salt stress (A) The phenotype of OE seedlings and WT on 1/2 MS medium; (B, C) The phenotype and survival rate of OE seedlings and WT on 1/2 MS medium containing 300 mM NaCl; (D) The phenotype of OE and WT callus mass incubated on medium containing 300 mM NaCl. Differences between mean values of groups were compared using t-test (“*”: p <0.05; “***”: p <0.001).
Figure 5.
The phenotype of OE lines in Arabidopsis and callus of grapevine under salt stress (A) The phenotype of OE seedlings and WT on 1/2 MS medium; (B, C) The phenotype and survival rate of OE seedlings and WT on 1/2 MS medium containing 300 mM NaCl; (D) The phenotype of OE and WT callus mass incubated on medium containing 300 mM NaCl. Differences between mean values of groups were compared using t-test (“*”: p <0.05; “***”: p <0.001).
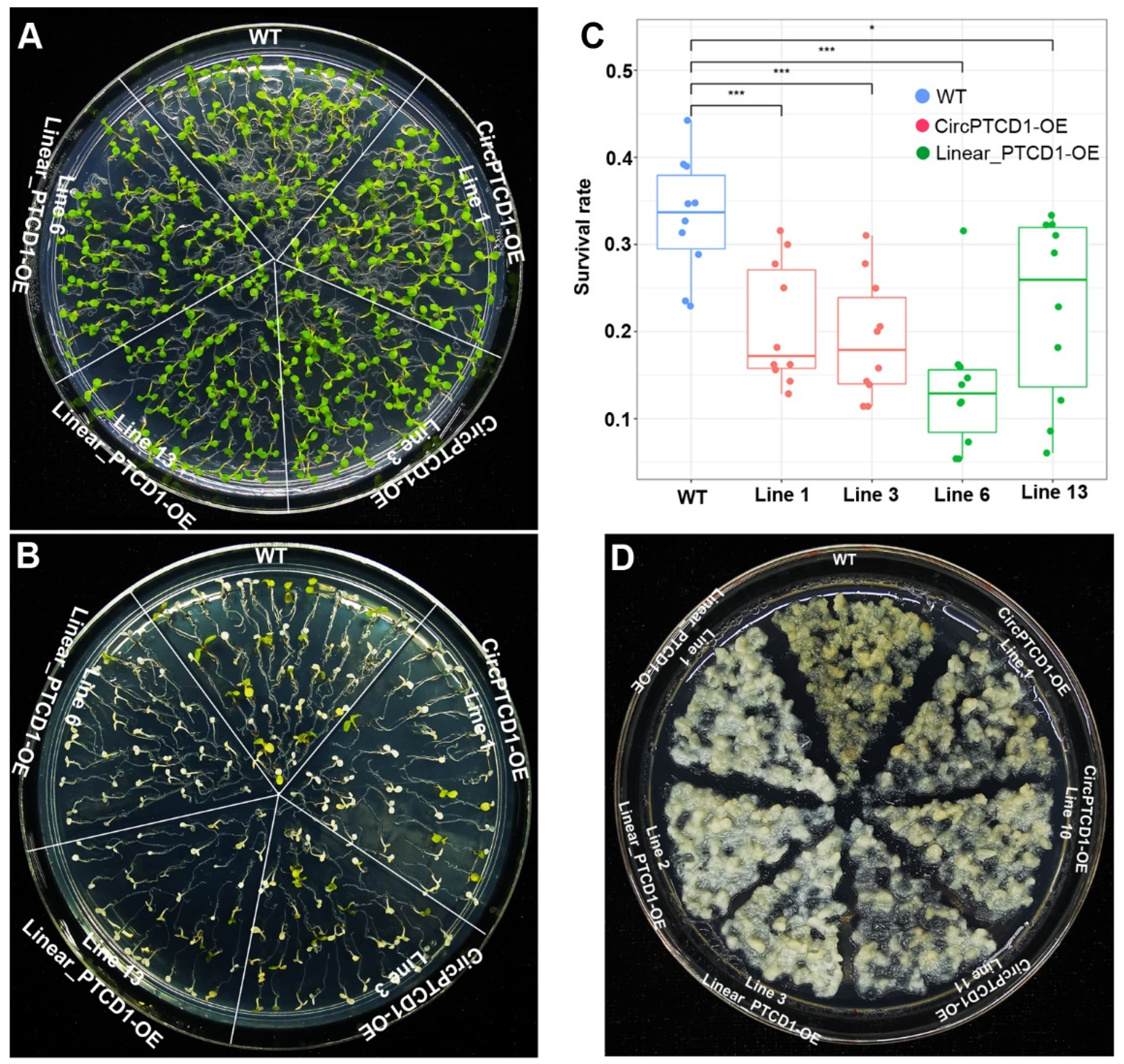
Figure 6.
The phenotype of OE lines in Arabidopsis and callus of grapevine under drought stress (A) The phenotype of a mouth old Arabidopsis during drought stress; (B) the growth of seedlings incubated on PEG-free medium; (C) the growth of seedlings incubated on medium containing PEG-8000; (D, E) the root length and fresh weight of rosette leaves of seedlings during PEG-8000 treatment; (F) the phenotype of grapevine callus mass incubated on medium containing PEG-8000; (G) the growth index of callus mass incubated on medium containing PEG-8000. Differences between mean values of groups were compared using t-test (“*”: p <0.05; “**”: p <0.01; “***”: p <0.001; n.s.: p >0.05).
Figure 6.
The phenotype of OE lines in Arabidopsis and callus of grapevine under drought stress (A) The phenotype of a mouth old Arabidopsis during drought stress; (B) the growth of seedlings incubated on PEG-free medium; (C) the growth of seedlings incubated on medium containing PEG-8000; (D, E) the root length and fresh weight of rosette leaves of seedlings during PEG-8000 treatment; (F) the phenotype of grapevine callus mass incubated on medium containing PEG-8000; (G) the growth index of callus mass incubated on medium containing PEG-8000. Differences between mean values of groups were compared using t-test (“*”: p <0.05; “**”: p <0.01; “***”: p <0.001; n.s.: p >0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



