
Submitted:
20 April 2023
Posted:
21 April 2023
You are already at the latest version
Abstract
Olive mill wastewater (OMW) has serious environmental problems considering its high organic matter, especially its phenolic compounds. The OMW application in crop protection is an alternative environmentally eco-friendly method reducing the chemicals pesticides impact on human health and environment. The present work aimed to study the antifungal activity of fresh and stored OMW and its ethyl acetate extract against phytopathogenic fungi: Syncephalastrum racemosum (S. racemosum), Paramyrothecium roridum (P. roridum), Fusarium oxysporum (F. oxysporum) and Verticilium dahlia (V. dahlia). The OMW was stored at 25 and 45 °C for 3 months. Fresh and stored OMW were used non-sterile, sterile and centrifuged. Phenolic and flavonoid compounds were extracted and identified by HPLC analysis. High inhibition of studied fungi was shown by fresh OMW and its derivative compounds, while OMW storage, sterilization, and centrifugation increased mycelium growth of studied fungi, especially S. racemosum that showed a relative resistance against stored OMW effect and its ethyl acetate extract. Under storage conditions, some phenolic and flavonoid compounds disappeared (resorcinol and vanillic acid), while the concentration of other compounds increased (gallic acid, chlorogenic acid and quercetin). This work highlighted the ability to use fresh OMW as a bio-agent to protect plants from fungi diseases.
Keywords:
Olive mill wastewater
; antifungal activity
; phytopathogenic fungi
; phenolic compounds
; storage
1. Introduction
Worldwide olive oil production is increasing annually by 5%, with olive mill wastewater (OMW) production estimated to be around 6 x 106 m3, 98% of which is in the Mediterranean basin [1]. Since 2015, Saudi Arabia has witnessed a 58.5% increase in olive oil production each year, with an annual production of 80,000 tons in the Kingdom of Saudi Arabia (KSA) in the northern regions of Al-Jouf, Al Qurayyat, Hail, and Tabuk cities, where climatic and geographical conditions are suitable for olive tree cultivation [2,3]. Therefore, OMW production in KSA is estimated at around 160,000 m3 [4].
OMW is characterized by high organic loads expressed in terms of chemical oxygen demand (COD), biological oxygen demand (BOD), total solids (TS), total suspended solids (TSS), total organic carbon (TOC), and phenolic compound concentrations. Phenolic compounds are responsible for OMW’s phytotoxic and antimicrobial activities, making it one of the most polluting wastewater generated by the food industry [5]. In addition, OMW contains soil fertilizing content due to its high mineral nutrient values. OMW’s biological activity is also characterized by its suppressive effects on plant pathogens. Many microorganisms are associated with plant diseases, including phytopathogenic fungi, which are causal agents of some of the world’s most serious plant diseases, capable of significantly reducing productivity on a large scale in agricultural fields. OMW can suppress the growth of these pathogens [6,7,8,9], as well as phytopathogenic bacterial and fungal species (Fusarium oxysporum f. sp. lycopersici, Pythium spp., Sclerotinia sclerotiorum, Verticillium dahliae [6,10], Phytophthora capsici, Rhizoctonia solani [11]), weed species (Amaranthus retroflexus, Solanum nigrum, Chenopodium album, and Sorghum halepense), and nematodes [11]. OMW does not harm crop growth when used appropriately regarding dose and timing for crop protection [12].
According to Jarboui et al. [13] and Magdich et al. [14], OMW contains phenolic compounds and free fatty acids that provide antimicrobial and phytotoxic properties. In particular, phenolic compounds exhibit antimicrobial activity, making them useful as natural plant protectors against fungal [15] and bacterial [16,17] diseases.
Furthermore, OMW can be used as a phytoprotective compound during the growth, harvest, and storage of vegetables and fruits. It is a promising way to prevent fruit and vegetable losses following an attack by saprophytic and phytopathogenic fungi [18,19]. In this regard, using OMW or phenolic compound extracts as alternatives to pesticides during the growing season is an appropriate approach to protecting humans and the environment from the harmful effects of pesticides frequently applied to plants.
Therefore, the present study aimed to investigate the antifungal activity of fresh and stored OMW, stored at 25 and 45 °C for three months, and its ethyl acetate extract against phytopathogenic fungi.
2. Materials and Methods
2.1. Sampling
The OMW was obtained from a three-phase continuous extraction factory in Al Jouf City, Saudi Arabia (29° 52’ 40.8108’’ N, 40° 6’ 15.5016’’ E) and underwent physicochemical characterization. The pH and electrical conductivity (EC) were measured directly using a pH meter (WTW inoLab 7110 model) and conductivity meter (CL 221 model), respectively. The TS content was determined after drying the OMW overnight at 105 °C [20]. The volatile solids (VS) content was gravimetrically assessed after incinerating the dry OMW at 550 °C for 4 h [20]. The ash content was calculated as the total and volatile solid contents. The TSS were determined gravimetrically after centrifuging the crude OMW at 4000 rpm for 15 min [20]. The COD was determined using the Knechtel method [21]. The BOD was measured by the manometric method with a pressure sensor using the OxiTop method. The TOC was measured with the Shimadzu TOC-500 analyzer after acidic hydrolysis of the sample with concentrated HCl [20]. The total phosphorus (TP) concentration was determined calorimetrically as a molybdovanadate phosphoric acid complex, while the total nitrogen (TN) concentration was determined by the Kjeldahl method [20]. The macro- and micro-elements were analyzed by an atomic absorption spectrophotometer (Hitachi Z-6100), and the polyphenol content was determined gravimetrically after extracting it with ethyl acetate. All OMW parameters were compared to the standard limits recommended by WHO [22] and the presidency of meteorology and environment in KSA [23]. All parameters were analyzed in triplicate.
2.2. Isolation and identification of phytopathogenic fungi
Phytopathogenic fungi were isolated from diseased plant organs (Table 1). To remove saprophyte microorganisms, surfaces of diseased plant organs were disinfected with 5% of chlorite solution (NaClO) and then, washed with sterile distilled water for 30 min. Under sterile conditions, plant fragments were cutted with sterile scalpel and then transferred onto Potato Dextrose Agar (PDA, Himedia, USA ). The Petri dishes were incubated at 28°C for 7 days. Isolated fungi were purified twice on PDA to confirm the purification of the selected isolates [24]. Isolated fungi were molecular identified.
Molecular identification of isolated fungi was performed by sequencing the internal transcribed spacer (ITS). Therefore, the genomic DNA of isolated fungi was extracted from a seven-day-old fungal culture grown on PDA. The fungal mass was scraped out with a fine spatula and then grinded in liquid nitrogen. 100 mg of mycelium was used for genomic DNA extractions using i-genomic™ Series DNA Extraction Mini Kit (iNtRON, Korean) specialized for extracting genomic DNA from the specific specimen.
The Internal Transcribed Spacer (ITS) gene amplification was performed using the ITS universal primer (Sigma Company, Germany). The universal primer, ITS1 (TCCGTAGGTGAACCTTGCGG) and ITS4 (TCCTCCGCTTATTGATGC) produced approximately 500 bp of amplicon products. PCR Master Mix (Promega, Netherlands) was used to amplify the ITS region. PCR mix without a DNA template was performed as a negative control. The PCR reaction conditions were 95°C for 2 min (1 cycle), followed by 35 cycles of denaturation at 95°C for 0.5 min, annealing at 52°C for 30 min, and extension at 72°C for 2 min, before a final extension at 72°C for 15 min.
The resulting purified PCR products were subjected to individual Basic Local Alignment Search Tool (BLAST) searches to verify their authenticity in GenBank. The reported sequences of the ITS rDNA gene were submitted to the GenBank database and assigned accession numbers MZ057792, MZ057761, OL913805, and MW830379, respectively, according to the fungi. The guanine-cytosine (GC) ratio was studied for the ITS regions of the four identified fungi, and calculations were performed using ACUA software [25]. Details regarding the identification, GC content, isolation source and disease symptoms of phytopathogenic fungi are presented in Table 1.
For the phylogenetic analyses, the sequences of the tested and the comparative isolates were aligned with highly similar sequences from the GenBank using the multiple sequence alignment (MUSCLE) software. The alignment of sequences was performed by ClustalX version 2.0 software [26]. Phylogenetic analysis of ITS rDNA derived from Neighbour-Joining method using MEGA 4.0 software.
2.3. OMW inhibition effect against phytopathogenic fungi
The OMW was used fresh and stored at 25 °C and 45 °C for three months. All samples were raw, centrifuged at 4000 rpm for 15 minutes, and sterilized by autoclaving at 121 °C for 20 minutes. To evaluate their potential to inhibit mycelium growth of the four studied phytopathogenic fungi, all OMW samples were diluted with PDA medium at ratios of 1/5 and 1/10. A 6-mm disk of mycelium from the periphery of seven-day-old fungal colonies was placed at the center of each plate and then, plates were incubated at 28 °C. PDA medium was used as a negative control. The mycelial growth inhibition was assessed by observing the point at which the mycelium growth of the control reached the periphery of the Petri dishes [27]. The inhibition percentage of mycelium growth of studied fungi were determined according to the following formula:
D: the diameter of fungal growth mycelium in the control
d: the diameter of fungal growth mycelium in Petri dish containing OMW.
2.4. Microbial count and water content determination
For microbial count, 10 ml of raw and centrifuged fresh and stored OMW at 25 °C and 45 °C for three months were suspended in sterile physiological water (90 ml). The suspension was used for microbial count expressed as total number of colonies forming units (cfu) according to ISO 7218 [28]. For each suspension, serial decimal dilutions were plated in triplicate on different agar media: Plate Count Agar (Pronadisa, Madrid - Spain), for the total count bacteria (TCB) incubated at 37 °C for 24 h, and PDA for yeasts and moulds enumeration, incubated at 28 °C for five days [29].
Water content gravimetrically determined before and after drying the studied OMW samples overnight at 105 °C [20].
2.5. Ethyl acetate extract effect against phytopathogenic fungi
Phenolic compounds were extracted three times using ethyl acetate from raw fresh, and stored OMW at 25 °C and 45 °C for three months. The pooled organic extracts were then filtered through a filter paper where sodium sulphate had been deposited, collected in 250 ml pear-shaped flask and the solvent removed in a rotavapor. The phenolic compounds were gravimetrically determined after weighing the pear-shaped flask initially and after solvent removal. The final concentration of phenolic compounds were estimated as 7.82 g ± 0.24, 8.54 g ± 0.36 and 9.66 g ± 0.82 of fresh and stored OMW at 25 °C and 45 °C respectively. After evaporation using a rotary evaporator, phenolic compounds were dissolved in 2 ml of methanol. The agar diffusion technique was used to determine the antifungal activity of OMW polyphenols against phytopathogenic fungi. 200 µl of each phenolic compound sample was poured onto the surface of the PDA medium, followed by incubation at 4 °C until complete adsorption in the medium. Then, a 6-mm disk of a 7-day-old fungal culture was transferred into the center of the PDA Petri dishes. PDA medium was used as a negative control. These were incubated at 28 °C until the mycelium growth of the control reached the Petri dishes periphery. The experiment was set with three replicates and repeated twice [30].
The inhibition percentage of tested fungi was determined according to the equation:
D: the diameter of fungal growth mycelium in the control
d: the diameter of fungal growth mycelium in Petri dish containing phenolic compounds.
2.6. Phenolic and flavonoid compounds analysis using HPLC method
Phenolic and flavonoid compounds were quantified and identified using reversed-phase High Performance Liquid Chromatography (HPLC) (Agilent 1200 series, Agilent, Santa Clara, CA, USA) equipped with a diode array detector and C18 column of 3 x 250 mm (Thermo Scientific, Waltham, USA). The mobile phase consisted of 0.05% trifluoroacetic acid in water (solvent A, Merck, Darmstadt, Germany) and acetonitrile (solvent B, HPLC grade ≥ 99.9%; Honeywell Seelze, Germany). The temperature was maintained at 40 °C, and the total running time was 20 min. The flow rate was 0.9 ml/min, and the injection volume was 10 µl. The mobile phase was consecutively programmed in a linear gradient as follows: 60–82% B over 20 min, followed by 5 min equilibration at 60% B. Detection was monitored at 280 nm, and the concentration of the identified compounds was determined based on the area of the relative peaks compared to the spectra of the commercial standards.
2.7. Statistical Analysis
The results were expressed as mean ± standard deviation using a one-way analysis of variance (ANOVA) test followed by a Student’s t-test, employing IBM SPSS 20 statistical software. Differences were considered significant when p ≤ 0.05. The results were expressed as means ± standard error (SE). All analyses were conducted in triplicate.
3. Results
3.1. Molecular identification of phytopathogenic fungi
Four phytopathogenic fungi were isolated from different diseased plant organs (Table 1). The high similarity of the sequence analyses was comparable to those of the reference strains available in the GenBank database. The ITS rDNA sequence alignment revealed that the isolates were S. racemosum, P. roridum, F. oxysporum, and V. dahliae. S. racemosum showed the lowest GC content (34%), while P. roridum showed the highest content (55.7%), which provided a reasonable prediction. Figure 1 shows the constructed Neighbour-joining tree based on the sequences of the four tested and twenty two comparative fungal strains obtained from GenBank belonging to four distinct species (S. racemosum, P. roridum, F. oxysporum, and V. dahliae) (Table 2). The results showed four groups. The sequences of each isolated strain were closely related to those in a similar group. The sequence of S. racemosum, P. roridum, F. oxysporum, and V. dahliae exhibited a similarity of 100% with the accession strains MW407961, GQ853401, MT530243, and EF377335 respectively.
3.2. Olive mill wastewater physico-chemical characterization
The physicochemical analysis of the OMW is presented in Table 3. The OMW pH was relatively acidic. The OMW had high organic loads (COD, BOD5, TOC, TS, VS, TSS, and total N) and high mineral content, especially in fertilizing elements (ash, EC, total P, Ca, Mg, K, and Na). The CODs/BOD5 rate (2.3) was lower than 2.5. Moreover, all the values of heavy metals in the OMW were within the standard limits recommended by WHO and the presidency of meteorology and environment in KSA.
3.3. Effect of OMW against phytopathogenic fungi
The suppressive effects of fresh and stored OMW at 25 °C and 45 °C for three months against phytopathogenic fungi were summarized in Table 4. The results indicated that all the studied OMW at 1/5 dilution showed the highest inhibition percentage for all the investigated phytopathogenic fungi compared to that at 1/10 dilution. Furthermore, non-sterile, raw, fresh OMW used at dilutions of 1/5 and 1/10 inhibited (100%) P. roridum, F. oxysporum, and V. dahliae. A similar effect was obtained with raw, non-sterile OMW stored at 25 °C for three months on the mycelium growth of P. roridum at dilutions of 1/5 and 1/10 and F. oxysporum at a dilution of 1/5. After storage at 45 °C for three months, raw, non-sterile OMW relatively decreased the inhibition percentage of all the studied fungi. The thermal treatment of OMW by autoclave strongly decreased the inhibition percentage of all the studied fungi, compared to centrifugation and non-sterilization processed OMW.
Furthermore, it was found that S. racemosum was significantly resistant (p < 0.05) to the effect of sterile raw fresh OMW and OMW stored at 25 °C and 45 °C under all the studied conditions. However, the inhibition percentage was not significant between P. roridum, F. oxysporum, and V. dahliae (p > 0.05).
In order to assess the effect of OMW humidity and microbial flora on fungal inhibition, water content, total count bacteria, and yeasts and molds enumeration were determined (Table 4). The results showed a decrease in the water content of fresh raw nonsterile OMW after storage at 45 °C more than 25 °C for three months. Centrifugation increases the water content in all raw OMW samples, while sterilization decreases it. Fresh OMW showed the highest number of the total count bacteria, while the highest number of yeast and molds was shown in OMW stored at 25 °C for three months. Centrifugation decreases the microbial flora count. After storage at 45 °C for three months, no microbial flora was detected in OMW.
3.4. Effect of ethyl acetate extract against phytopathogenic fungi
The inhibition percentages of the ethyl acetate extract on the mycelium growth of the tested phytopathogenic fungi are presented in Table 5. The fresh OMW ethyl acetate extract showed the highest inhibition growth [40% (S. racemosum) – 88% (F. oxysporum)] for all of the tested phytopathogenic fungi, while those extracted from OMW stored at 45 °C for 3 months exhibited the lowest inhibition percentages [0% (S. racemosum) – 55% (F. oxysporum)].”
Phenolic compounds extracted significantly negatively affected F. oxysporum, while those extracted from stored OMW positively affected S. racemosum mycelium growth.
3.5. Identification and quantification of phenolic compounds and flavonoids in fresh and stored OMW
Representative chromatograms of the ethyl acetate extract compounds from fresh and stored OMW obtained are shown in Figure 2. The fresh OMW chromatogram showed peaks corresponding to monomeric phenolic compounds and flavonoids. It revealed six main phenolic compounds and flavonoids in fresh OMW (Table 6). Quinic acid was the primary representative compound in fresh OMW. Storage of OMW at 25 °C for three months increased the concentrations of quinic acid, chlorogenic acid (5-caffeoylquinic acid), vanillic acid, and quercetin. Moreover, the apparition of cinnamic acid with the disappearance of gallic acid and resorcinol was observed. However, the most abundant phenolic compound in OMW stored at 45 °C was gallic acid, with a concentration of 5675 mg/l, while chlorogenic acid was the least detected phenolic compound, with a concentration of less than 2 mg/l.
4. Discussion
OMW is one of the most polluting wastewater in olive oil-producing countries, and the environmental problems associated with it are primarily due to its complex composition, which makes it difficult to treat using economically sustainable methods, thereby limiting its management and disposal. In Al Jouf, KSA, the OMW was found to be highly polluted by organic and mineral matters, while the CODs/BOD5 rate (2.3) was lower than 2.5, indicating that the OMW is biodegradable [31]. Therefore, OMW lacked heavy metal content that were within the standard limits recommended by WHO [22] and the presidency of meteorology and environment in KSA [23]. These findings are consistent with those reported by several authors studying OMW treatment in the same region [4,32,33] or in other parts of the world [5,34,35].
Managing OMW presents a challenging task for converting polluting wastewater into eco-friendly materials. A potential approach to achieving this is utilizing OMW’s aromatic compounds to manage plant diseases caused by pathogenic fungi, including Botrytis cinerea, Alternaria solani, Fusarium culmorum, Verticillium dahliae, and Sclerotinia sclerotiorum [6,36,37]. In this study, fresh OMW exhibited the highest inhibition of the growth of the studied phytopathogenic fungi. In contrast, storage and thermal treatment of OMW by autoclaving positively affected fungi mycelia growth. Interestingly, S. racemosum was significantly resistant (p< 0.05) to the effect of fresh sterile OMW and OMW stored at 25 °C and 45 °C under all studied conditions. Misra et al. [38] showed that S. racemosum was highly resistant in liquid media added with 2% of tannins.
Furthermore, the effect of storage on fungi was investigated by Al-Awadi and Al-Judaibi [39], who observed high inhibition of F. oxysporum after treatment with fresh camel urine (rich in phenolic compounds), while the inhibition was decreased by 22% when camel urine was stored for six months.
Indeed, long-time storage of OMW decreased their water content and therefore a reduction in the total count bacteria was reported. The yeasts and molds counts increased in OMW stored at 25 °C for three months related to the OMW acidic pH and their ability to adapt at low humidity [40]. After three months of storage at 45 °C, OMW becomes sterile. Furthermore, the thermal treatment by autoclaving sterilized the OMW, increasing the mycelium growth of phytopathogenic fungi compared to non-sterile OMW. This suggests that OMW’s microbial flora is vital in inhibiting phytopathogenic fungi. Several authors have reported similar results [10,41,42]. El-Masry et al. [41] and Bess [42] showed that the primary cause of the antifungal activity of compost and compost extracts was their richness in microflora. Sterilization of compost or compost extract, either by autoclaving or microfiltration (0.22 μm), destroyed their active microflora and caused a loss of their suppressive power [41]. Furthermore, high temperatures may destroy some phenolic compounds, reducing their antifungal activity. More recently, Cibeli et al. [10] reported that the decomposition of metabolites from OMW high-pressure homogenization strongly affected pathogenic fungi compared to the thermal treatment of OMW at 121 °C for 20 min.
The characterization of OMW revealed a high abundance of phenolic and flavonoid compounds, which possess antimicrobial properties, as previously reported in previous studies [8,13,43]. After storage at 25 °C and 45 °C for three months, the phenolic compound content increased, possibly due to decreased water content during prolonged storage at these temperatures. The increase in phenolic content could also be attributed to the inactivation of the phenoloxidase degradation enzyme of phenolic compounds. However, Feki et al. [44] observed a decrease in hydroxytyrosol derivative content and an increase in hydroxytyrosol concentration after four months of OMW storage. They suggested the addition of ethanol to stabilize OMW and prevent enzymatic and non-enzymatic oxidation reactions that could lead to the degradation or polymerization of phenolic compounds.
In our study, fresh OMW contained six major phenolic compounds, while OMW stored at 45 °C exhibited only three major phenolic compounds, with gallic acid being the dominant compound. The high inhibition effect ovoid of ethyl acetate extract of fresh OMW against studied fungi could be due to the lethal concentration of phenolic and flavonoid constituents. The disappearance of some phenolic and flavonoid compounds (quinic acid, resorcinol, vanillic acid, and quercetin) at 45 °C might be due to their rapid degradation during the first time of storage or their polymerization into polyphenolic compounds that are not detectable by HPLC analysis. Similarly, during the fermentation of naturally black olives, Romero et al. [45] reported the dominance of hydroxytyrosol in olive juice after 12 months of storage, despite, the presence of other compounds such as hydroxytyrosol, oleuropein, and tyrosol at time zero.
The antifungal activity of phenolic and flavonoid compounds could be related to different mechanisms exhibited by these compounds against phytopathogenic fungi. Therefore, Kulik et al. [46] and Oufensou et al. [47] mentioned that this bioactivity depends on the ability of phenolic and flavonoid compounds to affect fungal cellular membranes, acidification of cytosolic pH and deterioration of cellular ionic homeostasis. Similarly, Boutigny et al. [48] and Hu et al. [49] reported that the antifungal activity of phenolic compounds depends on their ability to disrupt the integrity of the plasma membrane and induce mitochondrial dysfunction, leading to metabolic stagnation. Martinez et al. [50] mentioned that chlorogenic acid totally inhibited mycelium growth and spore germination of S. sclerotium, F. solani, V. dahliae, B. cinerea and C. sojina. More recently, Calheiros et al. [51] reported that chlorogenic acid caused chitin, glucan and ergosterol reduction of the cell wall and the cell membrane of C. paropsilosis. Li et al. [52] showed that gallic acid inhibits the ergosterol biosynthesis of T. ruburum, while Bitencourt et al. [53] mentioned that quercetin reduces the ergosterol levels of T. ruburum. The efficacy of flavonoids as inhibitors of fungal growth has been referred to as their ability to react with nucleophilic amino acids in fungal proteins [54].
Several studies have reported the effect of temperature on the antioxidant activity and phenolic content of food products [55,56,57]. Rocha-Parra et al. [55] demonstrated that gallic acid was the most stable phenolic compound, and its content remained constant during the storage of encapsulated red wine at all water activity levels. Teles et al. [56] observed that the phenolic compounds and antioxidant activity of grape pomace dried at 60 °C were slightly higher than those observed for samples dried at 50 °C and 40 °C. This increase in phenolic content was attributed to the inhibition of phenoloxidase activity, the enzyme responsible for degrading phenolic compounds. However, drying temperatures of 40 °C and 50 °C were insufficient for inactivating the phenoloxidase enzymes. Terefe et al. [57] reported that exposure of the peroxidase enzyme to temperatures of 60 °C–70 °C for a prolonged time (more than 30 min) was sufficient for its inactivation.
The decrease in antifungal activity observed in the ethyl acetate fraction of OMW stored at 45 °C could be attributed to the sub-inhibitory concentration of gallic acid or the sterilization effect of OMW.
5. Conclusion
The present study demonstrated that long-term storage of OMW at 25 °C and 45 °C has a negative impact on its effect against phytopathogenic fungi. Moreover, the study highlighted an increase in the concentration of some phenolic compounds and the disappearance of others during the storage process. Additionally, storage of OMW led to decrease the water activity, the microbial flora, the inactivation of phenoloxidase, and the initiation of a polymerization process. Notably, fresh OMW and its ethyl acetate extract were found to protect plants against phytopathogenic fungi. Consequently, these OMW extracts could reduce the excessive use of chemical pesticides and hence prevent their impact on human health and the environment.
6. Patents
Author Contributions
R.J.; writing—original draft preparation, M.S.A.; Molecular analysis, S.M.N.M ; review and editing, H.B.; methodology and investigation. All authors have read and agreed to the published version of the manuscript.
Funding
“This research was funded by Deanship of Scientific Research at Jouf University, grant number DSR2020-01-510”
Institutional Review Board Statement
“Not applicable”
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors extend their appreciation to the Deanship of Scientific Research at Jouf University for funding this work through research grant No (DSR2020-01-510).
Conflicts of Interest
“The authors declare no conflict of interest.”
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. Crops 2020. Available online: http://www.fao.org/faostat/en/?#data/QC (accessed on 1 January 2021).
- Fraihat, S.; Gilbert-López, B.; Molina-Díaz, A.; Sabouni, I. Physicochemical characterization of olive oil from Al Jouf area of Saudi Arabia. Inter. J. Chem. Tech. Res. 2017, 10, 1004–1010. [Google Scholar]
- Hemida, M.H.; Ibrahim, A.A.E.; Al-Bahnsawy, R.M.; Al-Shathly, M.R. Influence of environmental factors on olive oil production and quality in the Northern region of Kingdom of Saudi Arabia. J. Am. Sci. 2014, 10, 61–66. [Google Scholar]
- Aly, A.A.; Hasan, Y.N.Y.; Al-Farraj, A.S. Olive mill wastewater treatment using a simple zeolite-based low-cost method. J. Environ. Manage. 2014, 145, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Jarboui, R.; Hadrich, B.; Gharsallah, N.; Ammar, E. Olive mill wastewater disposal in evaporation ponds in Sfax (Tunisia) - Moisture content effect on microbiological and physical chemical parameters. Biodegr. 2009, 20, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Vagelas, I.; Kalorizou, H.; Papachatzis, A.; Botu, M. Bioactivity of olive oil mill wastewater against plant pathogens and post-harvest diseases. Biotechnol. Biotechnol. Equip. 2016, 23, 1217–1219. [Google Scholar] [CrossRef]
- Silvan, J.M.; Pinto-Bustillos, M.A.; Vásquez-Ponce, P.; Prodanov, M.; Martinez-Rodriguez, A.J. Olive mill wastewater as a potential source of antibacterial and anti-inflammatory compounds against the food-borne pathogen Campylobacter. Innov. Food Sci. Emerg. Technol. 2019, 51, 177–185. [Google Scholar] [CrossRef]
- Russo, E.; Spallarossa, A.; Comite, A.; Pagliero, M.; Guida, P.; Belotti, V.; Caviglia, D.; Schito, A.M. Valorization and potential antimicrobial use of olive mill wastewater (OMW) from Italian olive oil production. Antioxidants 2022, 11, 903. [Google Scholar] [CrossRef]
- Pannucci, E.; Caracciolo, R.; Romani, A.; Cacciola, F.; Dugo, P.; Bernini, R.; Varvaro, L.; Santi, L. An hydroxytyrosol enriched extract from olive mill wastewaters exerts antioxidant activity and antimicrobial activity on Pseudomonas savastanoi pv. savastanoi and Agrobacterium tumefaciens. Nat. Prod. Res. 2021, 35, 2677–2684. [Google Scholar] [CrossRef]
- Cibelli, F.; Bevilacqua, A.; Raimondo, M.L.; Campaniello, D.; Carlucci, A.; Ciccarone, C.; Sinigaglia, M.; Corbo, M.R. Evaluation of fungal growth on olive-mill wastewaters treated at high temperature and by high-pressure homogenization. Front. Microbiol. 2017, 8, 2515. [Google Scholar] [CrossRef]
- Cayuela, M.L.; Millner, P.D.; Meyer, S.L.F.; Roig, A. Potential of olive mill waste and compost as biobased pesticides against weeds fungi and nematodes. Sci. Total Env. 2008, 399, 11–18. [Google Scholar] [CrossRef]
- El-Abbassi, A.; Saadaoui, N.; Kiai, H.; Raiti, J.; Hafidi, A. Potential applications of olive mill wastewater as biopesticide for crops protection. Sci. Total Env. 2017, 576, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Jarboui, R.; Sellami, F.; Kharroubi, A.; Gharsallah, N.; Ammar, E. Olive mill wastewater stabilization in open-air ponds: Impact on clay–sandy soil. Bioresour. Technol. 2008, 99, 7699–7708. [Google Scholar] [CrossRef] [PubMed]
- Magdich, S.; Jarboui, R.; Ben Rouina, B.; Boukhris, M.; Ammar, E. A yearly spraying of olive mill wastewater on agricultural soil over six successive years: Impact of different application rates on olive production, phenolic compounds, phytotoxicity and microbial counts. Sci. Total Env. 2012, 430, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L. changes in the chemical and biological profile of xylem sap from diseased cv. Sangiovese vines. Physiol. Mol. Plant Pathol. 2007, 71, 210–229. [Google Scholar] [CrossRef]
- Xia, E.; Deng, G.; Guo, Y.; Li, H. Biological activities of polyphenols from Grapes. Inter. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef]
- Baydar, N.G.; Sagdic, O.; Ozkan, G.; Cetin, S. Determination of antibacterial effects and total phenolic contents of grape (Vitis vinifera) seed extracts. Int. J. Food Sci. 2006, 41, 799–804. [Google Scholar] [CrossRef]
- Bonanomi, G.; Giorgi, V.; Del Sorbo, G.; Neri, D.; Scala, F. Olive mill residues affect saprophytic growth and disease incidence of foliar and soilborne plant fungal pathogens. Agric. Ecosyst. Environ. 2006, 115, 194–200. [Google Scholar] [CrossRef]
- Quaglia, M.; Moretti, Ch.; Cerri, M.; Linoci, G.; Taticchi, A. Effect of extracts of wastewater from olive milling in postharvest treatments of pomegranate fruit decay caused by Penicillium adametzioides. Postharvest Biol. Technol. 2016, 11, 26–34. [Google Scholar] [CrossRef]
- Japanese Standards Association JIS Handbook 1995. Wastewater Treatment Japanese Standards Association Tokyo.
- Knechtel, R.J. A more economical method for the determination of chemical oxygen demand. J. Water Pollut. Control Fed. 1987, 7, 25–29. [Google Scholar]
- World Health Organization (WHO). Guidelines for Drinking-Water Quality (3rd edn). World Health Organization Geneva. 2004.
- Presidency of Meteorology and Environment (PME). General environmental regulations and rules for implementation. Kingdom of Saudi Arabia 15 October 2001, p. 206.
- Shaima, M. N. M; Rania, H. T.; Hani, M. A. A.; Hassan, A. E. Novel biosynthesis of Ag-nanocomplex for controlling Verticillium wilt disease of olive tree. Arch. Phytopathol. Plant Prot. 2021, 1–19. [Google Scholar]
- Umashankar, V.; Arunkumar, V.; Dorairaj, S. ACUA: A software tool for automated codon usage analysis. Bioinformation 2007, 2, 62–63. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Onaran, A.; Bayram, M. Determination of antifungal activity and phenolic compounds of endemic Muscari aucheri (Boiss.) baker extract. J. Agric. Faculty Gazi. Osman Pasa. Univ. 2018, 35, 60–67. [Google Scholar] [CrossRef]
- International Standardisation Organisation, Règles générales pour les examens microbiologiques (ISO 7218), 1996.
- Association Française de la Normalisation (AFNOR), Microbiologie des aliments – Dénombrement des levures et moisissures par comptage des colonies à 25 °C – Méthode de routine (NF V08-059), 1995.
- Leontopoulos, S.V.; Giavasis, I.; Petrotos, K.; Kokkora, M.; Makridis, Ch. Effect of different formulations of polyphenolic compounds obtained from OMWW on the growth of several fungal plant and food borne pathogens. Studies in vitro and in vivo. Agric. Agric. Sci. Procedia 2015, 4, 327–337. [Google Scholar] [CrossRef]
- Płuciennik-Koropczuk, E.; Myszograj, S. New Approach in COD Fractionation Methods. Water 2019, 11, 1484. [Google Scholar] [CrossRef]
- Saleh, A.R.; Sabouni, I.; Salam, S.A.S. Physico-chemical and microbiological characterization of olive mill wastewater in Sakaka Aljouf region KSA. Int. J. Clim. Chang. Strateg. Manage. 2017, 5, 46–53. [Google Scholar]
- Alhajoj, A.; Alowaiesh, B. Innovative solutions for reduction of olive mill wastewater pollution. Desalin. Water Treat. 2019, 155, 48–54. [Google Scholar] [CrossRef]
- Paredes, C.; Cegarra, J.; Roig, A.; Sánchez-Monedero, M.A.; Berna, M.P. Characterization of olive mill wastewater (alpechin) and its sludge for agricultural purposes. Bioresour. Technol. 1999, 67, 111–115. [Google Scholar] [CrossRef]
- Arabi, M.; Elias, A.; Kamel, Z.; Ait younes, Y.; Mansouri, B.; Toumert, I. Characterization of olive mill wastewater and gamma irradiation effects on some parameters of its composition. J. Radioanal. Nucl. Chem. 2018, 317, 1095–1106. [Google Scholar] [CrossRef]
- Drais, M.I.; Pannucci, E.; Caracciolo, R.; Bernini, R.; Romani, A.; Santi, L.; Varvaro, L. Antifungal activity of hydroxytyrosol enriched extracts from olive mill waste against Verticillium dahliae the cause of Verticillium wilt of olive. Phytopathol. Mediterr. 2021, 60, 139–147. [Google Scholar] [CrossRef]
- Chatzistathis, T.; Kavvadias, V.; Sotiropoulos, T.; Papadakis, I.E. Organic fertilization and tree orchards. Agriculture 2021, 11, 692. [Google Scholar] [CrossRef]
- Misra, A.K.; Garg, N.; Yadav, K.K. First Report of Shell Soft Rot of Bael (Aegle marmelos) Caused by Syncephalastrum racemosum in North India. Plant Disease 2016, 100, 1779–1779. [Google Scholar] [CrossRef]
- Al-Awadi, A.; Al-Judaibi, A. Effects of heating and storage on the antifungal activity of camel urine. Clin. Microbiol. 2014, 3, 1000179. [Google Scholar]
- Jarboui, R.; Hadrich, B.; Gharsallah, N.; Ammar, E. Olive mill wastewater disposal in evaporation ponds in Sfax (Tunisia): moisture content effect on microbiological and physical chemical parameters. Biodegradation 2009, 20, 845–858. [Google Scholar] [CrossRef] [PubMed]
- El-Masry, M.H.; AI Khalil, M.S.; Hassouna, I.H.A.H. In situ and in vitro suppressive effect of agricultural composts and their water extracts on some phytopathogenic fungi. World J. Microbiol. Biotechnol. 2002, 18, 551–558. [Google Scholar] [CrossRef]
- Bess, V.H. Compost Teas and Compost Microbiology Understanding Compost Tea. BioCycle Reprints. 1999.
- Caballero-Guerrero, B.; Garrido-Fernandez, A.; Fermoso, F.G.; Rodríguez-Gutierrez, G.; Fernandez-Prior, M.A.; Reinhard, C.; Nystrom, L.; Benítez-Cabello, A.; Arroyo-Lopez, F.N. Antimicrobial effects of treated olive mill waste on foodborne pathogens. LWT-Food Sci. Technol. 2022, 164, 113628. [Google Scholar] [CrossRef]
- Feki, M.; Allouche, N.; Bouaziz, M.; Gargoubi, A.; Sayadi, S. Effect of storage of olive mill wastewaters on hydroxytyrosol concentration. Eur. J. Lipid Sci. Technol. 2006, 108, 1021–1027. [Google Scholar] [CrossRef]
- Romero, C.; Brenes, M.; Garcia, P.; Garcia, A.; Garrido, A. Polyphenol changes during fermentation of naturally black olives. J. Agric. Food Chem. 2004, 52, 1973–1979. [Google Scholar] [CrossRef]
- Kulik, T.; Stuper-Szablewska, K.; Bilska, K.; Bu´sko, M.; Ostrowska-Kołodziejczak, A.; Załuski, D.; Perkowski, J. Trans-cinnamic and chlorogenic acids affect the secondary metabolic profiles and ergosterol biosynthesis by Fusarium culmorum and F. graminearum sensu stricto. Toxins 2017, 9, 198. [Google Scholar] [CrossRef]
- Oufensou, S.; Balmas, V.; Azara, E.; Fabbri, D.; Dettori, M.A.; Schüller, C.; Zehetbauer, F.; Strauss, J.; Delogu, G.; Migheli, Q. Naturally occurring phenols modulate vegetative growth and deoxynivalenol biosynthesis in Fusarium graminearum. ACS Omega 2020, 5, 29407–29415. [Google Scholar] [CrossRef] [PubMed]
- Boutigny, A.L.; Barreau, C.; Atanasova-Pénichon, V.; Verdal-Bonnin, M.; Pinson-Gadais, L.; Richard-Forget, F. Ferulic acid, an efficient inhibitor of type B trichothecene biosynthesis and Tri gene expression in Fusarium liquid cultures. Mycol. Res. 2009, 113, 746–775. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang, J.; Kong, W.; Zhao, G.; Yang, M. Mechanisms of antifungal and anti-aflatoxigenic properties of essential oil derived from turmeric (Curcuma longa L.) on Aspergillus flavus. Food Chem. 2017, 220, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Martínez, G.; Regente, M.; Jacobi, S.; Del Rio, M.; Pinedo, M.; de la Canal, L. Chlorogenic acid is a fungicide active against phytopathogenic fungi. Pest. Biochem. Physiol. 2017, 140, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Calheiros, D.; Dias, M.I.; Calhelha, R.C.; Barros, L.; Ferreira, I.C.F.R.; Fernandes, C.; Gonçalves, T. Antifungal Activity of Spent Coffee Ground Extracts. Microorganisms 2023, 11, 242. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.J.; Liu, M.; Dawuti, G.; Dou, Q.; Ma, Y.; Liu, H.G.; Aibai, S. Antifungal activity of gallic acid in vitro and in vivo. Phytother. Res. 2017, 31, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, T.A.; Komoto, T.T.; Massaroto, B.G.; Miranda, C.E.S.; Beleboni, R.O.; Marins, M.; Fachin, A.L. Trans-chalcone and quercetin down-regulate fatty acid synthase gene expression and reduce ergosterol content in the human pathogenic dermatophyte Trichophyton rubrum. BMC Complement. Altern. Med. 2013, 13, 229. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef]
- Rocha-Parra, D.F; Lanari, M.C.; Zamora, M.C.; Chirife, J. Influence of storage conditions on phenolic compounds stability antioxidant capacity and colour of freeze-dried encapsulated red wine. LWT - Food Sci. Technol. 2016, 70, 162–170. [Google Scholar] [CrossRef]
- Teles, A.S.C.; Chávez, D.W.H.; Gomes, F.D.S.; Cabral, L.M.C.; Tonon, R.V. Effect of temperature on the degradation of bioactive compounds of Pinot Noir grape pomace during drying. Braz. J. Food Technol. Campinas 2018, 21, e2017059. [Google Scholar] [CrossRef]
- Terefe, N.S.; Delon, A.; Buckow, R.; Versteeg, C. Blueberry polyphenol oxidase: characterization and the kinetics of thermal and high-pressure activation and inactivation. Food Chem. 2015, 188, 193–200. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis of ITS rDNA derived from Neighbour-Joining method using MEGA 4.0 software; Diamond shapes represent our isolates and the remaining are reference sequences.
Figure 1.
Phylogenetic analysis of ITS rDNA derived from Neighbour-Joining method using MEGA 4.0 software; Diamond shapes represent our isolates and the remaining are reference sequences.

Figure 2.
HPLC chromatogram of phenolic and flavonoid compounds of fresh OMW (a); OMW stored at 25 °C for 3 months (b) and OMW stored at 45 °C for 3 months (c).
Figure 2.
HPLC chromatogram of phenolic and flavonoid compounds of fresh OMW (a); OMW stored at 25 °C for 3 months (b) and OMW stored at 45 °C for 3 months (c).
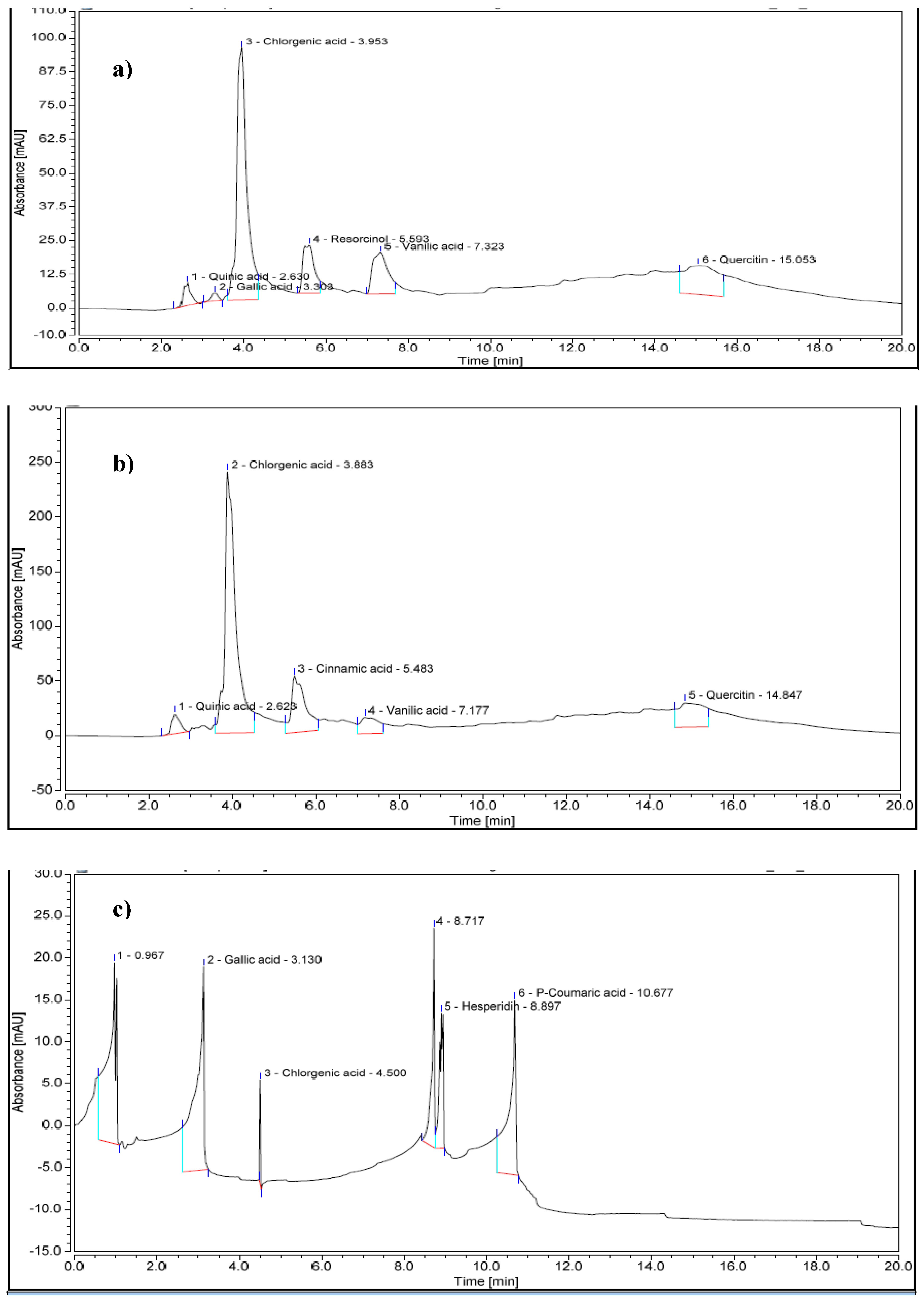

Table 1.
Studied phytopathogenic fungi.
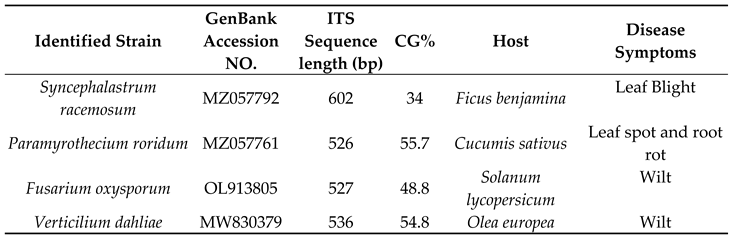 |
ITS: Internal transcribed spacer; GC%: Guanine-cytosine percent; Host: diseased plants from which fungi were isolated.
Table 2.
Comparative fungal strains from GenBank used for the phylogenetic analysis.
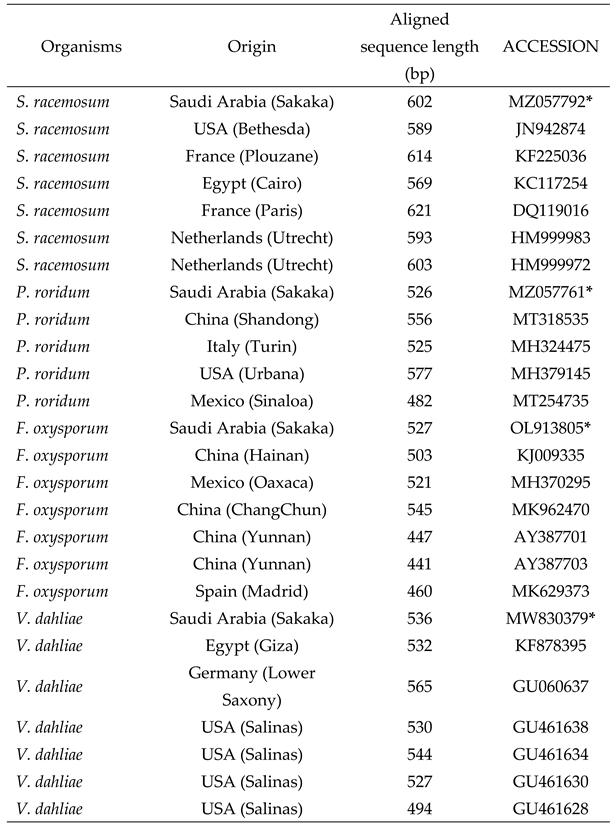 |
*: refereed to our studied fungi: S. racemosum: Syncephalastrum racemosum; P. roridum: Paramyrothecium roridum; F. oxysporum: Fusarium oxysporum; V. dahliae: Verticillium dahliae; bp: base pairs.
Table 3.
Physico-chemical characterization of olive mill wastewater (OMW).
| Parameter | OMW | Maximum limit (WHO) | Maximum limit (KSA) |
| Basic index | |||
| pH | 5.02 ± 0.50 | 6.5-8.0 | 6-9 |
| EC (mS/cm) | 13.52 ± 0.58 | 3000 | ND |
| TS (g/l) | 54.23 ± 1.20 | ND | ND |
| TSS (g/l) | 12.50 ± 0.85 | 500 | 15 |
| COD (g/l) | 76.00 ± 0.15 | ND | 150 |
| CODs (g/l) | 45.40 ± 0.10 | ND | ND |
| BOD5 (g/l) | 19.70 ± 0.05 | 5 | 25 mg/l |
| CODs/BOD5 | 2.30 | ND | ND |
| Organic content | |||
| VS (g/l) | 42.75 ± 1.25 | ND | ND |
| TOC (g/l) | 18.30 ± 1.20 | ND | 50 mg/l |
| Total N (g/l) | 1.50 ± 1.05 | ND | 5 mg/l |
| Total phenols (g/l) | 7.80 ± 0.08 | ND | 0.1 mg/l |
| Inorganic content | |||
| Ash (g/l) | 11.48 ± 0.88 | ND | ND |
| Total P (mg/l) | 76.87 ± 2.30 | < 5 | 1.0 |
| Calcium (mg/l) | 206.70 ± 1.24 | 10 | |
| Potassium (mg/l) | 5500 ± 50 | ND | ND |
| Sodium (mg/l) | 250 ± 2.50 | 919 | ND |
| Magnesium (mg/l) | 128.60 ± 2.00 | 60 | ND |
| Iron (mg/l) | 26.85 ± 2.65 | 5 | ND |
| Zinc (mg/l) | 1.2 ± 0.10 | 3 | 1.0 |
| Barium (mg/l) | 0.29 ± 1.02 | ND | ND |
| Boron (mg/l) | 0.64± 0.40 | ND | ND |
| Chromium (mg/l) | 0.017 ± 0.56 | 0.05 | 0.1 |
| Manganese (mg/l) | 0.166 ± 0.24 | 0.5 | ND |
| Copper (mg/l) | 0.12 ± 0.06 | 0.2 | 0.2 |
| Nickel (mg/l) | 0.087 ± 0.04 | 0.2 | 0.2 |
| Cadmium (mg/l) | 0.012 ± 0.01 | 0.01 | 0.02 |
| Molybdenum (mg/l) | 0.012 ± 0.01 | ND | ND |
| Lead (mg/l) | < 0.001 | 5 | 0.1 |
| Cobalt (mg/l) | < 0.001 | ND | ND |
| Vanadium (mg/l) | < 0.01 | ND | ND |
EC: electrical conductivity; TS: total solid; VS: volatile solids; TSS: total suspended solid; COD: chemical oxygen demand; CODs: soluble chemical oxygen demand; BOD5: biological oxygen demand: Total N: total nitrogen: Total P: total phosphorus; TOC: total organic carbon; ND: not determined. OMW physicochemical parameters were carried out in Triplicate and compared to the standard limits recommended by WHO and the presidency of meteorology and environment in KSA.
Table 4.
Suppressive effect of fresh and stored OMW against phytopathogenic fungi.
| OMW | Inhibition percentage of phytopathogenic fungi (%) | Water content (%) | Microbial flora (log10 N (ufc/ml)) |
|||||
| Dilution | S. racemosum | P. roridum | F. oxysporum | V. dahlia | ||||
| Fresh OMW | Raw non-sterile | 1/10 | 80a ± 2 | 100b | 100b | 100b | 94.16 ± 0.50 | TCB: 4.48 ± 0.02 Y&M: 3.50 ± 0.07 |
| 1/5 | 90a ± 4 | 100b | 100b | 100b | ||||
| Centrifuged non-sterile | 1/10 | 40a ± 1 | 94b ± 2 | 100b | 90b ± 2 | 97.25 ± 0.55 | TCB: 3.35 ± 0.04 Y&M: 2.30 ± 0.02 |
|
| 1/5 | 60a ± 3 | 100b | 100b | 100b | ||||
| Raw sterile | 1/10 | 0a | 46b ± 4 | 40b ± 1 | 20b ± 2 | 91.45 ± 0.26 | TCB: nd Y&M: nd |
|
| 1/5 | 0a | 70b ± 2 | 60b ± 2 | 50b ± 3 | ||||
| Stored at 25 °C for 3 months |
Raw non-sterile | 1/10 | 0a | 100b | 90ab± 1 | 90b± 2 | 88.56 ± 0.25 | TCB: 3.30 ± 0.20 Y&M: 4.80 ± 0.12 |
| 1/5 | 0a | 100b | 100b | 94b ± 4 | ||||
| Centrifuged non-sterile | 1/10 | 0a | 70b ± 4 | 80b ± 4 | 70b ± 2 | 90.50 ± 0.50 | TCB: 1.80 ± 0.20 Y&M: 2.80 ± 0.12 |
|
| 1/5 | 0a | 80b ± 3 | 90b ± 2 | 90b ± 4 | ||||
| Raw sterile | 1/10 | 0a | 60c ± 4 | 10b ± 1 | 20c± 2 | 86.35 ± 0.24 | TCB: nd Y&M: nd |
|
| 1/5 | 0a | 70c ± 2 | 36c ± 2 | 30c ± 2 | ||||
| Stored at 45 °C for 3 months |
Raw non-sterile | 1/10 | 0 a | 70b ± 3 | 70b ± 2 | 50b ± 3 | 70.24 ± 0.45 | TCB: nd Y&M: nd |
| 1/5 | 0 a | 80b ± 2 | 80b ± 4 | 60b ± 2 | ||||
| Centrifuged non-sterile | 1/10 | 0a | 60b ± 4 | 40b ± 1 | 40b ± 1 | 71.14 ± 0.24 | TCB: nd Y&M: nd |
|
| 1/5 | 0a | 66b ± 2 | 50b ± 2 | 54b ± 2 | ||||
| Raw sterile | 1/10 | 0a | 40c± 2 | 16c ± 1 | 30c ± 2 | 67.26 ± 0.85 | TCB: nd Y&M: nd |
|
| 1/5 | 0a | 50c ± 4 | 40c ± 1 | 50c ± 3 | ||||
Different letters (a - c) indicate significant differences (p < 0.05) between different OMW types on studied fungi; ufc: Unit Forming colony; TCB: Total count bacteria; Y&M: Yeasts and molds; nd: not detected.
Table 5.
Inhibition percentage (%) of ethyl acetate extract of fresh and stored OMW on mycelium growth of phytopathogenic fungi.
Table 5.
Inhibition percentage (%) of ethyl acetate extract of fresh and stored OMW on mycelium growth of phytopathogenic fungi.
| OMW | S. racemosum | P. roridum | F. oxysporum | V. dahliae |
| Fresh | 40a ± 2 | 70a ± 4 | 88a ± 2 | 80a ± 1 |
| Stored at 25 °C for 3 months |
0ab | 50ab ± 2 | 62ab ± 1 | 56ab ± 2 |
| Stored at 45 °C for 3 months |
0b | 48b ± 2 | 55b ± 2 | 44b ± 1 |
Different letters (a - b) indicate significant differences (p < 0.05) between different OMW types on studied fungi.
Table 6.
Identification of phenolic and flavonoid compounds in fresh and stored OMW.
| Peak name | Phenolic and flavonoid compounds concentration (mg/l) ± SD | ||
| Fresh OMW | Stored OMW at 25 °C | Stored OMW at 45 °C | |
| Quinic acid | 141.14 ± 2.35 | 335.10 ± 4.25 | nd |
| Gallic acid | 0.28 ± 0.08 | nd | 5675.54 ± 17.23 |
| Chlorogenic acid | 12.83 ± 0.56 | 35.88 ± 1.25 | 1.87 ± 0.20 |
| Cinnamic acid | nd | 18.14 ± 0.88 | nd |
| Resorcinol | 4.37 ± 0.84 | nd | nd |
| Vanillic acid | 6.04 ± 1.02 | 6.70 ± 0.52 | nd |
| Hesperidin | nd | nd | 1118.00 ± 10.00 |
| p-coumaric acid | nd | nd | 3784.24 ± 12.25 |
| Quercetin | 2.95 ± 0.30 | 4.77 ± 0.66 | nd |
nd: not detected; SD: Standard deviation; Note: the HPLC analysis was carried out in duplicate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



