Submitted:
20 April 2023
Posted:
21 April 2023
You are already at the latest version
Abstract
Gene therapy has seen major progress in recent years. Viral vectors have made a significant contribution through efficient engineering for improved delivery and safety. A large variety of indications such as cancer, cardiovascular, metabolic, hematological, neurological, muscular, ophthalmological, infectious diseases, and immunodeficiency have been targeted. Viral vectors based on adenoviruses, adeno-associated viruses, herpes simplex viruses, retroviruses including lentiviruses, alphaviruses, flaviviruses, measles viruses, rhabdoviruses, Newcastle disease virus, poxviruses, picornaviruses, reoviruses, and polyomaviruses have been used. Proof-of-concept has been demonstrated for different indications in animal models. Therapeutic efficacy has also been achieved in clinical trials. Several viral vector-based drugs have been approved for the treatment of cancer, and hematological, metabolic, and neurological diseases. Moreover, viral vector-based vaccines have been approved against COVID-19 and Ebola virus disease.
Keywords:
viral vectors
; gene therapy
; animal models
; clinical trials
; drug approval
1. Introduction
The introduction of genetic engineering in the 1980s provided novel alternatives for the field of biotechnology and played a crucial role in the establishment of gene therapy. Instead of relying on classic drug screening it became feasible to tackle disease on the gene level. Naturally, rapid whole genome sequencing strongly contributed to the improved understanding of gene structure and function, and the association with disease. In this context, it also became feasible to envision supplementation, correction, or modification of endogenous gene expression providing real therapeutic effect [1]. However, gene therapy applications have been expanded to include various forms of cancer therapy, including direct eradication of tumors, immunostimulatory approaches and vaccine development [2]. Additionally, gene silencing based on oligonucleotides [3] and RNA interference (RNAi) [4] have demonstrated promise. Recently, major technology breakthrough has allowed the addition of stem cell technologies [5], chimeric antigen receptor (CAR) T-cell therapy [6] and Clustered Regularly Interspaced Short Palindromic (CRISPR) [7] to the gene therapy repertoire.
In this review, the focus will be solely on viral vectors engineered for gene therapy. Previously, extensive reviews have described in detail the use of various viral vectors for a number of indications [8,9]. To avoid repetitiveness, different viral vectors are presented only briefly. Moreover, selected examples are only presented for different indication areas for major preclinical and clinical findings as well as for approved gene therapy-based drugs. Instead, the major focus here is to provide an overview of which types of viral vectors have been proven successful. Additionally, the areas which require improvement, especially for human use and issues related to complying with GMP production and regulatory demands will be discussed. Finally, challenges and perspectives of future viral vector-based gene therapy applications are presented.
2. Viral Vectors for Gene Therapy
Both DNA- and RNA-based viral vectors have been engineered for gene therapy applications (Figure 1 and Figure 2) (Table 1). Due to the differences in features of viruses, some of them have proven suitable for short-term expression for the treatment of acute diseases such as infectious diseases and cancers. In contrast, those viral vectors which are able to provide long-term expression have been applied for therapeutic intervention of inherited and chronic diseases such as hemophilia, sickle cell disease, muscular dystrophy, and immunodeficiencies. However, the classification is not very strict. For example, adenoviruses have been proven successful in the treatment of both acute and chronic diseases.
In principle, viral vectors which provide long-term expression are adeno-associated virus (AAV) [10], herpes simplex virus (HSV) [11], retroviruses (RV) [12], and lentiviruses (LV) [13]. Adenoviruses can also provide long-term expression, in some cases up to 7 years despite remaining extrachromosomal [14]. Similarly, AAV vectors can provide long-term expression without chromosomal integration [15] although AAV vectors have also been engineered for chromosomal integration [16]. Due to their reverse transcriptase activity, RV and LV can be integrated into the genome, which has provided up to 18 years of functional transgene expression in the case of X-linked severe combined immunodeficiency (SCID-X1) disease [17]. In the case of HSV, although the virus can remain in a latent stage for an extended time, HSV does not integrate into the host genome. A number of ssRNA viruses are characterized by high-level short-term transgene expression. Particularly, self-replicating RNA viruses such as alphaviruses [18], flaviviruses [19], paramyxoviruses [20], and rhabdoviruses [21] have been commonly used for vaccine development and cancer therapy. Moreover, due to its oncolytic properties of specific replication in tumor cells resulting in their killing, Newcastle disease virus (NDV) has been frequently used for cancer therapy [22]. Similarly, large poxviruses, especially the vaccinia virus (VV), have been frequently used for cancer immunotherapy [23]. Coxsackieviruses belonging to the picornaviruses family have been utilized to treat melanoma [24] and lymphoma [25]. Reoviruses carrying a dsRNA genome have shown efficacy in cancer therapy [26]. Finally, polyomaviruses have seen a broad range of applications including vaccine development and expression of suicide, chemoprotective, and anti-angiogenic genes for cancer therapy [27].
The use of the different viral vector expression systems also differs depending on several factors. In most cases, packaging cell lines and systems have been engineered to support the large-scale production of GMP-grade material used in clinical trials and for approved gene therapy products. For example, packaging systems have been constructed for adenoviruses [28], AAV [29], RV [30], and LV [31]. Moreover, packaging cell lines have been engineered for flaviviruses [32,33], paramyxoviruses [34], rhabdoviruses [35], and polyomaviruses [36]. In the case of self-replicating RNA viruses, excessive RNA replication has allowed their use as not only recombinant viral particles but also RNA replicons [37] or plasmid-based DNA replicons [38]. Related to RNA replicons, the delivery of their single-stranded RNA (ssRNA) genome, make them vulnerable to degradation affecting both delivery and transgene expression [39]. Encapsulation of RNA in lipid nanoparticles (LNPs) has demonstrated improved protection against degradation and has enhanced and prolonged transgene expression in vivo [40]. DNA replicons have been engineered by replacing the SP6 or T7 RNA polymerase promoter with a mammalian host cell compatible eukaryotic RNA polymerase type II CMV promoter [38], which allows direct application of DNA plasmids for transfection of mammalian cells or immunization in vivo. However, the drawback of this approach is the inferior transfection efficacy compared to viral infection and in contrast to RNA delivery, the requirement of nuclear delivery of DNA, which significantly reduces the delivery efficacy [41].
Oncolytic viruses have proven efficient for cancer therapy due to their specific replication in tumor cells leading to cell killing without causing much harm to normal tissue [42]. In addition to NDV [22], oncolytic vectors exist for HSV [43], alphaviruses [44], paramyxoviruses [45], and rhabdoviruses [46,47].
3. Gene Therapy Applications using Viral Vectors
Due to the large number of preclinical studies performed with viral vectors, essential examples of successful proof-of-concept studies in animal models and clinical studies in humans for different indications are presented below and summarized in Table 2.
3.1. Cancer Therapy
Cancer therapy has been commonly targeted due to its high frequency in the human population and also because of the fairly straight therapeutic approach of engineering vectors capable of killing tumor cells either after intratumoral or systemic administration. Generally, short-term high-level expression of anti-tumor or cytotoxic genes is sufficient, in fact desirable, to avoid any long-term effects of the treatment. Therefore, it is necessary to ensure that the expression is regulated, preferentially terminated, and not affecting normal tissue. In this context, targeted expression of tumor cells by oncolytic viruses [42], targeting tumor-specific protein structures [48], or utilization of tumor-specific promoters [49] have been useful.
For example, Ad vectors expressing the p53 tumor suppressor gene have been applied to restore the wild-type function of p53 in p53-inactivated tumors, which demonstrated significant suppression of tumor growth in nude mice with esophageal xenografts [50]. Furthermore, Ad vectors expressing the p53 gene were evaluated in a nude mouse model with intraperitoneal human ovarian cancer [51]. The treatment resulted in prolonged survival in mice. In clinical trials, Ad-p53-based therapy of patients with head and neck squamous cell carcinoma (HNSCC) has been proven safe and a promising approach [52]. Ad-p53 has been approved under the brand name GendicineTM in China for the treatment of head and neck cancer [53]. Furthermore, good safety and efficacy have been received in a combination of Ad-p53 and chemo- and radiotherapy [54].
Related to HSV, the oncolytic T-VEC system based on the expression of the granulocyte macrophage-colony stimulating factor (GM-CSF) has demonstrated enhanced tumor growth inhibition and prolonged survival in a melanoma mouse model after intratumoral injection [55]. HSV T-VEC has also been evaluated in phase II and III clinical trials, causing only minor adverse events, and enhanced therapeutic efficacy compared to GM-CSF administration [56]. HSV T-VEC has been approved under the brand name talimogene laherparevec for the treatment of advanced melanoma in the US, Europe, and Australia [57].
The replication-proficient retroviral (RRV) vector Toca 511 expressing the yeast cytosine deaminase (CD) gene prolonged survival in glioma-bearing mice [58]. Moreover, RV Toca 511 administration in patients with high-grade glioma (HGG) resulted in a statistically relevant extension of survival to 13.6 months in a phase I trial [59]. However, the overall survival did not show improvement in a phase II/III trial in HGG patients [60].
In the case of sarcoma therapy, the oncolytic reovirus (Reolysin) inhibited the proliferation and viability of the RD and RH30 rhabdosarcoma cell lines [61]. Additionally, tail vein injection of Reolysin inhibited the growth of tumor xenografts, which was further enhanced by a combination of radiation and cisplatin therapy in athymic nu/nu mice [61]. In clinical settings, Reolysin demonstrated clinical benefits in a phase I trial in patients with solid tumors leading to a partial response (PR) in one patient and stable disease (SD) in seven patients [62]. Furthermore, Reolysin combined with carboplatin and paclitaxel demonstrated good safety and potential efficacy in patients with advanced malignant melanoma in a phase II study [63]. Reolysin was granted orphan drug designation for malignant glioma [64] and a Fast Track designation for metastatic breast cancer [65] by the FDA.
Among paramyxoviruses, intratumoral injection of MV vectors expressing the carcinoembryonic antigen (CEA) delayed tumor growth and extended survival in mice carrying PC-3 prostate tumors [66]. In a phase I clinical trial, SD was observed in all nine patients with recurrent ovarian cancer after treatment with MV-CEA [67]. The overall survival time of 12.15 months was twice the expected time.
In the case of alphaviruses, VEE particles expressing the prostate-specific membrane antigen (PSMA) induced robust PSMA-specific immune responses in BALB/c and C57BL/6 mice [68]. The immunogenicity was enhanced after the administration of repeated doses of VEE-PSMA. Good safety and tolerability were seen in castrate-resistant metastatic prostate cancer (CRPC) patients in a phase I study [69]. Disappointingly, the immune responses were weak in the CRPC patients and require further optimization.
3.2. Cardiovascular and Metabolic Diseases
Gene therapy has also been applied for cardiovascular and metabolic diseases as presented by essential examples below and summarized in Table 3. The sarcoplasmic reticulum Ca2+ ATPase (SERCa2a) has been a common therapeutic target for the restoration of systolic and diastolic heart functions [70]. In this context, SERCa2a expressed from an Ad vector showed restored systolic and diastolic heart function in a rat model of heart failure [71]. In another study, AAV1-SERCa2a administration increased coronary blood flow in a pig heart failure model [72]. Protection against left ventricular dilation, improved systolic and diastolic functions, and reduced mortality rates were obtained in an ischemic rat heart failure model after the administration of LV-SERCa2a [73]. In a phase I trial in patients with heart failure, AAV1-SERCa2a improved functional, symptomatic, and ventricular/remodeling parameters [74]. Moreover, patients with class III/IV heart failure were subjected to a phase II trial, which resulted in improvements in a walking test, peak maximum oxygen consumption, and left ventricular endosystolic volume [75]. Furthermore, the number of cardiovascular events and deaths was reduced by AAV1-SERCa2a treatment in a phase IIa trial [76].
In the context of metabolic diseases, AAV-based expression of the human alpha-1 antitrypsin (hAAT) gene was evaluated in a nonobese diabetic (NOD) mouse model [77]. The treatment resulted in reduced intensity of insulitis, a decrease in insulin autoantibodies, and a lower frequency of diabetes mellitus type 1 (DTM1). In a phase I trial AAV2-hAAT was intramuscularly administered to patients with AAT deficiency showing a good safety profile and resulting in sustained AAT expression for at least one year [78]. Strong immunostaining was detected in AAT patients treated with AAV2-hAAT in a phase II follow-up study [79].
3.3. Hematological Diseases
Hematology has received plenty of attention for gene therapy applications [80] and recent development is described below and summarized in Table 4. Mutations in the factor VIII (FVIII) [81] and factor IX (FIX) [82] genes cause hemophilia A and B, respectively. In the context of hemophilia A, Ad vectors expressing the full-length FVIII gene generated physiological levels of FVIII in mice [83]. Due to the limited packaging capacity of AAV, the strategy has comprised of expression of the B-domain deleted (BDD) FVIII [84,85]. Moreover, the choice of AAV serotype strongly influences FVIII expression levels. For example, much higher FVIII levels were obtained from the AAV8 serotype than from AAV2, AAV3, AAV5 or AAV7 serotypes [86]. Moreover, expression of the canine BDD FVIII from AAV6 and AAV8 generated persistent and therapeutic levels of FVIII for more than 3 years in mice and dogs, while only transient expression was obtained for AAV2-based expression [87]. Another study in dogs demonstrated that AAV8-FVIII administration provided 1-2% of normal levels of FVIII resulting in the prevention of 90% of bleeding episodes [88]. In a 10-year follow-up study, in nine privately owned dogs, AAV8 and AAV9 produced 1.9-11.3% of normal levels of FVIII [89]. Additionally, FVIII has been expressed from an LV vector carrying a platelet-specific integrin alpha 2 b promoter (2bF8 LV) [90]. When bone marrow transduced with the LV vector was transplanted into mice, functional FVIII activity was detected, mice survived tail clipping, and the hemophilia A phenotype was corrected [90]. Related to clinical evaluation, a single administration of AAV-FVIII provided 8-60% of normal levels of FVIII in six hemophilia A patients in a phase I/II study [91]. Moreover, in a multiyear follow-up study, sustained benefits were recorded in hemophilia A patients receiving a single infusion of the AAV5-FVIII SQ variant [92]. For instance, bleeding events decreased, and prophylactic use of FVIII was unnecessary. AAV5-BDD FVIII has received conditional marketing approval from the European Medicines Agency (EMA) for the treatment of severe hemophilia A [93].
In the case of hemophilia B, expression of the FIX gene from an Ad vector resulted in long-term FIX expression in nude mice lasting for more than 300 days [94]. Moreover, complete correction of the hemophilia B phenotype was achieved in FIX-deficient hemophilia B dogs after administration of Ad vectors expressing the canine FIX (cFIX) gene [95]. Although the cFIX levels diminished within 3 weeks to 1-2% of the normal levels of FIX, the therapeutic FIX levels could be restored for at least 6 months by co-administration of the immunosuppressive agent cyclosporin A (CsA) [96]. Expression of FIX from an AAV8 vector resulted in 25-40% of normal FIX activity in two hemophilic dogs, while 200% activity was seen in a third dog [97]. Shortening of the clotting time and lack of bleeding episodes indicated a phenotypic correction in the hemophilic dogs. In a phase I study in 10 hemophilia B patients, AAV8-FIX produced 1-6% of normal levels of FIX for at least 3.2 years without the detection of any toxic effects of the treatment [98]. Moreover, the bleeding episodes were reduced by more than 90% and the use of prophylactic factor XI concentrates was unnecessary. In another approach, self-complimentary AAV2 vectors have been engineered, where the coding region generates dsDNA for immediate replication and transcription-independent of host cell DNA synthesis [99] and allowing utilization of low doses of scAAV2 vectors for circumvention of inflammatory responses [100]. Application of scAAV2 vectors for the expression of the FIX gene provided widespread moderate FIX levels in hepatocytes similar to normal livers, leading to the correction of coagulation function in FIX-deficient mice [100]. Administration of scAAV2-FIX to hemophilia B patients in a phase I trial demonstrated FIX expression for 7 years resulting in a significant reduction in bleeding episodes [91]. Moreover, the AAVS3 vector, comprising a synthetic capsid protein, was designed to enhance hepatocyte tropism and reduce immunogenicity [101]. The AAVS3 vector was applied for FIX expression in a phase I/II trial in hemophilia B patients [102]. Compared to normal FIX levels, 23-43% were obtained in 3 patients, 51-78% in 5 patients, and 260% in one patient, which lasted for at least 27 months. FIX expression has also been conducted from self-inactivating LV (SIN-LV) vectors resulting in long-term stable FIX expression in dogs [103].
Another hematological disease targeted by gene therapy comprises β-thalassemia, which is caused by mutations in the β-globin gene [111]. LV vectors such as LentiGlobin BB305 expresses the human hemoglobin T87Q mutant gene HbAT87Q [104]. In a phase I study, LentiGlobin BB305 administration allowed red blood cell transfusions to be stopped in 12 β-thalassemia patients with the β0/β0 genotype while the transfusion volume could be reduced by 73% in 9 other patients [104]. Moreover, based on interim results from a phase III study, sustained levels of HbAT87Q expression were obtained in patients with the non-β0/β0 genotype making them independent of transfusions [105]. Another approach for targeting β-thalassemia has been to use the mini-β-globin gene expressed from the GLOBE LV vector, which in a proof-of-concept study of intrahepatic in utero delivery in a humanized mouse model normalized the hematological phenotype in mice at 12-32 weeks of age [106]. Application of GLOBE LV-transduced stem cells resulted in rapid recovery in 3 adult and 6 pediatric β-thalassemia patients in a phase I/II study [107]. The outcome of the study was the complete discontinuation of transfusions in children, while it could be reduced in adults.
Sickle cell disease (SCD) has also been a target for gene therapy. As a single point mutation in the βA-globin results in the formation of abnormal hemoglobin [HbS (α2βS2)] responsible for SCD [112], one strategy has been to overexpress a βA-globin variant to prevent HbS polymerization [108]. For example, hematopoietic stem cells (HSCs) transduced with LV vectors expressing the anti-sickling β-globin gene showed expression for 10 months and accumulation of anti-sickling protein up to 52% of total hemoglobin in mice [108]. The previously described LentiGlobin BB305 vector expressing HbAT87Q was used for the transduction of bone marrow cells to a boy with the βS/βS genotype, which resulted in complete remission with correction of hemolysis and biologic hallmarks of the disease [109]. Transduction of CD34+ cells with the LentiGlobin vector did not cause any adverse events in three SCD patients in a phase I/II study [110]. Moreover, interim results demonstrated sustained production of anti-sickling hemoglobin HbAT87Q leading to the complete resolution of severe vaso-occlusive events after a single treatment with the LentiGlobin vector.
3.4. Neurological Disorders
Gene therapy has proven useful for the treatment of neurological disorders as described here and summarized in Table 5. For example, AAV vectors expressing the glutamic acid decarboxylase 65 (GAD65) showed improvement of symptoms related to Parkinson’s disease and relieved pain in rat models [113]. In a phase I study AAV-GAD particles were injected into the subthalamic nucleus of 11 male and one female patients with Parkinson’s disease [114]. The treatment was safe, and significant improvements in motor neuron functions were detected. Moreover, AAV and LV vectors have been utilized for the expression of the glial cell-derived neurotrophic factor (GDNF), which resulted in sustained transgene expression for 3-6 months and also promoted regeneration and significant functional recovery in 6-hydroxydopamine (6-OHDA)-lesioned rats [115]. Additionally, in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MTTP)-lesioned primates AAV-GDNF and LV-GDNF vectors showed neuroprotection, regeneration, and functional recovery [115]. Administration of LV-GDNF to the striatum and substantia nigra reversed functional and motor deficits and prevented nigrostriatal degradation in MTTP-lesioned macaques in another study [116].
AAV vectors have been used for expression of the human aromatic-L-amino acid (hAADC) and administered to primates showing approximately 50% improvement in L-Dopa responsiveness in a dose-ranging study [117]. In a phase I trial in patients with moderate to advanced Parkinson’s disease, AAV-hAADC treatment was well tolerated and provided significant improvement for at least two years in the Parkinson’s Disease Rating Scale (UPDRS) [118]. In another approach, the tyrosine hydroxylase (TH), the hAADC, and the GTP-cyclohydroxylase-1 (GCH-1) genes were expressed from separate AAV vectors [119]. Triple transduction into Parkinsonian rats enhanced the tetrahydrobiopterin (BH4) and dopamine production and also improved the rotational behavior for at least 12 months. In a phase I/II trial in Parkinson’s disease patients, LV-based expression of TH, hAADC, and GCH-1 (ProSavin) was safe and resulted in motor function improvement [120]. Moreover, Prosavin was administered to Parkinson’s disease patients in a phase I/II trial, which resulted in a long-lasting improved UPDRS score [121].
The inherited neurodegenerative Huntington’s disease is caused by a mutation in the huntingtin (HTT) gene, which has made it a valid target for gene therapy [122]. Expression of a microRNA (miRNA) specific for HTT from an AAV5 vector prevented almost completely the formation of mutant HTT aggregates and suppressed DARPP-32-associated neuronal dysfunction in a Huntington’s disease rat model [122]. Moreover, AAV5-miHTT therapy in transgenic Huntington’s disease minipigs reduced mutant HTT mRNA and protein levels significantly [123]. AAV5-miHTT vectors have been subjected to clinical evaluation in a phase I/II trial in adult patients with early-stage Huntington’s disease [124]. Bilateral injection of AAV5-miHTT (AMT-1309) showed good safety and tolerability profiles. In the context of the neuromuscular disease spinal muscular atrophy (SMA), AAV-based expression of the human survival motor neuron (hSMN) gene was evaluated in the central nervous system (CNS) in a mouse SMA model [125]. AAV8-hSMN improved muscle strength, coordination, and locomotion resulting in a significant extension of survival. Remarkable improvement in motor function and prolonged survival were observed in SMA patients in a phase I study [126]. It was also demonstrated that a single injection of AAV9-SMN into SMA patients in another phase I trial improved neuron function and extended the survival of mice [127]. Zolgensma, AAV9 vectors expressing the hSMN1 gene, has been approved for SMA treatment in children in the US, the EU and Canada [128].
3.5. Muscular Diseases
Different types of muscular dystrophy have been subjected to gene therapy applications [129]. Due to the large size of the dystrophin gene, “micro-dystrophin” cassettes (µDys) have been engineered for AAV-based gene therapy applications [130]. Evaluation of AAV6-µDys delivery to severely dystrophic mice reduced skeletal muscle pathology and extended the lifespan of mice by restoration of dystrophin levels in respiratory, cardiac, and limb musculature [131]. In a canine dystrophin model, AAV6-µDys delivery provided efficient dystrophin distribution for two years throughout different skeletal muscles [132]. Patients with Duchenne muscular dystrophy (DMD) have been subjected to the AAV-1 based rAAVrh74-µDys administration in a phase I/II trial [133]. Interim results demonstrated robust therapeutic µDys levels, correct localization of µDys protein, enhanced creatine kinase levels, and improved North Star Ambulatory Assessment (NSAA) scores in 4 treated patients [133]. In a phase I study in 4-12-year-old DMD patients, the AAV9-mini-dystrophin vector is currently under evaluation for safety, tolerability, expression and distribution of dystrophin, and muscle strength [134]. Additional phase I/II and phase III trials applying AAV vectors are currently in progress in DMD patients to evaluate safety, tolerability, and improvement of functional muscle strength (NCT03368742, NCT03375164, and NCT04281485). For example, a single dose of the rAAVrh74-µDys (Delandistrogene moxeparvovec) will be administered to two cohorts of DMD patients 3 months to 3 years and 4 to 7 years of age, respectively (NCT04281485). In another study, the application of the rAAVrh74-µDys vector was evaluated for total antibody binding in 101 DMD patients [135]. The rAAVrh74 serotype, originating from rhesus macaques, has been suggested to be less immunogenic than the AAV2, AAV5, and AAV9 serotypes although providing good transduction of muscle cells, which was confirmed by 86.1% of the DMD patients being seronegative for rAAVrh74 [135]. This finding supports the application of rAAVrh74 vectors for gene therapy on DMD and other neuromuscular diseases.
3.6. Immunodeficiency
Treatment of hereditary immunodeficiencies has provided breakthroughs in gene therapy (Table 6). For example, pediatric SCID patients treated with defective γ-retrovirus (γRV) Moloney murine leukemia virus expressing the interleukin-2 receptor gamma subunit (IL2RG) showed successful disease correction [236], which was hampered by the detection of few cases of leukemia due to the insertion of the therapeutic gene into the LMO2 proto-oncogene region [137]. Initially, γRV-IL2RG treatment generated normal T-cell counts and long-term clinical benefits in 8 out of 10 treated SCIX`D-X1 patients [136]. Moreover, in a long-term follow-up study, all but one patient showed normal growth and protection against SCID-X1-associated infections after 18 years [138]. However, although clinical benefits were obtained in 10 SCID-X1 patients, T-cell acute lymphoblastic leukemia (T-ALL) developed in patients 2-14 years later due to the integration of the γRV vector into either the LMO2 [136] or the CCDN2 [139] locus. To address the issue of chromosomal integration, SIN-γRV vectors have been constructed showing not a single case of leukemia in nine SCID-X1 patients treated with SIN-γRV [140]. Moreover, 44 SCID-X1 patients treated successfully with SIN-LV vectors showed no cases of leukemia [141].
SIN-LV vectors expressing the adenosine triphosphate-binding cassette transporter (ABCD1) gene transduced into patient-derived autologous CD34+ cells were used for the treatment of two patients with X-linked adrenoleukodystrophy (ALD) [140]. The treatment prevented progressive cerebral demyelination resulting in significant clinical benefits [140]. In the case of gene therapy for adenosine deaminase-severe combined immunodeficiency (ADA-SCID), SIN-γRV or SIN-LV vectors have demonstrated sustained ADA expression, metabolic correction, and high overall survival in more than 100 ADA-SCID patients [142,143].
3.7. Infectious Diseases
Vaccine development against infectious diseases has received enormous attention especially due to the recent COVID-19 pandemic (Table 7). In this context, safe and efficient vaccines against the severe acute respiratory syndrome-coronavirus-2 (SARS-CoV-2) have been developed for Ad vectors. For example, expression of the SARS-CoV-2 spike (S) protein from the chimpanzee Ad vector ChAdOx1 generated high vaccine efficacy in phase III clinical trials in healthy volunteers [144]. Similarly, the Ad5 serotype-based Ad5-S-nb2 vaccine has proven efficient in clinical studies [145]. To achieve efficacy, two doses of these COVID-19 vaccines are administered, not including additional booster vaccinations. In contrast, the Sputnik (rAd26-S/rAd5-S) vaccine is based on a prime vaccination with an Ad26 serotype vector expressing the SARS-CoV-2 S protein, followed by a booster vaccination with Ad5 serotype-based SARS-CoV-2 S expression [146]. The rAd26-S/rAd5-S regimen has demonstrated good efficacy in phase III studies [146]. Although the Ad26.COV2.S vaccine is based on an Ad26 serotype vector, it is different to the other Ad-based COVID-19 vaccines as a single dose is sufficient for protection against SARS-CoV-2 [147]. However, emerging variants of SARS-CoV-2 has required booster immunizations also for the Ad26.COV2.S vaccine. In any case, the favorable results from clinical evaluations have resulted in granting emergency use authorization (EUA) for the ChAdOx1 nCoV-19 vaccine in the UK in December 2020, the Ad26.COV2.S vaccine in February 2021 in the US and the Ad5-S-nb2 in China in February 2021 [148]. Although the rAd26-S/rAd5-S was approved in Russia in August 2020, it was surrounded by much controversy as it had only been evaluated in 76 volunteers at that time [149].
Related to other infectious diseases, clinical evaluation of the vesicular stomatitis virus (VSV)-based vaccine against Ebola virus disease (EVD), good safety and efficacy were demonstrated in two phase III trials in Guinea and Sierra Leone [150,151], which contributed to the approval of the VSV-ZEBOV vaccine under the brand name of Ervebo in 2020 [152].
3.8. Other Diseases
Ophthalmologic diseases have proven useful as targets for gene therapy due to the possibility of topic delivery of viral vectors. Briefly, the sFLT01 fusion protein comprising the VEGF and the Fc portion of the human IgG1 expressed from an AAV2 vector was subjected to intravitreal administration in an age-related macular degeneration (AMD) model in cynomolgus macaques, which resulted in localized long-term expression for at least 12 months of the transgene [153]. AAV2-sFL01 showed good safety and tolerability in 19 AMD patients in a phase I study [154]. In a phase II trial, no treatment-related serious adverse events were detected, but improved vision was seen in 11 AMD patients [155]. Moreover, AAV2 vectors expressing the NADH dehydrogenase protein subunit 4 (ND4) were subjected to a phase I study in patients with Leber’s hereditary optic neuropathy (LHON) leading to significant improvement [156]. AAV2-ND4 treatment showed moderate but statistically significant improvement in visual acuity in another phase I study in LHON patients [157]. AAV2 vectors have also been applied for the treatment of the retinal pigment epithelial-specific 65 kD protein (RPE65)-mediated inherited retinal dystrophy [158]. AAV2-RPE65 administration provided maximum vision improvement in inherited retinal dystrophy patients in a phase III trial [158]. Moreover, AAV2-RPE65 was approved under the brand name Luxturna for the treatment of patients with inherited retinal dystrophy in the US, Australia, and Canada [159].
The treatment of lung diseases has mainly focused on cystic fibrosis and overexpression of the cystic fibrosis transmembrane conductance regulator (CFTR) gene [160]. So far, only preclinical studies in macaques with AAV vectors [161], and in mice [162] and pigs [163] with human immunodeficiency virus (HIV) and feline immunodeficiency virus (FIV) vectors, respectively, have been conducted, demonstrating proof-of-concept expression of functional CFTR.
4. Challenges for Viral Vector-Based Gene Therapy
Although viral vectors have provided good safety and efficacy in both preclinical studies and clinical trials, there is certainly still a need for improvement. For example, despite vector engineering being used for generating less cytotoxic alphavirus vectors [164,165] additional vector modifications are desired. Trans-amplifying RNA (taRNA) vectors [166] have also been designed, which due to a significantly smaller vector size potentially require reduced vaccine doses causing fewer adverse events. Another possibility is to further explore the application of oncolytic viruses for cancer therapy [167]. Furthermore, the engineering of pseudoviruses with foreign immunogenic epitopes, full-length surface proteins, or tumor-associated antigens (TAAs) should be explored [168].
Another area of importance, especially related to cancer therapy and cancer vaccines, is the combination of viral vectors with experimental or approved drugs. For example, combination therapy of VEE particles expressing the HER2 gene with the immune checkpoint blockade (ICB) drug pembrolizumab has demonstrated superior efficacy compared to either component alone [169]. Moreover, different prime-boost strategies with viral vectors and conventional plasmid DNA have enhanced immune responses against Dengue virus [170].
Since the first gene therapy applications, gene delivery has been of concern both related to safety and efficacy. Although viral vectors have demonstrated delivery superiority compared to non-viral systems, the application of pseudoviruses can provide special cell targeting as described above. Moreover, the engineering of viral particles with cancer-specific epitopes in their surface structure also results in cell targeting [171]. Although polymer and lipid-based nanoparticles have been mainly used for nucleic acid delivery, examples of alphavirus encapsulation have resulted in improved delivery for oncolytic viruses [172] or passive tumor targeting [173] in animal models.
As is the case for all modern drug development, the process of product manufacturing has become more demanding during the last decades requiring the application of good manufacturing practice (GMP) for viral vector-based gene therapy products [174]. In this context, efficient packaging cell lines and systems have been engineered for most viral vectors such as Ad [28], AAV [29], HSV [175], RV [30], LV [31], flaviviruses [32,33], paramyxoviruses [34], rhabdoviruses [35], and polyomaviruses [36]. Although packaging cell lines have been developed for alphaviruses, their relatively low titers demand further improvements [176].
Another area of concern relates to regulatory issues of gene therapy. The relatively rapid development of gene therapy has caused some confusion regarding the regulatory classification of gene therapy products and vaccines, where for example, mRNA-based cancer vaccines have been categorized under gene therapy medicinal products (GTMPs) whereas vaccines against infectious diseases are excluded despite using the same basic technology and both belonging to immunological medicinal products [177]. Moreover, the FDA and the EMA seem to have different views on genetic manipulation and the components of gene therapy. While the FDA defines gene therapy as the utilization of recombinant DNA [178], the view of the EMA is broader including both DNA and RNA [179]. As the number of gene therapy-based clinical trials grows and new gene therapy products are approved, it is appropriate to resolve these regulatory issues as soon as possible.
5. Conclusions and Perspectives
In summary, proof-of-concept has been demonstrated for different types of viral vectors in animal models for a variety of diseases such as cancer, cardiovascular, metabolic, hematological, neurological, muscular, ophthalmological, infectious diseases, and immunodeficiency. Moreover, clinical evaluation has provided data on safety, tolerability, and efficacy for different indications. For example, clinical benefits and prolonged survival have been achieved resulting in the approval of Ad-based GendicineTM for head-and-neck cancer [53] and talimogene laherparevec (HSV T-VEC) for melanoma [57] treatment. AAV-based gene therapy applications in hemophilia A patients have provided clinical benefits, reduced bleeding episodes, and correction of phenotype in several studies [91,92]. It has led to conditional EMA marketing approval for severe hemophilia A [93]. Similarly, treatment of hemophilia B patients with AAV vectors resulted in significantly reduced bleeding episodes in clinical trials [91,98]. In the context of neurological disorders, Parkinson’s disease patients showed significant clinical benefits lasting for two years after treatment with AAV-hAADC vectors [118]. Moreover, SMA patients treated with AAV-SMN demonstrated improved motor function and prolonged survival [127]. The AAV-SMN vector was approved for the treatment of pediatric SMA patients in the US, the EU, and Canada under the brand name Zolgensma [128]. Among muscular diseases, several clinical trials have demonstrated significant long-term expression of dystrophin after treatment with AAV vectors [133,134]. Probably the greatest gene therapy success has been achieved in treating SCID-X1 [136,137,138,141] and ADA-SCID [143] patients with RV or LV vectors, which has resulted in a complete cure lasting for the whole follow-up period of 18 years [138]. Finally, vaccine development against infectious diseases has led to EUA for adenovirus-based COVID-19 vaccines [148,149] and approval of a VSV-based EVD vaccine [152].
Related to future aspects of viral vector-based gene therapy, it is anticipated that further vector development as described above will contribute to improved safety and efficacy. It seems that not only the combination of viral vectors with existing drugs will generate superior treatment options, but also the combination of different gene therapy technologies can be advantageous. For example, gene editing using the CRISPR-Cas9 strategy has been combined with the helper-dependent HDAd35 vector for the transduction of hematopoietic stem cells (HSCs), which resulted in complete phenotype correction of sickle cell disease in mice [180].
In the context of delivery, it is well recognized that successful transition from proof-of-concept studies in rodents has been difficult to reproduce in humans in clinical trials mainly for two reasons. First, the size difference between rodents and humans may play an important role and might require optimization of both delivery formulations and doses. Second, the induced tumors in rodents do not correspond to spontaneously occurring cancers in humans. For this reason, gene therapy programs would benefit from evaluation in dog models with naturally occurring tumors, which more closely resemble human cancers prior to conducting clinical studies in humans [181].
Personalized medicines have received much attention lately due to rapid next-generation sequence technology development and improved gene manipulation capacity based on gene editing methods. In the context of gene therapy, for example, DMD and rare muscular dystrophies like Limb Girdle Muscular Dystrophy (LGMD) require a personalized response [182]. It is facilitated by the increase in patient registries and combinatory therapies, which will contribute to future success in gene therapy applications.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Lundstrom, K. New era in gene therapy. In Novel Approaches and Strategies for Biologics, Vaccines and CancerTherapies; Elsevier: San Diego. California, USA, 2015; pp. 15–37. [Google Scholar]
- Ramirez-Montagut, T. Cancer vaccines. In Novel Approaches and Strategies for Biologics, Vaccines and Cancer Therapies; Elsevier: California, USA, 2015; pp. 365–88. [Google Scholar]
- Martinez, T.; Wright, N.; López-Fraga, M.; et al. Silencing human genetic diseases with oligonucleotide-based therapies. Hum. Genet. 2013, 132, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Bobbin, M.L.; Rossi, J.J. RNA interference (RNAi)-based therapeutics: delivering on the promise? Annu. Rev. Pharmacol. Toxicol. 2016, 56, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Anurogo, D.; Yuli Prasetyo Budi, N.; Thi Ngo, M.H.; et al. Cell and Gene Therapy for Anemia: Hematopoietic Stem Cells and Gene Editing. Int.J Mol. Sci. 2021, 22, 6275. [Google Scholar] [CrossRef] [PubMed]
- Sermer, D.; Brentjens, R. CAR-T cell therapy: full speed ahead. Hematol. Oncol. 2019, 37, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Lino, C.A.; Harper, J.C.; Carney, J.P.; et al. Delivering CRISPR: a review of the challenges and approaches. Drug Delivery 2018, 25, 1234–1257. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. Gene Therapy Cargoes Based on Viral Vector Delivery. Curr Gene Ther. 2023, 23, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. Viral Vectors in Gene Therapy: Where Do We Stand in 2023? Viruses 2023, 15, 698. [Google Scholar] [CrossRef] [PubMed]
- Samulski, R.; Muzycka, N. AAV-mediated gene therapy for research and therapeutic purposes. Annu. Rev. Virol. 2014, 1, 427–451. [Google Scholar] [CrossRef]
- Epstein, A.L.; Marconi, P.; Argnani, R.; Manservigi, A. HSV-1 derived recombinant and amplicon vectors for gene transfer and gene therapy. Curr. Gene Ther. 2005, 5, 445–458. [Google Scholar] [CrossRef]
- Lesbats, P.; Engelman, A.N.; Cherepanov, P. Retroviral DNA Integration. Chem. Rev. 2016, 116, 12730–12757. [Google Scholar] [CrossRef]
- Kay, M.A.; Glorioso, J.C.; Naldini, L. Viral vectors for gene therapy: The art of turning infectious agents into vehicles of therapeutics. Nat. Med. 2001, 7, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Brunetti-Pierri, N.; Ng, T.; Iannitti, D.; Cioffi, W.; Stapleton, G.; Law, M.; Breinholt, J.; Palmer, D.; Grove, N.; Rice, K.; et al. Transgene expression up to 7 years in nonhuman primates following hepatic transduction with helper-dependent adenoviral vectors. Hum. Gene Ther. 2013, 24, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Deyle, D.R.; Russell, D.W. Adeno-associated virus integration. Curr. Opin. Mol. Ther. 2009, 11, 442–447. [Google Scholar] [PubMed]
- Wang, Z.; Lisowski, L.; Finegold, M.J.; Nakai, H.; Kay, M.A.; Grompe, M. AAV Vectors Containing rDNA Homology Increased Chromosomal Integration and Transgene Persistence. Mol. Ther. 2012, 20, 1902–1911. [Google Scholar] [CrossRef] [PubMed]
- Cartier, N.; Hacein-Bey-Abina, S.; Bartholomae, C.C.; Veres, G.; Schmidt, M.; Kutschera, I.; Vidaud, M.; Abel, U.; Dal-Cortivo, L.; Caccavelli, L.; et al. Hematopoietic stem cell gene therapy with a lentiviral vector in X-linked adrenoleukodystrophy. Science 2009, 326, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.H.; Strauss, E.G. The alphaviruses; gene expression, replication and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef]
- Pijlman, G.P.; Suhrbier, A.; Khromykh, A.A. Kunjin virus replicons: An RNA-based, non-cytopathic viral vector system for protein production, vaccine and gene therapy applications. Exp. Opin. Biol. Ther. 2006, 6, 134–145. [Google Scholar] [CrossRef]
- Zhao, D.; Chen, P.; Yang, H.; Wu, Y.; Zeng, X.; Zhao, Y.; Wen, Y.; Zhao, X.; Liu, X.; Wei, Y.; et al. Live attenuated measles virus vaccine induces apoptosis and promotes tumor regression in lung cancer. Oncol. Rep. 2013, 29, 199–204. [Google Scholar] [CrossRef]
- Finke, S.; Conzelmann, K.K. Recombinant rhabdoviruses: Vectors for vaccine development and gene therapy. Curr. Top. Microbiol. Immunol. 2005, 292, 165–200. [Google Scholar]
- Ganar, K.; Das, M.; Sinha, S.; Kumar, S. Newcastle disease virus: Current status and our understanding. Virus Res. 2014, 184, 71–81. [Google Scholar] [CrossRef]
- Kwak, H.; Honig, H.; Kaufman, H.L. Poxviruses as vectors for cancer immunotherapy. Curr. Opin. Drug Discov. Devel. 2003, 6, 161–168. [Google Scholar] [PubMed]
- Shafren, D.R.; Au, G.G.; Nguyen, T.; Newcombe, N.G.; Haley, E.S.; Beagley, L.; Johansson, E.S.; Hersey, P.; Barry, R.D. Systemic therapy of malignant human melanoma tumors by a common cold-producing enterovirus, coxsackievirus a21. Clin. Cancer Res. 2014, 10, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Hadac, E.M.; Kelly, E.J.; Russell, S.J. Myeloma xenograft destruction by a nonviral vector delivering oncolytic infectious nucleic acid. Mol. Ther. 2011, 19, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Clements, D.; Helson, E.; Gujar, S.A.; Lee, P.W. Reovirus in cancer therapy: An evidence-based review. Oncol. Virother. 2014, 3, 69–82. [Google Scholar]
- Cordelier, P.; Bienvenu, C.; Lulka, H.; Marrache, F.; Bouisson, M.; Openheim, A.; Strayer, D.S.; Vaysse, N.; Pradayrol, L.; Buscail, L. Replication-deficient rSV40 mediate pancreatic gene transfer and long-term inhibition of tumor growth. Cancer Gene Ther. 2007, 14, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Fan, J.; Liao, J.; Zou, Y.; Song, D.; Liu, J.; Cui, J.; Liu, F.; Ma, C.; Hu, X.; et al. Engineering the rapid adenovirus production and amplification (RAPA) cell line to expedite the generation of recombinant adenoviruses. Cell Physiol. Biochem. 2017, 41, 2383–2398. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.A.; Chahal, P.S.; Audy, A.; Kamen, A.; Gilbert, R.; Gaillet, B. Manufacturing of recombinant adeno-associated viruses using mammalian expression platforms. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef]
- Berg, K.; Schäfer, V.N.; Barthnicki, N.; Eggenschwiler, R.; Cantz, T.; Stitz, J. Rapid establishment of stable retroviral packaging cells and recombinant susceptible target cell lines employing novel transposon vectors derived from Sleeping Beauty. Virology 2019, 531, 192–202. [Google Scholar] [CrossRef]
- Ferreira, M.V.; Cabral, E.T.; Coroadinha, A.S. Progress and Perspectives in the Development of Lentiviral Vector Producer. Cells Biotechnol. J. 2021, 16, e2000017. [Google Scholar] [CrossRef]
- Gherke, R.; Ecker, M.; Aberle, S.W.; Allison, S.L.; Heinz, F.X.; Mandl, C.W. Incorporation of tick-borne encephalitis virus replicons into virus-like particles by a packaging cell line. J. Virol. 2003, 77, 8924–8933. [Google Scholar] [CrossRef]
- Khromykh, A.A.; Varnavski, A.N.; Westaway, E.G. Encapsidation of the flavivirus Kunjin replicon RNA by using a complementation system providing Kunjin virus structural proteins in trans. J. Virol. 1998, 72, 5967–5977. [Google Scholar] [CrossRef] [PubMed]
- Lal, G.; Rajala, M. Engineering of measles virus to target cancer cells, an attempt. Intl. J. Infect. Dis. 2016, 45, 333–334. [Google Scholar] [CrossRef]
- Ito, N.; Takayama-Ito, M.; Yamada, K.; Hosokawa, J.; Sugiyama, M.; Minamoto, N. Improved recovery of rabies virus from cloned cDNA using a vaccinia virus-free reverse genetics system. Microbiol. Immunol. 2003, 47, 613–677. [Google Scholar] [CrossRef] [PubMed]
- Toscano, M.G.; van der Velden, J.; van der Werf, S.; Odijk, M.; Roque, A.; Camacho-Garcia, R.J.; Herrera-Gomez, I.G.; Mancini, I.; de Haan, P. Generation of a Vero-based packaging cell line to produce SV40 gene delivery vectors for use in clinical gene therapy studies. Mol. Ther. Methods Clin. Dev. 2017, 6, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Liljeström, P.; Garoff, H. A new generation of animal cell expression vectors based on the Semliki Forest virus replicon. Biotechnology 1991, 9, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- DiCiommo, P.D.; Bremner, R. Rapid, high level protein production using DNA-based Semliki Forest virus vectors. J. Biol. Chem. 1998, 273, 18060–18066. [Google Scholar] [CrossRef]
- Wadwha, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Takur, A. Opportunities and Challenges in the Delivery of mRNA-Based Vaccines. Pharmaceutics 2020, 12, 102. [Google Scholar] [CrossRef]
- Raouane, M.; Desmaele, D.; Urbinati, G.; Massaad-Massade, L.; Couvreur, P. Lipid conjugated oligonucleotides: a useful strategy for delivery. Bioconjug. Chem. 2012, 23, 1091–1104. [Google Scholar] [CrossRef]
- Bishop, C.J.; Majewski, R.L.; Guiriba, T.-R.M.; Wilson, D.R.; Bhise, N.S.; Quinones-Hinojosa, A.; Green, J.J. Quantification of Cellular and Nuclear Uptake Rates of Polymeric Gene Delivery Nnaoparticles and DNA Plasmids via Flow Cytometry. Acta Biomater. 2016, 37, 120–130. [Google Scholar] [CrossRef]
- Kaufman, H.L.; Kim, D.W.; DeRaffele, G.; Mitcham, J.; Coffin, R.S.; Kim-Schulze, S. Local and distant immunity induced by intralesional vaccination with an oncolytic herpes virus encoding GM-CSF in patients with stage IIIc and IV melanoma. Ann. Surg. Oncol. 2010, 17, 718–730. [Google Scholar] [CrossRef]
- Li, J.M.; Kao, K.C.; Li, L.F. Micro-RNA-145 regulates oncolytic herpes simplex virus-1 for selective killing of human non-small lung cancer cells. Virol. J. 2013, 10, 241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Y.; Tan, J.; Zhang, Y.; Wong, C.-W.; Lin, Z.; Liu, X.; Sander, M.; Yang, X.; Liang, L.; et al. Necroptotic virotherapy of oncolytic alphavirus M1 cooperated with Doxorubicin displays promising therapeutic efficacy in TNBC. Oncogene 2021, 40, 4783–4795. [Google Scholar] [CrossRef] [PubMed]
- Boisgerault, N.; Guillerme, J.-B.; Pouliquen, D.; Mesel-Lemoine, M.; Achard, C.; Combredet, C.; Fontenau, J.F.; Tangy, F.; Gré- goire, M. Natural oncolytic activity of live-attenuated measles virus against human lung and colorectal adenocarcinomas. Biomed. Res. Int. 2013, 2013, 387362. [Google Scholar] [CrossRef] [PubMed]
- Urbiola, C.; Santer, F.R.; Petersson, M.; van der Pluijm, G.; Horninger, W.; Erlmann, P.; Wollmann, G.; Kimpel, J.; Culig, Z.; von Laer, D. Oncolytic activity of the rhabdovirus VSV-GP against prostate cancer. Int. J. Cancer 2018, 143, 1786–1796. [Google Scholar] [CrossRef]
- Le Boeuf, F.; Selman, M.; Son, H.H.; Bergeron, A.; Chen, A.; Tsang, J.; Butterwick, D.; Arulanandam, R.; Forbes, N.E.; Tzelepis, F.; et al. Oncolytic Maraba Virus MG1 as a Treatment for Sarcoma. Int. J. Cancer 2017, 141, 1257–1264. [Google Scholar] [CrossRef]
- Liu, Y.; Deisseroth, A. Tumor vascular targeting therapy with viral vectors. Blood 2006, 107, 3027–3033. [Google Scholar] [CrossRef]
- Montaño-Samaniego, M.; Bravo-Estupiñan, D.M.; Méndez-Guerrero, O.; Alarcon-Hernandez, E.; Ibanez-Hernandez, M. Strategies for Targeting Gene Therapy in Cancer Cells with Tumor-Specific Promoters. Front. Oncol. 2020, 10, 605380. [Google Scholar] [CrossRef]
- Shimada, H.; Shimizu, T.; Ochiai, T.; Liu, T.L.; Sashiyama, H.; Nakamura, A.; Matusbara, H.; Gunji, Y.; Kobayashi, S.; Tagawa, M.; et al. Preclinical study of adenoviral p53 gene therapy for esophageal cancer. Surg. Today 2001, 3, 597–604. [Google Scholar] [CrossRef]
- von Grueningen, V.E.; Santoso, J.T.; Coleman, R.L.; Muller, C.Y.; Miller, D.S.; Mathis, J.M. In vivo studies of adenovirus-based p53 gene therapy for ovarian cancer. Gynecol. Oncol. 1998, 69, 197–204. [Google Scholar] [CrossRef]
- Tang, R.; Xu, Z. Gene therapy: a double-edged sword with great powers. Mol. Cell Biochem. 2020, 474, 73–81. [Google Scholar] [CrossRef]
- Räty, J.K.; Pikkarainen, J.T.; Wirth, T.; Ylä-Herttuala, S. Gene therapy: The first approved gene-based medicines, molecular mechanisms and clinical indications. Curr. Mol. Pharmacol. 2008, 1, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Li, L.; Li, D.; Liu, J.; Li, X.; Li, W.; Xu, X.; Zhang, M.J.; Chandler, L.A.; Lin, H.; et al. The first approved gene therapy product for cancer ad-p53 (Gendicine): 12 years in the clinic. Hum. Gene Ther. 2018, 29, 160–179. [Google Scholar] [CrossRef] [PubMed]
- Toda, M.; Martuza, R.L.; Rabkin, S.D. Tumor growth inhibition by intratumoral inoculation of defective herpes simplex virus vectors expressing granulocyte-macrophage colony-stimulating factor. Mol Ther. 2000, 2, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Puzanov, I.; Kelley, M.C. Talimogene laherparevec (T-VEC) for the treatment of advanced melanoma. Immunotherapy 2015, 7, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Rehman, H.; Silk, A.W.; Kane, M.P.; Kaufman, H.L. Into the clinic: Talimigene laherparevec (T-VEC), a first-in-class intratumoral oncolytic viral therapy. J. Immunother. Ther. Cancer 2016, 4, 53. [Google Scholar] [CrossRef]
- Huang, T.T.; Parab, S.; Burnett, R.; Diago, O.; Ostertag, D.; Hofman, F.M.; Lopez Espinoza, F.; Martin, B.; Ibanez, C.E.; Kasahara, N.; et al. Intravenous administration of retroviral replicating vector, Toca 511, demonstrates efficacy in orthotopic immune-competent mouse glioma model. Hum. Gene Ther. 2015, 26, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Cloughesy, T.F.; Landolfi, J.; Hogan, D.J.; Bloomfield, S.; Carter, B.; Chen, C.C.; Elder, J.B.; Kalkanis, S.N.; Kesari, S.; Lai, A.; et al. Phase I trial of vocimagine amiroretrorepvec and 5-fluorocytosine for recurrent high-grade glioma. Sci. Transl. Med. 2016, 8, 341ra75. [Google Scholar] [CrossRef]
- Tocagen Reports Results of Toca 5 Phase 3 Trial in Recurrent Brain Cancer. Tocagen. Published 12 September 2019. https://bit.ly/2lPm19v (accessed on 21 March 2023).
- Hingorani, P.; Zhang, W.; Lin, J.; Liu, L.; Guha, C.; Kolb, E.A. Systemic administration of reovirus (Reolysin) inhibits growth of human sarcoma xenografts. Cancer 2011, 117, 1764–1774. [Google Scholar] [CrossRef]
- Gollamudi, R.; Ghalib, M.H.; Desai, K.K.; Chaudhary, I.; Wong, B.; Einstein, M.; Coffey, M.; Gill, G.M.; Mettinger, K.; Mariadason, J.M.; et al. Intravenous administration of Reolysin, a live replication competent RNA virus is safe in patients with solid tumors. Invest. New Drugs 2010, 28, 641–649. [Google Scholar] [CrossRef]
- Mahalingam, D.; Fountzilas, C.; Moseley, J.; Noronha, N.; Tran, H.; Chakrabarty, R.; Selvaggi, G.; Coffey, M.; Thompson, B.; Sarantopoulos, J. A phase II study of REOLYSIN® (pelareorep) in combination with carboplatin and paclitaxel for patients with advanced malignant melanoma. Cancer Chemother Pharmacol. 2017, 79, 697–703. [Google Scholar] [CrossRef]
- Oncolytics Biotech (ONCY) Announces Receipt of FDA Orphan Drug Designation for REOLYSIN. April 2015. www.streetinsider.com (accessed on 21 March 2023).
- Oncolytics Biotech Inc. Announces FDA Fast Track Designation for REOLYSIN in Metastatic Breast Cancer”. www.newswire.ca (accessed on 21 March 2023).
- Msaouel, P.; Iankov, I.D.; Allen, C.; Morris, J.C.; von Messling, V.; Cattaneo, R.; Koutsilieris, M.; Russell, S.J.; Galanis, E. Engi- neered measles virus as a novel oncolytic therapy against prostate cancer. Prostate 2009, 69, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Hartmann, L.C.; Cliby, W.A.; Long, H.J.; Peethambaram, P.P.; Barrette, B.A.; Kaur, J.S.; Haluska, P.J. Jr.; Aderca, I.; Zollman, P.J.; et al. Phase I trial of intraperitoneal administration of an oncolytic measles virus strain engineered to express carcinoembryonic antigen for recurrent ovarian cancer. Cancer Res. 2010, 70, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Durso, R.J.; Andjelic, S.; Gardner, J.P.; Margitich, D.J.; Donovan, G.P.; Arrigale, R.R.; Wang, X.; Maughan, M.F.; Talarico, T.L.; Olmsted, R.A.; et al. A novel alphavirus vaccine encoding prostate-specific membrane antigen elicits potent cellular and humoral immune responses. Clin. Cancer Res. 2017, 13, 3999–4008. [Google Scholar] [CrossRef] [PubMed]
- Slovin, S.F.; Kehoe, M.; Durso, R.; Fernandez, C.; Olson, W.; Gao, J.P.; Israel, R.; Scher, H.I.; Morris, S. A phase I dose escalation trial of vaccine replicon particles (VRP) expressing prostate-specific membrane antigen (PSMA) in subjects with prostate cancer. Vaccine 2013, 31, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Gianni, D.; Chan, J.; Gwathmey, J.K.; del Monte, F.; Hajjar, R.J. SERCA2a in heart failure: role and therapeutic prospects. J. Bioenerg. Biomembr. 2005, 37, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, M.I.; del Monte, F.; Schmidt, U.; DiSalvo, T.S.; Kang, Z.B.; Matsui, T.; Guerrero, J.L.; Gwathmey, J.K.; Rosenzweig, A.; Hajjar, R.J. Adenoviral gene transfer of SERCa2a improves left-ventricular function in aortic-banded rats in transition to heart failure. Proc. Natl. Acad. Sci. USA 2000, 97, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Hadri, L.; Bobe, R.; Kawase, Y.; Ladage, D.; Ishikawa, K.; Atassi, F.; Lebeche, D.; Kranisa, E.G.; Leopold, J.A.; Lompre, A.-M-; et al. SERCA2a gene transfer enhances eNOS expression and activity in endothelial cells. Mol. Ther. 2010, 18, 1284–1292. [Google Scholar] [CrossRef]
- Niwano, K.; Arai, M.; Koitabashi, N.; Watanabe, A.; Ikeda, Y.; Miyoshi, H.; Kurabayashi, M. Lentiviral vector–mediated SERCA2 gene transfer protects against heart failure and left ventricular remodeling after myocardial infarction in rats. Mol. Ther. 2008, 16, 1026–1032. [Google Scholar] [CrossRef]
- Jaski, B.E.; Jessup, M.L.; Mancini, D.M.; Cappola, T.P.; Pauly, D.F.; Greenberg, B.; Borrow, K.; Dittrich, H.; Zsebo, K.M.; Hajjar, R.J. Calcium Up-Regulation by Percutaneous Administration of Gene Therapy In Cardiac Disease (CUPID) Trial Investigators. Calcium upregulation by percutaneous administration of gene therapy in cardiac disease (CUPID Trial), a first-in-human phase 1/2 clinical trial. J. Card. Fail. 2009, 15, 171–181. [Google Scholar]
- Jessup, M.; Greenberg, B.; Mancini, D.; Cappola, T.; Pauly, D.F.; Jaski, B.; Yaroshinsky, A.; Zsebo, K.M.; Dittrich, H.; Haijjar, H. Calcium Upregulation by Percutaneous Administration of Gene Therapy in Cardiac Disease (CUPID) Investigators. Calcium Upregulation by Percutaneous Administration of Gene Therapy in Cardiac Disease (CUPID): A phase 2 trial of intracoronary gene therapy of sarcoplasmic reticulum Ca2+-ATPase in patients with advanced heart failure. Circulation 2011, 124, 304–313. [Google Scholar]
- Zsebo, K.; Yaroshinsky, A.; Rudy, J.J.; Wagner, K.; Greenberg, B.; Jessup, M.; Hajjar, R.J. Long-term effects of AAV1/SERCA2a gene transfer in patients with severe heart failure: Analysis of recurrent cardiovascular events and mortality. Circ. Res. 2014, 114, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Goudy, K.; Campbell-Thompson, M.; Wasserfall, C.; Scott-Jorgensen, M.; Wang, J.; Tang, Q.; Crawford, J.M.; Ellis, T.M.; Atkinson, M.A.; et al. Recombinant adeno-associated virus-mediated alpha-1 antitrypsin gene therapy prevents type I diabetes in NOD mice. Gene Ther. 2004, 11, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Brantly, M.L.; Chulay, J.D.; Wang, L.; Muller, C.; Humphries, M.; Spencer, L.T.; Rouhani, F.; Conlon, T.J.; Calcedo, R.; Betts, M.R.; et al. Sustained transgene expression despite Tlymphocyte responses in a clinical trial of rAAV1-AAT gene therapy. Proc. Natl. Acad. Sci. USA 2009, 106, 16363–16368. [Google Scholar] [CrossRef]
- Flotte, T.R.; Trapnell, B.C.; Humphries, M.; Carey, B.; Calcedo, R.; Rouhani, F.; Campbell-Thompson, M.; Yachnis, A.T.; Sandhaus, R.A.; McElvaney, N.G.; et al. Phase 2 clinical trial of a recombinant adeno-associated viral vector expressing alphal- antitrypsin: Interim results. Hum. Gene Ther. 2011, 22, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. Gene Therapy in Hematology. In Comprehensive Hematology and Stem Cell Research; Elsevier: Chennai, India, 2023; in press. [Google Scholar]
- Gale, A.J.; Pellequer, J.L.; Getzoff, E.D.; Griffin, J.H. Structural basis for hemophilia A caused by mutations in the C domains of blood coagulation factor VIII. Thromb. Haemost. 2000, 83, 78–85. [Google Scholar] [CrossRef]
- Ludwig, M.; Sabharwal, A.K.; Brackmann, H.H.; Olek, K.; Smith, K.J.; Birktoft, J.J.; Bajaj, S.P. Hemophilia B caused by five different nondeletion mutations in the protease domain of factor IX. Blood 1992, 79, 1225–1232. [Google Scholar] [CrossRef]
- Balagué, C.; Zhou, J.; Dai, Y.; Alemany, R.; Josephs, S.F.; Andreason, G.; Hariharan, M.; Sethi, E.; Prokopenko, E.; Jan, H.Y.; et al. Sustained high-level expression of full-length human factor VIII and a restoration of clotting activity in hemophilic mice using a minimal adenovirus vector. Blood 2000, 95, 820–828. [Google Scholar] [CrossRef]
- Wang, L.; Herzog, R.W. AAV-mediated gene transfer for treatment of hemophilia. Curr. Gene Ther. 2005, 5, 349–360. [Google Scholar] [CrossRef]
- Sarkar, R.; Xiao, W.; Kazazian, H.H., Jr. A single adenoassociated virus (AAV)-murine factor FVIII. J. Thromb. Haemost. 2003, 1, 220–226. [Google Scholar] [CrossRef]
- Sarkar, R.; Tetreault, R.; Gao, G.; Wang, L.; Bell, P.; Chandler, R.; Wilson, J.M.; Kazazian, H.H., Jr. Total correction of hemophilia A mice with canine FVIII using an AAV 8 serotype. Blood 2004, 103, 1253–1260. [Google Scholar] [CrossRef]
- Jiang, H.; Lillicrap, D.; Patarroyo-White, S.; Liu, T.; Qian, X.; Scallan, C.D.; Powell, S.; Keller, T.; McMurray, M.; Labelle, A.; et al. Multiyear therapeutic benefit of AAV serotypes 2, 6, and 8 delivering factor VIII to hemophilia A mice and dogs. Blood 2006, 108, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Callan, M.B.; Haskins, M.E.; Wang, P.; Zhou, S.; High, K.A.; Arruda, V.R. Successful Phenotype Improvement following Gene Therapy for Severe Hemophilia A in Privately Owned Dogs. PLoS ONE 2016, 11, e0151800. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.N.; Everett, J.K.; Kafle, S.; Roche, A.M.; Raymond, H.E.; Leiby, J.; Wood, C.; Assenmacher, C.-A.; Merricks, E.P.; Long, C.T.; et al. A long-term study of AAV gene therapy in dogs with hemophilia A identifies clonal expansions of transduced liver cells. Nat. Biotechnol. 2021, 39, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Wilcox, D.A.; Fahs, S.A.; Fang, J.; Johnson, B.D.; Du, L.M.; Desai, D.; Montgomery, R.R. Lentivirus-mediated platelet-derived factor VIII gene therapy in murine haemophilia A. J. Tromb. Haemost. 2007, 5, 352–361. [Google Scholar] [CrossRef]
- Nathwani, A.C. Gene therapy for hemophilia. Hematol. Am. Soc. Hematol. Educ. Program 2019, 2019, 1–8. [Google Scholar] [CrossRef]
- Pasi, K.J.; Rangarajan, S.; Mitchell, N.; Lester, W.; Symington, E.; Madan, B.; Laffan, M.; Russell, C.B.; Li, M.; Pierce, G.F.; Wong, W.Y. Multiyear Follow-up of AAV5-hFVIII-SQ Gene Therapy for Hemophilia A. N. Engl. J. Med. 2020, 382, 29–40. [Google Scholar] [CrossRef]
- VandenDriessche, T.; Pipe, S.W.; Pierce, G.F.; Kaczmarek, R. First conditional marketing authorization approval in the European Union for hemophilia “A” gene therapy. Mol. Ther. 2022, 30, 3335–3336. [Google Scholar] [CrossRef]
- Dai, Y.; Schwarz, E.M.; Gu, D.; Zhang, W.W.; Sarvetnick, N.; Verma, I.M. Cellular and humoral immune responses to adenoviral vectors containing factor IX gene: Tolerization of factor IX and vector antigens allows long-term expression. Proc. Natl. Acad. Sci. USA 1995, 92, 1401–1405. [Google Scholar] [CrossRef]
- Kay, M.A.; Landen, C.N.; Rothenberg, S.R.; Taylor, L.A.; Leland, F.; Wiehle, S.; Fang, B.; Bellinger, D.; Finegold, M.; Thompson, A.R.; et al. In vivo hepatic gene therapy: Complete albeit transient correction of factor IX deficiency in hemophilia B dogs. Proc. Natl. Acad. Sci. USA 1994, 91, 2353–2357. [Google Scholar] [CrossRef]
- Fang, B.; Eisensmith, R.C.; Wang, H.; Kay, M.A.; Cross, R.E.; Landen, C.N.; Gordon, G.; Bellinger, D.A.; Read, M.S.; Hu, P.C.; et al. Gene therapy for hemophilia B: Host immunosuppression prolongs the therapeutic effect of adenovirus-mediated factor IX expression. Hum. Gene Ther. 1995, 6, 1039–1044. [Google Scholar] [CrossRef]
- Crudele, J.M.; Finn, J.D.; Siner, J.I.; Martin, N.B.; Niemeyer, G.P.; Zhou, S.; Mingozzi, F.; Lothrop, C.D. Jr.; Arruda, V.R. AAV liver expression of FIXPadua prevents and eradicates FIX inhibitor without increasing thrombogenicity in hemophilia B dogs and mice. Blood 2015, 125, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Nathwani, A.C.; Reiss, U.M.; Tuddenham, E.G.; Rosales, C.; Chowdary, P.; McIntosh, J.; Della Peruta, M.; Lheriteau, E.; Patel, N.; Raj, D.; Riddell, A.; et al. Long-term safety and efficacy of factor IX gene therapy in hemophilia B. N. Engl. J. Med. 2014, 371, 1994–2004. [Google Scholar] [CrossRef]
- McCarty, D.M.; Monahan, P.E.; Samulski, R.J. Self-complimentary recombinant adeno-associated virus (scAAV) vectors promote efficient transduction independently of DNA synthesis. Gene Ther. 2001, 8, 1248–1254. [Google Scholar] [CrossRef]
- Wu, Z.; Sun, J.; Zhang, T.; Yin, C.; Yin, F.; Van Dyke, T.; Samulski, R.J.; Monahan, R.E. Optimization of self-complementary AAV vectors for liver-directed expression results in sustained correction of hemophilia B at low vector dose. Mol. Ther. 2008, 16, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhu, L.; Zhang, J.; Guo, L.; Sun, X.; Huang, C.; Xu, K.; Zhang, Y.; Li, W.; Zhou, X. Rational engineering of adeno-associated virus capsid enhances human hepatocyte tropismm and reduced immunogenicity. Cell Prolif. 2022, 55, e13339. [Google Scholar] [CrossRef] [PubMed]
- Chowdary, P.; Shapiro, S.; Makris, M.; Evans, G.; Boyce, S.; Talks, K.; Dolan, G.; Reiss, U.; Phillips, M.; Riddell, A.; et al. Phase 1-2 Trial of AAVS3 Gene Therapy in Patients with Hemophilia B. N. Engl. J. Med. 2022, 387, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Cantore, A.; Ranzani, M.; Bartholomae, C.C.; Volpin, M.; Della Valle, P.; Sanvito, F.; Sergi Sergi, L.; Gallina, P.; Benedicenti, F.; Bellinger, D.; et al. Liver-directed lentiviral gene therapy in a dog model of hemophilia B. Sci. Transl. Med. 2015, 7, 277ra28. [Google Scholar] [CrossRef]
- Cavazzana, M.; Mavilio, F. Gene Therapy for Hemoglobinopathies. Hum. Gene Ther. 2018, 29, 1106–1113. [Google Scholar] [CrossRef]
- Lal, A.; Locatelli, F.; Kwiatkowski, J.L.; Porter, J.B.; Trasher, A.J.; Homgeng, S.; Sauer, M.G.; Thuret, I.; Lal, A.; Algeri, M.; et al. Northstar-3: Interim results from a phase 3 study evaluating lentiglobin gene therapy in patients with transfusion-dependent β-thalassemia and either a β0 or IVS-I-110 mutation at both alleles of the HBB gene. Blood 2019, 134, 815. [Google Scholar] [CrossRef]
- Shangaris, P.; Loukogeorkakis, S.P.; Subramaniam, S.; Flouri, C.; Jackson, L.H.; Wang, W.; Blundell, M.P.; Liu, S.; Eaton, S.; Bakhamis, N.; et al. In Utero Gene Therapy (IUGT) Using GLOBE Lentiviral Vector Phenotypically Corrects the Heterozygous Humanised Mouse Model and Its Progress Can Be Monitored Using MRI Techniques. Sci. Rep. 2019, 9, 11592. [Google Scholar] [CrossRef]
- Marktel, S.; Scaramuzza, S.; Cicalese, M.P.; Giglio, F.; Galimberti, S.; Lidonnici, M.R.; Calbi, V.; Assanelli, A.; Bernardo, M.E.; Rossi, C.; et al. Intrabone hematopoietic stem cell gene therapy for adult and pediatric patients affected by transfusion-depend- ent ss-thalassemia. Nat. Med. 2019, 25, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Pawliuk, R.; Westerman, K.A.; Fabry, M.E.; Payen, E.; Tighe, R.; Bouhassira, E.E.; Acharya, S.A.; Ellis, J.; London, I.M.; Eaves, C.J.; et al. Correction of sickle cell disease in transgenic mouse models by gene therapy. Science 2001, 294, 2368–2371. [Google Scholar] [CrossRef] [PubMed]
- Ribeil, J.-A.; Hacein-Bey-Abina, S.; Payen, E.; Magnani, A.; Semeraro, M.; Magrin, E.; Caccavelli, L.; Neven, B.; Bourget, P.; El Nemer, W.; et al. Gene therapy in a patient with sickle cell disease. N. Engl. J. Med. 2017, 376, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Kanter, J.; Walters, M.C.; Krishnamurti, L.; Mapara, M.Y.; Kwiatkowski, J.L.; Rifkin-Zenenberg, S.; Aygun, B.; Kasow, K.A.; Pierciey Jr., F.J.; Bonner, M.; et al. Biologic and Clinical Efficacy of LentiGlobin for Sickle Cell Disease. N. Engl. J. Med. 2022, 386, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Gambari, R. Alternative options for DNA-based experimental therapy of β-thalassemia. Expert Opin. Biol. Ther. 2012, 12, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Ingram, V. A specific chemical difference between the globins of normal human and sickle cell anemia hemoglobin. Nature 1956, 178, 792–794. [Google Scholar] [CrossRef]
- Kim, J.; Yoon, Y.S.; Lee, H.; Chang, W. AAV-GAD gene for rat models of neuropathic pain and Parkinson’s disease. Acta Neurochir. Suppl. 2008, 101, 99–105. [Google Scholar]
- Kaplitt, M.G.; Feigin, A.; Tang, C.; Fitzsimons, H.L.; Mattis, P.; Lawlor, P.A.; Bland, R.J.; Young, D.; Strybing, K.; Eidelberg, D.; et al. Safety and tolerability of gene therapy with an adeno-associated virus (AAV) borne GAD gene for Parkinson’s disease: an open label, phase I trial. Lancet 2007, 369, 2097–2105. [Google Scholar] [CrossRef]
- Björklund, A.; Kirik, D.; Rosenblad, C.; Georgievska, B.; Lundberg, C.; Mandel, R.J. Towards a neuroprotective gene therapy for Parkinson’s disease: Use of adenovirus, AAV and lentivirus vectors for gene transfer of GDNF to the nigrostriatal system in the rat Parkinson model. Brain Res. 2000, 886, 82–98. [Google Scholar] [CrossRef]
- Kordower, J.H.; Emborg, M.E.; Bloch, J.; Ma, S.Y.; Chu, Y.; Leventhal, L.; McBride, J.; Chen, E.Y.; Palfi, S.; Roitberg, B.Z.; et al. Neurodegeneration prevented by lentiviral vector delivery of GDNF in primate model of Parkinson’s disease. Science 2000, 290, 767–773. [Google Scholar] [CrossRef]
- Forsayeth, J.R.; Eberling, J.L.; Sanftner, L.M.; Zhen, Z.; Pivirotto, P.; Bringas, J.; Cunningham, J.; Bankiewicz, K.S. A dose-ranging study of AAV-hAADCtherapy in Parkinsonian monkeys. Mol. Ther. 2006, 14, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Eberling, J.L.; Jagust, W.J.; Christine, C.W.; Starr, P.; Larson, P.; Bankiewicz, K.S.; Aminoff, M.J. Results from a phase I safety trial of hAADC gene therapy for Parkinson disease. Neurology 2008, 70, 1980–1983. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Muramatsu, S.I.; Ikeguchi, K.; Fujimoto, K.I.; Fan, D.S.; Ogawa, M.; Mizukami, H.; Urabe, M.; Kume, A.; Nagatsu, I.; et al. Triple transduction with adeno-associated virus vectors expressing tyrosine hydroxylase, aromatic-L-amino acid decarboxylase, and GTP cyclohydrolase I for gene therapy for Parkinson’s disease. Hum. Gene Ther. 2000, 11, 1509–1519. [Google Scholar] [CrossRef]
- Palfi, S.; Gurruchaga, J.M.; Ralph, G.S.; Lepetit, H.; Lavisse, S.; Buttery, P.C.; Watts, C.; Miskin, J.; Kelleher, M; Deeley, S.; et al. Long-term safety and tolerability of ProSavin, a lentiviral vector-based gene therapy for Parkinson’s disease: A dose escalation, open-label, phase 1/2 trial. Lancet 2014, 383, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Palfi, S.; Gurruchaga, J.M.; Lepetit, H.; Howard, K; Ralph, G.S.; Mason, S.; Gouello, G.; Domenech, P.; Buttery, P.C.; Hantraye, P.; et al. Long-Term Follow-up of a Phase I/II of ProSavin, a Lentiviral Vector Gene Therapy for Parkinson’s Disease. Hum. Gene Ther. Clin. Dev. 2018, 29, 148–155. [Google Scholar] [CrossRef]
- Miniarikova, J.; Zimmer, V.; Martier, R.; Bouwers, C.C.; Pythoud, C.; Richetin, K.; Rey, M.; Lubelski, J.; Evers, M.M.; van Deventer, S.J.; et al. AAV5-miHTT gene therapy demonstrates suppression of huntingtin aggregation and neuronal dysfunction in a rat model of Huntington’s disease. Gene Ther. 2017, 24, 630–639. [Google Scholar] [CrossRef]
- Evers, M.M.; Miniarikova, J.; Juhas, S.; Vallès, A.; Bohuslavova, B.; Juhasova, J.; Skalnikova, H.K.; Vodicka, P.; Valekova, I.; Brouwers, C.; et al. AAV5-miHTT gene therapy demonstrates broad distribution and strong human mutant huntingtin lowering in Huntington’s disease minipig model. Mol. Ther. 2018, 26, 2163–2177. [Google Scholar] [CrossRef]
- Rodrigues, F.B.; Wild, E.J. Huntington’s Disease Clinical Trials Corner: April 2020. J. Huntington’s Dis. 2020, 9, 185–197. [Google Scholar] [CrossRef]
- Passini, M.A.; Bu, J.; Roskelley, E.M.; Richards, A.M.; Pablo Sardi, S.; O’Riordan, C.R.; Klinger, K.W.; Shihabuddin, L.S.; Cheng, S.H. CNS-targeted gene therapy improves survival and motor function in a mouse model of spinal muscular atrophy. J. Clin. Invest. 2010, 120, 1253–1264. [Google Scholar] [CrossRef]
- Pattali, R.; Mou, Y.; Li, X.-J. AAV9 vector: A novel modality in gene therapy for spinal muscular atrophy. Gene Ther. 2019, 26, 287–295. [Google Scholar] [CrossRef]
- Mendell, J.R.; Al-Zaidy, S.; Shell, R.; Arnold, W.D.; Rodino-Klapac, L.R.; Prior, T.W.; Lowes, L.; Alfano, L.; Berry, K.; Church, K.; et al. Single-dose gene-replacement therapy for spinal muscular atrophy. N. Engl. J. Med. 2017, 377, 1713–1722. [Google Scholar] [CrossRef] [PubMed]
- Hoy, S.M. Onasemnogene Abeparvovec First Global Approval. Drugs 2019, 79, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, J.R.; Chamberlain, J.S. Progress toward gene therapy for Duchenne muscular dystrophy. Mol. Ther. 2017, 25, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Yuasa, K.; Yoshimura, M.; Yokota, T.; Ikemoto, T.; Suzuki, M.; Dickson, G.; Miyagoe-Suzuki, Y.; Takeda, S. Micro- dystrophin cDNA ameliorates dystrophic phenotypes when introduced into mdx mice as a transgene. Biochem. Biophys. Res. Comm. 2002, 293, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Gregorevic, P.; Allen, J.M.; Minami, E.; Blankinship, M.J.; Haraguchi, M.; Meuse, L.; Finn, E.; Adams, M.E.; Froehner, S.C.; Murry, C.E.; et al. rAAV6-microdystrophin preserves muscle function and extends lifespan in severely dystrophic mice. Nat. Med. 2006, 12, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Storb, R.; Halbert, C.L.; Banks, G.B.; Butts, T.M.; Finn, E.E.; Allen, J.M.; Miller, A.D.; Chamberlain, J.S.; Tapscott, S.J. Successful regional delivery and long-term expression of a dystrophin gene in canine muscular dystrophy: A preclinical model for human therapies. Mol. Ther. 2012, 20, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Mendell, J.R.; Sahenk, Z.; Lehman, K.; Nease, C.; Lowes, L.P.; Miller, N.F.; Iammarino, M.A.; Alfano, L.N.; Nicholl, A.; Al-Zaidy, S.; et al. Assessment of systemic delivery of rAAVrh74. MHCK7.micro-dystrophin in children with Duchenne muscular dystrophy. JAMA Neurol. 2020, 77, 1122–1131. [Google Scholar] [CrossRef]
- Butterfield, R.; Shieh, P.; Geffen, D.; Yong, F.; Binks, M.; McDonnell, T.G.; Ryan, K.A.; Belluscio, B.; Neelakanten, S.; Levy, D.; et al. One year data from ambulatory boys in a phase 1b, open-label study of fordadistrogene movaparvovec (PF-06939926) for Duchenne muscular dystrophy (DMD). MDA Conference, Poster 53. www.mdaconference.org/abstract-library/one-year-data-from-ambulatory-boys-in-a-phase-1b-open-label-study-of-fordadistrogene-movaparvovec-pf-06939926-for-duchenne- muscular-dystrophy-dmd/.
- Goedeker, N.L.; Dharia, S.D.; Griffin, D.A.; Coy, J.; Truesdale, T.; Parikh, R.; Whitehouse, K.; Santra, S.; Asher, D.R.; Zaidman, C.M. Evaluation of rAAVrh74 gene therapy vector seroprevalence by measurement of total binding antibodies in patients with Duchenne muscular dystrophy. Ther. Adv. Neurol. Disord. 2023, 16, 17562864221149781. [Google Scholar] [CrossRef]
- Cavazzana-Calvo, M.; Hacein-Bey, S.; de Saint Basile, G.; Gross, F.; Yvon, E.; Nusbaum, P.; Selz, F.; Hue, C.; Certain, S.; Casa- nova, J.L.; Bousso, P.; et al. Gene therapy of human severe combined immunodeficiency (SCID)-X1 disease. Science 2000, 28, 669–672. [Google Scholar] [CrossRef]
- Hacein-Bey-Abina, S.; Garrigue, A.; Wang, G.P.; Soulier, J.; Lim, A.; Morillon, E.; Clappier, E.; Caccavelli, L.; Delabesse, E.; Beldjord, K.; et al. Insertional oncogenesis in 4 patients after retrovirus-mediated gene therapy of SCID-X1. J. Clin. Investig. 2008, 118, 3132–3142. [Google Scholar] [CrossRef]
- Fischer, A.; Hacein-Bey-Abina, S. Gene therapy for severe combined immunodeficiencies and beyond. J. Exp. Med. 2020, 217, e20190607. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, H.B.; Parsley, K.L.; Howe, S.; King, D.; Gilmour, K.C.; Sinclair, J.; Brouns, G.; Schmidt, M.; Von Kalle, C.; Barington, T.; et al. Gene therapy of X-linked severe combined immunodeficiency by use of a pseudotyped gammaretroviral vector. Lancet 2004, 364, 2181–2187. [Google Scholar] [CrossRef] [PubMed]
- Hacein-Bey-Abina, S.; Pai, S.Y.; Gaspar, H.B.; Armant, M.; Berry, C.C.; Blanche, S.; Bleesing, J.; Blondeau, J.; de Boer, H.; Buck- land, K.F.; et al. A modified γ-retrovirus vector for X-linked severe combined immunodeficiency. N. Engl. J. Med. 2014, 371, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Howe, S.J.; Mansour, M.R.; Schwarzwaelder, K.; Bartholomae, C.; Hubank, M.; Kempski, H.; Brugman, M.H.; Pike-Overzet, K.; Chatters, S.J.; de Ridder, D.; et al. Insertional mutagenesis combined with acquired somatic mutations causes leukemogenesis following gene therapy of SCID-X1 patients. J. Clin. Investig. 2008, 118, 3143–3150. [Google Scholar] [CrossRef] [PubMed]
- Kohn, D.B.; Hershfield, M.S.; Puck, J.M.; Aiuti, A.; Blincoe, A.; Gaspar, H.B.; Notarangelo, L.D.; Grunebaum, E. Consensus approach for the management of severe combined immune deficiency caused by adenosine deaminase deficiency. J. Allergy Clin. Immunol. 2019, 143, 852–863. [Google Scholar] [CrossRef]
- Kohn, D.B.; Booth, C.; Shaw, K.L.; Xu-Bayford, J.; Garabedian, E.; Trevisan, V.; Carbonaro-Sarracino, D.A.; Soni, K.; Terrazas, D.; Snell, K.; et al. Autologous Ex Vivo Lentiviral Gene Therapy for Adenosine Deaminase Deficiency. N. Engl. J. Med. 2021, 384, 2002–2013. [Google Scholar] [CrossRef]
- Ramasamy, M.N.; Minassian, A.M.; Ewer, K.J.; Flaxman, A.L.; Folegatti, P.M.; Owens, D.R.; Voysey, M.; Aley, P.K.; Angus, B.; Babbage, G.; et al. Safety and immunogenicity of ChAdOx1 nCov-19 vaccine adminsitered in a prime-boost regimen in young and old adults (COV002): A singe-blind, randomised, controlled phase 2/3 trial. Lancet 2020, 336, 1979–1993. [Google Scholar] [CrossRef]
- Zhu, F.C.; Guan, X.H.; Li, Y.H.; Huang, J.Y.; Jiang, T.; Hou, L.H.; Li, J.X.; Yang, B.F.; Wang, L.; Wang, W.J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A random- ised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S.; et al. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19: An interim analysis of a randomised controlled phase 3 in Russia. Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef]
- Stephenson, K.E.; Le Gars, M.; Sadoff, J.; de Groot, A.M.; Heerwegh, D.; Truyers, C.; Atyeo, C.; Loos, C.; Chandrashekar, A.; McMahan, K.; et al. Immunogenicity of the Ad26.COV2.S vaccine for COVID-19. JAMA 2021, 325, 1535–1544. [Google Scholar] [CrossRef]
- Lundstrom, K. Viral Vectors for COVID-19 Vaccine Development. Viruses 2021, 13, 317. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E. Russia’s fast-track coronavirus vaccine draws outrage over safety. Nature 2020, 584, 334–335. [Google Scholar] [CrossRef] [PubMed]
- Henao-Restrepo, A.M.; Longini, I.M.; Egger, M.; Dean, N.E.; Edmunds, W.J.; Camacho, A.; Carroll, M.W.; Doumbia, M.; Draguez, B.; Duraffour, S.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine expressing Ebola surface glycoprotein: Interim results from the Guinea ring vaccination cluster-randomised trial. Lancet 2015, 386, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Henao-Restrepo, A.M.; Camacho, A.; Longini, I.M.; Watson, C.H.; Edmunds, W.J.; Egger, M.; Carroll, M.W.; Dean, N.E.; Diatta, I.; Doumbia, M.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ca Suffit!). Lancet 2017, 389, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Ollmann Saphire, E. A vaccine against Ebola virus. Cell 2020, 181, 6. [Google Scholar] [CrossRef]
- Maclachlan, T.K.; Lukason, M.; Collins, M.; Munger, R.; Isenberger, E.; Rogers, C.; Malatos, S.; Dufresne, E.; Morris, J.; Calcedo, R.; et al. Preclinical safety evaluation of AAV2-sFFLT101- a gene therapy for age-related macular degeneration. Mol Ther. 2011, 19, 326–334. [Google Scholar] [CrossRef]
- Heier, J.S.; Kherani, S.; Desai, S.; Dugel, P.; Kaushal, S.; Cheng, S.H.; Delacono, C.; Purvis, A.; Richards, S.; Le-Halpere, A.; et al. Intravitreous injection of AAV2-sFLT01 in patients with advanced neovascular age-related macular degeneration: A phase I, open-label trial. Lancet 2017, 390, 50–61. [Google Scholar] [CrossRef]
- Constable, I.J.; Pierce, C.M.; Lai, C.-M.; Magno, A.L.; Degli-Esposti, M.A.; French, M.A.; McAllister, I.L.; Butler, S.; Barone, S.B.; Schwartz, S.D.; et al. Phase 2a randomized clinical trial: Safety and post hoc analysis of subretinal rAAV.sFLT-1 for wet age-related macular degeneration. EBioMedicine 2016, 14, 168–175. [Google Scholar] [CrossRef]
- Guy, J.; Feuer, W.J.; Davis, J.L.; Porciatti, V.; Gonzalez, P.J.; Koilkonda, R.D.; Yuan, H.; Hauswirth, W.W.; Lam, B.L. Gene therapy for Leder hereditary optic neuropathy: Low and medium-dose visual results. Ophthalmology 2017, 124, 1621–1634. [Google Scholar] [CrossRef]
- Vignal, C.; Uretsky, S.; Fitoussi, S.; Galy, A.; Blouin, L.; Girmens, J.-F.; Bidot, S.; Thomasson, N.; Bouquet, C.; Valero, S.; et al. Safety of rAAV2/2-ND4 gene therapy. Ophthalmology 2018, 125, 945–947. [Google Scholar] [CrossRef]
- Russell, S.; Bennett, J.; Wellman, J.A.; Chung, D.C.; Yu, Z.F.; Tillman, A.; Wittes, J.; Pappas, J.; Elci, O.; McCague, S.; et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65-mediated inherited retinal dystrophy: A randomised, controlled, open-label, phase 3 trial. Lancet 2017, 390, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Maguire, A.M.; Bennett, J.; Aleman, E.M.; Leroy, B.P.; Aleman, T.S. Clinical Perspective: Treating RPE65-Associated Retinal Dystrophy. Mol. Ther. 2021, 29, 442–463. [Google Scholar] [CrossRef]
- Flotte, T.R.; Afione, S.A.; Conrad, C.; McGrath, S.A.; Solow, R.; Oka, H.; Zeitlin, P.L.; Guggino, W.B.; Carter, B.J. Stable in vivo expression of the cystic fibrosis transmembrane conductance regulator with an adeno-associated virus vector. Proc. Natl. Acad. Sci. USA 1993, 90, 10613–10617. [Google Scholar] [CrossRef] [PubMed]
- Conrad, C.K.; Allen, S.S.; Afione, S.A.; Reynolds, T.C.; Beck, S.E.; Fee-Maki, M.; Barazza-Ortiz, X.; Adams, R.; Askin, F.B.; Carter, B.J.; et al. Safety of single-dose administration of an adeno-associated virus (AAV)-CFTR vector in the primate lung. Gene Ther. 1996, 3, 658–668. [Google Scholar] [PubMed]
- Limberis, M.; Anson, D.S.; Fuller, M.; Parsons, D.W. Recovery of airway cystic fibrosis transmembrane conductance regulator function in mice with cystic fibrosis after single-dose lentivirus-mediated gene transfer. Hum. Gene Ther. 2002, 13, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Cooney, A.L.; Abou Alaiwa, M.H.; Shah, V.S.; Bouzek, D.C.; Stroik, M.R.; Powers, L.S.; Gansemer, N.D.; Meyerholz, D.K.; Welsh, M.J.; Stoltz, D.A.; et al. Lentiviral-mediated phenotypic correction of cystic fibrosis pigs. JCI Insight 2016, 1, e88730. [Google Scholar] [CrossRef] [PubMed]
- Agapov, E.V.; Frolov, I.; Lindenbach, D.B.; Pragai, B.M.; Schlesinger, S.; Rice, C.M. Noncytopathic Sindbis virus RNA vectors for heterologous gene expression. Proc. Natl. Acad. Sci. USA 1998, 95, 12989–12994. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K.; Abenavoli, A.; Malgaroli, A.; Ehrengruber, M.U. Novel Semliki Forest virus vectors with reduced toxicity and temperature-sensitivity for long-term enhancement of transgene expression. Mol. Ther. 2003, 7, 202–209. [Google Scholar] [CrossRef]
- Perkovic, M.; Gawletta, S.; Hempel, T.; Brill, S.; Nett, E.; Sahin, U.; Beissert, T. A trans-amplifying RNA simplified to essential elements is highly replicative and robustly immunogenic in mice. Mol. Ther. 2023. [Google Scholar] [CrossRef]
- Lundstrom, K. Therapeutic Applications for Oncolytic Self-Replicating RNA Viruses. Int. J. Mol. Sci. 2022, 23, 15622. [Google Scholar] [CrossRef]
- Xiang, Q.; Li, L.; Wu, J.; Tian, M.; Fu, Y. Application of pseudovirus system in the development of vaccine, antiviral-drugs, and neutralizing antibodies. Microbiol. Res. 2022, 258, 126993. [Google Scholar] [CrossRef]
- Morse, M.A.; Crosby, E.J.; Force, J.; Osada, T.; Hobeika, A.C.; Hartman, Z.C.; Berglund, P.; Smith, J.; Lyerly, H.K. Clinical trials of self-replicating RNA-based cancer vaccines. Cancer Gene Ther. 2023, 10, 1–9. [Google Scholar]
- Chen, L.; Ewing, D.; Subramanian, H.; Block, K.; Rayner, J.; Alterson, K.D.; Sedegah, M.; Hayes, C.; Porter, K.; Raviprakash, K. A heterologous DNA prime-Venezuelan equine encephalitis virus replicon particle boost dengue vaccine regimen affords complete protection from virus challenge in cynomolgus macaques. J. Virol. 2007, 81, 11634–11639. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.K.; Burgess, A.W.; Clayton, A.H.; Scott, A.M. Targeting of a conformationally exposed, tumor-specific epitope of EGFR as a strategy for cancer therapy. Cancer Res. 2012, 72, 2924–2930. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, H.; Zou, H.; Tian, X.; Hu, J.; Qiu, P.; Ferreira, L. Liposome Encapsulation of Oncolytic Virus M1 To Reduce Immunogenicity and Immune Clearance in Vivo. Mol. Pharm. 2019, 16, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Boulikas, T.; Lundstrom, K.; Soling, P.C.; Warnke, P.C.; Rainov, N.G. Immunogene therapy of recurrent glioblastoma multiforme with a liposomally encapsulated replication-incompetent Semliki Forest virus vector carrying the human interleukin-12 gene—a phase I/II clinical protocol. J. Neuro-Oncol. 2003, 64, 147–154. [Google Scholar] [CrossRef]
- Van der Loo, J.C.; Wright, J.F. Progress and challenges in viral vector manufacturing. Hum. Mol. Genet. 2016, 25, R42–52. [Google Scholar] [CrossRef]
- Saeki, Y.; Fraefel, C.; Ichikawa, T.; Breakfield, X.O.; Chiocca, E.A. Improved helper virus-free packaging system for HSV amplicon vectors using an ICP27-deleted, oversized HSV-1 DNA in a bacterial artificial chromosome. Mol. Ther. 2001, 3, 591–601. [Google Scholar] [CrossRef]
- Polo, J.M.; Belli, B.A.; Driver, D.A.; Frolov, I.; Sherill, S.; Hariharan, M.J.; Townsend, K.; Perri, S.; Mento, S.J.; Jolly, D.J.; et al. Stable alphavirus packaging cell lines for Sindbis virus and Semliki Forest virus-derived vectors. Proc. Natl. Acad. Sci. USA 1999, 96, 4598–4603. [Google Scholar] [CrossRef]
- Guerriaud, M.; Kohli, E. RNA-based drugs and regulation: Toward a necessary evolution of the definitions issued from the European Union legislation. Front. Med. 2022, 9, 1012497. [Google Scholar] [CrossRef]
- Food and Drug Administration. Personal Communication: Questions About mRNA Drugs; Food and Drug Administration,: Silver Spring, MD:, 2022. [Google Scholar]
- Celis, P. Personal Communication: mRNA Drugs Regulation and Categorization.; European Medicines Agency,: ATMP Office of EMA Amsterdam, 2022. [Google Scholar]
- Li, C.; Wang, H.; Georgakopoulou, A.; Gil, S.; Yannaki, E.; Lieber, A. In Vivo HSC Gene Therapy Using a Bi-modular HDAd5/35++ Vector Cures Sickle Cell Disease in a Mouse Model. Mol. Ther. 2021, 29, 822–837. [Google Scholar] [CrossRef] [PubMed]
- Tarone, L.; Barutello, G.; Iussich, S.; Giacobino, D.; Quaglino, E.; Buracco, P.; Cavallo, F.; Riccardo, F. Naturally occurring cancers in pet dogs as pre-clinical models for cancer immunotherapy. Cancer Immunol. Immunother. 2019, 68, 1839–1853. [Google Scholar] [CrossRef] [PubMed]
- Barthélémy, F.; Wein, N. Personalized gene and cell therapy for Duchenne Muscular Dystrophy. Neuromusc. Disorders 2018, 28, 803–824. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Recombinant protein expression from viral vectors utilizing the host cell nucleus. Expression systems for adenovirus (Ad), polyomavirus (exemplified by simian virus 40 (SV40)), poxvirus (exemplified by vaccinia virus (VV)), herpes simplex virus (HSV), adeno-associated virus (AAV), and retrovirus/lentivirus (RV/LV). dsDNA, double-stranded DNA; ssDNA, single-stranded DNA; ssRNA, single-stranded RNA.
Figure 1.
Recombinant protein expression from viral vectors utilizing the host cell nucleus. Expression systems for adenovirus (Ad), polyomavirus (exemplified by simian virus 40 (SV40)), poxvirus (exemplified by vaccinia virus (VV)), herpes simplex virus (HSV), adeno-associated virus (AAV), and retrovirus/lentivirus (RV/LV). dsDNA, double-stranded DNA; ssDNA, single-stranded DNA; ssRNA, single-stranded RNA.
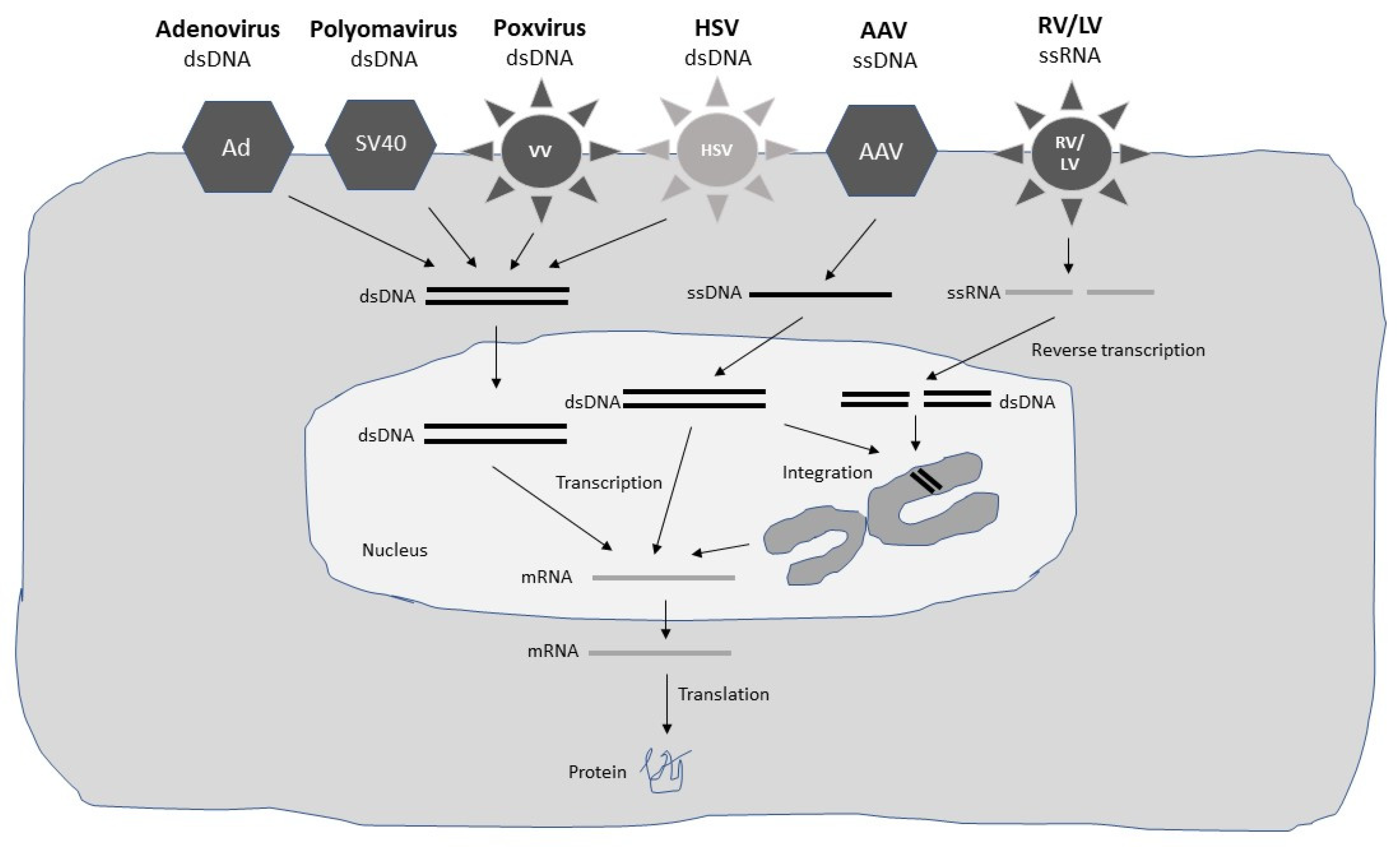
Figure 2.
Recombinant protein expression from viral vectors independent of the host cell nucleus. Expression systems for alphavirus/flavivirus (exemplified by Semliki Forest virus (SFV)), paramyxovirus/rhabdovirus (exemplified by vesicular stomatitis virus (VSV)), reovirus (Reo), Newcastle disease virus (NDV), Picornavirus exemplified by coxsackievirus A (CVA). dsRNA, double-stranded RNA; ssDNA; ssRNA+, positive single-stranded RNA; ssRNA-, negative single-stranded RNA.
Figure 2.
Recombinant protein expression from viral vectors independent of the host cell nucleus. Expression systems for alphavirus/flavivirus (exemplified by Semliki Forest virus (SFV)), paramyxovirus/rhabdovirus (exemplified by vesicular stomatitis virus (VSV)), reovirus (Reo), Newcastle disease virus (NDV), Picornavirus exemplified by coxsackievirus A (CVA). dsRNA, double-stranded RNA; ssDNA; ssRNA+, positive single-stranded RNA; ssRNA-, negative single-stranded RNA.
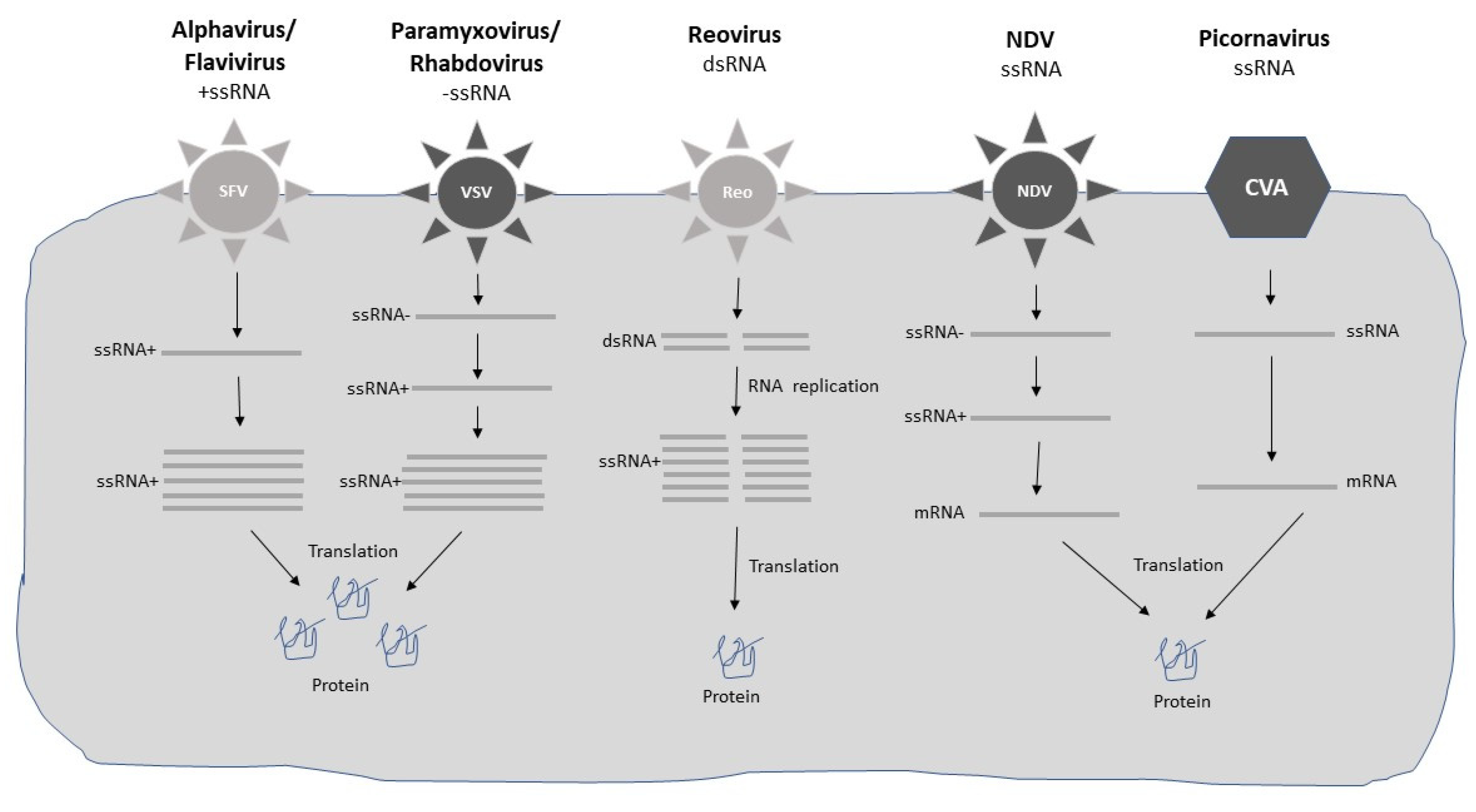

Table 1.
Examples of viral vectors applied for gene therapy.
| Virus | Genome | Capacity | Features |
|---|---|---|---|
| Ad | dsDNA | 7.5-37 kb | Extrachromosomal, short- and long-term expression [14] |
| AAV HSV RV LV Alphavirus Flavivirus Paramyxovirus Rhabdovirus NDV Poxvirus Picornavirus Reovirus Polyomavirus |
ssDNA dsDNA ssRNA ssRNA ssRNA ssRNA ssRNA ssRNA ssRNA dsDNA ssRNA dsRNA dsDNA |
4 kb 30-150 kb 8 kb 8 kb 8 kb 6 kb 6 kb 6 kb 4 kb > 30 kb 6 kb ND 17.7 kb |
Extrachromosomal, integration for long-term expression [10] Extrachromosomal, long-term expression, latency [11] Integration, long-term expression [12] Integration, long-term expression, transduction of non-dividing cells [13] Extreme short-term expression, RNA degradation [18] Extreme short-term expression, RNA degradation [19] Short-term expression, RNA degradation [20] Short-term expression, RNA degradation [21] Short-term expression, oncolytic activity [22] Extrachromosomal expression [23] Extrachromosomal expression [24,25] Extrachromosomal expression [24] Extrachromosomal expression [23] |
AAV, adeno-associated virus; Ad, adenovirus; HSV, herpes simplex virus; LV, lentivirus; NDV, Newcastle disease virus; RV, retrovirus.
Table 2.
Examples of viral vectors applied for cancer gene therapy.
| Cancer | Vector | Target | Findings |
|---|---|---|---|
| Esophageal | Ad | p53 | Significant tumor suppression of tumor growth in nude mice [50] |
| Ovarian HNSCC Head & neck Melanoma Melanoma Melanoma HGG HGG HGG Sarcoma Solid tumors Melanoma Glioma Breast Prostate Ovarian Prostate Prostate |
Ad Ad Ad HSV T-VEC HSV T-VEC HSV T-VEC RRV Toca 511 RRV Toca 511 RRV Toca 511 Reovirus Reovirus Reovirus Reovirus Reovirus MV MV VEE VEE |
p53 p53 p53 GM-CSF GM-CSF GM-CSF yCD yCD yCD Reolysin Reolysin Reolysin Reolysin Reolysin CEA CEA PSMA PSMA |
Prolonged survival in mice with implanted tumors [51] Safe and promising results in clinical trials [52] Approved as GendicineTM for head & neck cancer in China [53] Enhanced tumor growth inhibition, prolonged survival in mice [55] Minor adverse events, good therapeutic efficacy in Phase II and III [56] Approval for melanoma treatment in the US, Europa, and Australia [57] Extended survival in mice with orthotopic gliomas [58] Prolonged survival of 13.6 months in HGG patients in phase I [59] No overall extension of survival in HGG patients in phase II/III [60] Inhibition of tumor growth in nude mice [61] Clinical benefits in phase I: 1 PR, 7 SD [62] Good safety and clinical efficacy in melanoma patients in phase II [63] Orphan drug designation for malignant glioma [64] Fast Track designation for metastatic breast cancer [65] Delayed tumor growth, prolonged survival in mice [66] SD in all 9 patients, prolonged OS survival of 12.15 months in phase I [67] Robust PSMA-specific immune responses in mice [68] Good safety, and tolerability, but weak immunogenicity in phase I [69] |
AAV, adeno-associated virus; Ad, adenovirus; CEA, carcinoembryonic antigen; GM-CSF, granulocyte macrophage-colony stimulating factor; HGG, high-grade glioma; HNSCC, head & neck squamous cell carcinoma; HSV, herpes simplex virus; LV, lentivirus; MV, measles virus; NDV, Newcastle disease virus; OS, overall survival; PR, partial response, PSMA, prostate-specific membrane antigen; RRV, replicating retrovirus; RV, retrovirus; SD, stable disease; VEE, Venezuelan equine encephalitis virus; yCD, yeast cytosine deaminase.
Table 3.
Examples of viral vectors applied for gene therapy of cardiovascular and metabolic diseases.
Table 3.
Examples of viral vectors applied for gene therapy of cardiovascular and metabolic diseases.
| Disease | Vector | Target | Findings |
|---|---|---|---|
| Heart failure | Ad | SERCa2a | Restored systolic & diastolic heart functions rat heart model [71] |
| Heart failure Heart failure Heart failure Heart failure Heart failure DMT1 AAT AAT |
AAV1 LV AAV1 AAV1 AAV1 AAV2 AAV2 AAV2 |
SERCa2a SERCa2a SERCa2a SERCa2a SERCa2a hAAT hAAT hAAT |
Enhanced coronary blood flow in pig heart failure model [72] Improved systolic & diastolic functions, reduced mortality in rats [73] Improved functional, symptomatic and ventricular activity in phase I [74] Improved walking, peak oxygen consumption in phase II [75] Reduced cardiovascular events and deaths in Phase II [76] Reduced insulitis, insulin autoantibodies, and DTM1 in mice [77] Sustained AAT expression > 1 year in AAT patients in phase I [78] Immunostaining of AAT in AAT patients in phase II [79] |
AAV, adeno-associated virus; Ad, adenovirus; DMT1, diabetes mellitus type 1; HSV, herpes simplex virus; LV, lentivirus; MV, measles virus; NDV, Newcastle disease virus; OS, overall survival; PR, partial response, PSMA, prostate-specific membrane antigen; RRV, replicating retrovirus; RV, retrovirus; SD, stable disease; SERCa2a, sarcoplasmic reticulum Ca2+ ATPase; VEE, Venezuelan equine encephalitis virus; yCD, yeast cytosine deaminase.
Table 4.
Examples of viral vectors applied for gene therapy of hematological diseases.
| Disease | Vector | Target | Findings |
|---|---|---|---|
| Hemophilia A | Ad | FVIII | Expression of physiological levels of FVIII in mice [83] |
| Hemophilia A Hemophilia A Hemophilia A Hemophilia A Hemophilia A Hemophilia A Hemophilia A Hemophilia B Hemophilia B Hemophilia B Hemophilia B Hemophilia B Hemophilia B Hemophilia B Hemophilia B Hemophilia B β-thalassemia β-thalassemia β-thalassemia β-thalassemia SCD SCD SCD |
AAV6/AAV8 AAV8 AAV8/AAV9 LV-BM AAV AAV5 AAV5 Ad Ad Ad +CsA AAV8 AAV8 scAAV2 scAAV2 AAVS3 SIN-LV LentiGlobin LentiGlobin GLOBE LV GLOBE LV LV-HSC LentiGlobin LentiGlobin |
FVIII FVIII FVIII FVIII FVIII FVIII-SQ FVIII FIX cFIX cFIX FIX FIX FIX FIX FIX FIX HbAT87Q HbAT87Q Mini-β Mini-β βA globin HbAT87Q HbAT87Q |
Therapeutic FVIII levels lasting for > 3 years in dogs [87] 1-2% of normal FVIII levels, 90% reduction of bleeding in dogs [88] 1.9-11.3% of normal FVIII levels in dogs [89] Sustained FVIII activity, hemophilia A phenotype correction in mice [90] 8-60% of normal FVIII levels in hemophilia A patients in phase II [91] Clinical benefits: less bleeding, no need for prophylactic FVIII [92] Conditional EMA marketing approval for severe hemophilia A [93] Expression of FIX for > 300 days in mice [94] Complete correction of hemophilia B phenotype in dogs [95] Restored therapeutic FIX for 6 months with CsA in dogs [96] 25-200% FIX activity, correction of hemophilic phenotype in dogs [97] 1-6% of normal FIX, reduced bleeding episodes in patients in phase I [98] Correction of coagulation function in FIX-deficient mice [100] Significant reduction of bleeding episodes in hemophilia B patients [91] FIX expression for 27 months in hemophilia B patients in phase I/II [102] Stable long-term FIX expression in dogs [103] Blood transfusions terminated in β-thalassemia patients in phase I [104] Independence of transfusions in β-thalassemia patients in phase III [105] Normalized phenotype after in utero gene therapy [106] Reduced or no need for transfusion in β-thalassemia patients [107] Anti-sickling protein expression for 10 months in mice [108] Case report of complete remission of SCD patient [109] Complete resolution of vaso-occlusive events in SCD patients [110] |
AAV, adeno-associated virus; AAVS3, AAV vector with synthetic capsid; Ad, adenovirus; cFIX, canine factor IX; CsA, cyclosporin A; EMA, European Medicines Agency; FIX, factor IX; FVIII, factor VIII; LV, lentivirus; LV-BM, lentivirus transduced bone marrow; Mini-β, mini-β-globin gene; scAAV2, self-complementary AAV2; SCD, sickle cell disease; SIN-LV, self-inactivating LV.
Table 5.
Examples of viral vectors applied for gene therapy of neurological disorders.
| Disease | Vector | Target | Findings |
|---|---|---|---|
| PD PD PD PD PD PD PD PD PD PD PD PD HD HD HD SMA SMA SMA SMA |
AAV AAV AAV LV AAV LV LV AAV AAV AAV LV LV AAV5 AAV5 AAV5 AAV8 AAV8 AAV8 AAV9 |
GAD65 GAD GDNF GDNF GDNF GDNF GDNF hAADC hAADC TH, GCH, hAADC ProSavin ProSavin miHTT miHTT miHTT hSMN hSMN hSMN hSMN1 |
Reduced PD symptoms and relief of pain in rat models [113] Safe, improved motor neuron functions in PD patients in phase I [114] Regeneration, functional activity in 6-OHDA-lesioned rats [115] Regeneration, functional activity in 6-OHDA-lesioned rats [115] Regeneration, functional activity in MTTP-lesioned primates [115] Regeneration, functional activity in MTTP-lesioned primates [115] Reversed functional and motor deficits in macaques [116] 50% improvement in L-Dopa responsiveness in primates [117]. Significant improvement in PD patients for 2 years [118] Enhanced BH4 and dopamine production, improved rotational behavior in rats [119] Significant motor function improvement in PD patients [120] Long-term (4 years) motor function improvement in PD patients [121] Prevention of mutant HTT, decrease in neuronal dysfunction in rats [122] Reduced HTT mRNA and protein levels in transgenic minipigs [123] Good safety and tolerability in HD patients in phase I/II [124] Improved muscle strength, coordination and locomotion in mice [125] Improved motor function and prolonged survival in SMA patients [126] Improved motor function and prolonged survival in SMA patients [127] Approval for SMA patients in the US, the EU and Canada [128] |
AAV, adeno-associated virus; Ad, adenovirus; GAD65, glutamic acid decarboxylase 65; GCH, GTP-cyclohydroxylase-1; GDNF, glial-derived neurotrophic factor; hAADC, human aromatic-L-amino acid decarboxylase; HD, Huntington’s disease; LV, lentivirus; miHTT, huntingtin-specific microRNA; PD, Parkinson’s disease; ProSavin, TH/hAADC/GCH combination; SMA, spinal muscular atrophy; SMN, survival motor neuron; TH, tyrosine hydroxylase.
Table 6.
Examples of viral vectors applied for gene therapy of muscular diseases.
| Disease | Vector | Target | Findings | |
|---|---|---|---|---|
| Muscular | ||||
| Dystrophy | AAV6 | µDys | Restored Dys expression, reduced muscle pathology in mice [131] | |
| Dystrophy | AAV6 | µDys | Efficient Dys distribution in skeletal muscles in a canine model [132] | |
| DMD | rAAVrh74 | µDys | Therapeutic Dys levels, improved NSAA scores in 4 DMD patients [133] | |
| DMD | AAV9 | mini-Dys | Phase I in progress in 4-12-year-old DMD patients [134] | |
| Immunodeficiency | ||||
| SCID-X1 | γRV | IL2RG | SCID-X1 correction in pediatric patients [136], few leukemia cases [137] | |
| SCID-X1 | γRV | IL2RG | Long-term clinical benefits in 8 out of 10 patients [136] | |
| SCID-X1 | γRV | IL2RG | Normal growth, and protection against infections after 18 years [138] | |
| SCID-X1 | γRV | IL2RG | Unfavroable chromosomal integration causing T-ALL [139] | |
| SCID-X1 | SIN-γRV | IL2RG | No cases of leukemia in 9 treated SCID-X1 patients [141] | |
| SCID-X1 | SIN-LV | IL2RG | No cases of leukemia in 44 treated SCID-X1 patients [141] | |
| ALD | SIN-LV | ABCD1 | Prevention of cerebral demyelination, clinical benefits [140] | |
| ALD | SIN-γRV | ADA | Metabolic correction, high OS in ADA-SCID patients [142] | |
| ADA-SCID | SIN-LV | ADA | Metabolic correction, high OS in ADA-SCID patients [143] |
AAV, adeno-associated virus; ABCD1, adenosine triphosphate-binding cassette transporter; ADA, adenosine deaminase; ADA-SCID, adenosine deaminase-severe combined immunodeficiency; ALD, X-linked adrenoleukodystrophy; DMD, Duchenne muscular dystrophy; γRV, gamma retrovirus; µDys, micro-dystrophin; T-ALL, T-cell acute lymphoblastic leukemia.
Table 7.
Examples of viral vectors applied for gene therapy of infectious diseases.
| Disease | Vector | Target | Findings |
|---|---|---|---|
| COVID-19 | ChAdOx1 nCoV-19 | SARS-CoV-2 S | Good safety and 62-90% efficacy in Phase III [144] |
| COVID-19 COVID-19 COVID-19 COVID-19 COVID-19 COVID-19 COVID-19 EVD EVD EVD |
Ad5-S-nb2 rAd26-S/rAd5-S Ad26.COV2.S ChAdOx1 nCoV-19 Ad5-S-nb2 rAd26-S/rAd5-S Ad26.COV2.S VSV-ZEBOV VSV-ZEBOV VSV.ZEBOV |
SARS-CoV-2 S SARS-CoV-2 S SARS-CoV-2 S SARS-CoV-2 SARS-CoV-2 SARS-CoV-2 SARS-CoV-2 EBOV GP EBOV GP EBOV GP |
Good safety and efficacy in Phase III [145] Good safety and efficacy in Phase III [146] Good vaccine efficacy after a single dose [147] EUA in the UK in December 2020 [148] EUA in China in February 2021 [148] Approval in Russia in August 2020 [149] EUA in the US in February 2021 [148] Good safety and efficacy in Phase III [150] Good safety and efficacy in Phase III [151] Approval for EVD in 2020 [152] |
Ad, adenovirus; EBOV, Ebola virus; EUA, Emergency Use Authorization; EVD, Ebola virus disease; VSV, vesicular stomatitis virus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



