Submitted:
20 April 2023
Posted:
21 April 2023
You are already at the latest version
Abstract
Seed germination and seedling growth represent two vulnerable stages in a plant life cycle and it is important to determine how they will be affected by environmental changes associated with climate change. This study examines the impact of salinity levels (0, 50, 100, and 150 mM) at each of three temperatures (15, 25, and 35˚C) on seed germination and radicle growth for three dormant and three non-dormant varieties of conventional, glyphosate resistant, or reduced lignin alfalfa. The genetic basis and phenotypic plasticity for seed germination and seedling growth in response to salinity and temperature were determined. Both traits were phenotypically plastic with respect to temperature and salinity. Seed germination was 87.0% ± 1.7 at 25˚C, 0mM (no salinity), but decreased to 33.9% ± 3.9 germination at 35˚C and 150mM. Radicle length went from 36.7 ± 1.5 mm after four days at 25˚C, 0mM, to 10.5 ± 0.7 mm at 35˚C, 150mM. The phenotypic response brought seed germination and radicle length away from their optimum, but we detected standing genetic variation for both seed germination and radicle length. Selection to increase both traits at high temperature and salinity would facilitate alfalfa establishment under climate change.
Keywords:
Conventional alfalfa
; dormant variety
; glyphosate resistant alfalfa
; Medicago sativa
; non-dormant variety
; reduced lignin alfalfa
; salinity
; seed germination
; seedling growth
; temperature
1. Introduction
The impact of climate change on ecosystems, communities, and individual plants and animals is rapidly accumulating [1,2,3]. Abiotic factors such as water and temperature are expected to get modified under climate change, with an increase in temperature and a decrease in water availability predicted in some environments [4]. Heat stress and high salinity represent two major abiotic stresses for plants and higher temperature is often associated with greater water evaporation and increased soil salinity [5,6,7]. In warm climates, crop irrigation tends to increase soil salinity, creating an interaction between warm temperature, water availability, and soil salinity [6]. Higher temperature and increased salinity are expected to reduce crop yield and modify the areas available for crop cultivation [8,9,10].
Seed germination and seedling growth represent vulnerable stages in a plant life cycle. It is therefore important to examine the impact of heat stress and salinity on these life stages, as they could limit seedling establishment and negatively impact crop yield [9]. Heat stress affects the physiology and metabolism of plant cells by altering membrane permeability and affecting primary growth, such as cell differentiation, elongation, and expansion [11,12]. Extreme variation in temperature negatively impairs plant development and fruit set [5]. High saline levels in the soil affects the ability of plants to germinate and grow [13], and salinity affects crop production [7]. The surface of the soil, where seeds tend to germinate, is often more saline than the subsequent layers, where established plants grow [14].
Alfalfa (Medicago sativa) is widely cultivated as a forage crop, and ranks economically as the third or fourth most valuable field crop in the USA [15] with the 2017 production valued around $9.3 billion [16]. Successful establishment of alfalfa largely depends on successful germination where a radicle emerges that will develop as the primary root of the plant [17]. Approximately 50% of US alfalfa production is at least partly irrigated, and this percentage is expected to increase as water availability decreases. In some areas of the world, including China and Australia, alfalfa is mostly irrigated [15]. Irrigation can increase salinity which in turn has been shown to decrease seed germination and seedling growth of alfalfa cultivars [13,15,18,19,20]. The impact of salinity on alfalfa can be genotype dependent [21], and can vary with the developmental stage of the plant [15,19]. Alfalfa seed germination is also negatively impacted by higher temperatures, and the effects of temperature and salinity can interact with higher temperatures suppressing germination at higher salinity levels [14,22].
The genetic diversity of the alfalfa germplasm is high [23,24,25]; salt tolerance is polygenic, with some potential loci for salt tolerance identified [26]. Moreover, in addition to conventional varieties of alfalfa, genetically engineered (GE) varieties exist, for glyphosate resistance, and for reduced lignin content. The glyphosate resistant or Roundu Ready alfalfa has a CTP2-CP4 ESPS gene which confers resistance to glyphosate [27]. The reduced lignin alfalfa, released as HarvXtra, improves the digestibility of alfalfa to cows [28]. The reduction in lignin is attained via the downregulation of the gene encoding caffeoyl-CoA 3-O-methyltransferase (CCoAOMT), a key enzyme in the lignin biosynthesis pathway [28]. The reduced lignin alfalfa varieties also contain the glyphosate resistance allele. Within each of these three categories (conventional, glyphosate resistant, and reduced lignin), there exist both dormant and non-dormant varieties. Fall dormancy (FD) is scaled from 1-11 with 11 representing very non-dormant varieties. The latter have high crop yield under warm conditions, the largest height in the fall, and are not winter-hardy; in contrast, dormant varieties have reduced shoot elongation in the fall and survive the winter well [29].
The response of seed germination and radicle length to changes in temperature and salinity associated with climate change will depend on whether the traits are phenotypically plastic in response to these abiotic factors, and whether standing genetic variation exists for selection to act on [30,31,32]. Phenotypic plasticity provides a quick response to a change in an abiotic factor, while changes in a trait resulting from selection on standing genetic variation occurs more slowly [33]. In addition, selection can act on phenotypic plasticity if there exists standing genetic variation for phenotypic plasticity [34]. Phenotypic plasticity and selection on a trait can interact as a plant adjusts to new environmental conditions. For example, if phenotypic plasticity moves the trait towards the new optimum, the strength of natural selection on the trait is expected to decrease, leading to a slower rate of change toward the new optimum phenotype [35].
In the present study, we examine the impact of salinity level (0, 50, 100, 150 mM) on seed germination and radicle length, at each of three temperatures (15°C, 25°C, 35°C), for 18 alfalfa varieties or breeding stocks of different categories (conventional, glyphosate resistant, and reduced lignin alfalfa), and dormancy types (dormant and non-dormant). We predict increasing temperature and salinity will decrease seed germination and radicle length, but non-dormant varieties will outperform dormant varieties. By growing the same alfalfa varieties at different temperatures and salinity levels, this study examines the phenotypic response of seed germination and radicle length to changes in these environmental factors. In addition, by examining different dormant and non-dormant varieties of conventional and genetically modified alfalfa growing under similar conditions, this study examines whether standing genetic variation exist for these traits. With the impending impact of climate change on crops, it is important to determine how early stages in the alfalfa life cycle (seed germination and seedling growth) may be affected, and understand the venues available for alfalfa to adjust to the associated changes in abiotic factors, including temperature and salinity.
2. Results
2.1. Seed Germination
With temperature in the model, we observed a statistically significant main effect on seed germination for temperature, category, dormancy, and salinity (Table 1). Non-dormant varieties had slightly greater seed germination (74.9% ± 1.6) relative to dormant varieties (71.1 ± 2.3) (N= 108) (Table 1- dormancy). Because the varieties were grown under similar conditions, these results suggest the presence of genetic variation in seed germination between dormancy types.
Category also affected seed germination with reduced lignin breeding stocks having greater seed germination relative to conventional and glyphosate resistant alfalfa (Table 1; Figure 1A). However, we detected a statistically significant interaction between the effects of category and dormancy on seed germination (Table 1). The difference in seed germination between dormant and non-dormant varieties was statistically significant for glyphosate resistant varieties (p = 0.0042), borderline for the conventional varieties (p = 0.063), and non-significant for reduced lignin alfalfa (p = 0.58) (Figure 1B). These results further support the presence of genetic variation for seed germination.
Seed germination had a phenotypic response to both temperature (Table 1- temperature) and salinity (Table 1- salinity). Seed germination was lowest at 35°C (53.3% ± 2.8) (Figure 1C), and decreased with increasing salinity (Figure 1D). However, temperature and salinity interacted on their effects on seed germination such that the decrease in seed germination with increasing salinity was only statistically significant at 35°C (Table 1- temperature X salinity; Figure 2). Seed germination was lowest at 35°C and 150mM salinity (34%).
The phenotypic response of seed germination to temperature varied between dormancy types (Table 1- temperature X dormancy). This indicated the presence of genetic variation in the phenotypic response of seed germination to temperature. Non-dormant varieties only had greater seed germination relative to dormant varieties at 35°C and seed germination was low for both dormancy levels at 35°C (Figure 1E).
Figure 1.
Seed germination (%) (means ± se) per A. Category (N=72), B. Category and dormancy type (N= 36), C. Temperature (N= 72), D. Salinity (N= 54), and E. Temperature and dormancy type (N= 36). The variable N is the sample size. Different letters indicate statistically different values based on Tukey multiple means comparison tests.
Figure 1.
Seed germination (%) (means ± se) per A. Category (N=72), B. Category and dormancy type (N= 36), C. Temperature (N= 72), D. Salinity (N= 54), and E. Temperature and dormancy type (N= 36). The variable N is the sample size. Different letters indicate statistically different values based on Tukey multiple means comparison tests.
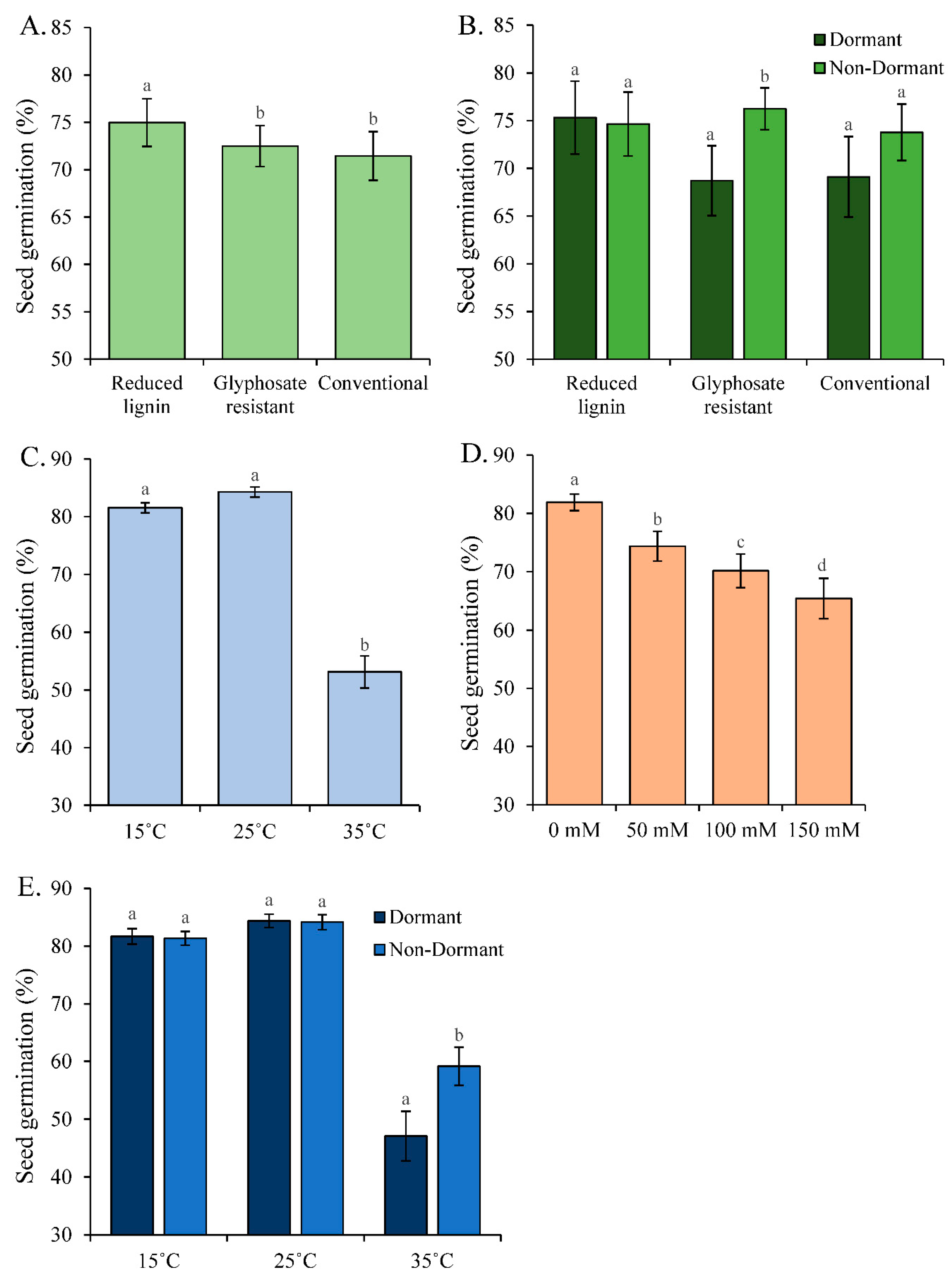
Figure 2.
Percent seed germination (mean +/ se) with increasing salinity at three temperatures.
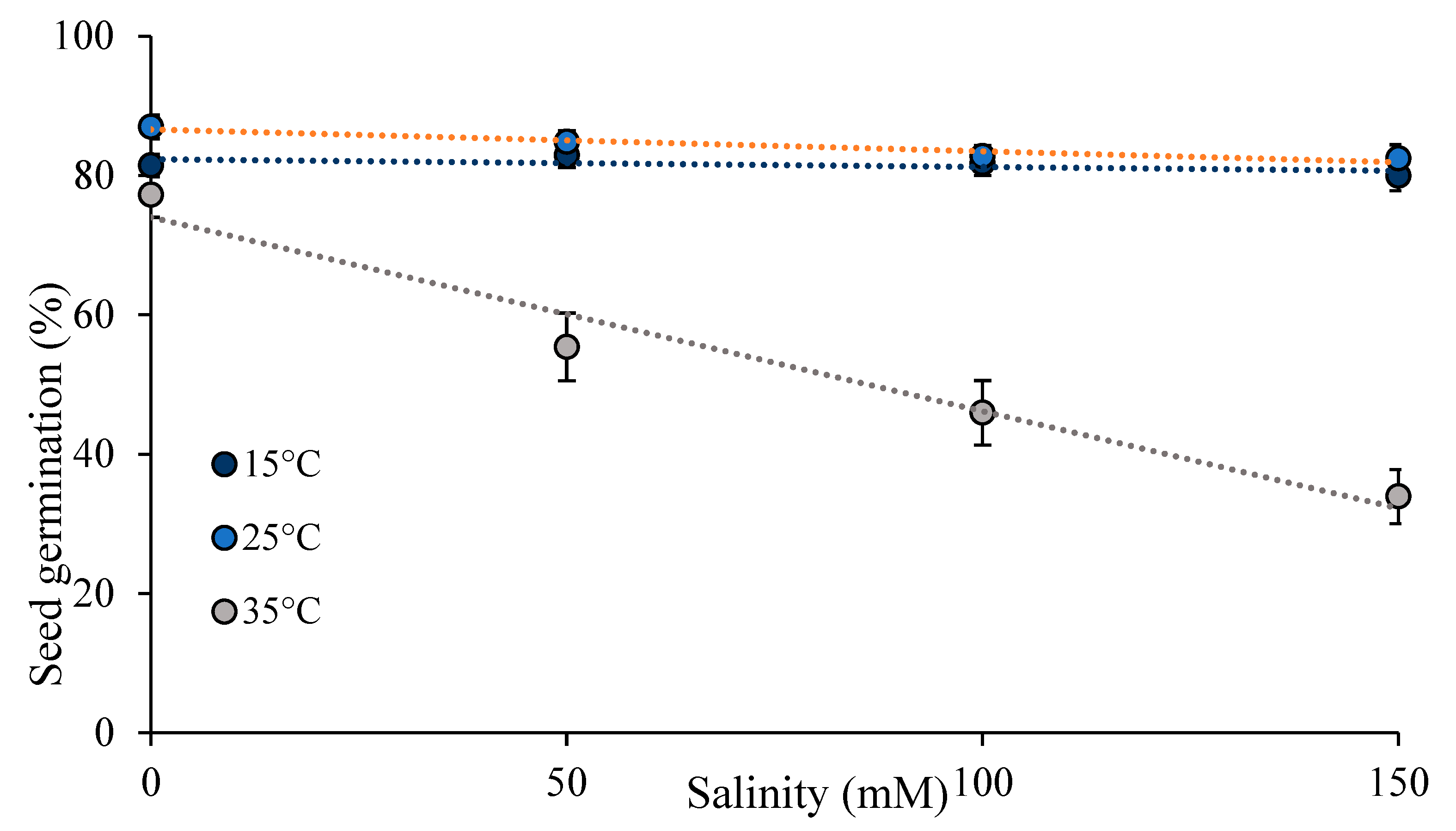
When we analyzed the results separately at each temperature, category affected seed germination at 15°C and 25°C, while dormancy type affected seed germination at 35°C. There was genetic variation in seed germination, and distinct responses of category and dormancy type at different temperatures. Only at 35°C did seed germination decrease as salinity levels increased, indicating variation in the phenotypic response to salinity with temperatures (Table S1). We did not observe any statistically significant interactions between category and dormancy type at any of the three temperatures.
2.2. Radicle Length
Temperature, dormancy, and salinity affected radicle length (Table 2). Non-dormant varieties had longer radicles (20.0 ± 0.91) than dormant varieties (18.8 ± 0.85). However, we detected a statistically significant interaction between the effects of category and dormancy on radicle length, suggesting genetic variation in radicle length between dormancy types for the different alfalfa categories (Table 2). While non-dormant varieties of conventional and glyphosate resistant alfalfa had longer radicles than dormant varieties, the reverse trend was detected for reduced lignin alfalfa (Figure 3A).
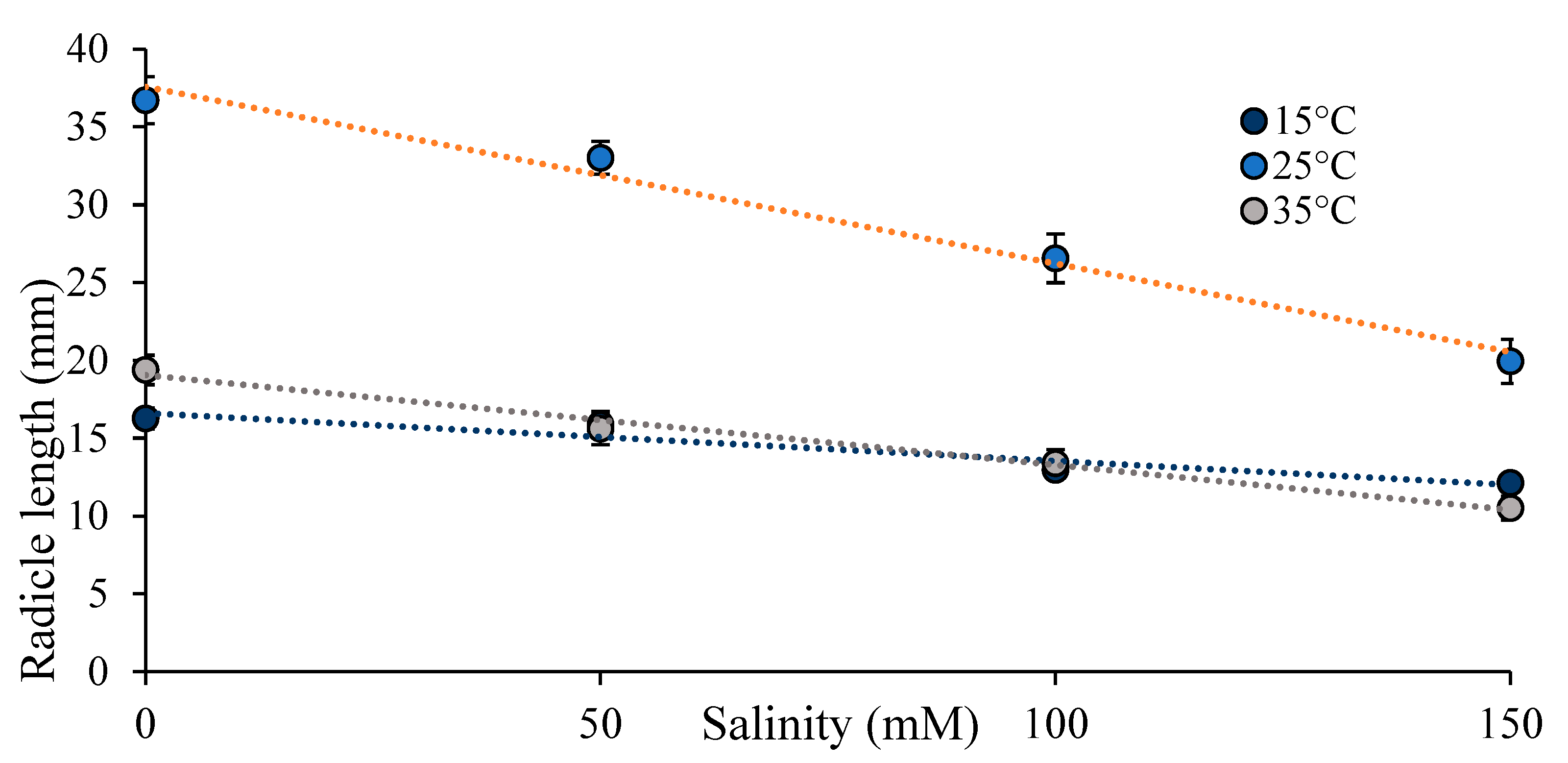
Radicle length was affected by temperature and by salinity (Table 2). There was a phenotypic response of radicle length to these two abiotic factors. The longest radicles after four days were detected at 25˚C (Figure 3B). Radicle length decreased with increasing salinity overall (Figure 3C). However, there was an interaction between the impact of temperature and salinity on radicle length, and the decline in radicle length with increasing salinity was steepest at 25˚C (Figure 4).
When each temperature was examined separately, statistically significant interactions were observed between the effects of category and dormancy on radicle length at all temperatures (Table S2). At all temperatures, radicle length decreased with increasing salinity, with variation in the phenotypic response among categories at 25˚C (Table S2).

Table 2.
The impact of temperature, category, dormancy and salinity and their interactions on radicle length based on linear models. Statistically significant factors are in bold.
Table 2.
The impact of temperature, category, dormancy and salinity and their interactions on radicle length based on linear models. Statistically significant factors are in bold.
| Effect | Num df | Den df | F value | Pr > F |
|---|---|---|---|---|
| Temperature | 2 | 144 | 299.58 | < 0.0001 |
| Category | 2 | 144 | 2.44 | 0.0911 |
| Dormancy | 1 | 144 | 5.69 | 0.0183 |
| Salinity | 3 | 144 | 74.59 | <.0001 |
| Temperature X Category | 4 | 144 | 0.62 | 0.6473 |
| Temperature X Dormancy | 2 | 144 | 0.78 | 0.4605 |
| Temperature X Salinity | 6 | 144 | 3.92 | 0.0012 |
| Category X Dormancy | 2 | 144 | 14.84 | <.0001 |
| Category X Salinity | 6 | 144 | 2.02 | 0.0673 |
| Dormancy X Salinity | 3 | 144 | 0.15 | 0.9281 |
| Temperature X Category X Salinity | 4 | 144 | 1.71 | 0.1512 |
| Temperature X Dormancy X Salinity | 6 | 144 | 0.72 | 0.6334 |
| Category X Dormancy X Salinity | 6 | 144 | 1.84 | 0.0957 |
| Temperature X Category X Dormancy X Salinity | 24 | 144 | 1.03 | 0.4352 |
3. Discussion
Anthropogenic climate changes are modifying temperature and precipitation patterns worldwide with consequences for crop production. We examine the potential for alfalfa to adjust to potential changes in temperature and salinity levels expected under climate change. We concentrate on early stages of the alfalfa life cycle, that is seed germination and seedling growth (radicle length after four days). Changes in abiotic factors often lead to a new optimum phenotype and we are examining the ways in which alfalfa might reach this new optimum. We quantified the phenotypic response of seed germination and radicle length to temperature and salinity, and determined whether genetic variation existed for these traits. If the phenotypic response brings the trait away from its optimum, then selection is necessary to bring the trait closer to the new optimum under the modified environmental conditions.
3.1. Seed Germination
Seeds from the same 18 varieties were exposed to all salinity levels and temperatures, which permitted the detection of the phenotypic response of seed germination to these two abiotic factors, also known as phenotypic plasticity. Seed germination was lowest at 35°C, with only 53% of seeds germinated, in contrast to over 80% at 15°C and 25°C, a 33.8% reduction in seed germination. Salinity overall reduced seed germination by 41.2%. However, the phenotypic response of seed germination to salinity levels varied with temperature; the decrease in seed germination with increasing salinity levels was only statistically significant at 35°C. At lower temperatures (15°C and 25°C), seed germination remained 80% or higher even at the stronger salinity level (150mM). At higher temperature (35°C), seed germination decreased from 77% in the absence of salinity to 34% at 150mM, a 55.8% reduction in seed germination for combined high temperature (35˚C) and high salinity (150mM) conditions. Comparing seed germination between 25°C and 0mM relative to 35°C and 150mM, we observed a 61% reduction in seed germination. The interaction between the effects of temperature and salinity on seed germination has been previously observed [14,21]. However, these earlier studies were performed on a limited number of alfalfa cultivars. In contrast, the current study included 18 varieties or breeding stocks, of both dormant and non-dormant types, for each of three alfalfa categories, conventional, glyphosate resistant, and reduced lignin alfalfa, enabling generalization of the results. The phenotypic response to high temperature (35°C) and high salinity (150mM) moved seed germination away from its high germination optimum, and selection is thus needed to enable better establishment of alfalfa under conditions of higher temperature and salinity expected under climate change in some regions.
Figure 3.
Radicle length (mm) (mean ± se) per A) Category and dormancy type, B) temperature, and C) Salinity. N is the sample size. Different letters indicate statistically. different treatments based on a Tukey means multiple comparison test.
Figure 3.
Radicle length (mm) (mean ± se) per A) Category and dormancy type, B) temperature, and C) Salinity. N is the sample size. Different letters indicate statistically. different treatments based on a Tukey means multiple comparison test.
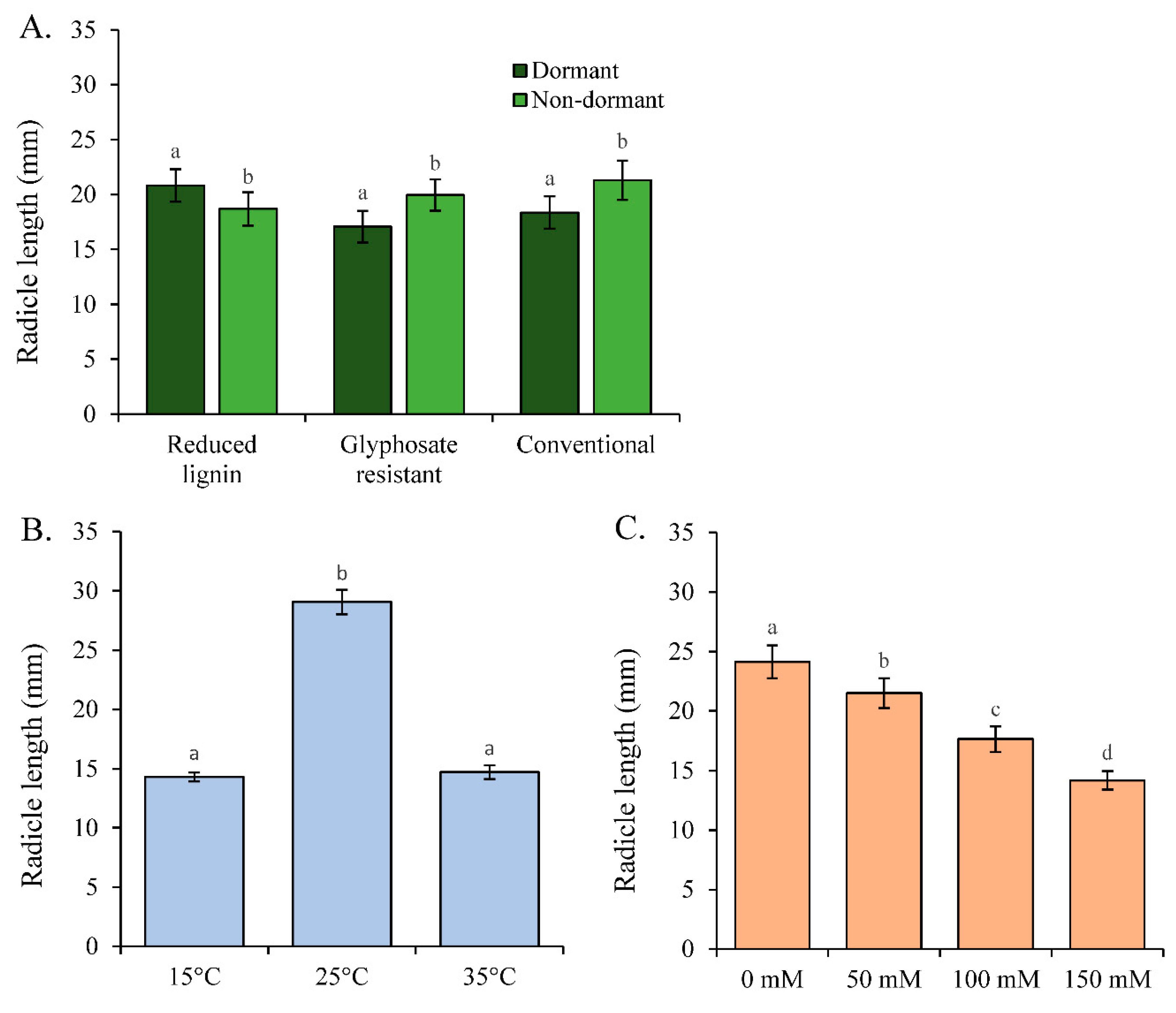
Figure 4.
Radicle length (mm) (mean ± se) with increasing salinity (mM) at three temperatures.
Standing genetic variation provides genetic material for selection to work on. Because all varieties were grown under similar environmental conditions, differences among dormancy types or alfalfa categories indicate the presence of genetic variation in the trait (genetic differentiation between dormancy type or category). We detected differences in seed germination between dormancy types, and an interaction with category. Non-dormant varieties had greater seed germination than dormant varieties overall, but the difference was statistically significant for glyphosate resistant varieties (p < 0.05), borderline (p = 0.06) for conventional varieties, and non-significant (p = 0.59) for reduced lignin breeding stocks. This genetic variation can be used to increase alfalfa seed germination under conditions of higher temperature and salinity. Further studies of reduced lignin alfalfa varieties are needed to determine whether the lack of difference between dormant and non-dormant varieties observed in this experiment is a consequence of using breeding stocks or if such difference is maintained in the reduced lignin varieties. Persistent differences would suggest different selection regimes to increase seed germination under higher temperature and salinity levels for the different alfalfa categories.
Genetic variation in the phenotypic response of seed germination to temperature, but not to salinity, was detected in this study. Seed germination was slightly greater for non-dormant (74.9% ± 1.6) relative to dormant varieties (71.1% ± 2.3) overall, but this difference was only statistically significant at 35°C (temperature X dormancy interaction). At 15°C and 25°C seed germination was similar for dormant and non-dormant varieties. This interaction between temperature and dormancy on their effects on seed germination indicates the presence of genetic variation in the phenotypic response of seed germination to temperature. Given the phenotypic response was a decrease in seed germination at high temperature for both non-dormant (59.2% ± 3.3) and dormant varieties (47.1% ± 4.3), the genetic variation in phenotypic plasticity could be used to lower phenotypic plasticity in response to temperature, while selecting for increased seed germination. With respect to salinity, we did not observe interactions between dormancy, or category, and salinity, indicating the absence of genetic variation in the phenotypic response of seed germination to salinity.
Because higher temperature and higher salinity often occur in concert, especially for irrigated alfalfa [6], a selection regime for increase seed germination under combined conditions of high temperature and high salinity would help alfalfa adapt to new environmental conditions associated with climate change. Selecting for higher seed germination and a low phenotypic response to temperature would seem most beneficial. For salinity, one cannot select for a lower phenotypic response, given the lack of genetic variation in phenotypic plasticity of seed germination in response to salinity. Besides selecting for higher seed germination, it is important to also select for high forage yield under conditions of high temperature and high salinity. Mature alfalfa plants have been shown to be more salt tolerant than previously thought, and can recover from periods of drought [15,36]. However, if plants cannot establish, the tolerance of mature plants to high temperature and high salinity becomes less relevant. Breeding for higher salt tolerance of alfalfa varieties has been an important goal for some time, and varieties with increased salt tolerance have been released in recent years (for example ‘Hi-Salt’ varieties by Alforex seeds). These varieties can reach greater forage yield on marginally saline soils [37]. However, salinity is only one of the abiotic factors expected to increase with climate change. Future studies should test these varieties under a range of temperatures and salinities, as was done in this study, to determine their phenotypic plasticity to these abiotic factors and further describe the standing genetic variation for seed germination and hay yield and for phenotypic plasticity itself. Testing varieties at temperatures higher than 35˚C but below 45˚C, a temperature that led to minimal germination, would also be of interest.
3.2. Radicle Length
Radicle length exhibited phenotypic plasticity to both temperature and salinity, with an interaction between the effects of temperature and salinity on radicle length. Radicles were much longer at 25˚C (29.1 ± 1.0 mm), compared to 15˚C (14.3 ± 0.37), or 35˚C (14.7 ±0.6), a 50% reduction in radicle length. Radicle length also responded to salinity, decreasing from 24.2 ± 1.4 mm at 0mM, to 14.2 ± 0.8 at 150mM, a 41.3% decrease. But the decrease with salinity varied with temperature. We observed a 45.8% decrease in radicle length from 0mM to 150mM at 25˚C, and a similar decrease at 35˚C (45.9%) but a lesser reduction at 15˚C (25.8%). Ideal growing conditions for alfalfa seedlings are at 25˚C with no salinity, and a 61% reduction in radicle length was observed at 35°C and 150mM. High temperature and high salinity moved radicle length away from its optimum.
We detected genetic variation in seedling growth (radicle length after four days), which is needed for selection to increase radicle length under conditions of high temperature and high salinity. Non-dormant varieties had longer radicles (20.0 ± 0.9 mm) relative to dormant varieties (18.8 ± 0.8 mm). However, the difference between non-dormant and dormant varieties varied with the alfalfa category. These was a statistically significant interaction between the effects of category and dormancy type on radicle length. While non-dormant varieties had longer radicles than dormant varieties in conventional and glyphosate resistant alfalfa, the opposite trend was observed in the reduced lignin breeding stocks, with longer radicle in dormant varieties. We did not detect any interaction between the impact of dormancy or category and either temperature or salinity, which reflects an absence of genetic variation in the phenotypic response of radicle length to these abiotic factors. Future studies should determine whether the different trend observed between dormancy types for reduced lignin alfalfa resulted from using seeds from breeding stocks as opposed to commercially available cultivars for the other two alfalfa categories. A persistent difference would affect the selection regimes for the different alfalfa categories.
3.3. Alfalfa under Climate Change
Alfalfa seeds germinate at the soil surface, and are therefore exposed to the higher temperature and higher salinity levels at the soil surface, relative to some other crops whose seeds germinate deeper in the ground. Ideal conditions for seed germination and seedling growth of alfalfa are at 25˚C with zero salinity. However, at 25˚C, increased salinity did not impact seed germination very much, although it slowed down seedling growth considerably. At temperature of 35˚C, seed germination significantly decreased (53.3% ± 2.8) and was even lower when combined with high salinity level (150mM) (34.0% ± 3.9). The seeds that germinated grew more slowly at 35˚C relative to 25˚C. Even if seed germination is not affected by lower temperatures (15˚C), seedlings grow more slower than at 25˚C. Selection for greater seed germination and seedling growth are needed for alfalfa to adapt to predicted high temperature and high salinity conditions that will be associated with climate change in some areas.
Genetic variation for seed germination and seedling growth in alfalfa is currently associated with dormancy type and category. To conserve plant genetic resources, alfalfa collections of wild relatives, landraces, and cultivars are held in various gene banks worldwide. The USDA-AARS National Plant Germplasm system, holds a gene bank of 13,000 unique individual alfalfa samples [38]. Such germplasm should be tested for the phenotypic response to temperature and salinity, and for genetic variation of traits of interest (seed germination, seedling growth, forage and seed yield), and we suggest using a design similar to the one introduced in this study. Genetic material with higher seed germination, seedling growth, and yield under high temperature, low water, and high salinity conditions, and with a low phenotypic response to these abiotic factors, would represent good genetic material to introduce into alfalfa cultivars to ensure alfalfa adapts to the new environments expected under climate change.
4. Materials and Methods
4.1. Plant Material
Medicago sativa, commonly known as alfalfa or lucerne, is a perennial plant that depends on bees for pollination and seed production [39]. Cultivated alfalfa is a tetraploid, self-compatible plant, with an average selfing rate of 12.2% in seed-production fields [40]. Three categories of alfalfa, conventional, glyphosate resistant (also known as Roundup Ready), and reduced lignin, were used in the experiment. Within each category, we tested three dormant and three non-dormant varieties (Table 3). Dormant alfalfa varieties have reduced shoot elongation in the fall, and are better adapted to colder conditions [5,29]. Non-dormant varieties continue to grow and reach their largest height in the fall, are generally not winter hardy, and are better adapted to warmer climates. Because multiple varieties of reduced lignin alfalfa (known as HarvXtra) were not available when this experiment took place, we used different breeding stocks. HarvXtra varieties combine the reduced lignin and glyphosate resistance traits. The name of the 18 alfalfa varieties or breeding stocks used in the experiment are summarized in Table 3, together with their category, dormancy type, and dormancy level. Seeds were provided by Forage Genetics International™.
4.2. Experimental Design
The effects of category (conventional, glyphosate resistant, and reduced lignin), dormancy type (dormant vs. non-dormant), and salinity levels (0, 50, 100, and 150 mM), on seed germination and radicle length were examined at three temperatures (15°C, 25°C, or 35°C) (Table 4). A fourth temperature (45°C) was tested in a preliminary study, but seed germination was extremely low (< 5.0%) and this treatment was not pursued. One incubator was set at 15°C, a second one at 25°C, and a third at 35°C, and within each incubator, seeds from the three categories, each with three dormant and three non-dormant alfalfa varieties, were tested under each of the four salinity levels (Table 4). The 0 mM solution consisted of distilled water and solutions of 50, 100, or 150 mM were prepared by diluting sodium chloride (NaCl) in distilled water. These salinity values are equivalent to 0, 2922, 5844 and 8766 ppm, respectively, and to values of 0, 4.56, 9.13 and 13.70 dS/m (milliSiemens per meter), assuming 640 ppm per 1 dS/m. As a comparison, seawater is approximately 599 mM with 35,000 ppm. One ppm is equivalent to 0.0001% NaCl. There were 216 cells in the experiment (3 temperatures X 3 categories X 2 dormancy levels X 4 salinity levels X 3 varieties) (Table 4). For each cell, one hundred seeds were placed in a petri dish between two layers of circular Fisherbrand™ Filter Paper Quantitative Q5. There were included 72 petri dishes and 7,200 seeds per incubator, for a total of 216 petri dishes and 21,600 seeds in the experiment.
Table 3.
The varieties or breeding stocks used in the experiment, their dormancy type and level, and category. A number was assigned to each of the varieties used in the experiment. Breeding stocks were used for reduced lignin alfalfa due to the unavailability of different HarvXtra varieties when the experiment was conducted.
Table 3.
The varieties or breeding stocks used in the experiment, their dormancy type and level, and category. A number was assigned to each of the varieties used in the experiment. Breeding stocks were used for reduced lignin alfalfa due to the unavailability of different HarvXtra varieties when the experiment was conducted.
| Category | Dormancy Type | Variety Name | Dormancy Level | Number |
|---|---|---|---|---|
| Conventional | Dormant | AmeriStand 427TQ | 4.3 | 1 |
| FSG 424 | 4 | 2 | ||
| Lightning Bolt | 4 | 3 | ||
| Non- Dormant | Fertillac 10 | 10 | 4 | |
| Salado | 9.5 | 5 | ||
| Sun Quest | 9 | 6 | ||
| Glyphosate | Dormant | 428RR | 4 | 7 |
| Resistant | Heritage RR | 4 | 8 | |
| WL 336HQ.RR | 3.2 | 9 | ||
| Non-Dormant | 6015R | 10 | 10 | |
| RRALF 9R100 | 9 | 11 | ||
| AmeriStand 835NTS RR | 8 | 12 | ||
| Reduced lignin | Dormant | HX FD-1 | NA | 13 |
| HX FD-2 | NA | 14 | ||
| HX FD-3 | NA | 15 | ||
| Non-Dormant | HX ND-1 | NA | 16 | |
| HX ND-2 | NA | 17 | ||
| HX ND-3 | NA | 18 |
4.3. Seed Germination and Radicle length Measurements
Petri dishes were checked daily and five mL of distilled water (0mM) or the respective NaCl solution was added when the double layered filter paper was getting dry. The number of germinated seeds per petri dish was recorded on days 1-4 following the day the experiment was set up, at the same time each day. Preliminary results indicated that little change in seed germination occurred after the fourth day. A seed was classified as germinated when the radicle length was greater than one mm. On each day, the number of seeds that had germinated in each petri dish were counted and recorded together with the total number of seeds per petri dish. The order of examination of petri dishes each day was randomly selected between and within incubators.
Radicle length (mm) was measured on the fourth day of the experiment for 10 germinated seeds per petri dish. The ten seeds were selected to represent the distribution of radicle lengths in a petri dish. Radicle length was measured using the program ImageJ [41]. A pilot study indicated no differences in radicle length when measured on the fourth day of the experiment relative to the fourth day following the day of germination for each specific seed. Therefore, to simplify the logistics of the experiment, we measured radicle length on the fourth day of the experiment. When fewer than ten seeds had germinated in a petri dish, radicle length was measured on all germinated seeds. The average radicle length was calculated for each petri dish.
Table 4.
Experimental design. There were three incubators, one set at 15°C, a second at 25°C, and a third at 35°C. Within each incubator, alfalfa seeds from three categories (conventional, glyphosate resistant, and reduced lignin), two dormancy types per category, and three varieties per dormancy type, were germinated and grown under four salinity levels (0, 50, 100, and 150 mM).
Table 4.
Experimental design. There were three incubators, one set at 15°C, a second at 25°C, and a third at 35°C. Within each incubator, alfalfa seeds from three categories (conventional, glyphosate resistant, and reduced lignin), two dormancy types per category, and three varieties per dormancy type, were germinated and grown under four salinity levels (0, 50, 100, and 150 mM).
| Temperature 15˚C | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Conventional | Glyphosate resistant | Reduced lignin | |||||||||||||||||||||
| Dormant | Non-Dormant | Dormant | Non-Dormant | Dormant | Non-Dormant | ||||||||||||||||||
| Salinity level (mM) | |||||||||||||||||||||||
| 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 |
| 1 | 1 | 1 | 1 | 4 | 4 | 4 | 4 | 7 | 7 | 7 | 7 | 10 | 10 | 10 | 10 | 13 | 13 | 13 | 13 | 16 | 16 | 16 | 16 |
| 2 | 2 | 2 | 2 | 5 | 5 | 5 | 5 | 8 | 8 | 8 | 8 | 11 | 11 | 11 | 11 | 14 | 14 | 14 | 14 | 17 | 17 | 17 | 17 |
| 3 | 3 | 3 | 3 | 6 | 6 | 6 | 6 | 9 | 9 | 9 | 9 | 12 | 12 | 12 | 12 | 15 | 15 | 15 | 15 | 18 | 18 | 18 | 18 |
| Temperature 25˚C | |||||||||||||||||||||||
| Conventional | Glyphosate resistant | Reduced lignin | |||||||||||||||||||||
| Dormant | Non-Dormant | Dormant | Non-Dormant | Dormant | Non-Dormant | ||||||||||||||||||
| Salinity level | |||||||||||||||||||||||
| 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 |
| 1 | 1 | 1 | 1 | 4 | 4 | 4 | 4 | 7 | 7 | 7 | 7 | 10 | 10 | 10 | 10 | 13 | 13 | 13 | 13 | 16 | 16 | 16 | 16 |
| 2 | 2 | 2 | 2 | 5 | 5 | 5 | 5 | 8 | 8 | 8 | 8 | 11 | 11 | 11 | 11 | 14 | 14 | 14 | 14 | 17 | 17 | 17 | 17 |
| 3 | 3 | 3 | 3 | 6 | 6 | 6 | 6 | 9 | 9 | 9 | 9 | 12 | 12 | 12 | 12 | 15 | 15 | 15 | 15 | 18 | 18 | 18 | 18 |
| Temperature 35˚C | |||||||||||||||||||||||
| Conventional | Glyphosate resistant | Reduced lignin | |||||||||||||||||||||
| Dormant | Non-Dormant | Dormant | Non-Dormant | Dormant | Non-Dormant | ||||||||||||||||||
| Salinity level | |||||||||||||||||||||||
| 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 | 0 | 50 | 100 | 150 |
| 1 | 1 | 1 | 1 | 4 | 4 | 4 | 4 | 7 | 7 | 7 | 7 | 10 | 10 | 10 | 10 | 13 | 13 | 13 | 13 | 16 | 16 | 16 | 16 |
| 2 | 2 | 2 | 2 | 5 | 5 | 5 | 5 | 8 | 8 | 8 | 8 | 11 | 11 | 11 | 11 | 14 | 14 | 14 | 14 | 17 | 17 | 17 | 17 |
| 3 | 3 | 3 | 3 | 6 | 6 | 6 | 6 | 9 | 9 | 9 | 9 | 12 | 12 | 12 | 12 | 15 | 15 | 15 | 15 | 18 | 18 | 18 | 18 |
Variety or breeding stock is the replicate in the experiment, with three replicates per salinity level within a category and dormancy type. The variety name associated with each variety number can be found in Table 3.
4.4. Statistical Analyses
We used a linear model to examine the impact of temperature, salinity, category, and dormancy type, together with their interactions, on seed germination or radicle length. Because the same varieties were grown under each treatment, a statistically significant effect of temperature or salinity in this study reflects a phenotypic response of seed germination or radicle length to the abiotic factor (phenotypic plasticity; Table 5). Because the varieties were grown under similar conditions, a category of dormancy type effect suggests genetic differentiation in seed germination or radicle length among category or dormancy type, respectively. Interaction between temperature and dormancy indicates a different phenotypic response to temperature between dormancy types. Table 5 summarizes the interpretation for the major factors and their interactions in the experiment.
Table 5.
Interpretation of factor effects in the full model. The factors include temperature, salinity, category, dormancy, and their interactions. .
Table 5.
Interpretation of factor effects in the full model. The factors include temperature, salinity, category, dormancy, and their interactions. .
| Factor | Interpretation |
|---|---|
| Temperature | Phenotypic plasticity to temperature |
| Salinity | Phenotypic plasticity to salinity |
| Category | Genetic differentiation in trait among categories |
| Dormancy | Genetic differentiation in trait among dormancy types |
| Temperature X Category | Genetic differentiation (category) in plastic response to temperature |
| Temperature X Dormancy | Genetic differentiation (dormancy) in plastic response to temperature |
| Temperature X Salinity | Phenotypic response to salinity varies with temperature |
| Category X Dormancy | Genetic differentiation in trait among dormancy types varies between categories |
| Salinity X Category | Genetic differentiation (category) in plastic response to salinity |
| Salinity X Dormancy | Genetic differentiation (dormancy) in plastic response to salinity |
| Temperature X Salinity X Category | Genetic differentiation (category) in plastic response to temperature X salinity |
| Temperature X Salinity X Dormancy | Genetic differentiation (dormancy) in plastic response to temperature X salinity |
| Salinity X Category X Dormancy | Genetic differentiation (category X dormancy) in plastic response to salinity |
| Temperature X Salinity X Category X Dormancy | Genetic differentiation (Category X dormancy) in plastic response to temperature X salinity |
Because we only had one incubator per temperature, we also analyzed the data separately at each temperature to consider potential issues with pseudo-replication. Variety (or breeding stock) was the replicate in the experiment. For statistically significant factors with more than two levels, multiple mean comparison tests were used to determine statistically significant differences among levels. Analyses were done in SAS v. 9.4 proc mixed. Results are presented for arcsin square root transformed seed germination, and for loge transformed radicle length because these transformations improved the models’ residuals.
5. Conclusions
Seed germination and seedling growth represent vulnerable stages in a plant life cycle. Both traits responded phenotypically, with drastic decreases in both seed germination and radicle length as temperature and salinity increased. The phenotypic response moved the traits away from their optimum seed germination or radicle length. Selection is therefore necessary to increase seed germination and radicle length under these harsher abiotic conditions. Some genetic variation exists for both traits among dormancy types and categories. In addition, there was genetic variation in the phenotypic response of seed germination to temperature, indicating that selection for lower levels of phenotypic plasticity is possible for that trait. Additional germplasm should be tested to further increase the genetic material available for selection of greater germination and better seedling growth in alfalfa under high temperature and high salinity conditions. Alfalfa establishment is currently quite poor under the high temperature and salinity levels tested here and needs to be improved via selection to permit alfalfa cultivars to adapt to new environmental conditions expected under climate change in many regions of the world.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: The impact of category, dormancy type, and salinity and their interactions on seed germination, at each of three temperatures (15˚C, 25˚C and 35˚C), based on linear models; Table S2: The impact of category, dormancy type, and salinity and their interactions on radicle length, at each of three temperatures (15˚C, 25˚C and 35˚C), based on linear models.
Author Contributions
Conceptualization, methodology and visualization J.B. and A.H.; validation, investigation, data curation, writing-review and editing, A.H.; formal analysis; resources, writing, supervision, project administration, and funding acquisition, J.B. All authors have read and agreed to the published version of the manuscript.
Funding
Funds for this research were provided by the Agricultural Research Service to J.B. The APC was funded by MDPI.
Data Availability Statement
The data presented in this study will be made openly available in [Dryad] at [doi], reference number [reference number] following acceptance of the manuscript.
Acknowledgments
We thank Emma Nelson for her help with data collection. Molly Dieterich Mabin prepared the figures. Austin Bauer and Marisa Brudny ran some preliminary trials.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Brunet, J.; Van Etten, M.L. The response of floral traits associated with pollinator attraction to environmental changes expected under anthropogenic climate change in high-altitude habitats. Int. J. Plant Sciences, 2019, 180, 929-933. https://www.journals.uchicago.edu/doi/pdfplus/10.1086/705591.
- Mooney, H.; Larigauderie, A.; Cesario, M.; Elmquis, T.; Hoegh-Guldberg, O.; Lavorel, S.; Mace, G.M.; Palmer, M.; Robert Scholes, R.; Tetsukazu Yahara, T. Biodiversity, climate change, and ecosystem services. COSUST .2009, 1, 46–54. [CrossRef]
- Trivedi, M.R.; Morecroftc, M.D.; Berrya, P.M.; Dawsond, T.P. Potential effects of climate change on plant communities in three montane nature reserves in Scotland, UK. Biol. Conserv. 2008, 141, 1665–1675.
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, R.; Jones, R. et al. Regional climate projections. In Climate change 2007: the physical science basis—contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L. eds. Cambridge University Press, New York. 2007. pp 847–940.
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [CrossRef]
- Ullah, A.; Bano, A.; Khan, N. Climate Change and Salinity Effects on Crops and Chemical Communication Between Plants and Plant Growth-Promoting Microorganisms Under Stress Front. Sustain. Food Syst., 2021, 5, 2021. | . [CrossRef]
- Li, J.; Ma, M.; Sun, Y.; Lu, P.; Shi, H.; Guo, Z.; Xhu, H. Comparative physiological and transcriptome profiles uncover salt tolerance mechanisms in alfalfa. Frontiers in Plant Science 2022, 13, 931619. [CrossRef]
- Porter, J.R. Rising temperatures are likely to reduce crop yields. Nature 2005, 436, 174. [CrossRef]
- Jagadish, S.V.K.; Way, D.A.; Sharkey, T.D. Scaling plant responses to high temperature from cell to ecosystem. Plant Cell Environ. 2021, 44, 1987–1991.
- Rising, J.; Devineni, N. Crop switching reduces agricultural losses from climate change in the United States by half under RCP 8.5. Nature Communications, 2020. 11, 4991. [CrossRef]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Palme, K.J.; Jansen, M.A.K. Stress-induced morphogenic responses: growing out of trouble? Trends Plant Sci. 2007, 12, 98–105.
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat Tolerance in Plants: An Overview. Environ. Exp. Bot. 2007, 61, 3.
- Ungar, I.A. Influence of Salinity and Temperature on Seed Germination. Ohio J. Sci. 1967, 67, 120–123.
- Esechie, H.A. Interaction of Salinity and Temperature on the Germination of Alfalfa CV CUF-101. Agronomie 1993, 13, 301–306.
- Putnam, D.; Meccage, E. Profitable alfalfa production sustains the environment. In Proceedings World Alfalfa Congress, San Diego, CA. 14-17 November 2022.
- NASS U.S. Department of Agriculture’s National Agricultural Statistics Service (NASS) 2017. https://www.nass.usda.gov/Publications/AgCensus/2017/index.php.
- Zhanwu, G.; Hui, Z.; Jicai, G.; Chunwu, Y.; Chunsheng, M.; Deli, W. Germination responses of Alfalfa (Medicago sativa L.) seeds to various salt-alkaline mixed stress. J. Agric. Res. 2001, 6, 3793–3803.
- Johnson, D.; Smith, S.; Dobrenz, A. Genetic and phenotypic relationships in response to NaC1 at different developmental stages in alfalfa. Theor Appl Genet. 1992, 83, 833–838.
- Scasta, J.D.; Trostle, C.L.; Foster, M.A. Evaluating Alfalfa (Medicago sativa L.) Cultivars for Salt Tolerance.
- Using Laboratory, Greenhouse and Field Methods. J. Agric. Sci. 2012, 4. [CrossRef]
- Waissman Assadian, N., Miyamoto, S. Salt Effects on alfalfa seedling emergence. Agron. J. 1987, 79, 710–714.
- Sandhu, D.; Cornacchione, M.V.; Ferreira J.F.S.; Suarez D.L. Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci. Rep. 2017, 7, 42958. [CrossRef]
- Stone, J.E.; Marx, D.B.; Dobrenz, A.K. Interaction of sodium chloride and temperature on germination of two alfalfa cultivars. Agron. J. 1979, 71, 425–427.
- Flajoulot, S.; Ronfort, J.; Baudouin, P.; Barre, P.; Huguet, T.; Huyghe C.; Julier, B. Genetic diversity among alfalfa (Medicago sativa) cultivars coming from a breeding program, using SSR markers. Theor Appl Genet 2005, 111, 1420–1429.
- Qiang, H.; Chen, Z.; Zhang, Z.; Wang, X.; Gao, H.; Wang, Z. Molecular diversity and population structure of a worldwide collection of cultivated tetraploid alfalfa (Medicago sativa subsp. sativa L.) germplasm as revealed by microsatellite markers. PLoS ONE 2015, 10, e0124592. [CrossRef]
- Annicchiarico, P.; Nazicari, N., Ananta, A.; Carelli, M.; Wei, Y.; Brummer, E.C. Assessment of cultivar distinctness in alfalfa: a comparison of genotyping-by-sequencing, simple-sequence repeat marker, and morphophysiological observations. Plant Genome, 2016, 9. [CrossRef]
- Yu, L.X.; Liu, X.; Boge, W.; Liu, X.P. Genome-wide association study identifies loci for salt tolerance during germination in autotetraploid alfalfa (Medicago sativa L.) using genotyping-by-sequencing. Front. Plant Sci. 2016, 7, 956. [CrossRef]
- Rogan, G.; Fitzpatrick, S. Petition for determination of nonregulated status: Roundup Ready alfalfa (Medicago sativa L.) events J101 and J163. USDA Petition 2004, (04-110), 101.
- Barros, J.; Temple, S.; Dixon, R.A. Development and commercialization of reduced lignin alfalfa. Curr. Opin.Biotechnol. 2019, 56, 48–54.
- Liu, Z.; Li, X.; Wang, Z.; Sun, Q. 2015. Contrasting Strategies of Alfalfa Stem Elongation in Response to Fall Dormancy in Early Growth Stage: The Tradeoff between Internode Length and Internode Number. PLOS ONE 2015, 10, e0135934. [CrossRef]
- Ghalambor, C.K.; McKay, J.K.; Carroll, S.P.; Reznick, D.N. Adaptive versus non-adaptive phenotypic plasticity and the pote tial for contemporary adaptation in new environments. Functional Ecology 2007, 21, 394‒407.
- Lande, R. Adaptation to an extraordinary environment by evolution of phenotypic plasticity and genetic assimilation. J. Evol. Biol. 2009, 22, 1435‒1446.
- Chevin, L.M.; Lande, R.; Mace, G.M. Adaptation, plasticity, and extinction in a changing environment: towards a predictive theory. PLOS Biology 2010, 8, 1‒8.
- Fox, R.J.; Donelson, J.M.; Schunter,C.R; Timothy, R.; Gaitan-Espitia, J.D. Beyond buying time: the role of plasticity in phenotypic adaptation to rapid environmental changes. Philos. Trans. R. Soc. B. 2019, 374, 20180174. [CrossRef]
- Laitinen, R.A.E.; Nikoloski, Z. Genetic basis of plasticity in plants. J. Exp. Bot. 2019, 70, 739–745.
- Chevin, L.M; Lande, R. When do adaptive plasticity and genetic evolution prevent extinction of a density-regulated popul tion? Evolution 2009, 64, 1143‒1150.
- Al-Farsi, S.M.; Nadaf, S.K.; Al-Sadi, A.M.; Ullah, A.; Farooq, M. Evaluation of indigenous Omani alfalfa landraces for morphology and forage yield under different levels of salt stress. Physiol Mol Biol Plants, 2020, 26, 1763–1772.
- Scott, K. High salinity soils take special considerations to raise alfalfa. High Plains Journal, 2020. https://www.hpj.com/crops/high-salinity-soils-take-special-considerations-to-raise-alfalfa/article_07d6fec8-a73a-11ea-baf9-9b539fd9730f.html, viewed 03-16-2023.
- Irish, B.M.; Greene, S.L. Germplasm collection, genetic resources, and gene pools in alfalfa. In: The Alfalfa Genome. Compendium of Plant Genomes. Yu LX., Kole C., Eds.; Springer. Cham, Switzerland. 2021. pp. 43–64.
- Bohart, G.E. Pollination of alfalfa and red clover. Annu Rev Entomol. 1957, 2, 355–389.
- Dieterich Mabin, M.E.; Brunet, J.; Riday, H.; Lehmann, L. Self-fertilization, inbreeding and yield in alfalfa seed production fields. Front.Plant Sci. 2021, 12, 700708. [CrossRef]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. 2014. Image Processing with ImageJ. Biophotonics Int., 2014, 11, 36–42.
Table 1.
The impact of temperature, category, dormancy and salinity and their interactions on seed germination based on linear models. Statistically significant factors are in bold.
Table 1.
The impact of temperature, category, dormancy and salinity and their interactions on seed germination based on linear models. Statistically significant factors are in bold.
| Effect | Num df | Den df | F value | Pr > F |
|---|---|---|---|---|
| Temperature | 2 | 144 | 190.27 | <0.0001 |
| Category | 2 | 144 | 4.26 | 0.0159 |
| Dormancy | 1 | 144 | 5.98 | 0.0157 |
| Salinity | 3 | 144 | 23.44 | <0.0001 |
| Temperature X Category | 4 | 144 | 1.23 | 0.299 |
| Temperature X Dormancy | 2 | 144 | 8.01 | 0.0005 |
| Temperature X Salinity | 6 | 144 | 14.54 | <0.0001 |
| Category X Dormancy | 2 | 144 | 3.14 | 0.0462 |
| Category X Salinity | 6 | 144 | 1.25 | 0.287 |
| Dormancy X Salinity | 3 | 144 | 0.22 | 0.882 |
| Temperature X Category X Salinity | 4 | 144 | 1.02 | 0.399 |
| Temperature X Dormancy X Salinity | 6 | 144 | 1.10 | 0.366 |
| Category X Dormancy X Salinity | 6 | 144 | 0.58 | 0.749 |
| Temperature X Category X Dormancy X Salinity | 24 | 144 | 0.55 | 0.957 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



