
Submitted:
25 April 2023
Posted:
26 April 2023
You are already at the latest version
Abstract
Soil nitrogen (N) is a common limiting factor where soil N cycling is a key component of agro-ecosystems. Soil N transformation processes are largely mediated by microbes and understanding bacteria involvement in soil N cycling in agricultural systems has both agronomic and environmental importance. This 2-yr field scale study examined the abundances and spatial distributions of total bacterial community (16s RNA), bacterial involved in nitrification (amoA), and de-nitrification (narG, nirK, and nosZ) and soil physicochemical properties of winter wheat (Triticum aestivum L.) – soybean (Glycene max L.) double cropping with 2-3 weeks of spring grazing (WGS) and without grazing (WS) and of tall fescue (Festuca arundinacea (L.) Schreb.) pasture (TF) managed to near natural conditions with similar grazing. The TF had a significantly higher abundance of 16S rRNA, amoA, narG, nirK, and nosZ genes than the WS and WGS, which had similar levels between themselves. Soil organic matter (OM) and soil pH had a stronger effect on the N-cycling bacteria gene abundance. All bacterial gene concentrations and soil pH showed non-random distribution patterns with 141-186 m range autocorrelation. These results indicate that biological N transformation processes are more important in natural agricultural systems and the abundance of N-cycling bacteria can be manipulated by field scale management strategies.
Keywords:
N cycling bacteria genes
; wheat-soybean cropping
; tall fescue pasturing
; spatial distribution
1. Introduction
Meeting the rising demand for food through increased productivity with concomitant reduction of environmental impacts is always challenging [1]. Sustainable intensification of conventional agriculture production systems has been identified as a measure of achieving higher production goals while assuring higher environmental quality standards [2]. Greater crop yields and animal production attained in recent decades were mainly derived from increasingly specialized and decoupled crop and livestock systems. However, this dissociation between crop and animal components poses significant threats to the sustainability of the food systems [3]. Integration of winter cereal crops for grain production or for dual use (grazing and grain production) into mono cropping systems can be a sustainable intensification strategy and an alternative to conventional decoupled crop and animal production [3]. Winter cereal crops with forage production capability can contribute additional positive agronomic and ecological benefits by providing forage sources for livestock and food for human consumption [4].
Winter wheat (Triticum aestivum L.) crop has been shown to be compatible with soybean (Glycene max L.) and winter wheat-soybean double cropping is a common practice in the southern corn-belt states in the United States [5] . Winter wheat as a dual-use crop can provide both grain yield and high-quality forage for livestock grazing in the spring [6,7] with no reduction in wheat grain yield compared to the winter wheat grown exclusively for grain production [8] . Winter wheat grown with soybean as a double crop did not affect the grain yield of subsequently planted soybean crop as well [9]. Winter wheat dual use crop offers unique economic benefits of both grain production and value that is added, as weight gain of cattle grazing on wheat crop. Tall fescue (Festuca arundinacea (L.) Schreb.) is an important cool-season perennial forage grass that is widely used for turf, forage production, and soil conservation [10]. Tall fescue pasture covers a significant area in the United States [11] and has the potential for growing on poor fertile soils with light management by occasional mowing [12].
Nitrogen (N) is an essential element for plant growth and is the one most demanded by plants [13]. The availability of N in soils is of primary importance in agro ecosystem productivity [14] and environmental quality [15]. Shifts in soil N status can be caused by variation in N transformations [16] which are largely mediated by the microbiota [17]. The diversity, richness, and composition of microbial communities associated with N-cycling affect nitrogen availability to crop and N loss from the agricultural ecosystem [18]. The major soil N transformations include mineralization of organic N, nitrification, denitrification, NH4 volatilization, and N2 fixation [19]. Mineralization of organic N in soils serves as the major source of N to plants for uptake and synthesis of biomass.
Nitrifying bacteria largely determine N availability in soils in terms of both inorganic N quantity and NO3-/ NH4+ balance. Nitrification converts N derived from mineralization of organic matter to nitrate which is suitable for plant uptake or further microbial cycling [20] thus is a fundamental component of soil N cycling and fertility. The initial step in the nitrification pathway, the oxidation of NH4+ to NO3- via NH2OH is completed by ammonia oxidizers, comprising ammonia oxidizing bacteria. Denitrifying bacteria are responsible for reducing soluble oxidized nitrogen compounds into gaseous N2O or N2 for energy conservation through a series of transformations. The denitrification process has received a greater attention because it accounts for significant losses of fertilizer nitrogen from agriculture soils. In addition, denitrification is also responsible for the emission of N2O, an important greenhouse gas with a global warming potential c.a. 250 times higher than carbon dioxide [21]. These effects serve to focus attention on the organisms involved in the biogeochemical transformation of nitrogen in the soil.
The key enzyme for aerobic ammonia oxidizers is ammonia monooxygenase. The gene coding for a subunit of this enzyme, amoA, can reflect the phylogeny of the ammonia oxidizers [22]. As this step is rate limiting, detection of amoA is widely used as a measure of the biological capacity for the entire nitrification process[23].The denitrification pathway consists of the sequential reduction of NO3- to N2 via the metalloenzymes nitrate reductase (NO3- to NO2-), nitrite reductase (NO2- to NO), nitric oxide reductase (NO to N2O), and nitrous oxide reductase ( N2O to N2) [24] encoded by narG, nirK /nirS, norB, and nosZ genes, respectively[25,26]. The concentration of total bacterial community can be quantified using 16S rRNA as a molecular marker[27].
The abundances of N cycling genes are related to process rate and substrate availability and microbial population concentration in some environments [17,28,29]. Different agricultural practices have shown impacting many microbial driven natural biogeochemical processes in soil including nitrogen cycling [30] and plant community [9], such that alterations in microbial community composition or in the abundance or activity of specific groups can alter nitrogen availability and nitrogen losses from the soil. In agricultural systems, the ecology of N geochemistry at a molecular level may be closely linked to soil factors. Previous studies have shown that shifts in structure of bacterial communities can be associated with soil properties including texture [31], soil pH [32]and soil N availability [33]. The ammonia oxidizing bacteria populations have greater abundance in agricultural soils (soils with N fertilizer inputs, higher soil disturbance) than in less disturbed systems [34]. Increase of soil pH by liming increased the abundance of amoA genes in pasture soils by 26% but did not affect narG copy numbers [17]. Environmental factors affecting denitrification, such as O2, pH, C availability, NO3- pools, etc., all act through the soil biological community[35]. Grazing can affect soil physical properties (e.g., compaction driven decreases in macro-porosity and pore space) which in turn influences habitat for soil microbiota and conditions affecting microbial processes (e.g., O2 limitation, REDOX, water filled pore spaces etc.). In addition, land management practices linked with agricultural production systems have shown affecting soil chemical, physical and biological parameters [36,37,38].
Evidence from several studies has shown that microbial communities can exhibit non-random spatial distribution patterns from centimeter to meter scale in terrestrial ecosystems [39,40,41]. Understanding distribution patterns of microbial communities and environmental determinants at field scale is important for assessing relative importance of local factors and land management practices on microbial communities and soil nutrient cycling processes that they are responsible for. Thus, characterizing spatial distribution patterns of N cycling bacteria in agricultural systems enables to better understand the ecology of N cycling bacteria communities at a scale compatible with land management strategies focusing on transformation related production improvement and mitigating negative environmental impacts of N transformations.
There exists a great opportunity for expanding productivity gains while achieving environmental goals of agricultural ecosystems by manipulating biological cycling of soil nutrients such as C, N, and P [15,42]. Thus, assessments conducted at microbial community level focusing on the microbial ecology of N cycling bacteria would facilitate realizing higher productivity gains while mitigating negative environmental effects through field scale management strategies. The overall objective of this experiment was to examine the N cycling bacteria gene concentrations in wheat-soybean cropping and in near natural tall fescue pasture systems. The specific objectives were 1. to investigate the effects of winter wheat grain-soybean cropping with (WGS) and without grazing (WS) and of tall fescue pasturing system (TF) managed to near natural conditions on the abundance of total soil bacteria (16S rRNA) and N cycling bacteria marker genes amoA , narG , nirK, and nosZ, and soil physicochemical properties. 2. to define field scale spatial distribution patterns of total bacteria and soil N cycling bacteria gene concentrations and soil properties. 3. to describe relationships between soil N cycling bacteria gene concentrations and soil properties.
2. Materials and Methods
2.1. Site management and experimental setup
The experiment was conducted at Western Kentucky University Agriculture Research and Education Complex, Bowling Green, Kentucky, USA (36055′42″ N, 86028″6″ W) during 2016–2018 cropping seasons in three experimental fields each with 3.6 ha extent (Figure 1). The soils of the experimental fields were dominated by Crider silt loam (fine-silty, mixed, active, mesic Typic Paleudalf) with Nolin silt loam (Fine-silty, mixed, active, mesic Dystric Fluventic Eutrudepts), and vertrees silty clay loam (Fine, mixed, semiactive, mesic Typic Paleudalfs) soils. The climatic conditions of the experimental site are presented in Figure S1 (a-b). Before initiating the experiment, the entire field area had been planted to tall fescue pasture for beef cattle grazing. Detailed description of the experimental fields and management practices were previously reported [8].
Briefly, two experimental fields were converted to winter wheat and summer soybean double cropping. In fall 2016 and 2017, soft red winter wheat cultivar (cv. Branson) was planted on those two fields as wheat grain crop (WS) and wheat (forage grazing + grain) dual use crop (WGS). The wheat crop fields were fertilized with 250 kg N ha-1 (di ammonium phosphate) as spilt application in the spring. The original tall fescue (Festuca arundinacea; cv. Kentucky 31) pasture stand in the 3rd field was managed to near natural condition (TF) with 2-3 mow downs, stockpiling, and light grazing for 2-3 weeks in spring seasons. Two different sets of cow/calf pairs of Angus breed (each consisted of eight pairs with similar body weights) grazed the TF or WGS fields for 2-3 weeks in the spring seasons of 2017 and 2018. The WS and WGS fields produced similar wheat grain yield (4.6 and 4.1 MG ha-1) and soybean grain yield (4.3 and 4.2 MG ha-1) with no significant differences between the WGS and WS systems [8].
Twelve georeferenced locations spaced at 40 m within TF and WGS experimental fields and eleven georeferenced locations spaced similarly from the WS field were used for soil sampling (Figure 1). The measurements of total bacteria, abundance of nitrogen cycling bacteria genes, and soil physicochemical properties of each sampling location were considered as an independent replication (pseudo replicates) for statistical purpose [43,44]. Six soil samples were collected around each georeferenced location to 10 cm depth using a 19 mm diameter soil probe (Oakfield Apparatus Co., Fond du Lac, Wisconsin) and were then combined into a single composite sample to represent each location. The composite soil samples were divided into two sub-samples where one was stored at -80 °C until bacterial DNA extraction and the other portion was air dried at 250C and used for chemical analysis.
2.2. Soil physicochemical analysis
Soil pH, soil organic matter (OM), total C, total N, ammonium N (NH4–N), and nitrate N (NO3–N) contents were measured on twelve soil samples collected from the TF and WGS fields and eleven soil samples collected from the WS field . Soil pH was measured using a glass electrode with a 1:1 soil/water ratio. Soil organic matter content was approximated by loss on ignition (LOI) method [45]. High-temperature combustion in a Vario MAX C-N analyzer (Elementar America Inc.) with 2-g soil sample [46] was used to measure total soil C and N contents. The NH4–N and NO3–N concentrations were determined by potassium chloride extraction and flow-injection colorimetric analysis with cadmium reduction [47] on a Lachat Quickchem FIA+ 8000 analyzer (USA Hach Co.). Dry soil bulk density was measured [48] at each georeferenced soil sampling location using a 173.4 cm3 compact slide hammer corer (AMS Samplers, American Falls, Idaho).
2.3. Quantification of total bacteria and N Cycling bacteria genes in soil samples
Metagenomic DNA was extracted from 500 mg of soil using the FastDNA Spin kit for soils (MP Biomedical, Santa Ana, CA) according to the manufacturer’s instructions. Real time quantitative PCR (qPCR) was run on Bio-Rad CFX 96 real-time PCR detection system (BioRad, Hercules, CA) to quantify the concentrations of the targeted genes by using published primers, probes, and protocols (Table S1) as previously described [49]. The primers and probes were obtained from Integrated DNA Technologies, Inc. (Coralville, Iowa). The qPCR assay was performed with Qiagen HotStarTaq master mix (Qiagen, Valencia, CA) in a total reaction volume of 25 μL. The assay consisted of 3 mM MgCl2, 600 nm each of the forward and reverse primers, 200 nm of probe, and 10 ng of sample DNA or the standard (ranging from 102 to 108 copies). Sample DNA was diluted in 1:200 ratio to reduce the effect of potential PCR inhibitors in the soil. A total 5 μL of standard DNA for the standard and 5 μL of the diluted sample DNA were used as a template in the qPCR reaction. Total bacterial concentration (16S rRNA gene), NH3–oxidizing bacteria (amoA), and NO3-reducing bacteria (narG), NO2- reducing bacteria (nirK), and N2O reducing bacteria (nosZ) gene copy numbers were quantified at all sampling locations in each experimental field.
2.4. Statistical analysis, geostatistical modeling, and spatial mapping
The gene copy numbers determined per gram of soil were transformed to log10 scale before statistical analysis. The statistical ANOVA model accounted four repeated measurements (spring and fall, 2017 and 2018) of log gene copy numbers g-1 soil and soil physicochemical properties measured at the twelve sampling locations of TF, WWGS and eleven locations of the WS field. The cropping system treatment was considered as the between subject factor for the analysis. The log gene copy numbers g-1 and soil properties were considered as dependent variables. To quantify the relationship between soil properties, and bacteria and N cycling bacterial genes, Pearson’s correlation coefficients were calculated. Step up (P = 0.02) multiple regression analysis (Table S2) was performed to describe the variation of bacterial gene abundances explained by soil properties. Statistical analyses were performed by SPSS 28 (IBM Corporation, Armonk, NY, USA). Post-hoc pair-wise multiple comparisons were performed by Bonferoni’s method at 5% level of significance. The present study used a pseudo-replicated experimental design [43,44] .
The spatial distribution patterns of total bacteria and N cycling bacterial gene concentrations and soil pH in the three experimental fields were examined together. Moran’s I index [50] was used to assess the presence of spatial autocorrelation for the abundance of total and different N cycling bacterial genes. The variables were modeled geostatistically [51] and variogram analysis was performed by GS+ version 9 software (Gamma Design Software, Plainwell, MI, USA). The variogram parameters (Table S3) were then used to calculate linear unbiased estimates at un-sampled locations as a weighted average of neighboring sampled points. Ordinary kriging in ArcGIS 10.1 (Environmental Systems Research Institute; Redlands, CA) and Geostatistical Analyst extension were used to interpolate and map the gene copy numbers g-1 (log) and soil pH across the experimental fields.
3. Results
Repeated measure ANOVA procedure revealed that cropping systems and time (season) had significant (P < 0.05) interaction effect on soil properties except for NH4-N content and soil bulk density. Significant interaction between cropping systems and time dynamics has not been detected for the abundance of total and N cycling bacterial genes. Both cropping systems and time (season) factors significantly influenced the abundances of total bacteria and N cycling bacterial genes.
3.1. The initial soil physicochemical properties
The initial (i.e., after preparing the fields for the experiment and before sowing the winter wheat in fall 2016) soil physicochemical properties are shown in Table 1. The initial levels of soil pH, OM, TN were similar among the cropping systems, but the soil NO3 -N (4.2 vs. 1.8 mg kg-1) and NH4-N (27.3 vs. 21.8-22.6 mg kg-1) levels were significantly higher in the TF than the two fields subsequently planted to winter wheat crop. On the other hand, the soil bulk density in the WS and WGS were significantly (P < 0.05) higher than the TF (1.38 - 1.42 vs. 1.29 g cm-3). However, all bacterial gene concentrations were not different among the cropping systems at the beginning of the experiment.
3.2. Effect of cropping systems on soil physicochemical properties
Mean soil physicochemical properties of cropping systems for the experimental period are presented in Tables 2 & 3. In this experiment, both WS and WGS soils planted to winter wheat-summer soybean double cropping with and without grazing in spring had significantly (P<0.05)higher soil bulk density than the TF soil. However, there was no difference for soil bulk density between the WS and WGS soils (Table 2).
During the spring 2017 season soil pH did not vary significantly among the cropping systems but significant difference appeared with the advancement of the growing period. The TF soils reported consistently higher soil pH > 6.0 whereas the soil pH in the WS and WGS dropped to < 6.0 from the fall 2017. Most of the time the TF had significantly higher soil pH than the WS and WGS soils while both the WS and WGS soils exhibited similar pH values. Soil OM and total C contents among the cropping systems followed similar within season distribution patterns. In general, the TF soils were rich in soil OM (24.7 -33.4 g kg-1) and total C (20.5-36.5 gkg-1) contents than the WS and WGS soils (OM, 20.3-31.5 g kg-1; total C, 17.6-31.5 g kg-1). The OM and total C levels remained similar in the wheat- soybean soils; WS and WGS. The total N contents of cropping system varied (1.92- 4.46 gkg-1) with no clear trend for the within season variability. Soil NH4-N content was not different among the cropping systems and mean content ranged 9.5 -10.1 mg kg-1. There were mixed responses for the soil NO3-N concentrations among the cropping systems. The WS and WGS soils contained NO3-N levels similar (spring/fall 2017) to, or higher than (spring 2018) or lower than (fall 2018) the TF. The overall NO3-N content during the experimental period ranged 3.3 -11.5 mg kg-1.

Table 3.
Mean soil chemical properties of the cropping systems.
| n | Spring 2017 | Fall 2017 | Spring 2018 | Fall 2018 | Mean | |
| pH | ||||||
| TF | 12 | 6.33 ± 0.14 a | 6.37 ± 0.14a | 6.28± 0.13a | 6.51 ± 0.15a | |
| WS | 11 | 6.06 ± 0.13a | 5.80 ± 0.14b | 5.75 ± 0.13b | 5.66 ± 0.16b | |
| WGS | 12 | 6.05 ± 0.14a | 5.83 ± 0.14b | 5.83 ± 0.13b | 5.77 ± 0.15b | |
| ------------------------------------ g kg-1------------------------------------ | ||||||
| OM | ||||||
| TF | 12 | 26.40 ± 1.00a | 24.78 ± 1.02a | 32.13 ± 1.14a | 33.47 ± 1.06a | |
| WS | 11 | 27.65 ± 1.03a | 23.32 ± 1.07ab | 31.54 ± 1.19a | 30.30 ± 1.10ab | |
| WGS | 12 | 23.15 ± 0.99b | 20.30 ± 1.03b | 26.66 ± 1.14b | 27.12 ± 1.06b | |
| TC | ||||||
| TF | 12 | 20.50 ± 0.98a | 36.50 ± 1.63a | 33.80 ± 1.4a | 28.68 ± 1.07a | |
| WS | 11 | 22.09 ± 1.02a | 30.96 ± 1.70ab | 31.99 ± 1.02a | 27.17 ± 1.12a | |
| WGS | 12 | 17.60 ± 0.98b | 28.32 ± 0.95b | 29.95 ± 1.83a | 25.24 ± 1.07a | |
| TN | ||||||
| TF | 12 | 2.36 ± 0.09a | 3.45 ± 0.14a | 3.67 ± 0.19b | 2.59 ± 0.07a | |
| WS | 11 | 2.34 ± 0.09a | 2.92 ± 0.15ab | 4.46 ± 0.20a | 2.59 ± 0.07a | |
| WGS | 12 | 1.92 ± 0.09b | 2.82 ± 0.15b | 4.19 ± 0.19ab | 2.22 ± 0.07b | |
| ------------------------------------ mg kg-1------------------------------------ | ||||||
| NH4-N | ||||||
| TF | 48 | - | - | - | - | 9.56 ± 0.94a |
| WS | 44 | - | - | - | - | 10.1 ± 1.05a |
| WGS | 48 | - | - | - | - | 9.54 ± 1.0a |
| NO3-N | ||||||
| TF | 12 | 5.79 ± 1.13a | 8.49 ± 1.1a | 5.77 ± 0.94b | 6.56 ± 0.45a | |
| WS | 11 | 3.33 ± 1.18a | 9.47 ± 1.14a | 11.51 ± 0.94a | 4.72 ± 0.45b | |
| WGS | 12 | 4.06 ± 1.13a | 8.74 ± 1.10a | 10.35 ± 0.9a | 4.17 ± 0.47b | |
Mean ± SE, Numbers with different letters for each soil variable within columns (seasons) are significantly different (P < 0.05).
3.3. Effect of cropping systems on the abundance of total and soil nitrogen cycling bacterial marker genes.
The abundances of 16S rRNA, amoA, narG, nirK, and nosZ gene copy numbers of WS, WGS, and TF treatments are presented in the Table 4. The total bacterial gene abundances among the cropping systems demarcated at > log 9.0 level. The nitrification and denitrification gene levels ranged 6.4 –7.9 log level. In general, tall fescue soils carried significantly higher abundances of total bacteria and all N cycling bacterial genes compared to the wheat-soybean cropping soils. Of the denitrification genes, nosZ gene reported the highest concentration for all cropping systems but with statistically similar levels.
3.4. Relationship between total and soil nitrogen cycling bacterial gene abundances and soil chemical properties.
Several soil chemical properties established significant correlations with the abundances of total and N cycling genes in the TF and wheat-soybean cropping systems (Table 5). In the TF system, soil OM emerged as the key factor controlling the abundances of all bacterial genes (r = 0.32 - 0.46) and the secondary factors were soil pH (amoA and narG; r = 0.37-0.46), NO3-N (amoA and narG; r = 0.30-0.39), and the NH4-N (16S rRNA, nirK, and nosZ; r = 0.31-0.53). Soil pH dominated as the key factor controlling the abundances of all bacterial genes (r = 0.25 - 0.59) in the WS and WGS systems. The other soil factors that established significant correlations with the abundances of bacterial genes in the WS and WGS soils were soil OM (16S rRNA, amoA, and narG; r = 0.25 – 0.32) and NH4-N (nirK and nosZ: r = 0.35 – 0.42).
Step-up multiple regression analysis (Table S2) revealed that in the wheat-soybean systems, 12% of the variation in 16S rRNA gene abundance and 25 – 40% variation in the N cycling bacterial genes were explained by the soil properties. The soil properties in the TF explained 15% variation in the total bacteria and 22%- 35% of variation in the N cycling bacterial genes (Supplementary Table S2).
Table 5.
Correlations between soil properties and the abundances of bacterial genes in the three cropping systems.
Table 5.
Correlations between soil properties and the abundances of bacterial genes in the three cropping systems.
| Cropping system/ bacterial gene | n | pH | OM | NO3-N | NH4-N |
| TF | 48 | ||||
| 16S rRNA | 0.20 | 0.34** | 0.18 | 0.31* | |
| amoA | 0.37** | 0.32* | 0.39** | 0.14 | |
| narG | 0.46** | 0.46** | 0.30* | 0.17 | |
| nirK | 0.18 | 0.37** | 0.18 | 0.53** | |
| nosZ | 0.20 | 0.35* | 0.13 | 0.52** | |
| WS/WGS | 92 | ||||
| 16S rRNA | 0.25* | 0.30** | 0.13 | 0.17 | |
| amoA | 0.59* | 0.25* | 0.16 | 0.12 | |
| narG | 0.46** | 0.32** | 0.14 | 0.18 | |
| nirK | 0.50** | 0.12 | 0.03 | 0.42** | |
| nosZ | 0.59** | 0.06 | 0.04 | 0.35** |
Pearson’s correlation coefficients significant * P < 0.05 and ** P < 0.00.
3.5. Spatial distribution of total and N cycling bacterial genes and soil chemical properties
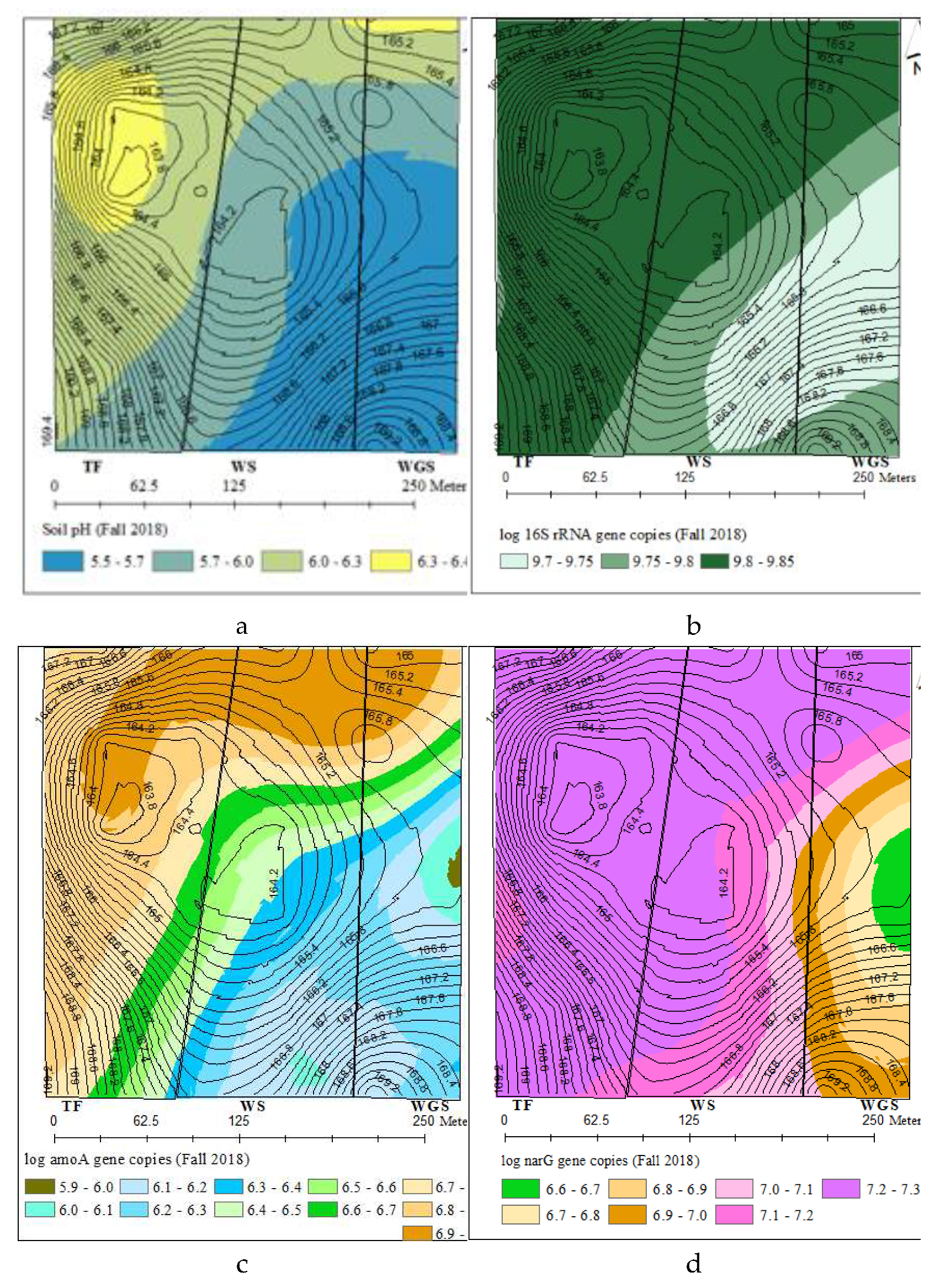
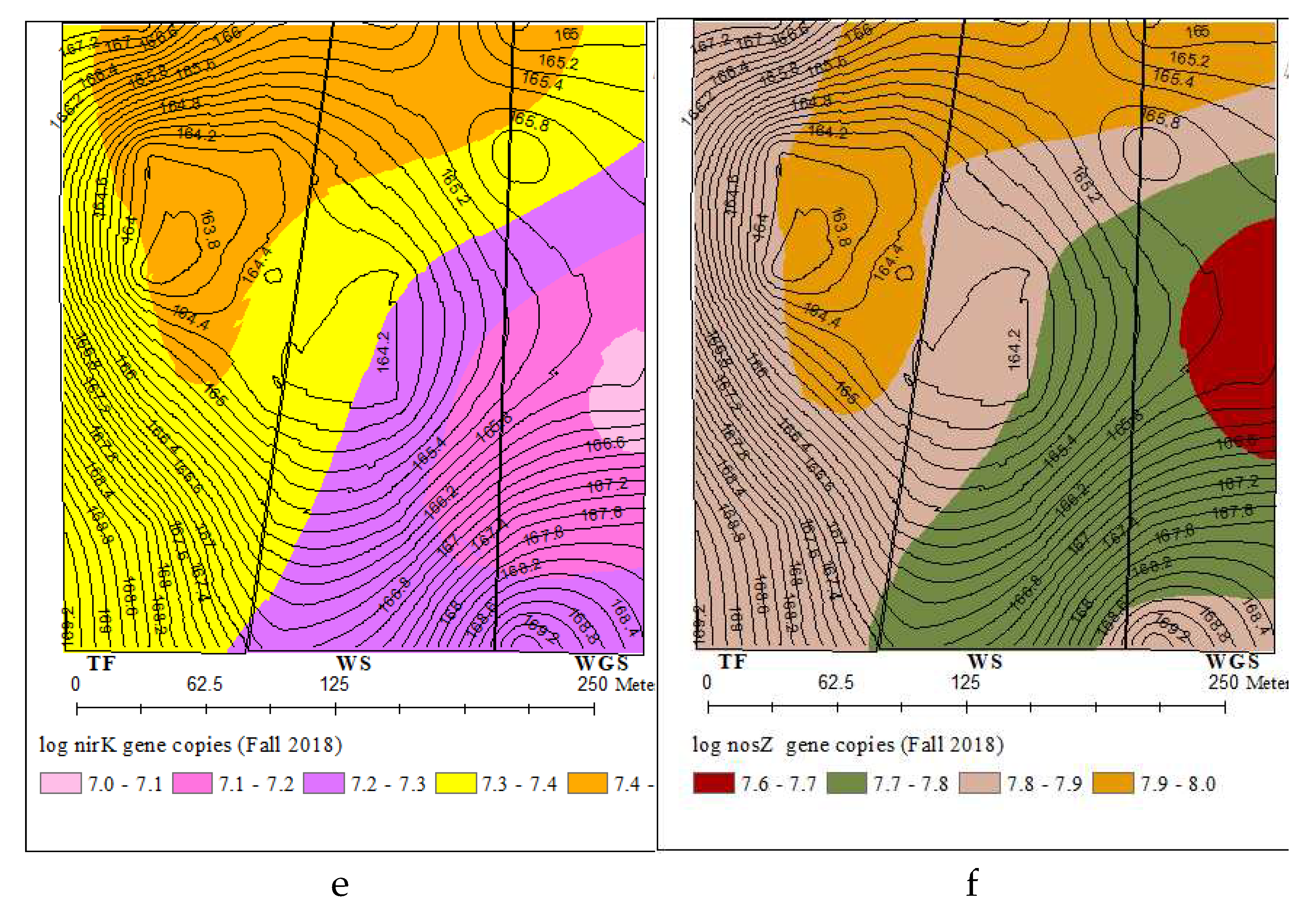
The spatial variability of total bacterial and N cycling bacterial gene abundances comprised over one order of magnitude (Figures 2b – 2f). The distribution of total bacteria (16SrRNA), nitrification (amoA) and denitrification (narG, nirK, and nosZ) genes described by different variogram models showed 141-187 m range spatial dependence (Table S.3). The kriged maps in Figures 2b-2f showed a south-east and north-west directional gradient for the distribution of all bacterial gene abundances. All bacterial genes were highly concentrated in the North and Northwest part of the landscape. Similarly soil pH also showed a spatial distribution pattern with high soil pH levels in the North and Northwest part of the landscape.
Figure 2.
Kriged maps for the spatial distributions of soil pH and bacterial genes across the three cropping systems. (a) soil pH, (b) 16S rRNA, (c) amoA, (d) narG, (e) nirK, (f) nosZ.
Figure 2.
Kriged maps for the spatial distributions of soil pH and bacterial genes across the three cropping systems. (a) soil pH, (b) 16S rRNA, (c) amoA, (d) narG, (e) nirK, (f) nosZ.
4. Discussion
Various management practices can have distinct influences on soil microbial communities in agricultural systems and their ecological functioning. Understanding ecology of bacterial communities responsible for nutrient cycling and their relationships with local soil physicochemical properties is beneficial for improving productivity while enhancing the sustainability of agricultural systems. This 2 yr. study examined the abundances and distribution of total bacteria(16S rRNA) and N cycling bacteria (amoA, narG, nirK, and nosZ) and soil properties of wheat– soybean cropping systems with 2-3 weeks of spring grazing (WGS) and without grazing (WS) and of tall fescue pasture system (TF) managed to near natural condition with similar grazing practice.
Before initiating the experiment, all the three experimental fields had same land use history of tall fescue pasturing for beef cattle grazing. The WS and WGS fields were grown to summer soybean crop before planting the first winter wheat crop for the experiment in fall 2016. We conclude that similar initial soil pH, OM, and TN contents among the WS, WGS, and the TF reflected the effects of uniform management history of the experimental fields. The more frequent use of farm machinery for various field operations of summer soybean cropping resulted in significantly higher soil bulk density[52] in the WS and WGS soils. In addition, active uptake of soil NH4-N and NO3-N by the summer soybean crop and low mineralization of organic N induced by high soil compaction[53] may have caused the depletion of pre experimental NH4-N and NO3-N levels in the WS and WGS compared to the TF. The similar initial fertility levels that existed in the cropping systems offered an equally favorable conditions for microbial growth, thus no difference was detected for the initial abundances of total bacteria and N cycling bacterial gene among the cropping systems.
The effects of different management disturbances of cropping systems were evident in this experiment. The TF soils that were managed to near natural conditions experienced with least disturbances (cultivation, fertilization, etc.) compared to the wheat-soybean cropping. Soil compaction, because of frequent cultivation is known to be an important problem that agriculture is facing [53]. Compaction disrupts soil’s physical integrity by modifying porosity and impeding gas, water, and nutrient movement and root growth in the soil profile [54]. In this experiment, both the WS and WGS soils that were planted to winter wheat-summer soybean double cropping were more exposed to farm machinery for cultivation compared to the near naturally managed TF soils. In the wheat-soybean cropping, the WGS soils experienced 2-3 weeks of cattle grazing on wheat in the spring seasons compared to the WS. The higher mean soil bulk density in the wheat-soybean cropping soils compared to the TF supported the effects of higher soil compaction by the extensive use of farm machinery for wheat-soybean cropping [52]. However, the additional exposure to cattle grazing for 2 -3 weeks in spring did not significantly increase soil bulk density of the WGS compared to the WS without grazing component. However, there was slightly higher soil bulk density in the WGS. The shorter duration (2-3 weeks) of the grazing period and the short lifespan for the effects of soil compaction[52] could have attributed the similar soil bulk density of the WS and WGS soils.
Different cropping and land use systems can pose significant effects on soil carbon and other soil nutrient contents, soil texture, and soil pH[55,56] arising mainly from differences in plant species and associated management practices. The winter wheat- summer soybean cropping especially the WGS had significantly different soil biogeochemical properties from the tall fescue pasturing (TF). The soil pH in the TF consistently remained > 6.0 throughout, most likely was due to the undisturbed and consistent near natural management practice. The drop in the soil pH in the WS and WGS to < 6.0 could be a result of ammonia-based fertilization to wheat-soybean cropping and to the soil acidification by ammonia fertilization[52]. The range of soil pH observed in the winter wheat-summer soybean soils was comparable to the levels (5.3 – 5.4) reported by [57] for wheat-soybean double crop soil. Low level of nitrification and release of small number of protons can elevate soil pH in more compacted than in less compacted soils [52]. Results for soil pH did not agree with[52] where more compacted WS and WGS soils (higher soil bulk density) were more acidic than the TF soils with lower soil bulk density. We suggest that soil compaction may not be a significant factor influencing soil pH in the WS and WGS systems, but it could be the ammonia-based N fertilization and resulting soil acidification[52].
Under the light near natural management, TF pasture growth before stockpiling was mow down to the soil. In the WS and WGS, winter wheat crop residues were removed from the fields in the form of bails and summer soybean crop residues were returned back to the soil. Thus there had been greater chance for higher input of crop residues to the TF soils compared to the WS and WGS. It has been shown that chemical composition of crop tissues influenced the decomposition rate of litter material [58,59]. The decomposition rate of plant residue is negatively correlated with the C: N ratio and hemicellulose content of plant tissues[58,60]. Soybean plant tissue has <15:1 C: N ratio and 100 g kg-1 DM hemicellulose content [59,60]. Analysis of crop residues from this study revealed (data not presented) that soybean crop residue contained 102-113 g kg-1 DM hemicellulose and tall fescue pasture tissue had 200-226 g kg-1 DM hemicellulose content. Based on the hemicellulose contents we expect higher decomposition potential for soybean crop residue than the TF pasture plant parts. Accordingly, we would expect that soils of WS and WGS to contain higher amounts of soil OM than the TF. In addition, soil compaction by farm machinery and grazing cattle resulted in poor aeration and hampered the mineralization of soil OM [53]. All the above facts suggest that wheat-soybean crop soils to contain higher soil OM than the TF soils. However, contrasting result from this study revealed that TF soils have higher soil OM levels than the WS and WGS soils. We postulate that loss of soil organic matter by cultivation [14] may have attributed the lower levels of soil OM detected in the WS and WGS soils. Thus, adoption of soil management practices with minimal disturbances would be beneficial to improve the soil OM levels in the wheat- soybean cropping systems.
Total soil C content among the cropping systems within seasons varied in a manner similar to the soil OM content. In general, the TF soils were rich in total C than the WS and WGS soils. The higher total C level in TF could be explained by the associated higher soil OM levels. the lower levels of total soil C in the WS and WGS soils could be explained by the potential microbial burning of soil C induced by N fertilization [61]. However, the levels of total C (25.2 – 31.5 g kg-1) reported in the wheat-soybean cropping soils of this experiment was 3-4 times higher than the 8.08-8.34 g kg-1 reported for winter wheat and soybean summer rotations [57].
The seasonal total soil N concentration variability was inconsistent among the cropping systems. In general, total N levels were similar between the TF and WS soils, but the levels in the TF was significantly (P>0.05) higher than the WGS soils, especially during the fall seasons. There is a greater potential for denitrification in the more compacted soil [62]. The slightly higher soil bulk density of the WGS soils and potentially higher denitrification losses could support the consistently lower total soil N concentration in the WGS soil compared to the WS soils. The concentration of total soil N observed in this experiment (0.5-0.94 g kg-1) was 1-2 times higher than the 2.22-2.92 g kg-1 previously reported for wheat-soybean crop rotations [57]. In general oxidation of ammonia is reduced in acidic soil condition [23] because of the exponential reduction in NH3 availability with decreasing pH, through ionization to NH4+ [63]. On the other hand, mineralization of organic N to NH4+ is less favored in compacted soils [53]. We suggest that soil pH and soil compaction collectively attributed the similar levels of soil NH4-N detected in all the cropping systems. The levels of soil NH4-N (9.5 -10.1 g kg-1) detected in the WS and WGS were comparable to the 8.0 g kg-1 reported by [57] for wheat-soybean crop rotations.
Soil compaction can affect denitrification mainly through limited supply of soil aeration and by the indirect effects on N and C transformation. Soil compaction reduces soil pore diameter, increases water-filled pore space which in turn restricts oxygen diffusion within the soil leading to denitrification [62]. Because of higher soil bulk density, we would expect higher denitrification and lower level of NO3- N in the WS and WGS soils compared to the TF soils. However, this scenario was evident only in the fall 2018 season. There had been mixed responses during rest of the seasons, where the WS and WGS soils contained NO3 level similar to (spring/fall 2017) or higher than (fall 2018) the TF. These mixed responses could have resulted from the seasonal climatic differences, management practices (N fertilization), and the crops that occupied. In this experiment we noticed 3.3-11.5 mg kg-1 soil NO3-N in the WS and WGS that was comparable to the levels (6.5-11.3 mg kg-1) reported by [57] for same crop rotation. Altogether tall fescue pasture system managed to near natural management conditions had higher soil pH, OM, and C concentration compared to the wheat-soybean cropping systems either with or without light spring grazing. However, the soil N dynamics in the tall fescue and wheat- soybean cropping systems are complex, unpredictable, and more likely be affected by management practices and local environmental factors.
Greater heterogeneity of above ground crop residues, senescent roots, and root exudates in agriculture systems along with other management practices can create more variable habitable resource niches in soil [60] , thus agriculture systems can have different soil microbial properties [64,65]. The similarity of cropping history (under tall fescue pasturing) resulted in similar initial levels of 16S rRNA, amoA, narG, nirK, and nosZ gene copies among the cropping systems. Although previous research [66,67] has shown higher microbial abundance in frequently disturbed soils than in weakly disturbed, it was noted that the TF soils with least disturbances (due to near natural management) had comparatively higher abundances of total and N cycling bacteria compared to the WS and WGS systems. The abundance of N cycling bacteria in some environments can be related to substrate availability [28]. Higher soil C storage provide a benign dwelling while supplying substrates for soil microbes [68] . Organic C compounds are suitable electron donors for biological metabolism and increase of organic C can stimulate the abundance of ammonia oxidizing and denitrifying bacteria [17,69]. We suspect that higher availability of labile organic substrate [70,71] from the OM in the TF system might have resulted in higher abundance of all bacteria. The mean 16S rRNA gene abundance observed in this experiment is consistent with the levels (108 -109) reported by [26] for agricultural soils.
Autotropic ammonia-oxidizing bacteria have shown growing faster in neutral or slightly alkaline media [23]. The TF field managed to near natural condition had comparatively higher soil pH close to neutral; reported higher abundances of amoA gene (log 7.01; 1.0 x 107) than the more acidic WS/WGS wheat-soybean systems. The level of amoA genes found in the TF was comparable to those previously reported by [72] for unfertilized arable soils. Some studies have shown that chemical fertilization increased abundance of ammonia-oxidizing bacteria [73], while negative impacts of chemical fertilization have also been observed [74,75] . The lower levels of amoA gene detected in the WS and WGS could have been derived from the low soil pH resultant from NH4 based N fertilization. The lower levels of amoA gene copies in the WS and WGS further indicate the low dependency of wheat-soybean cropping on biological nitrification. Both WS and WGS systems harbored similar concentrations of amoA genes and the levels were consistent with the concentrations found in fertilized soils [72].
Although, some of the between group differences were not significant abundance of all denitrification genes followed a general trend TF > WS > WGS and mimicked the trend observed for soil OM and total C variations [76]. Recent studies [77,78] have shown that some denitrifiers can have a truncated denitrification pathway and lack the nosZ gene encoding nitrous oxide reductase. However, in this experiment the nosZ gene accounted for the highest abundance among the denitrifiers. The nirK and nosZ gene levels detected in this experiment were consistent with the levels for agricultural soils planted to winter wheat, wheat, and corn crops [26]. There is evidence that cattle grazing can alter soil physical properties and conditions affecting the size of communities and microbial processes [54]. In addition, cattle grazing can modify nutrient availability through the deposition of urine and feces and stimulate denitrification. However, the effect of change of soil properties by cattle grazing on the abundance of denitrifying bacterial communities was not evident in this experiment. Both WS and WGS harbored similar concentrations of denitrifying bacteria. We presume that 2-3 weeks of grazing period did not alter soil physical or chemical properties in the WGS considerably to show any significant change in the levels of the denitrifying bacterial concentrations.
The significant correlations that existed for soil pH in the WS and WGS and soil OM contents in the WS and WGS with the abundance of all N cycling bacteria genes [17,73,76,79] suggests that different management practices in the cropping systems had influenced the abundance of N cycling bacteria gene concentrations. Frequent mow down of tall fescue pasture in the TF that added more soil organic matter and N fertilization (ammonium fertilizer) to wheat-soybean cropping that reduced soil pH could be identified as such management practices.
Characterization of bacterial distribution patterns at field-scale is important for understanding the ecology of bacterial communities at a scale compatible with land management strategies. The spatial variability of total and N cycling bacterial concentrations comprised over one order of magnitude with 141-187 m range spatial dependence and low nugget effects were consistent with the previous work by [39,41]. The range of spatial dependence observed for N cycling bacterial genes in this experiment was similar to the 130-140m range spatial auto correlation reported for ammonia oxidizing bacteria across a 44 ha field [80]. The similar spatial distribution patterns of total and N cycling bacteria indicates that the abundances of all bacterial genes were controlled by same edaphic factor/s. The significant correlation and similarity of kriged maps for N cycling bacterial concentration and soil pH further confirmed soil pH as one of the influential factors controlling the abundance of bacteria at field scale. There may exist consistent favorable niches for bacteria growth in the North and Northwest part of the landscape to harbor higher bacterial concentrations. The management history of experimental fields showed that regular placement of supplementary feeders for grazing cattle occurred in the north- northeast parts. The more frequent animal roaming, higher animal congregation, and intensive manure and urine input to soils would have favored the bacterial growth in those areas. These results altogether indicated that the abundance of bacteria involved in soil N cycling are driven mainly by local environmental gradients that can be controlled by local management practices.
5. Conclusions
Quantification of bacteria involved in soil N transformations in agricultural systems helps expand our understanding on response of N- cycling microbial population to respective environmental disturbances. This study quantified soil physicochemical properties and abundance of 16S rRNA gene, N cycling bacterial genes, amoA, narG, nirK, and nosZ in wheat – soybean crop rotations with 2-3 weeks of spring grazing (WGS) and without grazing (WS) and in tall fescue pasturing system managed to near natural condition with similar grazing (TF). We explored the influence of three cropping systems on the abundance of total and N cycling bacteria and soil physicochemical properties. In addition, we examined the relationships between soil properties and the abundance of bacteria and described field scale spatial distribution patterns. There existed a marked contrast of abundance of total and N cycling bacteria and soil properties between the cropping systems with higher abundances in the tall fescue pasture soils managed to near natural conditions. Soil OM and soil pH emerged as key factors to influence total soil bacteria and targeted N cycling gene abundances in the wheat-soybean and tall fescue pasture systems. Abundances of all bacteria genes of tall fescue and wheat-soybean cropping system soils were controlled by same edaphic factor/s which includes soil pH. There exists a potential to manipulate the abundance of N cycling bacteria by devising field scale management strategies targeting influential soil properties.
Supplementary Materials
Table S1: Quantitative real-time PCR assays; Table S2: Summary of step-up multiple linear regression between abundance of nitrogen cycling bacterial genes and soil properties (n=48); Table S3: Semi-variogram parameters of total and nitrogen cycling bacteria and mean soil pH ; Figure S1 (a-b): Climatic conditions of the site during the experimental period.
Author Contributions
Conceptualization, H.G. and A.N.; methodology, H.G., A.N, and G.A.; software, A.N.; formal analysis, A.N, G.A.; investigation, H.G, A.N, G.A., P.G, and K.R.; writing—original draft preparation, A.N.; writing—review and editing, H.G., A.N., G.A., P.G.,and K.R.; project administration, H.G. and P.G; funding acquisition, H.G.
Funding
This research was funded by the United States Department of Agriculture, through the USDA-ARS; Western Kentucky University cooperative research program (Project Nos. 5040-12630-006-00-D and 5040-12630-006-00-S).
Acknowledgments
We thank Mr. Rohan Parekh (U.S. Department of Agriculture, Agricultural Research Service, Food Animal Environmental Systems Research Unit, Bowling Green, Kentucky) for the technical assistance in microbial analysis laboratory work.
References
- Sanford: G., R.; Jackson, R. D.; Booth, E. G.; Hedtcke, J. L.; Picasso, V. , Perenniality and diversity drive output stability and resilience in a 26-year cropping systems experiment. Field Crops Research 2021, 263, 108071. [Google Scholar] [CrossRef]
- Smith, P. , Delivering food security without increasing pressure on land. Global food security 2013, 2(1), 18–23. [Google Scholar] [CrossRef]
- Lemaire, G.; Franzluebbers, A.; de Faccio Carvalho, P. C.; Dedieu, B., Integrated crop–livestock systems: Strategies to achieve synergy between agricultural production and environmental quality. Agriculture, Ecosystems & Environment 2014, 190, 4-8.
- Planisich, A.; Utsumi, S.; Larripa, M.; Galli, J. , Grazing of cover crops in integrated crop-livestock systems. Animal 2021, 15(1), 100054. [Google Scholar] [CrossRef] [PubMed]
- Moomaw, R. S.; Powell, T. A. , Multiple cropping systems in small grains in Northeast Nebraska. Journal of production agriculture 1990, 3(4), 569–576. [Google Scholar] [CrossRef]
- Horn, F. In Chemical composition of wheat pasture, Natl. Wheat Pasture Symp. Proc. GW Horn, ed. Oklahoma Agric. Exp. Stn., Stillwater, 1984; pp 47-54.
- Winterholler, S.; Lalman, D.; Hudson, M.; Ward, C.; Krehbiel, C.; Horn, G. , Performance, carcass characteristics, and economic analysis of calf-fed and wheat pasture yearling systems in the southern Great Plains. The Professional Animal Scientist 2008, 24(3), 232–238. [Google Scholar] [CrossRef]
- Netthisinghe, A.; Galloway, H.; DeGraves, F.; Agga, G. E.; Sistani, K. , Grain yield and beef cow–calf growth performance in dual-purpose and conventional grain wheat production systems and stockpiled tall fescue pasturing. Agronomy 2020, 10(10), 1543. [Google Scholar] [CrossRef]
- Kourtev, P.; Ehrenfeld, J.; Häggblom, M. , Experimental analysis of the effect of exotic and native plant species on the structure and function of soil microbial communities. Soil Biology and Biochemistry 2003, 35(7), 895–905. [Google Scholar] [CrossRef]
- Stuedemann, J. A.; Hoveland, C. S. , Fescue endophyte: History and impact on animal agriculture. Journal of Production Agriculture 1988, 1(1), 39–44. [Google Scholar] [CrossRef]
- Hoveland, C. S., Origin and history. Tall fescue for the twenty-first century 2009, 53, 1-10.
- Leuchtmann, A.; Clay, K. , Isozyme variation in the Acremonium/Epichloë fungal endophyte complex. Phytopathology 1990, 80(10), 1133–9. [Google Scholar] [CrossRef]
- Horvatic, J.; Peršić, V.; Kočić, A.; Čačić, L.; Has-Schoen, E. , Water quality and nutrient limitation in an area of the Danube River and an adjoining oxbow lake (1299 r. km): algal bioassay. Fresenius Environmental Bulletin 2009, 18 (1), 12-20.
- Tripathi, N.; Singh, R. S. , Influence of different land uses on soil nitrogen transformations after conversion from an Indian dry tropical forest. Catena 2009, 77(3), 216–223. [Google Scholar] [CrossRef]
- Orwin, K. H.; Wardle, D. A. , New indices for quantifying the resistance and resilience of soil biota to exogenous disturbances. Soil Biology and Biochemistry 2004, 36(11), 1907–1912. [Google Scholar] [CrossRef]
- Lang, M.; Cai, Z.-C.; Mary, B.; Hao, X.; Chang, S. X. , Land-use type and temperature affect gross nitrogen transformation rates in Chinese and Canadian soils. Plant and soil 2010, 334, 377–389. [Google Scholar] [CrossRef]
- Wakelin, S. A.; Gregg, A. L.; Simpson, R. J.; Li, G. D.; Riley, I. T.; McKay, A. C. , Pasture management clearly affects soil microbial community structure and N-cycling bacteria. Pedobiologia 2009, 52(4), 237–251. [Google Scholar] [CrossRef]
- Hayatsu, M.; Tago, K.; Saito, M. , Various players in the nitrogen cycle: diversity and functions of the microorganisms involved in nitrification and denitrification. Soil Science and Plant Nutrition 2008, 54(1), 33–45. [Google Scholar] [CrossRef]
- Reddy, K.; Patrick, W.; Broadbent, F. , Nitrogen transformations and loss in flooded soils and sediments. Critical Reviews in Environmental Science and Technology 1984, 13(4), 273–309. [Google Scholar] [CrossRef]
- Bolan, N. S.; Saggar, S.; Luo, J.; Bhandral, R.; Singh, J. , Gaseous emissions of nitrogen from grazed pastures: processes, measurements and modeling, environmental implications, and mitigation. Advances in agronomy 2004, 84(37), 120. [Google Scholar]
- Lashof, D. A.; Ahuja, D. R. , Relative contributions of greenhouse gas emissions to global warming. Nature 1990, 344(6266), 529–531. [Google Scholar] [CrossRef]
- Rotthauwe, J.-H.; Witzel, K.-P.; Liesack, W. , The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Applied and environmental microbiology 1997, 63(12), 4704–4712. [Google Scholar] [CrossRef]
- De Boer, W.; Kowalchuk, G. A. , Nitrification in acid soils: micro-organisms and mechanisms. Soil Biology and Biochemistry 2001, 33 (7-8), 853-866.
- Zumft, W. G. , Cell biology and molecular basis of denitrification. Microbiology and molecular biology reviews 1997, 61(4), 533–616. [Google Scholar]
- Chèneby, D.; Hallet, S.; Mondon, M.; Martin-Laurent, F.; Germon, J.; Philippot, L. , Genetic characterization of the nitrate reducing community based on narG nucleotide sequence analysis. Microbial ecology 2003, 46(1), 113–121. [Google Scholar] [CrossRef]
- Henry, S.; Bru, D.; Stres, B.; Hallet, S.; Philippot, L. , Quantitative detection of the nosZ gene, encoding nitrous oxide reductase, and comparison of the abundances of 16S rRNA, narG, nirK, and nosZ genes in soils. Applied and environmental microbiology 2006, 72(8), 5181–5189. [Google Scholar] [CrossRef] [PubMed]
- López-Gutiérrez, J. C.; Henry, S.; Hallet, S.; Martin-Laurent, F.; Catroux, G.; Philippot, L. , Quantification of a novel group of nitrate-reducing bacteria in the environment by real-time PCR. Journal of microbiological methods 2004, 57(3), 399–407. [Google Scholar] [CrossRef] [PubMed]
- Colloff, M.; Wakelin, S.; Gomez, D.; Rogers, S. , Detection of nitrogen cycle genes in soils for measuring the effects of changes in land use and management. Soil Biology and Biochemistry 2008, 40(7), 1637–1645. [Google Scholar] [CrossRef]
- Nicol, G. W.; Leininger, S.; Schleper, C.; Prosser, J. I. , The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environmental microbiology 2008, 10(11), 2966–2978. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, H. L.; Packer, A.; Bever, J. D.; Clay, K. , Grassroots ecology: plant–microbe–soil interactions as drivers of plant community structure and dynamics. Ecology 2003, 84(9), 2281–2291. [Google Scholar] [CrossRef]
- Girvan, M. S.; Bullimore, J.; Pretty, J. N.; Osborn, A. M.; Ball, A. S. , Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soils. Applied and environmental microbiology 2003, 69(3), 1800–1809. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R. B. , The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences 2006, 103(3), 626–631. [Google Scholar] [CrossRef]
- Frey, S. D.; Knorr, M.; Parrent, J. L.; Simpson, R. T. , Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests. Forest Ecology and Management 2004, 196(1), 159–171. [Google Scholar] [CrossRef]
- Hayden, H. L.; Drake, J.; Imhof, M.; Oxley, A. P.; Norng, S.; Mele, P. M. , The abundance of nitrogen cycle genes amoA and nifH depends on land-uses and soil types in South-Eastern Australia. Soil Biology and Biochemistry 2010, 42(10), 1774–1783. [Google Scholar] [CrossRef]
- Wallenstein, M. D.; Myrold, D. D.; Firestone, M.; Voytek, M. , Environmental controls on denitrifying communities and denitrification rates: insights from molecular methods. Ecological applications 2006, 16(6), 2143–2152. [Google Scholar] [CrossRef]
- Bissett, A.; Brown, M. V.; Siciliano, S. D.; Thrall, P. H. , Microbial community responses to anthropogenically induced environmental change: towards a systems approach. Ecology letters 2013, 16, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C. L.; Strickland, M. S.; Bradford, M. A.; Fierer, N. , The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biology and Biochemistry 2008, 40(9), 2407–2415. [Google Scholar] [CrossRef]
- Osborne, C. A.; Zwart, A. B.; Broadhurst, L. M.; Young, A. G.; Richardson, A. E. , The influence of sampling strategies and spatial variation on the detected soil bacterial communities under three different land-use types. FEMS Microbiology Ecology 2011, 78(1), 70–79. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R. B.; Mills, A. L. , Multi-scale variation in spatial heterogeneity for microbial community structure in an eastern Virginia agricultural field. FEMS microbiology ecology 2003, 44(3), 335–346. [Google Scholar] [CrossRef] [PubMed]
- Nunan, N.; Wu, K.; Young, I. M.; Crawford, J. W.; Ritz, K. , In situ spatial patterns of soil bacterial populations, mapped at multiple scales, in an arable soil. Microbial Ecology 2002, 44, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Ritz, K.; McNicol, J.; Nunan, N.; Grayston, S.; Millard, P.; Atkinson, D.; Gollotte, A.; Habeshaw, D.; Boag, B.; Clegg, C. , Spatial structure in soil chemical and microbiological properties in an upland grassland. FEMS Microbiology Ecology 2004, 49(2), 191–205. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V. , Soil Biology in Pasture Systems: Knowledge and Opportunity Audit. Meat and Livestock Australia: 2003.
- Hurlbert, S. H. , Pseudoreplication and the design of ecological field experiments. Ecological monographs 1984, 54(2), 187–211. [Google Scholar] [CrossRef]
- Wester, D. B., replication, randomization, and statistics in range research. Rangeland Ecology & Management/Journal of Range Management Archives 1992, 45 (3), 285-290.
- Rowell, D., Laboratory methods for studying mineralization. Soil science: Methods and Applications. Longman Scientific and Technical, Longman Group UK Ltd., Longman House, London, England 1994.
- Nelson, D. a.; Sommers, L. E., Total carbon, organic carbon, and organic matter. Methods of soil analysis: Part 2 chemical and microbiological properties 1983, 9, 539-579.
- Mulvaney, R. L., Nitrogen—inorganic forms. Methods of soil analysis: Part 3 Chemical methods 1996, 5, 1123-1184. 1123.
- Blake, G. ꎬ.; Hartge, K., Bulk density. Methods of soil analysis: Part 1 Physical and mineralogical methods 1986, 5, 363-375.
- Cook, K.; Ritchey, E.; Loughrin, J.; Haley, M.; Sistani, K.; Bolster, C. H. , Effect of turning frequency and season on composting materials from swine high-rise facilities. Waste Management 2015, 39, 86–95. [Google Scholar] [CrossRef]
- Moran, P. A., Notes on continuous stochastic phenomena. Biometrika 1950, 37 (1/2), 17-23.
- Isaaks, E. H.; Srivastava, R. M. , Applied geostatistics. Oxford university press New York: 1989; Vol. 561.
- Bhandral, R.; Saggar, S.; Bolan, N.; Hedley, M. , Transformation of nitrogen and nitrous oxide emission from grassland soils as affected by compaction. Soil and Tillage Research 2007, 94(2), 482–492. [Google Scholar] [CrossRef]
- De Neve, S.; Hofman, G. , Influence of soil compaction on carbon and nitrogen mineralization of soil organic matter and crop residues. Biology and fertility of soils 2000, 30, 544–549. [Google Scholar] [CrossRef]
- Greacen, E. L.; Sands, R. , Compaction of forest soils. A review. Soil Research 1980, 18(2), 163–189. [Google Scholar] [CrossRef]
- Murty, D.; Kirschbaum, M. U.; Mcmurtrie, R. E.; Mcgilvray, H. , Does conversion of forest to agricultural land change soil carbon and nitrogen? A review of the literature. Global change biology 2002, 8(2), 105–123. [Google Scholar] [CrossRef]
- Post, W. M.; Mann, L. Changes in soil organic carbon and nitrogen as a result of cultivation; Environmental System Science Data Infrastructure for a Virtual Ecosystem …: 2005.
- Sun, R.; Guo, X.; Wang, D.; Chu, H. , Effects of long-term application of chemical and organic fertilizers on the abundance of microbial communities involved in the nitrogen cycle. Applied Soil Ecology 2015, 95, 171–178. [Google Scholar] [CrossRef]
- Beyaert, R.; Paul Voroney, R. , Estimation of decay constants for crop residues measured over 15 years in conventional and reduced tillage systems in a coarse-textured soil in southern Ontario. Canadian journal of soil science 2011, 91(6), 985–995. [Google Scholar] [CrossRef]
- Broder, M.; Wagner, G. , Microbial colonization and decomposition of corn, wheat, and soybean residue. Soil Science Society of America Journal 1988, 52(1), 112–117. [Google Scholar] [CrossRef]
- McDaniel, M.; Grandy, A.; Tiemann, L.; Weintraub, M. , Crop rotation complexity regulates the decomposition of high and low quality residues. Soil Biology and Biochemistry 2014, 78, 243–254. [Google Scholar] [CrossRef]
- Williams, D. L.; Ineson, P.; Coward, P. , Temporal variations in nitrous oxide fluxes from urine-affected grassland. Soil Biology and Biochemistry 1999, 31(5), 779–788. [Google Scholar] [CrossRef]
- Rasiah, V.; Kay, B. , Legume N mineralization: effect of aeration and size distribution of water-filled pores. Soil Biology and Biochemistry 1998, 30(1), 89–96. [Google Scholar] [CrossRef]
- Frijlink, M. J.; Abee, T.; Laanbroek, H. J.; de Boer, W.; Konings, W. N. , The bioenergetics of ammonia and hydroxylamine oxidation in Nitrosomonas europaea at acid and alkaline pH. Archives of Microbiology 1992, 157, 194–199. [Google Scholar] [CrossRef]
- Ashworth, A.; DeBruyn, J.; Allen, F.; Radosevich, M.; Owens, P. , Microbial community structure is affected by cropping sequences and poultry litter under long-term no-tillage. Soil Biology and Biochemistry 2017, 114, 210–219. [Google Scholar] [CrossRef]
- Lauber, C. L.; Ramirez, K. S.; Aanderud, Z.; Lennon, J.; Fierer, N. , Temporal variability in soil microbial communities across land-use types. The ISME journal 2013, 7(8), 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.; Leemans, D.; Cook, R.; Hobbs, P. J. , Seasonality of the soil biota of grazed and ungrazed hill grasslands. Soil biology and Biochemistry 1997, 29(8), 1285–1294. [Google Scholar] [CrossRef]
- Bruns, M. A.; Stephen, J. R.; Kowalchuk, G. A.; Prosser, J. I.; Paul, E. A. , Comparative diversity of ammonia oxidizer 16S rRNA gene sequences in native, tilled, and successional soils. Applied and Environmental Microbiology 1999, 65(7), 2994–3000. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Martínez, K.; Suttle, K. B.; Brodie, E. L.; Power, M. E.; Andersen, G. L.; Banfield, J. F. , Despite strong seasonal responses, soil microbial consortia are more resilient to long-term changes in rainfall than overlying grassland. The ISME journal 2009, 3(6), 738–744. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Hallin, S.; Schloter, M. , Ecology of denitrifying prokaryotes in agricultural soil. Advances in agronomy 2007, 96, 249–305. [Google Scholar]
- Bardgett, R. D.; Wardle, D. A. , Herbivore-mediated linkages between aboveground and belowground communities. Ecology 2003, 84(9), 2258–2268. [Google Scholar] [CrossRef]
- Gaillard, V.; Chenu, C.; Recous, S.; Richard, G. , Carbon, nitrogen and microbial gradients induced by plant residues decomposing in soil. European Journal of Soil Science 1999, 50(4), 567–578. [Google Scholar] [CrossRef]
- Hermansson, A.; Lindgren, P.-E. , Quantification of ammonia-oxidizing bacteria in arable soil by real-time PCR. Applied and environmental microbiology 2001, 67(2), 972–976. [Google Scholar] [CrossRef]
- Fan, F.; Yang, Q.; Li, Z.; Wei, D.; Cui, X. a.; Liang, Y. , Impacts of organic and inorganic fertilizers on nitrification in a cold climate soil are linked to the bacterial ammonia oxidizer community. Microbial ecology 2011, 62, 982–990. [Google Scholar] [CrossRef]
- He, J. z.; Shen, J. p.; Zhang, L. m.; Zhu, Y. g.; Zheng, Y. m.; Xu, M. g.; Di, H. , Quantitative analyses of the abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices. Environmental microbiology 2007, 9(9), 2364–2374. [Google Scholar] [CrossRef]
- McAndrew, D.; Malhi, S. , Long-term N fertilization of a solonetzic soil: effects on chemical and biological properties. Soil Biology and Biochemistry 1992, 24(7), 619–623. [Google Scholar] [CrossRef]
- Miller, M.; Zebarth, B.; Dandie, C.; Burton, D.; Goyer, C.; Trevors, J. , Crop residue influence on denitrification, N2O emissions and denitrifier community abundance in soil. Soil Biology and Biochemistry 2008, 40(10), 2553–2562. [Google Scholar] [CrossRef]
- Jones, C. M.; Stres, B.; Rosenquist, M.; Hallin, S. , Phylogenetic analysis of nitrite, nitric oxide, and nitrous oxide respiratory enzymes reveal a complex evolutionary history for denitrification. Molecular biology and evolution 2008, 25(9), 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L., Denitrifying genes in bacterial and archaeal genomes. Biochimica et biophysica acta (BBA)-Gene structure and expression 2002, 1577 (3), 355-376.
- Bru, D.; Ramette, A.; Saby, N.; Dequiedt, S.; Ranjard, L.; Jolivet, C.; Arrouays, D.; Philippot, L. , Determinants of the distribution of nitrogen-cycling microbial communities at the landscape scale. The ISME journal 2011, 5(3), 532–542. [Google Scholar] [CrossRef]
- Wessén, E.; Söderström, M.; Stenberg, M.; Bru, D.; Hellman, M.; Welsh, A.; Thomsen, F.; Klemedtson, L.; Philippot, L.; Hallin, S. , Spatial distribution of ammonia-oxidizing bacteria and archaea across a 44-hectare farm related to ecosystem functioning. The ISME journal 2011, 5(7), 1213–1225. [Google Scholar] [CrossRef]
Figure 1.
The three experimental fields (tall fescue, winter wheat plus soybeans double cropping without grazing, and winter wheat plus soybeans double cropping with grazing) and geo-referenced soil sampling locations.
Figure 1.
The three experimental fields (tall fescue, winter wheat plus soybeans double cropping without grazing, and winter wheat plus soybeans double cropping with grazing) and geo-referenced soil sampling locations.

Table 1.
Mean initial (Fall 2016) levels of soil physicochemical properties, total bacteria, and N cycling bacterial gene concentrations at the experimental sites.
Table 1.
Mean initial (Fall 2016) levels of soil physicochemical properties, total bacteria, and N cycling bacterial gene concentrations at the experimental sites.
| TF | WS | WGS | |
| n | 12 | 11 | 12 |
| Soil properties | |||
| Bulk density (g cm-3) | 1.29 ± 0.03a | 1.38 ± 0.04ab | 1.42 ± 0.02b |
| pH | 6.62± 0.16a | 5.76 ± 0.29a | 5.81 ± 0.25a |
| ------------------------------- g kg-1------------------------------- | |||
| OM | 27.4 ± 1.62a | 24.9 ± 1.38a | 24.55 ± 0.86a |
| TN | 2.85 ± 0.09a | 2.96 ± 0.09a | 2.72 ± 0.08a |
| ------------------------------ mg kg-1------------------------------ | |||
| NO3-N | 4.19 ± 0.51a | 1.77 ± 0.24b | 1.82 ± 0.29b |
| NH4-N | 27.35 ± 1.77a | 22.6 ± 0.77b | 21.78 ± 1.01b |
| Bacterial genes | ----------------------- log10 gene copies ----------------------- | ||
| 16S rRNA | 10.07 ± 0.04a | 9.95 ± 0.04a | 9.94 ± 0.04a |
| amoA | 7.13 ± 0.10a | 6.64 ± 0.18a | 6.64 ± 0.11a |
| narG | 7.57 ± 0.12a | 7.13 ± 0.17a | 7.23 ± 0.06a |
| nirk | 7.74 ± 0.05a | 7.60 ± 0.08a | 7.52 ± 0.02a |
| nosZ | 8.21 ± 0.05a | 8.09 ± 0.09a | 7.94 ± 0.05a |
OM: soil organic matter, TN: total soil N, Mean ± SE, Numbers with different letters for each soil variable within rows are significantly different (P<0.05).
Table 2.
Mean soil bulk densities of the cropping systems during 2017-2018 .
| TF | WS | WGS | |
| n | 24 | 22 | 24 |
| -------------------------------- g cm-3 -------------------------------- | |||
| Bulk Density | 1.22± 0.02a | 1.34± 0.02b | 1.39± 0.02b |
Mean ± SE, Numbers with different letters within the row are significantly different (P < 0.05).
Table 4.
Mean bacterial gene copy numbers of cropping system treatments.
| TF | WS | WGS | |
| n | 12 | 11 | 12 |
| 16S rRNA | 9.87 ± 0.06a | 9.78 ± 0.06ab | 9.63 ± 0.06b |
| amoA | 7.01 ± 0.1a | 6.49 ± 0.1b | 6.42 ± 0.1b |
| narG | 7.25 ± 0.06a | 7.17 ± 0.06a | 6.88 ± 0.06b |
| nirK | 7.52 ± 0.04a | 7.35 ± 0.04b | 7.25 ± 0.04b |
| nosZ | 7.98 ± 0.05a | 7.89 ± 0.05a | 7.82 ± 0.05a |
Mean ± SE, Numbers with different letters for each bacterial gene within rows are significantly different (P<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



