
Submitted:
27 April 2023
Posted:
28 April 2023
You are already at the latest version
Abstract
The presence of Staphylococcus aureus in six dry-cured meat-processing facilities was investigated. S. aureus was detected in 3.8% of surfaces from five facilities. Prevalence was clearly higher during processing (4.8%) than after cleaning and disinfection (1.4%). Thirty eight isolates were typified by PFGE and MLST. Eleven sequence types (STs) were defined by MLST. ST30 (32%) and ST12 (24%), were the most abundant. Enterotoxin genes were detected in 53% of isolates. The enterotoxin A gene (sea) was present in all ST30 isolates, seb in one ST1 isolate and sec in two ST45 isolates. Sixteen isolates harbored the enterotoxin gene cluster (egc) with four variations in the sequence. The toxic shock syndrome toxin gene (tst) was detected in 82% of isolates. Regarding the antimicrobial resistance, twelve strains were susceptible to all the antibiotics tested (31.6%). However, 15.8% were resistant to three or more antimicrobials, and therefore multidrug-resistant. Our results showed that, in general, efficient cleaning and disinfection procedures were applied. Nonetheless, the presence of S. aureus with virulence determinants and resistance to antimicrobials, and particularly multidrug resistant MRSA ST398 strains might represent a potential health hazard for consumers, even though the pathogen was not isolated from final products.
Keywords:
MRSA
; enterotoxin
; virulence
; antibiotic resistance
1. Introduction
Staphylococcus aureus is one of the most common foodborne pathogens causing intoxication. Staphylococci can be introduced in the environment of food processing installations through various routes, as raw materials, food handlers or poor hygiene in food processing equipment. They are resistant to desiccation and can survive on different surfaces and resist sanitation forming biofilms [1]. S. aureus can also contaminate foods during preparation and processing, and temperature abuse conditions during transport and/or storage can allow bacterial growth and enterotoxin production. S. aureus is able to tolerate pH ranges from 4.5 to 9.0 and NaCl concentrations up to 9% and can grow and express virulence in a wide range of environmental conditions [2]. The pathogen causes food poisoning through the ingestion of heat stable staphylococcal enterotoxins (SE) preformed in food. Meat and meat products, milk and dairy products, bakery products, salads, etc., are commonly involved in staphylococcal food poisoning (SFP). Bacterial toxins represented the second most common causative agent of outbreaks in the European Union (EU) with 19.3% foodborne outbreaks in 2019 [3].
Staphylococcal enterotoxins are the main virulence factors associated with S. aureus and the primary cause of staphylococcal food poisoning. Most of the staphylococcal food poisoning outbreaks are classified as weak-evidence outbreaks, as only the classical enterotoxins SEA, SEB, SEC, SED and SEE can be detected commercially [4]. Together with these five SEs, new described enterotoxins and staphylococcal enterotoxin-like proteins have been characterized [5,6]. A high number of strains harbor the enterotoxin gene cluster (egc) [7], containing newer enterotoxin genes (seg, sei, sem, sen, seo and seu), and widely distributed in S. aureus isolated from foods and food handlers [8]. Genes encoding SE are located on mobile elements as plasmids, bacteriophages and pathogenicity islands, representing an additional risk factor in food intoxications due to the posible horizontal gene transfer. Additionally, S. aureus strains usually carry more than one SE gene [9]. Besides enterotoxins, S. aureus produce other virulence factors such as exfoliative toxins, toxic shock syndrome toxin, or the Panton-Valentine leukocidin (PVL) [10]. Staphylococcal enterotoxin A accounts for 80% of reported SFP cases, followed by enterotoxin B. Its worldwide predominance has been extensively documented [11].
Multidrug resistant strains have been found in SFP cases and isolated from foods. Multiple antibiotic-resistant strains of S. aureus are spreading rapidly around the world, which raises serious health concerns [12]. Methicillin resistant S. aureus (MRSA) is a major nosocomial emerging pathogen with increasing concern on the livestock industry. Livestock associated MRSA (LA-MRSA) are common colonizers of swine, and could be transmitted from production animals to humans [13]. MRSA ST398, the most prevalent lineage in Europe [14], has been increasingly isolated from meat and dairy products [15,16,17].
The incidence of S. aureus in the food processing environment and the characterization of the isolated strains will provide useful information in the control of SFP and contribute to improve strategies to eliminate the pathogen. The aim of this work was to investigate the prevalence of S. aureus from the environment and different products in six dry-cured meat processing facilities. Further, toxigenicity and antimicrobial resistance of isolated S. aureus was examined in order to evaluate the potential risk associated with the presence of this pathogen.
2. Materials and Methods
2.1. Sampling Procedure and Bacterial Isolation
A total of 720 samples from the environment and equipment surfaces and 82 from different product categories (ingredients, casings, meat batters and final products) were collected in six production facilities of dry-cured pork meat products (ham and traditional Spanish sausages) during a six month period. Environmental and equipment surfaces were sampled during processing (DP) and after cleaning and disinfection (ACD) which takes part before the beginning of the working day. Non-contact and food-contact surfaces were taken by means of pre-moistened sterile wipes (bioMérieux España SAU, Madrid, Spain) and 25 g samples were taken from products for analysis. Samples were kept at 4 ºC immediately after collection and analyzed within 24 h. Facilities were sampled twice at an interval of approximately 6 months.
Samples were homogenized in 0.1% peptone water, maintained for 1 h at 25 ºC and inoculated on Baird-Parker agar supplemented with tellurite egg yolk emulsion (Laboratorios Conda S.A., Madrid, Spain) and CHROMagar Staph aureus (Scharlab, Barcelona, Spain). The plates were incubated at 37 ºC for 24-48 h. Characteristic colonies were isolated and transferred to Brain Heart Infusion (Laboratorios Conda S.A.) for further identification. Coagulase test was carried out using rabbit plasma with EDTA (Biomerieux, France). S. aureus CECT976 (ATCC13565) and Staphylococcus epidermidis CECT231 were used as positive and negative control, respectively. Positive isolates were stored at -80 ºC in TSB supplemented with glycerol (30% v/v) until further analysis.
2.2. Genomic DNA Extraction and PCRs
A total of 124 coagulase positive isolates were selected for further identification. Genomic DNA from overnight cultures in BHI broth was extracted with the genomic DNA GeneJET PCR Purification Kit (Thermo Fisher Scientific, Waltham, MA, USA). The extracted DNA was quantified using Nanodrop, adjusted at 250 ng/μL and stored at -20 ºC.
Each PCR reaction mixture (20 μL) consisted of 2 μL of extracted bacterial DNA template, 10 μL of DNA AmpliTools Master Mix (2X) (Biotools, B & M Labs, S.A., Madrid, Spain), 0.8 μL of 5 mM of each primer (forward and reverse) and 6.4 μL of RNase/DNase-free water (Thermo Fisher Scientific). PCR amplifications were performed using a Mastercycler nexus gradient (Eppendorf, Hamburg, Germany). The amplification conditions were as follows: (1) initial denaturation at 95 ºC for 10 min, (2) 30-35 cycles of denaturation at 95 ºC for 30 s, with annealing temperature and time shown in Table S1, extension at 72 ºC for a variable time depending on the length of the amplicons, and (3) a final extension step at 72 ºC for 10 min. Amplified DNA fragments were separated by agarose gel electrophoresis in 1X TAE buffer stained with GelRed 1X solution. All primers in this study are listed in Table S1.
2.3. Pulsed-Field Gel Electrophoresis (PFGE) Typing
PFGE typification of the S. aureus isolates was determined following the Pulsenet protocol (https://www.cdc.gov/mrsa/pdf/ar_mras_PFGE_s_aureus.pdf). Digestion of the genomic DNA was performed with SmaI FastDigest (Thermo Fisher Scientific). XmaI (New England Biolabs Inc., Ipswich, MA, USA), an isoschizomer of SmaI not blocked by CpG methylation, was used for the digestion of genomic DNA of MRSA isolates. Salmonella ser. Braenderup H9812 digested with XbaI FastDigest (Thermo Fisher Scientific) was used as molecular size marker and included in every gel for standardization and comparison purposes. The restriction DNA fragments were separated using the polygonal contour clamped homogeneous electric field system CHEF DRII (Bio-Rad Laboratories, Hercules, CA, USA). Analysis of the PFGE patterns was performed using the BioNumerics software (Applied Maths NV, Sint-Martens-Latem, Belgium). Comparisons were performed using Dice similarity coefficient (Similarity of 1% and optimization of 1%). Dendrograms were constructed with the UPGMA algorithm.
2.4. Multilocus Sequence Typing (MLST)
MLST analysis was carried out as previously described [18]. In summary, fragments of seven housekeeping genes: arc, aroE, glpF, gmk, pta, tpi and yqil (Table S1) were amplified following the protocol accessible at https://pubmlst.org/organisms/staphylococcus-aureus/primers. PCR products were purified with GeneJet PCR Purification Kit (Thermo Scientific) following manufacturer’s specifications and sequenced by the Sanger Sequencing Service (Complutense University of Madrid, Spain). Clean sequences were queried in the database and corresponding allele numbers were assigned. The combination of seven alleles gave the Sequence Type (ST) for each isolate. New alleles or STs were assigned when necessary by the international database of MLST for S. aureus. Phylogenetic analysis of the different STs was performed using eBURST algorithm included in the software Phyloviz (http://www.phyloviz.net) and visualized in a minimum spanning tree.
2.5. Molecular Detection of Virulence Genes
S. aureus isolates were tested for the presence of enterotoxin genes (sea, seb, sec, sed, see, seg, seh, sei, sej, sek, sem, sen, seo, sep, seq, ser, seu, sev and sew). Other virulence factors investigated were leukocidin genes (lukS/F-PV), the toxic shock syndrome toxin gene (tst) and the biofilm associated gen icaA. Additionally, the presence of non-virulence penicillin resistance, β-lactamase gene (blaZ) was also investigated by PCR. All cleaned amplicons were sequenced by the Sanger Sequencing Service (Complutense University of Madrid, Spain). The enterotoxic gene cluster (egc) was completely amplified and sequenced using a new set of designed primers listed in Table S1.
2.6. Enterotoxins A-D Production
The production of classical enterotoxins SEA, SEB, SEC, and SED during the growth of S. aureus strains was assessed by reversed passive latex agglutination using the SET-RPLA Kit (Thermo Scientific™ Oxoid™ SET-RPLA Toxin Detection Kit, Thermo Fisher Scientific) according to the manufacturer’s instructions.
2.7. Antibiotic Susceptibility
Confirmed S. aureus isolates were tested for antibiotic resistance by the disc diffusion method (CLSI) using Mueller-Hinton agar (Laboratorios Conda S.A.) and commercially available Thermo Scientific™ Oxoid™ antimicrobial susceptibility discs (Thermo Fisher Scientific). The antimicrobials studied were penicillin G (PEN), ampicillin (AMP), amoxicillin with clavulanic acid (AMC), oxacillin (OXA), cefoxitin (FOX), clindamycin (CLD), erythromycin (ERY), chloramphenicol (CLF), tetracycline (TET), ciprofloxacin (CIP), gentamicin (GEN), vancomycin (VAN), and trimethoprim plus sulphamethoxazole (STX). S. aureus ATCC 25923 was used as reference strain for antimicrobial testing. The plates with antibiotic discs were incubated at 37 ºC for 18 h. Diameters of the inhibition zones were measured and the S. aureus strains were scored as susceptible, intermediate, or resistant according to the criteria of Clinical Laboratory Standards Institute [19].
3. Results
3.1. Prevalence of S. aureus in Dry-Cured Meat Products Producing Facilities
The presence of S. aureus was investigated in a total of 720 samples from the environment and equipment and 82 from different product categories in six dry-cured meat products processing facilities. S. aureus was found in five of the six sampled facilities. A total of 124 coagulase positive staphylococcal isolates confirmed as S. aureus were obtained from 38 positive samples. S. aureus was recovered (Table 1) from surfaces in five out of the six processing facilities with an overall incidence of 3.8%. Prevalence during processing was 4.8%, whereas diminished to 1.4% after cleaning and disinfection procedures. Food contact surfaces contamination was of 4.4%, slightly higher than 3.1% of positive non-food contact surfaces. Prevalence was higher in different product categories (13.4%) in 3 out the 4 facilities investigated, in batter and casings samples, although the pathogen was not detected in final products (Table 1). Isolation frequency varied among plants, between 8.0% (plant C) and 2.0% (plant E). Ten isolates (8%), harboring the mecA gene were adscribed to MRSA and recovered in four positive samples from plant A.Bulleted lists look like this:
3.2. Characterization of S. aureus Isolates
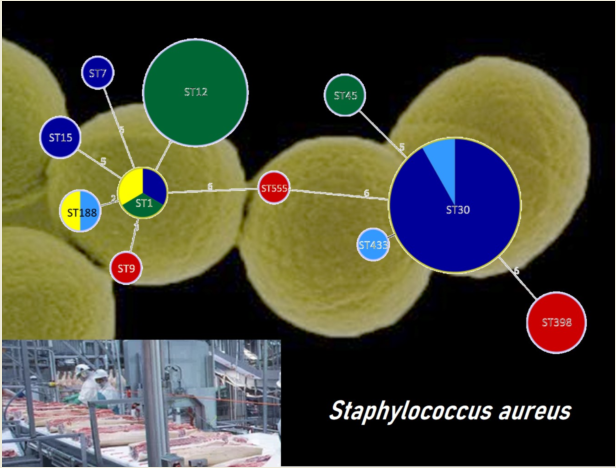
Results on PFGE characterization and the origin of 34 non-MRSA isolates from positive samples are shown in Figure 1. PFGE with SmaI revealed a total of 14 pulsotypes (PTs) showing high diversity. Two different pulsotypes were found in plant A (4% positive samples), PT2A during processing and PT11A in a drain in clean and disinfected installation. Plant B (6.8% positive samples) presented 4 different pulsotypes, with PT12B in 11 out of 15 positive samples. This PT was found in batter, casings and contact and non-contact surfaces during processing. Higher variability was detected in plant C (8% positive samples), with a majority of PT8C and PT9C in nine out of 12 positive samples, in batter, casings and contact and non-contact surfaces during processing. Positive samples from plant D (2.6%) presented three different pulsotypes, and two pulsotypes were found in plant E (2%). Only one common PT (3C and 3E) was detected at dry-cured ham deboning zones of two different processing facilities. The rest, as seen in Figure 1, were specific of each plant. MRSA isolates (all from plant A) were obtained from meat batter and contact surfaces during processing. PFGE typification of these isolates with XmaI digestion revealed a high degree of similarity (results not shown). Allelic profiles obtained by MLST allowed the definition of 11 different sequence types (STs) which were assigned to six clonal complexes (CCs) (Table 2). All MRSA isolates were adscribed to ST398. The most abundant ST was ST30 (32% of isolates), detected in plants B and D, followed by ST12 (24%), isolated from plant C and ST1 (8%) characterized in plants B, C and E. ST188 (5%) was detected in two plants whereas ST7, ST9, ST15, ST45, and ST433 were less abundant and only found in one plant. One isolate from a drain in plan A showed a new sequence type ST5554. ST398 (11%) was only detected in plant A. Isolates were distributed into seven CCs, CC1 (ST1, ST188, and ST9), CC7, CC12, CC15, CC30 (ST30 and ST433), CC45 and CC398. The phylogenetic relationships among the different populations, defined by MLST, are shown in Figure 2.
3.3. Detection of Virulence Genes
The frequence of virulence genes identified on the 38 isolates selected from the positive samples is shown in Table 2. SE genes were detected in 20 out the 38 strains investigated. Enterotoxin A gene sea was identified in all ST30 isolates from plant B and plant D, and one ST188 from plant D, seb in one ST1 isolate, sec in two ST45 isolates from plant C while sed and see were not amplified by PCR in any of the S. aureus investigated. Three ST1 isolates from plants B, C and E were positive for seh genes.
Sixteen out of the 38 isolates possessed the enterotoxin gene cluster (egc), belonging to ST9, ST30, ST45 and ST433. The complete egc was sequenced with the designed primers. Four different egc variants were found (Figure 3): ST433 and ST30 belonged to egc3 [11] or OMIUNG according to other classification [20]. The isolate D41 (ST30) possessed the egc3 but also two amino acid substitutions in SEO (D49E) and SEG (V230F) regarding deposited sequences in Genbank. ST9 isolate presented an egc1 type backbone, with the selw gene instead seu, which corresponds to an OMIWNG type. Amino acid substitutions were also found in SEO (E87K), SEN (K143R) and SEG (G131D), based on the amino acid sequences deposited in Genbank. The two ST45 isolates were similar to ST9, with an egc1 backbone, containing a selw gene (OMIWNG type) and amino acid substitutions in SEM (T63S, V64I and R115C) and SEI (Y103F). Genetic determinants of enterotoxin production were not detected in all ST398, ST12, ST15, ST5554, ST7 and one ST188 isolates. The presence of the tst gene was high, with 82% of isolates positive. All ST398, ST7, ST433 and one ST1 did not carry this gene. Sequencing of PCR product obtained with Panton-Valentine leukocidin primers for genes lukF and lukS revealed a sequence compatible with leukocidin ED in five out nine ST12 isolates from plant C. The icaA gene related with biofilm forming ability was present in all S. aureus isolates. On the other hand, staphylococcal enterotoxin A was produced by 13 isolates, 12 belonging to ST30 from plants C and D and one ST188 isolate from plant D. Only one isolate ST1 from plant B produced SEB and the two ST45 isolates from plant C produced SEC. None of the isolates tested produced SED.
3.4. Antimicrobial Resistance Profiles
The antibiotic susceptibility results are shown in Table 2. Twelve out of the 38 isolates (31.6 %) were susceptible to all the 13 antimicrobials tested. They were further typified as ST7, ST12, ST433 and ST5554. Three ST398-meticillin resistant (MRSA) isolates were also resistant to three or more antimicrobials and considered therefore multidrug resistant (MDR). Twenty-six isolates were resistant to penicillin (ST1, ST9, ST15, ST30, ST45, ST188 and ST398), seven to tetracycline (ST398, ST9 and ST15), four to erythromycin (ST398 and ST1), three to cefoxitin (ST398) and one to ciprofloxacin (ST398). Resistance to ampicilin, amoxicillin plus clavulanic acid, chloramphenicol, gentamycin, oxacillin and vancomycin was not observed by phenotypic analysis. The presence of blaZ gene encoding for β-lactamase was detected in all the isolates resistant to penicillin.
4. Discussion
S. aureus is frequently isolated from meat processing facilities, from contact and non-contact surfaces and from raw materials and different product categories. Contamination of meat products results from poor hygienic practices during processing and storage. In this study, overall prevalence of S. aureus in the environment and different categories of products from five out of six dry-cured meats processing facilities was low (4.7%). Prevalence was higher during processing than after cleaning and disinfection, although the pathogen was detected in clean surfaces (1.4%). S. aureus contamination was higher in batter and casings (13.4%) than in equipment surfaces, whereas the pathogen was not detected in final products.
Our prevalence results are in concordance with previous studies conducted in Spain, with a 3.2% in disinfected surfaces from different meat industries [21]. Higher incidence was observed in a cutting room with coagulase-positive S. aureus in 15.5% of equipment samples during cutting operation, and 31.8% of meat samples for dry-cured sausages [22]. Incidence reported by Gounadaki et al. [23] in food contact and non-contact surfaces of three of seven processing plants producing traditional fermented and/or dry sausages was 11.7%, whereas the pathogen was not detected in batter or final products. According to several authors [24,25], raw materials or ingredients are one of the main sources of S. aureus contamination in meat processing plants, data in agreement with our results on higher contamination in batters and casings. In our work, MRSA presence was relatively high (13.2%), although the pathogen was detected only in one of the industries investigated.
In general, MRSA contamination in meat is lower in Europe (3.2%) compared to other continents [26]. MRSA was detected in a 5% of RTE food samples positive for S. aureus [16], a value higher that the 1.3% in retail foods reported by Yang et al. [27]. Average incidence of S. aureus in retail meat including pork in China was 35% [17], similar to pork products in Spain (33.6%), with a high rate of MRSA found in 21.8% of samples, mainly in meat products with skin (ears and snout) [13].
The S. aureus isolates from the processing facilities investigated presented, in general, a high diversity of genotypes by PFGE. A large diversity has been already reported for S. aureus from the environment and food [28], and from clinical isolates [29]. In general, an association of pulsotypes (or clusters of pulsotypes) with the production plant was observed. For example, PT8C and PT9C with more than 90% similarity were isolated only in plant C.
MLST allowed the definition of 11 different sequence types (STs). As shown in Figure 2, the most abundant was ST30 (CC30) (32% of isolates), followed by ST12 (CC12) (24%). Most of the STs detected in our study have been previously characterized in in pork products from other countries: ST398 (35%), ST1, ST30, ST45, ST15 and ST9 in Denmark [30], and ST1 was predominant in samples from pork meat in US [31]. CC45 and CC1 predominated among MSSA isolates from pork meat samples [13]. On the other hand, MRSA ST398 is the major sequence type colonizing pigs in Europe [32], and predominated in pork meat samples in Spain [13]. In the present work, MRSA ST398 was not the dominant ST, but it was the majority in facility A. Different studies have found that S. aureus CC30 is predominant among human nasal carriers of the bacterium [33], and in cases of bacteremia in Denmark [30]. Although it has also been isolated from RTE food [4], it is not one of the major clonal complexes of porcine origin. We cannot rule out a human origin of those isolates.
The combination of PFGE and MLST revealed association between PTs and CCs with processing plants. Thus, PT1 belonged to CC45, only in plant C, PT10 with CC15 in plant B, PT12 with CC30 and plant B, among others. Taking together PFGE and MLST, we might ascertain that, in general, 1-3 clones are isolated at each plant.
Regarding the presence of enterotoxin genes, in the present work, 55% of S. aureus isolates carried one or more SE genes. Higher percentages (66%) were reported from industry surfaces [21], different food products (69%) [34], and from fermented pork sausages (60%) [35]. According to our results, SE genes were not detected in all ST398 and ST12 strains. SEA is the enterotoxin most frequently involved in SFP cases [11], while a lower number of cases are attributed to SEB, SEC, and SED. In our study, all the strains harboring classical SE genes (sea, seb and sec) effectively produced the corresponding toxin (SEA, SEB and SEC) as detected by agglutination test. This data confirms the virulence of these strains. Among the classic enterotoxins, SEB and SEE have been associated with infectious strains of bovine origin, mainly of ST188 [36,37]. Therefore it can be expected a low prevalence of those toxins amongst our isolates. In fact, only one ST1 SEB-producing isolate was characterized. Moreover, SED and SEE toxins or their genes were not detected.
The enterotoxin gene cluster egc (seg, sei, sem, sen, seo, seu/sew) was found in 37% of the isolates from CC30, CC45 and one ST9 isolate. This percentage is higher than previously reported from different sources, as 14.1% (food strains) [38] or 18.7% (RTE foods) [39]. However, higher percentages (50-70%) of egc in samples from healthy human carriers has been previously reported [40]. According to our results, the egc cluster was, in most cases, associated with classical SE genes, sea or sec, as previously observed [41,42]. Genes from egc as sei or seg have been linked to outbreaks [41]. Moreover, Schwendimann et al. [4] demonstrated that 75% egc positive strains expressed SEG and 100% SEI, indicating that these egc enterotoxins are involved in SFP. Therefore, these isolates are potentially pathogenic. Dicks et al. [20] found association between the egc type and clonal complexes of S. aureus. Thus, OMIWNG variant was present in CC1, CC5 and CC22. In our study, this variant was found in ST9, which belongs to CC1. We also found OMIWNG variant in ST45 (CC45), not described in the mentioned paper. The investigation of a higher number of strains worldwide could ascertain the relationship between egc variants and clonal complexes of S. aureus. Recombination between genes inside egc have been described (i.e. sel33 is a recombination between sew and sen [20]. Also, incomplete egc variants lacking any of the genes have been found. Thus, Song et al. [37] reported a 39.5% of egc strains lacking seu. In another study, most of the isolates of swine origin possessed an incomplete egc, lacking two of the genes [43]. Although our isolates possessed a complete egc, the absence of genes in egc regarding the pathogenicity of the strains needs to be further investigated.
Variations in the amino acid sequence of seo, sei, sem, sen and seg might constitute new genes. For example, a variant of seu firstly named seu2 is now considered sew.
The seh encoding SEH toxin was found in three (7.8%) CC1 isolates from three different industries. The seh gene seems to be restricted to CC1 isolates [29], and has been found in S. aureus from RTE foods [39,44]. SEH has been reported as cause of SFP cases, highlighting the importance of the detection of seh gene in foodborne outbreaks.
The toxin shock staphylococcal toxin gene tst, was found at high prevalence in the present study (63.2%). Xie et al. [29] observed the presence of tst gene in 48% clinical isolates from China, and by Argudín et al. [8] in 25.8% of isolates from food and food handlers in Spain. On the contrary, lower prevalence rates have been reported, with values of 2.1% [39], 7.2% [44] or 17% [16]. In another study, the detection rate of tst was high in MRSA ST9 strains from swine and human clinical isolates [45]. In our study, the presence of tst gene was detected in almost all CCs, not restricted to any specific ST. This high proportion of isolates expressing tst will need to be further investigated.
Concerning Panton-Valentine leukocidin, with the published primers [46], a length compatible amplicon (180 bp) was not obtained. However, the inespecific amplicons were sequenced, and a sequence compatible with lukED was observed in five ST12 isolates. This is probably a consequence of the similarity of the different leucocidin genes. In contrast, previous studies conducted in Spain have found high proportions of PVL, both in clinical and food isolates [8,47].
The intercellular gene cluster adhesion (ica) operon is one of the main factors involved in biofilm production by S. aureus [48]. Biofilm formation is a well-known mechanism for survival to disinfectants in the food industry [49]. The ica operon plays an important role in biofilm formation, specially through the exposition to NaCl [50], used as ingredient and preservative in dry-cured meat products. High salt concentrations might select the isolates with presence and activity of ica operon.
Notably, all MRSA ST398 isolates were negative for all tested enterotoxin, tst and pvl genes, in agreement with other MRSA results from slaughter pigs [51]. In the scientific literature, the detection of toxin encoding genes in MRSA CC398 is low, although they have been found colonizing or causing infections in humans [52,53,54].
The ability of S. aureus to acquire and develop resistance to multiple antibiotics that can be transmitted to humans by ingestion of contaminated food products is recognized worldwide. In the present study, the percentage of S. aureus resistant to antibiotics was high (71%), although higher percentages in meat have been recorded [17]. Values of resistance to three or more antibiotics (15.8%) were similar to 16.7% reported for S. aureus from RTE foods [40]. All ST12, ST5554, ST7 and ST433 isolates were susceptible to all tested antibiotics. Higher values were reported by Gutierrez et al. [21], with 70% of strains from food industry surfaces susceptible to 10 antibiotics tested. In our study, resistance to penicillin was observed in 68% of the isolates. Similar percentages have been reported in S. aureus from food or associated to food poisoning [34,42]. The presence of MDR strains is common among S. aureus isolates from meat and poultry samples [13]. Multiresistance to several classes of antimicrobial agents is also common in MRSA ST398 isolates and has been reported worldwide [55,56,57]. In our study, MRSA ST398 strains were resistant to two or more antibiotics, and only MRSA isolates were resistant to cefoxitin and ciprofloxacin. MRSA isolates from the present work showed also resistance to tetracycline, a common trait in S. aureus of animal origin [58]. This resistance seems to be acquired by livestock associated (LA)-MRSA CC398 after the introduction in livestock from human MSSA [59]. Presence of MDR is a matter of concern for the food industry, although in our study they were not detected in clean surfaces or final products.
In the present work, the pathogen was detected in a low number of samples after cleaning and disinfection. Our results showed a high variability in the environment, but in general, the cleaning and disinfection procedures were efficient. The highest contamination was recorded on meat batters that could contaminate surfaces during processing. Some points are critical for S. aureus presence and this knowledge is important for the improvement of hygiene control procedures. The presence of S. aureus with virulence determinants and resistance to antimicrobials represent a potential health hazard for consumers. In addition, multidrug resistant MRSA ST398 strains increases the risk for the spread of this pathogen.
5. Conclusions
S. aureus was found at low prevalence in the six pork meat industries sampled in this work. The isolates showed a wide genetic diversity, although some populations were detected in more than one processing plant. There was a clear reduction of S. aureus after the cleaning and disinfection procedures, observing a very low prevalence in clean surfaces, and no detection of the pathogen in final products. It is remarkable the appearance of MRSA isolates in one of the industries. The presence S. aureus with genetic determinants of enterotoxin production must be taken into account as a potential risk factor for food safety.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: List of primers used in this study.
Author Contributions
D.P.-B., M.D., A.G.-L., D.B., A.P.-B., P.G., M.M. and J.L.A. have made a substantial, direct, and intellectual contribution to the work, and approved it for publication. All authors have read and agreed to the published version of the manuscript.
Funding
This work has received financial support from projects RTA2017-00027-C03-01 and AT2016-003 (Spanish Ministry of Science and Innovation).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
FPI-SGIT2015-06 grant to A. Pérez-Baltar.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moretro, T.; Langsrud, S. Residential Bacteria on Surfaces in the Food Industry and Their Implications for Food Safety and Quality. Compr Rev Food Sci Food Saf 2017, 16, 1022–1041. [Google Scholar] [CrossRef]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet Mol Res 2003, 2, 63–76. [Google Scholar] [PubMed]
- EFSA. The European Union One Health 2019 Zoonoses Report 1831-4732; 2021; p. e06406. [Google Scholar]
- Schwendimann, L.; Merda, D.; Berger, T.; Denayer, S.; Feraudet-Tarisse, C.; Klaui, A.J.; Messio, S.; Mistou, M.Y.; Nia, Y.; Hennekinne, J.A.; et al. Staphylococcal Enterotoxin Gene Cluster: Prediction of Enterotoxin (SEG and SEI) Production and of the Source of Food Poisoning on the Basis of vSabeta Typing. Appl Environ Microbiol 2021, 87, e0266220. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, Y.; Cao, Y.; Yan, W.; Niu, X.; Zhou, L.; Chen, J.; Sun, Y.; Li, C.; Zhang, X.; et al. The Distribution of 18 Enterotoxin and Enterotoxin-Like Genes in Staphylococcus aureus Strains from Different Sources in East China. Foodborne Pathog Dis 2016, 13, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Miao, X.; Zhou, L.; Cui, B.; Zhang, J.; Xu, X.; Wu, C.; Peng, X.; Wang, X. Characterization of Oxacillin-Susceptible mecA-Positive Staphylococcus aureus from Food Poisoning Outbreaks and Retail Foods in China. Foodborne Pathog Dis 2020, 17, 728–734. [Google Scholar] [CrossRef]
- Jarraud, S.; Peyrat, M.A.; Lim, A.; Tristan, A.; Bes, M.; Mougel, C.; Etienne, J.; Vandenesch, F.; Bonneville, M.; Lina, G. egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J Immunol 2001, 166, 669–677. [Google Scholar] [CrossRef]
- Argudin, M.A.; Mendoza, M.C.; Gonzalez-Hevia, M.A.; Bances, M.; Guerra, B.; Rodicio, M.R. Genotypes, exotoxin gene content, and antimicrobial resistance of Staphylococcus aureus strains recovered from foods and food handlers. Appl Environ Microbiol 2012, 78, 2930–2935. [Google Scholar] [CrossRef]
- Rajkovic, A.; Jovanovic, J.; Monteiro, S.; Decleer, M.; Andjelkovic, M.; Foubert, A.; Beloglazova, N.; Tsilla, V.; Sas, B.; Madder, A.; et al. Detection of toxins involved in foodborne diseases caused by Gram-positive bacteria. Compr Rev Food Sci Food Saf 2020, 19, 1605–1657. [Google Scholar] [CrossRef]
- Doyle, M.E.; Hartmann, F.A.; Lee Wong, A.C. Methicillin-resistant staphylococci: implications for our food supply? Anim Health Res Rev 2012, 13, 157–180. [Google Scholar] [CrossRef]
- Argudin, M.A.; Mendoza, M.C.; Rodicio, M.R. Food poisoning and Staphylococcus aureus enterotoxins. Toxins (Basel) 2010, 2, 1751–1773. [Google Scholar] [CrossRef]
- Matyi, S.A.; Dupre, J.M.; Johnson, W.L.; Hoyt, P.R.; White, D.G.; Brody, T.; Odenwald, W.F.; Gustafson, J.E. Isolation and characterization of Staphylococcus aureus strains from a Paso del Norte dairy. J Dairy Sci 2013, 96, 3535–3542. [Google Scholar] [CrossRef] [PubMed]
- Mama, O.M.; Morales, L.; Ruiz-Ripa, L.; Zarazaga, M.; Torres, C. High prevalence of multidrug resistant S. aureus-CC398 and frequent detection of enterotoxin genes among non-CC398 S. aureus from pig-derived food in Spain. Int J Food Microbiol 2020, 320, 108510. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Assessment of the Public Health significance of meticillin resistant Staphylococcus aureus (MRSA) in animals and foods. EFSA J 2009, 7, 993. [Google Scholar] [CrossRef]
- de Boer, E.; Zwartkruis-Nahuis, J.T.; Wit, B.; Huijsdens, X.W.; de Neeling, A.J.; Bosch, T.; van Oosterom, R.A.; Vila, A.; Heuvelink, A.E. Prevalence of methicillin-resistant Staphylococcus aureus in meat. Int J Food Microbiol 2009, 134, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Parveen, S.; Rahman, M.; Huq, M.; Nabi, A.; Khan, Z.U.M.; Ahmed, N.; Wagenaar, J.A. Occurrence and Characterization of Methicillin Resistant Staphylococcus aureus in Processed Raw Foods and Ready-to-Eat Foods in an Urban Setting of a Developing Country. Front Microbiol 2019, 10, 503. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Wu, Q.; Zhang, J.; Zhang, F.; Yang, X.; Wu, H.; Zeng, H.; Chen, M.; Ding, Y.; et al. Staphylococcus aureus Isolated From Retail Meat and Meat Products in China: Incidence, Antibiotic Resistance and Genetic Diversity. Front Microbiol 2018, 9, 2767. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol 2000, 38, 1008–1015. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing: 29th Edition. Supplement M100 In CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, 2019. [Google Scholar]
- Dicks, J.; Turnbull, J.D.; Russell, J.; Parkhill, J.; Alexander, S. Genome Sequencing of a Historic Staphylococcus aureus Collection Reveals New Enterotoxin Genes and Sheds Light on the Evolution and Genomic Organization of This Key Virulence Gene Family. J Bacteriol 2021, 203. [Google Scholar] [CrossRef]
- Gutierrez, D.; Delgado, S.; Vazquez-Sanchez, D.; Martinez, B.; Cabo, M.L.; Rodriguez, A.; Herrera, J.J.; Garcia, P. Incidence of Staphylococcus aureus and analysis of associated bacterial communities on food industry surfaces. Appl Environ Microbiol 2012, 78, 8547–8554. [Google Scholar] [CrossRef]
- Pala, T.R.; Sevilla, A. Microbial contamination of carcasses, meat, and equipment from an Iberian pork cutting plant. J Food Prot 2004, 67, 1624–1629. [Google Scholar] [CrossRef]
- Gounadaki, A.S.; Skandamis, P.N.; Drosinos, E.H.; Nychas, G.J. Microbial ecology of food contact surfaces and products of small-scale facilities producing traditional sausages. Food Microbiol 2008, 25, 313–323. [Google Scholar] [CrossRef]
- Gelbicova, T.; Brodikova, K.; Karpiskova, R. Livestock-associated methicillin-resistant Staphylococcus aureus in Czech retailed ready-to-eat meat products. Int J Food Microbiol 2022, 374, 109727. [Google Scholar] [CrossRef]
- Korenova, J.; Reskova, Z.; Veghova, A.; Kuchta, T. Tracing Staphylococcus aureus in small and medium-sized food-processing factories on the basis of molecular sub-species typing. Int J Environ Health Res 2015, 25, 384–392. [Google Scholar] [CrossRef]
- Ou, Q.; Peng, Y.; Lin, D.; Bai, C.; Zhang, T.; Lin, J.; Ye, X.; Yao, Z. A Meta-Analysis of the Global Prevalence Rates of Staphylococcus aureus and Methicillin-Resistant S. aureus Contamination of Different Raw Meat Products. J Food Prot 2017, 763–774. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, J.; Yu, S.; Wu, Q.; Guo, W.; Huang, J.; Cai, S. Prevalence of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus in Retail Ready-to-Eat Foods in China. Front Microbiol 2016, 7, 816. [Google Scholar] [CrossRef]
- Adame-Gomez, R.; Castro-Alarcon, N.; Vences-Velazquez, A.; Toribio-Jimenez, J.; Perez-Valdespino, A.; Leyva-Vazquez, M.A.; Ramirez-Peralta, A. Genetic Diversity and Virulence Factors of S. aureus Isolated from Food, Humans, and Animals. Int J Microbiol 2020, 2020, 1048097. [Google Scholar] [CrossRef]
- Xie, Y.; He, Y.; Gehring, A.; Hu, Y.; Li, Q.; Tu, S.I.; Shi, X. Genotypes and toxin gene profiles of Staphylococcus aureus clinical isolates from China. PLoS One 2011, 6, e28276. [Google Scholar] [CrossRef]
- Li, H.; Andersen, P.S.; Stegger, M.; Sieber, R.N.; Ingmer, H.; Staubrand, N.; Dalsgaard, A.; Leisner, J.J. Antimicrobial Resistance and Virulence Gene Profiles of Methicillin-Resistant and -Susceptible Staphylococcus aureus From Food Products in Denmark. Front Microbiol 2019, 10, 2681. [Google Scholar] [CrossRef]
- Waters, A.E.; Contente-Cuomo, T.; Buchhagen, J.; Liu, C.M.; Watson, L.; Pearce, K.; Foster, J.T.; Bowers, J.; Driebe, E.M.; Engelthaler, D.M.; et al. Multidrug-Resistant Staphylococcus aureus in US Meat and Poultry. Clin Infect Dis 2011, 52, 1227–1230. [Google Scholar] [CrossRef]
- de Neeling, A.J.; van den Broek, M.J.; Spalburg, E.C.; van Santen-Verheuvel, M.G.; Dam-Deisz, W.D.; Boshuizen, H.C.; van de Giessen, A.W.; van Duijkeren, E.; Huijsdens, X.W. High prevalence of methicillin resistant Staphylococcus aureus in pigs. Vet Microbiol 2007, 122, 366–372. [Google Scholar] [CrossRef]
- Wattinger, L.; Stephan, R.; Layer, F.; Johler, S. Comparison of Staphylococcus aureus isolates associated with food intoxication with isolates from human nasal carriers and human infections. Eur J Clin Microbiol Infect Dis 2012, 31, 455–464. [Google Scholar] [CrossRef]
- Pereira, V.; Lopes, C.; Castro, A.; Silva, J.; Gibbs, P.; Teixeira, P. Characterization for enterotoxin production, virulence factors, and antibiotic susceptibility of Staphylococcus aureus isolates from various foods in Portugal. Food Microbiol 2009, 26, 278–282. [Google Scholar] [CrossRef]
- Sankomkai, W.; Boonyanugomol, W.; Kraisriwattana, K.; Nutchanon, J.; Boonsam, K.; Kaewbutra, S.; Wongboot, W. Characterisation of Classical Enterotoxins, Virulence Activity, and Antibiotic Susceptibility of Staphylococcus aureus Isolated from Thai Fermented Pork Sausages, Clinical Samples, and Healthy Carriers in Northeastern Thailand. J Vet Res 2020, 64, 289–297. [Google Scholar] [CrossRef]
- Song, Q.; Zhu, Z.; Chang, Y.; Shen, X.; Gao, H.; Yang, Y. Prevalence and Characteristics of Enterotoxin B-Producing Staphylococcus aureus Isolated from Food Sources: A Particular Cluster of ST188 Strains was Identified. J Food Sci 2016, 81, M715–M718. [Google Scholar] [CrossRef]
- Jung, H.R.; Lee, Y.J. Characterization of Virulence Factors in Enterotoxin-Producing Staphylococcus aureus from Bulk Tank Milk. Animals (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Song, M.; Shi, C.; Xu, X.; Shi, X. Molecular Typing and Virulence Gene Profiles of Enterotoxin Gene Cluster (egc)-Positive Staphylococcus aureus Isolates Obtained from Various Food and Clinical Specimens. Foodborne Pathog Dis 2016, 13, 592–601. [Google Scholar] [CrossRef]
- Mekhloufi, O.A.; Chieffi, D.; Hammoudi, A.; Bensefia, S.A.; Fanelli, F.; Fusco, V. Prevalence, Enterotoxigenic Potential and Antimicrobial Resistance of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Algerian Ready to Eat Foods. Toxins (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Chen, Q.; Xie, S. Genotypes, Enterotoxin Gene Profiles, and Antimicrobial Resistance of Staphylococcus aureus Associated with Foodborne Outbreaks in Hangzhou, China. Toxins (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Johler, S.; Giannini, P.; Jermini, M.; Hummerjohann, J.; Baumgartner, A.; Stephan, R. Further evidence for staphylococcal food poisoning outbreaks caused by egc-encoded enterotoxins. Toxins (Basel) 2015, 7, 997–1004. [Google Scholar] [CrossRef]
- Kerouanton, A.; Hennekinne, J.A.; Letertre, C.; Petit, L.; Chesneau, O.; Brisabois, A.; De Buyser, M.L. Characterization of Staphylococcus aureus strains associated with food poisoning outbreaks in France. Int J Food Microbiol 2007, 115, 369–375. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Cai, R.; Shi, L.; Li, C.; Yan, H. Prevalence of Enterotoxin Genes in Staphylococcus aureus Isolates from Pork Production. Foodborne Pathog Dis 2018, 15, 437–443. [Google Scholar] [CrossRef]
- Yang, X.; Yu, S.; Wu, Q.; Zhang, J.; Wu, S.; Rong, D. Multilocus Sequence Typing and Virulence-Associated Gene Profile Analysis of Staphylococcus aureus Isolates From Retail Ready-to-Eat Food in China. Front Microbiol 2018, 9, 197. [Google Scholar] [CrossRef]
- Wan, M.T.; Lauderdale, T.L.; Chou, C.C. Characteristics and virulence factors of livestock associated ST9 methicillin-resistant Staphylococcus aureus with a novel recombinant staphylocoagulase type. Vet Microbiol 2013, 162, 779–784. [Google Scholar] [CrossRef]
- Lina, G.; Piemont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin Infect Dis 1999, 29, 1128–1132. [Google Scholar] [CrossRef]
- Argudin, M.A.; Mendoza, M.C.; Mendez, F.J.; Martin, M.C.; Guerra, B.; Rodicio, M.R. Clonal complexes and diversity of exotoxin gene profiles in methicillin-resistant and methicillin-susceptible Staphylococcus aureus isolates from patients in a Spanish hospital. J Clin Microbiol 2009, 47, 2097–2105. [Google Scholar] [CrossRef]
- Rohde, H.; Knobloch, J.K.; Horstkotte, M.A.; Mack, D. Correlation of Staphylococcus aureus icaADBC genotype and biofilm expression phenotype. J Clin Microbiol 2001, 39, 4595–4596. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial Biofilms in the Food Industry-A Comprehensive Review. Int J Environ Res Public Health 2021, 18. [Google Scholar] [CrossRef]
- Lee, S.; Kim, S.; Lee, H.; Ha, J.; Lee, J.; Choi, Y.; Oh, H.; Yoon, Y.; Choi, K.H. icaA Gene of Staphylococcus aureus Responds to NaCl, Leading to Increased Biofilm Formation. J Food Prot 2018, 81, 412–416. [Google Scholar] [CrossRef]
- Gomez-Sanz, E.; Torres, C.; Lozano, C.; Fernandez-Perez, R.; Aspiroz, C.; Ruiz-Larrea, F.; Zarazaga, M. Detection, molecular characterization, and clonal diversity of methicillin-resistant Staphylococcus aureus CC398 and CC97 in Spanish slaughter pigs of different age groups. Foodborne Pathog Dis 2010, 7, 1269–1277. [Google Scholar] [CrossRef]
- Chen, C.; Wu, F. Livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) colonisation and infection among livestock workers and veterinarians: a systematic review and meta-analysis. Occup Environ Med 2020. [Google Scholar] [CrossRef]
- Cuny, C.; Kock, R.; Witte, W. Livestock associated MRSA (LA-MRSA) and its relevance for humans in Germany. Int J Med Microbiol 2013, 303, 331–337. [Google Scholar] [CrossRef]
- Sergelidis, D.; Angelidis, A.S. Methicillin-resistant Staphylococcus aureus: a controversial food-borne pathogen. Lett Appl Microbiol 2017, 64, 409–418. [Google Scholar] [CrossRef]
- Papadopoulos, P.; Papadopoulos, T.; Angelidis, A.S.; Kotzamanidis, C.; Zdragas, A.; Papa, A.; Filioussis, G.; Sergelidis, D. Prevalence, antimicrobial susceptibility and characterization of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus isolated from dairy industries in north-central and north-eastern Greece. Int J Food Microbiol 2019, 291, 35–41. [Google Scholar] [CrossRef]
- Parisi, A.; Caruso, M.; Normanno, G.; Latorre, L.; Sottili, R.; Miccolupo, A.; Fraccalvieri, R.; Santagada, G. Prevalence, antimicrobial susceptibility and molecular typing of Methicillin-Resistant Staphylococcus aureus (MRSA) in bulk tank milk from southern Italy. Food Microbiol 2016, 58, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Titouche, Y.; Hakem, A.; Houali, K.; Meheut, T.; Vingadassalon, N.; Ruiz-Ripa, L.; Salmi, D.; Chergui, A.; Chenouf, N.; Hennekinne, J.A.; et al. Emergence of methicillin-resistant Staphylococcus aureus (MRSA) ST8 in raw milk and traditional dairy products in the Tizi Ouzou area of Algeria. J Dairy Sci 2019, 102, 6876–6884. [Google Scholar] [CrossRef]
- Jones, C.H.; Tuckman, M.; Howe, A.Y.; Orlowski, M.; Mullen, S.; Chan, K.; Bradford, P.A. Diagnostic PCR analysis of the occurrence of methicillin and tetracycline resistance genes among Staphylococcus aureus isolates from phase 3 clinical trials of tigecycline for complicated skin and skin structure infections. Antimicrob Agents Chemother 2006, 50, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Price, L.B.; Stegger, M.; Hasman, H.; Aziz, M.; Larsen, J.; Andersen, P.S.; Pearson, T.; Waters, A.E.; Foster, J.T.; Schupp, J.; et al. Staphylococcus aureus CC398: host adaptation and emergence of methicillin resistance in livestock. mBio 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, A.B.; Skov, R.; Pallesen, L. Detection of low-level methicillin-resistant Staphylococcus aureus with commercially available tests. J Clin Microbiol 2003, 41, 3458. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Roth, R.; Peters, G. Rapid and specific detection of toxigenic Staphylococcus aureus: use of two multiplex PCR enzyme immunoassays for amplification and hybridization of staphylococcal enterotoxin genes, exfoliative toxin genes, and toxic shock syndrome toxin 1 gene. J Clin Microbiol 1998, 36, 2548–2553. [Google Scholar] [CrossRef] [PubMed]
- Monday, S.R.; Bohach, G.A. Use of multiplex PCR to detect classical and newly described pyrogenic toxin genes in staphylococcal isolates. J Clin Microbiol 1999, 37, 3411–3414. [Google Scholar] [CrossRef]
- McLauchlin, J.; Narayanan, G.L.; Mithani, V.; O'Neill, G. The detection of enterotoxins and toxic shock syndrome toxin genes in Staphylococcus aureus by polymerase chain reaction. J Food Prot 2000, 63, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Omoe, K.; Ishikawa, M.; Shimoda, Y.; Hu, D.L.; Ueda, S.; Shinagawa, K. Detection of seg, seh, and sei genes in Staphylococcus aureus isolates and determination of the enterotoxin productivities of S. aureus isolates Harboring seg, seh, or sei genes. J Clin Microbiol 2002, 40, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, P.; Nair, M.K.; Annamalai, T.; Venkitanarayanan, K.S. Phenotypic and genotypic characterization of bovine mastitis isolates of Staphylococcus aureus for biofilm formation. Vet Microbiol 2003, 92, 179–185. [Google Scholar] [CrossRef]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Nesme, X.; Etienne, J.; Vandenesch, F. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect Immun 2002, 70, 631–641. [Google Scholar] [CrossRef]
Figure 1.
Dendrogram of the SmaI profiles of 34 S. aureus isolates from surfaces, ingredients, meat batters and final products of dry-cured meat products processing facilities.
Figure 1.
Dendrogram of the SmaI profiles of 34 S. aureus isolates from surfaces, ingredients, meat batters and final products of dry-cured meat products processing facilities.
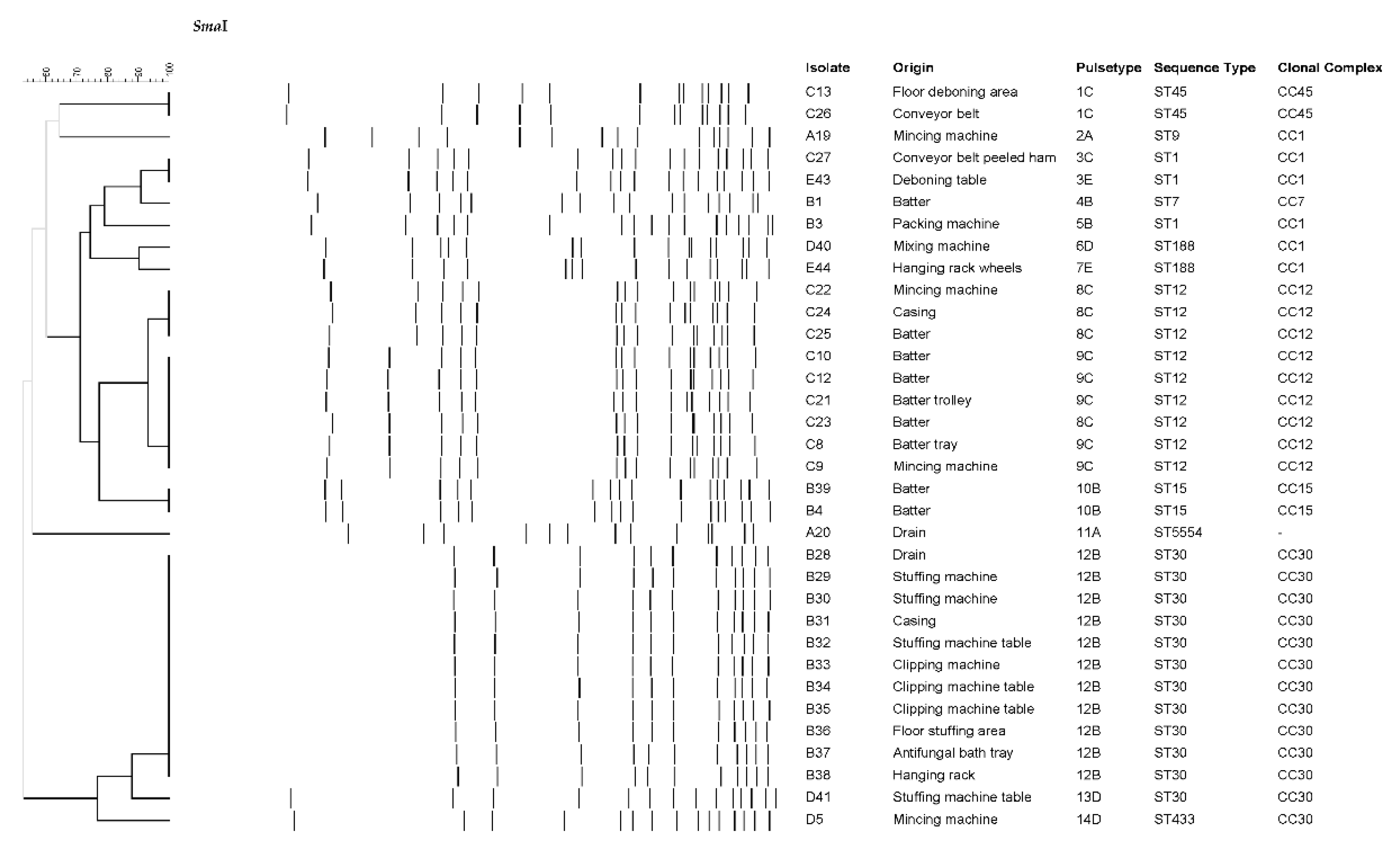
Figure 2.
Minimum Spanning Tree (MST) of the 38 S. aureus isolates from surfaces, ingredients, meat batters and final products of dry-cured meat products processing facilities. The STs are displayed as circles, proportional to the number of isolates. The origin (processing plant) of the isolates is shown with different colors. .
Figure 2.
Minimum Spanning Tree (MST) of the 38 S. aureus isolates from surfaces, ingredients, meat batters and final products of dry-cured meat products processing facilities. The STs are displayed as circles, proportional to the number of isolates. The origin (processing plant) of the isolates is shown with different colors. .
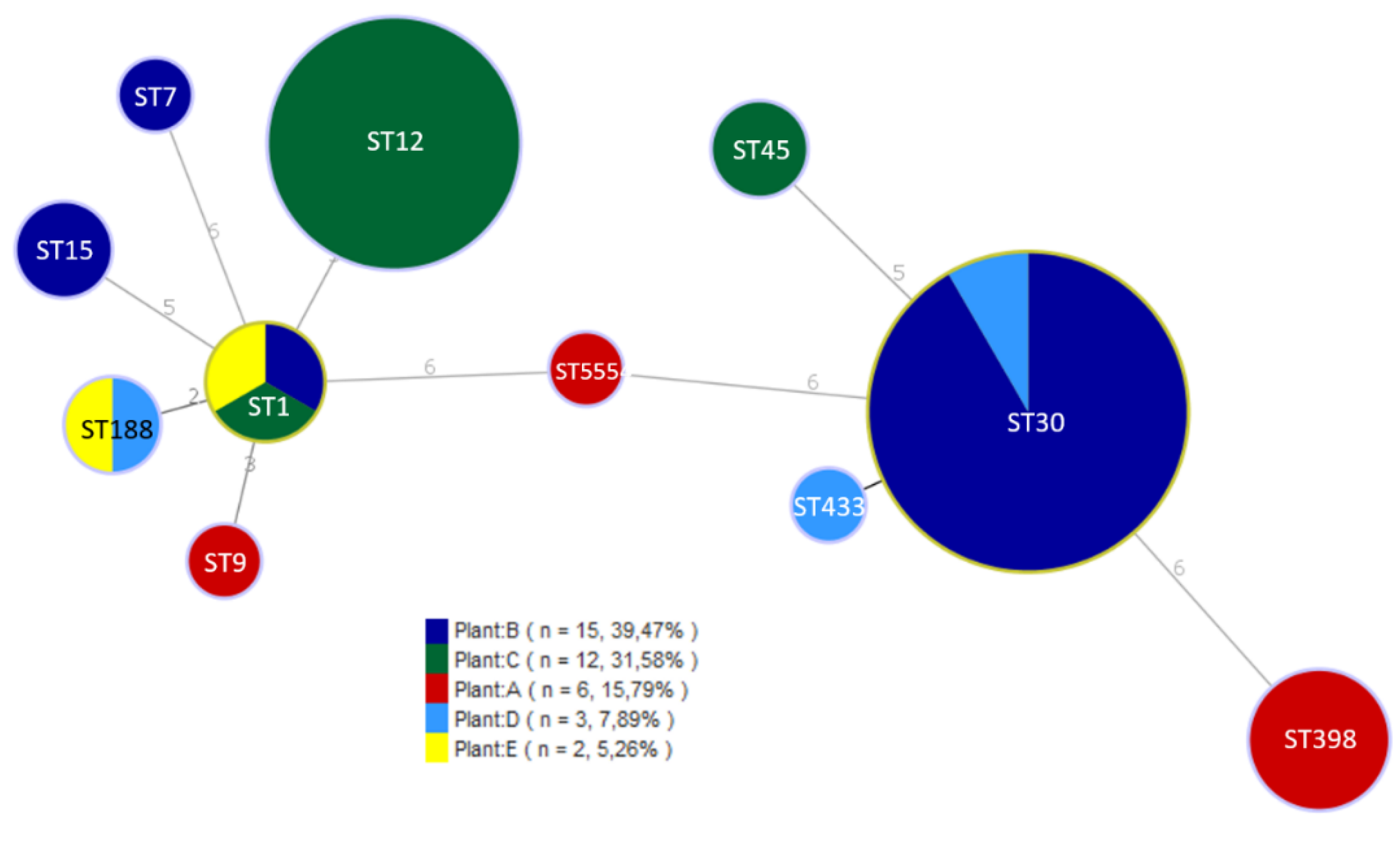
Figure 3.
Scheme of different egc types, along with the associated ST. The amino acid substitutions compared with the wild type (deposited sequence of amino acids) in the genes are also shown.
Figure 3.
Scheme of different egc types, along with the associated ST. The amino acid substitutions compared with the wild type (deposited sequence of amino acids) in the genes are also shown.
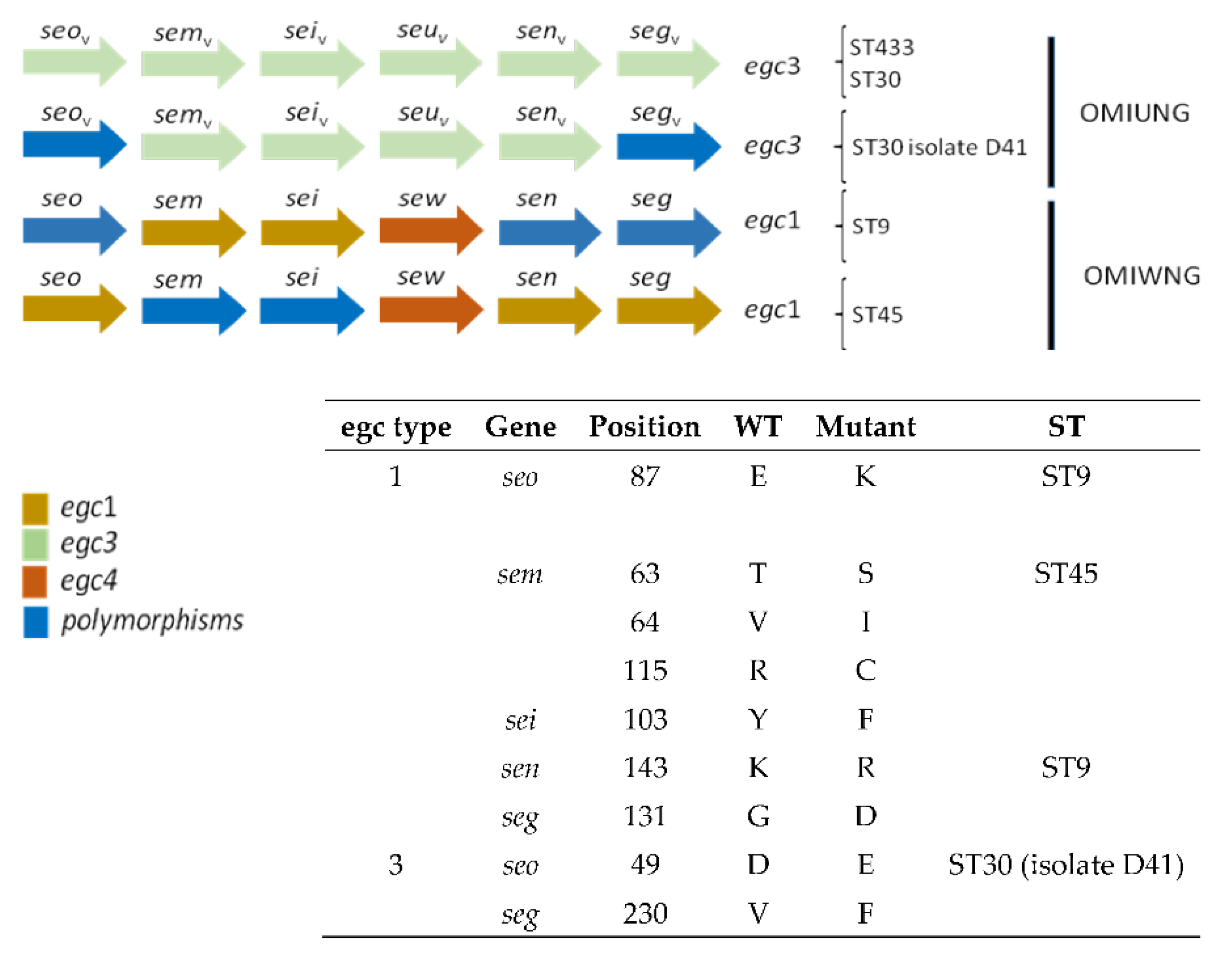

Table 1.
Prevalence of S. aureus in the environmental surfaces, ingredients, meat batters and final products from six dry-cured meat products processing facilities.
Table 1.
Prevalence of S. aureus in the environmental surfaces, ingredients, meat batters and final products from six dry-cured meat products processing facilities.
| Plant | ES | DP | ACD | FCS | NFCS | IMB | FP |
|---|---|---|---|---|---|---|---|
| No. positives/ No. samples |
No. positives/ No. samples |
No. positives/ No. samples |
No. positives/ No. samples |
No. positives/ No. samples |
No. positives/ No. samples |
No. positives/ No. samples |
|
| A | 4/123 | 3/122 | 1/1 | 3/49 | 1/74 | 2/18 | 0/4 |
| B | 11/183 | 11/129 | 0/54 | 6/98 | 5/85 | 4/29 | 0/7 |
| C | 7/139 | 6/71 | 1/68 | 4/60 | 3/79 | 5/11 | NA |
| D | 3/104 | 2/53 | 1/51 | 2/49 | 1/55 | 0/10 | 0/3 |
| E | 2/97 | 2/60 | 0/37 | 1/61 | 1/36 | NA | NA |
| F | 0/74 | 0/69 | 0/5 | 48 | 26 | NA | NA |
| Total | 27/720 | 24/504 | 3/216 | 16/365 | 11/355 | 11/68 | 0/14 |
ES, environmental surfaces; DP, during processing; ACD, after cleaning and disinfection; FCS, food contact surfaces; NFCS, non-food contact surfaces; IMB, ingredients, meat batters; FP, final products; NA, not analyzed.
Table 2.
Virulence and antimicrobial resistance of S. aureus strains isolated from surfaces, ingredients, meat batters and final products of six dry-cured meat products processing facilities.
Table 2.
Virulence and antimicrobial resistance of S. aureus strains isolated from surfaces, ingredients, meat batters and final products of six dry-cured meat products processing facilities.
| Isolate | Plant | ST | Toxin genes | Antibiotic resistance |
|---|---|---|---|---|
| A42 | A | ST398 | - | PEN, FOX, ERY, TET, CIP |
| A6 | A | ST398 | - | PEN, FOX, TET |
| A17 | A | ST398 | - | PEN, TET |
| A18 | A | ST398 | - | PEN, FOX, ERY, TET |
| A19 | A | ST9 | seg, sei, sem, sen, seo, sew, tst | PEN, TET |
| A20 | A | ST5554 | tst | - |
| B1 | B | ST7 | - | - |
| B3 | B | ST1 | seb, seh, tst | PEN |
| B4 | B | ST15 | tst | PEN, TET |
| B28 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B29 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B30 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B31 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B32 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B33 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B34 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B35 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B36 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B37 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B38 | B | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| B39 | B | ST15 | tst | PEN, TET |
| C8 | C | ST12 | tst | - |
| C9 | C | ST12 | tst | - |
| C10 | C | ST12 | tst | - |
| C12 | C | ST12 | tst | - |
| C13 | C | ST45 | sec, seg, sei, sem, sen, seo, sew, tst | PEN |
| C21 | C | ST12 | tst, lukED | - |
| C22 | C | ST12 | tst, lukED | - |
| C23 | C | ST12 | tst, lukED | - |
| C24 | C | ST12 | tst, lukED | - |
| C25 | C | ST12 | tst, lukED | - |
| C26 | C | ST45 | sec, seg, sei, sem, sen, seo, sew, tst | PEN |
| C27 | C | ST1 | seh, tst | PEN, ERY |
| D5 | D | ST433 | seg, sei, sem, sen, seo, seu | - |
| D40 | D | ST188 | sea, tst | PEN |
| D41 | D | ST30 | sea, seg, sei, sem, sen, seo, seu, tst | PEN |
| E43 | E | ST1 | seh | PEN, ERY |
| E44 | E | ST188 | tst | PEN |
PEN, penicillin; FOX, cefoxitin; CLD, clindamycin; ERY, erythromycin; TET, tetracycline; CIP, ciprofloxacin; GEN, gentamicin; STX, trimethoprim plus sulfamethoxazole.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



