
Submitted:
28 April 2023
Posted:
28 April 2023
You are already at the latest version
Abstract
The emergence of cloud computing, big data analytics, and machine learning has catalysed the use of remote sensing technologies to enable more timely land management of sustainability indicators such as ground cover and grassland biomass, given the uncertainty of future climate and drought conditions. Here, we examine the potential of “regenerative agriculture”, as an adaptive grazing management strategy to minimise bare ground exposure while maximising pasture biomass productivity. High-intensity sheep grazing treatments were conducted in small fields (less than 1 hectare) for short durations (typically less than 1 day). Paddocks were subsequently spelled to allow pasture biomass recovery (treatments comprising 3, 6, 9, 12, and 15 months) with each compared with control treatments with lighter stocking rates for longer periods (2,000 DSE). Pastures were composed of wallaby grass (Austrodanthonia species), kangaroo grass (Themeda triandra), Phalaris (Phalaris aquatica, and cocksfoot (Dactylis glomerata) were destructively sampled to estimate total standing dry matter (TSDM), standing green biomass, standing dry biomass and trampled biomass. We then invoked a machine learning model using Sentinel-2 imagery to quantify TSDM, standing green biomass and standing dry biomass. Faced with La Nina conditions, regenerative grazing did not significantly impact pasture productivity, with all treatments showing similar TSDM and green biomass. However, regenerative treatments significantly impacted litter fall and trampled material, with the high intensity grazing treatments causing more dry matter trampling, increasing litter, enhancing decomposition rates and surface organic matter. Pasture digestibility was greatest for treatments with minimal spelling (3 months), whereas both standing senescent and trampled material were significantly greater for the treatment with 15-month spell periods. Estimates of TSDM using machine learning with Sentinel-2 imagery underestimated TSDM in treatment plots but explained spatiotemporal variability associated within and across treatments. The root mean square error between the measured and modelled TSDM was 903 kg DM/ha, which was less than the variability measured in the field. We conclude that regenerative grazing with short recovery periods (3-6 months) are most conducive to increasing pasture production under high rainfall conditions, and we speculate that high intensity grazing is likely to positively impact on soil organic carbon through increased litterfall and trampling. Our study paves the way forward for using machine learning with satellite imagery to quantify pasture biomass at small scales, enabling management of pastures from afar.
Keywords:
remote sensing
; machine learning
; regenerative grazing
; grassland biomass
; total standing dry matter
; digital agriculture
; grazing management
; climate change
; Cibo Labs
; Sentinel-2
1. INTRODUCTION
Grasslands comprise key terrestrial ecosystems, providing feed and habitat for domesticated livestock and wildlife globally [1,2,3]. Grasslands allow significant carbon sequestration [4,5,6] in addition to existing carbon stocks they prevent from entering the atmosphere [7]. The resilience of grassland and land conditions to extreme drought and future climate requires an innovative agroecosystem approach that promotes functional biological drivers (such as soil microbial activities) and adaptive grazing management [8]. One such adaptive technique is using regenerative grazing principles [8,9] to stimulate ecosystem functions through short, intense grazing, adjustable stocking rate, and multi-paddock-system at the farm level (1 – 100 ha) with long rest periods allowing pasture biomass and land to recover. In regenerative grazing, residual biomass from the trampling effects of grazing livestock plays a significant role in reducing bare ground, enabling soil health (through soil microbial functionality), litter conversion, soil aggregation and porosity, and carbon sequestration [8,10]. Stimulation of organic microbial activities through residual biomass and trampling effects of grazing livestock contrasts with the unsustainable system of using conventional farm inputs (irrigation, synthetic fertilizers, etc.) [8]. In practice, information about regenerative grazing is based on anecdotal rather than evidence [10,11,12], thus, empirical research is needed to support this claim. Since the current information is not experimentally driven, available monitoring tools have not been tested to understand their usefulness to end-users. Due to large land areas and the dynamic and spatially variable nature of grazing [13,14], physical monitoring of grassland condition is often cumbersome, particularly where land areas are remote, large, and/or geographically challenging.
The rise of satellite imagery, cloud computing, big data analytics and machine learning has paved the way for innovative opportunities for land managers to remotely monitor crop, pasture or grassland biomass from afar [15,16]. In theory, availability of such technology would improve cost-efficiencies and timeliness of management by allowing monitoring of important sustainability indicators, such as ground cover, persistence and above-ground biomass [17] and, when coupled with decision-support tools that allow contrasting of agricultural management options [18,19,20], improvements in long-term sustainability. More timely monitoring of ground cover, vegetation, litter and biomass as influenced by land use [21] would also be expected to reduce labour associated with physical monitoring and improve farm business profitability [22,23]. Increasingly frequent global issues, such as conflict, COVID-19 [24], extreme weather events and climate change will likely catalyse demand for technologies for remote land management in the coming decades [17,25,26,27].
Conventional methods for monitoring pasture biomass and livestock utilisation (i.e., ground-based measurement and proximal sensing) are limited in terms of scope, and both spatial and temporal extent [28]. Previous research in Australia [29], United Kingdom [30], New Zealand [31], and the United States [32] has reported limitations of ground sampling approaches (i.e., visual, rising plate meter and destructive method by clipping) in quantifying the spatial variability of pasture biomass. By contrast, remote sensing provides timely spatiotemporal information that can predict availability of feed prior to grazing [30], allowing for feed budgeting. However, in most cases, remote sensing of pasture biomass is not process-driven (i.e., based on vegetation indices); often the use of such reflectance indices at small field scales (e.g., less than 50 hectares) is constrained by the resolution of the satellite imagery [30,33] and accurate calibration [34]. Remote sensing that considers process-based retrieval of pasture biomass and other biophysical variables may invoke site-specific modelling and machine learning techniques [35]. Although some successes have been reported, physical-based techniques such as radiative transfer modelling and light use efficiency modelling can be prohibitive as they may require a set of parametric rules for different study locations [36,37,38]. However, machine learning techniques including artificial neural networks (ANN) [16], random forest (RF) [39], and support vector machine (SVM) [32] are not site-specific and can be used to retrieve pasture biomass estimates [33]. ANN [16] was used to estimate pasture biomass leveraging multitemporal Sentinel-2 data collected over dairy farms in Tasmania [16]. The study showed that the accuracy of ANN improved when meteorological variables were included in the model; indeed, much process-based modelling is based primarily on longitudinal measurements of climate at a given site [2,34,40]. However, process-based applications are required as an operational service to support farm management - what is often known as a decision support system (DSS) [16,28,41] – and is often limited by the accuracy of site-specific soil characterisation [42,43].
Previous estimates of pasture biomass at the field (paddock) scale with machine learning algorithms has used standing green vegetation as a proxy to quantify the actual biomass from normalised difference vegetation index (NDVI) [32,39,44,45]. Information derived from NDVI can provide sufficient information about active photosynthetic [46] vegetation, whereas non-green senescent pasture species or dormant vegetation are often much more difficult to quantify due to their low reflectance in the near-infrared [47]. To successfully realise improved land-use sustainability through more timely, accurate biologically-intelligent monitoring of pasture sustainability indicators, more robust approaches are urgently needed [41,48,49,50]. This would also allow livestock farmers to better predict feed on offer (for total green and non-green forage) enabling planning of their stocking rate to maximise liveweight production while maximising environmental stewardship [42,43]. While a range of commercial technologies exist (e.g., Agroinsider, FORAGE, Cibo Labs, SPACETM and Pasture.io [48,50]), outputs from many of these applications are site specific and others have not been validated. This raises questions as to how well such applications predict pasture biomass outside their zone of calibration.
The launch of European Space Agency’s Sentinel-2 satellites has enhanced the development of “agricultural technology” or “Ag-tech” companies offering products aimed at quantifying land surface conditions. One such company – “Cibo Labs” (https://www.cibolabs.com.au/) - uses a predictive time series machine learning approach to derive spectral information from Sentinel-2 data about local properties at the field scale. Cibo Labs uses pasture cuts to train and validate the total standing dry matter (TSDM) model. Several thousand fields from farms across Australia are used to train a deep neural network (DNN). Cibo Labs uses the dropout regularisation method to reduce overfitting and computational costs, hence improving the generalisation of the DNN [51]. This is achieved by randomly dropping units (i.e., hidden and visible layers) to improve the neural network's performance during training. Hitherto the present study, Cibo Labs validated total standing dry matter (TSDM) estimates using 2,000 field measured samples collected over two years from across eastern and northern Australia. Thirty-three percent of field sites were used to train a three-layer, multilayer perceptron regression model (MPRM) using a 50% dropout and a maximum norm constraint [52,53,54]. The remainder of the field samples were used for validation. The model was trained with 100 iterations (~16,000 epochs) before reaching a termination criterion characterised by a median prediction error of 295 ± 8 kg DM/ha.
While such predictive accuracy was within the variability of measured data, the study was primarily conducted using measurements taken from low-latitude environments (the Northern part of Australia). Also, previous investigations of Cibo Labs' utility did not consider regenerative grazing principles implemented at the farm level. Therefore, it remains to be seen how well Cibo Labs performs in mid latitude environments such as the island state of Tasmania, where cloud cover in winter and spring is frequent [55], as well as examine if the tool can support regenerative grazing at the farm level. Clouds reduce spatial and temporal coverage by reducing target clarity and increasing time between clear useable images [15,16]. In the present study, we used a destructive sampling method to measure the total standing dry matter (kg DM/ha), equivalent to standing green and standing dry before and after grazing, with 3, 6, 9, 12 and 15 months of biomass regrowth. We applied regenerative grazing to the smaller plots of similar size (< 1 ha), while three plots of size 10 – 50 ha were used as controls (i.e., business-as-usual grazing). Our hypothesis was that the treatment plots or disturbance caused by the high stocking density would account for the TSDM variability. The key aim was to examine the effects of regenerative grazing on TSDM productivity in the plots and whether Sentinel-2 imagery and the Cibo Labs model could estimate the TSDM at the plot level. This was conducted by comparing Cibo Labs estimates of TSDM with destructively sampled pasture biomass for a site in south-eastern Tasmania subject to sheep grazing treatments.
Our objectives were to thus provide insight into: (1) the effects of regenerative grazing on TSDM productivity, consumption and trampling and (2) the usefulness of Sentinel-2 imagery and accuracy of the Cibo Labs model to estimate TSDM on effects of regenerative grazing at the farm level.
2. MATERIAL AND METHODS
2.1. Study site
The location for this study was south-eastern Tasmania, Australia. We worked on a case study farm (S 42° 30, E 147° 59) north of the town of Triabunna called ‘Okehampton’. The average annual rainfall at this location is 648 mm while the average annual minimum and maximum temperature are 7oC and 17oC, respectively [56]. Okehampton consists of 52 paddocks of sizes ranging from 1 - 138 ha covering an estimated area of 1446 ha (Figure 1). The botanical composition of fields comprises a mixture of native and sown pastures with mostly annual and perennial ryegrass (wallaby grass (Austrodanthonia species), kangaroo grass (Themeda triandra), Phalaris (Phalaris aquatic) and cocksfoot spear grass (Dactylis glomerata) [57]. The absence of irrigation and synthetic fertilizers on this site and the goal to stimulate pasture growth to improve livestock production, demand that agronomic systems implemented be sustainable, profitable, inclusive and enduring—especially given the uncertainty of future climate conditions in this region [18,40]. The farm has a history of sheep grazing but the field layouts have evolved over time to accommodate inclusive, intensive grazing management, conservation of biodiversity and environmental stewardship, including protection of endangered grass species and implementation of cultural burning practices informed by the local indigenous people (Pakana Services).
2.2. Regenerative grazing data collection
Twelve paddocks nominated by the case study farmer were used for the field sampling campaign. Biomass samples were collected by a local consultant from December 2021 through November 2022. Grazing was conducted for 1 day in the treatment plots (early morning to late evening). The control has “business-as-usual” grazing (Table 1). Pasture biomass fractions were generally quantified before grazing. Three plots [Vault control (VC), lower Bougainville (LB) and upper Bougainville (UB)] were used as controls following grazing regimes that were business-as-usual. These plots were grazed for longer period (weeks) at lower stocking rates (2,000 DSE) than the intensive treatments (i.e., the other seven paddocks) and allowed less time between subsequent grazing compared with intensively grazed paddocks. Control paddocks were larger in size compared with treatment plots. Treatment plots were stocked at the same rate while following adjusted stocking density (Table 1) and grazed for one day in consecutive days within the same week to minimise potential confounding effects of weather impacts on pasture growth, then rested for three, six, nine or twelve months before re-grazing. Treatment plots were conducted based on ‘regenerative’ principles that conduct short, intense grazing, with long rest periods allowing pastures to recover [25]. In contrast, control paddocks were grazed at lighter stocking rates, for longer durations, and allowed less time to recover (Table 1). Henceforth, the business-as-usual plots would be called BUA.
Pasture biomass was harvested to the ground level from five locations (quadrats) that were predetermined within each plot (from plot points with red layouts in Figure 1) using a battery-operated shearing handpiece and a 0.25 m2 quadrat (a square of 0.5 x 0.5 m). Standing biomass (green and dry) was cut prior to grazing while standing residuals (green and dry) and trampled biomass (green and dry) were taken post grazing, in a location immediately adjacent to the pre-grazing biomass harvest. Biomass was quickly placed in sealed, labelled plastic bags transported to a 4oC room in the laboratory where each bag was weighed after dung was excluded. The biomass was mixed, and using a quartering method, subsampled for separation and drying. Sub-samples of green and dry biomass were separated and then dried in a 60oC oven for at least 48 hours, before being weighed using a Mettler scale. This process was repeated for post-grazing biomass in some of the paddocks that were grazed. To account for the high volume of trampled biomass (i.e., biomass lying on the surface disturbed by the high density of sheep) this component was measured separately from the standing biomass (Table 1). Total standing dry matter (TSDM) was computed by the summation of green and dry biomass without trampled components. Thus, to determine the actual biomass utilised during a post-grazing event, we used Equation 1, and Equation 2 for total trampled dry matter (TTDM).
total standing dry matter (TSDM) - trampled residual = Biomass consumed
trampled green dry matter + trampled senesced dry matter = Total trampled dry matter (TTDM)
Since the sampled biomass collected from the five locations was done only once in each plot following a predetermined layout (plot points in Figure 1), we computed the mean for these locations to account for sampling error and tested if the treatment plots and grazing days have a statistically significant effect on biomass using ANOVA and general linear model. We developed a time series analysis for the treatment plots (including BUA) and compared with statistical outcomes.
The experiment was for twelve months, from December 2021 to November 2022, where the effects of short, intense grazing compared with the conventional grazing (control) on plot treatments for their drought resilience were observed. Hence, the experiment covered the four seasonal variations (summer, winter, autumn, and spring) in the study area. Summer is from December to February; autumn is from April to May; winter is from June to August; spring is from September to November [15].
2.3. Remote sensing
Estimates of TSDM were derived using the ‘PastureKey’ app within Cibo Labs, which is produced using 10 m resolution Sentinel-2 imagery provided by the European Space Agency (ESA). Only cloud-free pixels of Sentinel-2 imagery is used by Cibo Labs, and the application produces TSDM estimates for cloud-free paddocks every 5-days (Sentinel 2 revisit time). Cloudy pixels are detected and masked with the ‘Fmask’ algorithm [58]. Ten bands (b2, b3, b4, b5, b6, b7, b8, b8A, b11 and b12) of Sentinel-2 imagery were used to derive TSDM products. Using a predictive machine learning approach driven by deep neural networks (DNN), measured data are trained to predict TSDM within and across the paddock for every satellite revisit across a property. Cibo Labs uses Sentinel-2 bands from several thousand paddocks and dates of satellite imagery acquisition to train a three-layer, multilayer perceptron neural network regression model using a 20-50% dropout regularisation method. The dropout regularisation method addresses the problem of overfitting [51].
Pasture estimates in near real time are available from the PastureKey application within Cibo Labs. Hereafter, PastureKey application would be referred to as Cibo Labs for convenience. The multilayer perceptron model can learn in real-time, complementing the delivery of products to end-users in cloud optimised Geotiff (COG) format. Estimates of pasture biomass are available on demand or in a batch mode through a high-performance computing (HPC) environment.
2.4. Comparing measured pasture biomass with satellite estimates
On the account that Sentinel-2 could retrieve total standing green and dry matter from the plot with a size less than 1 ha, pasture estimates from Cibo Labs were evaluated by comparison with corresponding measured values for each time point (in each case using the most proximal Sentinel-2 imagery). Comparisons of measured against estimated data were assessed using, time series trendline and error bar, root mean square error and R2 following [14,40,59].
3. RESULTS
3.1. The effects of regenerative grazing on pasture productivity, consumption and trampling
In all treatments and BUA plots, pasture biomass removal through intensive and conventional (control) grazing typically shows biomass loss between pre-grazing (December 2021 - January 2022) and post-grazing (January - February) in phase 1 (Figure 3). This is also observed in phase 4, where Vault 2 and Vault 4 plots went through a post-grazing regime (Figure 3). The actual biomass consumed in the one-day grazing for all treatments is shown in Figure 3. As shown in Figure 3, the actual biomass utilised (see Equation 1) through grazing is low compared to the trampled biomass (see Equation 2) for all treatment plots (BUA inclusive). Therefore, trampling has a more significant effect on the TSDM than the actual grazing (i.e., consumption).
All treatments, including BUA plots, show similar temporal variability and trends of total standing biomass (Figure 4). A similar trend is observed in standing green DM and standing dry DM (Figure 5 and Figure 6). This indicates that the grazing intervals and the resting periods (3, 6, 9, 12 and 15 months) did not significantly influence biomass recovery or productivity in the experiment. For instance, after the first three months of rest in phase 1, where all treatment plots were grazed, pastures did not recover to the biomass level at the start of the experiment, likely due to seasonal variation in rainfall and temperature (Figure 5). However, following the consecutive increase in rainfall and temperature in winter through spring, total standing biomass increased, as observed in the Vaults and Bougainville 3 & 4 treatments (Figure 5). For example, the treatment plot (Vault 1) grazed only once (i.e., 12 months of rest) was similar to the Vault 4 plot, which was grazed every three months. In the same way, the Vault 5 plot that has not been grazed (i.e., 15 months of rest) is similar to the plot that was grazed every three months (Vault 4). In like manner, the Vault 4 treatment is similar to the Bougainville 2, 3 & 4 plots that were left ungrazed after phase 1 (Figure 3 and Figure 4). Only Bougainville 3 plot exceeded Vault 4 treatment in the TSDM during spring by 3,000 kg DM/ha. Therefore, biomass removal and recovery through regenerative grazing or conventional method does not influence Vaults and Bougainville treatments (Figure 3).
During spring, the TSDM in the Bougainville 1-4 plots varied due to rainfall, as shown in Figure 3 and Figure 4. The Bougainville 3 plot had more available TSDM than the Vault 5 treatment, as seen in Figure 4. Since the Bougainville plots are situated on a sloping hill, it is uncertain if their location played a role in the significant biomass growth observed in Bougainville 2, 3, and 4 during spring, as depicted in Figure 4. The Bougainville 1 plot had the lowest TSDM volume.
ANOVA and generalized linear models showed no significant association between plots and pasture biomass productivity (TSDM). However, there were significant differences when the date of grazing was used as an effect of treatment. Generally, the ANOVA test shows the effect of grazing date is statistically significant (p< 0.001) to the TSDM, while the post hoc Dunnett test does not show the level of interaction. Analysing the effects of dates of treatments and TSDM further with interaction using the GML model shows strong evidence of significant difference (p < 0.05) on 27th January 2022 by an estimated –11, 076 kg DM/ha compared to other grazing dates. The treatment plots associated with the 27 January grazing event include Bougainville 2, Bougainville 4, Vault 1, Vault 2, Vault 3, Vault 4 & Vault 5. In contrast, there is no statistical evidence that the BUA (Lower Bougainville, Upper Bougainville, and Vault Control) is significantly different (p> 0.1) to variability in biomass. Therefore, we conclude that the effect of regenerative treatments (i.e., short, intense grazing and rest periods) in the plots did not affect TSDM productivity and consumption. Only the trampling effect (surface disturbed by the high density of sheep) associated with the 27 January 2022 post-grazing event in phase 1 for Vaults (1, 2, 3, 4, & 5) and Bougainville 2 & 4 plots explained the variability in biomass. We conclude that regenerative grazing did not have effect on pasture biomass productivity in the wet year of 2022. All treatment plots have similar results.
The Vault 4 plot with three months grazing interval has the highest volume of standing green DM compared with Vault 5 with 15 months of resting interval.
The Vault 5 plot with 15 months of resting interval has the highest standing dry matter compared to other treatment plots as there was no grazing in this period.
3.2. Satellite estimate of pasture biomass
Cibo Labs (PastureKey application) utilises Sentinel-2 imagery to estimate TSDM, standing green DM, and standing dry DM in all the treatment plots, pre- and post-grazing (Figure 7 and Figure 8). The matchup of Sentinel-2 imagery with the measured biomass measurements ranges from 2 to 40 days. In summer (5 December 2021 to 13 January 2022), where six plots (Upper Bougainville, Vault 1 to 5) had a lag of 40 days between Sentinel-2 imagery and the measured measurements. A two day difference was experienced in autumn (between 3 and 5 July 2022).
Cibo Labs accounted for the variability in the TSDM in the treatment plots (between and within) but underestimated this value, compared with the measured TSDM (Figure 10). There are instances (phases 1-4) where the satellite estimated TSDM values closely (Bougainville 1, 2, & 4 and Vaults 1, 3 & 4) matched the measured points. For all the treatment plots excluding Upper Bougainville and Lower Bougainville which went through conventional grazing (control), the Cibo Labs passes through one or more error bars, indicating it is within an acceptable variability of the measured biomass. The plot (Vault 4) which went through repeated grazing treatment every three months is more closely associated with the measured variability than Vault 1 which was grazed only once, or Vault 5 with 15 months of rest (Figure 7). The Vault 5 treatment plot has the highest variance.
The measured TSDM collected for the post-grazing event in Vault 5 on 27th January 2022 shows the total trampled residual (trampled green DM and trampled dry DM) was zero which implies that biomass in this plot was lying on the surface due to the effect of high density of sheep (Figure 7). In this treatment, the biomass utilised was 355 kg DM/ha for one day of grazing [Total standing dry matter (TSDM) before grazing – Total trampled dry matter (TTDM) after grazing, (11,076 – 10,721= 355 kg DM/ha]. The unutilised trampled residual (green and dry) that was measured, 10,721 kg DM/ha has a corresponding estimate of 1,004 kg DM/ha from the Cibo Labs. It, therefore, implies that although Cibo Labs underestimates the TSDM, it can account for the trampled residual that is of high volume.
Sentinel-2 imagery integrated with the Cibo labs model has a better capability of estimating standing green DM than standing dry DM (Figure 8). Although estimates are within the variability of the measured data points, in phase 1 (except Vaults 1 & 2 and Bougainville 1 & 2) and phase 2 of the experiment, Cibo Lab overestimated standing green DM (Figure 8). No clear relationship exists between the measured and estimates (Figure 7) for the 7th of November 2022 (phase 4), similar to Figure 7. There is no correlation between the measured standing dry DM and Cibo Labs estimates. Cibo Labs underestimated standing dry DM- estimates are barely above the ground level (Figure 8).
3.3. Spatial maps derived from Sentinel—2 imagery and Cibo Labs model
Cibo Labs derived Sentinel-2 maps for the treatment plots at Okehampton, Triabunna, Tasmania show spatiotemporal changes and the variability (within and across) in pasture biomass levels in all the treatment plots including control for pre-grazing and post-grazing activities like the time series plot discussed in Figure 7. Cloud-free Sentinel-2 imagery to quantify the available pasture biomass against the ground measurement collected on 13th January 2022 before grazing the fields (paddocks) was on 5th December 2021. This makes a lag of 40 days between the available cloud-free satellite imagery and the ground measurement. All treatment plots started with more pasture biomass before grazing. All fields were grazed in phase 1 and left to rest for three months. All fields were grazed in phase 1 and left to rest for three months. After rest, the ground measurement collected on 26th April indicates the treatment plots have not recovered in autumn (proximal Sentinel-2 imagery available on 24th July). However, satellite imagery available on 3rd July against the measured pasture biomass collected on 5th July shows the plots (Vault and Bougainville) show increasing TSDM during winter. The map indicates Bougainville 1 (bottom) is the least-performing treatment plot with reference to phase 3 of the experiment. The maps correspond to Figure 3 and Figure 4 and the modelling time series in Figure 7. As shown earlier (Figure 3 and Figure 4), the map confirmed that Bougainville 1 is the least-performing plot.
4. DISCUSSION
4.1. The effects of regenerative grazing on pasture biomass productivity, consumption and trampling
This study examined the effect of regenerative grazing treatments (i.e., short, intense grazing and rest periods) with smaller plots (less than 1 ha) on pasture productivity, consumption, and trampling. In the treatment plots examined, regenerative grazing did not influence pasture biomass productivity in the wet year of 2022. All treatment plots, including the ones used for conventional grazing (control), have similar results (Figure 3, Figure 4, Figure 5 and Figure 6). ANOVA and generalized linear models (GLM) showed no significant association between treatment plots and pasture biomass productivity (TSDM). However, there were significant differences when the date of grazing was used as an effect of treatment (Section 3.1). GLM model shows a strong statistical significance exists only with the treatment plots (i.e., Vault 1, Vault 2, Vault 3, Vault 4, Vault 5, Bougainville 2, and Bougainville 4) associated with the post-grazing event of 27th January 2022 (Section 3.1). Therefore, this study concluded that the variability in the TSDM can only be explained with the treatment plots associated with post-grazing regime in phase 1 of the experiment. The time series charts in Figure 3, Figure 4, Figure 5 and Figure 6 confirm that although all treatment plots exhibited similar results, Vaults 4 and 5 showed significant variability with pasture biomass productivity. Similarly, the Bougainville 2, 3 & 4 plots benefited from rainfall to produce more biomass in the spring [60].
The effect of resting interval (3, 6, 9, 12, and 15 months) for TSDM to recover in the plots did not contribute to biomass variability (Figure 3 and Figure 4). The main effect of treatment in the plots is associated with the high stocking rate, which resulted in a high volume of trampling residual (i.e., 27 January 2022). This implies that the actual biomass utilised (i.e., TSDM minus trampling residual) for grazing in the treatment plots was significantly low (Figure 3). In all treatment plots (including the BUA), the recovery or productivity of TSDM from summer through spring due to increasing rainfall followed a similar pattern (Figure 4). This showed that the influence of weather contributed to biomass recovery in a similar way, thereby confounding the effect of other treatments. For example, there was no significant difference between the Vault 4 treatment, which was grazed every three months, and Vault 5 with 15 months of rest. Similarly, there was no significant difference between Vault 1 treatment with 12 months grazing plan and the Vault 4 plot (Figure 3 and Figure 4).
The present study has shown that although grazing through an intensive or conventional approach reduces pasture biomass [12], intensively grazed paddocks/fields through a regenerative strategy provide pastures with adaptive management for quick biomass recovery and reduction of bare ground. The plot (Vault 4) subjected to three months of resting interval utilised residual biomass from the trampling effects of grazing and optimum weather conditions to produce the highest volume of standing green DM over other treatment plots (Figure 5). Therefore, we conclude that Vault 4 is the treatment plot with the best pasture biomass productivity. In contrast, the Vault 5 treatment plot with 15 months of resting interval produced the highest standing dry DM compared to other plots (Figure 6). The pasture biomass produced is actively senescing from lack of utilisation.
We emphasise that the impact of favourable weather confounded the effect of treatments on pasture biomass variability or biomass recovery. Hence the resilience of pastures biomass to drought could not be established. A longer resting interval is not recommended in a situation like this with good weather conditions. An earlier study under a simulated environment of rainfall and other treatment variables considered a 30-day resting period insufficient to recover soil samples from trampling caused by intensive grazing rotation [61]. Although this and few other studies approached regenerative grazing in the sense of soil recovery [61,62,63,64], the same principle as the strategy employed here (pasture biomass utilisation) is used to stimulate microbial activities and soil functions. However, this study is the first to use an approach where the experiment conditions followed natural processes with no farm inputs (fertilizer, irrigation etc.) and a simulated environment. Our results indicate that post-grazing data provides an incentive to determine the effect of trampling, which according to the analysis in this study, is limited. Trampling residual data provides information about the actual biomass utilised by the grazing livestock, which in turn gives insight into liveweight gain [61]. Also, the actual biomass utilised for grazing is negligible compared to the trampled residual. Therefore, to minimise biomass wastage through trampling while achieving the regenerative grazing sustainability [8], future work will focus on adjusting the stocking density to accommodate more grazing days (3 to 5 days). This is because, in practice, one day of grazing may be infeasible [61] with limited land resources and logistical constraints. In addition, having 3 to 5 days of adjustable stocking rate instead of 1-day grazing would support a more effective intensive rotational grazing regime within a multipaddock system. Future research opportunities exist in understanding resting period that will be sustainable to recover pasture from trampling effect.
4.2. Satellite estimates of pasture biomass
In this study, we examined the usefulness and accuracy of PastureKey, an application from the Cibo Labs, and derived from 10 m resolution Sentinel-2 imagery estimates of total standing dry matter to support regenerative grazing at the farm level. The usefulness of the tool was examined with respect to capturing TSDM (standing green DM and standing dry DM) variability in the treatment and business-usual plots, similar to the one obtained by destructive sampling approach. The accuracy of the Cibo Labs (used instead of PastureKey for convenience) was then examined by performing regression analysis on the interacting variables (standing green DM, standing dry DM and total standing dry matter) with the sampled biomass.
Satellite estimates derived from the Cibo Labs model are within the sampled biomass's variability for all treatment plots except the control (Figure 7). There is a closer correlation and high variability in TSDM with the Vault 4 plot, which has three months of resting interval and grazing treatment than other treatment plots. The standard error bars show that the measure of variability with the sampled biomass (Figure 7) correlates with the post-grazing event of 27 January 2022 for treatment plots Vault 3, Vault 4, Vault 5, Bougainville 2, and Bougainville 4, similar to the statistical (GML model) result obtained in Section 3.1. Therefore, Cibo Labs derived from Sentinel-2 imagery can monitor the spatiotemporal variability associated with TSDM for all post-grazing events and the plot (Vault 4) with a regular regenerative grazing plan at the farm level. The Vault 5 plot with the 15-month resting interval has the highest degree of uncertainty compared with other plots. In addition, our findings reveal that Sentinel-2 imagery can account for the trampled residual as in Vault 5, where the TSDM is zero against the trampled biomass (high volume of lying biomass) for the post-grazing event on 27th January (Figure 7). The total trampled residual in this plot was 11,0721 kg DM/ha compared to 1,004 kg DM/ha of Cibo Labs estimated as TSDM. While the Cibo Labs Sentinel-2-derived model could provide useful information about regenerative grazing for the treatment plots the plots used for conventional grazing (BUA) (Lower Bougainville, Upper Bougainville, and Vault Control) are challenging to estimate (Figure 7).
Regarding the accuracy of the Cibo Labs estimates, the model underestimated the total TSDM in all treatment plots with MAE of 745 kg DM/ha and RMSE of 903 kg DM/ha (Figure 7 and Figure 9) and overestimated the standing green DM (Figure 8 and Figure 10). In addition, the model significantly underestimated the standing dry DM. (Figure 8, Figure 11 and Figure 13). The overestimation of the standing green DM and underestimation of the standing dry DM by the Cibo Lab model reveals that the model calibration is too sensitive to green vegetation and less to dry vegetation. In spring, when biomass growth reached optimum, the model underestimated TSDM in all plots but performed better in the Bougainville plots. The performance of the Cibo Lab model in Bougainville 2, 3 & 4 plots in spring is associated with the slopy hill, which influences the vegetation growth, distribution, and variations in biomass and productivity [65]. In the same way, the underestimation of TSDM in all plots in spring was caused by environmental conditions [16,66] (excess rainfall and soil type), which were not considered during model calibration. In general, the confounding influence of rainfall discussed in Sections 3.1 & 4.1 hardly substantiated any variability in the treatment plots [67]. The fact that there was no statistical interaction between the treatment plots themselves and TSDM except with the grazing dates where we found strong evidence of significant difference with 27 January 2022, shows that there would have been a better correlation between satellite estimates and measured biomass with more post-grazing events. However, the time series charts (Figure 7 and Figure 8) show Sentinel-2 imagery and predictive machine learning model can provide estimates of pasture biomass in monitoring regenerative grazing at the farm level. Such estimates are available as a spatial map providing management decisions per plot as an indication of available pasture biomass. Previous work has demonstrated the capability of machine learning to derive pasture estimates from Sentinel-2 imagery at the farm level [16,39], though not applied to regenerative grazing schemes.
4.3. The feasibility of using Sentinel-2 imagery to estimate total standing dry matter
Here, we demonstrated that Sentinel-2 imagery could be used to retrieve TSDM, standing green DM, and standing dry DM through a simple but powerful predictive machine learning to support regenerative grazing that considers a regular grazing and recovery period. To the best of our knowledge, this study is the first to examine this approach at the farm level. However, cloud constraints are one of the major limitations to model performance accuracy in this study. Tasmania is considered a medium-high latitude environment [55]. Despite using smaller fields of less than 1 ha, clouds over Tasmania hindered the consistent availability of Sentinel-2 imagery to feed the predictive machine learning model used in this study. The lag effect between the available Sentinel-2 imagery and the sampling date ranges from 2 to 40 days (Figure 14). Earlier work supports the argument that time lag effects between field sampling and data from the satellite are a potential source of error to model performance [68,69,70].
Although the model could retrieve TSDM through inherent Sentinel-2 SWIR band inclusion [30], the association between Cibo Labs TSDM and standing dry DM shows a weak relationship (Figure 11 and Figure 13). Summer is characterised by a high concentration of senescence (Figure 7, Figure 8 and Figure 13) intermixed with green vegetation. Hence, it is challenging to distinguish senesced from healthy vegetation despite the high spatial resolution of Sentinel-2 satellite [71].
A future optimisation study on estimating TSDM with a similar predictive machine learning model will consider more robust ground/field samples that will complement observation satellite data to improve accuracy. In the present study, the number of data available was limited to the summer of 2021 through the spring of 2022 on one farm. The review of [33] suggests that the accuracy of machine learning approaches in estimating aboveground biomass depends on the data source, the number of ground/field samples, pasture species composition, and addressing the errors associated with the algorithms. The improvement of pasture biomass prediction with the ANN algorithm from a similar study on five farms in Tasmania was based on the inclusion of more input parameters (meteorological data) to achieve 0.60 [16]. Sentinel-1 imagery, a synthetic aperture radar, is effective in addressing clouds constrain, saturation of optical instruments and in cases where limited field dataset is available to estimate pasture biomass [72]. In addition, the frequency of Sentinel-2 imagery has been enhanced by interpolating a daily revisit high resolution of Planet Lab to account for missing data[71,73].
5. CONCLUSION
Regenerative grazing in a wet year like 2022 did not have significant effect on pasture productivity in all the treatment plots examined. All treatment plots exhibited similar outcomes making the effects of intensive and short grazing duration (1-day) confounded because of the influence of rainfall. In the one-day grazing treatment, sheep could not exploit selective grazing, but rather the trampling of pasture biomass, which is caused by the disturbance from the high stocking density in the treatment plots. The trampled residual from the post-grazing event was found to be statistically significant, thus, providing an insight into the source of variability in the treatment plots. In the one-day grazing, an insignificant biomass volume was utilised. Therefore, being one of the pioneering studies in this field, there is an opportunity for future research to understand the effect of regenerative grazing in drought or in a year with moderate rainfall. More work is needed to understand the effects of more grazing days (3 to 5) to make regenerative grazing sustainable. Also, more robust data on post-grazing should be considered since it is the main effect in the current study.
This study demonstrated that a predictive machine learning model could be developed using Sentinel-2 time-series imagery to estimate TSDM, standing green DM, and standing dry DM to support regenerative grazing at the farm scale. Although the model underestimated TSDM in all the plots, it is within the variability of the measured biomass. Specifically, the model could explain the variability in biomass for the plot (Vault 4) with a regular grazing and recovery period. Also, the model could show the treatment plot (Vault 5) with the highest level of variance. In a follow-up paper, the underestimation of the TSDM by the model would be resolved using complementary satellite imagery (i.e., PlanetScope or radar) to address cloud constraints.
We conclude that in regenerative grazing, the productivity of TSDM is associated with plots having a short-term recovery interval than long-term in a wet year, and remote sensing with a predictive machine learning model can be used as a support tool to enable regenerative grazing management decisions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. The following are available: (i) list of reviewed paper used for analysis. (ii) list of articles referenced from the main papers.
Author Contributions
Conceptualization, Methodology, Formal Analysis, Investigation, Writing-Original Draft Preparation, M.O.G.; Visualisation, M.O.G, M.T.H, J.G.; Review and Editing, J.G.; Review and Editing, A.M.F.; Review and Editing, R.A.C.; Review and Editing, C.M.; Review and Editing; P.S.; Review and Editing, P.T.; Sampling design and field data collection, J.W, M.T.H.; Review and Editing, M.T.H.
Funding
This work was funded through the University of Tasmania, Tasmanian Institute of Agriculture.
Acknowledgments
Funding support for the Tasmanian Institute of Agriculture was provided through funding from the Australian Government’s Future Drought Fund and the University of Tasmania. The authors acknowledge project delivery and infrastructure from the Cape Herbert Pty Ltd, Australia. Special acknowledgment to Peter Ball, who collected the measured data and made it available.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harrison, M.T.; Cullen, B.R.; Mayberry, D.E.; Cowie, A.L.; Bilotto, F.; Badgery, W.B.; Liu, K.; Davison, T.; Christie, K.M.; Muleke, A.; et al. Carbon myopia: The urgent need for integrated social, economic and environmental action in the livestock sector. Glob. Chang. Biol. 2021, 27, 5726–5761. [Google Scholar] [CrossRef]
- Christie, K.M.; Smith, A.P.; Rawnsley, R.P.; Harrison, M.T.; Eckard, R.J. Simulated seasonal responses of grazed dairy pastures to nitrogen fertilizer in SE Australia: N loss and recovery. Agric. Syst. 2020, 182, 102847. [Google Scholar] [CrossRef]
- Thornton, P.K. Livestock production: recent trends, future prospects. Philos. Trans. R. Soc. London. Ser. B, Biol. Sci. 2010, 365, 2853–2867. [Google Scholar] [CrossRef]
- Farina, R.; Sándor, R.; Abdalla, M.; Álvaro-Fuentes, J.; Bechini, L.; Bolinder, M.A.; Brilli, L.; Chenu, C.; Clivot, H.; De Antoni Migliorati, M.; et al. Ensemble modelling, uncertainty and robust predictions of organic carbon in long-term bare-fallow soils. Glob. Chang. Biol. 2021, 27, 904–928. [Google Scholar] [CrossRef]
- Abberton, M. Grassland carbon sequestration : management , Proceedings of the Workshop sequestration in the mitigation of climate change. In Food and Agriculture Organisation; 2010; Vol. Vol. 11.
- Franzluebbers, A.J. Soil organic carbon in managed pastures of the southeastern United States of America. Grassl. carbon sequestration Manag. policy Econ. Proc. Work. role Grassl. carbon sequestration Mitig. Clim. Chang. 2010, 2007, 163–175. [Google Scholar]
- Henry, B.; Dalal, R.; Harrison, M.T.; Keating, B. Creating frameworks to foster soil carbon sequestration. 2022.
- Teague, R.; Kreuter, U. Managing Grazing to Restore Soil Health, Ecosystem Function, and Ecosystem Services. Front. Sustain. Food Syst. 2020, 4. [Google Scholar] [CrossRef]
- Díaz de Otálora, X.; Epelde, L.; Arranz, J.; Garbisu, C.; Ruiz, R.; Mandaluniz, N. Regenerative rotational grazing management of dairy sheep increases springtime grass production and topsoil carbon storage. Ecol. Indic. 2021, 125, 107484. [Google Scholar] [CrossRef]
- Teague, R.; Barnes, M. Grazing management that regenerates ecosystem function and grazingland livelihoods. African J. Range \& Forage Sci. 2017, 34, 77–86. [Google Scholar] [CrossRef]
- Spratt, E.; Jordan, J.; Winsten, J.; Huff, P.; van Schaik, C.; Jewett, J.G.; Filbert, M.; Luhman, J.; Meier, E.; Paine, L. Accelerating regenerative grazing to tackle farm, environmental, and societal challenges in the upper Midwest. J. Soil Water Conserv. 2021, 76, 15A–23A. [Google Scholar] [CrossRef]
- Öllerer, K.; Varga, A.; Kirby, K.; Demeter, L.; Biró, M.; Bölöni, J.; Molnár, Z. Beyond the obvious impact of domestic livestock grazing on temperate forest vegetation – A global review. Biol. Conserv. 2019, 237, 209–219. [Google Scholar] [CrossRef]
- Harrison, M.; Evans, J.; Moore, A.D. Using a mathematical framework to examine physiological changes in winter wheat after livestock grazing 2. Model validation and effects of grazing management. F. Crop. Res. 2012, 136, 127–137. [Google Scholar] [CrossRef]
- Harrison, M.T.; Evans, J.R.; Moore, A.D. Using a mathematical framework to examine physiological changes in winter wheat after livestock grazing: 1. Model derivation and coefficient calibration. F. Crop. Res. 2012, 136, 116–126. [Google Scholar] [CrossRef]
- Ara, I.; Harrison, M.T.; Whitehead, J.; Waldner, F.; Bridle, K.; Gilfedder, L.; Marques Da Silva, J.; Marques, F.; Rawnsley, R. Modelling seasonal pasture growth and botanical composition at the paddock scale with satellite imagery. In Silico Plants 2021, 3, 1–15. [Google Scholar] [CrossRef]
- Chen, Y.; Guerschman, J.; Shendryk, Y.; Henry, D.; Harrison, M.T. Estimating pasture biomass using sentinel-2 imagery and machine learning. Remote Sens. 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Snow, V.; Rodriguez, D.; Dynes, R.; Kaye-Blake, W.; Mallawaarachchi, T.; Zydenbos, S.; Cong, L.; Obadovic, I.; Agnew, R.; Amery, N.; et al. Resilience achieved via multiple compensating subsystems: The immediate impacts of COVID-19 control measures on the agri-food systems of Australia and New Zealand. Agric. Syst. 2021, 187, 103025. [Google Scholar] [CrossRef]
- Phelan, D.C.; Harrison, M.T.; McLean, G.; Cox, H.; Pembleton, K.G.; Dean, G.J.; Parsons, D.; do Amaral Richter, M.E.; Pengilley, G.; Hinton, S.J.; et al. Advancing a farmer decision support tool for agronomic decisions on rainfed and irrigated wheat cropping in Tasmania. Agric. Syst. 2018, 167, 113–124. [Google Scholar] [CrossRef]
- Ibrahim, A.; Harrison, M.; Meinke, H.; Fan, Y.; Johnson, P.; Zhou, M. A regulator of early flowering in barley (Hordeum vulgare L.). PLoS One 2018, 13, e0200722. [Google Scholar] [CrossRef]
- Ara, I.; Harrison, M.T.; Whitehead, J.; Waldner, F.; Bridle, K.; Gilfedder, L.; da Silva, J.; Marques, F.; Rawnsley, R. Modelling seasonal pasture growth and botanical composition at the paddock scale with satellite imagery. in silico Plants 2020, 3. [Google Scholar] [CrossRef]
- Liu, K.; Harrison, M.T.; Shabala, S.; Meinke, H.; Ahmed, I.; Zhang, Y.; Tian, X.; Zhou, M. The State of the Art in Modeling Waterlogging Impacts on Plants: What Do We Know and What Do We Need to Know. Earth’s Futur. 2020, 8, e2020EF001801. [Google Scholar] [CrossRef]
- Ho, C.K.M.; Jackson, T.; Harrison, M.T.; Eckard, R.J. Increasing ewe genetic fecundity improves whole-farm production and reduces greenhouse gas emissions intensities: 2. Economic performance. Anim. Prod. Sci. 2014, 54, 1248–1253. [Google Scholar] [CrossRef]
- Shahpari, S.; Allison, J.; Harrison, M.T.; Stanley, R. An Integrated Economic, Environmental and Social Approach to Agricultural Land-Use Planning. Land 2021, 10. [Google Scholar] [CrossRef]
- Fayemi, P.O.; Fayemi, O.E.; Joel, L.O.; Ogungbuyi, M.G. COVID-19 Syndrome: Nexus with Herbivory and Exposure Dynamics for Monitoring Livestock Welfare and Agro-Environment. Sustainability 2021, 13. [Google Scholar] [CrossRef]
- Langworthy, A.D.; Rawnsley, R.P.; Freeman, M.J.; Pembleton, K.G.; Corkrey, R.; Harrison, M.T.; Lane, P.A.; Henry, D.A. Potential of summer-active temperate (C3) perennial forages to mitigate the detrimental effects of supraoptimal temperatures on summer home-grown feed production in south-eastern Australian dairying regions. Crop Pasture Sci. 2018, 69, 808–820. [Google Scholar] [CrossRef]
- Harrison, M.T. Climate change benefits negated by extreme heat. Nat. Food 2021, 2, 855–856. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Harrison, M.T.; Archontoulis, S. V; Huth, N.; Yang, R.; Liu, D.L.; Yan, H.; Meinke, H.; Huber, I.; Feng, P. Climate change shifts forward flowering and reduces crop waterlogging stress. Environ. Res. Lett. 2021, 16, 94017. [Google Scholar] [CrossRef]
- Hudson, T.D.; Reeves, M.C.; Hall, S.A.; Yorgey, G.G.; Neibergs, J.S. Big landscapes meet big data: Informing grazing management in a variable and changing world. Rangelands 2021, 43, 17–28. [Google Scholar] [CrossRef]
- Trotter, M.G.; Lamb, D.W.; Donald, G.E.; Schneider, D.A. Evaluating an active optical sensor for quantifying and mapping green herbage mass and growth in a perennial grass pasture. Crop Pasture Sci. 2010, 61, 389–398. [Google Scholar] [CrossRef]
- Punalekar, S.M.; Verhoef, A.; Quaife, T.L.; Humphries, D.; Bermingham, L.; Reynolds, C.K. Application of Sentinel-2A data for pasture biomass monitoring using a physically based radiative transfer model. Remote Sens. Environ. 2018, 218, 207–220. [Google Scholar] [CrossRef]
- Edirisinghe, A.; Clark, D.; Waugh, D. Spatio-temporal modelling of biomass of intensively grazed perennial dairy pastures using multispectral remote sensing. Int. J. Appl. Earth Obs. Geoinf. 2012, 16, 5–16. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, X.; Bajgain, R.; Starks, P.; Steiner, J.; Doughty, R.B.; Chang, Q. Estimating leaf area index and aboveground biomass of grazing pastures using Sentinel-1, Sentinel-2 and Landsat images. ISPRS J. Photogramm. Remote Sens. 2019, 154, 189–201. [Google Scholar] [CrossRef]
- Morais, T.G.; Teixeira, R.F.M.; Figueiredo, M.; Domingos, T. The use of machine learning methods to estimate aboveground biomass of grasslands: A review. Ecol. Indic. 2021, 130, 108081. [Google Scholar] [CrossRef]
- Harrison, M.T.; Roggero, P.P.; Zavattaro, L. Simple, efficient and robust techniques for automatic multi-objective function parameterisation: Case studies of local and global optimisation using APSIM. Environ. Model. Softw. 2019, 117, 109–133. [Google Scholar] [CrossRef]
- Ali, I.; Greifeneder, F.; Stamenkovic, J.; Neumann, M.; Notarnicola, C. Review of Machine Learning Approaches for Biomass and Soil Moisture Retrievals from Remote Sensing Data. Remote Sens. 2015, 7, 16398–16421. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gamon, J.A. The need for a common basis for defining light-use efficiency: Implications for productivity estimation. Remote Sens. Environ. 2015, 156, 196–201. [Google Scholar] [CrossRef]
- Delegido, J.; Verrelst, J.; Rivera, J.P.; Ruiz-Verdú, A.; Moreno, J. Brown and green LAI mapping through spectral indices. Int. J. Appl. Earth Obs. Geoinf. 2015, 35, 350–358. [Google Scholar] [CrossRef]
- Bsaibes, A.; Courault, D.; Baret, F.; Weiss, M.; Olioso, A.; Jacob, F.; Hagolle, O.; Marloie, O.; Bertrand, N.; Desfond, V.; et al. Albedo and LAI estimates from FORMOSAT-2 data for crop monitoring. Remote Sens. Environ. 2009, 113, 716–729. [Google Scholar] [CrossRef]
- De Rosa, D.; Basso, B.; Fasiolo, M.; Friedl, J.; Fulkerson, B.; Grace, P.R.; Rowlings, D.W. Predicting pasture biomass using a statistical model and machine learning algorithm implemented with remotely sensed imagery. Comput. Electron. Agric. 2021, 180. [Google Scholar] [CrossRef]
- Ibrahim, A.; Harrison, M.T.; Meinke, H.; Zhou, M. Examining the yield potential of barley near-isogenic lines using a genotype by environment by management analysis. Eur. J. Agron. 2019, 105, 41–51. [Google Scholar] [CrossRef]
- Zhai, Z.; Martínez, J.F.; Beltran, V.; Martínez, N.L. Decision support systems for agriculture 4.0: Survey and challenges. Comput. Electron. Agric. 2020, 170, 105256. [Google Scholar] [CrossRef]
- Ge, Y.; Thomasson, J.A.; Sui, R. Remote sensing of soil properties in precision agriculture: A review. Front. Earth Sci. 2011, 5, 229–238. [Google Scholar] [CrossRef]
- Bilotto, F.; Harrison, M.T.; Migliorati, M.D.A.; Christie, K.M.; Rowlings, D.W.; Grace, P.R.; Smith, A.P.; Rawnsley, R.P.; Thorburn, P.J.; Eckard, R.J. Can seasonal soil N mineralisation trends be leveraged to enhance pasture growth? Sci. Total Environ. 2021, 772, 145031. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Cawkwell, F.; Dwyer, E.; Barrett, B.; Green, S. Satellite remote sensing of grasslands: From observation to management. J. Plant Ecol. 2016, 9, 649–671. [Google Scholar] [CrossRef]
- Smith, R.C.G.; Adams, M.; Gittins, S.; Gherardi, S.; Wood, D.; Maier, S.; Stovold, R.; Donald, G.; Khohkar, S.; Allen, A. Near real-time Feed On Offer (FOO) from MODIS for early season grazing management of Mediterranean annual pastures. Int. J. Remote Sens. 2011, 32, 4445–4460. [Google Scholar] [CrossRef]
- Dingaan, M.N. V; Tsubo, M. Improved assessment of pasture availability in semi-arid grassland of South Africa. Environ. Monit. Assess. 2019, 191. [Google Scholar] [CrossRef] [PubMed]
- Guerschman, J.P.; Hill, M.J.; Renzullo, L.J.; Barrett, D.J.; Marks, A.S.; Botha, E.J. Estimating fractional cover of photosynthetic vegetation, non-photosynthetic vegetation and bare soil in the Australian tropical savanna region upscaling the EO-1 Hyperion and MODIS sensors. Remote Sens. Environ. 2009, 113, 928–945. [Google Scholar] [CrossRef]
- Zhang, B.; Carter, J. FORAGE – An online system for generating and delivering property-scale decision support information for grazing land and environmental management. Comput. Electron. Agric. 2018, 150, 302–311. [Google Scholar] [CrossRef]
- Dong, S.; Shang, Z.; Gao, J.; Boone, R.B. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 287. [Google Scholar] [CrossRef]
- Donnelly, J.R.; Moore, A.D.; Freer, M. GRAZPLAN: Decision support systems for Australian grazing enterprises—I. Overview of the GRAZPLAN project, and a description of the MetAccess and LambAlive DSS. Agric. Syst. 1997, 54, 57–76. [Google Scholar] [CrossRef]
- Srivastava, N.; Hinton, G.; Krizhevsky, A.; Sutskever, I.; Salakhutdinov, R. Dropout: A simple way to prevent neural networks from overfitting. J. Mach. Learn. Res. 2014, 15, 1929–1958. [Google Scholar]
- Verrelst, J.; Malenovský, Z.; Van der Tol, C.; Camps-Valls, G.; Gastellu-Etchegorry, J.-P.; Lewis, P.; North, P.; Moreno, J. Quantifying Vegetation Biophysical Variables from Imaging Spectroscopy Data: A Review on Retrieval Methods. Surv. Geophys. 2019, 40, 589–629. [Google Scholar] [CrossRef]
- CRCSI Earth Observation: Data, Processing and Applications. Volume 1A: Data—Basics and Acquisition; Melbourne, Australia, 2018.
- Scarth, P.; Armston, J.; Flood, N.; Denham, R.; Collett, L.; Watson, F.; Trevithick, B.; Muir, J.; Goodwin, N.; Tindalla, D.; et al. OPERATIONAL APPLICATION OF THE LANDSAT TIMESERIES TO ADDRESS LARGE AREA LANDCOVER UNDERSTANDING. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, XL-3-W3, 571–575. [CrossRef]
- Roberts, D.; Mueller, N.; Mcintyre, A. High-Dimensional Pixel Composites From Earth Observation Time Series. IEEE Trans. Geosci. Remote Sens. 2017, 55, 6254–6264. [Google Scholar] [CrossRef]
- BoM Australia Government. Bureau of meteorology. Available online: http://www.bom.gov.au/climate/averages/tables/cw_092027.shtml (accessed on Oct 25, 2022).
- Macquarie Franklin Okehampton-optimising management of production and biodiversity assets, Devonport TAS; 2019.
- Zhu, Z.; Wang, S.; Woodcock, C.E. Improvement and expansion of the Fmask algorithm: cloud, cloud shadow, and snow detection for Landsats 4–7, 8, and Sentinel 2 images. Remote Sens. Environ. 2015, 159, 269–277. [Google Scholar] [CrossRef]
- Piñeiro, G.; Perelman, S.; Guerschman, J.P.; Paruelo, J.M. How to evaluate models: Observed vs. predicted or predicted vs. observed? Ecol. Modell. 2008, 216, 316–322. [Google Scholar] [CrossRef]
- Zhu, Q.; Lin, H. Influences of soil, terrain, and crop growth on soil moisture variation from transect to farm scales. Geoderma 2011, 163, 45–54. [Google Scholar] [CrossRef]
- Warren, S.D.; Thurow, T.L.; Blackburn, W.H.; Garza, N.E. The influence of livestock trampling under intensive rotation grazing on soil hydrologic characteristics. Rangel. Ecol. Manag. Range Manag. Arch. 1986, 39, 491–495. [Google Scholar] [CrossRef]
- Khatri-Chhetri, U.; Thompson, K.A.; Quideau, S.A.; Boyce, M.S.; Chang, S.X.; Kaliaskar, D.; Bork, E.W.; Carlyle, C.N. Adaptive multi-paddock grazing increases soil nutrient availability and bacteria to fungi ratio in grassland soils. Appl. Soil Ecol. 2022, 179, 104590. [Google Scholar] [CrossRef]
- van Eekeren, N.; Jongejans, E.; van Agtmaal, M.; Guo, Y.; van der Velden, M.; Versteeg, C.; Siepel, H. Microarthropod communities and their ecosystem services restore when permanent grassland with mowing or low-intensity grazing is installed. Agric. Ecosyst. Environ. 2022, 323, 107682. [Google Scholar] [CrossRef]
- Zwerts, J.A.; Prins, H.H.T.; Bomhoff, D.; Verhagen, I.; Swart, J.M.; de Boer, W.F. Competition between a Lawn-Forming Cynodon dactylon and a Tufted Grass Species Hyparrhenia hirta on a South-African Dystrophic Savanna. PLoS One 2015, 10, e0140789. [Google Scholar] [CrossRef]
- Ivanov, V.Y.; Bras, R.L.; Vivoni, E.R. file:///C:/Users/mgo/Downloads/sustainability-v13-i22_20230426.bib. Water Resour. Res. 2008, 44. [Google Scholar] [CrossRef]
- Barrachina, M.; Cristóbal, J.; Tulla, A.F. Estimating above-ground biomass on mountain meadows and pastures through remote sensing. Int. J. Appl. Earth Obs. Geoinf. 2015, 38, 184–192. [Google Scholar] [CrossRef]
- Andresen, J.A.; Alagarswamy, G.; Rotz, C.A.; Ritchie, J.T.; LeBaron, A.W. Weather Impacts on Maize, Soybean, and Alfalfa Production in the Great Lakes Region, 1895–1996. Agron. J. 2001, 93, 1059–1070. [Google Scholar] [CrossRef]
- Edirisinghe, A.; Hill, M.J.; Donald, G.E.; Hyder, M. Quantitative mapping of pasture biomass using satellite imagery. Int. J. Remote Sens. 2011, 32, 2699–2724. [Google Scholar] [CrossRef]
- Myrgiotis, V.; Harris, P.; Revill, A.; Sint, H.; Williams, M. Inferring management and predicting sub-field scale C dynamics in UK grasslands using biogeochemical modelling and satellite-derived leaf area data. Agric. For. Meteorol. 2021, 307, 108466. [Google Scholar] [CrossRef]
- Moore, C.E.; Beringer, J.; Donohue, R.J.; Evans, B.; Exbrayat, J.-F.; Hutley, L.B.; Tapper, N.J. Seasonal, interannual and decadal drivers of tree and grass productivity in an Australian tropical savanna. Glob. Chang. Biol. 2018, 24, 2530–2544. [Google Scholar] [CrossRef]
- Segarra, J.; Buchaillot, M.L.; Araus, J.L.; Kefauver, S.C. Remote Sensing for Precision Agriculture: Sentinel-2 Improved Features and Applications. Agronomy 2020, 10. [Google Scholar] [CrossRef]
- Crabbe, R.A.; Lamb, D.W.; Edwards, C.; Andersson, K.; Schneider, D. A Preliminary Investigation of the Potential of Sentinel-1 Radar to Estimate Pasture Biomass in a Grazed Pasture Landscape. Remote Sens. 2019, 11. [Google Scholar] [CrossRef]
- Sadeh, Y.; Zhu, X.; Dunkerley, D.; Walker, J.P.; Zhang, Y.; Rozenstein, O.; Manivasagam, V.S.; Chenu, K. Fusion of Sentinel-2 and PlanetScope time-series data into daily 3 m surface reflectance and wheat LAI monitoring. Int. J. Appl. Earth Obs. Geoinf. 2021, 96, 102260. [Google Scholar] [CrossRef]
Figure 1.
The study site (a) land use for Tasmania, (b) farm property with 52 paddocks, and (c) subplots used for field sampling, [three larger plots (10 ha, 14 ha and 54 ha) were used as controls, while treatment plots have similar sizes from 0.2 -0.4 ha]. The first six plots were located on a paddock called “Bougainville” located on a hill. Land use data in (a) was obtained from the Australian Government, Department of Agriculture, Fisheries and Forestry, land use and management (accessed on 10 October 2022).
Figure 1.
The study site (a) land use for Tasmania, (b) farm property with 52 paddocks, and (c) subplots used for field sampling, [three larger plots (10 ha, 14 ha and 54 ha) were used as controls, while treatment plots have similar sizes from 0.2 -0.4 ha]. The first six plots were located on a paddock called “Bougainville” located on a hill. Land use data in (a) was obtained from the Australian Government, Department of Agriculture, Fisheries and Forestry, land use and management (accessed on 10 October 2022).
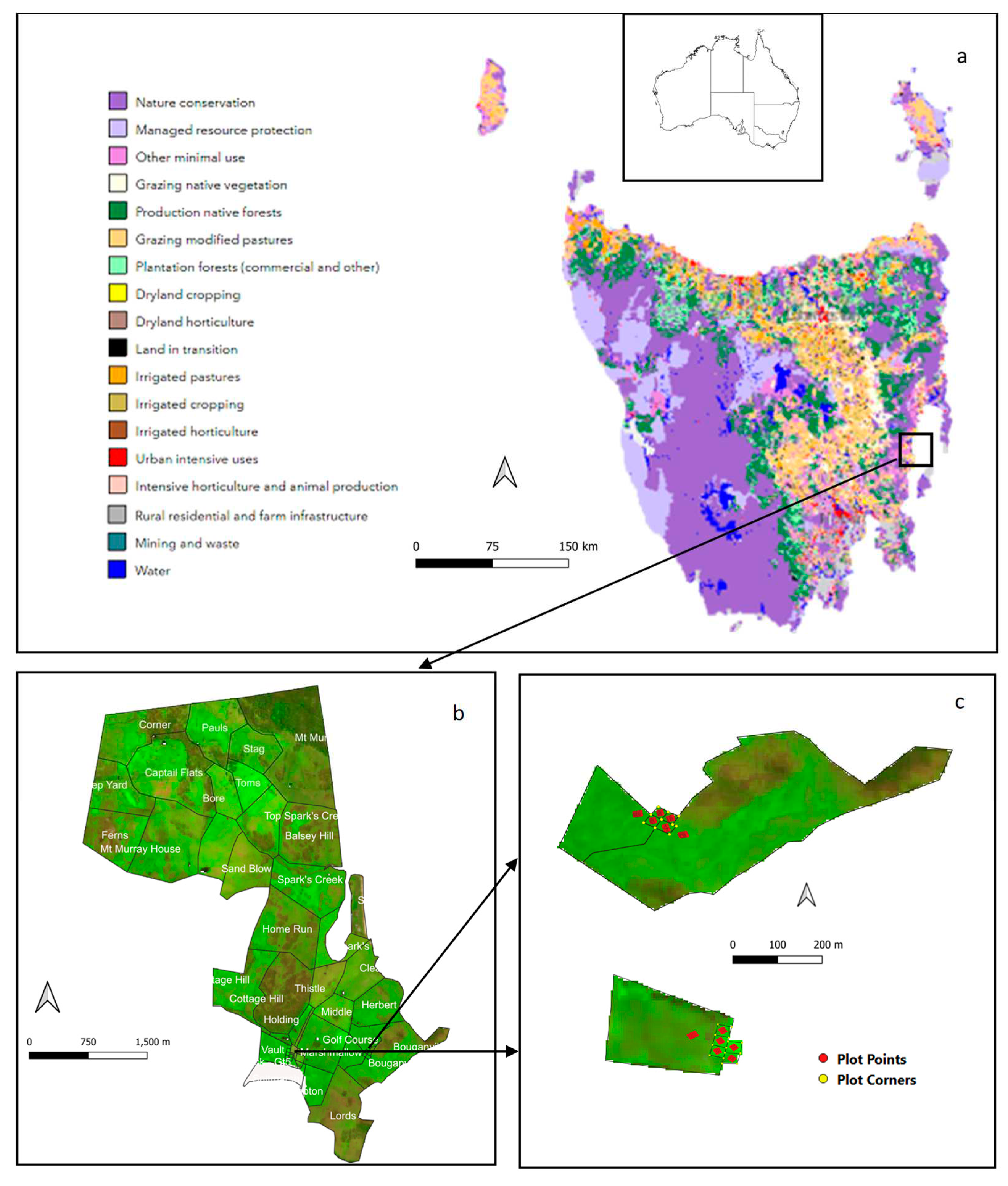
Figure 2.
Pasture biomass categories enumerated using destructive harvests at Okehampton, Tasmania, Australia. We measured (a) standing green biomass and (b) standing dry biomass prior to grazing; post-grazing we also measured (c) trampled green biomass and (d) trampled dry biomass. Photographs (a) and (b) were taken in autumn, (c) was taken in winter and (d) was taken in summer. We refer to the destructive sampling data herein as ‘measured’ data. Total standing dry matter (TSDM) was computed by the summation of green and dry standing biomass.
Figure 2.
Pasture biomass categories enumerated using destructive harvests at Okehampton, Tasmania, Australia. We measured (a) standing green biomass and (b) standing dry biomass prior to grazing; post-grazing we also measured (c) trampled green biomass and (d) trampled dry biomass. Photographs (a) and (b) were taken in autumn, (c) was taken in winter and (d) was taken in summer. We refer to the destructive sampling data herein as ‘measured’ data. Total standing dry matter (TSDM) was computed by the summation of green and dry standing biomass.

Figure 3.
Effects of grazing treatments on total standing dry matter (TSDM), computed as the sum of standing green DM, standing dry DM and trampled residual. The coloured small, dotted points show measurements obtained from five quadrats in each treatment plot; the large dot shows the mean for each plot.
Figure 3.
Effects of grazing treatments on total standing dry matter (TSDM), computed as the sum of standing green DM, standing dry DM and trampled residual. The coloured small, dotted points show measurements obtained from five quadrats in each treatment plot; the large dot shows the mean for each plot.
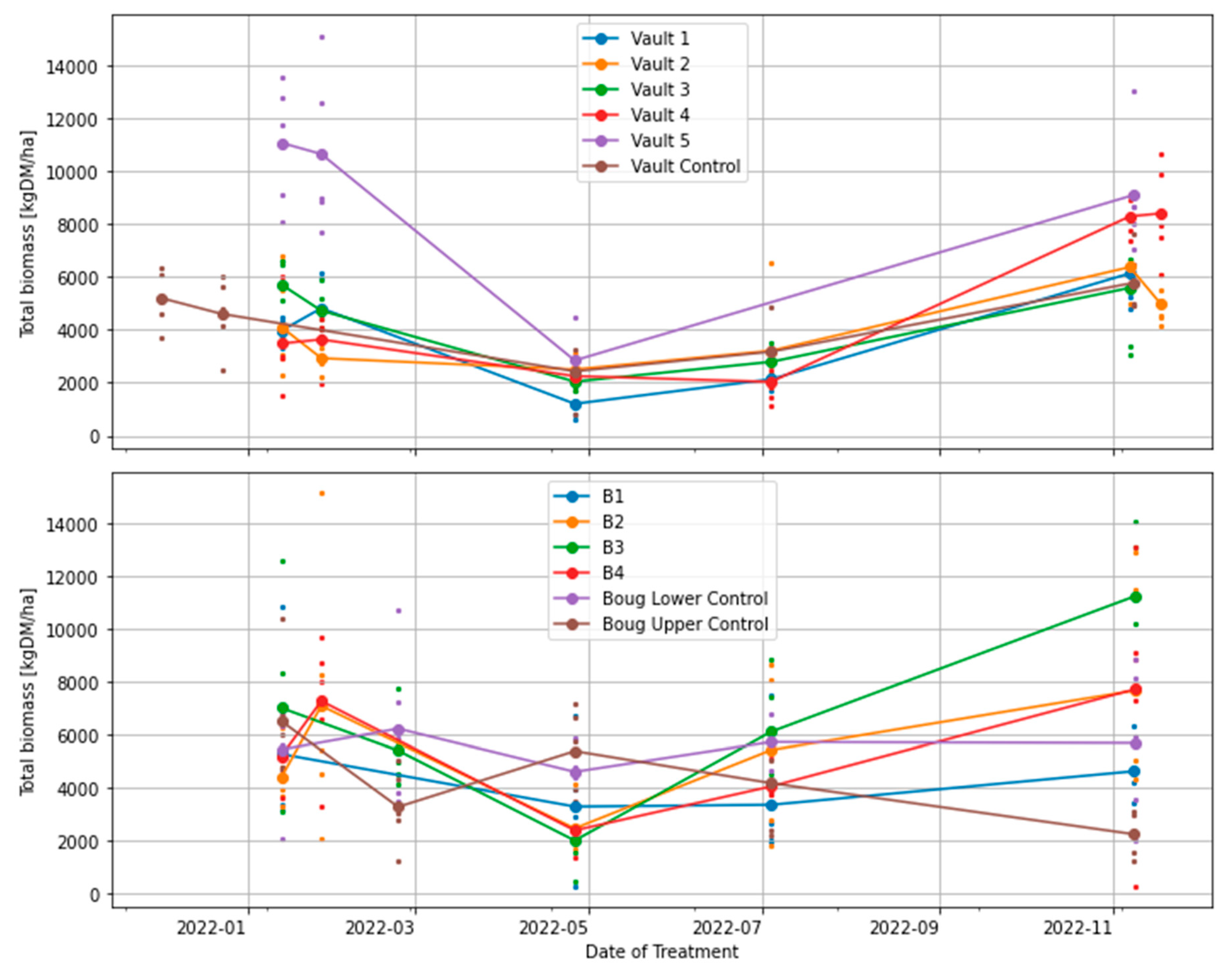
Figure 4.
Effects of treatments on the total standing dry matter. TSDM is computed as the summation of standing green DM and standing dry DM excluding the trampled residual.
Figure 4.
Effects of treatments on the total standing dry matter. TSDM is computed as the summation of standing green DM and standing dry DM excluding the trampled residual.
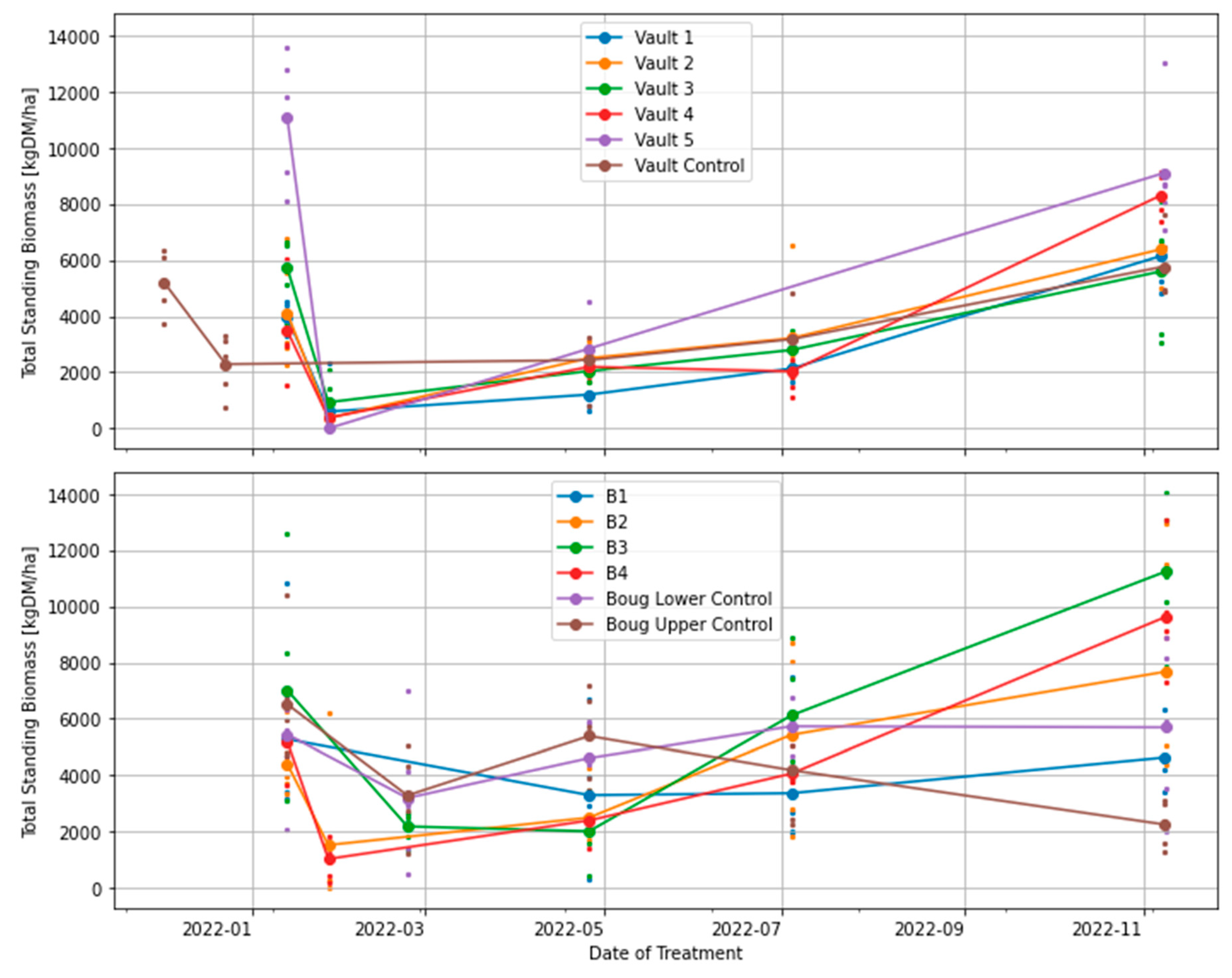
Figure 5.
Effects of treatment on the standing green biomass.
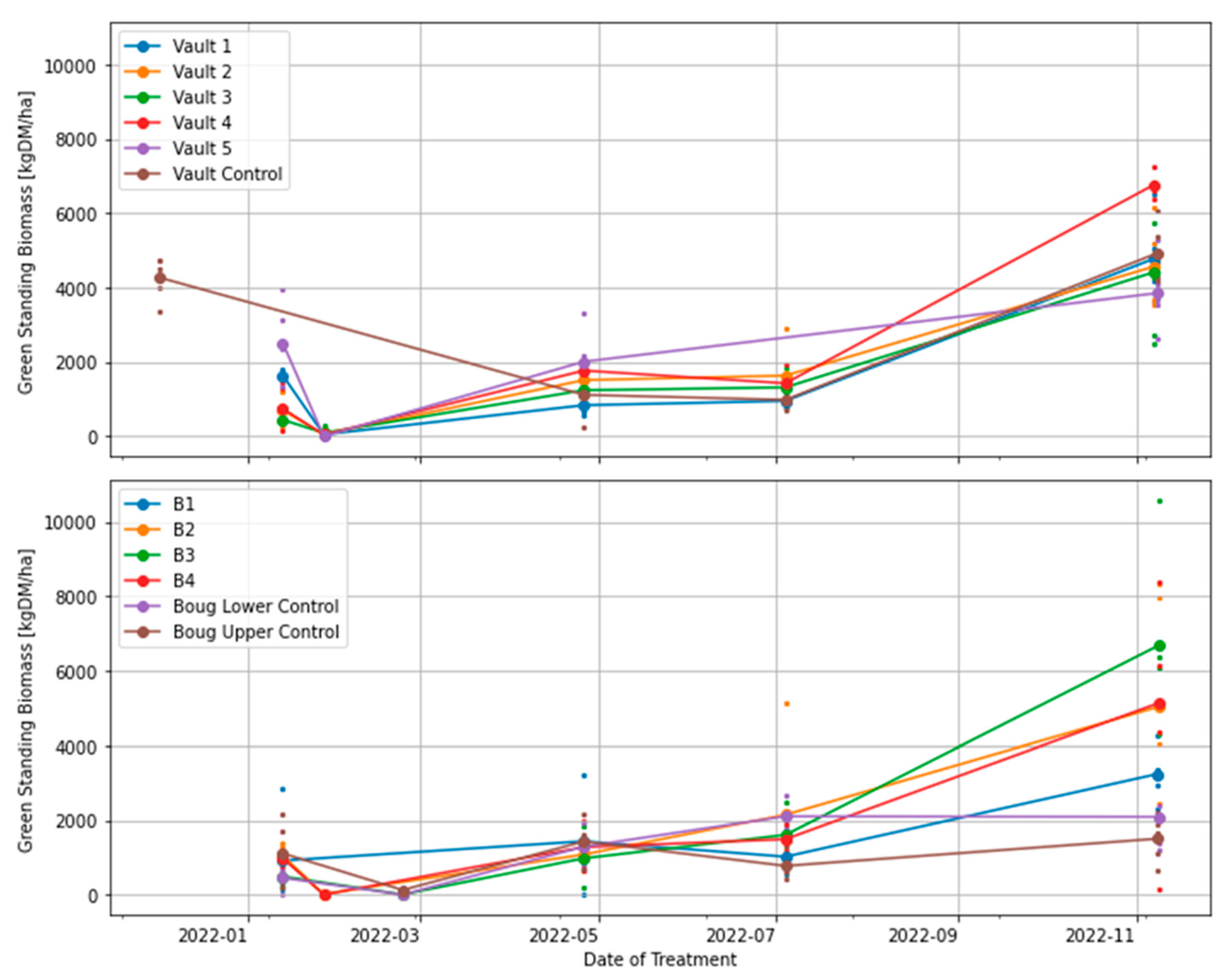
Figure 6.
Effects of treatment on the standing dry biomass.
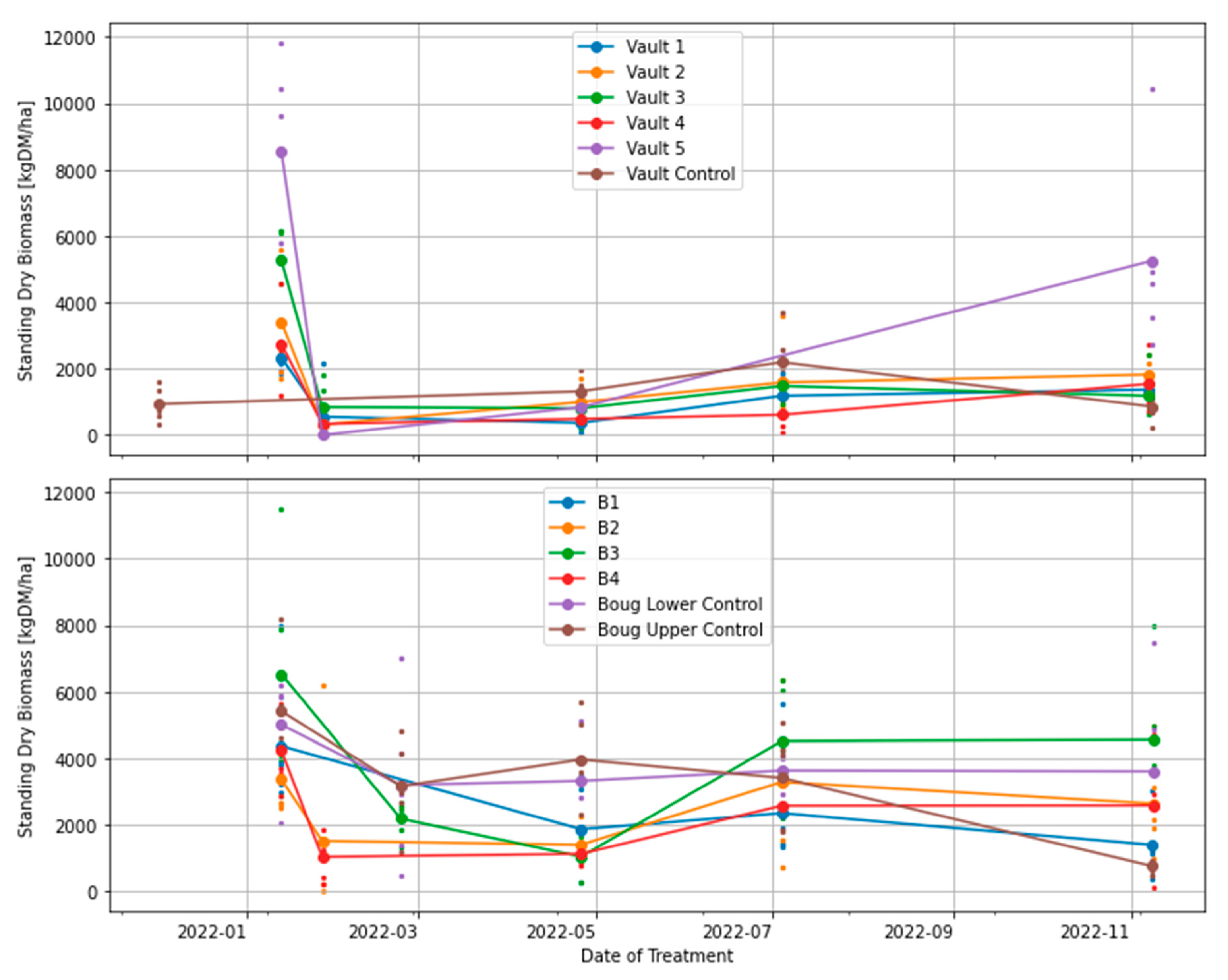
Figure 7.
Measured and estimated TSDM data at Okehampton, Triabunna, Tasmania. Trampled material is vegetation that was pushed against the ground by grazing which was measured in the phase 1 (Table 1) after post-grazing. The broken lines represent the measured TSDM while the blue solid line represents Cibo Labs estimated TSDM. There was only one day of grazing in the treatment plots and longer period in the control paddocks (Table 1). Bougainville 1 and 3 treatment plots were grazed as BUA at the start of the experiment and closed off. Each error bar symbolises measured data point equivalents to Cibo Labs estimate.
Figure 7.
Measured and estimated TSDM data at Okehampton, Triabunna, Tasmania. Trampled material is vegetation that was pushed against the ground by grazing which was measured in the phase 1 (Table 1) after post-grazing. The broken lines represent the measured TSDM while the blue solid line represents Cibo Labs estimated TSDM. There was only one day of grazing in the treatment plots and longer period in the control paddocks (Table 1). Bougainville 1 and 3 treatment plots were grazed as BUA at the start of the experiment and closed off. Each error bar symbolises measured data point equivalents to Cibo Labs estimate.
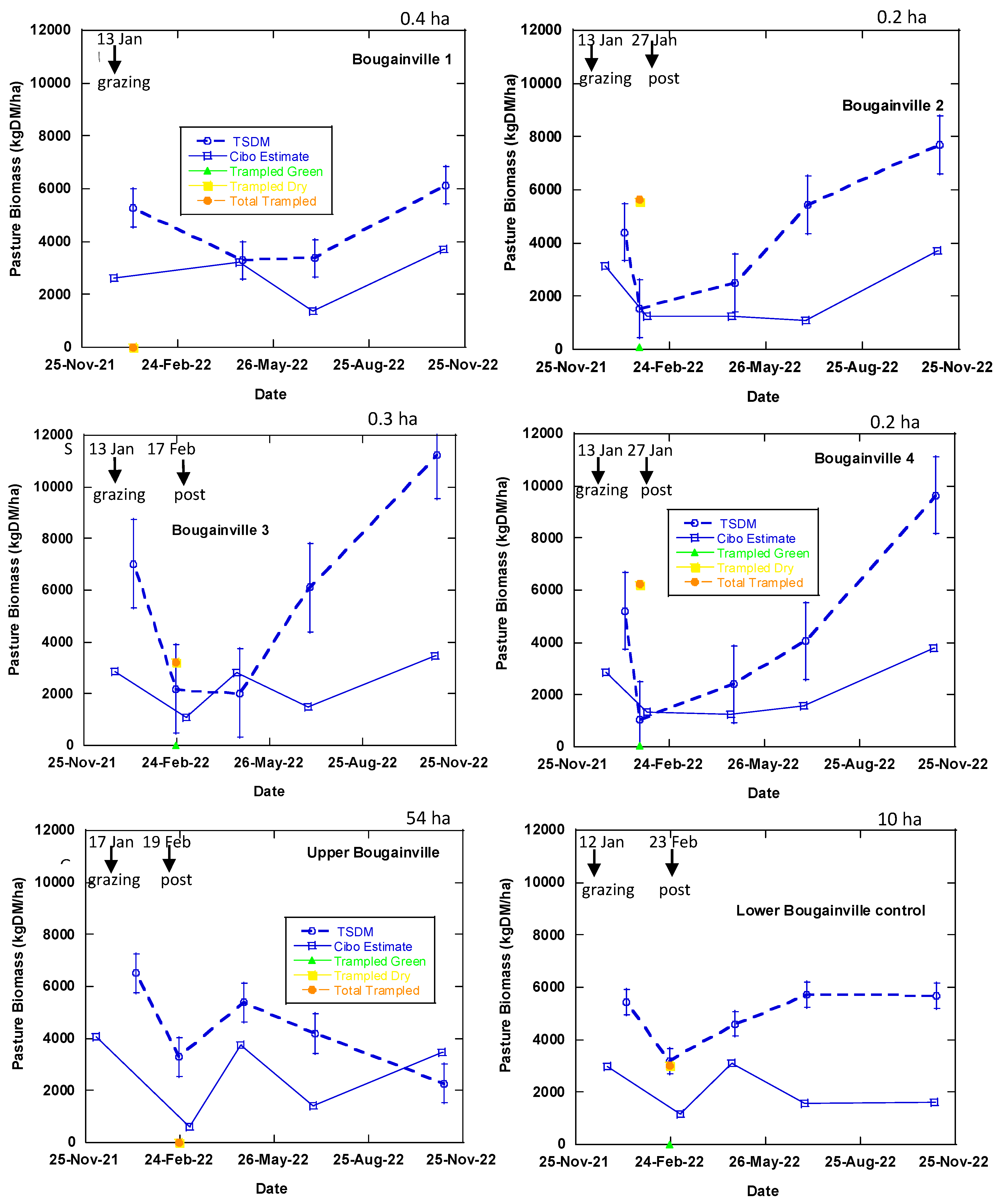
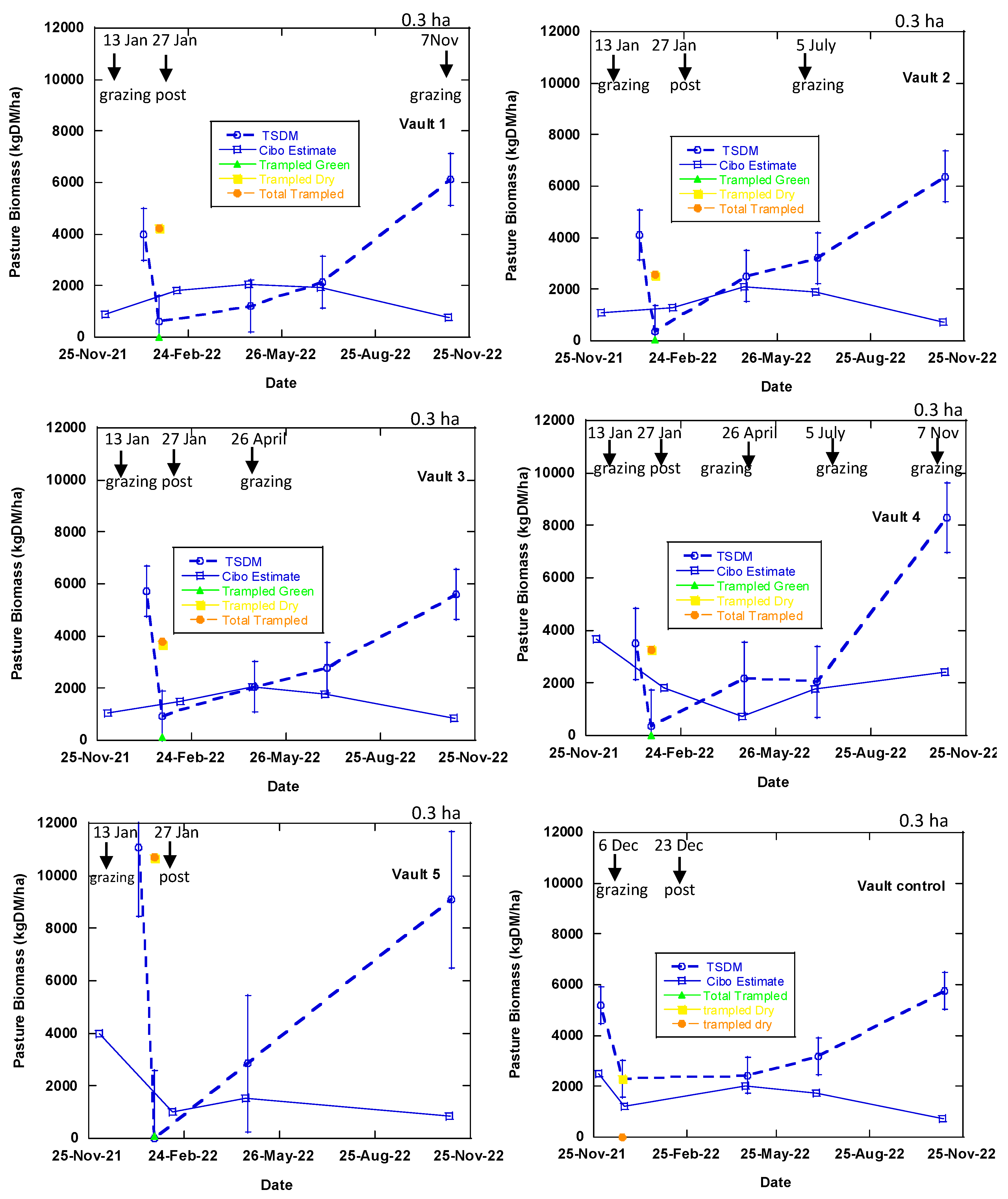
Figure 8.
Measured standing green and dry pasture biomass compared with the Cibo Labs estimates. The broken lines represent the measured green DM and dry DM while the blue solid line represents Cibo Labs estimated green DM and dry DM.
Figure 8.
Measured standing green and dry pasture biomass compared with the Cibo Labs estimates. The broken lines represent the measured green DM and dry DM while the blue solid line represents Cibo Labs estimated green DM and dry DM.
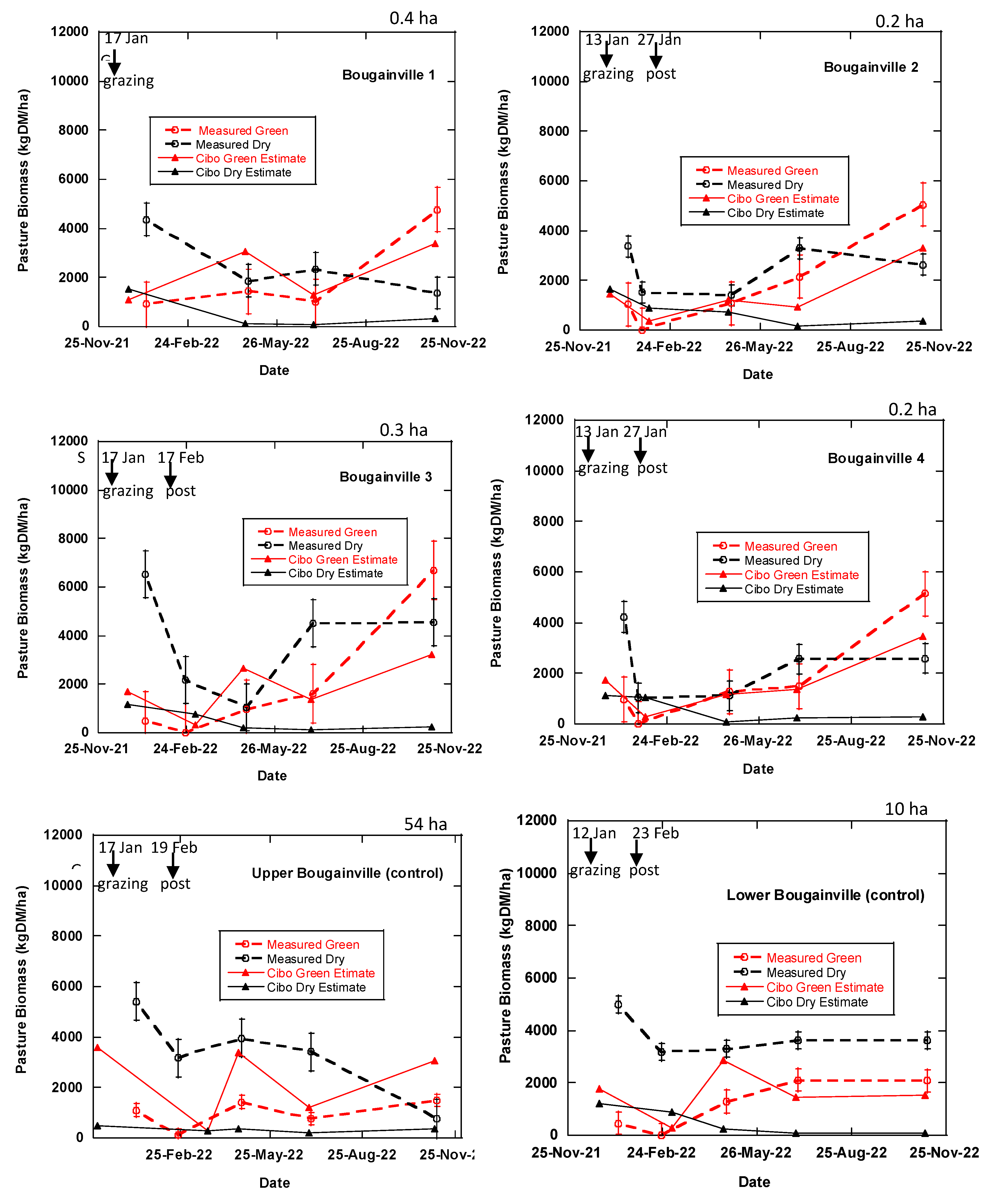
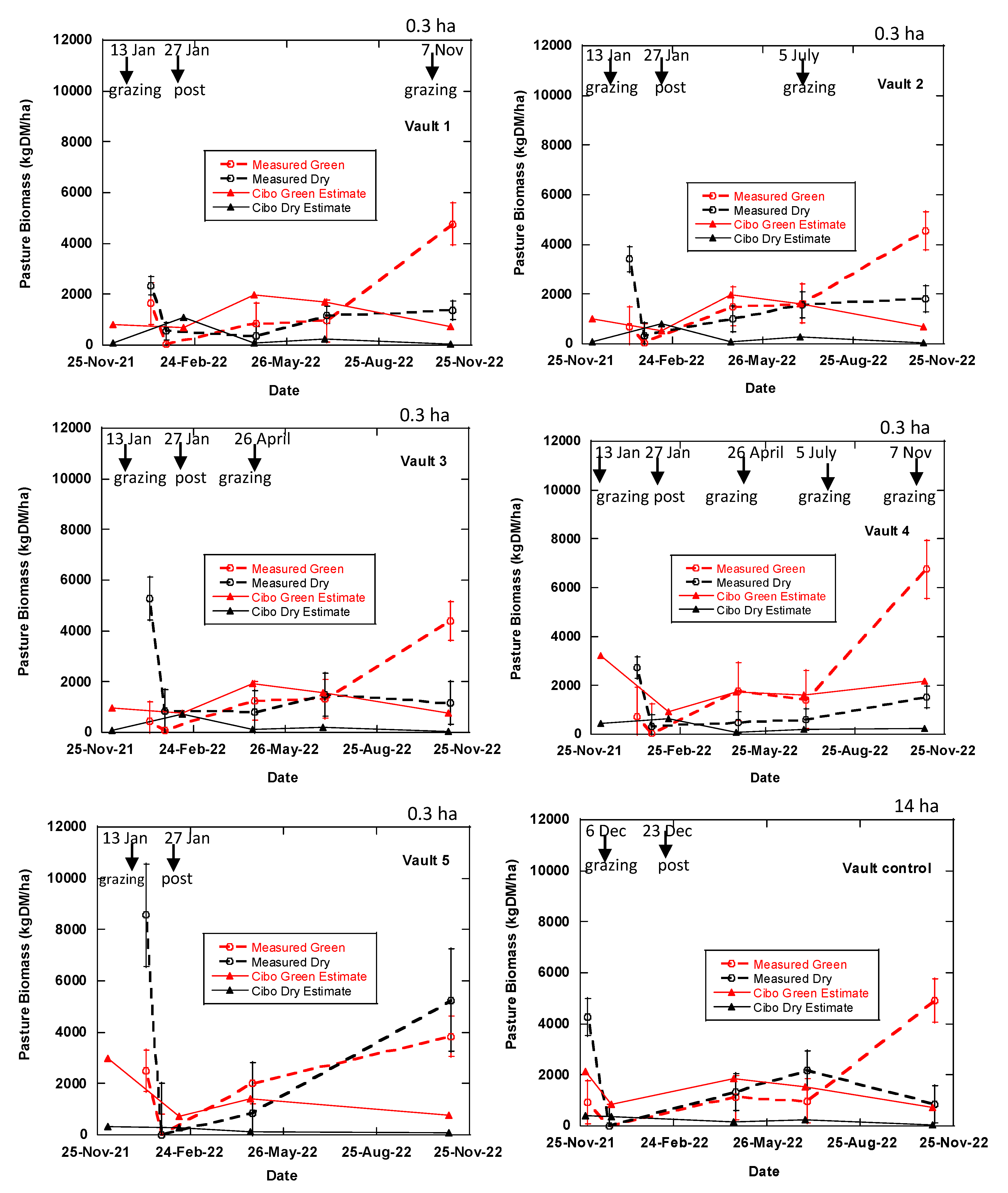
Figure 9.
Relationship between the measured total standing dry matter and Cibo Labs estimate. The equality line shows that estimates from Sentinel-2 integrated into the Cibo Lab model underestimated TSDM compared to the measured TSDM with MAE of 745 kg DM/ha and RMSE of 903 kg DM/ha.
Figure 9.
Relationship between the measured total standing dry matter and Cibo Labs estimate. The equality line shows that estimates from Sentinel-2 integrated into the Cibo Lab model underestimated TSDM compared to the measured TSDM with MAE of 745 kg DM/ha and RMSE of 903 kg DM/ha.
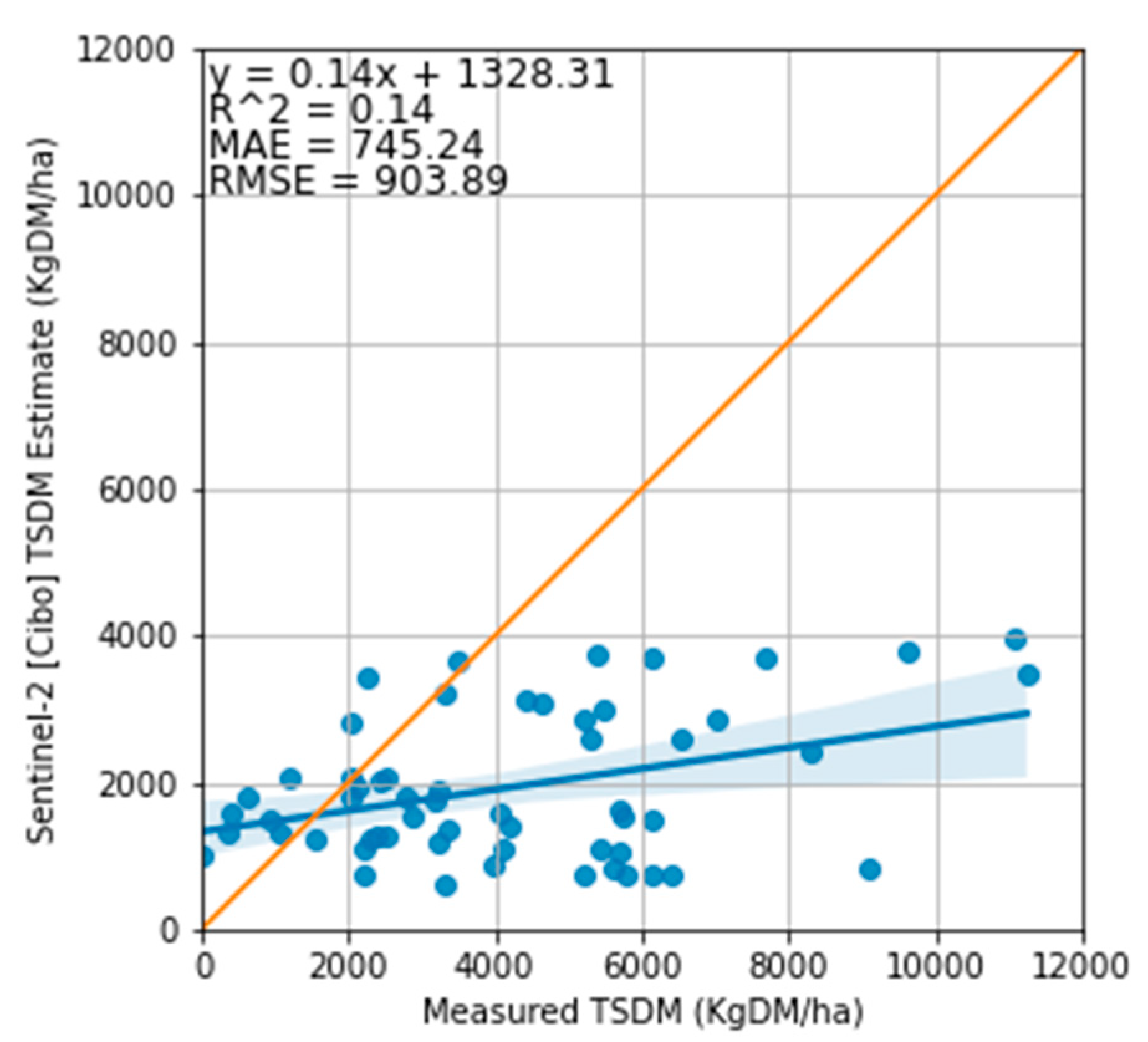
Figure 10.
Relationship between the measured standing green and Cibo Labs estimate. The regression 1:1 line shows that satellite estimates overestimated the standing green DM compared to the measured with MAE of 702 kg DM/ha and RMSE of 880 Kg DM/ha.
Figure 10.
Relationship between the measured standing green and Cibo Labs estimate. The regression 1:1 line shows that satellite estimates overestimated the standing green DM compared to the measured with MAE of 702 kg DM/ha and RMSE of 880 Kg DM/ha.
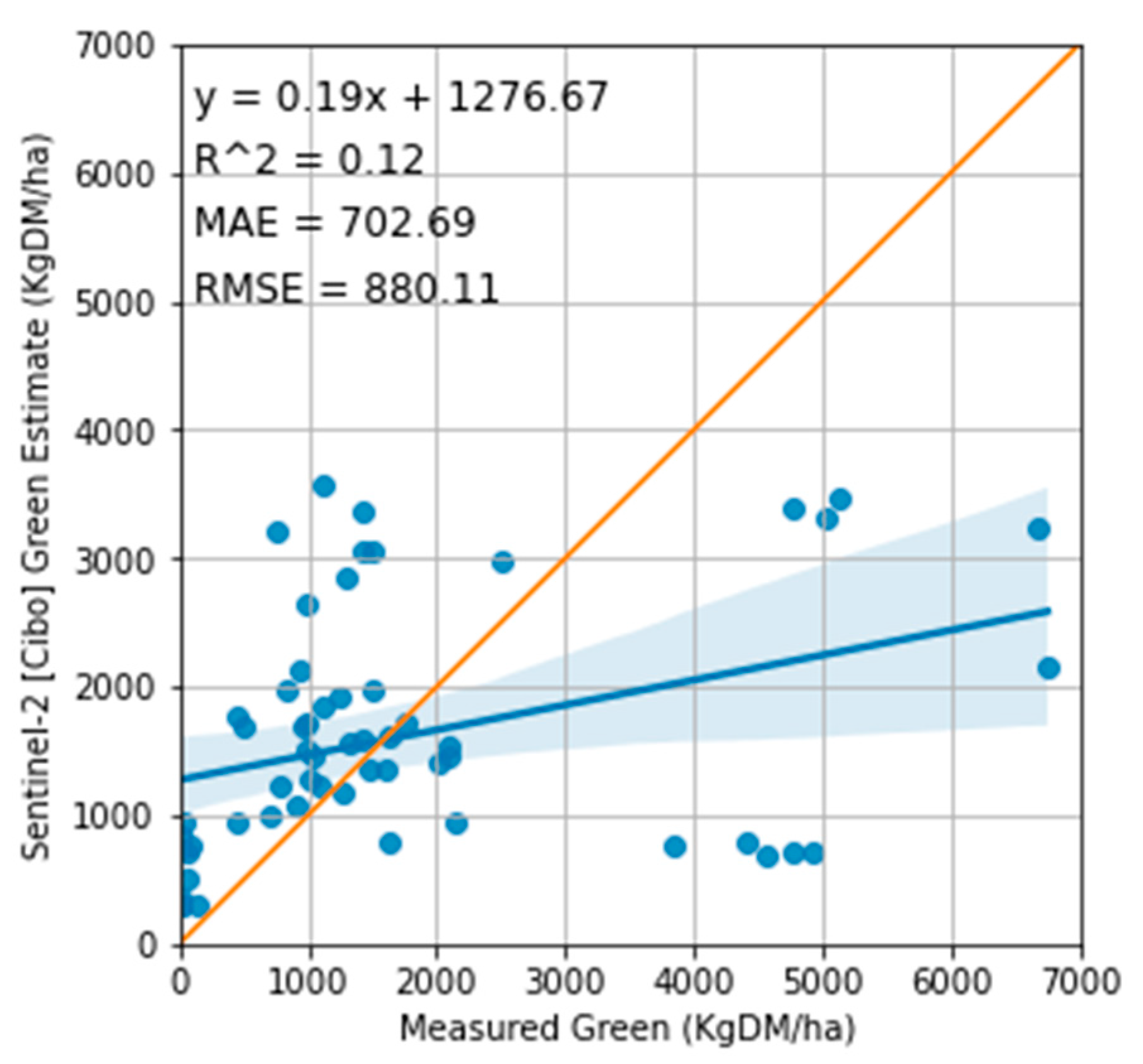
Figure 11.
Relationship between the measured standing dry and Cibo Labs estimate. Although the relationship between the estimated standing dry DM and measured TSDM shows underestimation, it thus lowers the MAE (297 kg DM/ha) and RMSE (388 kg DM/ha).
Figure 11.
Relationship between the measured standing dry and Cibo Labs estimate. Although the relationship between the estimated standing dry DM and measured TSDM shows underestimation, it thus lowers the MAE (297 kg DM/ha) and RMSE (388 kg DM/ha).
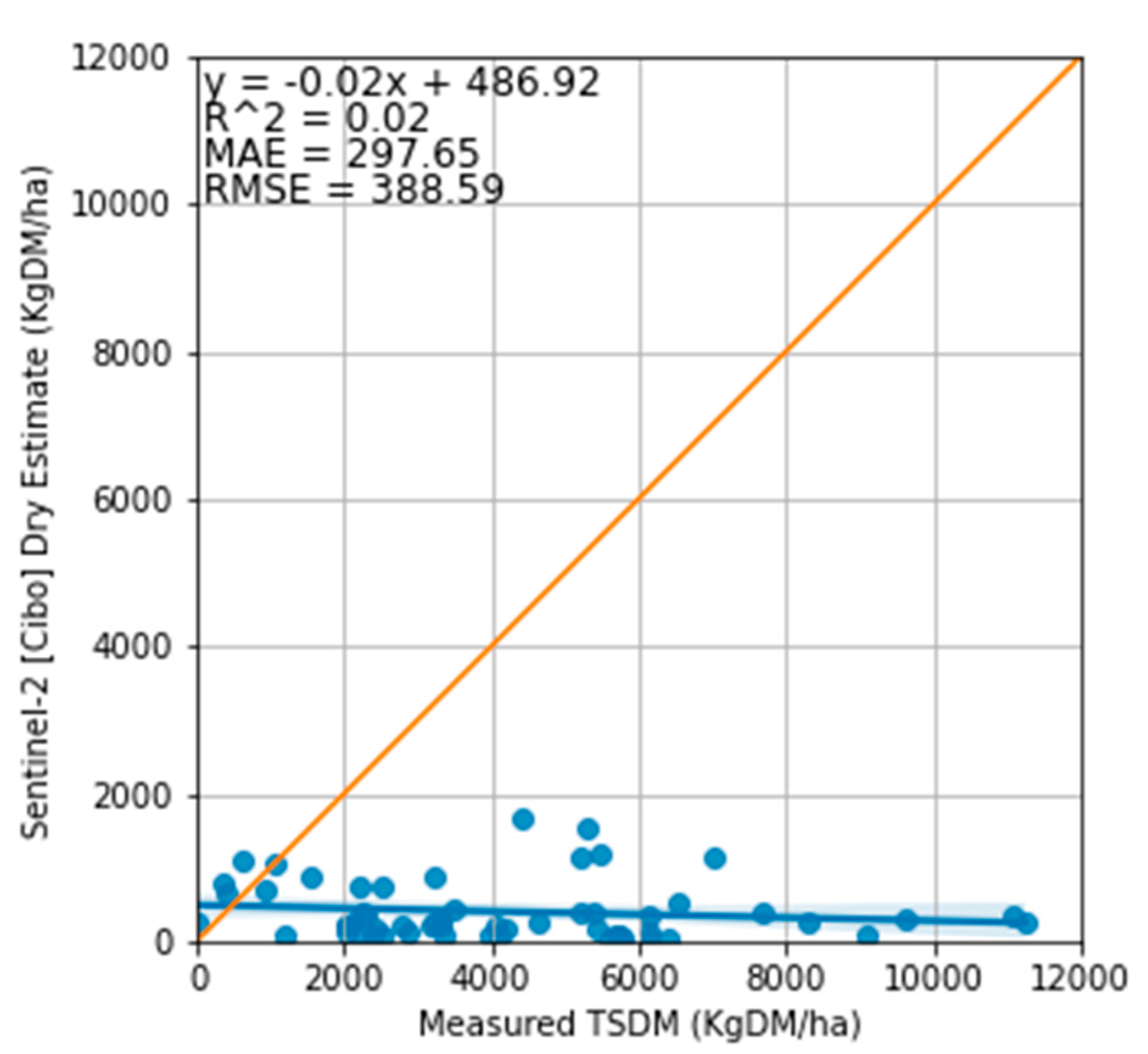
Figure 12.
Relationship between the measured total standing dry matter and Cibo Labs standing green estimate. The regression 1:1 line shows that a few estimated standing green DM from the Cibo Lab model were higher than the measured TSDM with the MAE of 685 kg DM/ha and RMSE of 850 kg DM/ha.
Figure 12.
Relationship between the measured total standing dry matter and Cibo Labs standing green estimate. The regression 1:1 line shows that a few estimated standing green DM from the Cibo Lab model were higher than the measured TSDM with the MAE of 685 kg DM/ha and RMSE of 850 kg DM/ha.
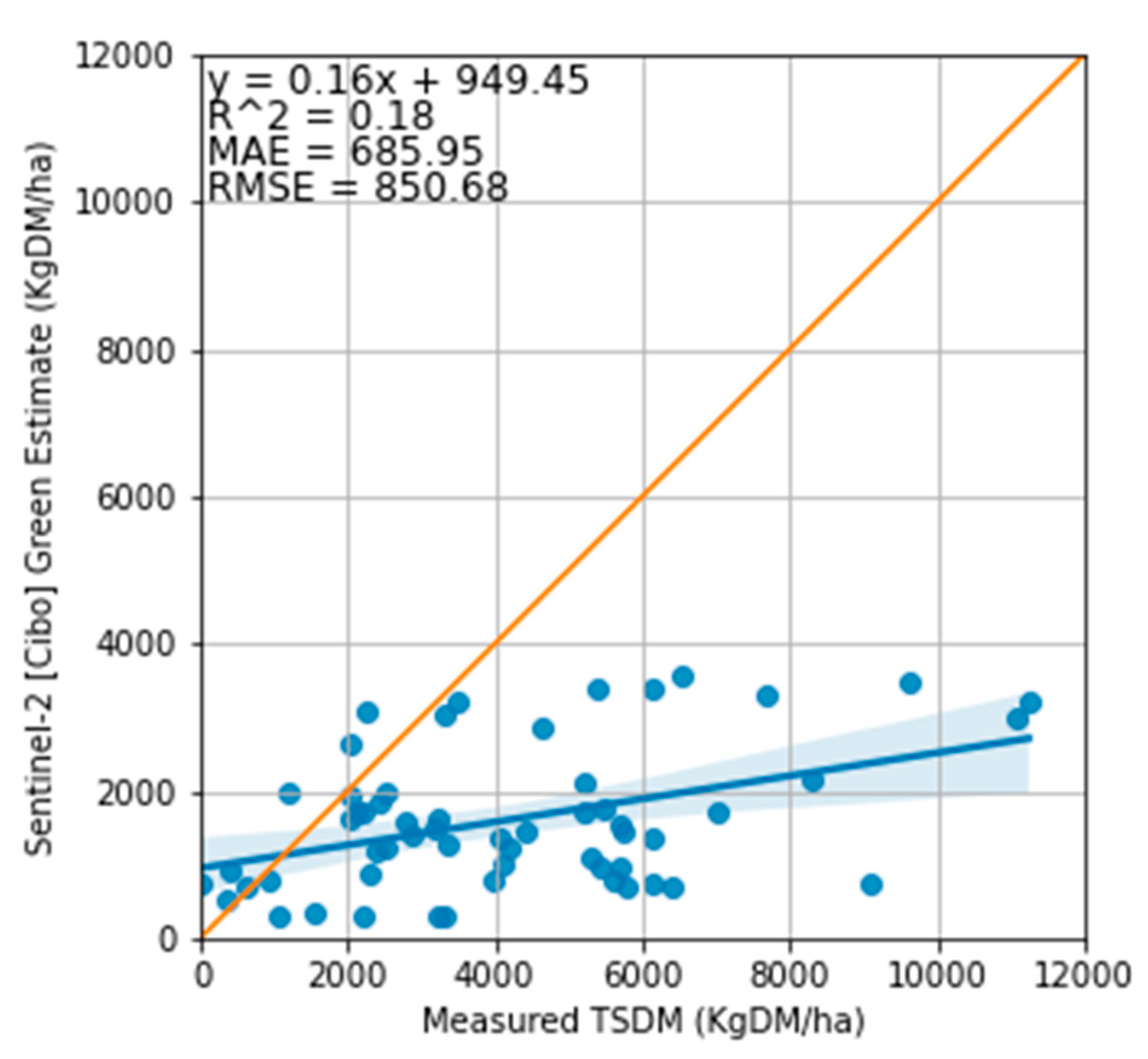
Figure 13.
Relationship between the measured total standing dry matter and Cibo Labs standing dry. The relationship shows a high concentration of Cibo Labs estimates barely above the ground level with the MAE of 302 kg DM/ha and RMSE of 385 kg DM/ha.
Figure 13.
Relationship between the measured total standing dry matter and Cibo Labs standing dry. The relationship shows a high concentration of Cibo Labs estimates barely above the ground level with the MAE of 302 kg DM/ha and RMSE of 385 kg DM/ha.
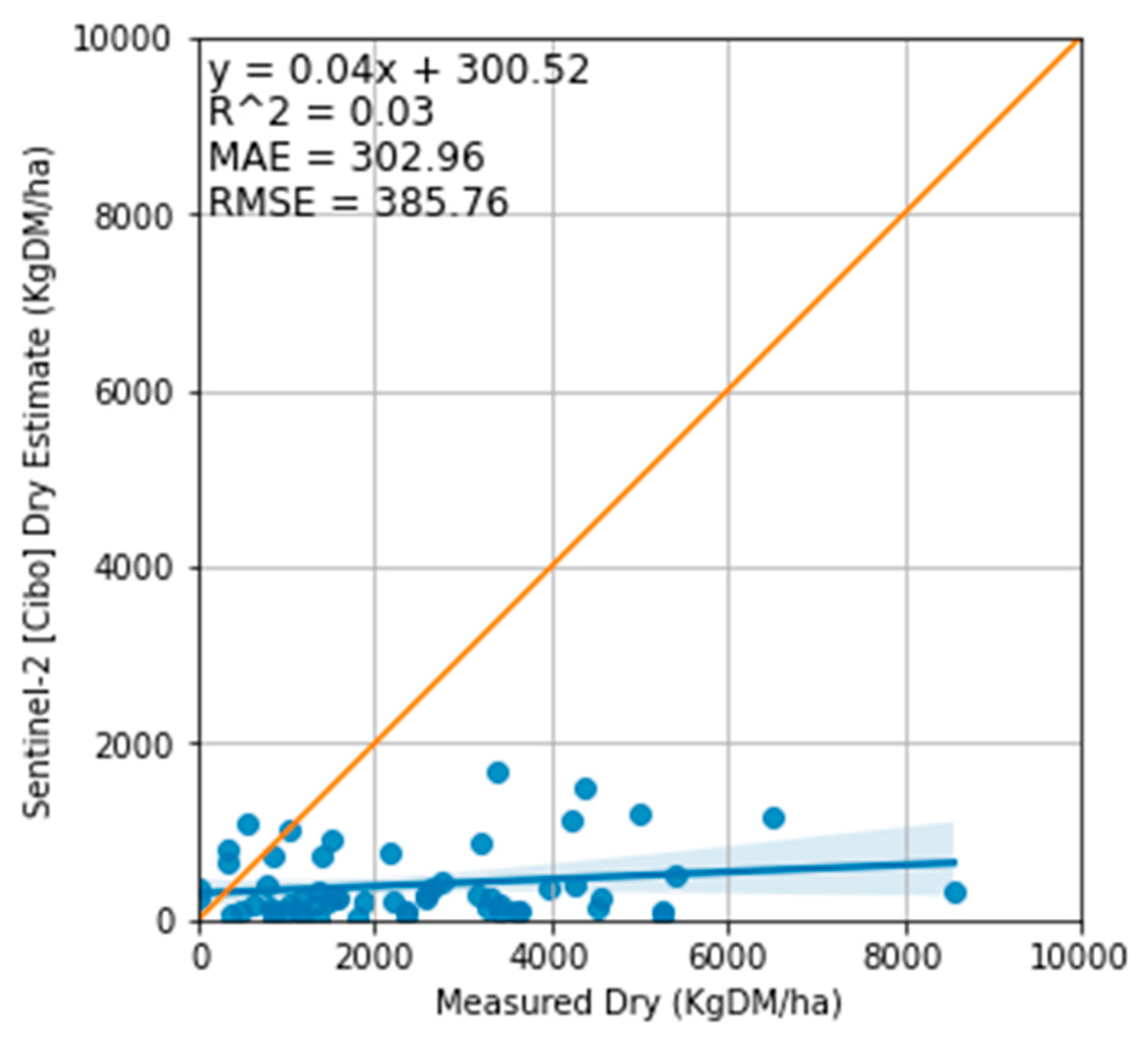
Figure 14.
Spatiotemporal variation in pasture biomass for all treatment plots. The smaller plots (expanded for better view) represent the regenerative grazing treatments while the bigger serve as the conventional (business-as-usual) grazing treatments. The Vaults (1 to 5) plots are at the lower-left expanded view of the panel, while Bougainville (1 to 4) plots are at the upper-right expanded view.
Figure 14.
Spatiotemporal variation in pasture biomass for all treatment plots. The smaller plots (expanded for better view) represent the regenerative grazing treatments while the bigger serve as the conventional (business-as-usual) grazing treatments. The Vaults (1 to 5) plots are at the lower-left expanded view of the panel, while Bougainville (1 to 4) plots are at the upper-right expanded view.
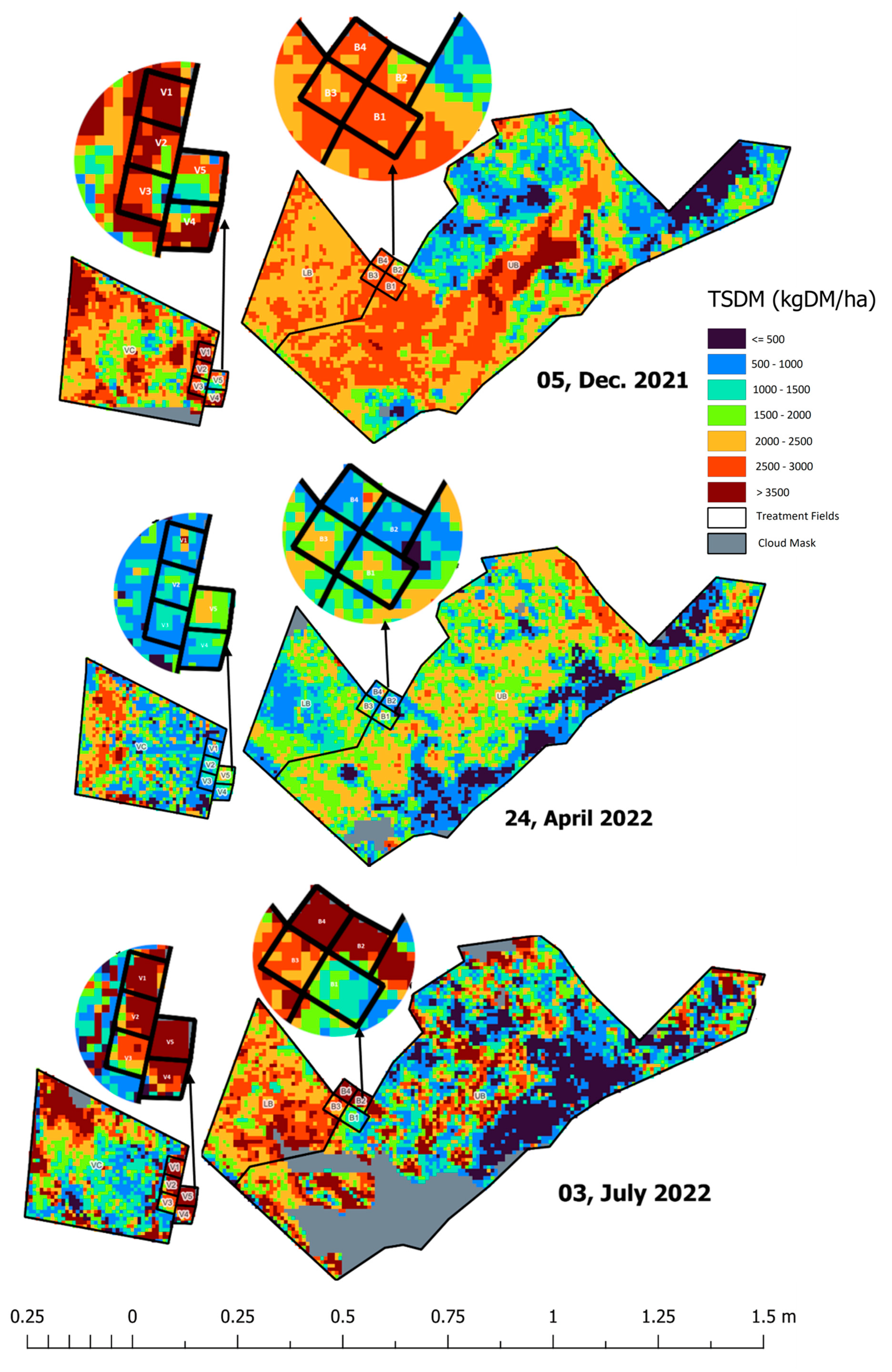

Table 1.
Experimental treatments and business-as-usual plots (controls). All plots were sampled and grazed in phase 1. Trampled residual was collected only for post-grazing. However, the Bougainville 1, 2, 3, and 4 plots have treatment plans different from the Vault treatments. In the beginning of the study, Bougainville 2 and 4 plots were subjected to intense grazing, similar to the Vault treatments, whereas Bougainville 1 and 3 plots were grazed in accordance with BUA. After phase 1, all four plots were closed and left ungrazed; however, the total standing dry matter was still measured every three months for all plots, regardless of grazing, to facilitate comparison. By examining the marked regions on the chart, you can determine when grazing occurred.
Table 1.
Experimental treatments and business-as-usual plots (controls). All plots were sampled and grazed in phase 1. Trampled residual was collected only for post-grazing. However, the Bougainville 1, 2, 3, and 4 plots have treatment plans different from the Vault treatments. In the beginning of the study, Bougainville 2 and 4 plots were subjected to intense grazing, similar to the Vault treatments, whereas Bougainville 1 and 3 plots were grazed in accordance with BUA. After phase 1, all four plots were closed and left ungrazed; however, the total standing dry matter was still measured every three months for all plots, regardless of grazing, to facilitate comparison. By examining the marked regions on the chart, you can determine when grazing occurred.
| Treatments | Plot | Phase 1 | Phase 2 | Phase 3 | Phase 4 | ||
| Size (ha) | Dec 2021 & Jan 2022 | Apr-22 | Jul-22 | Nov-22 | |||
| Grazing | Pre | Post | pre | pre | pre | ||
| Trampled after post-grazing | ✓ | V4 | |||||
| Stocking rate (DSE/ha) | 8000 | 6000 | 8800 | 8800 | |||
| BUA & Regenerative | Bougainville 1 (B1) | 0.4 | ✓ | ✓ | |||
| Regenerative | Bougainville (B2) | 0.2 | ✓ | ✓ | |||
| BUA & Regenerative | Bougainville (B3) | 0.3 | ✓ | ✓ | |||
| Regenerative | Bougainville (B4) | 0.2 | ✓ | ✓ | |||
| Control | Upper Bougainville (UB) | 54 | Business as usual | ||||
| Control | Lower Bougainville (LB) | 10 | Business as usual | ||||
| Regenerative | Vault 1 [12 months] | 0.3 | ✓ | ✓ | ✓ | ||
| Vault 2 [9 months] | 0.3 | ✓ | ✓ | ✓ | |||
| Vault 3 [6 months] | 0.3 | ✓ | ✓ | ✓ | |||
| Vault 4 [3 months] | 0.3 | ✓ | ✓ | ✓ | ✓ | ✓ | |
| Vault 5 [15 months] | 0.3 | ✓ | ✓ | ||||
| Control | Vault Control | 14 | Business as usual | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



