Submitted:
27 April 2023
Posted:
28 April 2023
You are already at the latest version
Abstract
The addition of mineral-solubilizing microbial inoculums is a kind of biological measure for vegetation restoration of rock mining areas. Its function is to accelerate soil weathering, improve soil fertility, improve the ability of plants to fix soil. Through understanding the response of plant rhizosphere microbial community to the mineral-solubilizing microbial inoculums, it is helpful to popularize the use of mineral-solubilizing microbial inoculums. However, little is known about the changes of plant rhizosphere soil microbial communities after the addition of mineral-solubilizing microbial inoculums. Therefore, the purpose of this study was to reveal the pasthways through which different mineral-solubilizing microbial inoculums positively affect underground part of R. pseudoacacia. A pot experiment was conducted to investigate the responses of rhizosphere soil bacterial and fungal communities in R. pseudoacacia by taking 32 samples from four different mineral-solubilizing microbial inoculums treatments. The results showed that the effect of mineral-solubilizing microbial inoculums on the structure of fungal community was greater than that of bacterial community. But the relative abundance of Proteobacteria was increased, which had a strong positive correlation with root nodulation. In terms of microbial diversity, mineral-solubilizing microbial inoculums had a greater effect on the diversity and evenness of bacterial community. It is worth noting that correlation analysis showed that Proteobacteria and Verrucomicrobia in bacteria and Ascomycota and Zoopagomycota in fungi were positively correlated with soil enzyme activity and plant growth. RDA analysis showed that the relative abundance of these two phyla in bacteria also had positive effects on plant root nodulation. Our results showed that the addition of mineral-solubilizing microbial inoculums can optimize the rhizosphere soil microbial community structure, promote R. pseudoacacia root nodulation, and enhance the nitrogen fixation capacity of plants. In addition, this study can provide a theoretical basis for the application of mineral-solubilizing microbial inoculums to a wide range of slope ecological restoration.
Keywords:
Bacterial communities
; Fungal communities
; Mineral-solubilizing microbial inoculums
; Robinia pseudoacacia L.
1. Introduction
Limestone is a crucial industrial raw material used in metallurgy, construction, chemical, light, and agricultural industries, as well as other specialized sectors [1,2]. As the steel and cement industries continue to develop, the importance of limestone is bound to increase [3,4]. However, a number of environmental issues that have a significant negative influence on land degradation and ecological danger might come from the irrational development and use of mineral resources, which can destroy ecosystems. [5,6]. To mitigate these issues, bioremediation measures such as mineral-solubilizing microbial inoculums have been developed to accelerate rock weathering [7-9]. Numerous studies have demonstrated that the inoculums can effectively promote plant growth, improve soil nutrient supply capacity, increase plant photosynthetic rate, enhance plant root tension, and improve plant nodulation [10-14]. As an environmentally friendly bioremediation measure, mineral-solubilizing microbial inoculums have wide-ranging applications and promising prospects.
The complex biochemical processes between microorganisms and plants achieve the dynamic balance of nutrient supply [15,16]. Although it is generally recognized that rhizosphere bacteria are crucial for plant health and nutrient uptake[17], our knowledge of the intricate plant-microbe interactions in the rhizosphere is still in its infancy. Plant rhizosphere soil microorganisms occupy the transition zone between the soil and root system, and they are the primary site of nutrient exchange. Therefore, the addition of mineral-solubilizing microbial inoculums will have a direct impact on the rhizosphere environment. R. pseudoacacia, which has a strong resistance to stress and a developed root system, is often used as a pioneer tree for soil consolidation and fertilizer conservation in slope ecological restoration [18]. Currently, research mainly focuses on the soil consolidation of R. pseudoacacia root systems and the growth of R. pseudoacacia as a building species on different slope sites [13]. However, exogenous microorganisms may augment, diminish, or have no effect on the native microbial activities. In order to comprehend rhizosphere nutrient dynamics, it is vital to understand how the structure of the soil’s microbial community responds to the addition of mineral-solubilizing microbial inoculums.
The goals of this study were to: (i) evaluate the alterations in the bacterial and fungal community and structure in the rhizosphere soil of R. pseudoacacia under various mineral-solubilizing inoculum treatments; (ii) examine potential mechanisms by which mineral-solubilizing microbial inoculums may change microbial communities and functions to affect plant growth during R. pseudoacacia development; and (iii) carry out a niche analysis of mineral- Our investigation can act as a blueprint for additional study targeted at effectively extending the use of soil spray-sowing technology. In addition, it offers a fundamental theoretical framework for the global ecological rehabilitation of mining regions.
2. Materials and methods
2.1. Study area
At Nanjing Forestry University, the pot experiments were carried out in an intelligent greenhouse where relative humidity and maximum photosynthetic radiation were managed. The pot water content was measured every other day and watered quantitatively to ensure the consistency of soil water content in the pot experiments.
2.2. Seed test material, microbial strains, and soil
The experimental Robinia pseudoacacia L seeds are provided by Tianhe nursery garden company in Jiangsu, China. After germination is promoted, the seedlings are cultured together in the seedling substrate. R. pseudoacacia is a pioneer tree species often used for ecological restoration of slopes. These plants were chosen for this study so that we could examine the impact of growth substrates and mineral-solubilizing microorganisms on the soil microbial population.
Bacillus thuringiensis (NL-11), Streptomyces thermocarboxydus (NL-1), and Gongronella butleri (NL-15) were isolated from soil surrounding weathered dolostones [7-9]. The bacterial strain NL-11, actinomycetes NL-1, and fungal strain NL-15 were cultured in a liquid medium, subjected to oscillation for 24 hours, and then fermented in a fermentation tank. At regular intervals, the microorganisms’ wet mass was calculated and recorded in order to create a curve graph. The bacteria were transferred to a sterilized plastic bottle and kept in a fridge before the curve initially peaked and started to drop. In order to evaluate the impacts on plant development and root nodulation, we picked these three inoculums and treatment approaches.
2.3. Pot experiment setup
Robinia pseudoacacia L. was used for each group, with eight replicates per group, totaling 32 pots. Each pot included 60 ml of mixed microorganisms in addition to 5 kg of nursery materials. The experiment consisted of four microbial groups: NL-11 (RPJ1), NL-11 + NL-15 (RPJ2), NL-1 + NL-11 + NL-15 (RPJ3), and a microbial liquid medium without microbes (CK).
The pot experiment began in December 2019, and sampling began in November 2020. Root and soil samples were removed, placed in low temperature heat insulation box and transferred to the laboratory for a short period of time to maintain freshness. Plant root nodules and rhizosphere soil were separated with sterilized tweezers for subsequent test.
2.4. Rhizosphere soil sampling
The R. pseudoacacia and soil samples were brought to the lab from the study location in a box with ice to maintain a consistent temperature. During the sampling period, most of the soil was removed by shaking and a small portion of the root soil was retained.
2.5. Rhizosphere soil physicochemical properties
The pH of the rhizosphere soil was measured using a glass electrode (PHS220-K, Mettler Shanghai, China) suspended in a 1 mol/L KCl solution (w: v, 1: 5). The carbon (TC) and nitrogen (TN) contents of the soil and biochar were measured using a various EL III elemental analyzer. 3,5-dinitro salicylic acid colorimetry was employed to measure the sucrase activity in soil, and the results were represented as mol glucose g-1 dry sample. Using urea as the substrate, soil urease activity was measured using indophenol colorimetry. The results were represented as μmol ammonium g-1 dry sample. Disodium phenyl phosphate colorimetry was used to measure the soil phosphatase activity, which was then reported as mol phenol per dry sample. Using 0.1 mol/L KMnO4, soil catalase activity was titrated for 20 minutes, and the findings were reported as mol KMnO4 g-1 dry sample.
2.6. Soil DNA extraction and amplification of sequencing
Total DNA was extracted from a total of 24 samples, with six samples obtained for each treatment. The extracted DNA samples underwent quality tests, and NanoDrop 2000 spectrophotometers were used to measure the quantities. Using the primers 338F (5’-ACTCCTACGGGAGGCAGCAG-3’) and 806R (5’-GGACTACHVGGGTWTCTAAT-3’), bacterial 16S rRNA gene segments (V3-V4) were amplified from the isolated DNA. Using the primers CS1-ITS3 (5′-ACACTGACGACATGGTTCTACACAHCGATGAAGAACGYRG-3′) and CS2-ITS4 (5′-TACGGTAGCAGAGACTTGGTCTTTCCTSCGCTTATTGATATGC-3′), the partial ITS region (ITS3-ITS4) of fungi was amplified: 27 cycles at 55 ℃ for 30 s. The size of the amplicons was confirmed using agarose gel electrophoresis. The bacterial and fungal raw high-throughput sequencing data were deposited in the NCBI Genbank database with the accession number SRA accession: PRJNA638789, respectively.
2.7. Data analysis and statistics
At a significance level of 5% (p < 0.05), the least significant difference (LSD) by Duncan’s multiple range test was performed to examine the significance of differences in soil parameters among four treatments. Correlational analysis was performed using the “corrr” R (4.2.2) package [19], and certain figures were generated using the “ggplot2” R (4.2.2) package [20]. Additionally, redundancy analysis (RDA) was conducted to examine the relationship between the relative abundance of the top 10 microbial phyla and plant nodulation. After demultiplexing, the resulting sequences were processed by FLASH (v1.2.11) [21], fastq (0.19.6)[22], the DADA2 [23], the Qiime2 [24] (version 2020.2).
3. Result
3.1. Soil properties under different mineral-solubilizing microbial inoculums treatments
There were significant differences observed in soil enzyme activity and pH values across the treatments of mineral-solubilizing microbial inoculums, which were assessed based on 8 soil characteristics. The PRJ1 treatment showed a considerable reduction in soil pH, and an increase in soil phosphatase and urease activity, compared to the control (CK) (P < 0.05). The PRJ2 treatment resulted in a significant improvement in soil sucrase activity and total carbon content (P < 0.05). However, there were no significant differences observed in other soil properties among the mineral-solubilizing microbial inoculum treatments (Table 1).
3.2. Soil microbial community composition under different mineral-solubilizing microbial inoculums treatments
After sequencing and quality filtering, a total of 2,753,009 bacterial and 2,537,567 fungal high-quality sequences were obtained from all 32 soil samples, with an average of 86,032 and 79,299 sequences per sample, respectively. These sequences were then clustered into 9,230 bacterial and 2,346 fungal OTUs. The Venn diagram analysis of bacterial OTUs indicated that 4,240 OTUs were shared across all four treatment groups, while RPJ1, RPJ2, RPJ3 and CK had 269, 344, 334, and 323 unique OTUs, respectively (Figure 1a). Similarly, the Venn diagram of fungal OTUs showed that 887 core OTUs were present in all four treatment groups, with 145, 173, 186, and 86 unique OTUs found in soils from RPJ1, RPJ2, RPJ3, and CK, respectively (Figure 1b).
The dilution curve is an essential tool for evaluating the quality of sequencing data in a sample library and its ability to cover all microbial groups. As the number of sequences increased in our investigation, the dilution curve for the bacteria and fungi (Figure S1) demonstrated that the curve for the sequencing samples tended to stabilize. This pattern suggests that the sequences have saturated and that further sequencing information has a smaller impact on the discovery of new OTUs. These findings collectively imply that the measured data amount was reasonable and that the sequencing data can accurately reflect the makeup of the soil microbial community structure.
In terms of the fungal community, Ascomycota was the most abundant phylum, followed by Basidiomycota, Chytridiomycota, and Zygomycota, covering 96.50, 95.92, 95.63, and 96.42% in CK, RPJ1, RPJ2, and RPJ3, respectively. Compared with CK, RPJ2 had a higher relative abundance of Ascomycota and Basidiomycota, while RPJ1 and RPJ3 had a higher relative abundance of Chytridiomycota and Zygomycota (Figure 2B). These results indicated that the rock-solubilizing microbial inoculums had different effects on the rhizosphere soil bacterial and fungal communities, which may be related to their different mineral solubilization capacities and the specific interactions between microbes and plants. Ascomycota, Basidiomycota, Mortierellomycota, and Chytridiomycota, which account for 31.23%, 34.67%, 48.41%, and 46.91% of the total fungal phyla, were found in CK, RPJ1, RPJ2, and RPJ3 at relative abundances larger than 1%. With a proportion of 44.15%, RPJ2 stood out as having the highest relative abundance of Ascomycota (Figure 2B).
3.3. Soil microbial community diversity under different treatments
Alpha diversity differences between the various treatments were determined by the abundance-based Shannon index, richness estimator Chao1, and microbial community coverage ACE index. The results indicated that RPJ2 significantly increased the soil bacterial Shannon and chao1 indices compared with CK (Figure 3A). RPJ3 significantly improved the soil fungi Shannon index compared with CK (Figure 3B). In order to further illustrate the impacts of mineral-solubilizing microbial inoculums on soil bacteria (Figure 4A) and fungal (Figure 4B) communities, principal coordinate analysis (PCoA) based on the Weighted UniFrac metric was carried out. The first two axes (PCoA1 and PCoA2) explained 46% and 45% of the cumulative variance, respectively (Figure 4A,B). The PCoA plots showed that there was no overlap among the microbial communities of CK, RPJ1, RPJ2, and RPJ3, indicating significant differences in the soil microbial communities under different mineral-solubilizing microbial inoculum treatments (Figure 4).
The relative abundance of particular bacteria and fungus that significantly differed among the treatments were identified using LEfSe analysis (Figure 5). When compared to the other treatments, Actinomarina, Promicromonspora, and Micromonospora had a considerably higher concentration of CK (Figure 5A). Comparing RPJ1 to the other treatments, Rubrobacter and Streptomycetes were highly enriched (Figure 5A). In comparison to the other treatments, Solirubrobacterales and Pseudonocardiaceae were significantly enriched in RPJ2 (Figure 5A). Comparing RPJ3 to the other treatments, acidimicrobiales were substantially more abundant (Figure 5A). In comparison to the other treatments, Macrophomina and Mycosphaerellaceae were substantially CK enriched (Figure 5B). Comparing Pezizales to the other treatments, RPJ1 was considerably enriched in Pezizales (Figure 5B). In comparison to the other treatments, Arthrographis, Eremomycetaceae, and Eurotiales had considerably higher RPJ2 concentrations (Figure 5B). In comparison to the other treatments, Pleosporales and Orbiliaceae were significantly enriched in RPJ3 (Figure 5B).
3.4. The correlations between plant nodulation and soil microbial community composition
Spearman’s correlation analyses revealed several significant associations. Specifically, the relative abundance of Verrucomicrobia and Planctomycetes showed a positive correlation with plant height (P < 0.1, Figure 6A), while Acidobacteria exhibited a negative correlation with soil catalase activity (P < 0.001, Figure 6A). In addition, Verrucomicrobia and Chloroflexi were positively correlated with soil sucrase activity (P < 0.01, Figure 6A) and soil total carbon (P < 0.01, Figure 6A), respectively. Moreover, the relative abundance of Chloroflexi showed a positive correlation with soil total nitrogen (P < 0.01, Figure 6A). On the other hand, the relative abundance of Rozellomycota was negatively correlated with plant leaf area (P < 0.001, Figure 6B) and soil catalase activity (P < 0.01, Figure 6B), while Zoopagomycota exhibited a positive correlation with soil catalase activity (P < 0.001, Figure 6B). Furthermore, Chytridiomycota was positively correlated with soil sucrase activity (P < 0.001, Figure 6B), while Ascomycota showed a positive correlation with soil urease activity (P < 0.001, Figure 6B) and soil total nitrogen (P < 0.01, Figure 6B). Additionally, the relative abundance of Blastocladiomycota was positively correlated with soil total nitrogen (P < 0.001, Figure 6B).
RDA analysis was employed to investigate the relationship between soil microbial community composition and plant nodulation (Figure 7). For soil bacterial communities, RDA1 and RDA2 accounted for 85.85% and 9.71% of the total variation, respectively (Figure 7A). These ten soil bacterial communities explained 95.56% of the variation in plant nodulation. Notably, the relative abundance of Proteobacteria, Verrucomicrobia, and Gemmatimonadetes exhibited significant positive correlations with plant nodulation (Figure 7A). Regarding soil fungal communities, 92.37% of the variation in plant nodulation was significantly explained by these communities. Specifically, the relative abundance of Ascomycota, Basidiomycota, and Zoopagomycota showed significant positive correlations with plant nodulation (Figure 7B).
4. Discussion
An increasing body of research has demonstrated the potential of external microorganisms to enhance crop resilience against abiotic stresses, such as drought, heat, and salinity, as well as biotic stresses, such as soil-borne pathogen [25-29]. These microorganisms can serve as mineral-solubilizing microbial inoculums to improve soil texture, with strong ecological adaptability that makes them ideal for use in revegetation initiatives.
4.1. Rhizosphere soil activity
In the process of ecosystem reconstruction, the plant rhizosphere soil, being an active nutrient exchange area, is often the focus of research to understand the mechanisms underlying plant growth changes [30-35]. Soil enzyme activity is a crucial indicator of soil quality [36]. Our study has shown that mineral-solubilizing microbial inoculums can enhance soil enzyme activity, thus improving the soil’s nutrient supply capacity and having a positive effect on promoting plant growth. Previous studies have demonstrated that the use of exogenous microorganisms can directly or indirectly increase the number of microorganisms and root exudates in the rhizosphere soil, contributing to the enhancement of soil enzyme activities [37-39]. These results concur with our findings..
The soil environment typically hosts a stable microbial community, and mining activities can disrupt the ecological balance and induce adaptations in the original microorganisms, leading to changes in microbial community structure and diversity [40-42]. Our study found that the addition of mineral-solubilizing microbial inoculums altered the structure of the rhizosphere soil microbial community, with the fungal community showing particularly significant changes. These findings are in line with previous research that has investigated the impact of exogenous microbial additives on microbial community structure [43,44].
The addition of mineral-solubilizing microbial inoculums had a significant effect on bacterial abundance and diversity during the potting experiment, indicating changes in bacterial α-diversity under the test conditions. However, the effect of microbial inoculums on soil microbial abundance and diversity was more controversial. Our study showed that there was no significant effect of bacterial agent addition on bacterial diversity. However, some studies on the effects of microbial agent addition have shown a reduction in the diversity of microbial communities in soil [45]. Therefore, it is suggested that microbial diversity may depend on specific site conditions and may vary with the input of microbial inoculums.
4.2. Effect of addition of mineral-solubilizing microbial inoculums on root nodulation of plant
In our study, we observed that mineral-solubilizing microbial inoculums had a positive impact on nodule number and total volume. Previous research has demonstrated that exogenous microorganisms can enhance plant root growth and improve their ability to withstand drought and infertile soil conditions [46]. In addition, the addition of exogenous substances like synthetic nodulation factors can increase the number of root nodules [47]. Our study achieved a similar effect by altering the rhizosphere soil microbial community through the action of mineral-solubilizing inoculums. We observed an increase in the relative abundance of Proteobacteria with the addition of mineral-solubilizing microbial inoculums. This is consistent with previous studies that found a positive correlation between the relative abundance of Proteobacteria and total carbon, total nitrogen, available potassium, and available phosphorus [48]. Proteobacteria also contain rhizobia, which are required for symbiotic nodulation in leguminous plants. Our findings suggest that an increase in the abundance of rhizobia in soil may lead to an increase in bacteria activated by nodulation factors in the root system, thereby promoting plant nodulation [49]. While plants play a leading role in regulating root nodulation, mineral-solubilizing microbial inoculums also play an important role in promoting nitrogen fixation and absorption during plant growth.
5. Conclusions
This study provides valuable information on the effects of adding mineral-solubilizing microbial inoculums to the rhizosphere soil of R. pseudoacacia, including changes in the taxonomic and functional characteristics of the microbial community. The addition of different mineral-solubilizing microbial inoculums had a significant impact on the relative abundance of fungal communities and bacterial diversity. The RPJ1 treatment was found to be the most effective in altering the rhizosphere soil microbial community of R. pseudoacacia. Additionally, our study revealed a mechanism for the effects of mineral-solubilizing microbial inoculums on promoting plant growth, specifically through the regulation of soil microbial community to increase the number and volume of R. pseudoacacia root nodules. Our findings demonstrate a linkage between enhanced symbiotic nitrogen fixation capacity of plants and the associated soil microbial community. The results of this study provide valuable insights into the impact of mineral-solubilizing microbial inoculums addition on plant growth for ecological restoration.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Zhaohui Jia; Methodology, Zhaohui Jia, Shilin Ma and Xuefei Cheng; Software, Zhaohui Jia and Chong Li; Validation, Zhaohui Jia; Formal analysis, Zhaohui Jia and Chong Li; Investigation, Xuefei Cheng and Jingchi Zhang; Resources, Shilin Ma; Data curation, Miaojing Meng, Xuefei Cheng, Hui Nie and Jingchi Zhang; Writing—original draft, Zhaohui Jia; Writing—review & editing, Chong Li, Shilin Ma, Xin Liu, Miaojing Meng and Jingchi Zhang. All authors have read and agreedto the published version of the manuscript.
Funding
This research was funded by the Jiangsu Science and Technology Plan Project [BE2022420]; the Innovation and Promotion of Forestry Science and Technology Program of Jiangsu Province [LYKJ (2021) 30]; the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD); and the Scientific Research Project of Baishanzu National Park (2021KFLY05).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful to Mr. Frank and Mrs. Leng for the editing of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mymrin, V.; Aibuldinov, E.K.; Alekseev, K.; Avanci, M.A.; Rolim, P.H.B.; Catai, R.E.; Carvalho, K.Q. Sustainable material manufacturing from hazardous bauxite red mud in composites with clay slate waste and lime production waste. The International Journal of Advanced Manufacturing Technology 2020, 111, 1375–1385. [Google Scholar] [CrossRef]
- Shehata, N.; Sayed, E.T.; Abdelkareem, M.A. Recent progress in environmentally friendly geopolymers: A review. Science of The Total Environment 2021, 762, 143166. [Google Scholar] [CrossRef]
- Chang, Z.; Long, G.; Zhou, J.L.; Ma, C. Valorization of sewage sludge in the fabrication of construction and building materials: A review. Resources, Conservation and Recycling 2020, 154, 104606. [Google Scholar] [CrossRef]
- Li, T.; Wang, A.; Xing, W.; Li, Y.; Zhou, Y. Assessing mineral extraction and trade in China from 1992 to 2015: A comparison of material flow analysis and exergoecological approach. Resources Policy 2019, 63, 101460. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Huang, D.; Wang, X. Evaluation Study of Ecological Resilience in Southern Red Soil Mining Areas Considering Rare Earth Mining Process. Sustainability 2023, 15, 2258. [Google Scholar] [CrossRef]
- Liu, W.; Lee, D.-R.; Wang, S.-Y.; Yu, H.-W. Assessing the ecological loss of mining areas in Taiwan. Environmental Monitoring and Assessment 2023, 195. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, J.; Wang, L.; Wang, Y. A rock-weathering bacterium isolated from rock surface and its role in ecological restoration on exposed carbonate rocks. Ecological Engineering 2017, 101, 162–169. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, J.; Guo, X.; Wang, Y.; Wang, Q. Isolation and characterisation of a rock solubilising fungus for application in mine-spoil reclamation. European Journal of Soil Biology 2017, 81, 76–82. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, J.; Guo, X. An Indigenous Soil Bacterium Facilitates the Mitigation of Rocky Desertification in Carbonate Mining Areas. Land Degradation & Development 2017, 28, 2222–2233. [Google Scholar] [CrossRef]
- Jia, Z.; Meng, M.; Li, C.; Zhang, B.; Zhai, L.; Liu, X.; Ma, S.; Cheng, X.; Zhang, J. Rock-Solubilizing Microbial Inoculums Have Enormous Potential as Ecological Remediation Agents to Promote Plant Growth. Forests 2021, 12, 357. [Google Scholar] [CrossRef]
- Li, C.; Jia, Z.; Peng, X.; Zhai, L.; Zhang, B.; Liu, X.; Zhang, J. Functions of mineral-solubilizing microbes and a water retaining agent for the remediation of abandoned mine sites. Science of The Total Environment 2021, 761, 143215. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jia, Z.; Zhai, L.; Zhang, B.; Peng, X.; Liu, X.; Zhang, J. Effects of Mineral-Solubilizing Microorganisms on Root Growth, Soil Nutrient Content, and Enzyme Activities in the Rhizosphere Soil of Robinia pseudoacacia. Forests 2021, 12, 60. [Google Scholar] [CrossRef]
- Li, C.; Zhaohui, J.; Yingdan, Y.; Xuefei, C.; Jichang, S.; Xinggang, T.; Yuhao, W.; Xiaonan, P.; Yiqiao, D.; Shilin, M.; et al. Effects of mineral-solubilizing microbial strains on the mechanical responses of roots and root-reinforced soil in external-soil spray seeding substrate. Science of The Total Environment 2020. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Yanwen, W.U.; Wang, L.; Jia, Z. Effects of Soil Bacteria Inoculation in Spray Seeding Matrix on Photosynthesis Characteristics and Chlorophyll Fluorescence Parameters of Amorpha fruticose. Research of Environmental Sciences 2017, 30, 902–910. [Google Scholar]
- Srivastava, S.; Randhawa, J.; Anand, V.; Bist, V.; Singh, P.; Rastogi, S.; Yadav, S.; Srivastava, S. Phosphate Starvation Responses in Plants and Microbe Mediated Phosphorus Recycling in Soil: A Review. INTERNATIONAL JOURNAL OF PLANT AND ENVIRONMENT 2022, 8, 25–37. [Google Scholar] [CrossRef]
- Girme, M.; Sarnobat, D. Effect of nutrient supply system on biometry and rhizosphere microflora of soybean, pigeon pea and sorghum. Agricultural Science Digest - A Research Journal 2018. [Google Scholar] [CrossRef]
- Sun, S.; Xue, R.; Liu, M.; Wang, L.; Zhang, W. Research progress and hotspot analysis of rhizosphere microorganisms based on bibliometrics from 2012 to 2021. Frontiers in Microbiology 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, Y.; Wang, Y.; Sun, Y.; Chen, Y. High stand density promotes soil organic carbon sequestration in Robinia pseudoacacia plantations in the hilly and gully region of the Loess Plateau in China. Agriculture, Ecosystems & Environment 2023, 343, 108256. [Google Scholar] [CrossRef]
- Jackson, S. Correlations in R [R package corrr version 0.2.1]. 2016.
- Wickham, H. ggplot2. Wiley Interdisciplinary Reviews: Computational Statistics 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: fast length adjustment of short reads to improve genome assemblies. BIOINFORMATICS 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- O'Halloran, D.M. fastQ_brew: module for analysis, preprocessing, and reformatting of FASTQ sequence data. BMC research notes 2017, 10, 275–275. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. NATURE METHODS 2016, 13, 581. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2 (vol 37, pg 852, 2019). NATURE BIOTECHNOLOGY 2019, 37, 1091–1091. [Google Scholar] [CrossRef]
- Buntić, A.; Tošić, S.; Knežević, M.; Jovković, M.; Sikiric, B.; Koković, N.; Saljnikov, E. Synergistic effect of Bacillus isolates and biomass ash on soil and plant quality: A preliminary pot- experiment with the analysis of potentially toxic elements. Zemljiste i Biljka 2021, 70, 42–55. [Google Scholar] [CrossRef]
- Chatterjee, S.; Mondal, K.; Chatterjee, S. Role of Soil Microbes in Soil Health and Stability Improvement. 2022; pp. 579-592. [CrossRef]
- Gong, D.; ll, S.; Xinghao, L.; Zhang, W.; Zhang, D.; Cai, J. Micro/Nanofabrication, Assembly, and Actuation Based on Microorganisms: Recent Advances and Perspectives. Small Structures 2023. [Google Scholar] [CrossRef]
- Kaboosi, E.; Rahimi, A.; Abdoli, M.; Ghabooli, M. Comparison of Serendipita indica Inoculums and a Commercial Biofertilizer Effects on Physiological Characteristics and Antioxidant Capacity of Maize Under Drought Stress. Journal of Soil Science and Plant Nutrition 2022, 23. [Google Scholar] [CrossRef]
- Treesubsuntorn, C.; Dhurakit, P.; Khaksar, G.; Thiravetyan, P. Effect of microorganisms on reducing cadmium uptake and toxicity in rice (Oryza sativa L.). Environmental Science and Pollution Research 2018, 25, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gang, L.; Nan, L.; Wang, N.; Yang, W. Effects of Soil Organic construction on Regional Surface ET and GPP of Coal mine goaf. Journal of Physics: Conference Series 2020, 1549, 022091. [Google Scholar] [CrossRef]
- Herrington, R.; Tibbett, M. Cradle-to-cradle mining: a future concept for inherently reconstructive mine systems? 2022; pp. 19–28. [Google Scholar] [CrossRef]
- Hu, J.; ye, B.; Bai, Z.; Feng, Y. Remote Sensing Monitoring of Vegetation Reclamation in the Antaibao Open-Pit Mine. Remote Sensing 2022, 14, 5634. [Google Scholar] [CrossRef]
- Metsaranta, J.; Beauchemin, S.; Langley, S.; Tisch, B.; Dale, P. Assessing the Long-Term Ecosystem Productivity Benefits and Potential Impacts of Forests Re-Established on a Mine Tailings Site. Forests 2018, 9, 707. [Google Scholar] [CrossRef]
- Pietrzykowski, M. Tree species selection and reaction to mine soil reconstructed at reforested post-mine sites: Central and eastern European experiences. Ecological Engineering: X 2019, 3, 100012. [Google Scholar] [CrossRef]
- Zhao, J.; ma, J.; Yang, Y.; Yu, H.; Zhang, S.; Chen, F. Response of Soil Microbial Community to Vegetation Reconstruction Modes in Mining Areas of the Loess Plateau, China. Frontiers in Microbiology 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yaying, L.; Yu, Y.; Yao, H. Effect of nonbiodegradable microplastics on soil respiration and enzyme activity: A meta-analysis. Applied Soil Ecology 2023, 184, 104770. [Google Scholar] [CrossRef]
- Huang, K.; Zhang, J.; Tang, G.; Bao, D.; Wang, T.; Kong, D. Impacts and mechanisms of biochar on soil microorganisms. Plant, Soil and Environment 2023, 69. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, D.; Cheng, H.; Ren, L.; Jin, X.; Fang, W.; Yan, D.; Li, Y.; Wang, Q.; Cao, A. Organic fertilizers activate soil enzyme activities and promote the recovery of soil beneficial microorganisms after dazomet fumigation. Journal of Environmental Management 2022, 309, 114666. [Google Scholar] [CrossRef]
- Umar, A.; Turaki, A.; Bagudo, A.; Yeldu, R. Production of Fibrinolytic Enzyme by Soil Actinobacteria. Proceedings of the Pakistan Academy of Sciences: B. Life and Environmental Sciences 2023, 60, 49–56. [Google Scholar] [CrossRef]
- Krause, S.; Näther, A.; Ortiz Cortes, V.; Mullins, E.; Kessel, G.; Lotz, B.; Tebbe, C. No Tangible Effects of Field-Grown Cisgenic Potatoes on Soil Microbial Communities. Frontiers in Bioengineering and Biotechnology 2020, 8. [Google Scholar] [CrossRef]
- Saghaï, A.; Wittorf, L.; Philippot, L.; Hallin, S. Loss in soil microbial diversity constrains microbiome selection and alters the abundance of N-cycling guilds in barley rhizosphere. Applied Soil Ecology 2021, 169, 104224. [Google Scholar] [CrossRef]
- Zhou, Z.; Ding, G.; Yu, M.-H.; Gao, G.; Wang, G. Diversity and Structural Variability of Bacterial Microbial Communities in Rhizocompartments of Desert Leguminous Plants; 2020. [CrossRef]
- Huang, L.; Bao, W.; Li, F.; Hu, H. Effects of soil structure and vegetation on microbial communities. Chinese Journal of Applied and Environmental Biology 2021, 27, 1725–1731. [Google Scholar] [CrossRef]
- Wang, Q.-Y.; Na, W.; Liu, Y.; Chen, G.; He, H.; Gao, J.; Zhuang, X.-L.; Zhuang, G.-Q. Microbial Community Structure of Soil Methanogens and Methanotrophs During Degradation and Restoration of Reed Wetlands in the Songnen Plain. Huan jing ke xue= Huanjing kexue / [bian ji, Zhongguo ke xue yuan huan jing ke xue wei yuan hui "Huan jing ke xue" bian ji wei yuan hui.] 2021, 42, 4968–4976. [Google Scholar] [CrossRef]
- Chen, J.; Mo, L.; Zhang, Z.; Nan, J.; Xu, D.; Chao, L.; Zhang, X.; Bao, Y. Evaluation of the ecological restoration of a coal mine dump by exploring the characteristics of microbial communities. Applied Soil Ecology 2020, 147, 103430. [Google Scholar] [CrossRef]
- Liu, X.-Q.; Xie, M.-M.; Hashem, A.; Abd Allah, E.F.; Wu, Q.-S. Arbuscular mycorrhizal fungi and rhizobia synergistically promote root colonization, plant growth, and nitrogen acquisition. Plant Growth Regulation 2023. [Google Scholar] [CrossRef]
- Ayala-Garcia, P.; Jiménez-Guerrero, I.; Jacott, C.; López-Baena, F.; Ollero, F.; del Cerro, P.; Pérez-Montaño, F. The Rhizobium tropici CIAT 899 NodD2 protein promotes symbiosis and extends rhizobial nodulation range by constitutive nodulation factor synthesis. Journal of Experimental Botany 2022, 73. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ullah, F.; Ahmad, R.; Shah, S.; Khan, A.; Adnan, M. Response of Soil Proteobacteria to Biochar Amendment in Sustainable Agriculture-A mini review. 2022, 1, 16–30. [Google Scholar] [CrossRef]
- Mondal, S.; Pramanik, K.; Pal, P.; Mitra, S.; Ghosh, S.; Mondal, T.; Soren, T.; Maiti, T. Multifaceted roles of root exudates in light of plant-microbe interaction. 2022; pp. 49-76. [CrossRef]
Figure 1.
Venn diagram of the number of unique and shared bacterial (A) and fungal (B) operational taxonomic units (OTUs) among the CK, RPJ1, RPJ2 and RPJ3 treatments based on a distance level of 97% similarity. Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
Figure 1.
Venn diagram of the number of unique and shared bacterial (A) and fungal (B) operational taxonomic units (OTUs) among the CK, RPJ1, RPJ2 and RPJ3 treatments based on a distance level of 97% similarity. Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
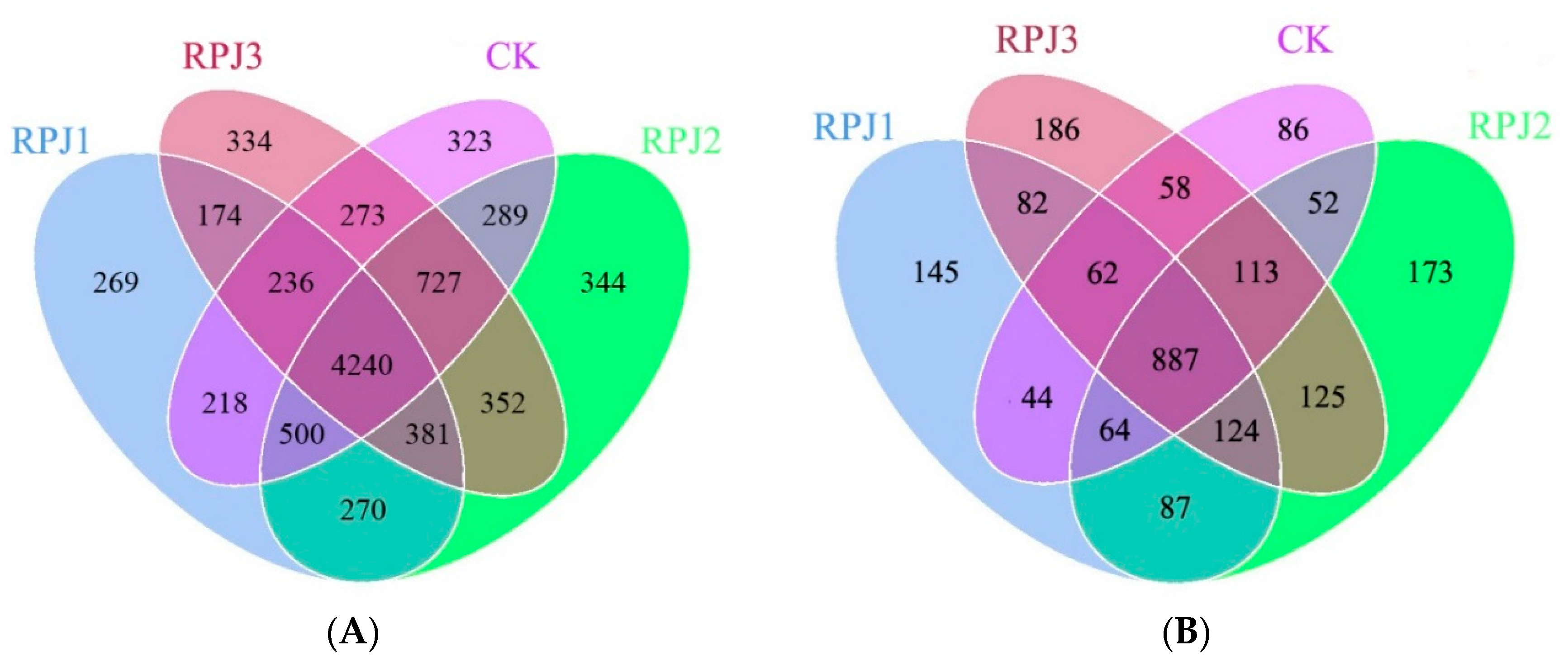
Figure 2.
The relative abundance(%) of the dominant bacterial phyla (A) and fungal phyla (B). Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
Figure 2.
The relative abundance(%) of the dominant bacterial phyla (A) and fungal phyla (B). Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
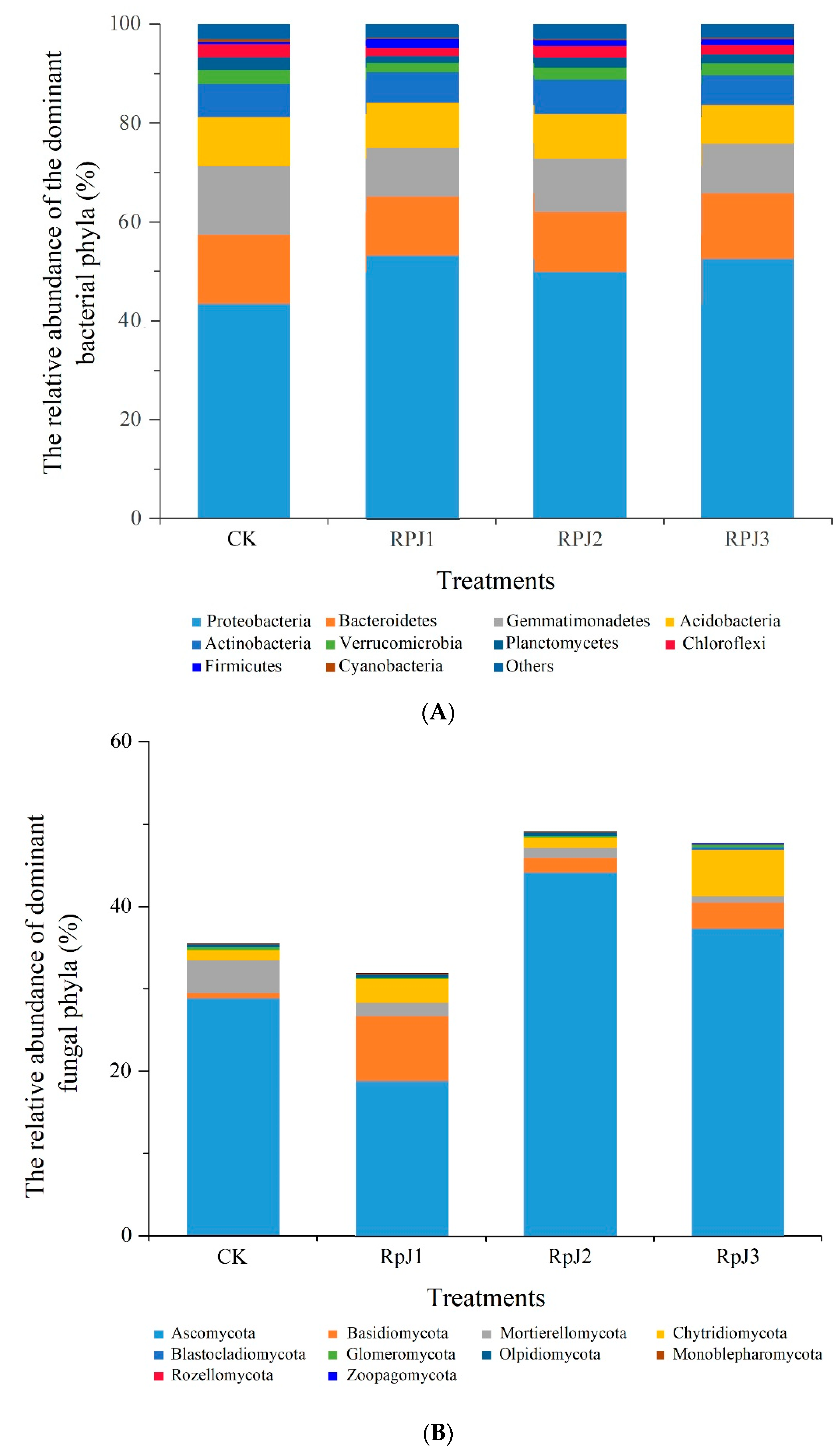
Figure 3.
Bacterial (A) and fungal (B) alpha diversity under different treatments. The ordinate on the left corresponds to the Shannon index, and the ordinate on the right corresponds to the Chao1 and ACE indices. Differences in alpha diversity between different treatments were compared with the Kruskal-Wallis test. *, p < 0.05; **, p < 0.01. Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
Figure 3.
Bacterial (A) and fungal (B) alpha diversity under different treatments. The ordinate on the left corresponds to the Shannon index, and the ordinate on the right corresponds to the Chao1 and ACE indices. Differences in alpha diversity between different treatments were compared with the Kruskal-Wallis test. *, p < 0.05; **, p < 0.01. Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
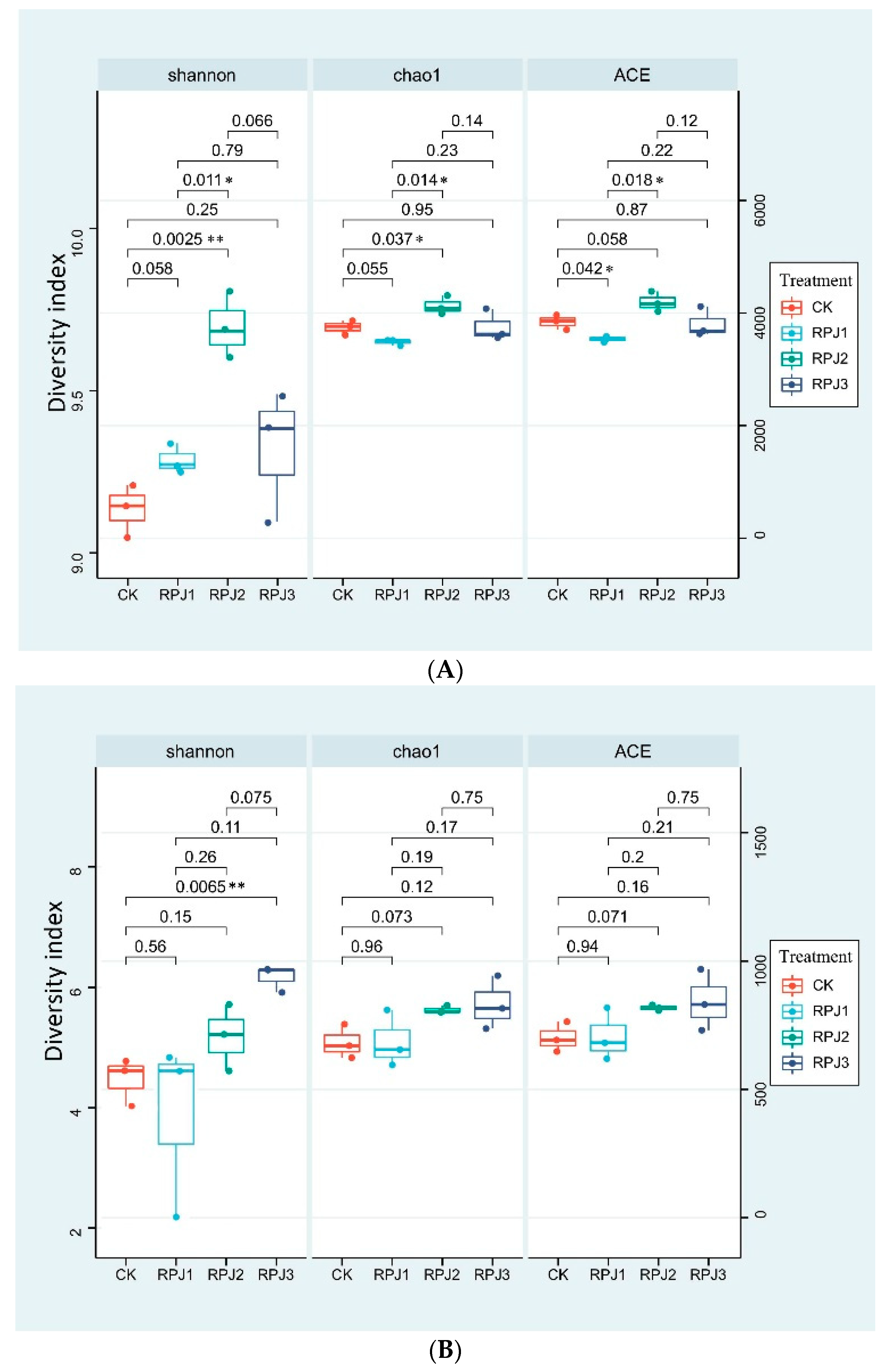
Figure 4.
Principal coordinates analysis (PCoA) of changes in the operational taxonomic units of rhizosphere soil bacterial (A) and fungi (B) based on Weighted UniFrac metric among all treatments. Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
Figure 4.
Principal coordinates analysis (PCoA) of changes in the operational taxonomic units of rhizosphere soil bacterial (A) and fungi (B) based on Weighted UniFrac metric among all treatments. Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
Figure 5.
Relationships among Plant growth, soil chemical properties and relative abundance of bacterial community(A) and fungal community (B). Significant correlations are marked in red (positive) and green (negative). *** indicates significant correlation at p<0.001** indicates significant correlation at p < 0.01; * indicates significant correlation at p < 0.05. SH, height; GD, basal diameter; LA, leaf areas; CAT, soil catalase activity; SUC, soil sucrase activity; HOS, soil alkaline phosphatase activity; URE, soil urease activity; pH, soil pH; C, soil total carbon ; N, soil total nitrogen; C/N, soil carbon to nitrogen ratio.
Figure 5.
Relationships among Plant growth, soil chemical properties and relative abundance of bacterial community(A) and fungal community (B). Significant correlations are marked in red (positive) and green (negative). *** indicates significant correlation at p<0.001** indicates significant correlation at p < 0.01; * indicates significant correlation at p < 0.05. SH, height; GD, basal diameter; LA, leaf areas; CAT, soil catalase activity; SUC, soil sucrase activity; HOS, soil alkaline phosphatase activity; URE, soil urease activity; pH, soil pH; C, soil total carbon ; N, soil total nitrogen; C/N, soil carbon to nitrogen ratio.
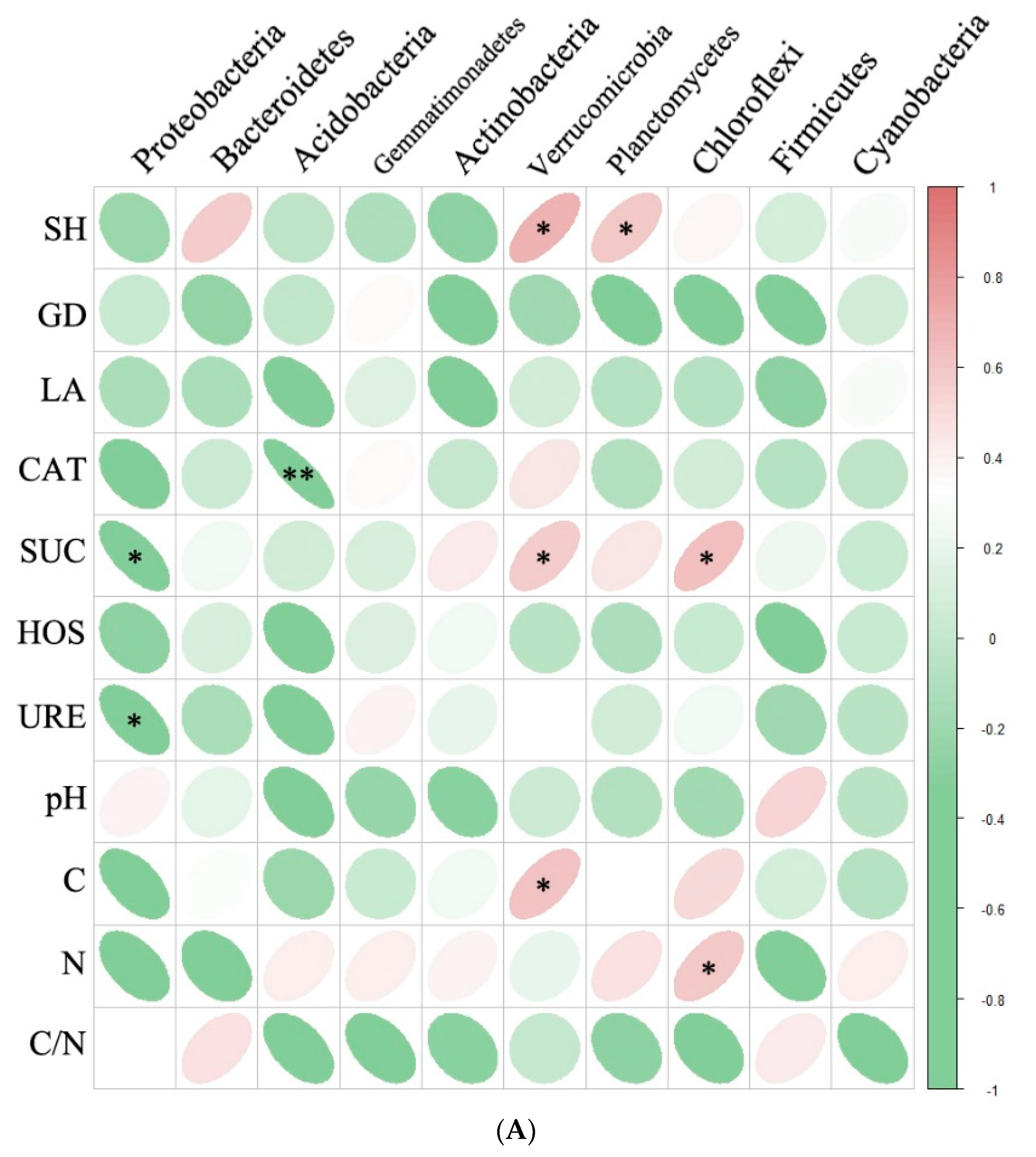
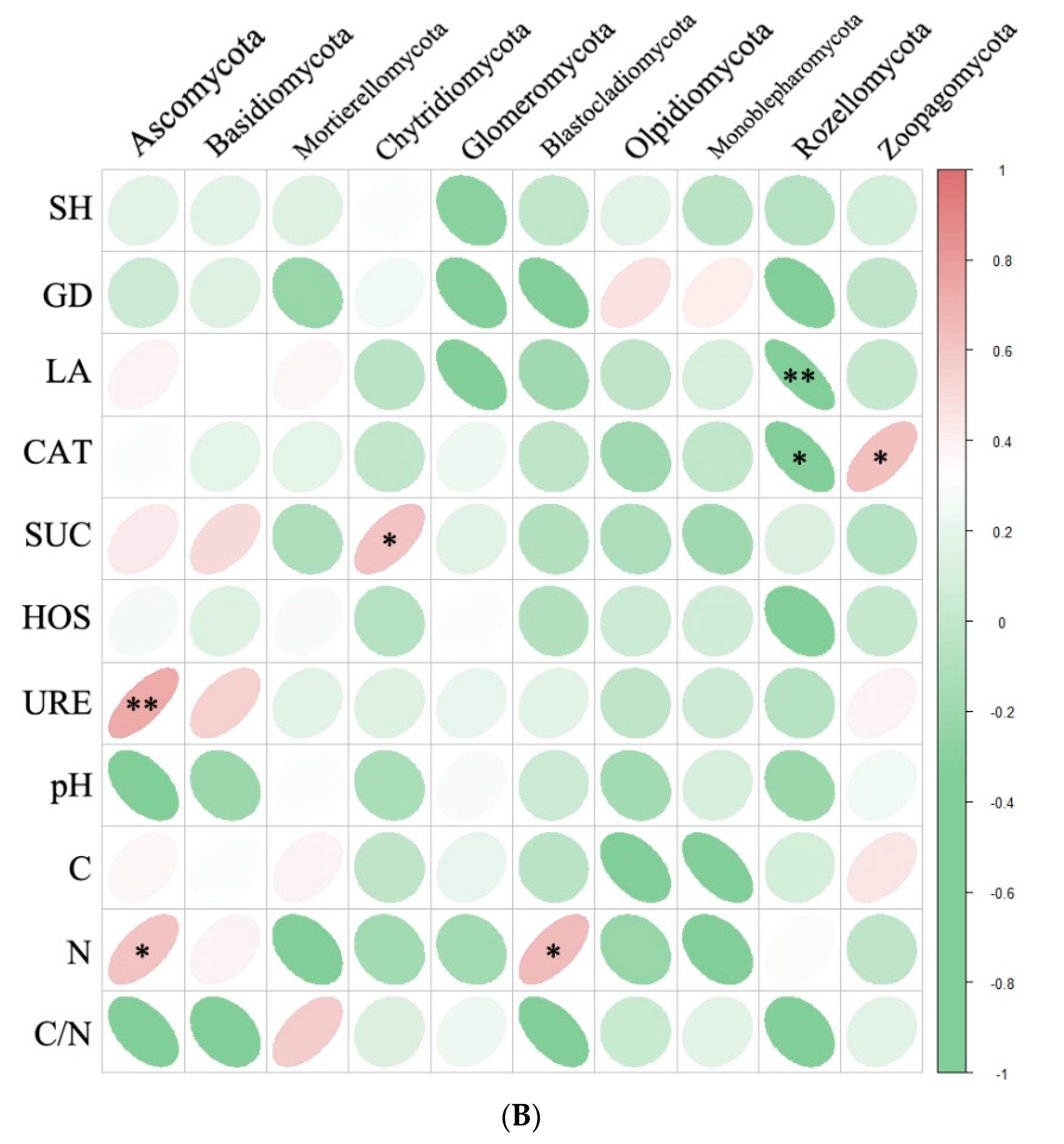
Figure 6.
Redundancy analysis (RDA) of plant nodulation and microbial community for bacterial (A) and fungal (B) in R. pseudoacacia rhizosphere. RNN, root nodules number; RNV, root nodules volume; RNDW, root nodules dry weight; RV, root volume; RNC, root nodules contribution.
Figure 6.
Redundancy analysis (RDA) of plant nodulation and microbial community for bacterial (A) and fungal (B) in R. pseudoacacia rhizosphere. RNN, root nodules number; RNV, root nodules volume; RNDW, root nodules dry weight; RV, root volume; RNC, root nodules contribution.
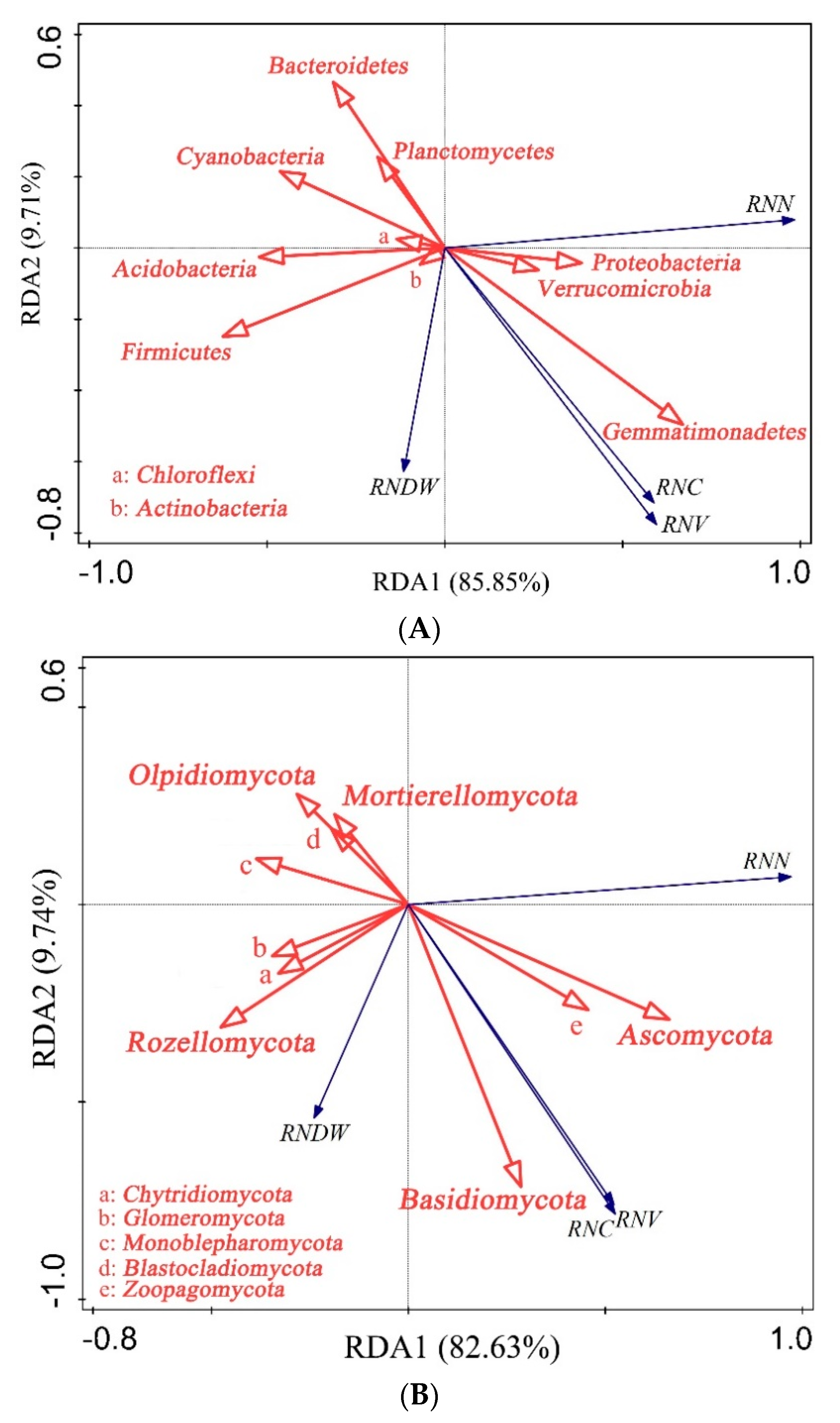
Figure 7.
Cladogram of rhizosphere soil bacteria (A) and fungi (B) in different treatments via LEfSe method identifies the significantly different abundant taxa. CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).
Figure 7.
Cladogram of rhizosphere soil bacteria (A) and fungi (B) in different treatments via LEfSe method identifies the significantly different abundant taxa. CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).

Table 1.
Soil properties in different treatments.
| Variables | CK | RPJ1 | RPJ2 | RPJ3 | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| CAT/(μmol/min) | 0.78a | 0.24 | 1.15a | 0.19 | 1.16a | 0.35 | 0.99a | 0.18 |
| SUC/(mg/g·24h) | 12.40b | 1.89 | 18.04ab | 4.88 | 26.57a | 15.21 | 17.71ab | 2.50 |
| HOS/(nmol/g·24h) | 338.31b | 21.40 | 375.08a | 20.64 | 365.38ab | 56.66 | 358.94ab | 50.54 |
| URE/(μg/g·24h) | 360.36b | 40.06 | 407.22a | 19.68 | 399.43ab | 2.50 | 378.16ab | 18.01 |
| pH value | 8.12a | 0.03 | 8.04b | 0.13 | 8.12a | 0.11 | 8.11a | 0.16 |
| TC/% | 7.37b | 0.32 | 7.77ab | 0.21 | 8.00a | 0.30 | 7.93ab | 0.35 |
| TN/% | 0.43a | 0.06 | 0.50a | 0.01 | 0.50a | 0.10 | 0.43a | 0.06 |
| C/N/% | 17.15a | 1.80 | 15.53a | 0.42 | 16.48a | 3.58 | 18.52a | 2.51 |
Note: Treatments: CK (control, add microbial liquid medium without microbes), RPJ1 (NL-11), RPJ2 (NL-11 + NL-15), and RPJ3 (NL-1 + NL-11 + NL-15).CAT: soil catalase activity; SUC: soil sucrose activity; HOS: soil alkaline phosphatase activity; URE: soil urease activity; pH value: soil pH; TC: total soil carbon; TN: total soil nitrogen; C/N: soil carbon to nitrogen ratio. Means are the average values of same treatment (n=3). Different letters indicate significance at a 0.05 probability level (p < 0.05) using the LSD test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



