
Submitted:
27 April 2023
Posted:
28 April 2023
You are already at the latest version
Abstract
Damping-off caused by Pythium aphanidermatum (Pa) is one of the most destructive diseases for watemelon seedlings. Application of biological control agents against Pa has attracted attention of many researchers for a long time. In this study, an actinomycete isolate JKTJ-3 with strong and broad-spectrum antifungal effect was screened from 23 bacterial isolates. Based on the morphological, cultural, physiological, and biochemical characteristics as well as the feature of 16S rDNA sequence, isolate JKTJ-3 was identified as Streptomyces murinus. We compared the biocontrol efficacy of isolate JKTJ-3 and its metabolites. The results revealed that seed and substrate treatments with the JKTJ-3 cultures showed a significant inhibitory effect on watermelon damping-off disease. Seed treatment with the JKTJ-3 cultural filtrates (CF) had higher control efficacy that of the fermentation cultures (FC). Treatment of the seeding substrate with the wheat grain cultures (WGC) of JKTJ-3 exhibited better control efficacy than that of the seeding substrate with the JKTJ-3 CF. Moreover, the JKTJ-3 WGC showed the preventive effect on suppression of the disease, and the efficacy increased with increase in the inoculation interval between the WGC and Pa. Production of the antifungal metabolite actinomycin D by isolate JKTJ-3 and cell wall-degrading enzymmes such as β-1,3-glucanase and chitosanase was probably the mechanisms for effective control of watermelon damping-off. It was showed for the first time that S. murinus can produce anti-oomycete substances including chitinase and actinomycin D. This is the first report about S. murinus used as biocontrol agent against watermelon Pa-induced damping-off.
Keywords:
Watermelon
; damping-off
; Pythium aphanidermatum
; Streptomyces murinus
; biocontrol
1. Introduction
Cucurbit damping-off disease is globally found at nursery beds, seedling losses have been reported ranging from 5% to 80% in infected areas [1,2]. Several pathogens in the genera Pythium, Rhizoctonia and Fusarium [3,4,5,6] are responsible for damping-off disease, of which Pythium is the most economically destructive pathogens with broad host range worldwide [7,8]. Once they were localized in a certain area, these pathogens will rapidly attack the seeds and seedlings, resulting in seed softening and rotting at pre-emergence stage and/or seedling damping off at post-emergence stage [9].
Varieties resistant to the pathogens of damping-off disease in cucurbit have seldom been reported. Fungicides are generally employed for chemical control against damping-off disease [10,11,12]. However, the overuse of fungicides has led to resistance of pathogens and failed to control Pythium damping-off disease [13,14,15]. Moreover, overuse of chemical agents threatens human health and causes ecological problems. Therefore, there is growing interest in developing alternative control methods, including biological control.
The beneficial microorganisms and their metabolites are used as biocontrol agents in biological control. Many naturally antagonistic bacteria and fungi have been used to suppress cucurbit Pythium damping-off [16,17,18,19]. Seed or substrate treatment with antagonistic bacteria or fungi is a good option for disease control. In [2] it was revealed that the biocontrol effect of the bacterial agent prepared from P. fluorescens and peat was better than that of the bacterial agent prepared from Bacillus and peat. Previous results had demonstrated that four actinomycete isolates mixed with the soil could significantly decrease the occurrence of cucumber damping-off disease, and their control effects were close to that of metalaxyl under pot culture [20]. Roberts et al. [21] reported the control effect of cucumber, melon, and pumpkin seed treatment with the cells and cell-free extracts of Serratia marcescens N4-5 against cucurbit Pythium damping-off. Huang et al. [22] also reported that the seed coating treatment or soil mixing treatment with Trichoderma effectively controlled damping-off disease. These biocontrol agents play a role in biocontrol by competition, producing antibiotic, inducing systemic resistance, and mycoparasitism [23,24,25,26].
China’s cucurbit planting area ranks the first in the world, of which the largest one is watermelon. The intensive seedling cultivation has played an important role in ensuring high yield and quality production of watermelon. Pythium damping-off is a disease frequently occurring during watermelon seedling raising, and P. aphanidermatum (Pa) is the dominant pathogen [27,28]. From 2016 to 2018, the author found that cucurbit damping-off happened in Hubei, China, with an incidence of 10%~30%, even over 30%, especially in scions cultivated by trays for cucurbit grafting seedling raising. 221 strains from Pythium damping-off in cucurbit seedlings was isolated, which was identified as P. aphanidermatum, P. ultimum, P. irregular, P. spinosum, and Pythium sp. Among these strains, P. aphanidermatum accounted for 52% of the total isolates and became the predominant species in Hubei [29]. However, the research on biological control of cucurbit damping-off disease mostly focuses on cucumbers, rather than watermelon.
Considering this, the current study investigated the biocontrol of watermelon damping-off caused by Pa. The objectives of the study were: (i) to screen a broad-spectrum antagonistic isolate with biocontrol potential against damping-off disease caused by Pa; (ii) to identify the antagonistic isolate based on the morphological, cultural, physiological, and biochemical, and taxonomic characteristics; (iii) to evaluate the prevention and control effects of seed and substrate treatment with different forms and different concentrations of the antagonistic isolate; and (iv) to reveal the potential biocontrol mechanisms of the antagonistic isolate.
2. Materials and Methods
2.1. Microbial isolates and cultural Conditions
A total of 36 microbial isolates were used in this study, including 10 actinomycete isolates (F46-1, F54, F77, H55-1a, JKTJ-3, V10a, V61a, V61b, W143, W143-1) isolated from watermelon plant rhizosphere in Wuhan, Hubei Province, 13 bacterial isolates (JKTJ-1, JKTJ-11, JKTJ-2, MKCC1, MKCC2, MKCC3, MKCC4, B.amy1, B.amy2, Pse2-22, Pse2-21, Lat6-4, S16-1) isolated from watermelon seeds, and 13 plant pathogens (Pa, Rhizoctonia solani, Colletotrichum gloeosporioides, Botrytis cinerea, Stagonosporopsis cucurbitacearum, Verticillium dahlia, Phomopsis vexans, Fusarium oxysporum f.sp. hiveum, Leptosphaeria biglobosa, Phomopsis asparagi, Fusarium solani, C. capsic, and Sclerotinia sclerotiorum). The plant, soil and seeds were collected and shaked in 100 mL sterile water for 5 min. Solution after two 10-fold dilutions were spreaded on nutrient agar (NA) (beef extract 3 g, peptone 10 g, NaCl 5 g, glucose 10 g, agar powder 13 g, water 1,000 mL, pH 7.0 to 7.2) and PDA potato dextrose agar medium (PDA, peeled potato 200 g, glucose 20 g, agar powder 13 g, water 1,000 mL). All plates were incubated at 28 °C to form single colony. Isolate JKTJ-3 was deposited at China Center for Type Culture Collection with registration number of CCTCC M 20211271. The cultural media included potato dextrose agar medium (PDA, peeled potato 200 g, glucose 20 g, agar powder 13 g, water 1,000 mL) and nutrient agar (NA) (beef extract 3 g, peptone 10 g, NaCl 5 g, glucose 10 g, agar powder 13 g, water 1,000 mL, pH 7.0 to 7.2). The microbial cultures were incubated at 28 °C in dark.
2.2. In vitro screening of biocontrol agents
Dual culture was used to screen the antagonists from 23 actinomycete and bacterial isolates against Pa by previously described method [30]. Briefly, a full ring of the candidate isolate spores were streaked on one side of the PDA dish (a diameter of 9 cm) with the spore line 3 cm away from the dish center, and cultured at 28 °C. After 2-day cultivation, a 6 mm-diameter fungal disc taken from the edge of a 2-day-old colony of Pa was placed on the center of PDA dish and cultured. PDA dish without actinomycetes or bacteria were used as controls. All the dishes were incubated at 25 °C for 48 h. Afterwards, the width of inhibition zones (distance between the pathogen mycelia and the candidate isolates) in each dish was measured.
Further broad-spectrum test of the screened actinomycete and bacterial isolates was carried out. The above-mentioned dual culture method was adopted with slight modification that the pathogens and the screened antagonists were simultaneously inoculated on the PDA dish. PDA dish without the screened antagonists was used as controls. All the dishes were cultured at 25 °C. When the phytopathogenic fungi covered the whole dish, the width of inhibition zone in each dish was measured.
2.3. In vivo screening of biocontrol agents
Considering that the antagonists screened in vitro tended to lose their activities in vivo, our preliminarily screened antagonistic actinomycetes were re-screened. The method of Paul et al. [31] and Li et al. [32] with some modification were used in this experiment. Spores were scraped off the surface of the 5-day-old colonies cultured at 28 °C on PDA dishes. The spore concentration was determined with a hemocytometer and then adjusted to 1 × 107 spores/mL. One mL of spore suspension of the antagonistic actinomycetes was inoculated into 100 mL of PDB (the same components with PDA excluding agar powder) and cultured at 28 °C in a 220 rpm rotary shaker for 4 days. The obtained fermentation culture (FC) was used as the antagonistic actinomycete inoculum. The substrate composed of peat (particle size, 0-7 mm; pH, 5.6; electrical conductivity (EC), 0.23 dS/m; Floragard, Germany) and perlite (particle size, 3-6 mm) at the ratio of 3:1 (v/v) was sterilized by autoclaving at 121 °C for 30 min. The mixture of the sterilized substrate and antagonistic actinomycete inoculum at the ratio of 5:1 (v/v) was used as the potting substrate, and put into pots (9 × 8 × 6 cm) at 200 mL per pot. Then a 5-mm-diameter mycelium disc of Pa (2-day-old) was placed at the depth of 1.5 cm below the center of the potting substrate. Watermelon, Citrullus lanatus vatiety ZAOJIA 8424 (Xinjiang Mingxin Kehong Agricultural Technology Co., Ltd., China), was used in this experiment. The seeds were surface sterilized in 1% (w/v) formaldehyde for 1 h and rinsed with running water, followed by 8 h soaking in sterile water. Then the seeds were wrapped with a towel and germinated at 28 °C for 36 h. The germinated seeds with about 1 cm of radicle length were selected, spread to each pot, and covered with the potting substrate (about 1.0 cm thick). The sterile substrate was respectively mixed with the 300-fold diluted solution of 70% hymexazol wettable powder (WP) (Shanxi Biaozheng Crop Science Co. Ltd., China) and sterile water with the latter used as controls. Each treatment was carried out in triplicates with 5 pots per replicate and 20 seeds per pot. All the pots were covered with a layer of plastic film to retain the moisture and incubated at 30 °C during pre-emergence stage and then cultivated during post-emergence stage in climate chamber (25 °C for 12 h light/ 18 °C for 12 h dark photoperiod, 90% humidity). The number of seed emergence was recorded at day 5 after sowing, and the number of the damping-off seedlings was counted at day 7 after sowing. The seedling damping-off incidence (DI) and seedling rate (SR) were calculated as follows [18], respectively.
where A and B indicated the total number of emergeed seeds and the number of seedlings with damping-off, respectively.
DI = 100 × B/20
SR = 100 × (A − B)/20
2.4. 16S rDNA sequencing
One mL of isolate JKTJ-3 spore suspension was inoculated into 100 mL PDB and cultured in at 220 rpm rotary shaker for 48 h. Subsequently, 2 mL of the cell suspension was centrifuged at 10000 rpm for 5 min, and the supernatant was discarded. The cells were resuspended in the sterile water and washed twice. The genomic DNA of isolate JKTJ-3 was extracted with bacterial genomic DNA extraction kit (Tiangen, model DP302-02, China) according to the manufacturers’ instruction. The DNA extract was stored at -20 °C. Subsequently, 16S rDNA gene was amplified by PCR in 25 μL system containing 1 μL PCR Mix (Aidlab Biotechnologies Co., Ltd., China), 1 μL of each primer (10 pmol/μL, Tsingke Biotechnology Co., Ltd., China), 0.5 μL template DNA (75 μg/mL), and 21.5 μL deionized water. The universal primer pair 27f (5′-AGA GTT TGA TCC TGG CTC AG-3′) and 1495r (5′-CTA CGG CTA CCT TGT TAC GA-3′) was used. The PCR was conducted as follows: pre-denaturation at 94 °C for 5 min, 30 cycles of denaturation at 94 °C for 1 min, annealing at 58 °C for 1 min, and extension at 72 °C for 2 min, followed by a final extension at 72 °C for 10 min. The PCR product was detected by 1% (w/v) of agarose gel electrophoresis. Afterwards, the target DNA fragments were recovered with DNA Recovery Kit (Axygen AP-GX-50, Axygen Biotechnology (Hangzhou) Co., Ltd., China), and connected to the pMD® 18-T vector (Takara Biotechnology (Dalian) Co., Ltd., China). Finally, the recombinant vectors were transformed into the competent cells of Escherichia coli JM109 (Takara Biotechnology (Dalian) Co., Ltd., China), and the positive clones containing the target DNA fragments were sequenced (Tsingke Biotechnology Co., Ltd., China). The 16S rDNA sequence of isolate JKTJ-3 was aligned against the EzBioCloud server and the GenBank databases to obtain the homology sequences. The multiple sequence alignment was performed by the CLUSTAL W program, and a phylogenetic tree was constructed by a maximum likelihood method using MEGA X software (Version 10.0.5). The support of each clade was determined by a bootstrap analysis with 1000 replications [33]. A matrix of pairwise distances was generated using Tamura-Nei model [34].
2.5. Determination of morphological, cultural and physiological features
The morphological characteristics of aerial hypha, spore hypha, and spore were observed by scanning electron microscope (SEM, Zeiss Sigma FESEM, Germany). The cultural characteristics of isolate JKTJ-3 on the ISP medium were determined, as previously reported [35,36]. Specifically, the aerial spore mass color, substrate mycelium pigmentation, and the soluble pigment production of isolate JKTJ-3 were recorded on yeast extract-malt extract agar (ISP-2), oatmeal agar (ISP-3), inorganic salts-starch agar (ISP-4), glycerol-asparagine agar (ISP-5), and peptone-yeast extract-iron agar (ISP-6), respectively. The pigment presence on Benett’s agar (BM) [37] and Gause’s No.1 [36] was also investigated. The growth characteristics of isolate JKTJ-3 were observed after 7-day incubation at 28 °C under dark condition.
Carbon source utilization, growth temperature, NaCl tolerance, and pH sensitivity of isolate JKTJ-3 were determined. A total of 8 carbon sources were used including D-glucose, sucrose, D-fructose, D-xylose, L-rhamnose, raffinose, L-arabinose, and D-mannitol. These carbon sources were prepared into 10% (w/v) solution and sterilized through a 0.22 μm syringe filter (Beijing Labgic Technology Co., Ltd., China), and then mixted with ISP-9 medium [36] to reach the final concentration of 1% (m/v) in the plate. The growth of isolate JKTJ-3 in ISP-2 medium was examined at the temperature of 10-45 °C and NaCl concentration of 0.5%-8% (w/v). The sensitivity of isolate JKTJ-3 to pH (3.0-8.0) was investigated in ISP-2 medium adjusted by 0.2 mol/L K2HPO4-HCl and 0.2 mol/L KH2PO4-K2HPO4. The growth characteristics were also recorded at day 7 after incubation at 28 °C in dark.
2.6. Determination of biocontrol efficacy of isolate JKTJ-3
A spore suspension of Streptomyce JKTJ-3 was prepared to reach a concentration of 107 spores/mL as described above. One mL of the JKTJ-3 spore suspension was inoculated into 100 mL of PDB and incubated in 220 rpm shaker for 24 h at 28 °C to obtain the isolate JKTJ-3 culture. One mL of obtained 24-h JKTJ-3 culture was incubated in 100 mL of PDB in 220 rpm shaker for 4 days to obtain the isolate JKTJ-3 fermentation culture (FC). FC was filtered with qualitative filter paper to obtain the crude filtrate. The crude filtrate was filtered with a 0.22 μm syringe filter to obtain the isolate JKTJ-3 cultural filtrate (CF). One mL of isolate JKTJ-3 spore suspension was inoculated into 100 g sterile wheat grain medium and incubated at 28 °C for 15 days to obtain the isolate JKTJ-3 wheat grain culture (WGC).
The control effects of different contents of JKTJ-3 CF or WGC on watermelon damping-off disease were examined. One liter of the sterile substrate (same in vivo screening preparation) was mixed with 10, 50, 100, 150, 200, 250, and 300 mL of the CF, respectively, corresponding to added with 290, 250, 200, 150, 100, 50, and 0 mL of sterile water to ensure the consistent water content in the substrate, namely, 10-300 mL CF per liter substrate (mL/L) including 10 mL/L, 50 mL/L, 100 mL/L, 150 mL/L, 200 mL/L, 250 mL/L, and 300 mL/L. Similarly, one liter of sterile substrate was mixed with 10, 20, 40, 80 and 160 g of JKTJ-3 wheat grain culture respectively to ensure 30% of the water content in the substrate, namely, 10-160 g WGC per liter seeding substrate (g/L) including 10, 20, 40, 80, and 160 g/L. One liter of the sterile substrate was mixed with 300 mL of 300-fold diluted solution 70% hymexazol WP as chemical treatment group, while one liter of sterile substrate was mixed with 300 mL of sterile water as control group. The substrate for each treatment was put into pots (9 × 8 × 6 cm) with 200 mL per pot. A 5 mm-diameter mycelium disc of Pa (25 °C for 48 h) was inoculated to a depth of about 1.5 cm below the center of the substrate. Each pot was sown with 20 germinated seeds and then covered with 1.0 cm-thick layer of the sterilized substrate. The damping-off incidence (DI) and seedling rate (SR) were determined, as described in the secondary screening experiment, and the seed emergence rate (ER) and control efficacy (CE) were calculated as follows, respectively.
where A represented the total number of seed emergence, while DIsw represented the damping-off incidence in the treatment of sterilized water (SW).
ER = 100 × A/20
CE = 100 × (DISW − DI)/DISW
The preventive effect of isolate JKTJ-3 CF and WGC on watermelon damping-off by substrate treatment was investigated. The CF and WGC of isolate JKTJ-3 was mixed with the sterilized substrate (same as secondary screening preparation) at the proportion of 30% (v/v) and 7.5% (w/v), respectively. All the substrates for WGC-treatment were added with 300 mL of sterile water to ensure the 30% water content in substrates. The 300-fold diluted solution 70% hymexazol WP and SW were respectively mixed with the sterile substrate at the proportion of 30% (v/v) and used as chemical treatment group and control group, respectively. The substrate for each treatment was put into pots (9 × 8 × 6 cm) with 200 mL per pot. Each pot was sown with 20 seeds (disinfected in 1% formaldehyde for 1 h, then soaked for 8 h), and then covered with a layer of about 1.0- cm-thick substrate. A 5 mm-diameter mycelium disc of P. aphanidermatum (48-hour-old) was inoculated to about 2.5 cm depth below the center of the substrate at day 0, 1, 2, and 3 after sowing, respectively. Seedling indices (ER, DI, SR, and RCE) were determined, as described in Section 2.3 and Section 2.6.
The seeds (sterilized in 1% formaldehyde (v/v) solution for 1 h, washed with tap water, and then rinsed 3 times with sterile water) were soaked in FC or in CF for 4 h, 8 h, and 12 h, respectively. Seeds treated with the sterile water for 4 h, 8 h and 12 h were used as controls. The sterile substrate (prepared in the same way as in secondary screening) was put into pots with 200 mL per pot. A 5 mm-diameter of mycelium disc of Pa (2-day-old) was inoculated to about 1.5 cm depth below the center of the substrate. Every pot was sown with 20 soaked seeds, and then covered with a layer of 1.0 cm-thick sterilized substrate. Each treatment was repeated for 3 times with 5 pots in one replicate. Seedling indices (ER, DI, SR, and RCE) were detected, as described in Section 2.3 and Section 2.6.
2.7. Determination of β-1,3-glucanase and chitosanase activities produced by JKTJ-3
One mL spore suspension (1 × 107 spores/mL) of isolate JKTJ-3 was inoculated into 100 mL PDB, chit-PDB (PDB added with mycelial homogenate of Pa, 25 g/L), SDM (soluble starch,10 g/L; K2HPO4, 0.3 g/L; MgCO3, 1 g/L; KNO3, 1 g/L; and NaCl, 0.5 g/L), and chit-SDM (SDM added with mycelial homogenate of Pa, 25 g/L), respectively. PDB, chit-PDB, SDM, and chit-SDM added with no spore suspensions were used as controls. The spore suspension of isolate JKTJ-3 was incubated in above-mentioned 4 media in 220 rpm shaker at 28 °C for 4 days to obtain 4 fermentation cultures (FCs), and these 4 FCs were flitered with 0.22 μm syringe filters. The activities of the β-1,3-glucanase and chitinase of the resultant filtrates were respectively determined with glucanase activity assay kit (AKSU038C, Beijing Boxbio Sci. & Technol. Co., Ltd., China) and chitinase activity assay kit (AKSU045C) according to the manufacturers’ instruction. Two enzyme activities were expressed as U/mL. Each treatment was performed in triplicates.
2.8. Determination of antifungal metabolites produced by JKTJ-3
The cultural filtrate of isolate JKTJ-3 was prepared by filtering a 4-day-old PDB fermentation culture through a 0.22 μm syringe filter, and the filtrate was extracted with equal-volume ethyl acetate and vacuum dried at 40 °C to obtain colloidal crude extract [38]. As previously described [6,39], the crude extract was analyzed by Waters 2695 ultra performance liquid chromatography (UPLC) coupled with Waters Micromass Quattro Micro® mass spectrometry in positive and negative ion mode with a heated electrospray ionization(ESI). Finally, the UV absorption spectrum and mass spectrum of the active substances in crude extract was analyzed by LC-MS software (Masslynx version 4.1). The obtained information was input to the Chapman compound database (version 2003) to obtain the chemical components and structures of active substances in crude extract.
2.9. Statistical Analysis
All data were processed and analyzed with WPS Excel 2019, and then Duncan’s new complex difference method in SAS v.9.0 (SAS Institute Inc., Cary, NC, Unite State) was used to conduct the analysis of variance (ANOVA), and the significant differences between different treatments in each experiment was compared at the level of p < 0.05.
3. Results
3.1. In vitro screening of antagonists
After 48-h culture, all the candidate bacterial isolates did not form an inhibition zone. All the candidate actinomycete isolates exhibited inhibitory effect on Pa (Figure 1). Among them, actinomycete isolate JKTJ-3 displayed the strongest antagonistic effect on Pa, with an inhibition zone width of 23.2 mm (Figure A1A), followed by isolates V61b, V10a, and H55-1a. The width of inhibition zone produced by these four isolates was significantly higher than that produced by other 6 isolates (p < 0.05). Therefore, the actinomycete isolates JKTJ-3, V61b, V10a, and H55-1a were selected for the subsequent determination of the antifugal spectrum. Isolate JKTJ-3 had the strongest inhibitory effect against 12 pathogenic fungi with the inhibition zone width ranging from 11.2 mm to 22.8 mm and an average inhibition zone width of 16.2 mm (Table 1, Figure A2). The average inhibition zone width of isolate V61b was 10.0 mm, which was inferior to that of isolate JKTJ-3. The antagonistic effect of actinomycete isolates V10a and H55-1a was relatively weak. Therefore, actinomycete isolates JKTJ-3 and V61b were selected for subsequent rescreening in vivo.
3.2. In vivo screening of biocontrol agents
The results showed an obvious decrease in the damping-off incidence under isolate JKTJ-3 treatment which was significantly lower than that under hymexazol and V61b treatments (p < 0.05) (Figure 2A, Figure A1B). Moreover, the seedling rate (SR) of isolate JKTJ-3 was also significantly higher than that of hymexazol and V61b treatments (p < 0.05) (Figure 2B). These results suggested that isolate JKTJ-3 had the potential for the biocontrol of watermelon damping-off caused by Pa, and thus this isolate was chosen for subsequent experiments.
3.3.Taxonomic identification of isolate JKTJ-3
Isolate JKTJ-3 could grow normally on media of ISP1-6, GS-1 and BM (Table 2). The aerial spore mass colors of this isolate presented gray-white, smoky pink (Figure 3A), and gray-brown on ISP 2-5 and BM media, but no aerial spore mass was formed on ISP-1, ISP-6, and GS-1media. The substrate hyphae of isolate JKTJ-3 were relatively straight with few branches (Figure 3B), and presented yellow and orange, and produced yellow and orange soluble pigment. The spore chains of this isolate were spiral (Figure 3C) and its spores were short rod-shaped or subglobose with the diameter of less than 1 µm and surface ridges (Figure 3D). The isolate JKTJ-3 could make use of D-glucose, sucrose, D-fructose, D-xylose, L-rhamnose, raffinose, L-arabinose, and D-mannitol, and this isolate could grow at 8% NaCl (w/v) or less. It could grow at the minimum pH of 3.5 with temperature range of 15 °C-43 °C. Other cultural, physiological, and biochemical characteristics were were shown in Table 2. The morphological and physiological features appeared similar to those of S. murinus.
To investigate taxonomic characteristics of isolate JKTJ-3, the fragment of 16S rDNA was sequenced, and the obtained nucleotide sequence was submitted to GenBank database (accession number: OK271440). The 26 type isolates with high homology and 1 outgroup isolate were downloaded from NCBI website, and a phylogenetic tree was constructed based on their 16S rDNA gene alignments with MEGA X software (Figure 4). The phylogenetic analysis demonstrated that isolate JKTJ-3 has a close relationship with S. murinus as well as with S. costaricanus, S. graminearus, and S. phaeogriseichromatogenes. Taking the results about morphological/physiological features and molecular phylogeny togather, isolate JKTJ-3 more likely belongs to S. murinus.
3.4. Biocontrol efficacy of JKTJ-3 by treatment of seeding substrate
To reveal whether there were differences in the biocontrol efficacy between different forms and different contents of JKTJ-3’s active ingredients, biocontrol efficacy of different concentrations of JKTJ-3 cultural filter (CF) and wheat grain culture (WGC) were determined by substrate treatment. Biocontrol efficacy of isolate JKTJ-3 CF was shown in Figure 5. The emergence rates of watermelon seedlings (Figure 5A) were increased and then decreased with the increase in the dosages of the JKTJ-3 CF from 10 mL/L to 300 mL/L, of which two dosages (100 and 150 mL/L) exhibited obviously higher emergence rate than the remaining dosages (p < 0.05). Seedling rate (Figure 5B) and seed emergence rate displayed similar trend, but 150 mL/L and 200 mL/L CF treatments showed significantly higher seedling rate than other treatments (p < 0.05). The damping-off incidence of watermelon seedlings (Figure 5C) were decreased and then increased with the increasing JKTJ-3’s CF, of which 150 mL/L and 200 mL/L CF treatments exhibited obviously lower damping-off incidence than other treatments (p < 0.05). The biocontrol efficacy (Figure 5D) under 150 mL/L CF treatment was 51.4%, which was significantly higher than 45.1% under 200 mL/L CF treatment, and biocontrol efficacy of these two dosages (150 and 200 mL/L) was significantly better than other treatments (p < 0.05). In addition, the biocontrol efficacys of JKTJ-3’s CF within 50-200 mL/L concentration range were much better than that of hymexazol treatment (p < 0.05).
There were differences in seed emergence rate, seedling rate, control efficacy, and damping-off incidence between JKTJ-3 CF and WGC. The seed emergence rates (Figure 6A) under the WGC treatments from 10 g/L to 160 g/L were higher than 91%, and the 160 g/L WGC treatment exhibited the minimum seed emergence rate, but 10-160 g/L WGC treatments exhibited no significant difference from the control in seed emergence rate. Seedling rates (Figure 6B) increased as the concentrations of WGC increased. The seedling damping-off incidences (Figure 6C) decreased with the increasing concentrations of WGC. Under 160 g/L WGC treatment, the damping-off incidence and seedling rate were 6.7% and 84.9%, respectively, both of which significantly different from those under other treatments (p < 0.05). The control efficacy of WGC treatment (Figure 6D, Figure A3) was within the range from 53.2% to 92.4%, which was significantly higher than that of hymexazol treatment (p < 0.05). The control efficacy of 160 g/L WGC treatment showed the best control efficacy against watermelon damping-off.
3.5. Protective efficacy of JKTJ-3 by treatment of seeding substrate
We further examined the protective effects of JKTJ-3 CF and WGC against Pa damping-off disease by substrate treatment. It could be seen from Table 3 that the emergence rate, seedling rate, and control efficacy of watermelon seedlings with JKTJ-3 CF by substrate treatment increased when the Pa inoculation interval time became longer, while damping-off incidence decreased. The lowest damping-off incidence, highest emergence rate and highest seedling rate were 34.7%, 95% and 62.0%, respectively, which was 40.7% lower, 13.7% higher, and 82.4% higher than that of simultaneous inoculation of JKTJ-3 CF and P. aphanidermatum (0 day interval), respectively. At the interval of 1-3 days, JKTJ-3 CF treatment exhibited the optimal damping-off incidence, seedling rate, and protectiveefficacy. Compared with that of fungicide hymexazol WP, the protectiveefficacy of JKTJ-3 CF at the interval of 1 d, 2 d and 3 d increased by 25.8%, 122.3%, and 614.6%, respectively.
Similar change trend was observed in WGC substrate treatment (Table 4, Figure A4). The highest seed emergence rate and highest seedling rate were 96.7% and 94.3% respectively, which was 9.5% higher and 36.3%% higher than that of simultaneous inoculation of JKTJ-3 WGC and P. aphanidermatum (0 day interval), respectively. Moreover, no seedling damping-off was observed under WGC treatment at the interval of 3 days. The damping-off incidence under WGC treatment at different inoculation intervals was significantly lower than that under control treatment and hymexazol WP treatment, while the seedling rate was significantly higher (p < 0.05). In the interval of 0-3 days, the protective effects under WGC treatment at different intervals was 36.7%-227.9% significantly higher than that under hymexazol WP treatment (p < 0.05). These findings suggested that Streptomyces JKTJ-3 CF and WGC treatments had a good protective effect on watermelon damping-off, and WGC exhibited better effect than CF.
3.6. Biocontrol efficacy of isolate JKTJ-3 by seed treatment
Further, we investigated the potential biocontrol efficacy of Streptomyces JKTJ-3 FC (fermentation culture) and CF (cultural filtrate) by seed treatment (Table 5). Compared with the control, seed treatment with JKTJ-3 FC and CF significantly (p < 0.05) reduced the damping-off incidence, but increased the seedling rate. The biocontrol efficacy of CF seed treatment ranged from 32.1% to 50.7%, while that of FC treatment was 14.7%-39.2%. In general, watermelon seed soaking treatment with both JKTJ-3 CF and FC could protect watermelon seedling against the pathogen P. aphanidermatumthe, but the control efficacy of JKTJ-3 CF was superior to that of JKTJ-3 FC.
3.7. Biocontrol mechanisms of isolate JKTJ-3
The activities of β-1,3-glucanase (Figure 7A) and chitinase (Figure 7B) were detected, and the results showed that after isolate JKTJ-3 was cultured in PDB medium and chit-PDB medium respectively, the activity of β-1,3-glucanase in PDB medium was 6.39 U/ml, and that of chitinase in chit-PDB medium was 4.07 U/mL, which were higher than those in SDM medium and chit-SDM medium, respectively.
The active compounds of JKTJ-3 ethyl acetate crude extract were detected by LC-MS. The results showed that a compound at 6.62 min showed a strong ultra violet (UV) absorption peak (Figure 8A) corresponding to a characteristic absorption peak at 442 nm (Figure 8B). In the mode diagram of positive and negative ion flow, the two corresponding mass spectra peaks at 1255.64 m/z (Figure 8C) and 1256.29 m/z (Figure 8D) exhibited high abundances (M-1 = 1255.64, M + Na (23) = 1256.29). According to the mass spectrum rules, the molecular weight of the compound was about 1256.0. The molecular weight and UV absorption peak wavelength of the active compounds were input to Chapman Database, and a polypeptide antibiotic with similar properties to actinomycin D was obtained. The molecular formula and molecular weight of this compound was C62H86N12O16 and 1255.42, respectively, and a characteristic absorption peak was observed at the wavelength of 442 nm, which was consistent with the mass spectrometry analysis data. The above results indicated that actinomycin D was one of the active substances of JKTJ-3 ethyl acetate crude extract.
4. Discussion
Some studies have reported the application of microorganisms to the biological control of Pythium damping-off [20,40,41]. In this study, we obtained actinomycete isolate JKTJ-3 with the strongest antagonistic activity against important pathogenic groups causing watermelon damping-off such as Pa, R. solani, F. oxysporum, and F. solani. Our in vivo experiment data showed that actinomycete isolate JKTJ-3 could prevent the watermelon seedlings from Pa-induced damping-off disease. Thus, actinomycete isolate JKTJ-3 would be a potential biocontrol agent and deserve further study on its accurate identification.
Based on 16S rDNA sequences of genus Streptomyces, it is difficult to accurately clarify isolate JKTJ-3 as one of the four reported species, including S. costaricanus, S. murinus, S. graminearus, and S. phaeogriseichromatoge. This result was in line with that previous reports [42,43]. Furthermore, we separately discussed the morphological /physiological characteristics among four species. According to taxonomic characteristics described in Manual of Streptomyces Identification [44], the isolate S. graminearus does not produce soluble pigments in GS-1 medium, which was inconsistent with our findings of the isolate JKTJ-3, indicating that JKTJ-3 did not belong to S. graminearus species. Goodfellow et al. [45] have reported that the type isolate S. phaeogriseichromatogenes NRRL 2834 can not grow at 40 °C and above, and it can use neither D-raffinose nor L-rhamnose as sole carbon sources, but our data showed that isolate JKTJ-3 could grow at 43 °C, and it did utilize D-raffinose and L-rhamnose, suggesting that JKTJ-3 did not belong to S. phaeogriseichromatogenes species. et al. [46] have revealed that the type isolate S. costaricanus CR-43 (ATCC 55274) did not produce soluble pigments in ISP-3 medium or ISP-4 medium; it can not use sucrose, L-arabinose, D-raffinose, or L-rhamnose, and it can not grow in the presence of over 5% NaCl (w/v), which was quite different from our observation of isolate JKTJ-3, implying that JKTJ-3 did not belong to S. costaricanus species. As Esnard et al. [46] described, the type isolate S. murinus NRRL B-2286 can produce soluble pigments in ISP-2, ISP-3, ISP-4, and ISP-5 media, but not in ISP-6 medium; it can utilize sucrose, L-arabinose, cottonseed sugar, D-rhamnose as only carbon source; and it can grow at 7% NaCl (w/v), which was highly consistent with our findings of the isolate JKTJ-3. Based on above comparison of the four species, isolate JKTJ-3 was identified as S. murinus.
As previously reported, S. murinus contributed to produce immobilized glucose isomerase, xylose isomerase and others in Food Engineering [47,48]. Suzuki et al. [49] found a new cell aggregation factor called SAF in the mycelia of S. murinus. Fang et al. [50] reported that S. murinus had AMP deaminase genes, which could become the gene source for constructing constitutive expression of AMP deaminase. To the best of our knowledge, this is the first report of S. murinus in the biocontrol of watermelon Pa damping-off. Moreover, there are few studies on biocontrol of watermelon damping-off disease. S. murinus JKTJ-3 may become a new biocontrol agent against damping-off.
Many researchers refer to damping-off as nursery disease which is usually associated to soil- or seed-born pathogens [9] Biological seed treatment and substrate treatment are two frequently used methods for controlling Pythium damping-off [51,52,53]. In this study, comprehensive biocontrol strategy was adopted using S. murinus JKTJ-3 by seed treatment and substrate treatment with the fermentation culture (FC), cultural filtrate (CF), and wheat grain culture (WGC). Watermelon seed soaking treatment with S. murinus JKTJ-3 inhibited watermelon damping-off. The biocontrol efficacy was affected by the active compounds of JKTJ-3 and the soaking time. The biocontrol efficacy of substrate treatment was better than that of seed soaking treatment. We also found that the biocontrol efficacy of S. murinus JKTJ-3 WGC was better than that of JKTJ-3 CF.
In addition, the protective efficacy of the JKTJ-3′s cultural filtrate (CF) and wheat grain culture (WGC) was improved as Pa inoculation interval increased. As previously reported, the propagules of Pythium can propagate and colonize very rapidly under suitable temperature and humidity, thus escaping natural antagonism [54]. Our data showed that the control efficacy of substrate treatments with Pa at 1-3 d inoculation interval was better than that at simultaneous inoculation (namely, 0 d inoculation interval). In other words, early inoculation of S. murinus JKTJ-3 active ingredient, may exhibite an advantage in its colonization in the substrate, seeds and seedling, or in inhibiting or killing pathogens before pathogens propagated massively, which eventually reducing the occurance of watermelon damping-off disease. Thus, early inoculation of the biocontrol agent could increase the control efficacy of S. murinus JKTJ-3, which is consistent with Becker and Schwinn reported [55].
Currently, there have been few reports on the biocontrol mechanism of S. murinus. Antagonism, as an action mechanism of controlling plant disease, mainly depends on antibiotics to inhibit the growth and metabolism of the pathogens [56,57]. Another molecular mechanism of antagonism lies in that a variety of enzymes inhibit, degrade and hydrolyze other pathogens [58]. In this study, a large inhibition zone was found to be formed between the colonies of phytopathogenic fungi and isolate JKTJ-3, indicating that isolate JKTJ-3 could produce antibiotics. Our enzyme activity determination indicated that S. murinus JKTJ-3 did secrete β-1,3-glucanase and chitinase. β-1,3-glucanase and chitinase activities can degrade the cell wall of pathogenic fungi, which has been identified as a main biocontrol mechanism in many studies [59]. Our LC-MC analysis confirmed that S. murinus JKTJ-3 produced actinomycin D. Zajkowicz et al. [60] have reported that actinomycin D has antimicrobial and antiviral effects as well as anticancer and antiumor effects. Lei et al. [61] have revealed that S. antibioticus can strongly inhibit several pathogenic bacteria and fungi by producing actinomycin D. The result indicated that S. murinus JKTJ-3 produced multiple metabolites including chitinase, and actinomycin D, which might be part of the potential mechanism of isolate JKTJ-3′s biocontrol against watermelon damping-off disease. The specific functions of these metabolites in the biocontrol remain to be further investigated.
Overall, S. murinus JKTJ-3 efficiently inhibited several damping-off pathogens of cucurbit crops in vitro, further confirming its ability to control Pa damping-off of watermelon. The isolate JKTJ-3 produced β-1, 3-glucanase, chitinase, and actinomycin D which are probably the mechanisms for biocontrol of watermelon damping-off. S. murinus JKTJ-3 and its active ingredien as biocontrol factors are a newly discovered which need to further clarify the biocontrol mechanisms.
5. Conclusions
Importantly, our findings clearly provide valuable evidence in determining the importance and practicability to utilize S. murinus as a new promising biocontrol agent in controlling seedling damping-off.
Author Contributions
Conceptualization, Mihong Ge and Xiang Cai; Formal analysis, Huan Liang and Juhong Zhu; Investigation, Mihong Ge and Dehuan Wang; Writing—original draft, Mihong Ge; Writing—review & editing, Xianfeng Shi and Guoqing Li.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/nuccore/OK271440.
Acknowledgments
We thank the kind help from Prof. Z. Y. Wan (National Engineering Research Center for Biopesticides, Hubei Academy of Agricultural Sciences, Wuhan, China) for LC-MS analysis of actinomycin D in this work.
Conflicts of Interest
The authors declare no conflflict of interest.
Appendix A
Figure A1.
Screening of two antagonistic actinomycete isolates against Pa. A, in vitro screening of 2 candidate actinomycete isolates against Pa; B, in vivo screening of 2 antagonistic actinomycete isolates against watermelon Pa damping-off.
Figure A1.
Screening of two antagonistic actinomycete isolates against Pa. A, in vitro screening of 2 candidate actinomycete isolates against Pa; B, in vivo screening of 2 antagonistic actinomycete isolates against watermelon Pa damping-off.
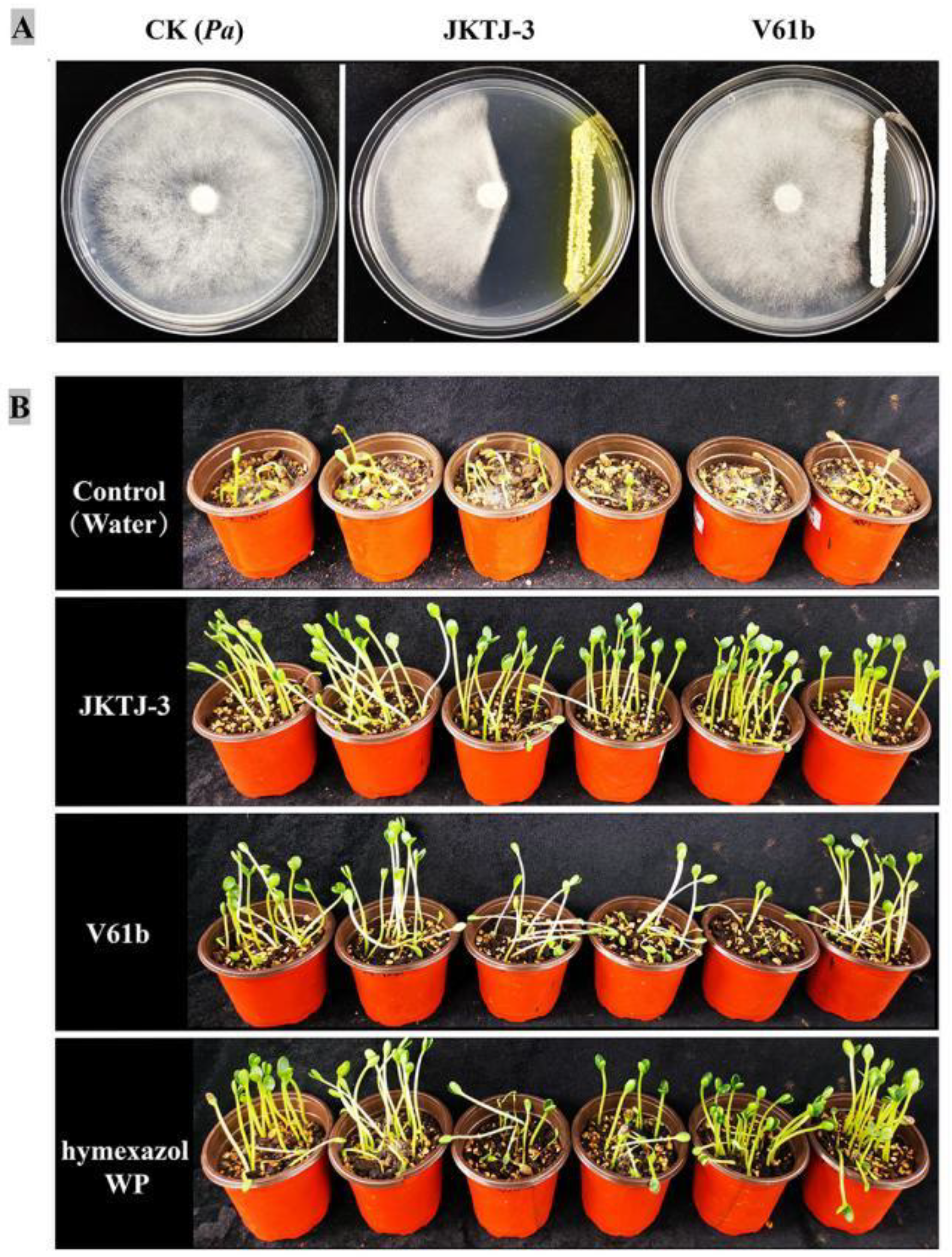
Figure A2.
Antagonistic effects of candidate actinomycete isolate JKTJ-3 on 12 phytopathogenic fungi.
Figure A2.
Antagonistic effects of candidate actinomycete isolate JKTJ-3 on 12 phytopathogenic fungi.

Figure A3.
The biocontrol effects of different contents of isolate JKTJ-3 wheat grain culture (WGC) by treatment of seeding substrate.
Figure A3.
The biocontrol effects of different contents of isolate JKTJ-3 wheat grain culture (WGC) by treatment of seeding substrate.

Figure A4.
The control efficacy of isolate JKTJ-3 wheat grain culture (WGC) by substrate treatments with Pa at 0-3 d inoculation interval.
Figure A4.
The control efficacy of isolate JKTJ-3 wheat grain culture (WGC) by substrate treatments with Pa at 0-3 d inoculation interval.

References
- Tojo, M.; Fujita, Y.; Awad, H.M.; Ichitani, T. Preparation of Pythium Inocula Using Bentgrass Seeds for Glasshouse Studies. Annual Report of the Kansai Plant Protection Society 1993, 35, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Georgakopoulos, D.G.; Fiddaman, P.; Leifert, C.; Malathrakis, N.E. Biological Control of Cucumber and Sugar Beet Damping-off Caused by Pythium ultimum with Bacterial and Fungal Antagonists. J. Appl. Microbiol Appl. 2010, 92, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Haenseler, C.M. Control of Seed Decay and Damping-off of Cucumbers. New Jersey Agric. 1930, 254–264. [Google Scholar]
- Burgieł, Z.J. Research on Possibilities of Utilization of Chosen Brassicaceae Plants in Protection of Cucumber against Damping-off Caused by Rhizoctonia solani Kuhn and Fusarium culmorum (W.G. Smith) Sacc. J. Polish. Bot. Soc. 2005, 58, 171–178. [Google Scholar] [CrossRef]
- Al-Fadhal, F.A.; AL-Abedy, A.N.; Alkhafije, D.A. Isolation and Molecular Identification of Rhizoctonia solani and Fusarium solani Isolated from Cucumber (Cucumis sativus L.) and Their Control Feasibility by Pseudomonas fluorescens and Bacillus subtilis. Egypt. J. Biol. Pest Control 2019, 29, 1–11. [Google Scholar] [CrossRef]
- Wan, Z.Y.; Fang, W.; Wu, Z.Y.; Zhang, Y.N.; Zhang, Z.G.; Wang, K.M.; et al. Isolation and Identification of Active Substances from Broad-spectrum Antifungal Streptomyces HBERC-57169. Biotic Resources 2019, 41, 516–523, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Mckeen, C.D. Arasan as a Seed and Soil Treatment for the Control of Damping-off in Certain Vegetables. Sci. Agri. 1950, 30, 261–270. [Google Scholar] [CrossRef]
- Arora, H.; Sharma, A.; Sharma, S.; Haron, F.F.; Datta, R. Pythium Damping-Off and Root Rot of Capsicum annuum L.: Impacts, Diagnosis, and Management. Microorganisms 2021, 9, 823. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Dürr, C.; Schwanck, A.A.; Robin, M.-H.; Sarthou, J.-P.; Cellier, V.; Messéan, A.; Aubertot, J.-N. Integrated Management of Damping-off Diseases. A Review. Agon. Sustain. Dev. 2017, 37, 1–25. [Google Scholar] [CrossRef]
- Abdelrahim, A.M.; Abusurrieh, A.A. Chemical Control of Pythium Debaryanum, the Causal Organism of Damping-off in Cucumber. Arab. J. Plant Prot. 1991, 1, 14–18. [Google Scholar]
- Palakshappa, M.G.; Lokesh, M.S.; Parameshwarappa, K.G. Efficacy of Ridomil Gold (Metalaxyl M+Mancozeb (4+64 WP)) against Chilli Damping off Caused by Pythium aphanidermatum. Karnataka J. Agric. Sci. 2010, 23, 445–446. [Google Scholar]
- Begum, S.; Devi, R.; Singh, N.I. Management of Damping-off of Vegetable Seedlings Caused by Rhizoctonia solani. Plant Protection Association of India 2014, 42, 248–254. [Google Scholar]
- Cohen, Y.; Reuveni, M. Occurrence of Metalaxyl-Resistant Isolates of Phytophthora infestans in Potato Fields in Israel. Phytopathology 1983, 73, 925–927. [Google Scholar] [CrossRef]
- White, J.G.; Stanghellini, M.E.; Ayoubi, L.M. Variation in the Sensitivity to Metalaxyl of Pythium spp. Isolated from Carrot and Other Sources. Ann. Appl. Biol. 2010, 113, 269–277. [Google Scholar] [CrossRef]
- Ma, Z.; Michailides, T.J. Advances in Understanding Molecular Mechanisms of Fungicide Resistance and Molecular Detection of Resistant Genotypes in Phytopathogenic Fungi. Crop Prot. 2005, 24, 853–863. [Google Scholar] [CrossRef]
- Hwangbo, H.; Kim, K.-Y.; Choi, H.-S. Effects of Biocontrol Agents on Suppression of Damping-off in Cucumis sativus L. Caused by Rhizoctonia Solani. Hortic. Environ. Biote. 2016, 57, 191–196. [Google Scholar] [CrossRef]
- Roberts, D.P.; Selmer, K.; Lupitskyy, R.; Rice, C.; Buyer, J.S.; Maul, J.E.; Lakshman, D.K.; DeSouza, J. Seed Treatment with Prodigiosin Controls Damping-off of Cucumber Caused by Pythium ultimum. AMB Express 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Sadeghi, A.; Koobaz, P.; Azimi, H.; Karimi, E.; Akbari, A.R. Plant Growth Promotion and Suppression of Phytophthora drechsleri Damping-off in Cucumber by Cellulase-Producing Streptomyces. BioControl 2017, 62, 805–819. [Google Scholar] [CrossRef]
- Awad, L.K.; Fayyadh, M.A. The Activity of Some Actinomycetes Isolates in Control of Cucumber Damping off Disease Caused by Rhizoctiona solani and Pythium sp. Basrah J. Agr. Sci. 2018, 31, 11–23. [Google Scholar] [CrossRef]
- El-Tarabily, K.A. Rhizosphere-Competent Isolates of Streptomycete and Non-Streptomycete Actinomycetes Capable of Producing Cell-Wall-Degrading Enzymes to Control Pythium aphanidermatum Damping-off Disease of Cucumber. Botany 2006, 84, 211–222. [Google Scholar] [CrossRef]
- Roberts, D.; McKenna, L.; Lakshman, D.; Meyer, S.; Kong, H.; De Souza, J.; Lydon, J.; Baker, C.; Buyer, J.; Chung, S. Suppression of Damping-off of Cucumber Caused by Pythium ultimum with Live Cells and Extracts of Serratia marcescens N4-5. Soil Biol. Biochem. 2007, 39, 2275–2288. [Google Scholar] [CrossRef]
- Huang, X.; Chen, L.; Ran, W.; Shen, Q.; Yang, X. Trichoderma harzianum Strain SQR-T37 and Its Bio-Organic Fertilizer Could Control Rhizoctonia solani Damping-off Disease in Cucumber Seedlings Mainly by the Mycoparasitism. Appl. Microb. Biot. 2011, 91, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Valois, D.; Fayad, K.; Barasubiye, T.; Garon, M.; Dery, C.; Brzezinski, R.; Beaulieu, C. Glucanolytic Actinomycetes Antagonistic to Phytophthora fragariae Var. Rubi, the Causal Agent of Raspberry Root Rot. Appl. Environ. Microb. 1996, 62, 1630–1635. [Google Scholar] [CrossRef] [PubMed]
- Samac, D.A.; Willert, A.M.; McBride, M.J.; Kinkel, L.L. Effects of Antibiotic-Producing Streptomyces on Nodulation and Leaf Spot in Alfalfa. Appl. Soil Ecol. 2003, 22, 55–66. [Google Scholar] [CrossRef]
- Hata, E.M.; Yusof, M.T.; Zulperi, D. Induction of Systemic Resistance against Bacterial Leaf Streak Disease and Growth Promotion in Rice Plant by Streptomyces shenzhenesis TKSC3 and Streptomyces sp. SS8. Plant Pathology. J. 2021, 37, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Baharlouei, A.; Sharifi-Sirchi, G.R.; Shahidi, B.G.H. Identification of an Antifungal Chitinase from a Potential Biocontrol Agent, Streptomyces plicatus Strain 101, and Its New Antagonistic Spectrum of Activity. Philipp. Agric. Sci. 2010, 93, 439–445. [Google Scholar] [CrossRef]
- Njoroge, S.M.; Riley, M.B.; Keinath, A.P. Effect of Incorporation of Brassica spp. Residues on Population Densities of Soilborne Microorganisms and on Damping-off and Fusarium Wilt of Watermelon. Plant Dis. 2008, 92, 287–294. [Google Scholar] [CrossRef]
- Al-Sa’di, A.M.; Drenth, A.; Deadman, M.; De Cock, A.W.A.M.; Aitken, E. Molecular Characterization and Pathogenicity of Pythium Species Associated with Damping-off in Greenhouse Cucumber (Cucumis Sativus) in Oman. Plant Patho. 2007, 56, 140–149. [Google Scholar] [CrossRef]
- Ge, M.H. Identification of the pathogen of vegetable Pythium damping-off and screening and application of novel high-efficiency biocontrol agents. [D]. Huazhong Agricultural University: Wuhan, China, 2022. [Google Scholar]
- Shakeel, Q.; Lyu, A.; Zhang, J.; Wu, M.; Li, G.; Hsiang, T.; Yang, L. Biocontrol of Aspergillus flavus on Peanut Kernels Using Streptomyces yanglinensis 3-10. Frontiers microbiol. 2018, 9, 1049. [Google Scholar] [CrossRef]
- Paul, B.; Charles, R.; Bhatnagar, T. Biological Control of Pythium mamillatum Causing Damping-off of Cucumber Seedlings by a Soil Bacterium, Bacillus Mycoides. Microbiol. Res. 1995, 150, 71–75. [Google Scholar] [CrossRef]
- Li, B.; Ravnskov, S.; Xie, G.; Larsen, J. Biocontrol of Pythium Damping-off in Cucumber by Arbuscular Mycorrhiza-Associated Bacteria from the Genus Paenibacillus. Biocontrol 2007, 52, 863–875. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Pridham, T.; Gottlieb, D. The Utilization of Carbon Compounds by Some Actinomycetales as an Aid for Species Determination. J. Bacteriol. 1948, 56, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Shirling, E.B.; Gottlieb, D. Methods for Characterization of Streptomyces Species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef]
- Jones, K.L. Fresh Isolates of Actinomycetes in Which the Presence of Sporogenous Aerial Mycelia Is a Fluctuating Characteristic. J. Bacteriol. 1949, 57, 141–145. [Google Scholar] [CrossRef]
- Shakeel, Q.; Lyu, A.; Jing, Z.; Wu, M.; Chen, S.; Chen, W.; Li, G.; Long, Y. Optimization of the Cultural Medium and Conditions for Production of Antifungal Substances by Streptomyces platensis 3-10 and Evaluation of Its Efficacy in Suppression of Clubroot Disease (Plasmodiophora brassicae) of Oilseed Rape. Biol. Control 2016, 101, 59–68. [Google Scholar] [CrossRef]
- Wan, Z.Y.; Fang, W.; Zhang, Y.N.; Zhang, Z.G.; Wu, Z.Y.; Wang, K.M.; et al. Isolation and identification of antifungal active compounds of actinomycete HBERC-39158. Hubei Agr Sci 2020, 59, 140–144, (In Chinese with English abstract). [Google Scholar]
- Aljarah, N. The Activity of Metarhizium sp. to Control Pythium aphanidermatum Causal Agent of Cucumber Damping off under Greenhouse Conditions. Int. J. Sci. Res. 2017, 6, 1098–1101. [Google Scholar] [CrossRef]
- Halo, B.A.; Al-Yahyai, R.A.; Maharachchikumbura, S.S.; Al-Sadi, A.M. Talaromyces Variabilis Interferes with Pythium aphanidermatum Growth and Suppresses Pythium-Induced Damping-off of Cucumbers and Tomatoes. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Labeda, D.; Goodfellow, M.; Brown, R.; Ward, A.; Lanoot, B.; Vanncanneyt, M.; Swings, J.; Kim, S.-B.; Liu, Z.; Chun, J.; et al. Phylogenetic Study of the Species within the Family Streptomycetaceae. Antonie Van Leeuwenhoek 2012, 101, 73–104. [Google Scholar] [CrossRef]
- Komaki, H. Reclassification of Streptomyces costaricanus and Streptomyces phaeogriseichromatogenes as Later Heterotypic Synonyms of Streptomyces murinus. Int. J. Syst. Evol. Microbiol. 2021, 71, 004638. [Google Scholar] [CrossRef]
- Actiomycete Taxonmic Group; Institute of Microbiology of Chinese Academy of Sciences. Manual of Streptomyces Identification; Science Press: Beijing, China, 1975; p. 336. [Google Scholar]
- Goodfellow, M.; Kumar, Y.; Labeda, D.P.; Sembiring, L. The Streptomyces violaceusniger Clade: A Home for Streptomycetes with Rugose Ornamented Spores. Antonie van Leeuwenhoek 2007, 92, 173–199. [Google Scholar] [CrossRef]
- Esnard, J.; Potter, T.L.; Zuckerman, B.M. Streptomyces costaricanus sp. nov., Isolated from Nematode-Suppressive Soil. Int. J. Syst. Bacteriol. 1995, 45, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, O.; Karlsen, L.; Nielsen, N.; Pedersen, S.; Rugh, S. A New Immobilized Glucose Isomerase with High Productivity Produced by a Strain of Streptomyces murinus. Starch-Stärke 1988, 40, 307–313. [Google Scholar] [CrossRef]
- Pastinen, O.; Visuri, K.; Schoemaker, H.E.; Leisola, M. Novel Reactions of Xylose Isomerase from Streptomyces rubiginosus. Enzyme Microb. Tech. 1999, 25, 695–700. [Google Scholar] [CrossRef]
- Suzuki, K.; Kosai, M.; Yokomizo, K.; Uyeda, M.; Shibata, M. SAF, a New Cell Aggregation Factor Produced by Streptomyces murinus Strain No. A-2805. Agri. Biol. Chem. 1987, 51, 3017–3025. [Google Scholar] [CrossRef]
- Fang, W.; Zhang, L.; Gu, Z.; Ding, C.; Shi, G. Constitutive Expression of AMP Deaminase from Streptomyces murinus in Pichia pastoris GS115 Using the GAP Promoter. Microbiol Chin 2014, 10, 2022–2028. [Google Scholar] [CrossRef]
- Bowers, J.; Parke, J.L. Epidemiology of Pythium Damping-off and Aphanomyces Root Rot of Peas after Seed Treatment with Bacterial Agents for Biological Control. Phytopathology 1993, 83, 1466–1473. [Google Scholar] [CrossRef]
- Lewis, J.; Larkin, R.; Rogers, D. A Formulation of Trichoderma and Gliocladium to Reduce Damping-off Caused by Rhizoctonia solani and Saprophytic Growth of the Pathogen in Soilless Mix. Plant Dis. 1998, 82, 501–506. [Google Scholar] [CrossRef]
- Roberts, D.P.; Lakshman, D.K.; McKenna, L.F.; Emche, S.E.; Maul, J.E.; Bauchan, G. Seed Treatment with Ethanol Extract of Serratia marcescens Is Compatible with Trichoderma Isolates for Control of Damping-off of Cucumber Caused by Pythium ultimum. Plant Dis. 2016, 100, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Callan, N.W.; Mathre, D.; Miller, J.B. Bio-Priming Seed Treatment for Biological Control of Pythium ultimum Preemergence Damping-off in Sh-2 Sweet Corn. Plant Dis. 1990, 74, 368–372. [Google Scholar] [CrossRef]
- Becker, J.O.; Schwinn, F.J. Control of Soil-Borne Pathogens with Living Bacteria and Fungi: Status and Outlook. Pesticide Sci. 1993, 37, 355–363. [Google Scholar] [CrossRef]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes Benefaction Role in Soil and Plant Health. Microb. pathogenesis 2017, 111, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Liotti, R.G.; da Silva Figueiredo, M.I.; Soares, M.A. Streptomyces griseocarneus R132 Controls Phytopathogens and Promotes Growth of Pepper (Capsicum Annuum). Biol. Control 2019, 138, 104065. [Google Scholar] [CrossRef]
- Hong, T.-Y.; Cheng, C.-W.; Huang, J.-W.; Meng, M. Isolation and Biochemical Characterization of an Endo-1,3-β-Glucanase from Streptomyces sioyaensis Containing a C-Terminal Family 6 Carbohydrate-Binding Module That Binds to 1,3-β-Glucan. Microbiology 2002, 148, 1151–1159. [Google Scholar] [CrossRef]
- Dukare, A.S.; Paul, S.; Nambi, V.E.; Gupta, R.K.; Singh, R.; Sharma, K.; Vishwakarma, R.K. Exploitation of Microbial Antagonists for the Control of Postharvest Diseases of Fruits: A Review. Crit. Rev. Food Sci. 2019, 59, 1498–1513. [Google Scholar] [CrossRef]
- Zajkowicz, A.; Gdowicz-Kłosok, A.; Krześniak, M.; Ścieglińska, D.; Rusin, M. Actinomycin D and Nutlin-3a Synergistically Promote Phosphorylation of P53 on Serine 46 in Cancer Cell Lines of Different Origin. Cellular signalling 2015, 27, 1677–1687. [Google Scholar] [CrossRef]
- Lei, J.; Duan, J.; Ma, H.; Li, J.; Li, H.; Yang, Z. Screening, Identification and Optimized Fermentation Condition of an Actinomycete Strain against Pseudomonas olanacearum. Chin. J. Appl. Environ. Biol. 2010, 16, 79–83. [Google Scholar] [CrossRef]
Figure 1.
In vitro screening of 10 candidate actinomycete isolates against Pa.
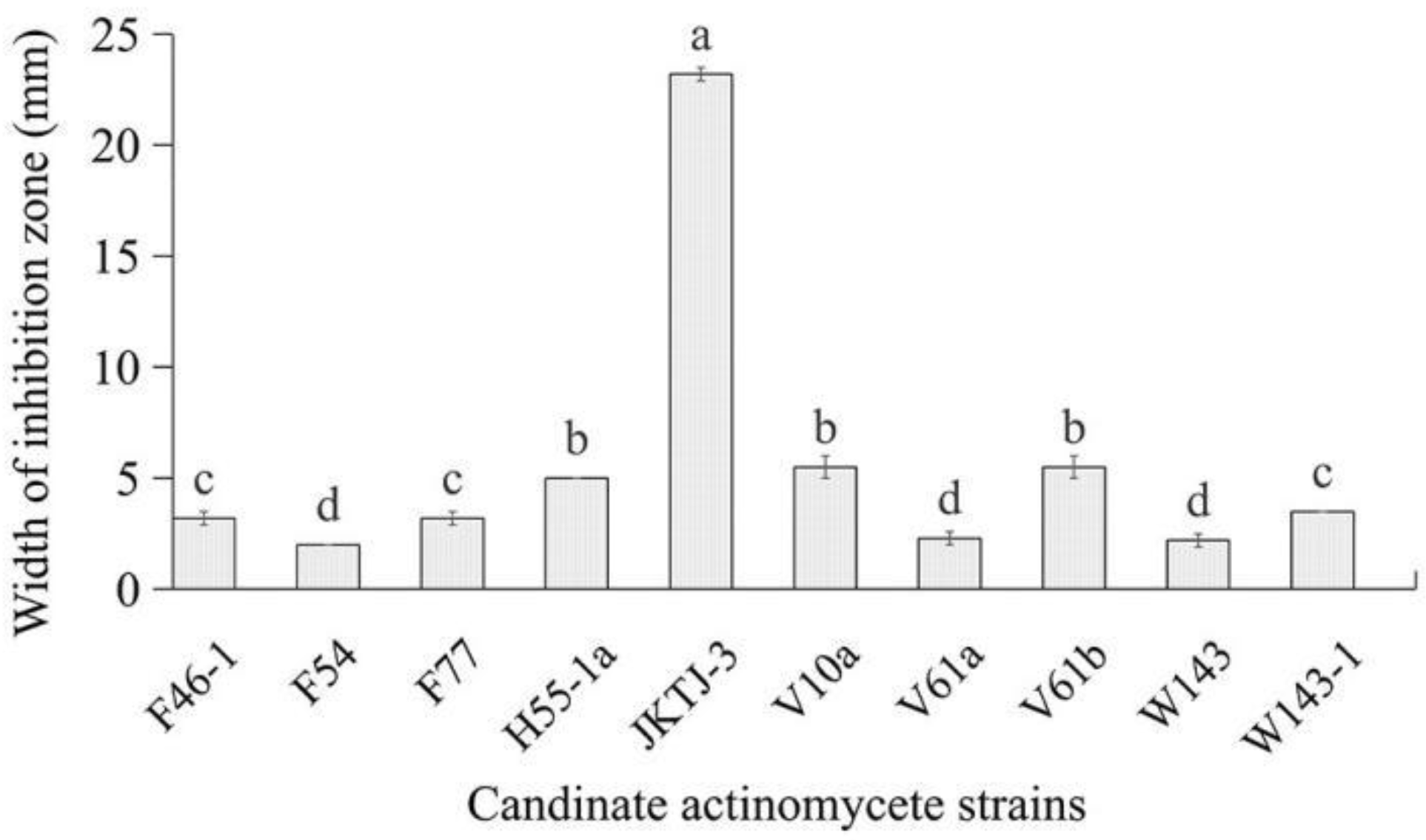
Figure 2.
In vivo screening of 2 antagonistic actinomycete isolates against watermelon Pa damping-off.
Figure 2.
In vivo screening of 2 antagonistic actinomycete isolates against watermelon Pa damping-off.
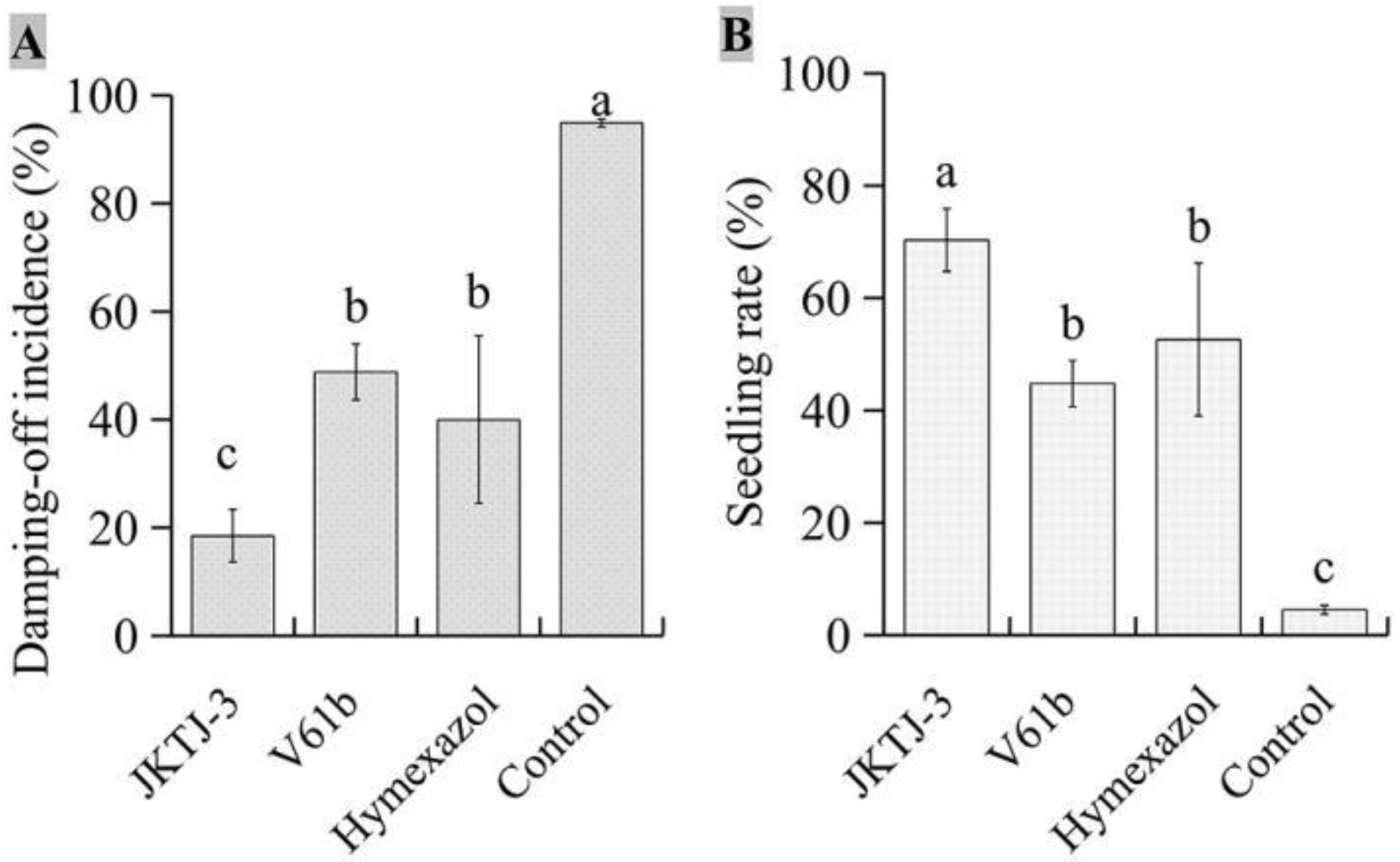
Figure 3.
The morphological characteristics of aerial hypha, spore hypha, and spore of isolate JKTJ-3 A, colony morphology (ISP-2, 28 °C, 7d); B, substrate mycelium (2 µm, SEM), few branches on the substrate mycelium; C, spiral spore (SS) chains (1 µm, SEM); D, ridged spore (RS) (200 nm, SEM).
Figure 3.
The morphological characteristics of aerial hypha, spore hypha, and spore of isolate JKTJ-3 A, colony morphology (ISP-2, 28 °C, 7d); B, substrate mycelium (2 µm, SEM), few branches on the substrate mycelium; C, spiral spore (SS) chains (1 µm, SEM); D, ridged spore (RS) (200 nm, SEM).
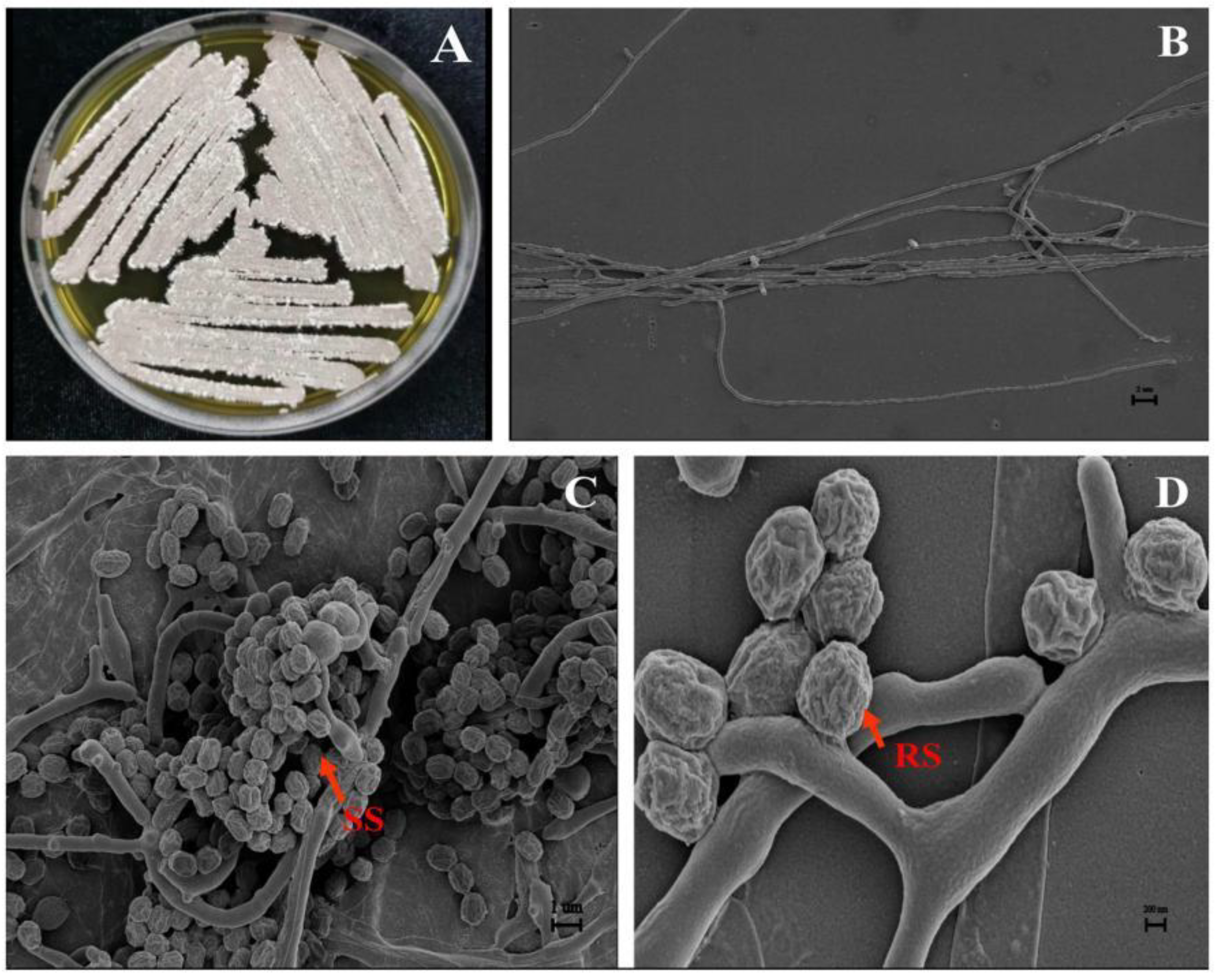
Figure 4.
Phylogenetic tree inferred by the Maximun Likelihood method based on 16S rDNA sequences. The bootstrap values of the Maximum-likelihood analysis (n = 1,000) over 50% were shown in the tree. The scale bar indicated 0.5% nucleotides variation per site.
Figure 4.
Phylogenetic tree inferred by the Maximun Likelihood method based on 16S rDNA sequences. The bootstrap values of the Maximum-likelihood analysis (n = 1,000) over 50% were shown in the tree. The scale bar indicated 0.5% nucleotides variation per site.

Figure 5.
The biocontrol effects of different concentrations of isolate JKTJ-3 cultural filtrate (CF) by treatment of seeding substrate. A, emergence rate; B, seedling rate; C, damping-off incidence; D, control efficacy.
Figure 5.
The biocontrol effects of different concentrations of isolate JKTJ-3 cultural filtrate (CF) by treatment of seeding substrate. A, emergence rate; B, seedling rate; C, damping-off incidence; D, control efficacy.
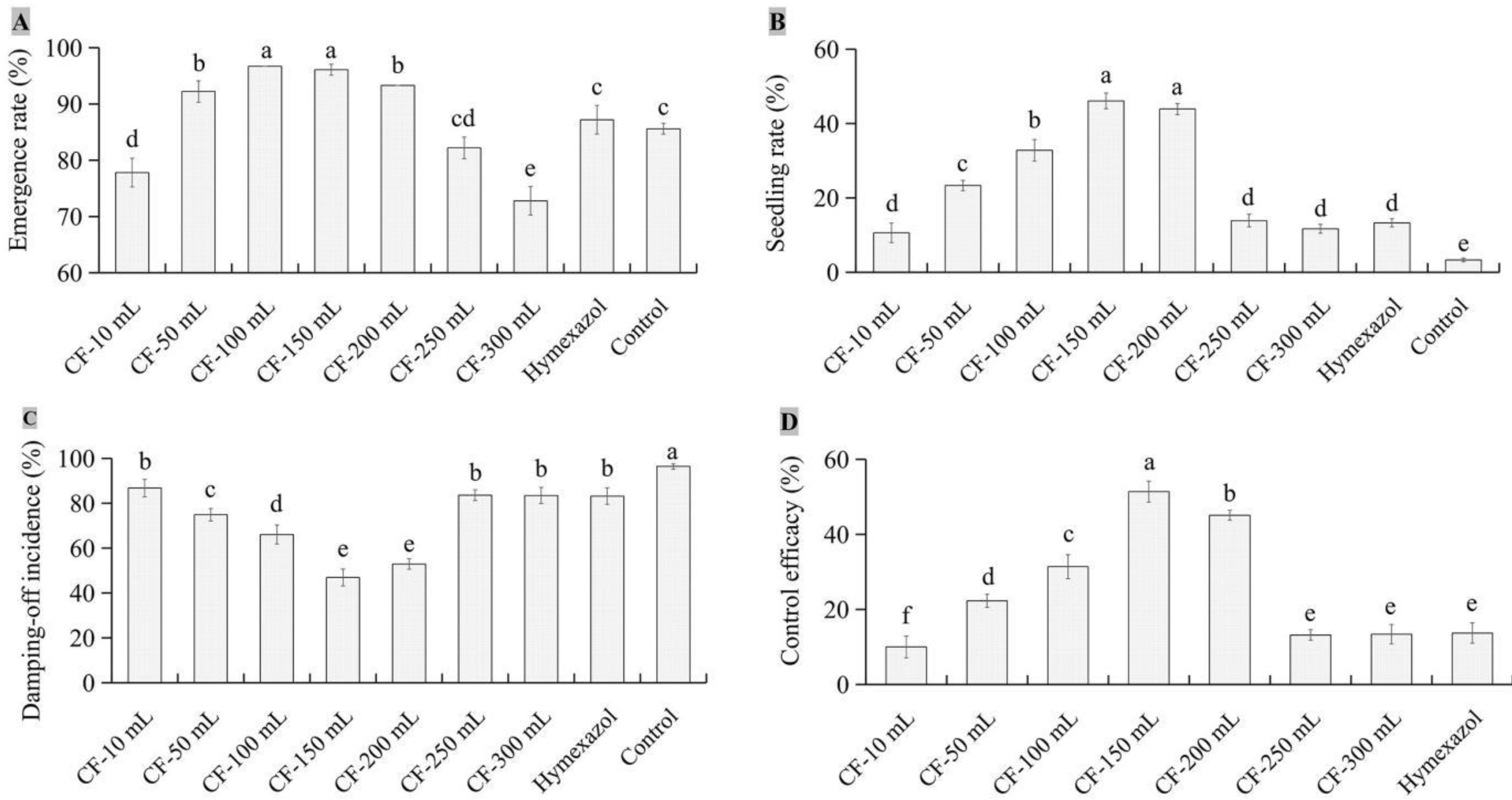
Figure 6.
The biocontrol effects of different contents of isolate JKTJ-3 wheat grain culture (WGC) by treatment of seeding substrate. A, emergence rate; B, seedling rate; C, damping-off incidence; D, control efficacy.
Figure 6.
The biocontrol effects of different contents of isolate JKTJ-3 wheat grain culture (WGC) by treatment of seeding substrate. A, emergence rate; B, seedling rate; C, damping-off incidence; D, control efficacy.
Figure 7.
Analysis of β-1,3-glucanase and chitinase activities A, β-1,3-glucanase activity; B, chitinase activity.
Figure 7.
Analysis of β-1,3-glucanase and chitinase activities A, β-1,3-glucanase activity; B, chitinase activity.
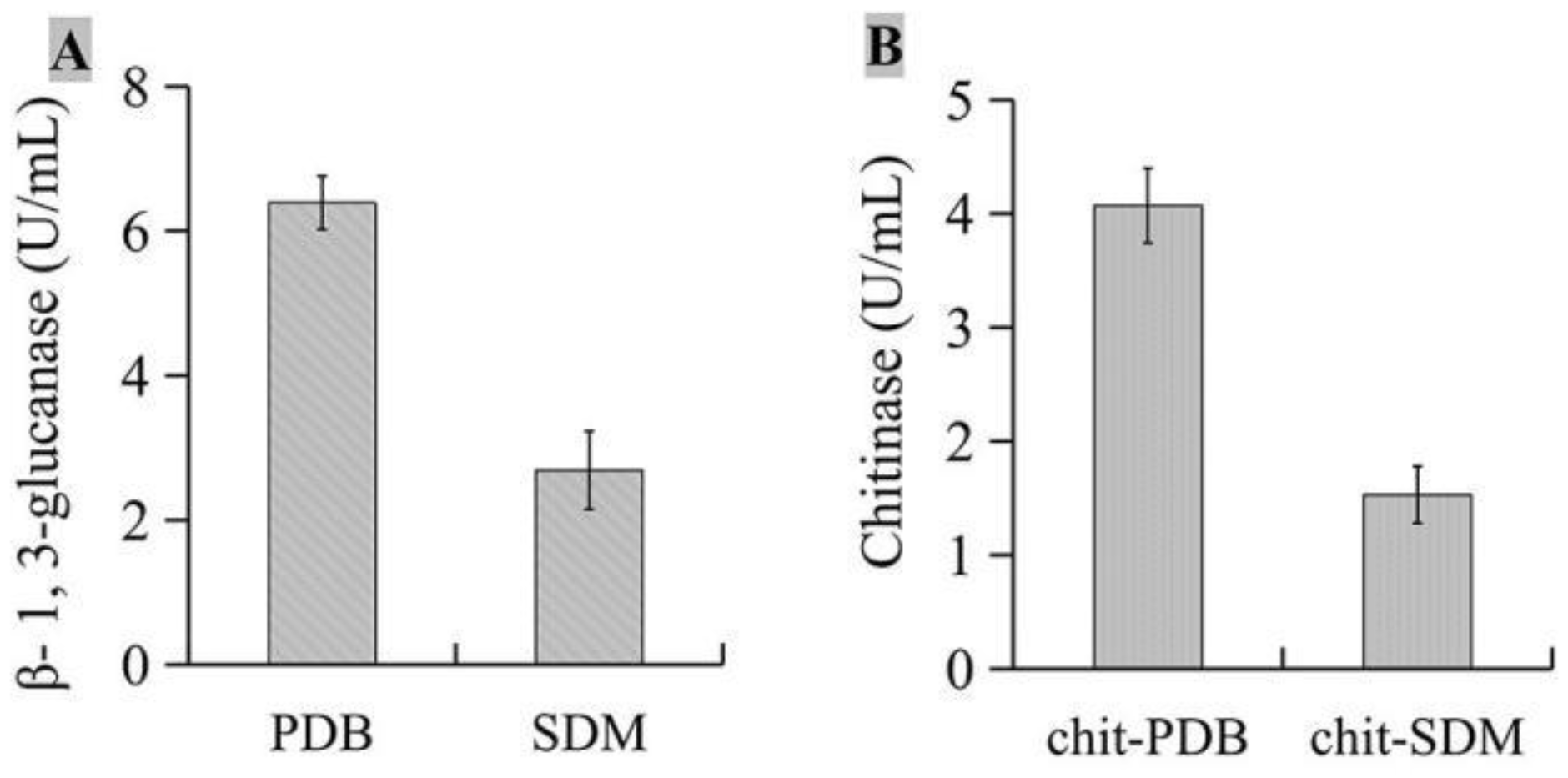
Figure 8.
Analysis of active substances of isolate JKTJ-3 ethyl acetate extract by HPLC-MS. A, HPLC-MS spectrum of ultraviolet absorption of JKTJ-3 ethyl acetate extract; B, ultraviolet absorption spectrum of an active substance; C, negative ion flow; D, positive ion flow.
Figure 8.
Analysis of active substances of isolate JKTJ-3 ethyl acetate extract by HPLC-MS. A, HPLC-MS spectrum of ultraviolet absorption of JKTJ-3 ethyl acetate extract; B, ultraviolet absorption spectrum of an active substance; C, negative ion flow; D, positive ion flow.
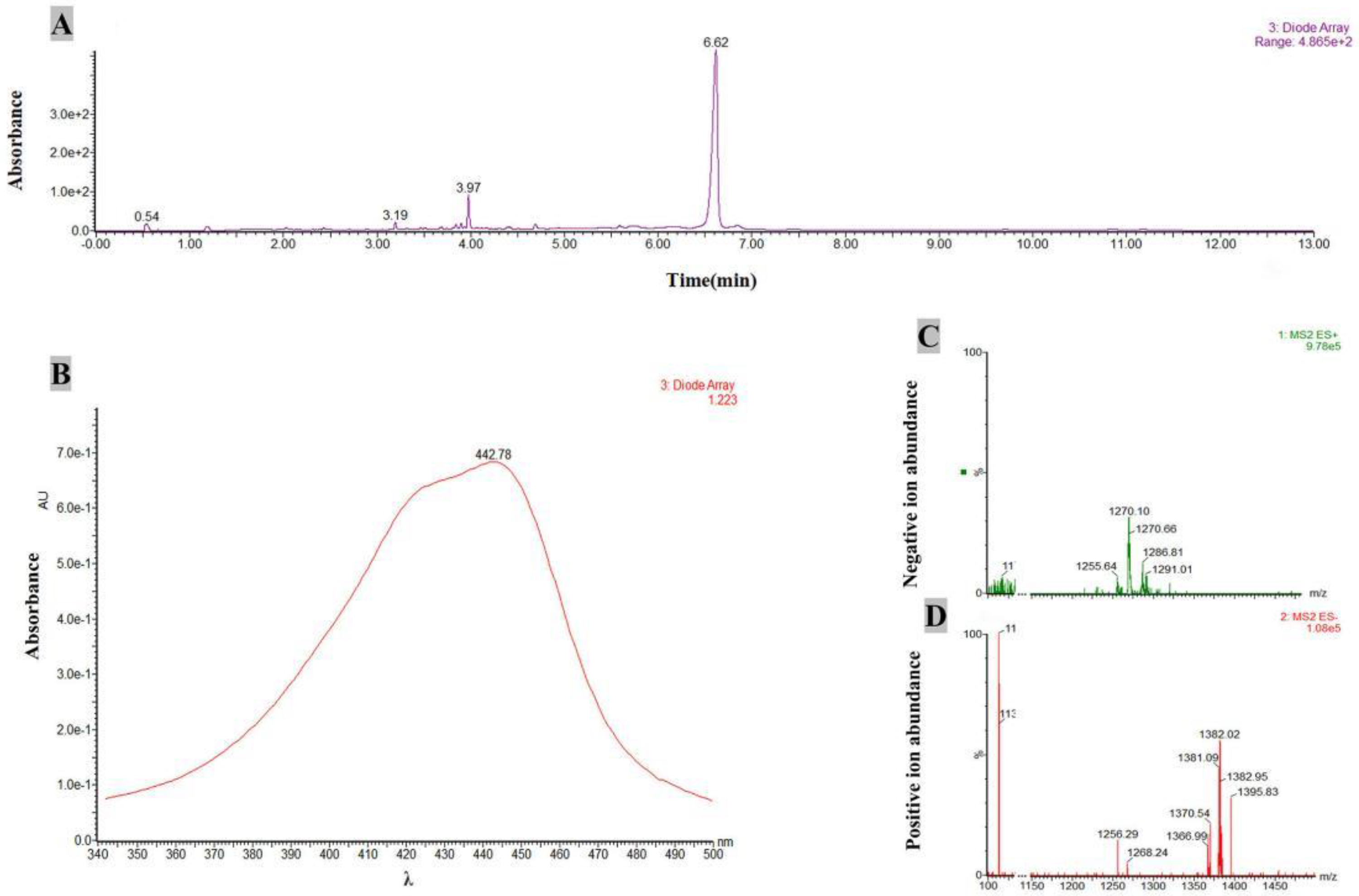

Table 1.
Comparison of 4 actinomycete isolates against 12 phytopathogenic fungi.
| Pathogen | Width of inhibition zone (mm) | |||
|---|---|---|---|---|
| JKTJ-3 | H55-1a | V10a | V61b | |
| Rhizoctonia solani | 21.0 ± 0.3a1 | 0 ± 0c1 | 0 ± 0c1 | 16.0 ± 0.2b1 |
| Stagonosporopsis cucurbitacearum | 11.3 ± 0.2a | 4.7 ± 0.5c | 5.7 ± 0.5c | 9.6 ± 0.5b |
| Fusarium oxysporum f. sp. hiveum | 12.1 ± 0.2a | 0 ± 0c | 4.3 ± 0.4b | 4.6 ± 0.3b |
| F. solani | 14.5 ± 0.2a | 3.2 ± 0.6c | 4.2 ± 0.4c | 7.8 ± 0.7b |
| Botrytis cinerea | 11.2 ± 0.3a | 1.3 ± 0.4c | 6.7 ± 0.3b | 8.7 ± 0.5b |
| Colletotrichum gloeosporioides | 18.3 ± 0.4a | 5.5 ± 0.3c | 7.8 ± 0.5b | 8.5 ± 0.2b |
| C. capsic | 12.7 ± 0.1a | 4.8 ± 0.2c | 8.3 ± 0.3b | 8.8± 0.3b |
| Sclerotinia sclerotiorum | 17.8 ± 0.3a | 6.7 ± 0.3c | 0 ± 0d | 11.8 ± 0.4b |
| Verticillium dahliae | 22.8 ± 0.2a | 15.7 ± 0.4b | 8.8 ± 0.3c | 15.8 ± 0.6b |
| Leptosphaeria biglobosa | 21.3 ± 0.4a | 16.5 ± 0.2b | 9.8 ± 0.1c | 17.8 ± 0.3b |
| Phomopsis vexans | 15.2 ± 0.2a | 6.2 ± 0.5c | 2.8 ± 0.2d | 9.7 ± 0.4b |
| Phomopsis asparagi | 16.8 ± 0.5a | 0 ± 0d | 8.2 ± 0.3b | 6.0 ± 0.5c |
1 Means ± S.D. followed with the same letters within each column were not significantly different (p > 0.05)
Table 2.
Cultural, physiological, and biochemical characteristics of isolate JKTJ-3.
| Characteristic | Growth1 | Substrate mycelia color | Aerial mycelia color2 | Sporulation3 | Pigment |
|---|---|---|---|---|---|
| Growth characteristics | |||||
| ISP-1 | ++ | Orange | - | NS | Orange |
| ISP-2 | +++ | Orange | GP/SP/DB | AS | Orange |
| ISP-3 | + | Orange | W | SS | Yellow |
| ISP-4 | ++ | Orange | W | SS | Yellow |
| ISP-5 | +++ | Orange | W/GP | AS | Orange |
| ISP-6 | ++ | Orange | - | NS | Orange |
| GS-1 | + | Yellow | - | NS | Yellow |
| BM | +++ | Orange | W/GP/DB | AS | Orange |
| Utilization of carbon sources | |||||
| D-Glucose | + | Not determined | Not determined | SS | Pale |
| Sucrose | ++ | Not determined | Not determined | SS | Pale |
| L-Arabinose | + | Not determined | Not determined | SS | Pale |
| D-Fructose | ++ | Not determined | Not determined | DS | Yellow |
| D-Xylose | +++ | Not determined | Not determined | SS | Yellow |
| D-Mannitol | +++ | Not determined | Not determined | SS | Yellow |
| Raffinose | + | Not determined | Not determined | SS | Pale |
| Rhamnose | ++ | Not determined | Not determined | SS | Pale |
| Growth response to NaCl | |||||
| NaCl (≤2%) | +++ | Not determined | Not determined | AS | Orange to yellow |
| NaCl (5%-8%) | + | Not determined | Not determined | NS | Yellow |
| NaCl (>8%) | - | Not determined | Not determined | NS | Pale |
| Growth response to pH | |||||
| pH 3.5 | + | Not determined | Not determined | NS | Pale |
| pH 4-7 | ++/+++ | Not determined | Not determined | SS | Yellow |
| pH 7.5-8.0 | + | Not determined | Not determined | NS | Yellow |
1 Growth: +, sparse mycelia; ++, dense mycelia; +++, highly-dense mycelia; -, no growth. 2 Aerial mycelia color: -, no aerial hyphae; W, whitish; GP, grayish pink; DB, dark brown; SP, smoky purple. 3 Sporulation: SS, sparse sporulation; DS, dense sporulation; AS, abundant sporulation; NS, no sporulation.
Table 3.
Protective efficacy of isolate JKTJ-3 cultural filtrate (CF) by treatment of seeding substrate.
Table 3.
Protective efficacy of isolate JKTJ-3 cultural filtrate (CF) by treatment of seeding substrate.
| Inoculation interval | Treatment | Seed emergence rate (%) | Damping-off incidence(%) | Seedling rate(%) | Control efficacy (%) |
|---|---|---|---|---|---|
| 0 d | JKTJ-3 CF | 82.0 b1 | 58.5 b1 | 34.0 b1 | 39.0 b1 |
| hymexazol WP | 90.0 a | 47.8 c | 47.0 a | 50.2 a | |
| Control | 75.0 c | 96.0 a | 3.0 c | - | |
| 1 d | JKTJ-3 CF | 90.0 a | 40.0 c | 54.0 a | 57.5 a |
| hymexazol WP | 90.0 a | 51.1 b | 44.0 b | 45.7 b | |
| Control | 86.0 b | 94.2 a | 5.0 c | - | |
| 2 d | JKTJ-3 CF | 95.0 a | 35.8 c | 61.0 a | 63.8 a |
| hymexazol WP | 95.0 a | 70.5 b | 28.0 b | 28.7 b | |
| Control | 90.0 b | 98.9 a | 1.0 c | - | |
| 3 d | JKTJ-3 CF | 95.0 a | 34.7 c | 62.0 a | 63.3 a |
| hymexazol WP | 95.0 a | 86.3 b | 13.0 b | 8.9 b | |
| Control | 95.0 a | 94.7 a | 5.0 c | - |
1 The values followed with the same letters within each column for each assay were not significantly different (p > 0.05).
Table 4.
Protective efficacy of isolate JKTJ-3 wheat grain culture (WGC) by treatment of seeding substrate.
Table 4.
Protective efficacy of isolate JKTJ-3 wheat grain culture (WGC) by treatment of seeding substrate.
| Inoculation interval |
Treatments | Seed emergence rate (%) | Damping-off incidence (%) | Seedling rate (%) | Control efficacy (%) |
|---|---|---|---|---|---|
| 0 d | JKTJ-3 WGC | 88.3 b1 | 21.7 c1 | 69.2 a1 | 75.6 a1 |
| hymexazol WP | 91.7 a | 40.0 b | 52.0 b | 55.3 b | |
| Control | 86.7 b | 89.3 a | 9.8 c | - | |
| 1 d | JKTJ-3 WGC | 87.5 a | 10.0 c | 78.8 a | 88.6 a |
| hymexazol WP | 89.2 a | 47.5 b | 43.3 b | 46.2 b | |
| Control | 82.5 b | 88.5 a | 10.3 c | - | |
| 2 d | JKTJ-3 WGC | 90.8 b | 0.8c | 90.1 a | 99.2 a |
| hymexazol WP | 93.3 a | 54.2b | 41.6 b | 38.3 b | |
| Control | 90.8 b | 87.8a | 11.4 c | - | |
| 3 d | JKTJ-3 WGC | 96.7 a | 0.0 c | 94.3 a | 100.0 a |
| hymexazol WP | 97.5 a | 59.8 b | 38.8 b | 30.5 b | |
| Control | 94.3 b | 86.3 a | 13.3 c |
1 The values followed with the same letters within each column for each inoculation interval treatmment were not significantly different (p > 0.05).
Table 5.
Bioconrol efficacy of isolate JKTJ-3 fermentation culture (FC) and cultural filtrate (CF) by treatment of seed-soaking.
Table 5.
Bioconrol efficacy of isolate JKTJ-3 fermentation culture (FC) and cultural filtrate (CF) by treatment of seed-soaking.
| Seed-soaking time | Treatment | Seed emergence rate (%) | Damping-off incidence (%) | Seedling rate (%) | Control efficacy (%) |
|---|---|---|---|---|---|
| 4 h | JKTJ-3 FC | 92.0 bc1 | 55.7 de1 | 44.3 c1 | 39.2 bc1 |
| JKTJ-3 CF | 91.0 c | 45.1 f | 54.9 a | 50.7 a | |
| Control | 95.0 b | 91.6 a | 8.4 g | ||
| 8 h | JKTJ-3 FC | 94.0 bc | 76.6 b | 23.4 ef | 14.7 e |
| JKTJ-3 CF | 95.0 b | 50.5 e | 49.5 b | 43.7 b | |
| Control | 93.0 bc | 89.8 a | 10.2 g | ||
| 12 h | JKTJ-3 FC | 95.0 b | 72.6 b | 27.4 e | 22.3 d |
| JKTJ-3 CF | 99.0 a | 63.5 c | 36.5 d | 32.1 c | |
| Control | 92.0 bc | 93.5 a | 6.5 g |
1 The values followed with the same letters within each column for each seed-soaking trial were not significantly different (p > 0.05) according to least significance test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



