
Submitted:
27 April 2023
Posted:
28 April 2023
You are already at the latest version
Abstract
Newcastle disease (ND) is a highly contagious and usually causes severe illness that affects Aves all over the world, including domestic poultry. Depending on the virus's virulence, it can impact the nervous, respiratory, and digestive systems and causes up to 100% mortality. The chIFITM genes are activated in response to viral infection. The current study was conducted to quantify the mRNA of chIFITM genes in vitro in response to ND viral infection. It also examined its ability to inhibit ND virus replication in Chicken Embryo Fibroblast (CEF) cells of Aseel and Kadaknath breeds. Results from the study showed that the expression of all chIFITM genes was significantly upregulated throughout the period in the infected CEF cells of both breeds compared to uninfected CEF cells. In CEF cells of the Kadaknath breed, elevated levels of expression of the chIFITM3 gene dramatically reduced the ND viral growth and the viral load was 60% lower than in CEF cells of the Aseel breed. The expression level of the chIFITMs in Kadaknath ranged from 2.39 to 11.68 log2 folds higher than that of control CEFs, and was consistently (p<0.01) higher than Aseel CEFs. Similar to this, IFN-γ gene expresses strongly quickly and peaks at 13.9 log2 fold at 48 hpi. The result suggests that the Kadaknath chicken breed may have a higher level of disease tolerance compared to the Aseel chicken.
Keywords:
ISG
; IFITM gene
; Mx
; Interferon
; CEF
; Aseel
; Kadaknath
; Newcastle disease
; viral load
1. Introduction
Newcastle disease (ND) has been globally distributed and the causative agent, Newcastle Disease virus (NDV) belongs to the Paramyxoviridae family, genus Avulavirus and is designated as Avian Paramyxovirus-1 (APMV-1). NDV is a non-segmented, negative sense, single-stranded enveloped RNA virus with approximately 15kb genome encodes namely, hemagglutinin–neuraminidase (HN), nucleoprotein (NP), fusion (F), phosphoprotein (P), matrix (M), RNA-dependent RNA polymerase (L) [1]. ND virus has a wide range of hosts and infection was reported in 250 Avian species in the world by either natural or experimental mechanisms [2]. Depending on the viral pathotype, the incidence of ND that affects poultry manifests as gastrointestinal, respiratory and neurological conditions that can result in up to 100% mortality [3,4]. ND has a high impact on the poultry industry through heavy economic loss aroused due to heavy mortality and production loss and also due to extensive attention to the prevention and treatment of this disease such as standard vaccination protocols and biosecurity measures [5]. It was calculated that from thirteen layer farms in the Gujarat state of India suffered a total economic loss of $4,588 (₹ 37,19,223) per year [5]. Various disease preventive strategies and vaccination are not effective due to the complex genetic diversity of viruses. So, it is necessary to develop a chicken population with disease resistant naturally.
During viral infection Type I interferon defense mechanism are triggered to express a set of genes against viral infections known as interferon-stimulated genes (ISGs) [6,7]. The IFITM (interferon-inducible transmembrane) gene is one of these ISGs and has been shown to prevent the propagation of several highly virulent viral pathogens, such as the coronavirus responsible for the severe acute respiratory syndrome (SARS), the filoviruses Marburg and Ebola, the influenza A viruses (IAVs), and flaviviruses (dengue virus) [8,9,10]. Scientists discovered that the chIFITM gene expression has a negative correlation with the emergence of influenza virus and its titre in in vitro study, indicating that chIFITMs1 ,2 and 3 had a functional role in the management of viral infections [11,12]. In chicken, IFITM genes were located in Chromosome 5 and found in two loci, one containing the various numbers of immune-related (IR)-IFITM (IFITM 1, 2, 3) genes and IFITM5 gene and IFITM10 gene in another loci [13,14]. Lanz et al. found that swIFITMs (swine IFITMs) had a dose-dependent restriction against IAV after infecting porcine HEK293-T cells with IAV A/WSN/33 (WSN) for 24 or 48 hours [15]. swIFITM2 and -3 were expressed at late endosomes and have most potent antiviral activity against IAV in porcine cells. Furthermore, no swIFITM5 expression was detected in any of the tested cell lines and concludes that the IFITM5 do not have significant role in immune function [15]. Further, the knockdown of IFITM3 in DF-1 cells by siRNA increased the infectivity of a vesicular stomatitis virus G protein-pseudo-typed lentiviral vector [16].
Hence this study was undertaken to examine the chIFITM gene expression pattern in two chicken breeds of India, Aseel and Kadaknath, whose levels of viral resistance are high. So, we selected the model of chicken embryo fibroblasts (CEFs) to observe chicken IFITM gene expression in vitro following infection with NDVs. We compared the expression of IFN γ and Mx gene in response to NDV infection by quantitative real-time polymerase chain reaction (qRT-PCR).
In addition to providing prospects for a deeper understanding of viral resistance, analysis of these genes in chickens offers potential strategies for preventing viruses in poultry farming. It may be possible to do a selective breeding program in poultry breeds for increased resistance against viral infections. So it needs to discover characteristics of resistance and understand how they operate. Further, The observation may have useful implications in terms of vaccine production. Many vaccines are produced in embryonated hen’s eggs or continuous avian cell lines. However, it is well established that the rate-determining step in the manufacture of numerous vaccines is the induction of antiviral immune responses that prevent the replication of vaccine viruses and the high cost involved in maintain and producing specific pathogen free (SFP) eggs from chicken.
2. Materials and Methods
2.1. Ethics Statement
The experiment was conducted with the approval of the Institutional Bio-safety Committee (IBSC) of TANUVAS-Veterinary College and Research Institute, Namakkal, Tamil Nadu, India (Approval Lr. No. 1764/VCRI-NKL/IBSC/2022 dated 11.05.2022 of the Dean, VCRI, Namakkal).
2.2. Chicken Embryo Fibroblast Cells
CEF cells were prepared from 9-10 days old SPF chicken embryos of Aseel and Kadaknath (Department of Poultry Science, Veterinary College and Research Institute, TNUVAS, Namakkal) as previously described method [17]. Fibroblastic cells were isolated from the respective embryo by removing the head and viscera, then cut into small pieces using sterile scissors and forces. The remaining tissues were washed with PBS and trypsinized for 5 minutes with 0.25% trypsin and magnetic stir. Allow pieces to settle, collect supernatant, centrifuge at 1000 rpm for 5 min, resuspend pellet in growth medium containing Dulbecco’s modified Eagle’s medium (DMEM) (Hi-media, Cat. No.AL007S) supplemented with 10 % fetal bovine serum (FBS) ((Hi-media, Cat. No. RM112-500ML), 1% antibiotics, Antimycotic solution (100x), stabilized (Sigma, Cat.No. A5955) 37 °C with 5 % CO2 for 24hrs.
2.3. Virus
The velogenic genotype XIII, NDV strain isolated from a field ND outbreak by Poultry Disease Diagnosis and Surveillance Laboratory, TANUVAS, Namakkal, India was used in this study. The ND viruse was inoculated into the allantoic cavity of 10-day-old SPF-embryonated White Leghorn chicken eggs and incubated at 37°C for 72 hrs as per the standard procedures of the Office International Des Epizooties [18]. After 72 hrs, inoculated eggs were chilled for 30 mins and then allantoic fluid was collected under sterile condition and stored at −80°C for further use. ND virus titers were quantified by Haemagglutination assay (HA) and confirmed by PCR amplification of ND viral F-gene using the primer to amplify 356 bp amplicon (Figure 1) using the primer pair of NDVF (5’- GCAGCTGCAGGGATTGTGGT – 3’) and NDVR (5’-TCTTTGAGCAGGAGGATGTTG – 3’) with the cycle condition of initial denaturation 95°C for 3 min followed by 35 cycle of 94°C for 45s, 52°C for 45s, 72°C for 45s and final extension 72°C for 5min [19].
2.4. Tissue Culture Infection Doses (TCID50)
The viral infective dose was measured by TCID50. CEF cells of SPF-embryonated White leghorn chicken eggs were cultured in 96-well plates and were incubated with the cell supernatants of different groups, which had 10-fold serially diluted viral suspension. Each dilution had five replicates. One h after NDV infection, the supernatants were replaced with DMEM containing 2% fetal bovine serum (FBS). Then incubated at 37 °C and was observed daily for CPE scoring and continued scoring daily till the control wells started dying. The TCID50 value was determined using Spearman-Karber’s method [20].
2.5. Viral Infection
CEFs prepared from Aseel and Kadaknath SPF chicken embryos were seeded 24 h prior to infection in a 25 cm2 Tissue culture Flask (T25) (Hi-media, India) at a cell density of approximately 7 × 105 cells/flask. The CEF cells in triplicate were infected with velogenic genotype XIII, NDV strains to circulate in Tamil Nadu, South India [21]. The ND viral suspension was diluted to 50 % tissue culture infective dose (TCID50) of 106/ml and 0.5ml of viral suspension (TCID50) was added into the flask and allowed for viral adsorption by incubating at 37oC in a humidified atmosphere containing 5 % CO2 for 1 hr. Afterwards, the growth medium was replaced with DMEM supplemented with 2 % FBS. Uninfected cells were regarded as control samples. CEF cells were harvested from uninfected control and infected CEF cells at 3, 6, 12, 24 and 48 hours post-infected (hpi) and stored at −80°C for RNA extraction. Virus load in CEF cells were quantified by an absolute quantification method [22] and gene expression by relative quantification 2−ΔΔCt method [23].
2.6. RNA Extraction and cDNA Synthesis
Total RNA was extracted from the infected and uninfected control group CEF cells of both breeds at each time point by Trizol method using RNAiso Plus, Takara, (Cat. # 9109)) (Total RNA extraction reagent). RNA in each sample was quantified by using Thermo Scientific's Nanodrop spectrophotometer (Thermo Scientific, USA). Approximately, 1 µg RNA from each sample was used for complementary DNA (cDNA) synthesis by using iScript cDNA Synthes kit (Bio-Rad, Cat # 1708891) according to the manufacturer’s protocol which follows a method of Reverse Transcription (RT) random primer.
2.7. Primer Pairs Design
RT-qPCR primers for chIFITMs genes were designed using Primer-BlAST with a length of 20 to 23 bases and amplicon sizes ranging from 115 to 196 bp. The sequences of these genes were obtained from NCBI (https://www.ncbi.nlm.nih.gov). The primer specificity of each gene was verified using 2.5% agarose gel electrophoresis and melting curve analysis. To validate the specificity of each primer pair, it was verified by in-silico PCR with the NCBI sequence database using NCBI PRIMER BLAST.
2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
A total of 4 targeted genes (chIFITM1, 2, 3 and 5) expression pattern in ND virus infected and uninfected control CEF cells of both breeds were studied alone with 2 Positive immune-related genes (IFN γ and Mx ) and 1 housekeeping gene (β Actin) (Table 1). The relative expression of specific gene mRNA was quantified and the absolute quantification of viral load was done by a real-time thermal cycler (IIIumina Real-time machine, USA). All reactions were performed in a nuclease-free 48-well qRT-PCR Illumina plate with sealer. The qRT-PCR response was done with the final volume of 20 µl using 10 µl of SYBR Green PCR Master Mix Kit (Bio-rad), 10 pmol of each forward and reverse primer, and 1µl of cDNA. The cycle condition of qRT-PCR was initial denaturation at 95oC for 10 mins, followed by 40 cycles of 95 °C for 10s, and 60 °C (β Actin, IFN γ chIFITM1 and Mx genes) to 62 °C (chIFITM2, 3 and 5) for 45s and 72 °C for 15s. Further, the target and β-actin genes' respective cycle threshold (Ct) values were computed. Using the 2−ΔΔCt approach, the relative fold change of the target genes in the infected groups was calculated using the delta Ct of the uninfected control group [23].
2.9. Standard Curve Analysis for Detection of Viral Load
Standard curve was constructed by linear regression method to do absolute quantification of viral load in the samples [22]. Ten-fold serially diluted T-NP plasmid (20 ng/µl, A260/280 ratio = 1.80) with known concentration was used as a standard to construct a standard curve and to obtain a linear regression equation. A standard curve was then generated by plotting 𝐶𝑞 values against logarithmic of plasmid copies numbers in the standard. A correlation between NDV-specific nucleoprotein (NP) gene copy numbers and Cq values as found by using Pla-rt13 and Pla-rt14 primer pairs specific for nucleoprotein (NP) region of NDV genome (Table 1). The qPCR cycle condition was initially 95°C for 5 mins, followed by 40 cycles of 95°C for 15s, 60°C for 30s and finally 72°C for 30s. Melting curve analysis was used to know the specificity of qPCR primers. A partial regression equation was obtained as 𝑌 = −2.25x + 14.09 (Figure 1). R2 value ranging from 0.994 to 0.932 and it indicates strong and linear relationships between the Cq value and number of gene copies in the sample.
2.10. Statistical Analysis
The t-test and one-way ANOVA was conducted to analyze real time PCR data. The mean (n = 3 per time point/breed for each infected and control) and standard error of the mean are in log2(2-ΔΔCt) used to express data and log10 (viral copies) used to express viral load. R software version 4.2.1 was used to analyze data and a p-value < 0.05 was used to determine statistical significance. Further, the R program was also used to create the graphical illustrations of the results.
3. Results
3.1. NDV Infection-Induced Cytopathic Changes and Viral Load
In the current study, we have determined the IFITM gene expression against the live NDV (velogenic genotype XIII, NDV strains) in the CEF cells. Figure 2 highlights the normal morphology of the CEF cells derived from the 9–10-day-old SPF embryonated chicken eggs. The effect of NDV infection in the chicken embryo fibroblast cells was examined under an inverted microscope for their morphology to determine the cytopathic effect (CPE) at 3, 6, 12, 24 and 48 hpi and compared with uninfected control cells. The cell monolayer was intact and morphologically similar to the control group till 3 and 6 hpi in Aseel and Kadaknath respectively. However, at 6 and 12 hpi, morphological changes was observed which are typical of cytopathic effect (CPE) viz., cell rounding, a fusion of infected cells to form syncytia cells and detachment of cells from monolayer followed by cell death were noticed under the light microscope in both breeds (Figure 2).
Further, NDV infected CEF cells also showed morphological alterations typical of apoptosis, rounding of a cell and cytoplasm vacuolation was noticed in both breeds. But cell rounding starts from 6h and progressed and at 48h, complete cell detachment was observed in Aseel CEF cells. However, it was delayed in the case of Kadaknath CEFs, cell rounding starts from 12 h and progressed, and at 48h cell detachment progressed. This revealed that CPE advancement related to virus load and time of infected cells, in addition to confirming virus infection and its replication in infected cells.
Viral load was calculated by standard curve analysis using the T-NP plasmid as a standard and we observed a steady increase in viral load till 12h and 24h respectively in Aseel and Kadaknath CEF cells (Figure 3). In the case of Aseel CEF, viral load was significantly (p<0.01) higher than Kadaknath at 3 hpi (2.01 log10), 6 hpi (2.75 log10) and at 12 hpi (5.38 log10). In contrast, in Kadaknath CEF, the viral load was significantly (p<0.01) lower when compared to Aseel at each point 3 hpi (1.01 log10), 6 hpi (1.59 log10) and 12 hpi (2.15 log10). But there is no significant difference found at 24 hpi ( 4.54 and 4.63 log10) and 48 hpi (2.98 and 2.95 log10) between Aseel and Kadaknath CEF respectively. In Aseel, the viral load peaked at 12 hours and the largest load was observed at approximately 5.3 log10 viral copies in Aseel, while it was significantly (p<0.001) low as 2.15 log10 viral copies as in Kadaknath CEF cells at 12 hpi. Whereas, in Kadaknath highest load was recorded at 24 hours and found as 4.63 log10 viral copies (Figure 3d). In the later hours, the viral production gradually decreased. However, the viral load in the Aseel CEF was still higher than in Kadaknath CEF cells at any time, confirming that Kadaknath cells potentially restrict the multiplication of ND viruses compared to Aseel CEF cells.
3.2. Expression Analysis of chIFITM Gene in Newcastle Disease Virus Infected CEF Cells
The relative expression of chIFITM genes in the control (uninfected) and infected cells were quantified by qRT-PCR. Our results showed that the expression of the chIFITM 1, 2, 3, 5, IFN-γ and Mx genes are in a time-dependent fashion (Table 2 and Figure 3a,b). After ND viral infection of the CEF cells, the mRNA levels of all four selected genes (chIFITM 1, 2, 3, 5) and positive immune-related genes (IFN-γ and Mx ) were gradually increased reaching a peak at different hours post-infection (hpi) in both breeds. In comparison to the calibrator (uninfected-control CEF cells), the relative expression of IFN- γ reaches peck at 48 hpi as 2.59 log2 fold and 13.9 log2 fold higher in Aseel and Kadaknath, respectively. The results show that the chIFITMs are expressed at basal levels in CEF cells of both breeds. It also demonstrates that the breeds that were studied exhibit various patterns of expression. Compared to Kadaknath CEF, expression of the chIFITMs gene is lower and more variable in Aseel.
In Aseel, only at 12 hpi expression of the chIFITM2 gene was dramatically increased, whereas chIFITM5 upregulation begins at 6 hpi and continues until 24 hpi. In addition, the highest expression in chIFITM2 and 5 genes were recorded as 3.25 log2 fold and 3.82 log2 fold respectively at 6 hpi and 24 hpi. The chIFITM3 gene was strongly expressed from 3 to 12 hpi, with the maximum level being 3.46 log2 fold at 6 hpi. Like chIFITM3, chIFITM1 upregulation starts at 3 hpi, but it continues strongly until 48 hpi and reaches a maximum of 6 log2 fold at 6 hpi. Mx and chIFITM3 exhibit comparable patterns of expression, Mx gene reaching its highest level of expression at 3 hpi as a 4.18 log2 fold increase.
As opposed to Aseel, Kadaknath CEF cells express all chIFITMs genes strongly and severalfold significantly higher from 3 to 48 hours after ND virus infection. At 6 hpi, the highest expression of chIFITM2 was detected. chIFITM1, 5 and Mx were at 24 hpi, but chIFITM2 and IFN-γ at 48 hpi. In Kadaknath, the expression level among the chIFITMs ranged from 2.39 to 11.68 log2 folds more than that of control CEFs, which was over the entire time span, significantly (p<0.01) higher than Aseel CEFs (Figure 3c). Similarly, IFN-γ expresses strongly from the beginning and reaches maximum as 13.9 log2 fold at 48 hpi. The expression levels of chIFITM1 and 3 were found to be higher than the other chIFITMs in Aseel and Kadaknath respectively.
4. Discussion
To measure the level of chIFITM gene expression against NDV in the current work, we employed the velogenic genotype XIII of NDV strain that was used to infect CEF cells. The ND viral load increases, prompting the chicken embryo fibroblast (CEFs) cells to express more chIFN-γ significantly [25]. In Aseel's CEF cells, there was noticeable upregulation starting at 6 hpi and continuing until 48 hpi. In contrast, in Kadaknath CEF cells, chIFN-γ expression began to increase significantly at 3 hpi and peaked (13.9±0.49 log2 fold) like Aseel CEF at 48 hpi. Further, it was significantly (p<0.001) several folds higher than Aseel. However, interferons (IFNs), a vital component of innate immune signaling, serve as the first line of defense against invading viruses [26]. Therefore, it consistently and strongly expressed against the ND virus [27]. chIFITMs and Mx are members of the interferon-stimulating gene (ISG) group (Mycovirus resistant gene) [28,29]. Chicken has five members of the IFITM family: chIFITM1, chIFITM2, chIFITM3, chIFITM5, and chIFITM10. These protein genes are activated and made to express themselves by type I and type II IFNs, signifying the start of the innate host response [13,30]. Recent studies confirm that STAT/IRF signaling pathways activate IFITM gene expression together with other ISGs during infection and inflammation [31]. This study examines the relative mRNA expression profile of chicken IFITMs after Newcastle disease virus (NDV) infection in vitro with a focus on how the cells react during the early stages of NDV infection.
We observed significant upregulation of chIFITMs, chIFN-γ and Mx in the CEF cells of both breeds. A significant viral load indicated the presence of a replicating virus in the CEF cells. chIFITM1, 2, 3, and 5 are noticeably and gradually upregulated in both breeds of CEFs after NDV infection. In Kadaknath CEF cells, chIFITMs and IFN-γ expressions were relatively high with statistical significance (p<0.001) from 3 to 48 hours post-infection compared to control uninfected. Researchers also found that high quantities of chIFITM1, 2, and 3 are expressed in CEFs from 4 to 24 hours after H9N2 infection.[32]. While CEF cells from Aseel took longer to exhibit strong mRNA expression of chIFN-γ and chIFITM genes strongly expressed at 6 hours post-infection. In Kadaknath CEF, among the IR-IFITM family members, chIFITM3 (11.68 log2 folds) has the greatest expression observed at 6 hours post-infection, followed by chIFITM2 (8.89 log2 folds) at 48 hours post-infection, chIFITM1 (7.79 log2 folds) and chIFITM5 (7.04 log2 folds) at 24 hours post-infection [13,15]. But in Aseel, IFITM1 (6.00 log2 folds) came first, then chIFITM3 (3.46 log2 folds), and then chIFITM2 (3.25 log2 folds) at 6hpi, lastly chIFITM5 (3.82 log2 folds) at 24hpi. In our finding that Kadaknath expressed large levels of IFN-γ and stimulated high levels of chIFITMs compared to Aseel is supported by studies from other scientists that IFN treated CEFs expressed high levels of chIFITMs [13,30,32]. It is known that chIFITM 1, 2, and 3 prevent the replication of a variety of RNA viruses that enter the host cell through endocytosis [8]. Infected CEF of Kadaknath showed a significant (p<0.01) and robust overexpression of all chIFITMs started from 3hpi when compared to the control. Whereas similarly in Aseel chIFITM genes upregulation starts from at 3 hours post-infection, compared to the control. A Significant and high level Mx gene response has been observed at 3 hours post-infection (4.18 log2 folds) and then decreased level of expression similar to H9N2 infected CEFs [32] and it was delayed in Kadaknath at 24 hours post-infection (3.57 log2 folds) [31]. Mx gene was used as positive control gene because it is a one of the well-known IFN-stimulating and it highly expressed as restriction factor of influenza A viral infection [33].
Results from other publications' findings support in a similar way that the expression of chIFITM-2, 3, and Mx significantly increased after H3N8 infection, and this increase started at 6 hours after infection. Although there was a reduction in Mx expression at 12 hours after infection and both chIFITM2 and 3 were significantly elevated. At 6 hours after infection, chIFITM1 2, 3, and 5 and Mx expression significantly increased and persisted for 24 hours in H5N3-infected CEF cells [32]. Like this, IFITMs were constantly and significantly upregulated in Kadaknath CEF cells throughout the study, which may be related to the increased level of IFN-γ gene expression. In contrast, Aseel CEFs expressed low levels of IFN-γ and chIFITM genes investigated. This is supported by the findings of other papers. Thus, it was hypothesized that interferons would activate and upregulate the expression of chIFITM in CEFs based on evidence from researchers Whitehead and Smith et al. [14,32]. It was further demonstrated that it is possible to inhibit virus replication by simply preventing access into a cell as evidenced by the production of the IFITMs following IFN treatment [26,30].
We measured the log10 viral copies of NDV at 3-, 6-, 12-, 24- and 48-hours post-infection in CEF cells and contrasted both breeds. The viral load steadily increased from 3 hours post-infection itself in both breeds and the viral load was significantly (p<0.01) lower in Kadaknath when compared to Aseel from 3- to 12- hours post-infection and also overall load through the infected period. The outcomes showed that the ND viral load at 12 hours post-infection in Kadaknath CEF (2.15 log10) cells was reduced by 60% in comparison to Aseel CEF (5.38 log10) cells. Inversely proportional to viral load, Kadaknath showed significantly (p<0.01) high expression of all the chIFITM genes at all the periods than Aseel. Like this, Blyth et al. found that overexpressing chIFITM3 reduces influenza H6N2 and H1N9 strain infection in DF-1 cells by 30 to 40% [34]. Similarly to this, in vitro overexpression of chIFITM3 limits the multiplication of the influenza virus by 55% [14]. Infectious Bursal viral (IBV) strains of QX, M41-CK and Beaudette infection significantly upregulates all IR-chIFITM genes at 24 hpi [13]. Scientist concluded that chIFITM2 and 3 greatly decrease the lyssavirus infection [13,14].
However, by altering the characteristics of cellular membranes and blocking the cell surface receptors to restrict viral entry, the Interferon-inducible transmembrane proteins (IFITMs) prevent many harmful viruses from infecting cells and causing infection [35,36,37]. This ultimately prevents viral fusion [36]. Several reports confirm that the IFITMs effectively control the RNA viruses such as Avian influenza A virus (IAV), Lyssaviruses [14], Infectious Bronchitis virus (IBV) [13] and Avian Reovirus multiplication (ARV) [30], which follows the endosomal pathway to enter the host cell membrane for multiplication.
Microscopic analysis of the infected cells showed that the monolayer remained intact, much like in mock-infected cells, and that no CPE occurred until 3 h post-infection (Aseel) to 6 h post-infection (Kadaknath). However, after 6 to 12 hpi, the light-microscopy analysis revealed morphological changes indicative of CPE, including rounding, the fusing of infected cells to form syncytia, and the detachment of cells from the monolayer followed by cell death. Cellular rounding, membrane blebbing, cytoplasm vacuolation, nuclear condensation, and nuclear envelope collapse are among the morphological changes brought on by NDV infection. This result is congruent with what has been documented in other publications [38]. The CPE in the current investigation, however, showed that CEF cells of both breeds began exhibiting CPE sooner than had been previously reported. According to Li et al., overexpression of IFITM3 inhibited the inflammatory response of PF15 cells and is crucial to the TLR4-NF-B signalling pathway, which is implicated in the inflammatory response [39]. In Kadaknath, chIFITM3 expression levels are consistently high (p<0.01), with a log2 fold range of 10.08 to 11.68 in a contrast to 1.00 to 3.46 in Aseel. As a result, compared to Aseel, the Kadaknath CEF cells had a delayed cytopathic effect and cell death. Elevated IFITM gene expression inhibits the spread of infections by restricting host cell proliferation. It is also involved in inhibiting cell adhesion and controlling cell growth [40]. Additionally, Anjum et al. noticed a decrease in cytopathic effects in chIFN-treated CEF cells when they were infected with ND and AIV [41]. Similarly, in Kadaknath CEF cells expressed high level of IFN-γ and delayed cytopathic changes was observed.
5. Conclusions
In this study: we have shown the variability in the magnitude of chIFITMs mRNA expression between breeds during the course of Newcastle disease Viral-infection. Such variation suggests that the chIFITM response may be breed-dependent and intragenic factor-dependent. Our data suggest CEF cells start expressing all chIFITM genes significantly in the early stage of infection, regardless of the breed of the chicken. Elevated levels of expression of chIFITMs in Kadaknath CEF cells restrict the viral multiplication compared to the Aseel CEFs. Together the result shows chIFITMs plays a critical role in restricting the ND virus multiplication. In addition, it has been shown that the basal level of IFN-γ expression will impact the chIFITM gene expression. Therefore, we have revealed that viral entry is restricted depending on the level of chIFITMs expression and the expression depends on other factors. This study was conducted in vitro and more experiments are necessary to clarify the underlying mechanism in controlling the viral diseases in chicken. In future, In-Ovo and In vivo studies will be required to the better understand the role of this gene in immune system.
Author Contributions
Conceptualization, M.M., A.K.T. and A.R; methodology, M.M., and A.K.T.; software, M.M., and V.B.R.; validation, N.M., K.S., and M.S.; formal analysis, A.K.T.; investigation, M.M.; resources, V.G.; data curation, M.M. and A.K.T.; writing—original draft preparation, M.M., and A.K.T.; writing—review and editing, N.M., M.S., A.R., S.O.P and K.S.; supervision, A.K.T., N.M., M.S., and K.S; project administration, N.M., K.S., and M.S.; funding acquisition, N.M., and M.S.: All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by TANUVAS, India. (USO. No. 20652/A1/2020 and No.273/A1/2021, dated 26.02.2021 of Registrar, TANUVAS, Chennai – 51, India.)
Institutional Review Board Statement
The experiment was conducted with the approval of the Institutional Bio-safety Committee (IBSC) of TANUVAS-Veterinary College and Research Institute, Namakkal, Tamil Nadu, India (Approval Lr. No. 1764/VCRI-NKL/IBSC/2022 dated 11.05.2022 of the Dean, VCRI, Namakkal).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data set created and analysed in the current study will be made available on reasonable request.
Acknowledgments
The authors gratefully acknowledge the faculties of the Poultry Disease Diagnosis and Surveillance Laboratory (PDDSL), TANUVAS, Namakkal, India, for their support and for laboratory facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chambers, P.; Millar, N.S.; Bingham, R.W.; Emmerson, P.T. Molecular Cloning of Complementary DNA to Newcastle Disease Virus, and Nucleotide Sequence Analysis of the Junction between the Genes Encoding the Haemagglutinin-Neuraminidase and the Large Protein. J. Gen. Virol. 1986, 67 Pt 3, 475–486. [Google Scholar] [CrossRef]
- O.I.E. Newcastle Disease. Chapter 2.3.14; In, Ed.; World Organisation for Animal Health: Paris, 2012. [Google Scholar]
- Qosimah, D.; Murwani, S.; Sudjarwo, E.; Lesmana, M.A. Effect of Newcastle Disease Virus Level of Infection on Embryonic Length, Embryonic Death, and Protein Profile Changes. Vet. World 2018, 11, 1316–1320. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.A.; Parks, G.D. Paramyxoviridae: The Viruses and Their Replication. In Fields virology; Fields, B.N., Knipe, D.N., Howley, P.M., Eds.; Lippincott, Williams, and Wilkins, 2007; pp. 1449–1496.
- Kalaria, V.A.; Prajapati, K.S.; Javia, B.B.; Bhadaniya, A.R.; Fefar, D.T.; Vagh, A.A.; Trangadiya, B.J.; Padodara, R.J.; Mokaria, K.N.; Kumbhani, T.R. An Economical Impact of Newcastle Disease Outbreaks in Various Commercial Broiler Chicken Farms During 2020- 21 in Gujarat, India. Int J Curr Microbiol App Sci 2021, 10, 411–420. [Google Scholar]
- Aristizábal, B.; González, Á. Innate Immune System; Anaya, J.M., Shoenfeld, Y., Rojas-Villarraga, A., Eds.; Autoimmunity: Bogota (Colombia, 2013. [Google Scholar]
- Dai, M.; Xie, T.; Liao, M. Systematic Identification of Chicken Type I, II and III Interferon-Stimulated Genes. Vet Res 2020, 51, 70. [Google Scholar] [CrossRef] [PubMed]
- Brass, A.L.; Huang, I.-C.; Benita, Y.; John, S.P.; Krishnan, M.N.; Feeley, E.M.; Ryan, B.J.; Weyer, J.L.; van der Weyden, L.; Fikrig, E.; et al. The IFITM Proteins Mediate Cellular Resistance to Influenza A H1N1 Virus, West Nile Virus, and Dengue Virus. Cell 2009, 139, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Weidner, J.M.; Qing, M.; Pan, X.-B.; Guo, H.; Xu, C.; Zhang, X.; Birk, A.; Chang, J.; Shi, P.-Y.; et al. Identification of Five Interferon-Induced Cellular Proteins That Inhibit West Nile Virus and Dengue Virus Infections. J. Virol. 2010, 84, 8332–8341. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.-C.; Bailey, C.C.; Weyer, J.L.; Radoshitzky, S.R.; Becker, M.M.; Chiang, J.J.; Brass, A.L.; Ahmed, A.A.; Chi, X.; Dong, L.; et al. Distinct Patterns of IFITM-Mediated Restriction of Filoviruses, SARS Coronavirus, and Influenza A Virus. PLOS Pathog. 2011, 7, e1001258. [Google Scholar] [CrossRef]
- Feeley, E.M.; Sims, J.S.; John, S.P.; Chin, C.R.; Pertel, T.; Chen, L.-M.; Gaiha, G.D.; Ryan, B.J.; Donis, R.O.; Elledge, S.J.; et al. IFITM3 Inhibits Influenza A Virus Infection by Preventing Cytosolic Entry. PLoS Pathog. 2011, 7, e1002337. [Google Scholar] [CrossRef]
- Fife, M.; Moore, J. IFITM Knockdown/Knockout Technology for Vaccine Production.
- Steyn, A.; Keep, S.; Bickerton, E.; Fife, M. The Characterization of ChIFITMs in Avian Coronavirus Infection In Vivo, Ex Vivo and In Vitro. Charact. ChIFITMs Avian Coronavirus Infect. Vivo Ex Vivo Vitro Genes 2020, 11, 918. [Google Scholar] [CrossRef]
- Smith, S.E.; Gibson, M.S.; Wash, R.S.; Ferrara, F.; Wright, E.; Temperton, N.; Kellam, P.; Fife, M. Chicken Interferon-Inducible Transmembrane Protein 3 Restricts Influenza Viruses and Lyssaviruses In Vitro. J. Virol. 2013, 87, 12957–12966. [Google Scholar] [CrossRef]
- Lanz, C.; Yángüez, E.; Andenmatten, D.; Stertz, S. Correction for Lanz et al., Swine Interferon-Inducible Transmembrane Proteins Potently Inhibit Influenza A Virus Replication. J. Virol. 2015, 89, 2988. [Google Scholar] [CrossRef] [PubMed]
- Kidani, S.; Okuzaki, Y.; Kaneoka, H.; Asai, S.; Murakami, S.; Murase, Y.; Iijima, S.; Nishijima, K. Expression of Interferon-Inducible Transmembrane Proteins in the Chicken and Possible Role in Prevention of Viral Infections. Cytotechnology 2017, 69, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Hitchner, B.; Donermuch, H.; Graham Purchas, H.; William, E. Isolation and Identification of Pathogens, 2nd ed.American Association of Avian Pathologists, Inc, 1980. [Google Scholar]
- Office International des Epizooties (OIE) OIE Terrestrial Manual 2021 2021.
- Toyoda, T.; Sakaguchi, T.; Hirota, H.; Gotoh, B.; Kuma, K.; Miyataj, T.; Nagai, Y. Newcastle Disease Virus Evolution: II. Lack of Gene Recombination in Generating Virulent and Avirulent Strains. Virology 1989, 169, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Ogbole, O.O.; Akinleye, T.E.; Segun, P.A.; Faleye, T.C.; Adeniji, A.J. In Vitro Antiviral Activity of Twenty-Seven Medicinal Plant Extracts from Southwest Nigeria against Three Serotypes of Echoviruses. Virol. J. 2018, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Gowthaman, V.; Ganesan, V.; Gopala Krishna Murthy, T.R.; Nair, S.; Yegavinti, N.; Saraswathy, P.V.; Suresh Kumar, G.; Udhayavel, S.; Senthilvel, K.; Subbiah, M. Molecular Phylogenetics of Newcastle Disease Viruses Isolated from Vaccinated Flocks during Outbreaks in Southern India Reveals Circulation of a Novel Sub-Genotype. Transbound. Emerg. Dis. 2019, 66, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Yu, Y.; Yu, S.; Zhan, Y.; Wei, N.; Song, C.; Sun, Y.; Tan, L.; Ding, C. Development of Strand-Specific Real-Time RT-PCR to Distinguish Viral RNAs during Newcastle Disease Virus Infection. Sci. World J. 2014, 934851. [Google Scholar] [CrossRef] [PubMed]
- Pfaf, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res 2001, 29, 45. [Google Scholar] [CrossRef]
- Ramakrishnan, S.; Annamalai, A.; Sachan, S.; Kumar, A.; Sharma, B.K.; Govindaraj, E.; Chellappa, M.M.; Dey, S.; Krishnaswamy, N. Synergy of Lipopolysaccharide and Resiquimod on Type I Interferon, pro-Inflammatory Cytokine, Th1 and Th2 Response in Chicken Peripheral Blood Mononuclear Cells. Mol. Immunol. 2015, 64, 177–182. [Google Scholar] [CrossRef]
- Susta, L.; Cornax, I.; Diel, D.G.; Garcia, S.C.; Miller, P.J.; Liu, X.; Hu, S.; Brown, C.C.; Afonso, C.L. Expression of Interferon Gamma by a Highly Virulent Strain of Newcastle Disease Virus Decreases Its Pathogenicity in Chickens. Microb Pathog 2013, 61–62, 73–83. [Google Scholar] [CrossRef]
- Yang, X.; Arslan, M.; Liu, X.; Song, H.; Du, M.; Li, Y.; Zhang, Z. IFN-γ Establishes Interferon-Stimulated Gene-Mediated Antiviral State against Newcastle Disease Virus in Chicken Fibroblasts. Acta Biochim Biophys Sin Shanghai 2020, 52, 268–280. [Google Scholar] [CrossRef]
- Bashir, K.; Kappala, D.; Singh, Y. Combination of TLR2 and TLR3 Agonists Derepress Infectious Bursal Disease Virus Vaccine-Induced Immunosuppression in the Chicken. Sci Rep 2019, 9, 8197. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Qu, L.J.; Hou, Z.C.; Yao, J.F.; Xu, G.Y.; Yang, N. Genomic Structure and Diversity of the Chicken Mx Gene. Poult. Sci. 2007, 86, 786–789. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, J.; Winkler, C.A.; P, A.; Guo, J.-T. IFITM Genes, Variants, and Their Roles in the Control and Pathogenesis of Viral Infections. Front Microbiol 2019, 9, 3228. [Google Scholar] [CrossRef] [PubMed]
- Q., W.; X., Y.; C., L.; H., D.; C, W. Distribution of IFITM3 in Yellow-Feathered Broilers and Inhibition of Avian Reovirus Multiplication by IFITM3. Braz. J. Poult. Sci. 2018, 20, 377–386. [CrossRef]
- Friedlová, N.; Zavadil Kokáš, F.; Hupp, T.R.; Vojtěšek, B.; Nekulová, M. IFITM Protein Regulation and Functions: Far beyond the Fight against Viruses. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, T.J. Characterisation of Chicken Interferon-Inducible Transmembrane Proteins: Locus Architecture, Gene Expression and Viral Restriction. Doctoral thesis (Ph. D), 2018.
- Haller, O.; Kochs, G. Mx Genes: Host Determinants Controlling Influenza Virus Infection and Trans-Species Transmission. Hum. Genet. 2020, 139, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Blyth, G.A.D.; Chan, W.F.; Webster, R.G.; Magor, K.E. Duck Interferon-Inducible Transmembrane Protein 3 Mediates Restriction of Influenza Viruses. J. Virol. 2016, 90, 103–116. [Google Scholar] [CrossRef]
- Perreira, J.M.; Chin, C.R.; Feeley, E.M.; Brass, A.L. IFITMs Restrict the Replication of Multiple Pathogenic Viruses. J. Mol. Biol. 2013, 425, 4937–4955. [Google Scholar] [CrossRef]
- Desai, T.M.; Marin, M.; Chin, C.R.; Savidis, G.; Brass, A.L.; Melikyan, G.B. IFITM3 Restricts Influenza A Virus Entry by Blocking the Formation of Fusion Pores Following Virus-Endosome Hemifusion. PLoS Pathog 2014, 10, 1004048. [Google Scholar] [CrossRef]
- Amini, B.O.S.; Choi, Y.J.; Lee, J.H.; Shi, M.; Huang, I.C.; Farzan, M.; Jung, J.U. The Antiviral Effector IFITM3 Disrupts Intracellular Cholesterol Homeostasis to Block Viral Entry. 2013, 452–464.
- Ravindra, P.V.; Tiwari, A.K.; Ratta, B.; Chaturvedi, U.; Palia, S.K.; Chauhan, R.S. Newcastle Disease Virus-Induced Cytopathic Effect in Infected Cells Is Caused by Apoptosis. Virus Res 2009, 141, 13–20. [Google Scholar] [CrossRef]
- Li, X.; Su, S.; Cui, N.; Zhou, H.; Liu, X.; Cui, Z. Transcriptome Analysis of Chicken Embryo Fibroblast Cell Infected with Marek’s Disease Virus of GX0101∆LTR. Braz. J. Poult. Sci. 2017, 19, 179–184. [Google Scholar] [CrossRef]
- Siegrist, F.; Ebeling, M.; Certa, U. The Small Interferon-Induced Transmembrane Genes and Proteins. J Interferon Cytokine Res 2011, 31, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Anjum, F.R.; Rahman, S.U.; Aslam, M.A.; S, Q.A. Antiviral Potential and Stability Analysis of Chicken Interferon-α Produced by Newcastle Disease Virus in Chicken Embryo Fibroblast Cells. Vet Med-Czech 2021, 66, 197–207. [Google Scholar] [CrossRef]
Figure 1.
Newcastle disease virus detection and quantification. (a) PCR amplification of a 356 bp long F-gene amplicon specific to the ND virus. (b, c, d) Real-time quantitative PCR assay creation for absolute quantification. (b) Amplification plot of serially diluted T-NP plasmid standard. (c) Melting curve, (d) Standard curve for ND viral load detection with linear regression equation 𝑌 = −2.25x + 14.09 and R2 score value 0.994.
Figure 1.
Newcastle disease virus detection and quantification. (a) PCR amplification of a 356 bp long F-gene amplicon specific to the ND virus. (b, c, d) Real-time quantitative PCR assay creation for absolute quantification. (b) Amplification plot of serially diluted T-NP plasmid standard. (c) Melting curve, (d) Standard curve for ND viral load detection with linear regression equation 𝑌 = −2.25x + 14.09 and R2 score value 0.994.
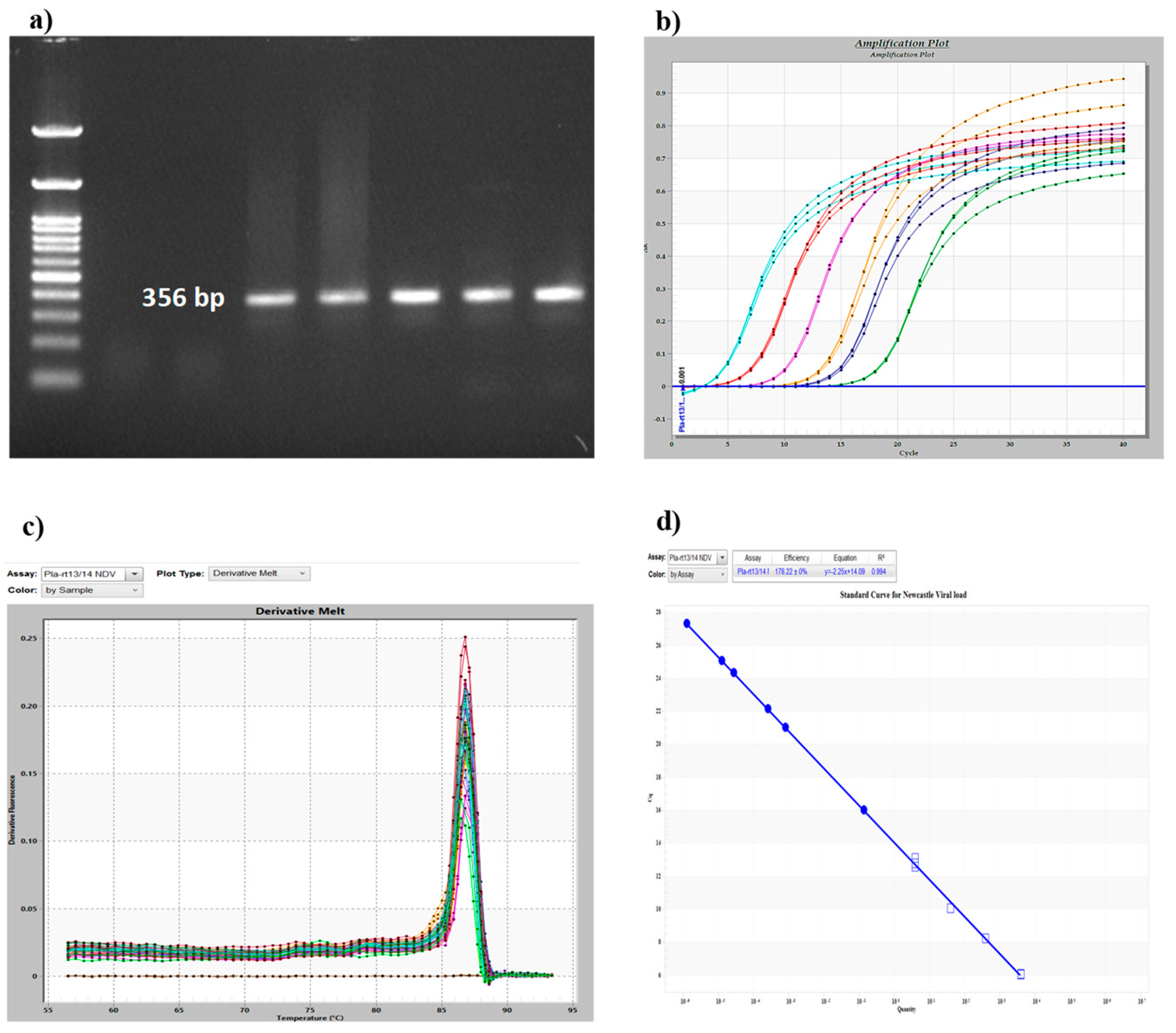
Figure 2.
Observations of Chicken embryo fibroblast cell cultures under an inverted microscope exhibiting at 20X (a, b), and 40X (c, d), phase contrast objectives. (a) and (b). Uninfected and Newcastle disease virus infected CEF cell at different hours post infection (hpi). (a) Kadaknath CEF showing delayed cytopathic changes, cell rounding noticed at 12hrs and (b) Aseel CEF started showing cytopathic changes such us cell rounding noticed at 6hrs. (c) Normal CEF cells and (d) Cytopathic effect of ND virus in CEF showing multinucleated cells (Syncytia).
Figure 3.
Gene expression analysis and viral load in CEF of Aseel and Kadaknath. (a, and b) Heatmap and Boxplot respectively, illustrating the various levels of gene expression at different hours post-infection (hpi) of ND virus infection. (c) Boxplot showing the cumulative gene expression of each gene in the Kadaknath and Aseel throughout time. (d) A line graph depicts the absolute quantity of viral load based on T-NP plasmid standards at various time point in CEF cells.
Figure 3.
Gene expression analysis and viral load in CEF of Aseel and Kadaknath. (a, and b) Heatmap and Boxplot respectively, illustrating the various levels of gene expression at different hours post-infection (hpi) of ND virus infection. (c) Boxplot showing the cumulative gene expression of each gene in the Kadaknath and Aseel throughout time. (d) A line graph depicts the absolute quantity of viral load based on T-NP plasmid standards at various time point in CEF cells.
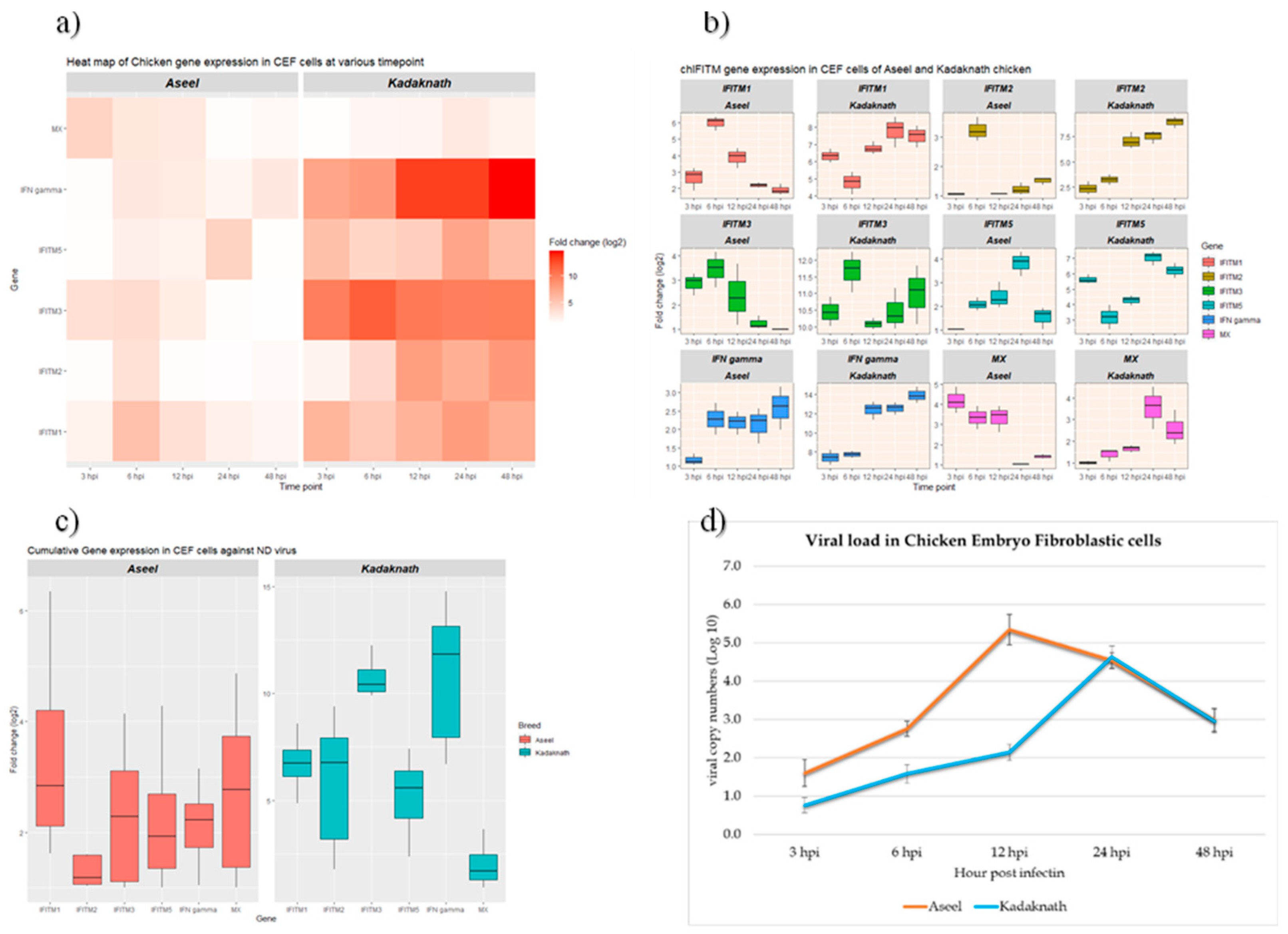

Table 1.
List of primer sequences for qRT-PCR.
| Gene | Primer | Sequence (5'->3') | Reference |
|---|---|---|---|
| chIFITM1 | FP | GCAGGATGTGACCACCACTA | NM_001350059.2 |
| RP | CTTCGCTGTCCTCCCATAGC | ||
| chIFITM2 | FP | AACAGGCGGAGGTGAGCAT | NM_001350058.2 |
| RP | AAGATGAGCGAGGGGAAGCA | ||
| chIFITM3 | FP | CGTGAAGTCCAGGGATCGCA | NM_001350061.2 |
| RP | GCAACCAGGGCGATGATGAG | ||
| chIFITM5 | FP | CCAACCCCACTTCTGGACGA | NM_001199498.1 |
| RP | ATCACTCCGAAGGGCACGAC | ||
| chMx | FP | GTCCAAGAGGCTGAATAACAGAG | NM_204609 |
| RP | GTCGGATCTTTCTGTCATATTGG | ||
| chIFN-γ | FP | TGAGCCAGATTGTTTCGATG | [24] |
| RP | CTTGGCCAGGTCCATGATA | ||
| chβ-Actin | FP | TATGTGCAAGGCCGGTTTC | |
| RP | TGTCTTTCTGGCCCATACCAA | ||
| NDV-NP | Pla-rt13 | CAACAATAGGAGTGGAGTGTCTGA | [22] |
| Pla-rt14 | CAGGGTATCGGTGATGTCTTCT |
Table 2.
Chicken embryo fibroblastic cells' gene expression fold changes (log2 (2-ΔΔCt)) in response to the Newcastle disease virus infection.
Table 2.
Chicken embryo fibroblastic cells' gene expression fold changes (log2 (2-ΔΔCt)) in response to the Newcastle disease virus infection.
| Breed | Time point | IFITM1 | IFITM2 | IFITM3 | IFITM5 | IFN-γ | MX |
|---|---|---|---|---|---|---|---|
| 3 hpi | 2.64±0.40c | 1.06±0.01c | 2.87±0.26b | 1.01±0.00b | 1.17±0.09c | 4.18±0.38a | |
| 6 hpi | 6.00±0.25a | 3.25±0.24a | 3.46±0.42a | 2.07±0.16b | 2.28±0.24ab | 3.35±0.33b | |
| Aseel | 12 hpi | 3.88±0.35b | 1.06±0.00c | 2.38±0.71b | 2.41±0.32b | 2.18±0.18b | 3.34±0.38b |
| 24 hpi | 2.19±0.09c | 1.22±0.12bc | 1.26±0.16c | 3.82±0.30a | 2.13±0.28b | 1.03±0.01c | |
| 48 hpi | 1.91±0.20cd | 1.51±0.08b | 1.00±0.00c | 1.53±0.27c | 2.59±0.34a | 1.43±0.06c | |
| 3 hpi | 6.33±0.24b | 2.39±0.38d | 10.44±0.26b | 5.63±0.17b | 7.46±0.45c | 1.01±0.05c | |
| 6 hpi | 4.77±0.38c | 3.25±0.29c | 11.68±0.36a | 3.18±0.46c | 7.77±0.21c | 1.38±0.16c | |
| Kadaknath | 12 hpi | 6.77±0.21b | 7.02±0.47b | 10.08±0.10b | 4.28±0.19bc | 12.38±0.54b | 1.66±0.10c |
| 24 hpi | 7.79±0.52a | 7.48±0.36b | 10.47±0.36ab | 7.04±0.27a | 12.56±0.39b | 3.57±0.56a | |
| 48 hpi | 7.48±0.36a | 8.89±0.33a | 11.00±0.51a | 6.22±0.27a | 13.9±0.49a | 2.56±0.46b |
Mean value with different superscript shows significant different at p<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



