Submitted:
28 April 2023
Posted:
29 April 2023
You are already at the latest version
Abstract
Ants of the genus Solenopsis are important invasive pests in the world. Due to their high dispersion capacity, aggressive behavior and because they feed on various sources of vegetable and animal protein, these insects have caused serious damage to public health and agriculture. The control has usually been carried out through the use of organosynthetic insecticides. However, due to the ecological and environmental impacts caused by the use these products, more sustainable alternatives have been sought for their control. Faced with these problems and due to the scarcity of registered products for the control of these insects, we aimed with this study to synthesize and evaluate the anticidal potential of carvacrol derivatives. The lethal and sublethal effects caused by these derivatives were compared with the essential oil of Lippia gracilis (50.7% carvacrol) and with the base molecule — carvacrol. Carvacryl benzoate was the most toxic derivative to Solenopsis sp. with an LD50 of 3.20 g/ mg. This compound was about 2 and 7.6 times, more toxic than carvacrol at the doses needed to kill 50 and 90% of populations, respectively. The workers of Solenopsis sp. showed a rapid reduction in survival when exposed to carvacrol (LT50 = 8.43h) and carvacryl benzoate (LT50 = 8.87h). Insects treated with sublethal doses of the compounds did not show significant effects on self-cleaning, allogrooming and aggregation, with the exception of those treated with L. gracilis essential oil. The oil increased self-cleaning and reduced allogrooming and aggregation. Ants treated with carvacrol and carvacryl benzoate showed greater distance and speed when compared to the control. These compounds decreased meanders and angular velocity. When live workers were exposed to dead individuals at the LD90 of the compounds, carvacryl benzoate was the derivative that most reduced insects survival, due to horizontal transfer. These results show the potential of the carvacrol derivative — carvacryl benzoate — for the alternative management of ants of the genus Solenopsis sp.
Keywords:
Insecta
; pest control
; ecotoxicology
; behavior
; biological products
; germoplasm
; essential oil
; insecticide
; necrophoresis
1. Introduction
Ants of the genus Solenopsis Westwood (Hymenoptera: Formicidae) — known as fire ants or red fire ants [1,2] — originate from South America, but have spread to various parts of the world, being found in countries such as the United States [3], South Korea [4], Japan, China and New Zealand [5]. These ants are considered invasive, mainly in disturbed habitats [6,7]. Several features make Solenopsis sp. good invaders, such as: high dispersal capacity [8], ample diet [9], high aggressiveness [1,10], in addition to being good competitors [11] and having large colonies.
The invasion of Solenopsis sp. in new areas makes management difficult, generation major impacts on public health and agriculture [5,12,13]. In general, it is estimated that the economic costs caused by ants of the genus Solenopsis, in invaded areas, can reach about 3.49 billion dollars [14]. In agricultural areas, ants cause direct damage to several crops [2,3]; in addition to indirect damage, via reduction of natural biological control of populations of sucking insect pests. This occur through mutualistic interactions with sucking insects (eg aphids and cochineal), where ants perform pest defense against predators, in exchange for honeydew (a sugary substance) [2,15,16]. Because they are very aggressive, they can also attack several species causing ecological disturbances [3]. In relation to public health, they can contribute to the dissemination of pathogens in hospital environments [17]; and their bites can cause irritation and, in more severe cases, anaphylaxis and breathing difficulties [8,9,10,11,12,13,14,15,16,17,18,19]. In addition, they can cause damage to equipament and buildings [14].
Such disorders caused by Solenopsis sp. require control measures with direct applications on anthills and adjacent areas [20]. Attempts at control are mainly made with organosynthetic insecticides [9,21] from the pyrethroid group [22], neonicotinoids and growth regulators [23]. The products used can be slow-acting (eg toxic baits) or fast-acting (eg contact insecticides); the use of repellents and fumigants is also common [17]. However, the persistent and inappropriate use of these products can lead to ecological disturbances, either directly or through residual action [9,24,25,26].
In this context, plant essential oils (EOs) and their constituents can be effective as insecticides and act more sustainably [27]. The monoterpene carvacrol, for example, has already know bioactivity on the tick Rhipicephalus (Boophilus) microplus [28] and insect pests such as the termite Cryptotermes brevis [29] and the larvae Diaphania hyalinata [30]. Additionally, such compounds are interesting chemical models for the synthesis of new derivatives that may have their bioactivity Enhanced [31]. Due to the wide variety of constituents present in plant EOs, they can cause different effects on insects by acting as repellents, attractants and oviposition deters [24,31]; in addition to promoting behavioral changes that can alter insect communication [29,30,31,32,33,34,35,36,37]. Such effects may be desirable for controlling eusocial insects, for which interindividual communication is of paramount importance for colony cohesion [33,38].
Prophylactic strategies are recognized for promoting the first line of defense of eusocial insects against pathogens, increasing colony health and viability [40,41]. Among such strategies, necrophoresis (removal of bodies) and allogrooming (interindividual cleaning) can reduce infection within colonies. Such behaviors are adaptative, since colonies often have high densities of genetically related individuals, constantly involved in collaborative activities [42]. However, studies report that contacts between colonial individuals can contribute to the transfer of insecticides, increasing the spread of contaminants within the nest. This horizontal transfer of insecticides may thus constitute an important control mode [43,44].
2. Materials and Methods
2.1. Insects
Ant workers of the genus Solenopsis were collected using attractive food baits (animal protein) distributed near nests located in a home environment in the municipality of Aracaju, SE, Brazil (10° 53′ S, 37° 06′ W). The ants were collected with the aid of a soft bristle brush, placed in 200 mL plastic pots, covered with organza fabric and sent to the Integrated Pest Management Laboratory located at the Federal University of Sergipe (UFS), São Cristóvão, SE, Brazil (10° 54′ S, 37° 04′ W).
The insects were then placed in plastic containers (11 cm x 7.5 cm), covered with plastic film (G-útil Guarufilme, Guarulhos, SP, Brazil) and kept under controlled conditions (27 ± 2 °C, 70 ± 5% RH and 12 hour photoperiod) for 24h for acclimatization, until the experiments were carried out. During this period, only distilled water was provided to the workers.
2.2. Synthesis of carvacrol derivatives
The derivatives were synthesized at the Pharmaceutical Chemistry Laboratory of the UFS University through nucleophilic nuclear substitution and esterification reactions. Nine compounds were synthesized from the base molecule carvacrol: the esters carvacryl acetate, carvacryl benzoate, carvacryl butyrate, carvacryl hexanoate, carvacryl isobutyrate, carvacryl isovalerate, carvacryl pivaloate, carvacryl ethyl ether and carvacryl trichloroacetate (Figure 1).
The ether compound was synthesized according to the method described in Collen [47]. The esters, however, followed the methodologies of Dolly & Barba, Ben Arfa et al., Morais et al. [48,49,50], using THF (tetrahydrofuran) as solvent [31].
The reactions were followed by thin layer chromatography (TLC), verified in ultraviolet light of 256 ηm and compared with the starting material (carvacrol). Derivatives were purified using sílica gel 60 in the stationary phase and pure hexane in the mobile phase. Melting points were determined on a Logen Scientific melting point apparatus without correction. The Chemicals used in the reactions and the monoterpene carvacrol were obtained from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA).
2.3. Bioassays
The synthetic derivatives were compared with the precursor molecule carvacrol and with the EO of L. gracilis, rich in carvacrol (50,7%) [30]. The EO of L. gracilis was obtained of plants that are maintained at the UFS Active Germplasm Bank – located at the experimental farm – UFS Rural Campus, São Cristóvão, SE, Brazil (10° 55′ S, 37° 11′ W). The extraction, identification and quantification of compounds was performed as described in Santos et al. [51].
The bioassays were conducted in climate-controlled rooms with controlled conditions (27 ± 2 °C, 70 ± 5% RH and 12-hour photoperiod) at the Integrated Pest Management Laboratory at UFS. The compounds used were diluted in acetone solvent (Panreac, UV-IR-HPLC-GPC PAI-ACS, 99,9% purity) and applied topically (0.5 µL) in the prothorax region of the workers, with the aid of a Hamilton® microsyringe (Reno, NV, USA). In the negative control, only acetone was used.
In order to facilitate the application of the compounds, the ants were immobilized in a freezer at -19 °C (Eletrolux®, Curitiba, PR, Brazil) for 90 s. Preliminary tests have shown that this method does not affect the survival and behavior of ants. To determine the doses to be applied, Fifty workers were weighed on a precision analytical balance (Shimadzu, AUW220D) to obtain the average mass of the insects.
In the acute toxicity bioassays, the treatments used were the EO of L. gracilis, the base molecule carvacrol and the nine synthetic derivatives. In the behavioral bioassays, EO of L. gracilis, carvacrol and its most toxic synthetic derivative (carvacryl benzoate) were used.
2.3.1. Acute toxicity
The bioassays to determine the dose-response curves and lethal doses were conducted in a completely randomized design with eight replications per treatment. The experimental unit consisted of a group of six to seven worker ants of Solenopsis sp. of the same size, totaling 4144 individuals. Initially, tests were carried out with three doses (1, 5 and 10 µg of substance/mg of insect) in order to obtain mortalities between 5 and 95%. Intermediate doses were then tested to establish dose-response curves.
The ants were placed in a Petri dish (Global Trade Technology, Monte Alto, SP, Brazil) (6 x 1.5 cm) lined with filter paper (Unfil) moistened with 0.4 mL of distilled water and covered with plastic film (G-util Guarufilme, Guarulhos, SP, Brazil). Mortality assessment was performed 48h after setting up the bioassays. The criterion was based on the mobility of ants after being stimulated with a soft bristle brush. Insects that did not move after the stimulus were considered dead.
In the bioassays to obtain survival curves and lethal time of the compounds on workers of Solenopsis sp. the LDs90 obtained in the acute toxicity bioassays were used. The procedures were similar to those used in toxicity bioassays, with the exception of doses, number of repetitions and evaluation time. Fifteen repetitions were performed per treatment, totaling 420 individuals. The first mortality assessments were performed every 10 min until completing 2h; then every 30 min until completing 6 h; followed by 1 h assessments during the first 12 h and every 2 h until reaching the first 24 h. Subsequently, evaluations were carried out at intervals of 4 hours in the following 12 hours and every 12 hours until completing 100 hours.
2.3.2. Individual and collective behavior
For this experiment, we observed the individual behavior of Solenopsis sp. treated (individual behavior) (n = 1) and the interaction of a group of ants not exposed to the compounds (6 ants) with a treated one (collective behavior) (n = 7).
The experimental designs were completely randomized, with 60 repetitions per treatment. The workers of Solenopsis sp. were exposed to acetone (negative control) and to the LDs30 of the compounds determined in the acute toxicity bioassays. Before being exposed to the treatments, the ants were placed in a Petri dish (6 x 1.5 cm) lined with filter paper moistened with 0.4 mL of distilled water for 5 min for acclimatization.
In the experiment to evaluate individual behavior, after acclimatization and immobilization the ants were treated and placed in Petri dishes individually, totaling 240 individuals. After 1 min of exposure, self-cleaning behavior was counted. As for the collective behavior experiment, after acclimatization and immobilization, an ant was randomly removed from the group of seven ants and marked with non-toxic yellow paint (Acrilex Tintas Especiais S.A., São Bernardo do Campo, São Paulo, Brazil). After 3 min of marking with the ink, the ant received the treatment and after 1 min it was relocated in the Petri dish with the other untreated ants. One min after the relocation on the board, the counting of the antenation, cleaning and aggregation behaviors began. Preliminarly tests indicated that the paint did not affect the survival and behavior of the ants.
In both bioassays, observations were performed for one continuous minute, with a one-minute interval between them, during a period of 10 minutes, totaling 5 minutes of observation for each insect. Overall, behaviors were recorded for 2400 min [2 bioassays x 4 treatments x 60 repetitions x 5 min of observation].
2.3.3. Walking behavior
The individual walking behavior was evaluated with workers of Solenopsis sp. submitted to the LDs30 of each treatment and acetone (negative control). The experiments were carried out in a completely randomized design, with 60 repetitions per treatment, totaling 240 individuals. The bioassays were performed in Petri dish arenas (6 x 1.5 cm), lined with moistened filter paper and covered with plastic film.
After 1 min of placing the individuals in the arena, recording began for a period of 600 s using a vídeo camera (Panasonic SD5 Superdynamic – model WV-CP504), equipped with a Spacecom lens (1/3” 3-8 mm) attached to a computer. The distance traveled (mm), velocity (mm/s), meander (°/mm) and angular velocity (°/s) were captured in Ethovision XT software (version 8.5; Noldus Integration System, Sterling, VA) and data analyzed using Studio 9 software (Pinnacle Systems, Moutain View, CA).
2.3.4. Horizontal transfer of compounds
The aim of this experiment was to investigate whether the compounds used in this study can be transferred from dead ants to live ants that were not subjected to the treatments. Two proportions were used in the experiments, [1:20] = 1 dead ant to 20 live ants and [1:5] = 4 dead ants to 20 live ants. The design was completely randomized with ten repetitions, totaling 800 live individuals and 200 cadavers. The bioassays were performed in Petri dish arenas (6 x 1.5 cm), lined with filter paper moistened with 0.4 mL of distilled water and covered with film plastic.
Dead ants were obtained after 24 h of exposure of the workers to the LDs90 of the compounds. For the control, cadavers were obtained after application of acetone and freezing in a freezer at -19 °C for 30 min. The number of live and dead ants was evaluated after 1, 2, 4, 6, 10, 14, 26, 38, 50, 62, 74, 86, 98, 110 and 122 hours after setting up the experiments.
2.4. Statistical analysis
Mortality data were submitted to Probit analysis to determine the dose-response curve for each treatment using the PROC PROBIT procedure in SAS (SAS Institute, 2008). From these curves, the lethal doses necessary to cause 30, 50 and 90% mortality (LD30, LD50 and LD90) and their respective confidence limits at 95% probability (LC95) were obtained. The determination of the most toxic synthetic derivative was performed using the criterion of non-overlapping confidence intervals with the origin of the interval in the LDs50.
For analysis of survival, horizontal transfer and their respective lethal times, Kaplan-Meier estimators were used through the Log-Rank test (SigmaPlot, version 14). From this analysis, survival curves and lethal times (LT50) were obtained for each treatment. The Holm-Sidak multiple coparison method was used at a significance level of 0.05 to compare the curves (SigmaPlot, version 14). The LT50 were compared by the criterion of non-overlapping confidence intervals with the origin of the interval.
The data from the behavioral bioassays (individual, collective and walking) (n = 60) were analyzed by parametric tests, considering that the data distribution is close to the Gaussian distribution (Central limit theorem) and, therefore, meets the assumptions of the analysis of variance (ANOVA). Thus, these data were initially submitted to ANOVA followed by Dunnett’s test (p < 0.05) to verify differences between treatments in relation to the control. All behavioral data analyzes were performed using SigmaPlot software, version 14.
3. Results
3.1. Acute toxicity
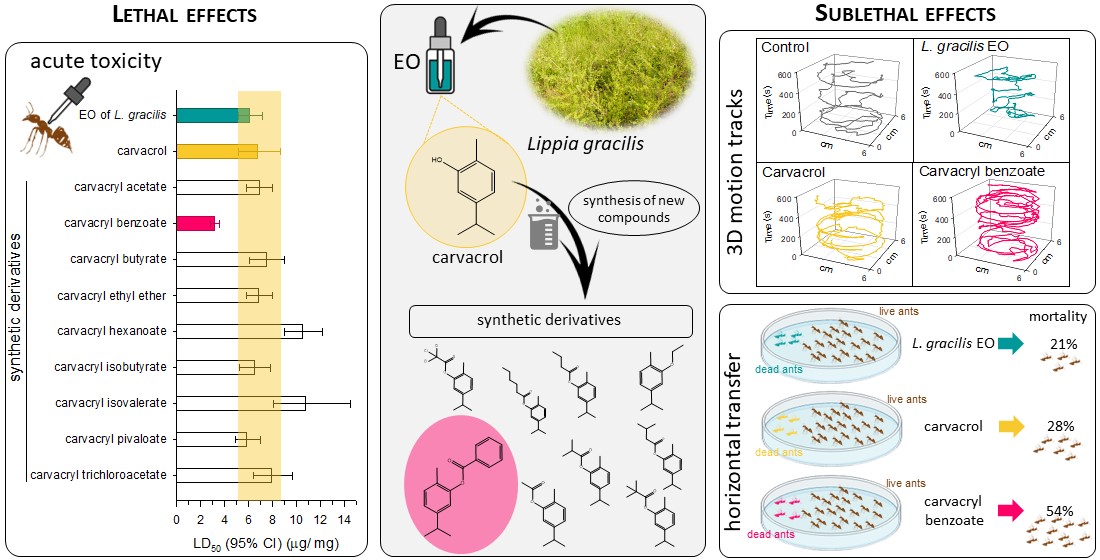
All synthetic carvacrol derivatives were bioactive against Solenopsis sp. The doses needed to kill 50% of ant populations ranged from 3.2 to 10.8 µg/mg (Table 1). Carvacryl benzoate was the most toxic derivative for ants (3.2 µg/mg; CI95%: 2.86-3.57). Its LD50 was about 2 times lower than those observed for the essential oil of L. gracilis (6.11 µg/mg; CI95%: 5.16-7.20) and its major compound carvacrol (6.79 µg/mg; CI95%: 5.19-8.71). The other synthetic derivatives showed similar or lower toxicities than the base molecule carvacrol (Table 1).
The greatest slope of the dose-response curves was observed for carvacryl benzoate (3.2 ± 0.24), which resulted in greater toxicities at higher DLs. This compound was 3.2 and 7.6 times, more toxic than EO from L. gracilis and carvacrol at the dose considered standard control in Brazil (LD90) [53,54], respectively (Table 1).
Survival of Solenopsis sp. exposed to the LDs90 of the EO of L. gracilis, its major compound carvacrol and the most toxic synthetic derivative (carvacryl benzoate), was significantly reduced over time (χ2 = 359.4; d.f. = 3; p < 0.001) (Figure 2). The workers of Solenopsis sp. showed a rapid reduction in survival when exposed to carvacrol (LT50 = 8.43 h; CI95% = 5.37-11.50) and carvacryl benzoate (LT50 = 8.87 h; CI95% = 7.04-10.7) (Figure 2). In less than 60 h all individuals died. There was no significant difference between the survival curves for ants exposed to these two compounds (P = 0.279). On the other hand, the EO of L. gracilis caused slower mortality in Solenopsis sp. It took 39.6 h (CI95% = 32.7-46.5) for this oil to kill 50% of populations (Figure 2). Despite reducing survival more slowly, the oil caused 100% mortality within 84 h of exposure (Figure 2).
3.2. Individual and collective behavior
Data on individual self-cleaning behavior (F3;236 = 10.35; P < 0.001) and collective allogrooming (F3;236 = 5.23; P < 0.002) and aggregation (F3;236 = 4.62; P < 0.004) from Solenopsis sp. varied between treatments. The antennation behavior did not vary between treatments (F3;236 = 0.66; P < 0.577).
3.3. Walking behavior
The distance covered (F3;236 = 9.32; P < 0.001) and the speed (F3;236 = 8.73; P < 0.001) of Solenopsis sp. varied between treatments (Figure 4). Workers treated with carvacrol and carvacryl benzoate increased the distance covered (Figure 4A) and speed (Figure 4B).
Individuals treated with the LD30 of these compounds increased the distance walked by 38 and 27% in relation to the control (817.7 mm), respectively (Figure 4A). The displacement behavior of the ants treated with the EO of L. gracilis did not differ from the control (Figure 4).
Meander behavior (F3;236 = 14.07; P < 0.001) and angular velocity (F3;236 = 7.63; P < 0.001) of Solenopsis sp. varied between treatments (Figure 5). The normal walking activity (control) of Solenopsis sp. involves greater change in direction of movement by distance traveled (meander = 89.6 ± 7.7 °/mm) and by time (angular velocity = 26.4 ± 2.1 °/s) (Figure 5). Workers treated with carvacrol and carvacryl benzoate reduced meander (62.2 ± 7.1 and 36.1 ± 4.6 °/mm) and angular velocity (20.0 ± 2.1 and 14.7 ± 1.4 °/s) versus the control, respectively (Figure 5). The movement of workers treated with these two compounds was more circular around the edge of the arena and with little change in direction (2D and 3D graphics) (Figure 5). The EO of L. gracilis did not affect these parameters (Figure 5).
3.4. Horizontal transfer of compounds
In general, mixing dead ants submitted to the LD90 of the compounds with live ants reduced the survival of workers of Solenopsis sp. over time (χ2 = 200.8; d.f. = 7; p < 0.001) (Figure 6). There was no significant difference between the survival curves of EO from L. gracilis (P = 0.833) and carvacrol (P = 0.951) in the two proportions (1:20 and 1:5) (Figure 6).
The greatest horizontal transfer occurred when the ants were treated with carvacryl benzoate and the corpses were placed together with live ants in a 1:5 ratio. At the end of the experiment, this compound caused death in 54% of the ants, requiring 78.5 h (CI95% = 71.9-85.1) for the compound to kill 50% of the populations (Figure 6).
4. Discussion
Plant essential oils and their compounds are considered biorational and environmentally safe products [55]. Here we show that the EO of L. gracilis, its major compound carvacrol and the synthetic derivative compound carvacryl benzoate, showed relevant lethal and sublethal effects for the control of ants of the genus Solenopsis.
Compounds from EOs of plants act on several sites of action such as octopamine receptors, sodium channels, acetylcholine receptors, glutamate-mediated chloride channels and GABA receptors [55], having potential for use in integrated pest management programs [56]. Carvacrol has already proven to be toxic to several pests such as the common bug Cimex lectularius Linnaeus [57], the housefly Musca domestica [58], the corn weevil Sitophilus zeamais Motschulsky [59], the cotton bollworm Helicoverpa armigera Hübner [60] and the larvae Diaphania hyalinata [30]. Although there is no consensus regarding its molecular mechanism and its main target of action, it is believed that carvacrol inhibits the activity of the enzyme acetylcholinesterase [61]. It is known, however, that carvacrol also acts as a positive allosteric modulator in the GABA receptor, enhancing the uptake of Cl- anions in the presence of GABA. Thus, this xenobiotic can act on different sites of action [55] and have multiple targets [62].
Carvacrol is a phenolic and aromatic monoterpene [63], considered a good precursor molecule [31,64]. Studies on synthetic derivatives make it possible to assess which structural properties of a molecule are involved in its activity; thus allowing the Discovery of new synthetic routes and the large-scale production of bioactive molecules [66]. Among the synthetic carvacrol derivatives used in this study, carvacryl benzoate was the most toxic, showing a lower LD50 than that of the parent compound (carvacrol). The intensity of the toxic effect may be related to chain length, functional groups and substituent position (eg meta- or ortho-) [67]. Our results suggest that the substitution of the hydroxyl in the ortho position of carvacrol increases the toxic action on workers of Solenopsis sp.
Although all compounds were toxic to Solenopsis sp. workers, mortality occurred more quickly with carvacrol and its derivative carvacryl benzoate (LT50 ~ 9h); while the mortality of ants treated with EO from L. gracilis was slower (LT50 ~ 40h). This difference may be related not only to the toxicity of the compound itself, but also to the interaction with sublethal effects that can alter the behavior of ants. Through interindividual behaviors such as allogrooming and aggregation, contaminated ants can transfer toxic compounds to other members of the colony. Here we found that ants treated with EO from L. gracilis increased self-cleaning behaviors and reduced allogrooming, possibly avoiding contact with other nestmates. Additionally, EO also caused less aggregation, which may be related to a possible repellent effect of EO on ants, resulting in repulsion between individuals. Although these behaviors reduce contact between ants, these results indicate the possibility of using EO from L. gracilis as a repellent for Solenopsis sp. Repellent action is a desirable effect to prevent the presence of ants in unwanted places [65,66]. In turn, carvacrol and its derivative carvacryl benzoate did not change self-cleaning and alogrooming behaviors in relation to the control, which may result in a rapid horizontal contamination within the colony. This horizontal transfer is of great relevance for the management of ants [43,44], especially when the location of the anthill is difficult to access [67].
The horizontal transfer, allied to the high toxicity of carvacryl benzoate, had a relevant effect, increasing the speed of death of both ants: donor and recipient. This effect was even more intensified when there was a higher cadaver: healthy worker ratio. Thus, since carvacryl benzoate was more toxic and could result in a greater number of corpses inside the nest, this compound could result in a high horizontal transfer rate between workers of Solenopsis sp. In colonies of this species, the oldest workers ate the most exposed to the action of insecticides, as they are close to the nest and in the foraging areas [68]. The impact of contaminants on these ants can result in a lower ability of the colony to obtain food, with lower rates of recruitment and energy exchange among nestmates.
The sublethal effects can also interfere with foraging by altering the walking ability and orientation of Solenopsis sp. workers. Our results show that the normal activity of Solenopsis sp. involves low walking speed and changes in movement direction (average angle the subject turned during the vídeo) by distance covered (meander - °/s) and by time (angular velocity - °/s). Possibly this behavior is related to the pattern of exploration of the environment by these ants.
Animals that walk with greater changes in the direction of movement increase the change of finding nearby resources, while fewer meanders and low angular velocity may be related to the search for more distant resources [69]. The exposure of ants to carvacrol and carvacryl benzoate caused an increase in the distance covered and walking speed, as well as a reduction in the number of meanders and angular speed; likely making this behavior more directional. Some authors point out that the increase in walking speed could indicate an attempt to escape by ants, in order to avoid the contaminant [70,71]. Future studies could evaluate how such compounds interfere in the way ants explore the environment and in their ability to acquire energy.
In conclusion, the results obtained in this work suggest an avoidance behavior of nestmates treated with the EO of L. gracilis; and demonstrate the activity of carvacrol and specially its derivative carvacryl benzoate on mortality, behavioral changes and spread of compounds among nestmates. Thus, our work demonstrates the potential of these compounds for the development of new products for the management of Solenopsis ants.
Author Contributions
Conceptualization, A.P.A.A., J.O.D. and L.B.; methodology, J.E.S., M.C.P. and L.B.; validation, L.B.; formal analysis, J.E.S. and L.B.; investigation, J.O.D., J.E.S., V.S.A., G.S.M. and T.B.B.; resources, S.C.H.C. and A.F.B.; data curation, J.O.D., V.S.A., G.S.M. and T.B.B.; writing—original draft preparation, J.O.D. and J.E.S.; writing—review and editing A.P.A.A. and L.B.; visualization, S.C.H.C., A.P.A.A., A.F.B., M.C.P.; supervision, L.B.; project administration, L.B.; funding acquisition, S.C.H.C., A.F.B. and L.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico – Brasil (CNPq), the Fundação de Apoio à Pesquisa e a Inovação Tecnológica do Estado de Sergipe (FAPITEC/SE) – Brasil, the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001, and the Financiadora de Estudos e Projetos – Brasil (FINEP).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
The authors acknowledge Universidade Federal de Sergipe and funding sources for the financial support in publishing this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pitts, J.P.; Mchugh, J.V.; Ross, K.G. Cladistic analysis of the fire ants of the Solenopsis saevissima species-group (Hymenoptera: Formicidae). Zoologica Scripta. 2005, 34, 493-505. [CrossRef]
- Chan, K.H.; Guénard, B. Ecological and socio-economic impacts of the red import fire ant, Solenopsis invicta (Hymenoptera: Formicidae), on urban agricultural ecosystems. Urban Ecosystems. 2020, 23,1-12. [CrossRef]
- Wylie, F.R.; Janssen-May, S. Red imported fire ant in Australia: what if we lose the war? Ecological Management & Restoration. 2017, 18, 32-44. [CrossRef]
- Lyu, D.P.; Lee, H.S. The red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae: Myrmicinae) discovered in Busan sea port, Korea. Korean journal of applied entomology. 2017, 56, 437-438. [CrossRef]
- Xu, Y.; Vargo, E.L.; Tsuji, K.; Wylie, R. Exotic Ants of the Asia-Pacific: Invasion, National Response, and Ongoing Needs. Annual Review of Entomology. 2022, 67, 27-42. [CrossRef]
- LeBrun, E.G.; Plowes, R.M.; Gilbert, L.E. Imported fire ants near the edge of their range: disturbance and moisture determine prevalence and impact of an invasive social insect. Journal of Animal Ecology. 2012, 81, 884-895. [CrossRef]
- Tschinkel, W. R. The fire ants. Harvard University Press, 2013.
- Drees, B.M.; Calixto, A.A.; Nester, P.R. Integrated pest management concepts for red imported fire ants Solenopsis invicta (Hymenoptera: Formicidae). Insect Science. 2013, 20, 429-438. [CrossRef]
- Du, Y.; Zhou, A.; Chen, J. 2021. Olfactory and behavioral responses to acetate esters in red imported fire ant Solenopsis invicta. Pest Management Science. 2021, 77, 1371-1382. [CrossRef]
- Lard, C.F.; Schmidt, J.; Morris, B.; Estes, L.; Ryan, C.; Bergquist, D. 2006. An economic impact of imported fire ants in the United States of America. Texas A&M University, Department of Agricultural Economics, Texas Agricultural Experiment Station, College Station, TX. 2006.
- Xu, C.; Su, J.; Qu, X.; Zhou, A. Ant-mealybug mutualism modulates the performance of co-occurring herbivores. Scientific reports. 2019, 9, 1-11. [CrossRef]
- Gruber, M.A.M.; Janssen-May, S.; Santoro, D.; Cooling, M.; Wylie, R. Predicting socio-economic and biodiversity impacts of invasive species: Red Imported Fire Ant in the developing western Pacific. Ecological Management & Restoration. 2021, 22, 89-99. [CrossRef]
- Park, Y.; Vatanparast, M.; Sajjadian, S.M. Pathogenicity of Beauveria bassiana ANU1 to the red imported fire ant, Solenopsis invicta workers in Korea. Journal of Asia-Pacific Entomology. 2022, 25, 101913. [CrossRef]
- Angulo, E.; Hoffman, B.D.; Ballesteros-Mejia, L.; Taheri, A.; Balzani, P.; Bang, A.; Renault, D.; Cordonnier, M.; Bellard, C.; Diagne, C.; Ahmed, D. A.; Watari, Y.; Courchamp, F. Economic costs of invasive alien ants worldwide. Biological invasions. 2022, 24, 2041-2060. [CrossRef]
- Xu, T.; Xu, M.; Lu, Y.; Zhang, W.; Sun, J.; Zeng, R.; Turlings, T. C. J.; Chen, L. A trail pheromone mediates the mutualism between ants and aphids. Current Biology. 2021, 31, 4738-4747. [CrossRef]
- Montgomery, M.P.; Vanderwoude, C.; Lintermans, M.; Lynch, A.J.J. The Little Fire Ant (Hymenoptera: Formicidae): A Global Perspective. Annals of the Entomological Society of America. 2022, 115, 427-448. [CrossRef]
- Chen, J.; OI, D.H. Naturally occurring compounds/materials as alternatives to synthetic chemical insecticides for use in fire ant management. Insects. 2020, 11, 758. [CrossRef]
- Guillet, C.; Martin, O.Y.; Meincke, C.; Joerg, L.; Schmid-Grendelmeier, P. Part I: Insect stings and bites—Beyond the realm of bee and wasp allergies. Allergo Journal International. 2022, 31, 183-193. [CrossRef]
- Vinson, S.B. Impact of the invasion of the imported fire ant. Insect Science. 2013, 20, 439-455. [CrossRef]
- Fu, J.T.; Tang, L.; Li, W.S.; Wang, K.; Cheng, D.M.; Zhang, Z.X. Fumigant toxicity and repellence activity of camphor essential oil from Cinnamonum camphora Siebold against Solenopsis invicta workers (Hymenoptera: Formicidae). Journal of Insect Science. 2015, 15, 129. [CrossRef]
- Li, Y.; Yu, S.; Huang, J.; Wang, Z.; Zeng, Y.; Wu, X.; Han, K.; Zhou, H.; Wang, G.; Yu, Z. 2022. Study of behavioral, electrophysiological response, and the active compounds of the essential oils from six kinds of flowers against Solenopsis invicta Buren (Hymenoptera: Formicidae). Industrial Crops and Products. 2022, 188, 115603. [CrossRef]
- Wen, C.; Chen, J.; Qin, W. Q.; Chen, X.; Cai, J. C.; Wen, J. B.; Wen, X. J.; Wang, C. 2021. Red imported fire ants (Hymenoptera: Formicidae) cover inaccessible surfaces with particles to facilitate food search and transportation. Insect Science. 2021, 28, 1816-1828. [CrossRef]
- Tsai, C.C.; Hung, S.H.; Lin, X.R.; Huang, R.N. Herbal plants as alternatives for the management of the red imported fire ant, Solenopsis invicta. Journal of Applied Entomology. 2022, 146, 975-989. [CrossRef]
- Song, Z.; Wang, Y.; Li, C.; Tan, Y.; Wu, J.; Zhang, Z. Fumigant toxicity and behavioral inhibition of garlic against red imported fire ants (Solenopsis invicta). Environmental Science and Pollution Research. 2022, 30, 1889-1897. [CrossRef]
- Xiao, C.X.; Tan, Y. T.; Wang, F. F.; Wu, Q.H.; Qin, Q.; Zhang, Z.X. The Fumigating Activity of Litsea cubeba oil and Citral on Solenopsis invicta. Sociobiology. 2020, 67, 41-47. [CrossRef]
- Li, Y.; Yu, S.; Huang, J.; Wang, Z.; Zeng, Y.; Wu, X.; Han, K.; Zhou, H.; Wang, G.; Yu, Z. Study of behavioral, electrophysiological response, and the active compounds of the essential oils from six kinds of flowers against Solenopsis invicta Buren (Hymenoptera: Formicidae). Industrial Crops and Products. 2022, 188, 115603. [CrossRef]
- Mostafiz, M.M.; Hassan, E.; Lee, K.Y. Methyl Benzoate as a Promising, Environmentally Safe Insecticide: Current Status and Future Perspectives. Agriculture. 2022, 12, 378. [CrossRef]
- Cruz, E. M.O.; Costa-Junior, L. M.; Pinto, J.A.O.; Santos, D.A, de Araujo, S.A.; Arrigoni-Blank, M.F; Bacci, L.; Alves, P.B.; Cavalcanti, SCH.; Blank, A. F. Acaricidal activity of Lippia gracilis essential oil and its major constituents on the tick Rhipicephalus (Boophilus) microplus. Veterinary Parasitology. 2013, 195, 198-202. [CrossRef]
- Santos, A.A.; de Oliveira, B.M.S.; Melo, C.R.; Lima, A.P.S.; Santana, E.D.R.; Blank, A.F.; Picanço, M.C.; Araújo, A.P.A.; Cristaldo, P.F.; Bacci, L. Sub-lethal effects of essential oil of Lippia sidoides on drywood termite Cryptotermes brevis (Blattodea: Termitoidea). Ecotoxicology and Environmental Safety. 2017, 145, 436-441. [CrossRef]
- Melo, C.R.; Picanço, M.C.; Santos, A.A.; Santos, I.B.; Pimentel, M.F.; Santos, A.C.C.; Blank, A.F.; Araújo, A.P.A.; Cristaldo, P.F.; Bacci, L. Toxicity of essential oils of Lippia gracilis chemotypes and their major compounds on Diaphania hyalinata and non-target species. Crop Protection. 2018, 104, 47-51. [CrossRef]
- Silva, V.B.; Travassos, D.L.; Nepel, A.; Barison, A.; Costa, E.V.; Scotti, L.; Scotti, M.T.; Mendonça-Junior, F.J.B.; Santos, R.L.C.; Cavalcanti, S.C.H. Synthesis and chemometrics of thymol and carvacrol derivatives as larvicides against Aedes aegypti. Journal of Arthropod-Borne Diseases. 2017, 11, 315-330.
- Santos, A.C.C., Cristaldo, P.F, Araújo, A.P.A., Melo, CR.; Lima, A.P.S., Santana, E.D.R.; Oliveira, B.M.S.; Oliviera, J.W.S.; Vieira, J.S.; Blank, A.F.; Bacci, L. Apis mellifera (Insecta: Hymenoptera) in the target of neonicotinoids: A one-way ticket? Bioinsecticides can be an alternative. Ecotoxicology and Environmental Safety. 2018,163, 28-36. [CrossRef]
- Costa, A.C.F.; Cavalcanti, S.C.H.; Santana, A.S.; Lima, A.P.S.; Santana, E.D.R.; Brito, T. B.; Oliveira, R.R.B.; Macêdo, N.A.; Cristaldo, P.F.; Bacci, L. Formicidal activity of indole derivatives on Atta opaciceps (Borgmeier): lethal, behavioral and locomotive effects. Journal of Applied Entomology. 2019, 143, 58-68. [CrossRef]
- Lima, A.P.S.; Santana, E.D.R.; Santos, A.C.C.; Silva, J.E.; Ribeiro, G.T.; Pinheiro, A.M, Santos, I.T.B.F.; Blank, A.F.; Araújo, A.P.A.; Bacci, L. Insecticide activity of botanical compounds against Spodoptera frugiperda and selectivity to the predatory bug Podisus nigrispinus. Crop protection. 2020, 136, 105230. [CrossRef]
- Feitosa-Alcantara, R.B., Bacci, L., Blank, A.F., Alves, P.B., Silva, I.M.A., Soares, C.A.; Sampaio, T.S.; Nogueira, P.C.L.; Arrigoni-Blank, M.F. Essential oils of Hyptis pectinata chemotypes: isolation, binary mixtures and acute toxicity on leaf-cutting ants. Molecules, 2017, 22, 621. [CrossRef]
- Rocha, A. G.; Oliveira, B.M.S.; Melo, C.R.; Sampaio, T.S.; Blank, A.F.; Lima, A.D.; Nunes, R.S.; Araújo, A.P.A.; Cristaldo, P.F.; Bacci, L. Lethal effect and behavioral responses of leaf-cutting ants to essential oil of Pogostemon cablin (Lamiaceae) and its nanoformulation. Neotropical entomology. 2018, 47, 769-779. [CrossRef]
- Oliveira, B.M.S.; Melo, C.R.; Santos, A.C.C.; Nascimento, L.F.A.; Nízio, D.A.C.; Cristaldo, P.F.; Blank, A.F.; Bacci, L. Essential oils from Varronia curassavica (Cordiaceae) accessions and their compounds (E)-caryophyllene and α-humulene as an alternative to control Dorymyrmex thoracius (Formicidae: Dolichoderinae). Environmental Science and Pollution Research. 2019, 26, 6602-6612. [CrossRef]
- Rytter, W.; Shik, J.Z. Liquid foraging behaviour in leafcutting ants: the lunchbox hypothesis. Animal Behaviour. 2016, 117, 179-186. [CrossRef]
- Müller, C. Impacts of sublethal insecticide exposure on insects—Facts and knowledge gaps. Basic and Applied Ecology. 2018, 30, 1-10. [CrossRef]
- Della Lucia, T.M.C.; Souza, D.J. Importância e história de vida das formigas-cortadeiras. In: Della Lucia, T.M.C. (Ed.). Formigas-cortadeiras: da bioecologia ao manejo. Viçosa: Editora UFV, 2011, 13-26.
- Della Lucia, T.M.C.; Amaral, K.D. Past and current strategies for the control of leaf-cutting ants in Brazil. In: Forest Pest and Disease Management in Latin America: Modern Perspectives in Natural Forests and Exotic Plantations. 2020, 31-43. [CrossRef]
- Diez, L; Lejeune, P; Detrain, C. Keep the nest clean: survival advantages of corpse removal in ants. Biology letters. 2014, 10, 20140306. [CrossRef]
- Liang, M.R.; Shuang, Y.M.; Deng, J.F.; Peng, L.Y.; Zhang, S.Q.; Zhang, C.; Xu, Y.J.; Lu, Y.Y.; Wang, L. Toxicity and horizontal transfer of bifenthrin and dimefluthrin against the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae), and the efficacy of their dust applications in the field. Journal of Integrative Agriculture. 2022. [CrossRef]
- Zhang, L; Wang, L.; Chen, J.; Zhang, J.; He, Y.; Lu, Y.; Cai, J.; Chen, X.; Wen, X.; Xu, Z.; Wang, C. Toxicity, horizontal transfer, and physiological and behavioral effects of cycloxaprid against Solenopsis invicta (Hymenoptera: Formicidae). Pest Management Science. 2022, 78, 2228-2239. [CrossRef]
- Santos, M. C.; Teodoro, A. V.; Menezes, M. S.; Pinto-Zevallos, D. M.; Arrigoni-Blank, M. F.; Oliveira, E. M. C.; Sampaio, T. S.; Farias, A. P.; Coelho, C. R.; Blank, A. F. Bioactivity of essential oil from Lippia gracilis Schauer against two major coconut pest mites and toxicity to a non-target predator. Crop protection. 2019 125,104913. [CrossRef]
- Blank, A. F.; de Fátima Arrigoni-Blank, M.; Bacci, L.; Costa Junior, L. M.; Castro Nizio, D. A. Chemical diversity and insecticidal and anti-tick properties of essential oils of plants from northeast Brazil. In: Essential Oil Research: Trends in Biosynthesis, Analytics, Industrial Applications and Biotechnological Production. 2019, 235-258. [CrossRef]
- Coolen, H.K.A.C, Meeuwis, J.A.M.; Vanleeuwen, P.W.N.M, Nolte, R.J.M. Substrate selective catalysis by rhodium metal-lohosts. J Am Chem Soc. 1995, 117, 11906–11913. [CrossRef]
- Ben Arfa, A.; Combes, S.; Preziosi-Belloy, L.; Gontard, N.; Chalier. Antimicrobial activity of carvacrol related to its chemical structure. Lett Appl Microbiol. 2006, 43, 149–154. [CrossRef]
- Dolly, B. B.; Barba, F. Cathodic reduction of hydroxycarbonyl compound trichloroacetyl esters. Tetrahedron. 2003, 59, 9161–9165. [CrossRef]
- Morais, S.M.; Vila-Nova, N.S, Bevilaqua, C.M.L, Rondon, F.C, Lobo, C.H, Moura, A.D.A.N, Sales, A.D.; Rodrigues, A.P.R.; Figuereido, J.R.; Campello, C.C.; Wilson, M.E.; Andrade, H.F. Thymol and eugenol derivatives as potential antileishmanial agents. Bioorg Med Chem. 2014, 22, 21,6250–6255. [CrossRef]
- Santos, C.P.; Pinto, J.A.O.; dos Santos, C.A.; Cruz, E.M.O.; Arrigoni-Blank, M.F.; Andrade, T.M.; Santos, D.A.; Alves, P.B.; Blank, A.F. Harvest time and geographical origin affect the essential oil of Lippia gracilis Schauer. Industrial Crops and Products. 2016, 79, 205-210. [CrossRef]
- SAS/STAT User’s Guide. Cary, NC, USA. 2008.
- Santos, N.C.; Silva, J.E.D, Santos, A.C.C, Dantas, J.D.O, Tavares, S.R.S.A, Andrade, V.S.; Oliveira, S.D.S.; Blank, A.F.; Araújo, A.P.A.; Bacci, L. Bioactivity of essential oils from Croton grewioides and its major compounds: toxicity to soybean looper Chrysodeixis includens and selectivity to the predatory stink bug Podisus nigrispinus. Environmental Science and Pollution Research. 2022, 30, 1-12. [CrossRef]
- Albuquerque, E.L.; Lima, J.K.; Souza, F.H.; Silva, I.M.; Santos, A.A.; Araújo, A.P.A.; Blank, A.F.; Lima, R.F.; Alves, P.B.; Bacci, L. Insecticidal and repellence activity of the essential oil of Pogostemon cablin against urban ants species. Acta tropica. 2013. 127, 181-186. [CrossRef]
- Tong, F.; Coats, J.R. Effects of monoterpenoid insecticides on [3H]-TBOB binding in house fly GABA receptor and 36Cl – uptake in American cockroach ventral nerve cord. Pestic. Biochem. Physiol. 2010, 98, 317-324. [CrossRef]
- Oladipupo, S.O.; Hu, X.P.; Appel, A.G. Essential oils in urban insect management-A review. Journal of Economic Entomology. 2022, 115, 1375-1408. [CrossRef]
- Gaire, S.; Scharf, M.E.; Gondhalekar, A.D. Synergistic toxicity interactions between plant essential oil components against the common bed bug (Cimex lectularius L.). Insects. 2020, 11, 133. [CrossRef]
- Xie, Y.; Huang, Q.; Rao, Y.; Hong, L.; Zhang, D. Efficacy of Origanum vulgare essential oil and carvacrol against the housefly, Musca domestica L (Diptera: Muscidae). Environmental Science and Pollution Research. 2019, 26, 23824-23831. [CrossRef]
- Rodríguez, A.; Beato, M.; Usseglio, V.L.; Camina, J.; Zygadlo, J.A.; Dambolena, J.S.; Zunino, M.P. Phenolic compounds as controllers of Sitophilus zeamais: A look at the structure-activity relationship. Journal of Stored Products Research. 2022, 99, 102038. [CrossRef]
- Gong, X.; Ren, Y. Larvicidal and ovicidal activity of carvacrol, p-cymene, and γ-terpinene from Origanum vulgare essential oil against the cotton bollworm, Helicoverpa armigera (Hübner). Environmental Science and Pollution Research. 2020, 27, 18708-18716. [CrossRef]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pesticide Biochemistry and Physiology. 2012, 102, 124-128. [CrossRef]
- Lu, X.; Weng, H.; Li, C.; He, J.; Zhang, X.; Ma, Z. Efficacy of essential oil from Mosla chinensis Maxim. cv. Jiangxiangru and its three main components against insect pests. Industrial Crops and Products. 2020, 147, 112237. [CrossRef]
- Staudt, A.; Brack, Y.; Itabaiana Jr, I.; Leal, I.C.R. 2022. Biocatalytic synthesis of monoterpene esters–A review study on the phylogenetic evolution of biocatalysts. Molecular Catalysis. 2022, 528, 112464. [CrossRef]
- Natal, C.M.; Fernandes, M.J.G.; Pinto, N.F.; Pereira, R.B.; Vieira, T.F.; Rodrigues, A.R.O.; Pereira, D.M.; Souza, S.F.; Fortes, A.G.; Castanheira, E.M.S.; Gonçalves, M.S.T. New carvacrol and thymol derivatives as potential insecticides: Synthesis, biological activity, computational studies and nanoencapsulation. RSC advances. 2021, 11, 54, 34024-34035. [CrossRef]
- Kafle, L.; Shih, C.J. Toxicity and repellency of compounds from clove (Syzygium aromaticum) to red imported fire ants Solenopsis invicta (Hymenoptera: Formicidae). Journal of Economic Entomology. 2013, 106, 131-135. [CrossRef]
- Chen, J.; Cantrell, C.L.; Duke, S.O.; Allen, M.L. Repellency of callicarpenal and intermedeol against workers of imported fire ants (Hymenoptera: Formicidae). Journal of economic entomology. 2008, 101, 265-271. [CrossRef]
- Buczkowski, G. Trap–treat–release: horizontal transfer of fipronil in field colonies of black carpenter ants, Camponotus pennsylvanicus. Pest management Science. 2019, 75, 2195-2201. [CrossRef]
- Qi, Y.X.; Zeng, T.; Wang, L.; Lu, Y.Y. Biogenic amine signaling systems in the red imported fire ant, Solenopsis invicta–possible contributors to worker division of labor General and Comparative Endocrinology. 2018, 262, 59-70. [CrossRef]
- Adams, C.; Schenker, J.; Weston, P., Gut, L.; Miller, J. Path meander of male codling moths (Cydia pomonella) foraging for sex pheromone plumes: Field validation of a novel method for quantifying path meander of random movers developed using computer simulations. Insects. 2020, 11, 549. [CrossRef]
- Oliveira, B.M.; Melo, C.R.; Santos, A.C.; Nascimento, L.F.; Nízio, D.A.; Cristaldo, P.F.; Blank, A.F.; Bacci, L. Essential oils from Varronia curassavica (Cordiaceae) accessions and their compounds (E)-caryophyllene and α-humulene as an alternative to control Dorymyrmex thoracius (Formicidae: Dolichoderinae). Environmental Science and Pollution Research. 2019, 26, 6602-6612. [CrossRef]
- Melo, C.R.; Blank, A.F.; Oliveira, B.M.S.; Santos, A.C.C.; Cristaldo, P.F.; Araújo, A.P.A.; Bacci, L. Formicidal activity of essential oils of Myrcia lundiana chemotypes on Acromyrmex balzani. Crop Protection. 2021, 139, 105343. [CrossRef]
Figure 1.
Synthetic derivatives of carvacrol [31].
Figure 1.
Synthetic derivatives of carvacrol [31].
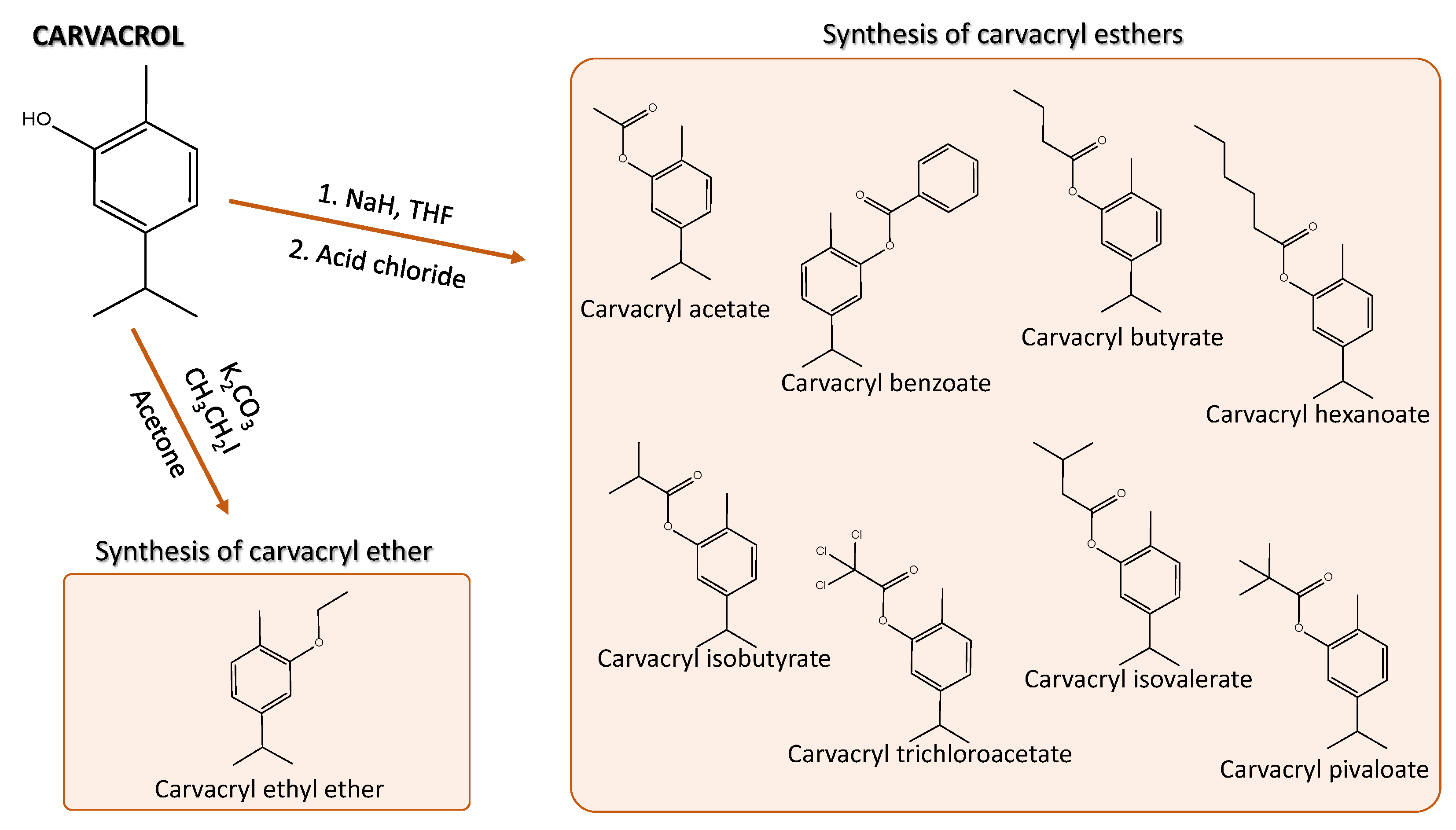
Figure 2.
Survival curves and lethal time (LT50) of Solenopsis sp. exposed to Lippia gracilis essential oil (green), carvacrol (yellow) (main oil compound) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol). Survival curves followed by the same letter do not differ significantly by the Holm-Sidak method (P > 0.05). Squares indicate the mean (± confidence interval to 95%) of the LT50 (lethal time required to kill 50% of the population).
Figure 2.
Survival curves and lethal time (LT50) of Solenopsis sp. exposed to Lippia gracilis essential oil (green), carvacrol (yellow) (main oil compound) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol). Survival curves followed by the same letter do not differ significantly by the Holm-Sidak method (P > 0.05). Squares indicate the mean (± confidence interval to 95%) of the LT50 (lethal time required to kill 50% of the population).
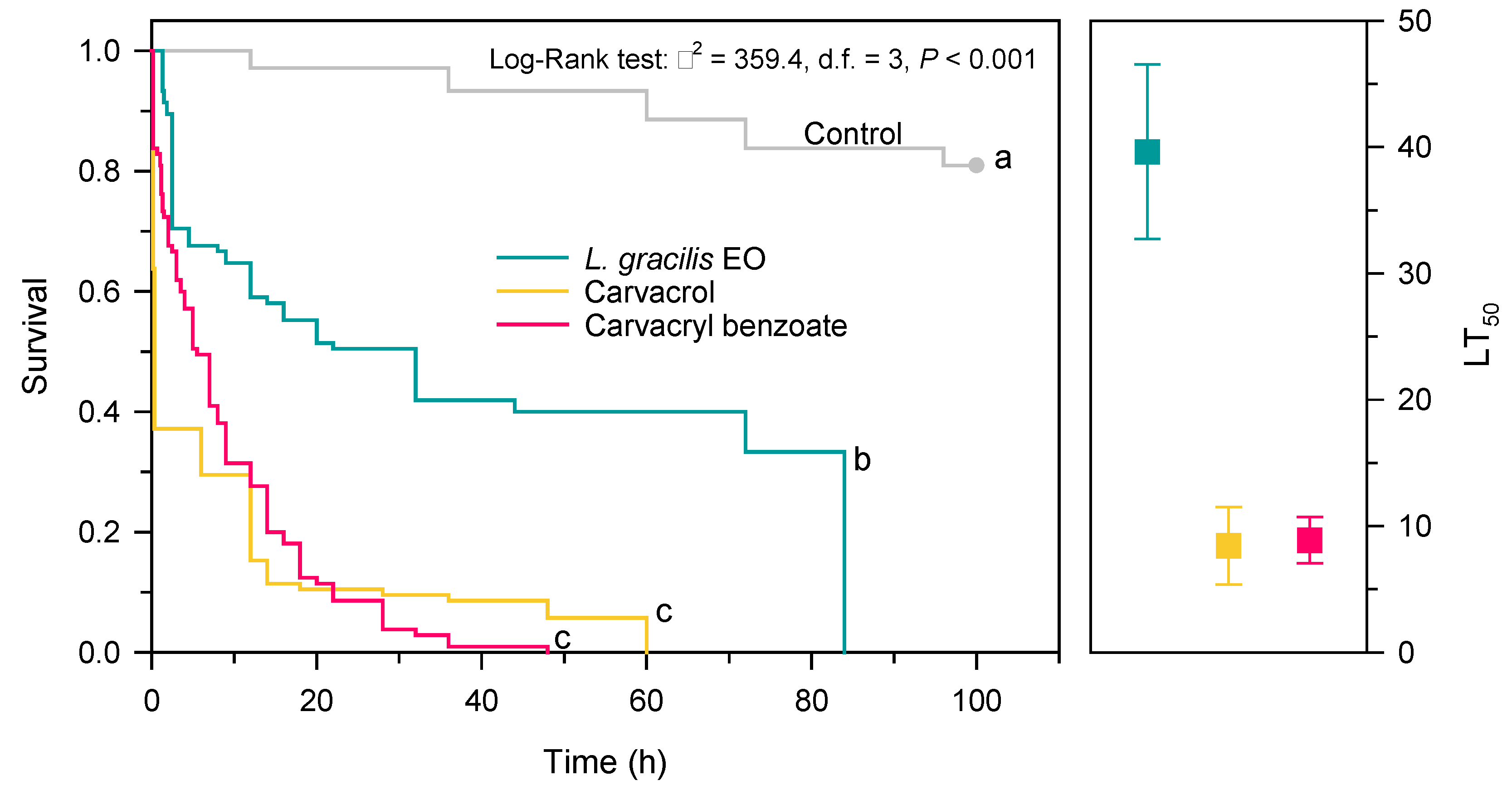
Figure 3.
Individual cleaning (A) and collective cleaning and aggregation behavior (B) of Solenopsis sp. exposed to the LD30s of the essential oil of Lippia gracilis (green), carvacrol (yellow) (major compound in the oil) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol). Means (± standard error) of the histograms followed by an asterisk differ from the control by Dunnett’s test (P < 0.05).
Figure 3.
Individual cleaning (A) and collective cleaning and aggregation behavior (B) of Solenopsis sp. exposed to the LD30s of the essential oil of Lippia gracilis (green), carvacrol (yellow) (major compound in the oil) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol). Means (± standard error) of the histograms followed by an asterisk differ from the control by Dunnett’s test (P < 0.05).
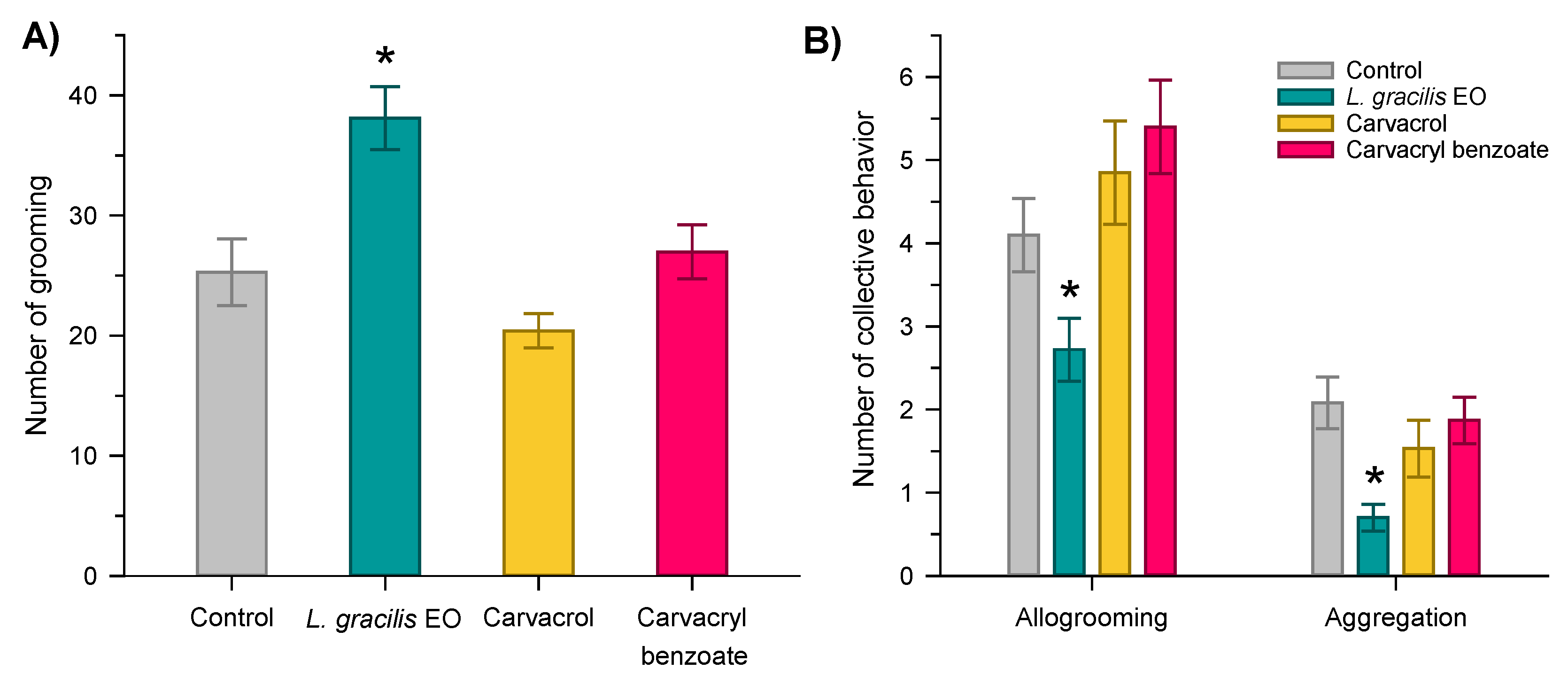
Figure 4.
Distance covered (mm) (A) and walking speed (mm/s) (B) of individual workers of the genus Solenopsis sp. exposed to the LD30s of Lippia gracilis essential oil (green), carvacrol (yellow) (major compound in the oil) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol) for 600 s in 6 x 6 cm arenas. Histograms with means (± standard error) followed by an asterisk differ from the control by Dunnett’s test (P < 0.05). Circles and the shaded area in the Radar plot represent the mean and standard error, respectively.
Figure 4.
Distance covered (mm) (A) and walking speed (mm/s) (B) of individual workers of the genus Solenopsis sp. exposed to the LD30s of Lippia gracilis essential oil (green), carvacrol (yellow) (major compound in the oil) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol) for 600 s in 6 x 6 cm arenas. Histograms with means (± standard error) followed by an asterisk differ from the control by Dunnett’s test (P < 0.05). Circles and the shaded area in the Radar plot represent the mean and standard error, respectively.
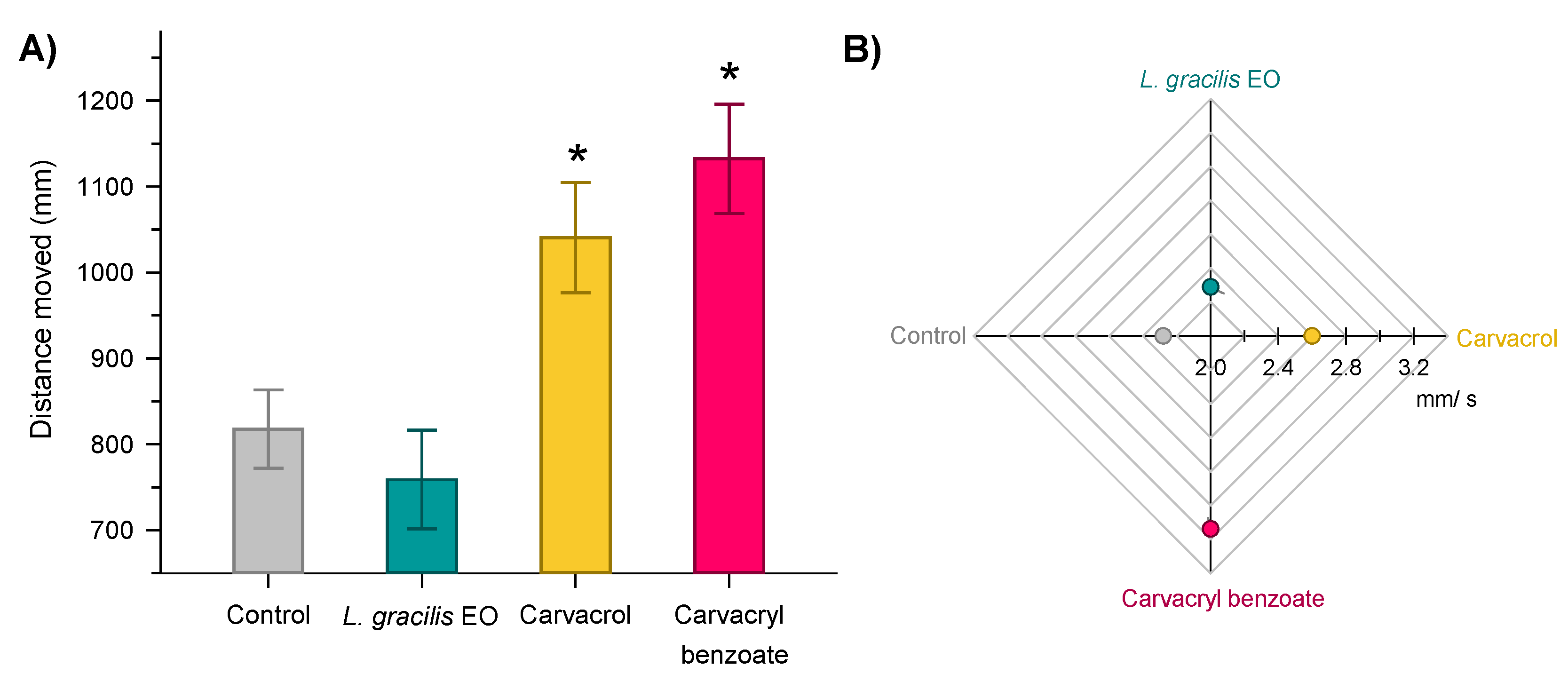
Figure 5.
Meander behavior (o/ mm), angular velocity (o/ s) and 2D and 3D walking paths of workers of the genus Solenopsis sp. exposed to LDs30 of Lippia gracilis essential oil (green), carvacrol (yellow) (major compound in the oil) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol) for 600 s in 6 x 6 cm arenas. Histograms with means (± standard error) followed by an asterisk, differ from the control by Dunnett’s test (P < 0.05).
Figure 5.
Meander behavior (o/ mm), angular velocity (o/ s) and 2D and 3D walking paths of workers of the genus Solenopsis sp. exposed to LDs30 of Lippia gracilis essential oil (green), carvacrol (yellow) (major compound in the oil) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol) for 600 s in 6 x 6 cm arenas. Histograms with means (± standard error) followed by an asterisk, differ from the control by Dunnett’s test (P < 0.05).
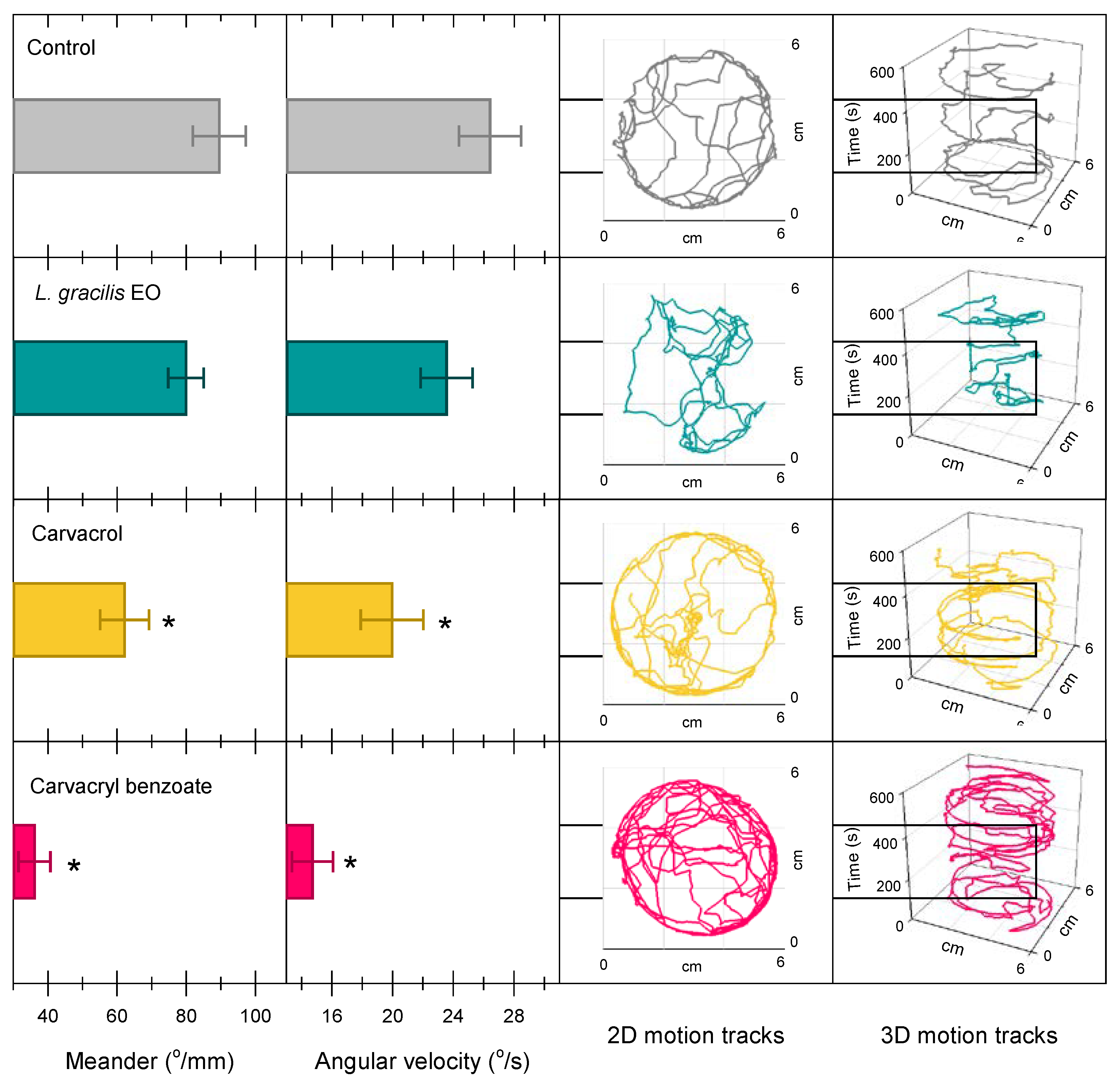
Figure 6.
Survival curves and lethal time (LT50) of the necrophoresis behavior of Solenopsis sp. exposed to corpses of ants subjected to LDs90 of Lippia gracilis essential oil (green), carvacrol (yellow) (major compound of the oil) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol) in two proportions [cadaver: live workers]. [1:20] = 1 corpse: 20 live workers and [1: 5] = 1 corpse: 5 live workers. Survival curves followed by the same letter do not differ significantly by the Holm-Sidak method (P > 0.05). Squares indicate the mean (confidence interval ± 95%) of the LT50 (lethal time required to kill 50% of the population).
Figure 6.
Survival curves and lethal time (LT50) of the necrophoresis behavior of Solenopsis sp. exposed to corpses of ants subjected to LDs90 of Lippia gracilis essential oil (green), carvacrol (yellow) (major compound of the oil) and carvacryl benzoate (red) (synthetic derivative of the more toxic carvacrol) in two proportions [cadaver: live workers]. [1:20] = 1 corpse: 20 live workers and [1: 5] = 1 corpse: 5 live workers. Survival curves followed by the same letter do not differ significantly by the Holm-Sidak method (P > 0.05). Squares indicate the mean (confidence interval ± 95%) of the LT50 (lethal time required to kill 50% of the population).
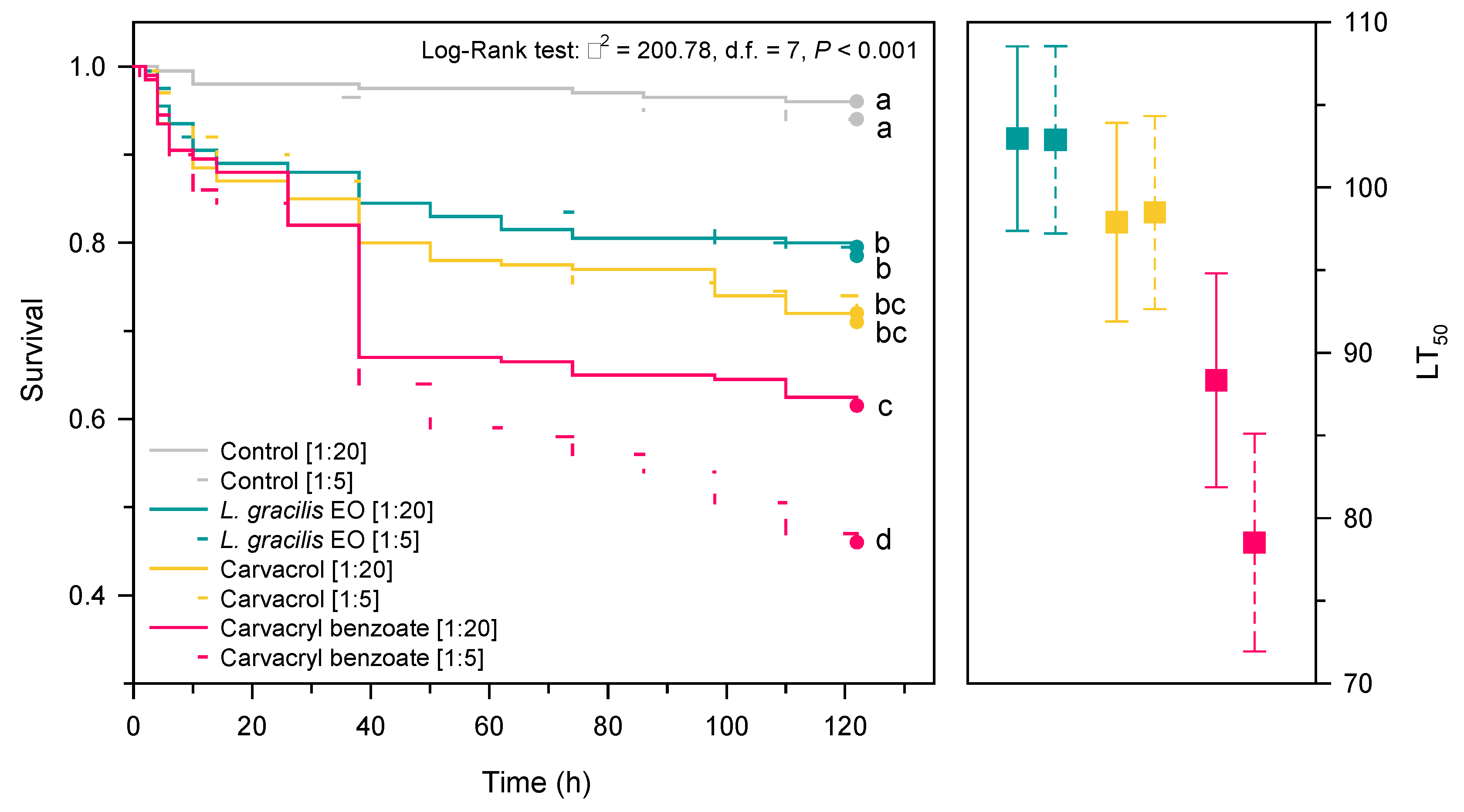

Table 1.
Acute toxicity after 48 h of topical exposure to the essential oil of Lippia gracilis, its major compound carvacrol and nine synthetic derivatives of carvacrol on workers of the ant Solenopsis sp.
Table 1.
Acute toxicity after 48 h of topical exposure to the essential oil of Lippia gracilis, its major compound carvacrol and nine synthetic derivatives of carvacrol on workers of the ant Solenopsis sp.
| Compounds | n1 | LD30 (CI 95%)2 |
LD50 (CI 95%)2 |
LD90 (CI 95%)2 |
Slope ± SE |
χ2 | P |
|---|---|---|---|---|---|---|---|
| -------------------- μg/ mg -------------------- | |||||||
| Essential oil of Lippia gracilis | 336 | 3.42 (2.66-4.14) |
6.11 (5.16-7.20) |
25.2 (19.0-38.3) |
2.08 ±0.23 |
7.41 | 0.12 |
| Carvacrol | 336 | 2.76 (1.81-3.75) |
6.79 (5.19-8.71) |
60.9 (39.2-118) |
1.35 ±0.16 |
0.76 | 0.94 |
| Carvacryl acetate | 392 | 4.09 (3.17-4.94) |
6.92 (5.86-7.97) |
24.9 (20.2-33.5) |
2.30 ±0.24 |
6.39 | 0.27 |
| Carvacryl benzoate | 448 | 2.21 (1.92-2.49) |
3.20 (2.86-3.57) |
7.97 (6.90-9.53) |
3.24 ±0.24 |
4.81 | 0.57 |
| Carvacryl butyrate | 392 | 3.73 (2.67-4.75) |
7.50 (6.09-8.99) |
41.2 (30.5-64.0) |
1.73 ±0.19 |
8.62 | 0.13 |
| Carvacryl ethyl ether | 448 | 3.81 (3.12-4.51) |
6.87 (5.88-8.04) |
29.1 (22.7-40.3) |
2.05 ±0.17 |
7.96 | 0.24 |
| Carvacryl hexanoate | 336 | 6.25 (5.02-7.39) |
10.5 (8.99-12.2) |
37.1 (28.9-53.3) |
2.33 ±0.25 |
6.46 | 0.17 |
| Carvacryl isobutyrate | 336 | 3.46 (2.45-4.40) |
6.54 (5.28-7.84) |
30.9 (23.3-47.4) |
1.90 ±0.22 |
3.54 | 0.47 |
| Carvacryl isovalerate | 392 | 6.16 (3.79-8.18) |
10.8 (8.10-14.5) |
42.3 (26.8-112) |
2.16 ±0.33 |
9.92 | 0.08 |
| Carvacryl pivaloate | 392 | 3.05 (2.41-3.70) |
5.85 (4.88-7.03) |
28.7 (21.4-43.0) |
1.85 ±0.17 |
6.80 | 0.24 |
| Carvacryl trichloroacetate | 336 | 4.05 (3.00-5.12) |
7.92 (6.39-9.66) |
40.7 (30.4-60.7) |
1.80 ±0.18 |
5.99 | 0.20 |
1number of insects tested. 2 lethal doses required to kill 30, 50 and 90% of populations and their respective confidence intervals to 95%.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



