
Submitted:
29 April 2023
Posted:
30 April 2023
You are already at the latest version
Abstract
In the present paper, we describe several coleoid jaws discovered in the lower Toarcian black shales, cropping out along the Vilyui River (Yakutia, Russia). This is the first record of a Lower Jurassic coleoid jaw outside Europe and the first report of such a finding from the Mesozoic of Siberia. The described coleoid jaws demonstrate the same mode of preservation and morphology as the coeval jaws previously reported from Europe. Their preservation in Siberia became possible due to the widespread occurrence of black shale facies associated with the early Toarcian oceanic anoxic event (TOAE).
Keywords:
Yakutia
; Mesozoic
; palaeobiology
; cephalopoda
1. Introduction
Despite the worldwide distribution of Toarcian black shale facies, suitable for preserving non-mineralized coleoid remains, such as their beaks, very few such findings were reported so far. For the first time, belemnite jaws from the lower Toarcian Posidonienschiefer Formation of Germany were reported and figured by Riegraf and Hauff [1] and Reitner and Urlichs [2], and additional specimens were later provided by Schlegelmilch [3]. Recently, jaws of a putative diplobelid Clarkeiteutis of the Toarcian age were figured by Jenny et al. [4]. However, all these findings were poorly preserved, and details of their shapes are unclear. Better preserved but also not clearly visible beaks are known for the octobrachian coleoids – vampyropods Parabelopeltis and Jeletzkyteuthis [5]. From the territory of Russia, the Toarcian coleoid jaws have never been described or depicted to date, although both in Siberia and in the northeastern Russia, the marine Toarсian is the most extensively studied and geographically expanding wider than any other Lower Jurassic stage [6]. In the present paper, we provide the first report of coleoid jaws from the black shale strata, corresponding to the Toarcian oceanic anoxic event (TOAE), and discuss their possible affinity.
2. Geological setting
The upper Pliensbachian and lower Toarcian deposits of the Vilyui River basin (Yakutia, Russia) crop out along the Vilyui River banks in its middle and upper reaches and are also known from Vilyui tributaries (Tyung, Markha, and Sinyaya rivers) [7,8,9].
The Upper Pliensbachian Tyung Formation consists of silts, sands, and sandstones with a total thickness of ca. 40 m and is subdivided into 3 members. Ammonite occurrences are uncommon, and only the lower member of the Tyung Formation is characterized by records of Amaltheus [7]. Nevertheless, detailed subdivision of this formation can be carried out using bivalve assemblages [9], as they are the most common and diverse macrofossils met across the formation. Bladed glendonites showing gradual size increase upwards from ~0,5 to 30 cm in length are typical for the middle and upper parts of the Tyung Formation throughout the Vilyui River basin [10] and were previously used for geological mapping purposes [8]. There are records of belemnites from the lower part of the formation [11], and from its top ([12]; as ‘Sardanga Formation’); however, other authors who visited the same sections reported the absence of belemnites in the same strata (Nalnjaeva in [13], p. 142). Currently, the consensus is that the belemnites migrated to the Northern Siberian seas in the Early Toarcian, quickly forming endemic lineages and reaching the maximum diversity up to the late Toarcian - early Aalenian time [14,15].
The overlying lower Toarcian Suntar Formation (up to 60 m, subdivided into two members) is separated by a remarkable discontinuity but seemingly without any biostratigraphic gap. The lower Toarcian strata are represented chiefly by mudstones (in the lowermost part, mainly thin-laminated clay intercalated with unfrequent silty surfaces, or ‘black shales’) with numerous carbonate concretions. The lower 10-20 m of the succession contain rare bivalve findings. Until now, only a single occurrence of the ammonite was reported from the lowermost part of the Suntar Formation ([7], Figure 3): this specimen, identified as Pliensbachian Paltarpites argutus in the original publication, was later re-identified as Toarcian Harpoceras by M.S. Mesezhnikov (see [16]). Also, the ‘black shale’ strata at the base of the Suntar Fm contain numerous poorly preserved belemnite rostra ([8]; as ‘fourth member of the Pliensbachian’). Relatively common ammonites (Dactylioceras of D. commune group, Osperleioceras, and Zugodactylites) and very numerous bivalves (mainly Dacryomya, Tancredia, Lenoceramus) and belemnites (Catateuthis, Acrocoelites, ‘Nannobelus,’ Clastoteuthis, Brachybelus, Odontobelus) occur in the upper part of the Suntar formation [16,17,18], with bivalves and belemnites often forming allochthonous accumulations. Besides mollusks, the Pliensbachian-Toarcian boundary beds of the Vilyui River basin are known for their ichthyosaurian and plesiosaurian occurrences [19].
As can be seen from this short overview, the Pliensbachian-lower Toarcian sequence at Vilyui River is poorly characterized by index macrofossils, and thus the stage boundary is primarily defined by using bivalves. Conventionally the Pliensbachian-Toarican boundary in Siberia is aligned with the base of the organic-rich shale interval, widely spread in the region and known as ‘Kiterbyut horizon’ [15]; in the Vilyui River sections, it corresponds to the boundary between Tyung and Suntar Formations. However, as the black shale event in different parts of the world starts slightly above the base of the Toarcian [20,21] and there is no ground to assume its earlier appearance in Siberia, it can not be excluded that the real chronostratigraphic boundary between the Pliensbachian and the Toarcian falls into the upper part of the Tyung Formation, as was proposed by Repin [22].
2. Material
During the 2021 fieldwork on the Pliensbachian-Toarchian sequence of the Vilyui River (Yakutia; Figure 1), three slabs with coleoid jaws were collected from large carbonate concretions weathered out from the lower Toarcian black shales and lying loose on the beach at the exposure 6b. The base of the concretions, having a width of up to 1-1.5 m and height of 20 cm, is located at 1,2 m above the Pliensbachian/Toarcian boundary and can be traced throughout all the studied outcrops near the Sardanga settlement (sections 4a-d, 6b and 7 in Figure 2). The black shales unit is barren of macrofossils, except for belemnite rostra, frequently occurring in the lower 50 cm of the unit and scarcely above. They are poorly preserved and undeterminable, however, having elongated shapes typical for the genera Passaloteuthis and Acrocoelites.
Carbonate concretions, which occur between 0,5 and 1,2 m above the base of black shales (Figure 3), yielded infrequent bivalves (mainly Kedonella and Tancredia) and common fish scales. Smaller concretions from the level 1 m above the Pliensbachian/Toarcian boundary in exposure 4d were found to contain numerous onychites. Although the lowermost part of black shales throughout the Vilyui River basin lacks ammonites, the age of the studied unit can be deducted through bivalve occurrences, as Kedonella mytileformis collected from the jaw-bearing level is restricted to two lowermost lower Toarcian ammonite zones [23].
Our observations on the belemnite distribution across the succession confirm Repin’s statement [22] on the first occurrence of belemnites within the ‘Pliensbachian.’ Near the top of the Tyung Formation, there is a level with poorly preserved belemnite rostra of the genus ?Passaloteuthis accumulated at the intraformational discontinuity surface within the sandstone. This level is the earliest appearance of belemnites within the studied succession, which may reflect the early episode of mass migration of belemnites into the epicontinental seas of northeastern Eurasia in the late Pliensbachian-early Toarcian time.
Figure 3.
Field photos of the Pliensbachian-Toarcian boundary in section 4b. A – general view on the boundary beds; large carbonate concretions 1,2 m above the boundary are well-visible; B – level of first appearance of belemnites within the succession (topmost part of the Pliensbachian) is visible right from hammer.
Figure 3.
Field photos of the Pliensbachian-Toarcian boundary in section 4b. A – general view on the boundary beds; large carbonate concretions 1,2 m above the boundary are well-visible; B – level of first appearance of belemnites within the succession (topmost part of the Pliensbachian) is visible right from hammer.

3. Description of coleoid jaws
All three specimens are preserved as a flattened imprint on the bedding surface and its counterpart.
Specimen No. 1 (Figure 4.1) is an upper jaw having a length of 14 mm and a height of 9 mm. A well-preserved external plate of the jaw (hood) demonstrates a prominent anterior tip (rostrum) and both lateral wings behind it. The inner plate is partly visible along the lower-rear part of the wings, but its rear part is not preserved. The rostrum is robust and pointed, achieving almost 5 mm in length. Its inner margin is relatively straight and the angle between the rostrum and wings (jaw angle) is acute, circa 65°.
Specimen No. 2 (Figure 4.2) is an upper jaw having a length of 12 mm and a height of 7 mm. As in specimen No. 1, the outer plate of the jaw has a prominent rostrum, and one lateral wing is well-preserved on the bedding surface. The inner plate of the jaw is visible on the back of the wing and behind the hood, but its rear part is not preserved. In contrast to the first specimen, the inner edge of the rostrum looks not straight but slightly concave, which makes the rostrum slightly curved. The angle between the rostrum and wing is relatively straight and again close to 65°. Both upper jaws are slightly different from each other, however it is not clear whether they belonged to various coleoid species or whether the differences are due to slightly various positions of the specimens in the rock.
Specimen No. 3 (Figure 4.2) is small (5x4 mm in size) and has a sub-square overall shape. It shows a curved outer plate and a narrower inner plate translucent throughout it. The edges of the specimen appear to have been incompletely preserved, making it difficult to understand the original shape. Tentatively we interpret it as an incomplete lower jaw – however, insufficient preservation leaves room for doubt in this case.
4. Discussion
4.1. Early Toarcian palaeoenvironments and taphonomy
Like in other regions with OAE-related facies recorded, in the Vilyui River basin the lowermost Toarcian strata show evidence of deposition under oxygen-depleted environments. These are lack of bioturbation and fine parallel lamination, clearly visible both in shales and concretions (Figure 5), as well as taphonomical characters of macrofossils and general appearance of the assemblage. Bivalve specimens here are rare and mainly belong to the genus Kedonella, which had a pseudoplanctonic lifestyle, attaching to floating wood, ammonites, and other objects ([24,25]; in these papers, Kedonella was classified as Pseudomytiloides). However, the presence of Tancredia, an infaunal suspension feeder which needed well-oxygenated seawater [26] but which is tolerant to low salinity environments, suggests that near-bottom water at least sometimes exhibited normal oxygen contents. Belemnites, although numerous in the topmost part of the Tyung Formation and near the base of the Suntar Formation, become rarer and rarer upwards the black shale interval, completely disappearing above it. This may indicate the transformation of oxygen-depleted marine environments, still providing suitable habitats for nektonic organisms in the upper part of the water column into low-salinity environments and facies upward the succession.
Finally, it should be noted that in contrast to other well-studied TOAE-related sections worldwide and in other parts of Siberia, the Viluyi succession shows numerous indicators of shallow depths, the proximity of land, and reduced salinities in most intervals. This explains why ammonite occurrences are uncommon in the studied area (and why they are more numerous in the offshore parts of the Vilyui Sea basin, available for study in the sections along the Tyung and Markha rivers).
All the organic coleoid remains (beaks, hooks) from the ‘black shale interval’ of the Vilyui River sections were collected from the carbonate nodules. In contrast, careful excavation and examination of the bedding surfaces within the shales provided no additional findings. The belemnite rostra, sometimes met in the black shale interval, are always heavily weathered up to the unidentifiable state (making problematic even the generic attribution), being recrystallized into the whitish calcite. The observed preservation of belemnite rostra results from seasonal weathering in severe climates with heavy snow cover, melting in spring and leaking deeply into the black shales along the silt interbeds. As a result, most carbonate fossils as well as fragile organic fossils, were destroyed, making the ‘black shales’ at Vilyui useless for surface collecting. In turn, carbonate concretions served as natural sarcophaguses protecting embedded chitinous elements from weathering.
4.2. Comparison of described coleoid jaws with known Mesozoic records and their possible affinities
The jaws described herein were found isolated from cephalopod remains. Therefore, the main basis for understanding the affinity of these jaws is their shape and structure, as well as some general assumptions. Three large groups of cephalopods inhabited the Toarcian seas: Nautilida, Ammonoidea, and Coleoidea, all having well-developed jaw apparatuses. However, starting at least from the Middle Triassic, nautilids demonstrate calcitic elements (rhyncholites and conchorhynchs) in their jaws [27]. Such calcitic elements are absent in the studied specimens; thus nautilid affinity can be confidently excluded. The jaws also could not belong to ammonoids, since ammonoid jaws never had protruding wings on the sides of the external lamella [28]. Therefore, the hosts of jaws belonged to Coleoidea.
The overall shape of the jaws No.1 and No.2, and particularly the presence of a long, pointed rostrum, indicates that the jaws belonged to decabrachian coleoids, characterized by such prominent rostra. In contrast, the tips of the octobrachian jaws are usually shorter and more rounded. Among the decabrachian jaws published to date, the new findings are very similar to isolated upper jaws from the Bathonian and Callovian of European Russia, for which belemnite affinity was tentatively proposed [32,33]. Our jaws are also similar to an upper jaw from the Callovian of Poland ([34], Figure 1C), which also was interpreted as probably belonging to a belemnitid coleoid. However, they differ from the stratigraphically close belemnitid upper jaws from the Aalenian of Switzerland, and from the specimens from the Kimmeridgian of Germany, which have very thin and strongly curved rostra [35,36]. It should be additionally highlighted that the straight and robust rostra of the Toarcian jaws are similar to the giant coleoid jaws of presumed teuthid affinity, described from the Turonian (Upper Cretaceous) of Japan ([37], Figure 1), from where true belemnites are not known [38]. However, it is possible that the Early Jurassic and Late Cretaceous jaws could belong to different decabrachian taxa, which had similar feeding strategies and thus their similarity in convergent. It can be assumed that more robust and straight beaks could be used to hunt more sluggish but better-protected prey than thin and curved varieties.
Among Toarcian decabrachian coleoids, several taxonomic groups can be treated as the hypothetical hosts of the beaks. Belemnites are the most attractive candidates to be considered as beak hosts, as they are the only coleoid group clearly present within the black shale interval. However, considering the huge preservation potential of their massive calcitic rostra, it should be noted that only a single belemnite rostrum was found within the same concretionary level as beaks, while beak findings were numerous.
The lower Toarcian interval is also well-known for highly diversified and abundant gladius-bearing coleoids (e.g. [5,29,30]) as well as for peculiar phragmocone-bearing taxa such as Clarkeiteuthis [4,31], the position of which within the coleoid system is not completely clear. As can be judged from the abundance of such fossils in the Toarcian Lagerstättes in Europe and Canada [29,30], these coleoid groups had a sub-global distribution in the Toarcian. However, due to taphonomical constraints, they had almost no chance of being preserved as fossils in the Vilyui River sections.
5. Conclusions
For the first time, well-preserved jaws of decabrachian coleoids are reported from the Lower Jurassic (Toarcian) of Siberia. The preservational constraints of the Vilyui River sections, along with the scarce records of this type of fossils in literature, prevent us from their unequivocal attribution to any certain taxon.
Author Contributions
Conceptualization, M.R.; fieldwork, M.R., A.I., O.L.; methodology, M.R., A.I., A.M., O.L.; investigation, M.R., A.I., A.M.; writing—original draft preparation, M.R., A.I., A.M.; writing—review and editing, M.R., A.I., A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Russian Government Program FMMG-2021-0003 (Geological Institute of RAS, Moscow). Field works were funded by Russian Foundation for Basic Research grant number 21-55-15015.
Data Availability Statement
Studied specimens are deposited in the collection of Aprelevka branch of All-Russian Research Geological Petroleum Institute (Aprelevka, Moscow area, Russia).
Acknowledgments
Special thanks to the Sardanga and Suntar settlements administration for their help during the fieldwork and to Nikolay Zverkov for his assistance during the field works.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Riegraf, W.; Hauff, R. Belemnitenfunde mit Weichkörper, Fangarmen und Gladius aus dem Untertoarcium (Posidonienschiefer) und Unteraalenium (Opalinuston) Süd-Westdeutschlands. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 1983, 165, 466–483. [Google Scholar]
- Reitner, J.; Urlichs, M. Echte Weichteilbelemniten aus dem Untertoarcium (Posidonienschiefer) Südwestdeutschlands. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 1983, 165, 450–465. [Google Scholar]
- Schlegelmilch, R. Die Belemniten des Süddeutschen Jura. Gustav Fischer: Stuttgart, Jena, Lübeck, Ulm, Germany, 1998; 151 S. [CrossRef]
- Jenny, D.; Fuchs, D.; Arkhipkin, A.I.; Hauff, R.B.; Fritschi, B.; Klug, C. Predatory behaviour and taphonomy of a Jurassic belemnoid coleoid (Diplobelida, Cephalopoda). Scientific reports 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Klug, C.; Schweigert, G.; Fuchs, D.; De Baets, K. Distraction sinking and fossilized coleoid predatory behaviour from the German Early Jurassic. Swiss Journal of Palaeontology 2021, 140, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kalacheva, E.D. Toarcian. Geological Society of America Special Paper 1988, 223, 14–18. [Google Scholar] [CrossRef]
- Koshelkina, Z.V. New data on the stratigraphy of the Lower Jurassic marine deposits along the Vilyuy river. Izvestia of the Academy of Sciences of the USSR. Geologic Series 1961, 8, 78–86. [Google Scholar]
- Kirina, T.I. Stratigraphy of the Lower Jurassic deposits of the western part of Vilyui Syneclise. Transactions of VNIGRI 1966, 249, 18–71. (in Russian). [Google Scholar]
- Knjazev, V.G.; Devyatov, V.P.; Schurygin, B.N. Stratigraphy and paleogeography of the Early Jurassic of the east of the Siberian Platform. Yakut Scientific Center SB AS USSR: Yakutsk, Russia, 1991; 100 p. (in Russian).
- Rogov, M.; Ershova, V.; Gaina, C.; Vereshchagin, O.; Vasileva, K.; Mikhailova, K.; Krylov, A. Glendonites throughout the Phanerozoic. Earth-Science Reviews 2023, 241, 104430. [Google Scholar] [CrossRef]
- Koshelkina, Z.V. Stratrigraphy and bivalves of the Jurassic deposits of Vilyui Synclise and Priverchoyanye depression. Transactionsof the SKVNII 1963, 5, 1–219. (in Russian). [Google Scholar]
- Repin, Yu.S. Peculiarities of the structure of the Jurassic section in the middle reaches of the Vilyui River. In Upper Paleozoic and Mesozoic reference horizons of the northern European part of the USSR and Siberia (a collection of scientific papers). VNIGRI: Leningrad, Russia, 1983; pp. 64–74. (in Russian).
- Sachs, V.N.; Nalnjaeva, T.I. The Early and Middle Jurassic belemnites of the North of the USSR. Nannobelinae, Passaloteuthinae and Hastitidae. Nauka: Leningrad, USSR, 1970; 228 pp. (in Russian).
- Meledina, S.V.; Shurygin, B.N.; Dzyuba, O.S. Stages in development of mollusks, paleobiogeography of Boreal seas in the Early-Middle Jurassic and zonal scales of Siberia. Russian Geology and Geophysics 2005, 46, 239–255. [Google Scholar]
- Nikitenko, B.L.; Shurygin, B.N.; Knyazev, V.G.; Meledina, S.V.; Dzyuba, O.S.; Lebedeva, N.K.; Peshchevitskaya, E.B.; Glinskikh, L.A.; Goryacheva, A.A.; Khafaeva, S.N. Jurassic and Cretaceous stratigraphy of the Anabar area (Arctic Siberia, Laptev Sea coast) and the Boreal zonal standard. Russian Geology and Geophysics 2013, 54, 808–837. [Google Scholar] [CrossRef]
- Dagis, A.A. Toarcian ammonites (Dactylioceratidae) of the north of Siberia. Trans. Inst. Geol. Geophys. Sib. Branch Ac. Sci. USSR 1968, 40, 1–108. (in Russian). [Google Scholar]
- Krymgolts, G.Ya.; Tazikhin, N.N. Ammonites of Suntar and some questions of stratigraphy of Jurassic deposits of the Vilyui syneclise. Transactions of VNIGRI 1963, 220, 205–219. (in Russian). [Google Scholar]
- Nalnjaeva, T.I. Biostratigraphic subdivisions in the Lower and Middle Jurassic according to belemnites. Trans. Inst. Geol. Geophys. Sib. Branch Ac. Sci. USSR 1986, 648, 111–118. (in Russian). [Google Scholar]
- Zverkov, N.G.; Grigoriev, D.V.; Danilov, I.G. Early Jurassic palaeopolar marine reptiles of Siberia. Geological Magazine 2021, 158, 1305–1322. [Google Scholar] [CrossRef]
- Riegraf, W.; Werner, G.; Lorcher, F. Der Posidonienschiefer Biostratigraphie, Fauna und Fazies des südwestdeutschen Untertoarciums (Lias epsilon). Enke-Verlag: Stuttgart, Germany, 1984; pp. 1–195.
- Martindale, R.C.; Them, T.R.; Gill, B.C.; Marroquín, S.M.; Knoll, A.H. A new Early Jurassic (ca. 183 Ma) fossil Lagerstätte from Ya Ha Tinda, Alberta, Canada. Geology 2017, 45, 255–258. [Google Scholar] [CrossRef]
- Repin, Yu. S. The aspects of the geological structure of the Jurassic succession in the middle reaches of the Vilyui river. In Upper Paleozoic and Mesozoic reference horizons of the North of the European Part of the USSR and Siberia (collection of scientific papers). VNIGRI: Leningrad, USSR, 1983; pp. 64–74. (in Russian).
- Lutikov, O.A.; Arp, G. A Boreal Toarcian Biochronological Zonation Based on Bivalve Mollusks of the Genus Meleagrinella Whitfield, 1885. Stratigraphy and Geological Correlation 2023, 31, 49–70. [Google Scholar] [CrossRef]
- Schmid–Röhl, A.; Röhl, H.J. Overgrowth on ammonite conchs: environmental implications for the Lower Toarcian Posidonia Shale. Palaeontology 2003, 46, 339–352. [Google Scholar] [CrossRef]
- Röhl, H. J.; Schmid-Röhl, A. Lower Toarcian (Upper Liassic) black shales of the Central European epicontinental basin: a sequence stratigraphic case study from the SW German Posidonia Shale. SEPM Special Publication 2005, 82, 165–189. [Google Scholar] [CrossRef]
- Zakharov, V.A.; Shurygin. B.N. Biogeography, facies and stratigraphy of the Middle Jurassic of Soviet Arctic (by bivalve molluscs). Trans. Inst. Geol. Geophys. Sib. Branch Ac. Sci. USSR 1978, 352, 1–206. (in Russian). [Google Scholar]
- Klug, C. Functional morphology and taphonomy of nautiloid beaks from the Middle Triassic of southern Germany. Acta Palaeontologica Polonica 2001, 46, 43–68. [Google Scholar]
- Tanabe, K.; Kruta, I.; Landman, N.H.; Ammonoid buccal mass and jaw apparatus. In Ammonoid Paleobiology: From Anatomy to Ecology. Topics in Geobiology, 43. Klug, C., Korn, D., De Baets, K., Kruta, I., Mapes, R. H., Eds. Springer: Dordrecht, Germany, 2015; pp. 439–494. [CrossRef]
- Donovan, D.T.; Fuchs, D. Part M, Chapter 13: Fossilized soft tissues in Coleoidea. Treatise Online 2016, 73, 1–30. [Google Scholar] [CrossRef]
- Marroquín, S. M.; Martindale, R. C.; Fuchs, D. New records of the late Pliensbachian to early Toarcian (Early Jurassic) gladius-bearing coleoid cephalopods from the Ya Ha Tinda Lagerstätte, Canada. Papers in Palaeontology 2018, 4, 245–276. [Google Scholar] [CrossRef]
- Fuchs, D.; Donovan, D. T.; Keupp, H. Taxonomic revision of “Onychoteuthis” conocauda Quenstedt, 1849 (Cephalopoda: Coleoidea). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 2013, 270, 245–255. [Google Scholar] [CrossRef]
- Keupp, H.; Mitta, V.V. Cephalopod jaws from the Middle Jurassic of central Russia. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 2013, 270, 23–54. [Google Scholar] [CrossRef]
- Mironenko, A.A.; Mitta, V.V. On new records of cephalopod jaws from the Upper Bajocian (Middle Jurassic) of the Northern Caucasus (Russia, Karachay-Cherkessia). Paleontological Journal 2020, 54, 477–486. [Google Scholar] [CrossRef]
- Dzik, J. Uncalcified cephalopod jaws from the Middle Jurassic of Poland. Neues Jahrbuch für Geologie und Paläontologie Monatshefte 1986, 7, 405–417. [Google Scholar] [CrossRef]
- Klug, C.; Etter, W.; Hoffmann, R.; Fuchs, D.; De Baets, K. Jaws of a large belemnite and an ammonite from the Aalenian (Middle Jurassic) of Switzerland. Swiss Journal of Palaeontology 2020, 139, 1–11. [Google Scholar] [CrossRef]
- Klug, C.; Schweigert, G.; Fuchs, D.; Dietl, G. First record of a belemnite preserved with beaks, arms and ink sac from the Nusplingen Lithographic Limestone (Kimmeridgian, SW Germany). Lethaia 2010, 43, 445–456. [Google Scholar] [CrossRef]
- Tanabe, K.; Hikida, Y.; Iba, Y. Two coleoid jaws from the Upper Cretaceous of Hokkaido, Japan. Journal of Paleontology 2006, 80, 138–145. [Google Scholar] [CrossRef]
- Iba, Y.; Mutterlose, J.; Tanabe, K.; Sano, S.; Misaki, A.; Terabe, K. Belemnite extinction and the origin of modern cephalopods 35 m.y. prior to the Cretaceous−Paleogene event. Geology 2011, 39, 483–486. [Google Scholar] [CrossRef]
Figure 1.
Maps showing the location of studied Lower Jurassic outcrops. A – map of the region; B – map of the Vilyui river upper flows; C – satellite photo and the position of fossil localities near Sardanga settlement.
Figure 1.
Maps showing the location of studied Lower Jurassic outcrops. A – map of the region; B – map of the Vilyui river upper flows; C – satellite photo and the position of fossil localities near Sardanga settlement.
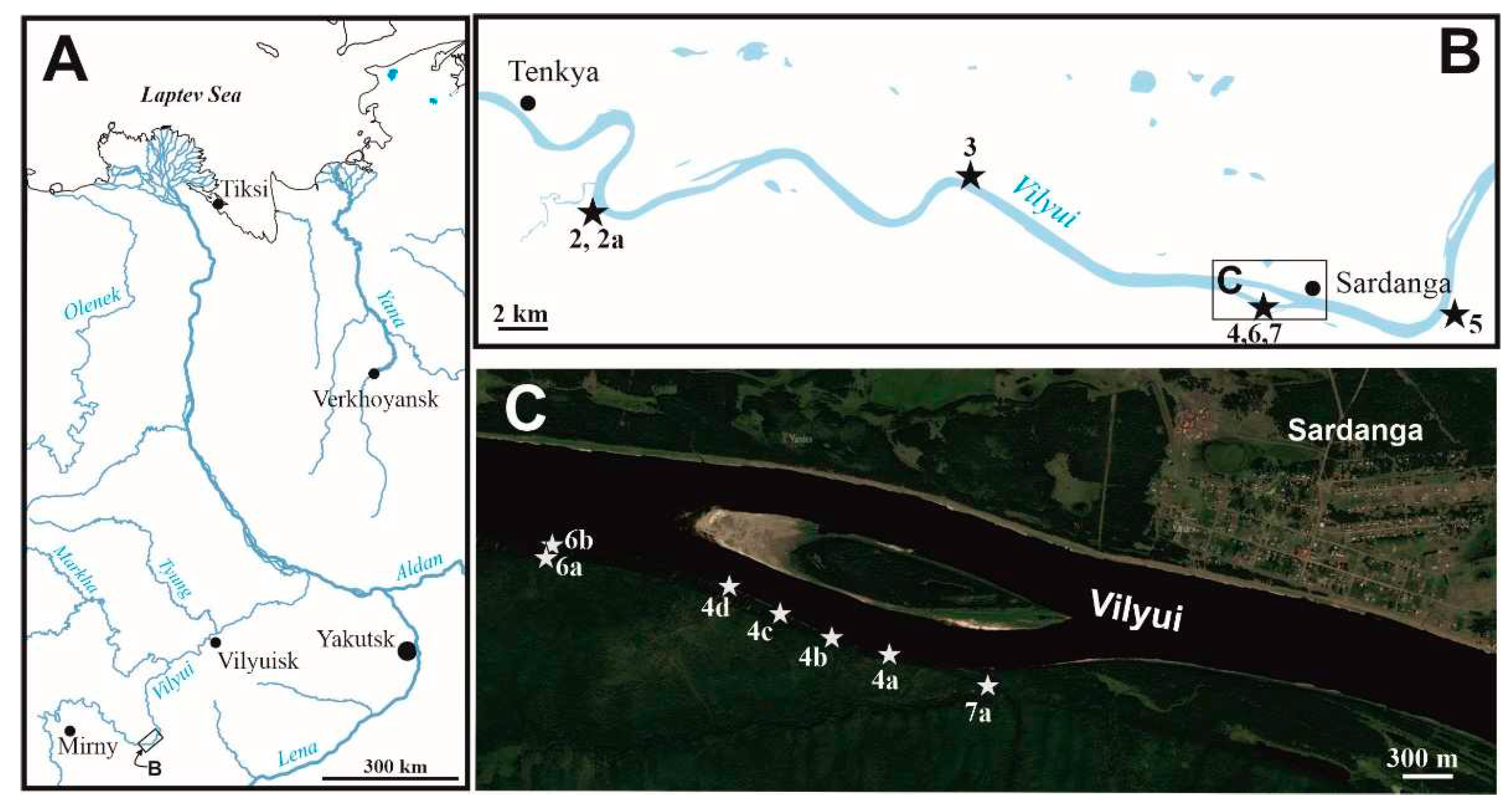
Figure 2.
Logs of studied sections and their biostratigraphic and lithostratigraphic subdivision.

Figure 4.
Coleoid jaws collected from bed 13 of section 6b. 1 A, B – upper jaw, specimen MAR 3/1a and its counterpart, MAR 3/1b; 2 A, B – upper jaw, specimen MAR 3/2a and its counterpart, MAR 3/2b; 3 A, B – lower? jaw, specimen MAR 3/3a and its counterpart, MAR 3/3b.
Figure 4.
Coleoid jaws collected from bed 13 of section 6b. 1 A, B – upper jaw, specimen MAR 3/1a and its counterpart, MAR 3/1b; 2 A, B – upper jaw, specimen MAR 3/2a and its counterpart, MAR 3/2b; 3 A, B – lower? jaw, specimen MAR 3/3a and its counterpart, MAR 3/3b.

Figure 5.
Thin section of the jaw-bearing concretion, showing clearly visible fine lamination.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



