
Submitted:
02 May 2023
Posted:
03 May 2023
You are already at the latest version
Abstract
The number of published studies on curcuminoids in cancer research, including its lead molecule curcumin and synthetic analogs, has been increasing substantially during the past two decades. Insights on the diversity of inhibitory effects they produced on a multitude of pathways involved in carcinogenesis and tumor progression were provided. As this wealth of data was obtained in a setting of various experimental and clinical data, this review first aimed at presenting a chronology of discoveries and an update on their complex in vivo effects. Secondly, there are many interesting questions linked to their pleiotropic effects. One of them, which is a growing research topic, relates to their ability to modulate metabolic reprogramming. This review will also cover the use of curcuminoids as chemosensitizing molecules, that can be combined with several anticancer drugs for reversing the phenomenon of multidrug resistance. Finally, current investigations in these three complementary research fields raise several important questions that will be put in the prospects for the future research related to the importance of these molecules in cancer research.
Keywords:
curcuminoids
; curcumin
; cancer
; signaling pathways
; metabolic reprogramming
; chemosensitization.
1. Introduction
Turmeric (Curcuma longa) was used for thousands of years in traditional Indian and Eastern Asian medicine. Its cultivation in the Middle East was documented since the 18th century BC in the gardens of Babylon, well before its transfer to Africa, mainly through Arabic influences [1]. Finally spread to Europe thanks to Marco Polo, and then to America and Oceania via the international spice trade, it is one of the best examples of how the exchange of knowledge between different parts of the world benefited humankind in improving the health of millions of people. Among natural anticancer products, it also represents a fascinating and continuous link between past, present, and future [1]. Its benefits as a food additive for the prevention of many diseases worldwide started to be documented through the report of the antibacterial action of its main component, curcumin, in 1949 [2]. However, a short report published in 1876 [3] demonstrated that a great part of empirical knowledge accumulated over centuries in Asia was not fully valorized between these two dates. Nevertheless, its potential anticancer activity was originally demonstrated against the development of tumors in animals [4]. The relief produced by a topical application of curcumin ointment in patients with external cancerous lesions was also revealed [5].
At the beginning of 2023, the number of publications related to the use of turmeric’s main phytochemical as anticancer agent reached 7,267 in the Pubmed database. Although curcumin represents “the leading” molecule, a significant part of these articles refers to “curcuminoids” [6]. The term also covers for synthetic chemical molecules, that are produced for optimizing pharmacological potency [7]. Synthetic chemistry based on curcumin represents an exciting field of research. It has been developing since 2008 [8], and continues to grow [9]. Ten years ago, the diversity of inhibitory effects produced by these molecules on a multitude of pathways involved in carcinogenesis and tumor progression started to be documented. This background was the milestone for interesting prospects in the search for alternative and less toxic therapies against the most common malignancy worldwide and the leading cause of cancer-related deaths, lung cancer [10]. However, at that date, most data were preclinical, including many in vitro studies. This weakness was overcome during the last decade, through insights provided by numerous in vivo and ex vivo investigations conducted on diverse experimental tumor models in laboratory rodents, or on clinical tumor tissue explants. Thus, this review will first aim at providing an update in this field. The potential of curcuminoids to reverse the deleterious consequences of the epithelial-to-mesenchymal transition (EMT) in invasive cancers was also recently reviewed [11]. However, among the many remaining unanswered questions regarding pleiotropic effects of curcuminoids, investigations focused on their suppressive effects on metabolic reprogramming first documented ten years ago [12] represents a current growing research topic [13].
To avoid the consequences of curcumin’s hydrophobicity and low bioavailability, considerable efforts have been made in the production of a great diversity of drug delivery systems, which were recently extensively reviewed [14,15,16,17]. To reverse multidrug resistance, curcuminoids canbe used in combination with many other drugs as chemosensitizer in cancer chemotherapy [18]. Thus, this crucial research field that continuously opens increasing potential will also be covered in this review. In this review, we have tried to profile available data very carefully, especially when interpreting data with respect to cell lines used, as there is an increasing problem with propagating and publishing results using the cell lines that are commonly contaminated with HeLa [19].
2. Evidence of multi-targeted action of curcuminoids against cancers invivo, historical overview and recent findings
The main signaling pathways / pharmacological actions affected by curcuminoids in cancer are summarized in Table 1.
2.1. Historical overview
Three successive research phases can be distinguished when studying thefindings reported in in vivo models, over the decades that followed the pioneering works of Kuttan et al [4,5].
2.1.1. Chemopreventive properties
At the beginning of the 1990s, the role of curcumin as a potent inhibitor of arachidonic acid-induced inflammation in vivo in mouse skin was revealed, with an associated suppression of chemical-induced tumor promotion [20]. In parallel, another research, that included rats fed by curcumin and then exposed to two different carcinogens administered intraperitoneally (i.p.) demonstrated its strong antimutagenic action in vivo [21]. Subsequently, studies on chemical-induced forestomach and skin tumors in mice reported that the anticarcinogenic activity of different natural curcuminoids could be explained in part by an alteration of the activation of carcinogen metabolism [22]. An important discovery was a significant decrease in the number of chemical-induced mammary tumors and in vivo formation of adducts in rats that were receiving curcumin intraperitoneally [23]. The chemopreventive properties of curcumin given orally during the initiation and/or post-initiation stage against azoxymethane-induced colon tumors in rats were also correlated to an increased rate of apoptosis in tumor cells (shrinkage, nuclear condensation, and presence of apoptotic bodies) [24]. The previously mentioned inhibition of arachidonic acid metabolism was then attributed to a stimulatory effect of curcuminon the expression of genes encoding three stress proteins, Hsp27, Hsp70 and αB crystallin [25]. Finally, the action of orally administered curcumin during the promotion/progression stage of colon carcinogenesis in rats was demonstrated, and the modulation of apoptosis associated with molecular action of curcumin was confirmed in colon tumors [26]. During that period, the apoptotic process in cancer cells, induced by curcumin, was also reported to involve the activation of caspase-3 and the generation of reactive oxygen species (ROS) in a rat model of histiocytic tumor [27].
2.1.2. Modulation of immune functions and inhibition of signaling pathways
The second phase started with an important finding in the field of immuno-oncology. Using the AK-5 rat tumor model (histiocytoma), Bhaumik et al. revealed a differential activation status in host macrophages and NK cells induced by curcumin during the spontaneous regression of the tumor after subcutaneous injection of AK-5 cells [28]. This study represented a bridge between investigations focused on the pharmacology of these molecules, that were previously conducted separately in the fields of inflammation/immunology and oncology. Other reports made a functional links between the modulation of lymphocyte-mediated immune functions produced by curcumin and the induction of curcumin-related antitumor response in a mouse strain that spontaneously develop multiple intestinal adenomas [29]. Another breakthrough was the demonstration of curcumin’s action in protection of the host (tumor-bearing mice) from tumor-induced immunosuppression and toxicity [30].
The hypothesis that curcumin may suppress tumor promotion through inhibition of different signal transduction pathways was also proposed, while questions regarding the pharmacological activities of its major metabolites, after biotransformation, emerged [31]. In this context, one of the first review of the anti-carcinogenic activity of curcumin in animal models suggested that its action includes suppression of oncogenes expression and impairment of the ubiquitin-proteasome pathway [32]. Curcumin was reported to induce histones H3 and H4 hypoacetylation in glioblastoma cells, leading to hypoacetylation-enhanced apoptotic cell death. On the other hand, its application induced neurogenesis of neural progenitor cells (NPCs), through suppressing differentiation into astrocytes while promoting differentiation into neurons. These twoneurogenic phenomena were confirmed in vivo [33]. Interestingly, these findingson the anti-carcinogenic effects of curcumin resonated with preliminary studies conducted 12 years earlier, in China [34]. The same year, the inhibition of NF-κB activation and increased apoptotic rate was documented after i.p. administration of curcumin to nude mice bearing xenografts of human prostate cancer (PC-3 cells) [35]. This work was closely related to the previous research conducted by another research group, showing the inhibition of angiogenesis associated with orally given curcumin in another type of prostate cancer (LNCap) grown in nude mice [36]. Evidence of a decreased expression of cyclin D1 and phospho-IκB-α after curcumin treatment of mice bearing head and neck squamous cell carcinoma was published four years later [37]. Over the next 8 years, these discoveries were further confirmed in many other rodent and human tumor models, and the number of reports published annually increased exponentially. Meanwhile, the signaling pathways affected by curcuminoids were extended to TP53, MAPKs, Akt, Notch-1, Nrf2, JAK/STAT, Wnt/β-Catenin, AMPK/COX-2, and others [38]. From 2008, these in vitro data started to extend into the clinic through clinical trials [39], which have also been recently reviewed [13].
2.1.3. Extension of preclinical studies and first clinical trials
This stage corresponded to an extension of investigations in the field of immuno-oncology, joined with the research focused on diverse signaling pathways previously shown to be targeted by curcuminoids. During that period, the rationale of using a single cancer drug for a single target was also questioned [40]. Attempts to use curcumin combined with immunization, using soluble proteins isolated from viable tumor cells, led to an increase of the anti-tumor humoral immune response and inhibition of B16-R melanoma cells growth in mice [40]. Such research approaches led to the assessment of the potential of this phytochemical to enhance the cytotoxicity of CD8+ T cells against lymphoma cells in mice, and to increase the accumulation of transferred T cells in target tumors and caspase 3 mediated apoptosis [41]. Immunocompetent rats bearing a syngeneic peritoneal mesothelioma tumor model and treated with multiple injections of curcuminhad residual tumors infiltrated with many CD8+ T cells and decreased intensity of immunohistochemical staining of IL-6 and vimentin [42]. Immunohistochemical analyses of xenografted tumor biopsies of curcumin-treated mice bearing a murine mesothelioma revealed apoptosis that was stimulated via an increase of the pro-apoptotic signals mediated by phosphorylated p38, MAPK and CARP-1 proteins [43].
Important insights into curcumin-induced apoptosis in a model of human liposarcoma included discovery of ER stress which preceded the apoptosis, and was induced by interaction with the sarcoplasmic/endoplasmic reticulum Ca2+-ATPase 2 (SERCA2) [44]. The same process of activation of the ER stress signaling pathway was further confirmed for a synthetic curcuminoid,in a human model of lung carcinoma in mice [45]. Finally, the inhibition of SERCA2 activity by a second generation synthetic curcuminoid was later confirmed in two different models of triple negative breast cancer in mice.An induction of SERC2-related autophagy in triple negative breast cells tumor tissues was also reported,after applying RL71, a second-generation curcumin analog [46].
Meanwhile, the understanding of the mechanisms of inhibition of other signaling pathways targeted by curcuminoids was improved. In ovarian carcinoma-bearing mice, curcumin treatment was shown to suppress pathways mediated by NF-κB and phosphorylated STAT3. These phenomena were associated with a decrease in VEGF, IL-8 and MMP-9 [47]. Curcumin-related reduction of the metastatic potential of prostate xenografts in mice is connected with inhibition of the feedback loop between NF-κB and the proinflammatory cytokines CXCL1/2 [48]. Neutrophil elastase was identified as a direct protein-target of curcumin in the process of lung cancer angiogenesis. Targeting neutrophil elastase was shown to have a negative impact on the signaling cascade: HIF1α (hypoxia inducible factor 1 subunit alpha)/mTOR/VEGF/VEGFR [49].
During that period, curcumin was shown to inhibit the growth of a great diversity of human tumors in xenotransplant or orthotransplant animal models. These studies were reviewed together with the results of the first clinical trials [50]. Moreover, curcumin was shown to affectthe growthof a rat experimental glioblastoma [51]. Head and neck squamous cell carcinoma represented one of the first cancer type in which curcumin application showed promising features, relevant for the proposed therapeutic activity [52]. These effects were systematically reviewed [53]. Insights into curcumin’s effect on mammary cancer progression were provided through the demonstration of its synergistic action with β-interferon-induced upregulation of the anti-oncogenic protein GRIM-19 and inhibition of STAT3 transcription [54]. The increase of the reactive oxygen species and activation of FOXO3a, a member of the Forkhead Box Class O transcription factor, was reported as closely related to the apoptosis induced by a curcumin synthetic analog T63 in a lung cancer model in mice [55]. With a different curcumin analog, C-150, another report demonstrated that the modulation of the Notch/Akt signaling pathway was involved in curcumin-mediated effects in vivo [56]. An interesting observation was the enriched membrane localization of β-catenin, which correlated with the inhibition of tumor growth of a prostate cancer xenograft [57]. Subsequently, a confirmation of the suppression of Wnt/ β-catenin signaling pathway by curcumin was provided, showing a decreased expression of Wnt3a, LRP6, phosphor-LRP6, and phospho-β-catenin, C-myc and surviving in gastric carcinoma xenografts [58]. Another independent study highlighted curcumin-related inhibition of both, STAT3 and β-catenin in gastric carcinogenesis [59]. This inhibition of STAT3, which was previously well documented in the chemoprevention of lung cancer [60], was investigated in more detail through hybridization of curcumin with a synthetic moiety to increase the ROS level. Using this hybrid, the treatment of nude mice bearing human MCF-7 breast cancer cells led to suppression of p-STAT3 and a decrease in Ki67 immunostaining in the tumor tissues [61]. Regarding tp53, curcumin administration led to an increased expression and nuclear accumulation of tp53, and upregulation of p21 in mice bearing tp53 wild-type H460 xenografts, but not in those bearing tp53-null H1299. It was later shown that curcumin has an activating effect on the axis: tp53-miR-192-5p/215-XIAP in lung cancer [62], which is relevant, and contributes to apoptosis. Another work complemented these findings by showing curcumin induced apoptosis of head and neck squamous cell carcinoma in vivo, through ATM/Chk2/tp53-dependent pathway [63].
To end up, three other major breakthroughs characterized this period. The first one was the discovery of curcumin’s ability to inhibit the proteasome activity, a discovery which was originally obtained in colon cancer and confirmed later in other cancer types [64]. The second was a confirmation of the functional connection between the inhibition of NF-κB pathway and epigenetic modulation induced by curcumin, through downregulation of DNA methyltransferase 1, which was discovered in a mouse model of acute myeloid leukemia [65]. The third finding is curcumin’s effect on ATP synthase [66], a crucial point related to reprogramming of tumor metabolism, which will be developed in the second part of this review.
2.2. Recent findings, and new developments
Among recent findings, important improvements were reported at the interface of immune system and cancer cells, through the research in immuno-oncology. Curcumin treatment contributed to reinvigorate defective T cells via multi-level immune checkpoint axis suppression, in xenograft tumor models of head and neck cancer in nude mice [67]. Our understanding of the complexity of curcumin’s actions against this cancertypealso benefited from ex vivo studies, on tumor tissue explants from patients. These studies confirmed curcumin’s role in the inhibition of NF-κB nuclear translocation and its immune stimulatory effect in the tumor microenvironment [68]. Using the same methodological approach, another interesting study showed how curcumin changed immune cell composition and localization in colorectal cancer and adenoma patient-derived explants [69]. In a liver cancer xenograft model in mice, curcumin treatment impacted another population of immune cells involved in immunosuppression, myeloid-derived suppressor cells (MDSCs), inhibited the TLR4/NF-κB signaling pathway, and the expression of inflammatory factors, including IL-6, IL-1β, prostaglandin E2 and cyclooxygenase-2 (COX2) [70]. These works were complemented by the demonstration that a curcumin analog prevented the growth of a melanoma through the reduction of the population of Foxp3+Tregs tumor-infiltrating lymphocytes [71]. At this immuno-oncology interface, other insights were provided through the combined use of a model of aggressive peritoneal mesothelioma in immunocompetent rats [42] and high-throughput proteomic analyses applied to formalin-fixed paraffin-embedded tissues. In a first step, an analysis of abundance changes in 1411 liver proteins identified a set of proteins associated with the curcumin-induced immune response that included the purine nucleoside phosphorylase (PNPH), the enzyme that is involved in T cell functions [72]. Secondly, comparison of the mesenteric lymph node and liver proteomes obtained from the same rats, revealed a high level of similarity with respect to abundance of PNPH [73]. Finally, proteome analyses of tumor tissues from curcumin-treated rats revealed that there are specific changes measurable through abundance of 22 proteins regulating the tumor microenvironment, including a continuous rise in caveolin-1, a protein regulating immune cell infiltration, T cell activation and dendritic cell maturation [74].
Improvements are apparent when one follows on the diversity of signaling pathways targeted by these molecules. The inhibition of growth and invasion of a human monocytic leukemia in mice was joined with alteration of MAPK and MMP signaling [75]. In a glioblastoma model, the inhibitory action of curcumin on tumorigenesis in vivo, was characterized by an inhibition of the p-AKT/mTOR pathway, but also an enhancement of the expression of the tumor suppressor protein tp53 [76].The mechanism of inhibition of the Wnt signaling pathway induced by curcumin in vivo was reported to involve the downregulation of axin2, a negative feedback regulator of this pathway [77]. The knowledge on the role of non-coding RNAs has been also progressing. In this field, a first insight related to curcumin was provided through discovery of his influence on the axis that includes hsa_circ_0007580 (circ-PRKCA), miR-384 and integrin subunit beta 1 (ITGB1), in the non-small cell lung cancer (NSCLC) model. Curcumin decreases circ-PRKCA, a sponge for miR-384, resulting in miR-384 increase and ITGB1 decrease, which, all together, leads to a decrease of biological aggressiveness of cancer cells) [78]. A second report documented the inactivation of the JAK2/STAT3 pathway in a liver cancer model, in a similar fashion: a curcumin analog, GL63, contribute to a decreased expression of circZNF83, a sponge for miR-324-5p, whose target is cyclin-dependent kinase 16 (CDK16) [79].
Important findings were also revealed in fouradditional fields. In relation to mitochondria dysfunction, a first one corresponded to the generation of ROS and subsequent YAP mediated JNK activation produced by a curcumin analog (while low protein levels of YAP were observed in breast cancer tissues) [80]. The accumulation of ROS, which, together with intracellular iron, represents a hallmark of ferroptosis, was reported to be specific for the action of curcumin in lung cancer,in vivo [81]. To explain the strong effect of both, curcumin and inhibitors of the mitochondrial sodium-calcium-lithium exchanger (NCLX) on colon cancer in vivo, a common mitochondria calcium overload was suggested to lead to mitochondrial membrane depolarization [82]. A second axis, represented by the activation of ER stress produced by curcuminoids, received additional insights through the report that demonstrated an increase of the expression of ATF4 and CHOP [83]. Two enhanced signaling pathways, PERK-eIF2a and IRE1a-XBP1, were also identified in glioma xenografts in mice following radiotherapy-induced immunogenic cell death [84]. A third research field was represented by curcumin’s inhibitory effect on proteasome activity, which, at the molecular level, benefited from the demonstration of the involvement of the p300/miR-142-3p/PSMB5 axis [85]. In the fourth research field, that included transcription regulation, curcumin was found to produce a reduction of the expression of the histone methyltransferase EZH2 in a xenograft mouse model of myelodysplastic syndrome [86]. An activation of Nrf2 protein and expression of its target, HMOX-1, was also observed in vivo, in mice after topical treatment [87]. In a work dedicated to the understanding of the mechanism of interleukin 17-A mediated acute lung injury in mice, a proteomic analysis revealed that the increased level of several minichromosome maintenance proteins associated with increased level of IL-17A, can be reversed by curcumin [88].

Table 1.
Main in vivo studies reporting curcumin effects in experimental tumor models and in cancer patients.
Table 1.
Main in vivo studies reporting curcumin effects in experimental tumor models and in cancer patients.
| Major findings | Tumor type | Adm. route | Hist. | Ref |
|---|---|---|---|---|
| sympomatic relief, ↓ lesion size | Patients with external cancers | topical | - | [5] |
| tumor growth inhibition | Head & neck carcinoma (M) | topical | - | [37] |
| ↓ CD31 and HIF-1α expression | Head & neck carcinoma (M) | p.o. | + | [63] |
| ↑ IFN-γ and granzyme expression | Head & neck carcinoma (M) | i.p. | + | [67] |
| ↑ T-cell proliferation, ↓ PD-1 expression | Head & Neck carcinoma (M) | i.p. | + | [85] |
| humoral response (+ immunization) | Melanoma B16-R (M) | i.p. | - | [40] |
| ↓ ATP-synthase activity, ATP/AMP ratio | Melanoma B16-R (M) | i.v. | + | [66] |
| ↓ Foxp3+ Tregs in tumor | Melanoma B16-F10 (M) | i.p. GO-Y030 * | - | [71] |
| ↓tumor formation | Dalton’s lymphoma (M) | i.p. (liposomes) | - | [4] |
| downregulation of DNMT1 | Acute myeloid leukemia (M) | i.p. | - | [65] |
| ↓ MMP2, MMP9, vimentin expression | Monocytic leukaemia (M) | i.p. | + | [75] |
| ↓ EZH2, H3K4me3, H3K27me3 | Myelodysplastic syndrome (M) | - | - | [86] |
| apoptosis induction | Colon adenocarcinoma (R) | p.o. (diet) | + | [24] |
| inhibition of colon tumorigenesis | Colon adenocarcinoma (R) | p.o. (diet) | - | [26] |
| tumor prevention, ↑ CD4+ T-cells, B cells | Genetic colon cancer (M) | p.o. (diet) | + | [29] |
| inhibition of proteasome activity | Colon cancer HCT-116 (M) | p.o. | + | [64] |
| ↓ PCNA, β-catenin, Axin-2 | Colon cancer (M) | ? | + | [77] |
| ↑oxidative stress, mitochondrial Ca2+ | Colorectal cancer (M) | i.p. | + | [82] |
| inhibition of tumor-induced immune cell ↓ |
Ehrlich’s ascites carcinoma (M) | p.o. | + | [30] |
| no peritoneal bulge + survival | Histiocytic tumor AK-5 (R) | i.p. | - | [27] |
| ER stress-associated apoptosis | Liposarcoma (M) | i.p. | + | [44] |
| ↓ MDSCs, lL-6, Il-1β, GM-CSF secretion | Hepatocarcinoma HepG2 (M) | p.o. | + | [70] |
| inactivation of JAK2/STAT3 pathway | Hepatocellular carcinoma (M) | i.p. GL63 * | - | [79] |
| suppression of Wnt/ β-catenin signaling | Gastric carcinoma (M) | p.o. | - | [58] |
| inhibition of β-catenin and STAT3 | Genetic gastric cancer (M) | p.o. (diet) GO-Y031* |
+ | [59] |
| activation of ER stress pathway | Adrenocortical carcinoma (M) | i.p. | + | [83] |
| induction of cleaved caspase-3 and PARP | Prostate cancer PC-3 (M) | i.p. | + | [35] |
| ↑apoptosis, ↓ tumor angiogenesis | Prostate cancer LNCaP (M) | p.o. (diet) | + | [36] |
| ↓lung metastasis | Prostate cancer PC-3 (M) | p.o. (diet) | + | [48] |
| ↑membrane localization of β-catenin | Prostate cancer C4-2 (M) | intratumoral | + | [57] |
| NF-κB and p-STAT3 suppression, ↓ Il-8 | Ovarian cancer (M) | p.o. | + | [47] |
| synergically↑ IFN-β induced apoptosis | Breast cancer (M) | p.o. | - | [54] |
| ↓tumor incidence, DNA adducts | Mammary carcinoma (R) | i.p. | - | [23] |
| inhibition of SERCA2, ER stress | Breast cancer (M) | i.p. (RL71) * | + | [46] |
| inhibition of STAT3 phosphorylation | Breast cancer (M) | i.p. Curcumin-BTP hybrid * | + | [61] |
| N-cadherin, MMP2, MMP9 suppressed | Breast cancer (M) | i.p. WZ35 * | + | [80] |
| ↓ HIF1α/mTOR/VEGF cascade | Lewis lung cancer (M) | ? | - | [49] |
| ↑ expression of FOXO3a, p27, p21 | Lung cancer A549 (M) | i.p. | + | [55] |
| activation of p53-miR-192-5p/215-XIAP | Lung cancer (M) | p.o. | + | [62] |
| ↓circ-PRKCA, ITGB1 expression | Lung cancer A549 (M) | p.o. | - | [78] |
| induction of ferroptosis via autophagy | Lewis Lung carcinoma (M) | i.p. | + | [81] |
| immune response (CD8+ T cells), ↓ Il-6 | Mesothelioma (R) | i.p. | + | [42] |
| ↑p38, MAPK and CARP-1 | Patient mesothelioma (M) | p.o. | + | [43] |
| proteome changes (liver invasion) | Mesothelioma (R) | i.p. | + | [72] |
| proteome changes (residual tumors) | Mesothelioma (R) | i.p. | + | [74] |
| ↓tumor volume, hemorrhage | Glioblastoma C6 (R) | i.p. | + | [51] |
| ↑ PTEN and P53 expression | Glioblastoma U87 (M) | i.p. | + | [76] |
| ↑ HSP70, ER stress, immune cells | Glioma GL261 (M) | i.p. | + | [84] |
* Curcumin analogs. p.o. per os, i.p., intraperitoneal; i.v., intravenous injection. Hist., histological data included (+), not included (-). Animal models, M (mouse), R (rat).
3. Curcumin and chemosensitization through metabolic reprogramming
In 2013, Vishvakarma et al. were shown decreased response of cancer cells (DL – Dalton’s lymphoma) to cisplatin and methotrexate if they were exposed to high concentration glucose. This phenomenon was associated with high production of lactate, acidification of the medium, and increased expression of one of glucose transporters, GLUT1. Although the authors did not perform mechanistic studies, they concluded that increased rate of cancer cell’s glycolysis contributes to therapy response [89]. In a recently published paper [90], availability of glucose (up to 30 mM) and overexpression of GLUT1 were shown to be crucial for development of chemoresistance against doxorubicine and methotrexate in liver cancer originating cells, HepG2.
The metabolic reprogramming is a well-known feature of the cancer cell, which develops for securing numerous cancer-specific features. Cancer cell must sustain activity of its biosynthetic machinery to support its own replication capacity and high mitotic rate. Its ability for metabolic reprogramming highly depends on availability of two major growth-supporting substrates: glucose and glutamine.
In non-transformed cells, the precise regulation of glucoseintake is under strict control of growth factors [91] that develop a molecular communication network which extends to PI3K/AKT/mTOR pathway and results in an increased expression of transporters for glucose and amino acids [92]. The activity of this pathway is sufficient to increase the size of resting cells (G0). Malignant cells do not have to rely on this physiological regulation. Instead, their specific oncogenotype allow them to adapt their metabolism with regard to available nutrients in order to sustain their biomass and proliferation capacity through glycolysis.
Glycolysis takes place in the cytoplasm through a cascade of several enzymatic reactions, producing pyruvate and only two molecules of ATP. If oxygenation is sufficient, non-transformed cell convert pyruvate into Acetyl-CoA and directs it into the TCA cycle. If there is no sufficient oxygen, lactate dehydrogenase A (LDH-A), the enzyme that is commonly overexpressed in malignant tumors [93] converts pyruvate into lactate. It has been known for a while that promoter of LDH-A has a binding site for HIF-1α [94] which, especially in hypoxic environment, positively regulates the activity of the LDH-A gene [95]. Malignant cells, even if there is a sufficient supply of oxygen, commonly avoid coupling glycolysis with TCA (a phenomenon known as Warburg effect) with a consequential increase oflactate production and low production of energy. This, however, does not exclude them from utilizing oxidative phosphorylation (OXPHOS), which is preferable metabolic process in non-transformed cells. None of these metabolic pathways is a unique event, for these cells: the Warburg effect may occur also in non-transformed cells, and is now recognized as a metabolic state in which cell meets short-time scale energy demands [96]. Genetic setting of malignant cell, in combination with a complex epigenome, directs activity of genes involved in cellular metabolism that enables cancer cells to produce different nutrients (metabolic flexibility) and process them in various ways (metabolic plasticity) [97].
Although traditionally considered as a „metabolic waste“, lactate – produced from pyruvate, is now recognized as a oncometabolite which is involved in development of resistance to radio-/chemotherapy [98]. However, lactate is only one of many factors that contributes to development and maintenance of cancer hallmarks, including chemoresistance. It is the whole specific oncometabolic feature of the cancer cell that allows and favors a straightforward glycolytic conversion of glucose into pyruvate into quite complexed branching points. That is possible because every step of glycolysis results in occurrence of a specific intermediary product which cancer cell can, in dependence on its genomic constitution, use for its own highly demanding biosynthetic needs, while keeping the energy production at a very low level, with consequential increased uptake of glucose. The activity of specific branch, of which only two are presented on Figure 1, is determined through rate-limiting enzymes and balanced equilibrium of respective intermediates [99].
Curcumin, through its pleiotropic mode of action has a potential for influencing glycolytic process and most, if not all the metabolic branching points. In this, no doubt, broad and complex part of cancer cell biology, we tried to connect data related to select processes involved in cancer cell metabolism, resistance to applied therapy and beneficial effects of curcumin application that is associated with specific metabolic features of cancer cells.
3.3. Curcumin: Glycolysis and Lactate Production
Improvement of therapeutic efficacy of chemotherapeutics in the presence of curcumin in various in vitro models, as well as in experimental animals, has been documented and reviewed numerous times [100]. Recent reports indicate curcumin’s strong influence ondecrease of lactate production and/or inability of cancer cell to excrete lactate. Once excreted, lactate acidifies the tumor microenvironment, and contributes to aggressive behavior of malignant tumors [98]. In vitro model of liver cancer and T cell lymphoma (HepG2 and HUT78) was explored for showing that curcumin, already in low concentration (5μM/24 hours), significantly reduces lactate concentration in the medium and, through that mechanism, contributes to increased sensitivity of HepG2 cells to doxorubicin and methotrexate. External addition of 20 mM lactate reversed the phenotype toward chemoresistance [90], and was associated with increased activity of genes coding for proteins whose oncogenic potential was shown in various cancer models (STAT3 and HIF-1α), earlier mentioned LDHA, lactate receptor - HCAR1 (hydroxycarboxylic acid receptor 1/GPR81), and ABCB1 (ATP binding cassette subfamily B member 1; also known as MDR1 or P-glycoprotein). In 2017, Wagner et al showed that lactate, acting through its receptor – HCAR1, strongly increases expression and activity of ABCB1 through yet unknown molecular mechanism, resulting in PKC-dependent decreased doxorubicin sensitivity of the HeLa cervical cancer cell line [101]. In that scenario, chemoresistance related to increased production of lactate may be negatively influenced by curcumin not only through curcumin’s negative regulation of HCAR1 [101], but also through a negative regulation of the GLUT1 transporter and consequential decreased cellular availability of glucose [102].
Proteomic study performed after electric pulse application of 50 μM curcumin (EP_Cur)to triple negative breast cancer (TNBC) cell line MDA-MB-231 demonstrated decreased level of glycolytic enzymes and, at the same time, increase of enzymes involved in oxydative phosphorylation, associated with dramatic decrease of cellular lactate [103]. This discovery clearly demonstrates multilevel acting of curcumin. In colon cancer cell lines HT-29 and HCT116, application of high concentration of curcumin (40 μM) for 24 hours resulted in significant decrease of lactate production (39.1% and 34.5%, respectively) [104].
As shown on Figure 1, the first step of glycolysis is catalyzed by hexokinase. There are four highly homologous hexokinase isoforms in mammalian cells. AKT potentiates hexokinase activity [105], resulting in phosphorylation of glucose molecules, and their retention in the cell.
Figure 1.
Curcumin modulates glycolytic metablic pathway and has a potential to modulate de novo serine synthesis pathway (SSP). Blue arrows:decrease of transcript/protein/enzyme activity. Stars: direct binding of curcumin to the the target protein.
Figure 1.
Curcumin modulates glycolytic metablic pathway and has a potential to modulate de novo serine synthesis pathway (SSP). Blue arrows:decrease of transcript/protein/enzyme activity. Stars: direct binding of curcumin to the the target protein.
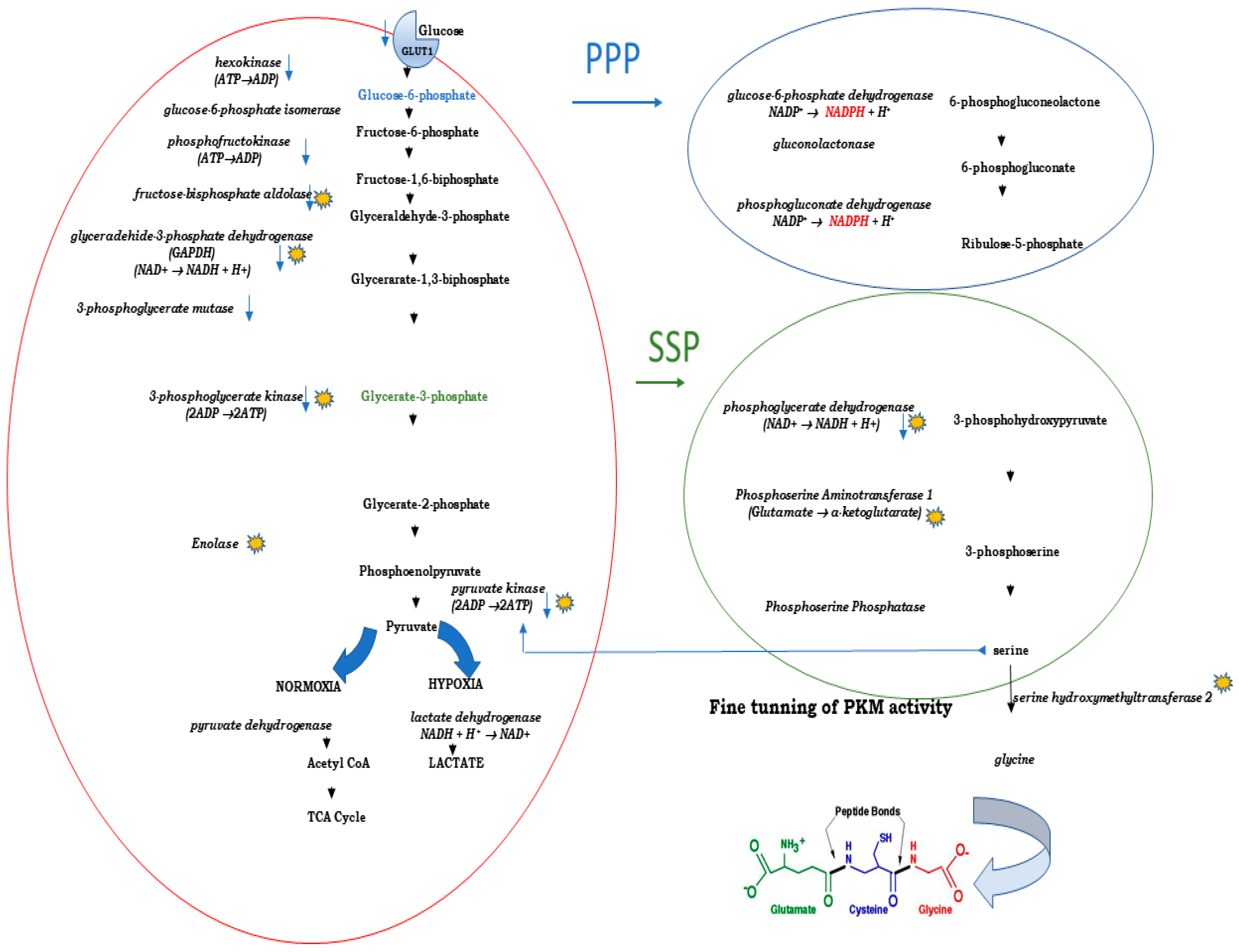
Hexokinase 2 (HEX2) was shown to be highly expressed in various malignant tumors [106]. At least in vitro, the level of its expression is significantly different in triple-negative breast cancer TNBC (model: MDA-MB-231; high expression) compared to estrogen receptor positive breast cancer cells (model: MCF-7; low expression). High expression of HEX2 in TNBC is under direct transcriptional control of overexpressed transcription factor SLUG. High expression of SLUG and HEX2 strongly associates with resistance of MDA-MB-231 to 4-hydroxytamoxifen (4-OHT). The resistance can be ameliorated through combined application of curcumin and 4-OHT, leading to apoptotic death [107]. In colon cancer cell lines, HT-29 and HCT116, application of 40 μM curcumin for 24 hours resulted in decreased rate of glycolysis, associated with significant decrease of HEX2, and its dissociation from the outer mitochondrial membrane [104].
There is a strong evidence that, in cancer cells, curcumin fatally affects oxidative phosphorylation and contributes to energetic deficit through inhibiting ATP-synthase activity, which is associated with significant increase of ROS (Reactive Oxygen Species) [66]. Expectedly so, it was hypothesized that, although exposed to curcumin,transformed cells may be able to replenish lack of ATP through switching to a less efficient energy producing metabolic pathway - glycolysis. When testing enzymatic activity of glycolytic enzymes and extracellular concentration of lactate in four murine cancer cell lines (L1210 - lymphocytic leukemia, 4T1- breast, B16 murine melanoma and CT26 - colon), it was demonstrated that cells exposed to curcumin have a significant decrease of activity of hexokinase, lactate dehydrogenase, phosphofructokinase and pyruvate kinase (which may indicate the switch from active PKM1 to less active PKM2 – will be discussed later) activity, in all, but melanoma-originating B16. At least for phosphofructokinase and lactate dehydrogenase A/B, this may be a consequence of their decreased cellular level – as demonstrated in some other experimental models [108]. There was a clear distinction between neuroectoderm-originating melanoma B16 and other three cell lines, as all measured glycolytic parameters only in B16 cell line become significantly increased, probably reflecting its specific origin.
In two human cell lines originating from glioblastoma (U-87 MG) and neuroblastoma (SH-SY5Y), exposure to curcumin and its analogue, MS13, induced significant change in the level of various proteins including few involved in metabolic regulation: glyceraldehyde-3-phosphate dehydrogenase (GAPDH; decrease in both MS13-treated cell lines) and phosphoglycerate kinase 1 (PGK1) in MS13-treated SH-SY5Y [109]. In human leukemia cell lines, K562 and LAMA84, exposure to curcumin was associated with severe change of numerous proteins (143 up-regulated and 234 down-regulated). Among decrased proteins werephosphoglycerate mutase 1, phosphoglycerate kinase 1,D-3-phosphoglycerate dehydrogenase (a rate limiting enzyme of serine de novo biosynthetic pathway) and pyruvate kinase (PKM) [110]. Curcumin can directly target metabolic enzymes. It was shown in another study, based on cell-permeable clickable curcumin probe and quantitative chemical proteomics in colon cancer cell line, HCT116 [111]. A stringent profiling revealed the following metabolic enzymes as curcumin binding partners: pyruvate kinase isozymes M1/M2 , fructose-bisphosphate aldolase A, glyceraldehyde-3-phosphate dehydrogenase, alpha-enolase (enolase 1), L-lactate dehydrogenase A and B chains, phosphoglycerate kinase 1, D-3-phosphoglycerate dehydrogenase, mitochondrial serine hydroxymethyltransferase (SHMT2).
It is very interesting that phosphoglycerate kinase 1, in addition for being presented as crucumin’s binding partner, turned to be listed as downregulated in all proteomic studies we could find, related to cancer and curcumin. As recently reviewed, high expression of phosphoglycerate kinase 1 is positively associated with chemoresistance in all cancer models explored so far [112]. In 2000, Elson et al showed that HIF-1α, PGK1, GLUT1 and VEGF occur in the earliest phase (early-stage hyperplasia) of multistage epidermal carcinogenesis [113]. Only four years later, Li et al. showed that PGK1 has the HIF-1α binding site in its promoter and that transcriptional activity of PGK1, under hypoxic condition, depends on HIF1α [114]. This transcription factor has been associated with chemoresistance in various types of cancer [115]. Extracellular ATP can stimulate hypoxia-inducible factor (HIF) signaling and contribute to breast cancer cell resistance even under normoxic condition [116]. The most recent data show that STAT-3 – mediated ALDOA (fructose-bisphosphate aldolase A, binding to HIF1α (which itself contributes to multiple resistance in cancer) strongly contributes to development of chemoresistance against cisplatin (models: breast cancer cell line MCF-7 and xenografts MDA-MB-231 [117].
In these scenarios, curcumin indeed may – due to its pleiotropism, add to chemosensitization through synchronous actions at several molecular levels. For example: a) through downregulation of ALDOA [103], b) through negativeinfluence on HIF1α, as recently reviewed [118], c) through inhibitory effect on STAT3 (direct binding of curcumin to STAT3 has been discovered a few years ago [119]). All these data clearly show that metabolic reprogramming of cancer cells, when in favor of glycolysis and lactate production, strongly increases chemoresistance of tumor cells. They also show a very strong, functional interconnection between the metabolic status of the cell and various signaling pathways that are not traditionally considered „metabolic“. This shows that none of the hallmark of cancer should be considered as an isolated entity because molecules and processes related to cancer hallmarks are shared among various signaling pathways and one pleiotropic molecule may indeed affect several of them at the same time.
3.2. Curcumin and Lactate Excretion
Glycolytically produced lactate must be excreted from the cell, where it contributes to extratumoral acidosis. It is known that malignant tumors maintain their hallmarks through maintenance of intracellular alkality (pHi ≥ 7.4) and extracellular acidity (pHe ~6.7-7.1) [120]. These values are significantly different from values related to differentiated cells (pHi ~7.2; pHe ~ 7.4). This cancer-related phenomenon, known as „pH gradient reversal“ was recognized as a possible vulnerability that may be triggered by therapy, back in 1996 [121] and was recently reviewed [122]. Lactate shuttle, that is needed for establishment of synergistic metabolism between glycolytic tumor cells andoxidative tumor cells,is highly dependent on activity of lactate transporters, MCT1 and MCT4 (SLC16A1 solute carrier family 16 members 1 and 4). In order to explore the effect of chemosensitization of the whole extract of the C. Longa, and then each active compound separately: curcumin, biscurcumin and demethoxycurcumin, with respect to cancer cell response to 5-FU and lactate metabolism, Li et al. [123] used 5-FU resistant colon cancer cell line, HCT8, previously shown to overexpress MCT1 [124]. The authors were able to show that the chemosensitizing effect of the whole extract, in this experimental model, partially relates to a significantly decreased expression of MCT1 and high increase of intracellular lactate (three- and four-fold change), after application of curcumin and biscurcumin, respectively. It is expected that so significant increase of intracellular lactate changes the intracellular pH (pHi), creating a scenario which may have a potential for being detrimental for cancer cell (toxic acidosis). In another model, application of curcumin was shown to have a potential for shifting intracellular pH of cancer cell toward acidic through decreasing both, MCT1 and Na+/H+ antiporter NHE1 (SLC9A1 - solute carrier family 9 member A1 [125], which also may contribute to curcumin’s chemosensitizingproperties.
As recently shown, increased intracellular acidity induced by hypoxia plus MCT1/2 inhibition significantly compromises the survival of MCF7 breast cancer cells. The effect was amplified when glyceraldehyde-3-phosphate dehydrogenase (GAPDH), was silenced [126]. Whether exposure to curcumin produces exactly the same mode of action remains to be explored. Obviously, pleiotropic action of curcumin can target cancer cell metabolism at many levels that are closely related and interconnected.
3.3. Curcumin and Pentose Phosphate Pathway (PPP)
Although not providing adenosine 5′-triphosphate (ATP) for cellular energy demands, high activity of PPP in cancer cells has a strong contribution to a reducing cellular power (through production of NADPH) and ribonucleotides synthesis (through production of ribulose 5- phosphate (R5P) [127]. Production of NADPH through PPP is needed for generation of reduced glutathione (GSH) which allows for alleviation of oxidative stress through successful removal of ROS [128]. This oxidative branch of PPP relates to the activity of two enzymes, glucose-6-phosphate dehydrogenase (G6P) and phosphogluconate dehydrogenase (PGD).
As recently reviewed, the increased activity of PPP relates to cisplatin chemoresistance that can be developed through several mechanisms [129] The NADPH generated in the PPP counteracts, forexample, a high cisplatinum cytotoxicity associated with a high production of ROS. Expectedly so, inhibition of the two enzymes related to NADPH generation, G6P and PGD, makes cancer cells sensitive to cisplatin [130,131].
There are no data on related to the selective effect of curcumin on G6PD specifically in cancer cells. However, feeding animals with curcumin (2%, w/v) increases activity of G6PD in mice liver and kidney tissues for 89% and 67%, respectively, contributing to systemic defense against oxidative stress, as expected [132], in a chemopreventive, and not therapeutic, setting.
3.4. Curcumin, Pyruvate Kinase and Serine Synthesis Pathway (SSP)
In cancer cells, biosynthesis of serine (SSP – Serine Byosinthesis Pathway) depends on several interconnected factors: a) availability of the precursor: 3-phosphogylcerate (glycerate 3-phosphate; labeled green on Figure 1) whose generation is dependent on glucose intake and presence of metabolically inert pyruvate kinase 2 (PKM2), b) ATF4-directed expression of three metabolic enzymes: 1) PHGDH - Phosphoglycerate-3-Dehydrogenase; 2) PSAT1-Phosphoserine Aminotransferase 1, and 3) PSPH-Phosphoserine Phosphatase.
There are numerous studies demonstrating an increase of each of these enzymes, in various malignant tumors. There is also a study showing the increased protein level of all three proteins (fold change: PHGDH: + 3.62; PSAT 1: + 6.94: PSPH: + 1.27) in lung cancer tissue, when compared to corresponding non-tumorous tissue [133]. In cancer cells, serine has important role associated with activity of pyruvate kinase (PKM).
Pyruvate kinase catalyses the last glycolytic step: transfer of phosphate group from phosphoenolpyruvate (PEP) to ADP for generation of ATP and pyruvate. Contrary to isoenzyme PKM1, which is highly expressed in tissues with high energetic needs (heart, muscle, brain), expression of PKM2 is common in malignant tumors. It is considered as cancer-specific isoenzyme. The difference between PKM1 and PKM2 depends on alternative splicing of the primary transcript of the PKM gene.
The PKM2 activity depends on a highly complex, serine-dependent allosteric regulation: contrary to PKM2 tetramers that are strong catalyzers (as are PKM1 tetramers),PKM2 specific dimers are catalytically inert and they support biosynthetic processes [134]. Lack of serine favors accumulation of inert PKM2 dimers, decreased generation of pyruvate, and accumulation of glycolytic intermediary products of which one – 3PG enters the SSP (Figure 1). De novo synthesized serine may be converted to glycine throughactivity of mitochondrialserine hydroxymethyltransferase 2 (SHMT2). Glycine is then used for synthesis of glutathione (GSH) and purine nucleotides.
Thus, in its essence, the active SSP is beneficial for cancer cell not only with respect to serine supply (when needed) , but also for development of resistance to therapy, due to its positive impact on GSH synthesis which is needed for cellular defensive response against therapy-induced oxidative stress. Curcumin binds to SHMT2 [135] and there is a high probability that the binding impacts both, the structure and the catalytic activity of the active form of SHMT2 [136] which was recently shown to drive resistance to 5-FU in colorectal model of cancer [137].
Increased level/activity of enzymes directly and indirectly involved in SSP and pyruvate kinase activity have been shown in various experimental models, and in native tumors. The importance of some of them for development of resistance to therapy was also shown in variouscancer models. However, available data is not necessarily unequivocal and asks for a careful interpretation with respect to thespecifics of cancer origin: high expression of PKM2 was shown to be associated with resistance to cisplatin in bladder cancer [138], but, on the other hand, it enhances response to cisplatin in cervical cancer through a complex interaction with the mTOR signaling pathway [139]. Although an indicator of worse clinical prognosis in breast cancer patients, high expression of PKM2 was shown to strongly associate with a positive therapy response (epirubicin and 5-fluorouracil), resulting in longer disease free survival and overall survival [140]. Most of studies related to the role of PKM2 in the setting of cancer therapy were recently reviewed [141].
The published studies demonstrate that curcumin’s effect on PKM2 are not entirely conclusive: Sidiqqui et al. [142] demonstrated that curcumin decreases the PKM2 level at the level of mRNA and protein in cell lines of different origin (H1299 – non-small cell lung cancer; MCF7 – breast cancer; HeLa- cervical cancer). There are data showing that the type of response may be cell-type specific: for example curcumin was shown to decrease PKM2 mRNA level in only one (Cal 27), among three tested cell lines originating from head and neck carcinoma (Cal 27, FaDu, Detroit 562) [143]. Yadav et al have shown that curcumin influences the splicing of the PKM transcript in favor of PKM1 and, through that mechanism reduces the level of PKM2 transcript and corresponding level of inert PKM dimers [144].
The limiting enzyme for serine de novo synthesis is NAD+-consuming PHGDH. Its high expression in cancer was originally shown to be associated with amplification of the chromosomal locus 1p12, where the gene resides, in melanomas and breast cancer [145]. During the last ten years, serine was recognized as a very important molecule involved in cancer cell metabolism, and the rate-limiting enzyme for it synthesis, the PHGDH, came in the focus of research, asa potential therapeutic target. Many various PHGDH inhibitors have been developed and tested, but are still notmaking their way into the clinic [146].
In tissues with extremely low serine availability (brain tissue), inhibition of PHGDH was shown to be very beneficial therapeutic approach: TNBC originating metastases cannot develop in brains of animals with inhibited activity of PHGDH [147]. High level of PHGDH relates to resistance to cisplatin, 5-FU and Sorafenib in ovarian cancer [148], colorectal cancer [149] and liver cancer [150], respectively. Resistance to other types of cancer therapy, mediated through PHGDH, was reviewed recently [151]. Although commonly presented in a simple way, the metabolic scenario in which PHGDH exerts its pro-tumorigenic activity is multilevel and highly complex.
As recently demonstrated [152], a strong association between relapse of disease in a cohort of ovarian cancer patients receiving cisplatinum and, at the same time,having a low level of PHGDH, was well-documented. The phenomenon was explored on a molecular level: when exposed to cisplatin, cancer cells balance available NAD+ between PARP-1 (PARP-1 activity is needed for DNA repair) and PHGDH in favor of repairing mechanism mediated by PARP1and, instead activating SSP, taking serine from the medium. Indeed, in experimental animals, a diet with decreased content of serine/glycine combined with selective inhibition of PHGDH was shown to have a very potent effect with respect to inhibition of cancer growth, in vitro, and in vivo [153].
PHGDH is shown to be curcumin binding partner in three experimental systems: benign schwannoma [154], HeLa (cervical cancer) [135], and HCT116 colon cancer cells [111]. One would expect that curcumin bound to PHGDH enzyme NAD+ pocket abolishes its activity and negatively influences pro-proliferative SSP.Although there are many data which show an association of PSAT1 and biological behavior of malignant tumors, there are no many data on its involvement in chemoresistance. That is surprising because the activity of this second enzyme in serine de novo biosynthesis was originally shown to be included in development of chemoresistance to FOLFIRI treatment (leucovorin, 5-FU and irinotecan) in a small cohort of colon cancer patients, already in 2008 [155]. The most recent data indicates its association with tumor immune microenvironment [156]. In glioblastoma multiforme, regorafenib (inhibitor of multiple tyrosine kinase) exertes its effect through stabilization of PSAT1, leading the malignant cell into autophagy. In this scenario, regorafenib may induce autophagy only if high level of PSAT1 exists in the cell [157].
The third third SSP enzyme, phosphoserine phosphatase, is associated with poor therapeutic response of ER+ breast cancers to tamoxifen [158] and acts as an oncogene in various types of malignant tumors, including non-small cell lung carcinoma (NSCLC) [159].
In 2015, Chiang et al have shown that exposure to curcumin reduced gene activity of all three SSP enzymes: PHGDH (-3.19), PSAT1 (-2.27) and PSPH (-2.20), using the model of lung cancer cell line, NCI-H460 [160].
4. Curcumin as a chemosensitizer in conventional chemotherapy
4.1. Chemosensitization: need of the hour.
Natural products from diverse sources encompass a unique arena of chemical space that overlaps extensively with pharmaceuticals, which are not found in synthetic chemical libraries. These natural compounds have been evaluated for long time for their ability to selectively target different kinds of ailments, including cancer. Most of the anticancer drugs used in chemotherapy are natural products derived compounds [161,162]. Nowadays, most of the currently available chemotherapeutics fail to accomplish the expected outcome. Upon prolonged exposure to the chemotherapeutics, the emergence of inherited and acquired chemoresistance due to the up regulation of major survival signals is regarded as the main cause for this drawback. Combination chemotherapy is currently used for various cancer types, and this approach is more effective compared to single agent treatment [163]. Multidrug resistance (MDR), the ability of tumor cells to develop resistance to a broad range of structurally and functionally unrelated drugs is also a major hindrance in the success of combination chemotherapy. Apart from the classic mechanisms of MDR development (over-expression of drug efflux pumps and increased activity of DNA repair machinery), alterations at the level of apoptos control serve as a crucial mechanism for the induction of drug resistance [161]. Increasing the dosage of the drug can certainly help in evading the condition, but serious side effects limit the success of clinical outcome of chemotherapy. It may also lower patients’ quality of life, and, in some cases, even results in discontinuation of chemotherapy [164].
Thus, chemosensitization can be a choice that absolutely matches the need. Chemosensitization can be defined as a process by which a non-toxic compound of either natural or synthetic origin sensitizes the cancer cells to a cytotoxic therapeutic agent without affecting the efficacy of the same. This has an added advantage of minimizing the dosage of the chemotherapeutic and thereby decreasing the side effects. Additionally, this strategy is more economic while considering the cost of currently available chemotherapeutic drugs, especially in the case of developing countries, where the rate of cancer incidence is relatively high. Since chemoresistance is a tightly regulated process under the control of multiple survival pathways, the inhibition of any single molecule may not be sufficient to circumvent the phenomenon. Hence, compounds that can simultaneously modulate multiple survival signaling pathways might provide a better therapeutic outcome than that of individual inhibitors. Several phytochemicals have been shown to modulate multiple pathways involved in chemoresistance and, hence, are assumed to be of better chemosensitizing efficacy.
4.2. Curcumin: the celebrity among nutraceuticals
From ancient days onwards, Indian and Chinese traditional medicine had made use of combinations of medicinal herbs. Among those, flavonoids are a large subgroup of the family of natural polyphenolic compounds, which are mostly the part of secondary metabolism in plants [165,166]. Most of the plant polyphenols including curcumin, resveratrol, genistein, quercetin, epigallocatechin gallate (EGCG), luteolin, apigenin, chrysin, tannic acid etc. are dietary compounds which are part of our day today diet. Amongst the wide range of natural polyphenols, curcumin is the most studied natural compound with perfect documentation of its therapeutic effect in large number of disease conditions including cancer [167,168].
Curcumin is well known for its chemopreventive [169,170,171,172,173,174,175,52], as well as chemosensitizing [176,177,178,179,180,181,182,183,161,18], efficacy against cancer, together with anti-inflammatory, antioxidant and antibacterial activities [184,185]. We have attempted to compile the studies in which curcumin is used as a chemosensitizer [161].
4.3. Molecular targets of curcumin as a chemosensitizer
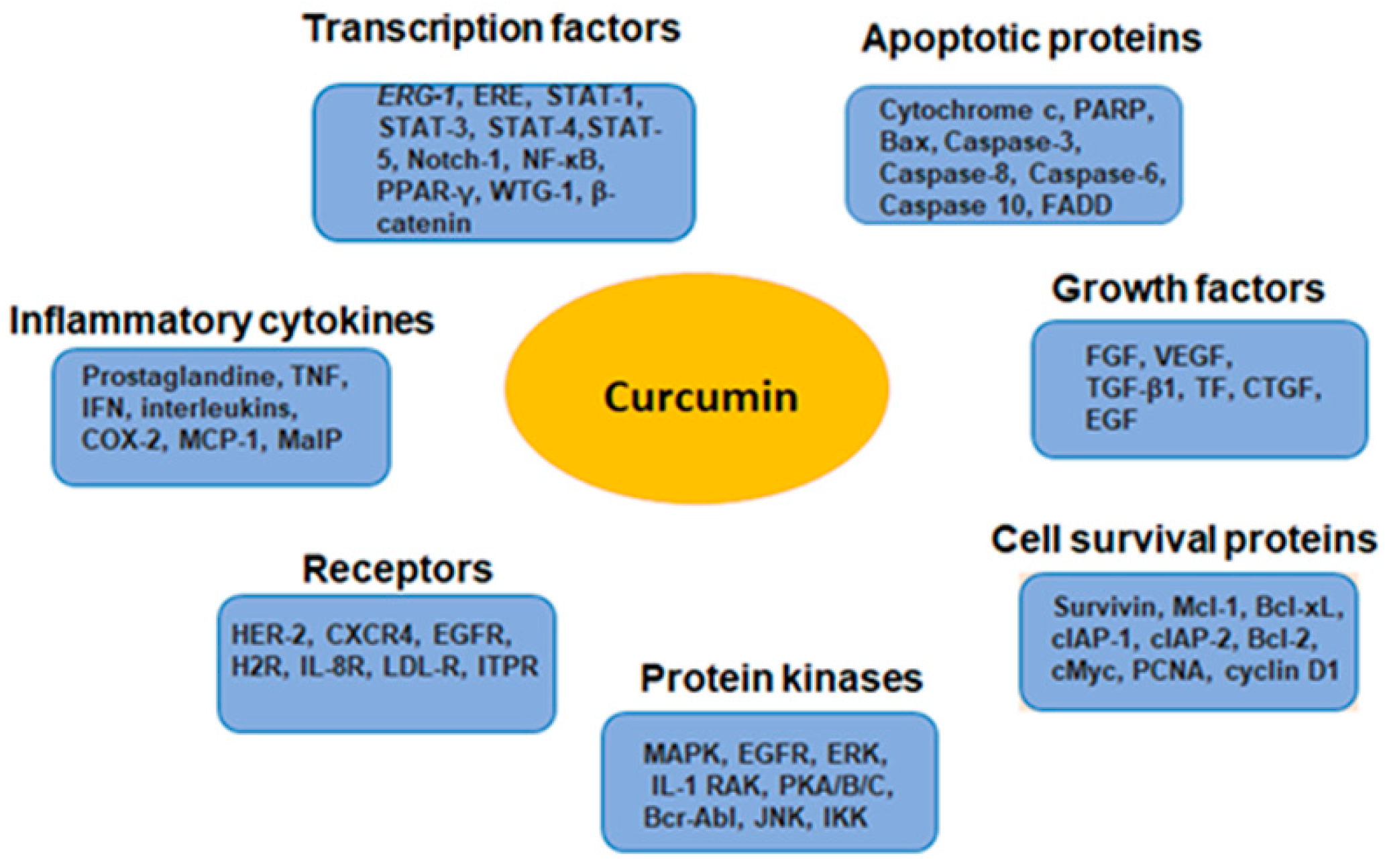
Curcumin exerts its anticancer effect through targeting various regulatory molecules, including protein kinases, transcription factors, receptors, enzymes, growth factors, cell cycle, and apoptosis-related molecules, as well as miroRNAs. Several reports have shown the effect of curcumin on variety of key molecular signaling pathways such as NF-ĸB, MAPK, PI3K/Akt/mTOR, JAK/STAT, Wnt/β-catenin etc. [186,187]. It also possesses modulatory effects on apoptotic, metastatic and cell cycle pathways involved in cancer development and progression [177]. The major molecular targets that regulate the chemosensitizing efficacy of curcumin are depicted in Figure 2.
Figure 2.
Pleiotropic effects of curcumin relevant for its chemosensitizing activity.
4.4. Curcumin as a chemosensitizer in adjuvant chemotherapy
Several in vitro, in vivo and clinical trials have shown the chemosensitizing efficacy of curcumin in combination with current chemotherapeutic drugs. Curcumin is reported to potentiate the antitumor effects gemcitabine via down-regulation of COX-2 and phospho-extracellular signal-regulated kinase1/2 (ERK1/2) levels in pancreatic adenocarcinoma cells [188], as well as through the inhibition of gemcitabine-induced NF-κB and its downstream targets, in an orthotopic model of pancreatic cancer [189]. A phase II clinical trial also evaluated the effectiveness of the combination [190]. Studies have shown that curcumin sensitize breast cancer cells to 5-FU-mediated chemotherapy through the inhibition of 5-FU-induced up-regulation of thymidylate synthase (TS) both in vitro [191] and in vivo [178], irrespective of the receptor status. It was also shown that the antitumor effects of paclitaxel could be enhanced by curcumin in cervical cancer cells through the down-regulation of paclitaxel-induced activation of NF-κB, Akt, and Bcl-2 [18,182,183]. The ability of curcumin to augment the antitumor effect of capecitabine in human colorectal cancer by modulating cyclin D1, COX-2, matrix metallopeptidase 9 (MMP-9), VEGF, and C-X-C chemokine receptor type 4 (CXCR4), has been assessed by using an orthotopic mouse model [192]. Curcumin has been also shown to sensitize prostate cancer cells to the cytotoxic effect of 5-FU, through a tp53-independent cell-cycle arrest and the downregulation of constitutive NF-κB activation [193]. Curcumin enhances the cytotoxic effect of 5-FU and oxaliplatin in colon cancer cells through the downregulation of COX-2 and the modulation of EGFR and insulin-like growth factor 1 receptor (IGF-1R) [194]. Both in vitro [195] and in vivo [196] data shows that curcumin-mediated inhibition of NF-κB activation enhances the sensitivity of prostate cancer cells to TRAIL-induced apoptosis. It has been reported that the combination of curcumin and doxorubicin could enhance the sensitivity of breast cancer cells through inhibition of ABC subfamily B member 4 (ABCB4) activity [197]. Down-regulation of IAPs by curcumin has also been reported to enhance the effect of cisplatin in hepatic cancer cells [198]. Recent reports also suggested that the combination of curcumin and paclitaxel could inhibit the ALDH-1 and paclitaxel-induced Pgp-1 expression in breast cancer cells. The combination resulted in upregulation of Bax, caspase-7, and caspase-9 along with downregulation of Bcl-2 expression in treated cells [199]. Curcumin has been shown to inhibit the FA/ BRCA pathway and sensitizes ovarian cancer cells to cisplatin-induced apoptosis [200]. It has shown that inhibition of HER2 and reduction of NF-κB activation by the combination of curcumin and its derivatives with doxorubicin enhance the toxicity of doxorubicin in resistant breast cancer cells [201]. Studies have illustrated that co-treatment with curcumin and cisplatin sensitizes breast cancer cells to cisplatin through the activation of autophagy pathway. The key mechanism underlying the curcumin-mediated chemosensitization was found to be the downregulation of CCAT1 expression and inactivation of PI3K/Akt/mTOR pathway, in treated breast cancer cells [202]. Tan & Norhaizan showed that a combination of camptothecin and curcumin-loaded cationic polymeric nanoparticle increased intracellular drug concentrations and synergistic effects of the drugs in the colon-26 cells [203].
5. Discussion
The three sections included in this article, which reviewed the main recent findings reported in the literature in some major areas of this research topic, raised several questions and prospects that need to be discussed.
A first question related to the multiple signaling pathways and components of the tumor microenvironment targeted by curcuminoids is: What kind of cognitive tools would be required for understanding its complex, pleiotropic mode of action against the hallmarks of cancer? Although a wealth of data has been accumulated both in vitro and in vivo during the past decades, many points are still lacking in our vision of how the complex regulatory networks governing the tumor microenvironment functions are modulated. To give just one example, the recent discovery showed the way curcuminoids act against the deleterious effects of cancer associated fibroblasts (CAFs), in invasive cancers [11]. New developments continue to add more complexity in our understanding of the diverse functions of this TME component [204]. CAFs increase matrix stiffness, promoting EMT, and, as a conesquence, the physics of interactions at biological interfaces is likely to be changed under their influence. Cell-cell interfaces include transient interactions between mobile immune cells, but also long-lived cell-cell contacts, in which matrix stiffness may play a crucial role. However the role of water, which indeed represents itself as an active matrix, has never been considered in this scheme [205]. At the subcellular level, this important parameter is underrepresented in the molecular biology of the cell, despite major breakthroughs published in the field [206]. Fortunately, recent developments and the emergence of innovative tools applied to cancer research [207,208] could contribute in answering this fundamental question in the future. Another related observation relates to the the fact that curcuminoids, through their simultaneous action on multiple molecular targets, belong to the field of polypharmacology [209]. Thus, understanding their mode of action required a more sophisticated view than the consideration of a single signaling pathway. Many questions come with this challenge, and there is a great potential for answering them through applying new approaches [210,211,212].
A second question is: What resource would be best suited for the experimental study of curcuminoids effects against cancers? To deal with the limitations of the cancer cell lines monolayers, advances in in vitro 3D culture technologies are being continuously reported. However, even the success of organoids in cancer research [213] still raises questions related to the mode of investigation that should be applied for exploring the host’s immune response modulated by curcuminoid treatment in vivo.? There is a high level of complexity of the ecosystems represented by neoplastic tissues, comprising a heterogeneous population of tumor cells, and a multitude of immune and non-immune cells communicating through a plethora of systemic mediators, and embedded in an extracellular matrix [214]. Thus, the modelling represents an almost impossible task. This situation is even more complicated by the fact that most stromal cells are exposed to dynamic changes during malignant progression [215].
Another question is related to an increasing number of reports demonstrating the role of interactions between curcumin given orally and gut microbiota in the generation of active metabolites. This observation led to the “low bioavailability/high bioactivity paradox” that concerns not only curcuminoids but most dietary polyphenols [216]. In this new research field, enzymatic modifications of curcumin by bacteria have already been established and reviewed [217], and the consequences of the regulation of intestinal microbiota by curcumin with respect to chemotherapeutic treatment start to be investigated [218]. The question for the future of this research is: How to take advantage of this process for optimizing current therapeutic strategies when combination treatments are used?
The increasing number of reports published each year on these molecules leads to many interesting prospects that could be summarized in three main points. First, new active natural molecules belonging to this family are continuously being discovered in the world. To mention just a few, in the search for new marine anticancer drugs, and in a context of global climate change that could affect bioresources [219], the potential of new molecules extracted from marine invertebrates that have an ability to reverse the immune-escape phenotype in metastatic tumors just started [220]. With respect to the third question discussed above, new biologically active derivatives produced through microbial transformation of curcuminoids (diarylheptanoids) isolated from natural resources are also studied [221].
In parallel, new synthetic curcumin analogs are still being produced, some of them exhibiting innovative therapeutic potential against cancers. In this field, improvements are emerging in the identification of curcumin derivatives with improved binding affinities for some growth factor receptors [222]. Using another methodological approach, Pandya et al, studied the exact binding pattern that one analog of interest formed with the c-myc DNA sequence [223]. The way curcumin analogs boost the efficacy of anti-immune checkpoint inhibitors also represents a new research field under development [71].
Whether curcumin, or curcumin analogs are selected for investigations of their impact on the different populations of immune cells involved in immunosuppressive microenvironment [224], there is a growing interest for the design and use of innovative nanoformulations [225]. Finally, among phytocompounds used to design and develop new drugs, curcuminoids are also being increasingly used as models with a potential for improving our understanding of their multiple mode of actions, using computational approaches such as virtual screening, target prediction, molecular dynamic and pharmacophore modelling [226].
Author Contributions
Conceptualization, D.L.P., K.G.T., R.J.A.; writing—original draft preparation, D.L.P., K.G.T., R.J.A.; writing—review and editing, D.L.P., K.G.T., R.J.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gerelene, J. Turmeric, The Golden Spice: From Asia to Africa. Open Access Journal of Archaeology & Anthropology (OAJAA) 2020, 2. [Google Scholar] [CrossRef]
- Schraufstatter, E.; Bernt, H. Antibacterial action of curcumin and related compounds. Nature 1949, 164, 456. [Google Scholar] [CrossRef] [PubMed]
- Ally, B. Turmeric Flower as a Remedy for Gonorrhoea. Ind Med Gaz 1876, 11, 273. [Google Scholar] [PubMed]
- Kuttan, R.; Bhanumathy, P.; Nirmala, K.; George, M.C. Potential anticancer activity of turmeric (Curcuma longa). Cancer Lett 1985, 29, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Kuttan, R.; Sudheeran, P.C.; Josph, C.D. Turmeric and curcumin as topical agents in cancer therapy. Tumori 1987, 73, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Ruby, A.J.; Kuttan, G.; Babu, K.D.; Rajasekharan, K.N.; Kuttan, R. Anti-tumour and antioxidant activity of natural curcuminoids. Cancer Lett 1995, 94, 79–83. [Google Scholar] [CrossRef]
- Anto, R.J.; George, J.; Babu, K.V.; Rajasekharan, K.N.; Kuttan, R. Antimutagenic and anticarcinogenic activity of natural and synthetic curcuminoids. Mutat Res 1996, 370, 127–131. [Google Scholar] [CrossRef]
- Steward, W.P.; Gescher, A.J. Curcumin in cancer management: recent results of analogue design and clinical studies and desirable future research. Mol Nutr Food Res 2008, 52, 1005–1009. [Google Scholar] [CrossRef]
- Rodrigues, F.C.; Kumar, N.A.; Thakur, G. The potency of heterocyclic curcumin analogues: An evidence-based review. Pharmacol Res 2021, 166, 105489. [Google Scholar] [CrossRef]
- Mehta, H.J.; Patel, V.; Sadikot, R.T. Curcumin and lung cancer--a review. Target Oncol 2014, 9, 295–310. [Google Scholar] [CrossRef]
- Pouliquen, D.L.; Boissard, A.; Henry, C.; Coqueret, O.; Guette, C. Curcuminoids as Modulators of EMT in Invasive Cancers: A Review of Molecular Targets With the Contribution of Malignant Mesothelioma Studies. Front Pharmacol 2022, 13, 934534. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, R.A.; Garcia-Smith, R.; Dorsey, J.; Griffith, J.K.; Bisoffi, M.; Trujillo, K.A. Tumor necrosis factor alpha induces Warburg-like metabolism and is reversed by anti-inflammatory curcumin in breast epithelial cells. Int J Cancer 2013, 133, 2504–2510. [Google Scholar] [CrossRef] [PubMed]
- Troselj, K.G.; Samarzija, I.; Tomljanovic, M.; Kujundzic, R.N.; Dakovic, N.; Mojzes, A. Implementing Curcumin in Translational Oncology Research. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Amekyeh, H.; Alkhader, E.; Sabra, R.; Billa, N. Prospects of Curcumin Nanoformulations in Cancer Management. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, M.; van Nostrum, C.F.; Kok, R.J.; Storm, G.; Hennink, W.E.; Heger, M. Utility of Intravenous Curcumin Nanodelivery Systems for Improving In Vivo Pharmacokinetics and Anticancer Pharmacodynamics. Mol Pharm 2022, 19, 3057–3074. [Google Scholar] [CrossRef] [PubMed]
- Maleki Dizaj, S.; Alipour, M.; Dalir Abdolahinia, E.; Ahmadian, E.; Eftekhari, A.; Forouhandeh, H.; Rahbar Saadat, Y.; Sharifi, S.; Zununi Vahed, S. Curcumin nanoformulations: Beneficial nanomedicine against cancer. Phytother Res 2022, 36, 1156–1181. [Google Scholar] [CrossRef] [PubMed]
- Obeid, M.A.; Alsaadi, M.; Aljabali, A.A. Recent updates in curcumin delivery. J Liposome Res 2022, 10.1080/08982104.2022.2086567, 1-12. [CrossRef]
- Bava, S.V.; Puliyappadamba, V.T.; Deepti, A.; Nair, A.; Karunagaran, D.; Anto, R.J. Sensitization of taxol-induced apoptosis by curcumin involves down- regulation of nuclear factor-kappaB and the serine/threonine kinase Akt and is independent of tubulin polymerization. J Biol Chem 2005, 280, 6301–6308. [Google Scholar] [CrossRef]
- Horbach, S.; Halffman, W. The ghosts of HeLa: How cell line misidentification contaminates the scientific literature. PLoS One 2017, 12, e0186281. [Google Scholar] [CrossRef]
- Conney, A.H.; Lysz, T.; Ferraro, T.; Abidi, T.F.; Manchand, P.S.; Laskin, J.D.; Huang, M.T. Inhibitory effect of curcumin and some related dietary compounds on tumor promotion and arachidonic acid metabolism in mouse skin. Adv Enzyme Regul 1991, 31, 385–396. [Google Scholar] [CrossRef]
- Polasa, K.; Sesikaran, B.; Krishna, T.P.; Krishnaswamy, K. Turmeric (Curcuma longa)-induced reduction in urinary mutagens. Food Chem Toxicol 1991, 29, 699–706. [Google Scholar] [CrossRef]
- Nagabhushan, M.; Bhide, S.V. Curcumin as an inhibitor of cancer. J Am Coll Nutr 1992, 11, 192–198. [Google Scholar] [CrossRef]
- Singletary, K.; MacDonald, C.; Wallig, M.; Fisher, C. Inhibition of 7,12-dimethylbenz[a]anthracene (DMBA)-induced mammary tumorigenesis and DMBA-DNA adduct formation by curcumin. Cancer Lett 1996, 103, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Samaha, H.S.; Kelloff, G.J.; Steele, V.; Rao, C.V.; Reddy, B.S. Modulation of apoptosis by sulindac, curcumin, phenylethyl-3-methylcaffeate, and 6-phenylhexyl isothiocyanate: apoptotic index as a biomarker in colon cancer chemoprevention and promotion. Cancer Res 1997, 57, 1301–1305. [Google Scholar] [PubMed]
- Kato, K.; Ito, H.; Kamei, K.; Iwamoto, I. Stimulation of the stress-induced expression of stress proteins by curcumin in cultured cells and in rat tissues in vivo. Cell Stress Chaperones 1998, 3, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Kawamori, T.; Lubet, R.; Steele, V.E.; Kelloff, G.J.; Kaskey, R.B.; Rao, C.V.; Reddy, B.S. Chemopreventive effect of curcumin, a naturally occurring anti- inflammatory agent, during the promotion/progression stages of colon cancer. Cancer Res 1999, 59, 597–601. [Google Scholar] [PubMed]
- Khar, A.; Ali, A.M.; Pardhasaradhi, B.V.; Begum, Z.; Anjum, R. Antitumor activity of curcumin is mediated through the induction of apoptosis in AK-5 tumor cells. FEBS Lett 1999, 445, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Bhaumik, S.; Jyothi, M.D.; Khar, A. Differential modulation of nitric oxide production by curcumin in host macrophages and NK cells. FEBS Lett 2000, 483, 78–82. [Google Scholar] [CrossRef]
- Churchill, M.; Chadburn, A.; Bilinski, R.T.; Bertagnolli, M.M. Inhibition of intestinal tumors by curcumin is associated with changes in the intestinal immune cell profile. J Surg Res 2000, 89, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Bhattacharyya, S.; Choudhuri, T.; Datta, G.K.; Das, T.; Sa, G. Amelioration of immune cell number depletion and potentiation of depressed detoxification system of tumor-bearing mice by curcumin. Cancer Detect Prev 2005, 29, 470–478. [Google Scholar] [CrossRef]
- Lin, J.K.; Pan, M.H.; Lin-Shiau, S.Y. Recent studies on the biofunctions and biotransformations of curcumin. Biofactors 2000, 13, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.K. Suppression of protein kinase C and nuclear oncogene expression as possible action mechanisms of cancer chemoprevention by Curcumin. Arch Pharm Res 2004, 27, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.K.; Cha, S.H.; Jeon, H.G. Curcumin-induced histone hypoacetylation enhances caspase-3-dependent glioma cell death and neurogenesis of neural progenitor cells. Stem Cells Dev 2006, 15, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Han, R. Highlight on the studies of anticancer drugs derived from plants in China. Stem Cells 1994, 12, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Khor, T.O.; Keum, Y.S.; Lin, W.; Kim, J.H.; Hu, R.; Shen, G.; Xu, C.; Gopalakrishnan, A.; Reddy, B.; Zheng, X.; et al. Combined inhibitory effects of curcumin and phenethyl isothiocyanate on the growth of human PC-3 prostate xenografts in immunodeficient mice. Cancer Res 2006, 66, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Dorai, T.; Cao, Y.C.; Dorai, B.; Buttyan, R.; Katz, A.E. Therapeutic potential of curcumin in human prostate cancer. III. Curcumin inhibits proliferation, induces apoptosis, and inhibits angiogenesis of LNCaP prostate cancer cells in vivo. Prostate 2001, 47, 293–303. [Google Scholar] [CrossRef] [PubMed]
- LoTempio, M.M.; Veena, M.S.; Steele, H.L.; Ramamurthy, B.; Ramalingam, T.S.; Cohen, A.N.; Chakrabarti, R.; Srivatsan, E.S.; Wang, M.B. Curcumin suppresses growth of head and neck squamous cell carcinoma. Clin Cancer Res 2005, 11, 6994–7002. [Google Scholar] [CrossRef]
- Shehzad, A.; Lee, Y.S. Molecular mechanisms of curcumin action: signal transduction. Biofactors 2013, 39, 27–36. [Google Scholar] [CrossRef]
- Calabrese, V.; Bates, T.E.; Mancuso, C.; Cornelius, C.; Ventimiglia, B.; Cambria, M.T.; Di Renzo, L.; De Lorenzo, A.; Dinkova-Kostova, A.T. Curcumin and the cellular stress response in free radical-related diseases. Mol Nutr Food Res 2008, 52, 1062–1073. [Google Scholar] [CrossRef]
- Odot, J.; Albert, P.; Carlier, A.; Tarpin, M.; Devy, J.; Madoulet, C. In vitro and in vivo anti-tumoral effect of curcumin against melanoma cells. Int J Cancer 2004, 111, 381–387. [Google Scholar] [CrossRef]
- Chang, Y.F.; Chuang, H.Y.; Hsu, C.H.; Liu, R.S.; Gambhir, S.S.; Hwang, J.J. Immunomodulation of curcumin on adoptive therapy with T cell functional imaging in mice. Cancer Prev Res (Phila) 2012, 5, 444–452. [Google Scholar] [CrossRef]
- Pouliquen, D.L.; Nawrocki-Raby, B.; Nader, J.; Blandin, S.; Robard, M.; Birembaut, P.; Grégoire, M. Evaluation of intracavitary administration of curcumin for the treatment of sarcomatoid mesothelioma. Oncotarget 2017, 8, 57552–57573. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Rishi, A.K.; Wu, W.; Polin, L.; Sharma, S.; Levi, E.; Albelda, S.; Pass, H.I.; Wali, A. Curcumin suppresses growth of mesothelioma cells in vitro and in vivo, in part, by stimulating apoptosis. Mol Cell Biochem 2011, 357, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Song, R.; Shen, Y.; Sun, Y.; Gu, Y.; Shu, Y.; Xu, Q. Targeting sarcoplasmic/endoplasmic reticulum Ca(2)+-ATPase 2 by curcumin induces ER stress-associated apoptosis for treating human liposarcoma. Mol Cancer Ther 2011, 10, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiao, J.; Zhou, H.; Yang, S.; Wu, X.; Jiang, C.; Zhao, Y.; Liang, D.; Li, X.; Liang, G. A novel monocarbonyl analogue of curcumin, (1E,4E)-1,5-bis(2,3-dimethoxyphenyl)penta-1,4-dien-3-one, induced cancer cell H460 apoptosis via activation of endoplasmic reticulum stress signaling pathway. J Med Chem 2011, 54, 3768–3778. [Google Scholar] [CrossRef]
- Gao, J.; Fan, M.; Peng, S.; Zhang, M.; Xiang, G.; Li, X.; Guo, W.; Sun, Y.; Wu, X.; Wu, X.; et al. Small-molecule RL71-triggered excessive autophagic cell death as a potential therapeutic strategy in triple-negative breast cancer. Cell Death Dis 2017, 8, e3049. [Google Scholar] [CrossRef]
- Lin, Y.G.; Kunnumakkara, A.B.; Nair, A.; Merritt, W.M.; Han, L.Y.; Armaiz-Pena, G.N.; Kamat, A.A.; Spannuth, W.A.; Gershenson, D.M.; Lutgendorf, S.K.; et al. Curcumin inhibits tumor growth and angiogenesis in ovarian carcinoma by targeting the nuclear factor-kappaB pathway. Clin Cancer Res 2007, 13, 3423–3430. [Google Scholar] [CrossRef]
- Killian, P.H.; Kronski, E.; Michalik, K.M.; Barbieri, O.; Astigiano, S.; Sommerhoff, C.P.; Pfeffer, U.; Nerlich, A.G.; Bachmeier, B.E. Curcumin inhibits prostate cancer metastasis in vivo by targeting the inflammatory cytokines CXCL1 and -2. Carcinogenesis 2012, 33, 2507–2519. [Google Scholar] [CrossRef]
- Fan, S.; Xu, Y.; Li, X.; Tie, L.; Pan, Y.; Li, X. Opposite angiogenic outcome of curcumin against ischemia and Lewis lung cancer models: in silico, in vitro and in vivo studies. Biochim Biophys Acta 2014, 1842, 1742–1754. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Anand, P.; Aggarwal, B.B. Curcumin inhibits proliferation, invasion, angiogenesis and metastasis of different cancers through interaction with multiple cell signaling proteins. Cancer Lett 2008, 269, 199–225. [Google Scholar] [CrossRef]
- Zanotto-Filho, A.; Braganhol, E.; Edelweiss, M.I.; Behr, G.A.; Zanin, R.; Schroder, R.; Simoes-Pires, A.; Battastini, A.M.; Moreira, J.C. The curry spice curcumin selectively inhibits cancer cells growth in vitro and in preclinical model of glioblastoma. J Nutr Biochem 2012, 23, 591–601. [Google Scholar] [CrossRef]
- Wilken, R.; Veena, M.S.; Wang, M.B.; Srivatsan, E.S. Curcumin: A review of anti-cancer properties and therapeutic activity in head and neck squamous cell carcinoma. Mol Cancer 2011, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Borges, G.A.; Rego, D.F.; Assad, D.X.; Coletta, R.D.; De Luca Canto, G.; Guerra, E.N. In vivo and in vitro effects of curcumin on head and neck carcinoma: a systematic review. J Oral Pathol Med 2017, 46, 3–20. [Google Scholar] [CrossRef]
- Ren, M.; Wang, Y.; Wu, X.; Ge, S.; Wang, B. Curcumin synergistically increases effects of beta-interferon and retinoic acid on breast cancer cells in vitro and in vivo by up- regulation of GRIM-19 through STAT3-dependent and STAT3-independent pathways. J Drug Target 2017, 25, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhou, B.H.; Qiu, X.; Wang, H.S.; Zhang, F.; Fang, R.; Wang, X.F.; Cai, S.H.; Du, J.; Bu, X.Z. T63, a new 4-arylidene curcumin analogue, induces cell cycle arrest and apoptosis through activation of the reactive oxygen species-FOXO3a pathway in lung cancer cells. Free Radic Biol Med 2012, 53, 2204–2217. [Google Scholar] [CrossRef] [PubMed]
- Hackler, L., Jr.; Ozsvari, B.; Gyuris, M.; Sipos, P.; Fabian, G.; Molnar, E.; Marton, A.; Farago, N.; Mihaly, J.; Nagy, L.I.; et al. The Curcumin Analog C-150, Influencing NF-kappaB, UPR and Akt/Notch Pathways Has Potent Anticancer Activity In Vitro and In Vivo. PLoS One 2016, 11, e0149832. [Google Scholar] [CrossRef]
- Sundram, V.; Chauhan, S.C.; Ebeling, M.; Jaggi, M. Curcumin attenuates beta-catenin signaling in prostate cancer cells through activation of protein kinase D1. PLoS One 2012, 7, e35368. [Google Scholar] [CrossRef]
- Zheng, R.; Deng, Q.; Liu, Y.; Zhao, P. Curcumin Inhibits Gastric Carcinoma Cell Growth and Induces Apoptosis by Suppressing the Wnt/beta-Catenin Signaling Pathway. Med Sci Monit 2017, 23, 163–171. [Google Scholar] [CrossRef]
- Uehara, Y.; Inoue, M.; Fukuda, K.; Yamakoshi, H.; Hosoi, Y.; Kanda, H.; Oshima, M.; Iwabuchi, Y.; Shibata, H. Inhibition of beta-catenin and STAT3 with a curcumin analog suppresses gastric carcinogenesis in vivo. Gastric Cancer 2015, 18, 774–783. [Google Scholar] [CrossRef]
- Alexandrow, M.G.; Song, L.J.; Altiok, S.; Gray, J.; Haura, E.B.; Kumar, N.B. Curcumin: a novel Stat3 pathway inhibitor for chemoprevention of lung cancer. Eur J Cancer Prev 2012, 21, 407–412. [Google Scholar] [CrossRef]
- Zhang, W.; Guo, J.; Li, S.; Ma, T.; Xu, D.; Han, C.; Liu, F.; Yu, W.; Kong, L. Discovery of monocarbonyl curcumin-BTP hybrids as STAT3 inhibitors for drug-sensitive and drug-resistant breast cancer therapy. Sci Rep 2017, 7, 46352. [Google Scholar] [CrossRef]
- Ye, M.; Zhang, J.; Zhang, J.; Miao, Q.; Yao, L.; Zhang, J. Curcumin promotes apoptosis by activating the p53-miR-192-5p/215-XIAP pathway in non-small cell lung cancer. Cancer Lett 2015, 357, 196–205. [Google Scholar] [CrossRef]
- Hu, A.; Huang, J.J.; Zhang, J.F.; Dai, W.J.; Li, R.L.; Lu, Z.Y.; Duan, J.L.; Li, J.P.; Chen, X.P.; Fan, J.P.; et al. Curcumin induces G2/M cell cycle arrest and apoptosis of head and neck squamous cell carcinoma in vitro and in vivo through ATM/Chk2/p53-dependent pathway. Oncotarget 2017, 8, 50747–50760. [Google Scholar] [CrossRef]
- Milacic, V.; Banerjee, S.; Landis-Piwowar, K.R.; Sarkar, F.H.; Majumdar, A.P.; Dou, Q.P. Curcumin inhibits the proteasome activity in human colon cancer cells in vitro and in vivo. Cancer Res 2008, 68, 7283–7292. [Google Scholar] [CrossRef]
- Yu, J.; Peng, Y.; Wu, L.C.; Xie, Z.; Deng, Y.; Hughes, T.; He, S.; Mo, X.; Chiu, M.; Wang, Q.E.; et al. Curcumin down-regulates DNA methyltransferase 1 and plays an anti- leukemic role in acute myeloid leukemia. PLoS One 2013, 8, e55934. [Google Scholar] [CrossRef]
- Bianchi, G.; Ravera, S.; Traverso, C.; Amaro, A.; Piaggio, F.; Emionite, L.; Bachetti, T.; Pfeffer, U.; Raffaghello, L. Curcumin induces a fatal energetic impairment in tumor cells in vitro and in vivo by inhibiting ATP-synthase activity. Carcinogenesis 2018, 39, 1141–1150. [Google Scholar] [CrossRef]
- Liu, L.; Lim, M.A.; Jung, S.N.; Oh, C.; Won, H.R.; Jin, Y.L.; Piao, Y.; Kim, H.J.; Chang, J.W.; Koo, B.S. The effect of Curcumin on multi-level immune checkpoint blockade and T cell dysfunction in head and neck cancer. Phytomedicine 2021, 92, 153758. [Google Scholar] [CrossRef]
- Kotting, C.; Hofmann, L.; Lotfi, R.; Engelhardt, D.; Laban, S.; Schuler, P.J.; Hoffmann, T.K.; Brunner, C.; Theodoraki, M.N. Immune-Stimulatory Effects of Curcumin on the Tumor Microenvironment in Head and Neck Squamous Cell Carcinoma. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Miles, G.J.; Demetriou, C.; Sidat, Z.; Foreman, N.; West, K.; Karmokar, A.; Howells, L.; Pritchard, C.; Thomas, A.L.; et al. Ex vivo explant model of adenoma and colorectal cancer to explore mechanisms of action and patient response to cancer prevention therapies. Mutagenesis 2022, 37, 227–237. [Google Scholar] [CrossRef]
- Tian, S.; Liao, L.; Zhou, Q.; Huang, X.; Zheng, P.; Guo, Y.; Deng, T.; Tian, X. Curcumin inhibits the growth of liver cancer by impairing myeloid-derived suppressor cells in murine tumor tissues. Oncol Lett 2021, 21, 286. [Google Scholar] [CrossRef] [PubMed]
- MaruYama, T.; Kobayashi, S.; Nakatsukasa, H.; Moritoki, Y.; Taguchi, D.; Sunagawa, Y.; Morimoto, T.; Asao, A.; Jin, W.; Owada, Y.; et al. The Curcumin Analog GO-Y030 Controls the Generation and Stability of Regulatory T Cells. Front Immunol 2021, 12, 687669. [Google Scholar] [CrossRef]
- Pouliquen, D.L.; Boissard, A.; Henry, C.; Blandin, S.; Richomme, P.; Coqueret, O.; Guette, C. Curcumin Treatment Identifies Therapeutic Targets within Biomarkers of Liver Colonization by Highly Invasive Mesothelioma Cells-Potential Links with Sarcomas. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Pouliquen, D.L.; Boissard, A.; Henry, C.; Blandin, S.; Coqueret, O.; Guette, C. Lymphoid Organ Proteomes Identify Therapeutic Efficacy Biomarkers Following the Intracavitary Administration of Curcumin in a Highly Invasive Rat Model of Peritoneal Mesothelioma. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Pouliquen, D.L.; Malloci, M.; Boissard, A.; Henry, C.; Guette, C. Proteomes of Residual Tumors in Curcumin-Treated Rats Reveal Changes in Microenvironment/Malignant Cell Crosstalk in a Highly Invasive Model of Mesothelioma. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Zhu, G.; Shen, Q.; Jiang, H.; Ji, O.; Zhu, L.; Zhang, L. Curcumin inhibited the growth and invasion of human monocytic leukaemia SHI-1 cells in vivo by altering MAPK and MMP signalling. Pharm Biol 2020, 58, 25–34. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, F.; Liao, W.; Yu, L.; Hu, Z.; Li, M.; Xia, H. Curcumin suppresses glioblastoma cell proliferation by p-AKT/mTOR pathway and increases the PTEN expression. Arch Biochem Biophys 2020, 689, 108412. [Google Scholar] [CrossRef]
- Hao, J.; Dai, X.; Gao, J.; Li, Y.; Hou, Z.; Chang, Z.; Wang, Y. Curcumin suppresses colorectal tumorigenesis via the Wnt/beta-catenin signaling pathway by downregulating Axin2. Oncol Lett 2021, 21, 186. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Zhang, Y.; Wang, Z. Curcumin suppresses the malignancy of non-small cell lung cancer by modulating the circ-PRKCA/miR-384/ITGB1 pathway. Biomed Pharmacother 2021, 138, 111439. [Google Scholar] [CrossRef]
- Zhao, J.A.; Nie, W.; Dong, L.; Liu, W.; Wei, W. A curcumin analog GL63 inhibits the malignant behaviors of hepatocellular carcinoma by inactivating the JAK2/STAT3 signaling pathway via the circular RNA zinc finger protein 83/microRNA-324-5p/cyclin-dependent kinase 16 axis. J Gastroenterol Hepatol 2021, 36, 2967–2977. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.; Tao, Z.; Zhao, L.; Zhu, Z.; Wu, W.; He, Y.; Chen, H.; Zheng, B.; Huang, X.; et al. Curcumin derivative WZ35 inhibits tumor cell growth via ROS-YAP-JNK signaling pathway in breast cancer. J Exp Clin Cancer Res 2019, 38, 460. [Google Scholar] [CrossRef]
- Tang, X.; Ding, H.; Liang, M.; Chen, X.; Yan, Y.; Wan, N.; Chen, Q.; Zhang, J.; Cao, J. Curcumin induces ferroptosis in non-small-cell lung cancer via activating autophagy. Thorac Cancer 2021, 12, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Gueguinou, M.; Ibrahim, S.; Bourgeais, J.; Robert, A.; Pathak, T.; Zhang, X.; Crottes, D.; Dupuy, J.; Ternant, D.; Monbet, V.; et al. Curcumin and NCLX inhibitors share anti-tumoral mechanisms in microsatellite-instability-driven colorectal cancer. Cell Mol Life Sci 2022, 79, 284. [Google Scholar] [CrossRef]
- Huang, X.; Liang, C.; Yang, H.; Li, X.; Deng, X.; Liang, X.; Li, L.; Huang, Z.; Lu, D.; Ma, Y.; et al. Curcumin induces apoptosis and inhibits the growth of adrenocortical carcinoma: Identification of potential candidate genes and pathways by transcriptome analysis. Oncol Lett 2021, 21, 476. [Google Scholar] [CrossRef]
- Xiu, Z.; Sun, T.; Yang, Y.; He, Y.; Yang, S.; Xue, X.; Yang, W. Curcumin Enhanced Ionizing Radiation-Induced Immunogenic Cell Death in Glioma Cells through Endoplasmic Reticulum Stress Signaling Pathways. Oxid Med Cell Longev 2022, 2022, 5424411. [Google Scholar] [CrossRef]
- Liu, L.; Fu, Y.; Zheng, Y.; Ma, M.; Wang, C. Curcumin inhibits proteasome activity in triple-negative breast cancer cells through regulating p300/miR-142-3p/PSMB5 axis. Phytomedicine 2020, 78, 153312. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, X.; Wang, Z.; Huang, L.; Meng, F.; Hu, L.; Chen, Y.; Wei, J. Anti-cancer Effects of Curcumin on Myelodysplastic Syndrome through the Inhibition of Enhancer of Zeste Homolog-2 (EZH2). Curr Cancer Drug Targets 2019, 19, 729–741. [Google Scholar] [CrossRef]
- Shin, J.W.; Chun, K.S.; Kim, D.H.; Kim, S.J.; Kim, S.H.; Cho, N.C.; Na, H.K.; Surh, Y.J. Curcumin induces stabilization of Nrf2 protein through Keap1 cysteine modification. Biochem Pharmacol 2020, 173, 113820. [Google Scholar] [CrossRef]
- Shaikh, S.B.; Najar, M.A.; Prasad, T.S.K.; Bhandary, Y.P. Comparative protein profiling reveals the inhibitory role of curcumin on IL-17A mediated minichromosome maintenance (MCM) proteins as novel putative markers for acute lung injury in vivo. Biomed Pharmacother 2021, 141, 111715. [Google Scholar] [CrossRef]
- Vishvakarma, N.K.; Kumar, A.; Singh, V.; Singh, S.M. Hyperglycemia of tumor microenvironment modulates stage-dependent tumor progression and multidrug resistance: implication of cell survival regulatory molecules and altered glucose transport. Mol Carcinog 2013, 52, 932–945. [Google Scholar] [CrossRef]
- Soni, V.K.; Mehta, A.; Ratre, Y.K.; Chandra, V.; Shukla, D.; Kumar, A.; Vishvakarma, N.K. Counteracting Action of Curcumin on High Glucose-Induced Chemoresistance in Hepatic Carcinoma Cells. Front Oncol 2021, 11, 738961. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Plas, D.R.; Rathmell, J.C.; Fox, C.J.; Harris, M.H.; Thompson, C.B. Growth factors can influence cell growth and survival through effects on glucose metabolism. Mol Cell Biol 2001, 21, 5899–5912. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.B.; Bielska, A.A. Growth factors stimulate anabolic metabolism by directing nutrient uptake. J Biol Chem 2019, 294, 17883–17888. [Google Scholar] [CrossRef]
- Feng, Y.; Xiong, Y.; Qiao, T.; Li, X.; Jia, L.; Han, Y. Lactate dehydrogenase A: A key player in carcinogenesis and potential target in cancer therapy. Cancer Med 2018, 7, 6124–6136. [Google Scholar] [CrossRef]
- Firth, J.D.; Ebert, B.L.; Ratcliffe, P.J. Hypoxic regulation of lactate dehydrogenase A. Interaction between hypoxia-inducible factor 1 and cAMP response elements. J Biol Chem 1995, 270, 21021–21027. [Google Scholar] [CrossRef]
- Cui, X.G.; Han, Z.T.; He, S.H.; Wu, X.D.; Chen, T.R.; Shao, C.H.; Chen, D.L.; Su, N.; Chen, Y.M.; Wang, T.; et al. HIF1/2alpha mediates hypoxia-induced LDHA expression in human pancreatic cancer cells. Oncotarget 2017, 8, 24840–24852. [Google Scholar] [CrossRef]
- Epstein, T.; Xu, L.; Gillies, R.J.; Gatenby, R.A. Separation of metabolic supply and demand: aerobic glycolysis as a normal physiological response to fluctuating energetic demands in the membrane. Cancer Metab 2014, 2, 7. [Google Scholar] [CrossRef]
- Fendt, S.M.; Frezza, C.; Erez, A. Targeting Metabolic Plasticity and Flexibility Dynamics for Cancer Therapy. Cancer Discov 2020, 10, 1797–1807. [Google Scholar] [CrossRef]
- de la Cruz-Lopez, K.G.; Castro-Munoz, L.J.; Reyes-Hernandez, D.O.; Garcia-Carranca, A.; Manzo-Merino, J. Lactate in the Regulation of Tumor Microenvironment and Therapeutic Approaches. Front Oncol 2019, 9, 1143. [Google Scholar] [CrossRef]
- Wittig, R.; Coy, J.F. The role of glucose metabolism and glucose-associated signalling in cancer. Perspect Medicin Chem 2008, 1, 64–82. [Google Scholar] [CrossRef]
- Troselj, K.G.; Kujundzic, R.N. Curcumin in combined cancer therapy. Curr Pharm Des 2014, 20, 6682–6696. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Kania, K.D.; Blauz, A.; Ciszewski, W.M. The lactate receptor (HCAR1/GPR81) contributes to doxorubicin chemoresistance via ABCB1 transporter up-regulation in human cervical cancer HeLa cells. J Physiol Pharmacol 2017, 68, 555–564. [Google Scholar] [PubMed]
- Gunnink, L.K.; Alabi, O.D.; Kuiper, B.D.; Gunnink, S.M.; Schuiteman, S.J.; Strohbehn, L.E.; Hamilton, K.E.; Wrobel, K.E.; Louters, L.L. Curcumin directly inhibits the transport activity of GLUT1. Biochimie 2016, 125, 179–185. [Google Scholar] [CrossRef]
- Mittal, L.; Aryal, U.K.; Camarillo, I.G.; Raman, V.; Sundararajan, R. Effective electrochemotherapy with curcumin in MDA-MB-231-human, triple negative breast cancer cells: A global proteomics study. Bioelectrochemistry 2020, 131, 107350. [Google Scholar] [CrossRef]
- Wang, K.; Fan, H.; Chen, Q.; Ma, G.; Zhu, M.; Zhang, X.; Zhang, Y.; Yu, J. Curcumin inhibits aerobic glycolysis and induces mitochondrial-mediated apoptosis through hexokinase II in human colorectal cancer cells in vitro. Anticancer Drugs 2015, 26, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Majewski, N.; Nogueira, V.; Bhaskar, P.; Coy, P.E.; Skeen, J.E.; Gottlob, K.; Chandel, N.S.; Thompson, C.B.; Robey, R.B.; Hay, N. Hexokinase-mitochondria interaction mediated by Akt is required to inhibit apoptosis in the presence or absence of Bax and Bak. Mol Cell 2004, 16, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Ciscato, F.; Ferrone, L.; Masgras, I.; Laquatra, C.; Rasola, A. Hexokinase 2 in Cancer: A Prima Donna Playing Multiple Characters. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Li, J.; Ding, F.; Wu, G.; Yang, Q.; Sun, Y.; Zhang, Z.; Dong, T.; Tian, X. Curcumin suppresses 4-hydroxytamoxifen resistance in breast cancer cells by targeting SLUG/Hexokinase 2 pathway. Biochem Biophys Res Commun 2016, 473, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Nocito, M.C.; Avena, P.; Zavaglia, L.; De Luca, A.; Chimento, A.; Hamad, T.; La Padula, D.; Stancati, D.; Hantel, C.; Sirianni, R.; et al. Adrenocortical Carcinoma (ACC) Cells Rewire Their Metabolism to Overcome Curcumin Antitumoral Effects Opening a Window of Opportunity to Improve Treatment. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Q.; Rajadurai, P.; Abas, F.; Othman, I.; Naidu, R. Proteomic Analysis on Anti-Proliferative and Apoptosis Effects of Curcumin Analog, 1,5-bis(4-Hydroxy-3-Methyoxyphenyl)-1,4-Pentadiene-3-One-Treated Human Glioblastoma and Neuroblastoma Cells. Front Mol Biosci 2021, 8, 645856. [Google Scholar] [CrossRef]
- Monteleone, F.; Taverna, S.; Alessandro, R.; Fontana, S. SWATH-MS based quantitative proteomics analysis reveals that curcumin alters the metabolic enzyme profile of CML cells by affecting the activity of miR-22/IPO7/HIF-1alpha axis. J Exp Clin Cancer Res 2018, 37, 170. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Zhang, C.J.; Wong, Y.K.; Lim, T.K.; Hua, Z.C.; Liu, B.; Tannenbaum, S.R.; Shen, H.M.; Lin, Q. In situ Proteomic Profiling of Curcumin Targets in HCT116 Colon Cancer Cell Line. Sci Rep 2016, 6, 22146. [Google Scholar] [CrossRef]
- He, Y.; Luo, Y.; Zhang, D.; Wang, X.; Zhang, P.; Li, H.; Ejaz, S.; Liang, S. PGK1-mediated cancer progression and drug resistance. Am J Cancer Res 2019, 9, 2280–2302. [Google Scholar] [PubMed]
- Elson, D.A.; Ryan, H.E.; Snow, J.W.; Johnson, R.; Arbeit, J.M. Coordinate up-regulation of hypoxia inducible factor (HIF)-1alpha and HIF-1 target genes during multi-stage epidermal carcinogenesis and wound healing. Cancer Res 2000, 60, 6189–6195. [Google Scholar]
- Li, H.; Ko, H.P.; Whitlock, J.P. Induction of phosphoglycerate kinase 1 gene expression by hypoxia. Roles of Arnt and HIF1alpha. J Biol Chem 1996, 271, 21262–21267. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol Cancer 2019, 18, 157. [Google Scholar] [CrossRef]
- Yang, H.; Geng, Y.H.; Wang, P.; Yang, H.; Zhou, Y.T.; Zhang, H.Q.; He, H.Y.; Fang, W.G.; Tian, X.X. Extracellular ATP promotes breast cancer invasion and chemoresistance via SOX9 signaling. Oncogene 2020, 39, 5795–5810. [Google Scholar] [CrossRef]
- Yang, H.; Geng, Y.H.; Wang, P.; Zhang, H.Q.; Fang, W.G.; Tian, X.X. Extracellular ATP promotes breast cancer chemoresistance via HIF-1alpha signaling. Cell Death Dis 2022, 13, 199. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, A.; Atkin, S.L.; Majeed, M.; Sahebkar, A. Effects of curcumin on hypoxia-inducible factor as a new therapeutic target. Pharmacol Res 2018, 137, 159–169. [Google Scholar] [CrossRef]
- Hahn, Y.I.; Kim, S.J.; Choi, B.Y.; Cho, K.C.; Bandu, R.; Kim, K.P.; Kim, D.H.; Kim, W.; Park, J.S.; Han, B.W.; et al. Curcumin interacts directly with the Cysteine 259 residue of STAT3 and induces apoptosis in H-Ras transformed human mammary epithelial cells. Sci Rep 2018, 8, 6409. [Google Scholar] [CrossRef]
- Webb, B.A.; Chimenti, M.; Jacobson, M.P.; Barber, D.L. Dysregulated pH: a perfect storm for cancer progression. Nat Rev Cancer 2011, 11, 671–677. [Google Scholar] [CrossRef]
- Gerweck, L.E.; Seetharaman, K. Cellular pH gradient in tumor versus normal tissue: potential exploitation for the treatment of cancer. Cancer Res 1996, 56, 1194–1198. [Google Scholar]
- Koltai, T. Cancer: fundamentals behind pH targeting and the double-edged approach. Onco Targets Ther 2016, 9, 6343–6360. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-F.Z., Y-L.; Guo, D-X.; Zhao, C-J.; Yao, Y-F.; Lin, Y-Q.; Wang, S-Q. Biochemometric approach combined with 1D CSSF-TOCSY for the identification of sensitization agents in Curcuma longa L. and prediction of their action mechanisms. Microchemical Journal 2022, 181, 107727. [CrossRef]
- Dong, S.; Liang, S.; Cheng, Z.; Zhang, X.; Luo, L.; Li, L.; Zhang, W.; Li, S.; Xu, Q.; Zhong, M.; et al. ROS/PI3K/Akt and Wnt/beta-catenin signalings activate HIF-1alpha-induced metabolic reprogramming to impart 5-fluorouracil resistance in colorectal cancer. J Exp Clin Cancer Res 2022, 41, 15. [Google Scholar] [CrossRef]
- Kim, S.W.; Cha, M.J.; Lee, S.K.; Song, B.W.; Jin, X.; Lee, J.M.; Park, J.H.; Lee, J.D. Curcumin Treatment in Combination with Glucose Restriction Inhibits Intracellular Alkalinization and Tumor Growth in Hepatoma Cells. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Persi, E.; Duran-Frigola, M.; Damaghi, M.; Roush, W.R.; Aloy, P.; Cleveland, J.L.; Gillies, R.J.; Ruppin, E. Systems analysis of intracellular pH vulnerabilities for cancer therapy. Nat Commun 2018, 9, 2997. [Google Scholar] [CrossRef]
- Patra, K.C.; Hay, N. The pentose phosphate pathway and cancer. Trends Biochem Sci 2014, 39, 347–354. [Google Scholar] [CrossRef]
- Kuehne, A.; Emmert, H.; Soehle, J.; Winnefeld, M.; Fischer, F.; Wenck, H.; Gallinat, S.; Terstegen, L.; Lucius, R.; Hildebrand, J.; et al. Acute Activation of Oxidative Pentose Phosphate Pathway as First-Line Response to Oxidative Stress in Human Skin Cells. Mol Cell 2015, 59, 359–371. [Google Scholar] [CrossRef]
- Giacomini, I.; Ragazzi, E.; Pasut, G.; Montopoli, M. The Pentose Phosphate Pathway and Its Involvement in Cisplatin Resistance. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Catanzaro, D.; Gaude, E.; Orso, G.; Giordano, C.; Guzzo, G.; Rasola, A.; Ragazzi, E.; Caparrotta, L.; Frezza, C.; Montopoli, M. Inhibition of glucose-6-phosphate dehydrogenase sensitizes cisplatin- resistant cells to death. Oncotarget 2015, 6, 30102–30114. [Google Scholar] [CrossRef]
- Hong, W.; Cai, P.; Xu, C.; Cao, D.; Yu, W.; Zhao, Z.; Huang, M.; Jin, J. Inhibition of Glucose-6-Phosphate Dehydrogenase Reverses Cisplatin Resistance in Lung Cancer Cells via the Redox System. Front Pharmacol 2018, 9, 43. [Google Scholar] [CrossRef]
- Iqbal, M.; Sharma, S.D.; Okazaki, Y.; Fujisawa, M.; Okada, S. Dietary supplementation of curcumin enhances antioxidant and phase II metabolizing enzymes in ddY male mice: possible role in protection against chemical carcinogenesis and toxicity. Pharmacol Toxicol 2003, 92, 33–38. [Google Scholar] [CrossRef]
- Li, L.; Wei, Y.; To, C.; Zhu, C.Q.; Tong, J.; Pham, N.A.; Taylor, P.; Ignatchenko, V.; Ignatchenko, A.; Zhang, W.; et al. Integrated omic analysis of lung cancer reveals metabolism proteome signatures with prognostic impact. Nat Commun 2014, 5, 5469. [Google Scholar] [CrossRef]
- Chaneton, B.; Hillmann, P.; Zheng, L.; Martin, A.C.L.; Maddocks, O.D.K.; Chokkathukalam, A.; Coyle, J.E.; Jankevics, A.; Holding, F.P.; Vousden, K.H.; et al. Serine is a natural ligand and allosteric activator of pyruvate kinase M2. Nature 2012, 491, 458–462. [Google Scholar] [CrossRef]
- Abegg, D.; Frei, R.; Cerato, L.; Prasad Hari, D.; Wang, C.; Waser, J.; Adibekian, A. Proteome-Wide Profiling of Targets of Cysteine reactive Small Molecules by Using Ethynyl Benziodoxolone Reagents. Angew Chem Int Ed Engl 2015, 54, 10852–10857. [Google Scholar] [CrossRef] [PubMed]
- Kujundžić, R.N.; Stepanić, V.; Milković, L.; Gašparović, A.; Tomljanović, M.; Trošelj, K.G. Curcumin and its Potential for Systemic Targeting of Inflamm-Aging and Metabolic Reprogramming in Cancer. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Pranzini, E.; Pardella, E.; Muccillo, L.; Leo, A.; Nesi, I.; Santi, A.; Parri, M.; Zhang, T.; Uribe, A.H.; Lottini, T.; et al. SHMT2-mediated mitochondrial serine metabolism drives 5-FU resistance by fueling nucleotide biosynthesis. Cell Rep 2022, 40, 111233. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, F.; Wu, X.R. Inhibition of Pyruvate Kinase M2 Markedly Reduces Chemoresistance of Advanced Bladder Cancer to Cisplatin. Sci Rep 2017, 7, 45983. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wu, J.; Zhang, W.; Luo, H.; Shen, Z.; Cheng, H.; Zhu, X. PKM2 enhances chemosensitivity to cisplatin through interaction with the mTOR pathway in cervical cancer. Sci Rep 2016, 6, 30788. [Google Scholar] [CrossRef]
- Lin, Y.; Lv, F.; Liu, F.; Guo, X.; Fan, Y.; Gu, F.; Gu, J.; Fu, L. High Expression of Pyruvate Kinase M2 is Associated with Chemosensitivity to Epirubicin and 5-Fluorouracil in Breast Cancer. J Cancer 2015, 6, 1130–1139. [Google Scholar] [CrossRef]
- Su, Q.; Luo, S.; Tan, Q.; Deng, J.; Zhou, S.; Peng, M.; Tao, T.; Yang, X. The role of pyruvate kinase M2 in anticancer therapeutic treatments. Oncol Lett 2019, 18, 5663–5672. [Google Scholar] [CrossRef]
- Siddiqui, F.A.; Prakasam, G.; Chattopadhyay, S.; Rehman, A.U.; Padder, R.A.; Ansari, M.A.; Irshad, R.; Mangalhara, K.; Bamezai, R.N.K.; Husain, M.; et al. Curcumin decreases Warburg effect in cancer cells by down-regulating pyruvate kinase M2 via mTOR-HIF1alpha inhibition. Sci Rep 2018, 8, 8323. [Google Scholar] [CrossRef] [PubMed]
- Mojzeš, A.; Tomljanović, M.; Milković, L.; Kujundžić, R.N.; Gašparović, A.; Trošelj, K.G. Cell-Type Specific Metabolic Response of Cancer Cells to Curcumin. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Yadav, S.; Bhagat, S.D.; Gupta, A.; Samaiya, A.; Srivastava, A.; Shukla, S. Dietary-phytochemical mediated reversion of cancer-specific splicing inhibits Warburg effect in head and neck cancer. BMC Cancer 2019, 19, 1031. [Google Scholar] [CrossRef]
- Locasale, J.W.; Grassian, A.R.; Melman, T.; Lyssiotis, C.A.; Mattaini, K.R.; Bass, A.J.; Heffron, G.; Metallo, C.M.; Muranen, T.; Sharfi, H.; et al. Phosphoglycerate dehydrogenase diverts glycolytic flux and contributes to oncogenesis. Nat Genet 2011, 43, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Spillier, Q.; Frederick, R. Phosphoglycerate dehydrogenase (PHGDH) inhibitors: a comprehensive review 2015-2020. Expert Opin Ther Pat 2021, 31, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Ngo, B.; Kim, E.; Osorio-Vasquez, V.; Doll, S.; Bustraan, S.; Liang, R.J.; Luengo, A.; Davidson, S.M.; Ali, A.; Ferraro, G.B.; et al. Limited Environmental Serine and Glycine Confer Brain Metastasis Sensitivity to PHGDH Inhibition. Cancer Discov 2020, 10, 1352–1373. [Google Scholar] [CrossRef] [PubMed]
- Bi, F.; An, Y.; Sun, T.; You, Y.; Yang, Q. PHGDH Is Upregulated at Translational Level and Implicated in Platin- Resistant in Ovarian Cancer Cells. Front Oncol 2021, 11, 643129. [Google Scholar] [CrossRef]
- Zou, S.; Qin, B.; Yang, Z.; Wang, W.; Zhang, J.; Zhang, Y.; Meng, M.; Feng, J.; Xie, Y.; Fang, L.; et al. CSN6 Mediates Nucleotide Metabolism to Promote Tumor Development and Chemoresistance in Colorectal Cancer. Cancer Res 2023, 83, 414–427. [Google Scholar] [CrossRef]
- Wei, L.; Lee, D.; Law, C.T.; Zhang, M.S.; Shen, J.; Chin, D.W.; Zhang, A.; Tsang, F.H.; Wong, C.L.; Ng, I.O.; et al. Genome-wide CRISPR/Cas9 library screening identified PHGDH as a critical driver for Sorafenib resistance in HCC. Nat Commun 2019, 10, 4681. [Google Scholar] [CrossRef]
- Rathore, R.; Schutt, C.R.; Van Tine, B.A. PHGDH as a mechanism for resistance in metabolically-driven cancers. Cancer Drug Resist 2020, 3, 762–774. [Google Scholar] [CrossRef]
- Van Nyen, T.; Planque, M.; van Wagensveld, L.; Duarte, J.A.G.; Zaal, E.A.; Talebi, A.; Rossi, M.; Korner, P.R.; Rizzotto, L.; Moens, S.; et al. Serine metabolism remodeling after platinum-based chemotherapy identifies vulnerabilities in a subgroup of resistant ovarian cancers. Nat Commun 2022, 13, 4578. [Google Scholar] [CrossRef] [PubMed]
- Tajan, M.; Hennequart, M.; Cheung, E.C.; Zani, F.; Hock, A.K.; Legrave, N.; Maddocks, O.D.K.; Ridgway, R.A.; Athineos, D.; Suárez-Bonnet, A.; et al. Serine synthesis pathway inhibition cooperates with dietary serine and glycine limitation for cancer therapy. Nat Commun 2021, 12, 366. [Google Scholar] [CrossRef] [PubMed]
- Angelo, L.S.; Maxwell, D.S.; Wu, J.Y.; Sun, D.; Hawke, D.H.; McCutcheon, I.E.; Slopis, J.M.; Peng, Z.; Bornmann, W.G.; Kurzrock, R. Binding partners for curcumin in human schwannoma cells: biologic implications. Bioorg Med Chem 2013, 21, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Vie, N.; Copois, V.; Bascoul-Mollevi, C.; Denis, V.; Bec, N.; Robert, B.; Fraslon, C.; Conseiller, E.; Molina, F.; Larroque, C.; et al. Overexpression of phosphoserine aminotransferase PSAT1 stimulates cell growth and increases chemoresistance of colon cancer cells. Mol Cancer 2008, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Cui, H.; Tu, W.; Li, L.; Gao, Y.; Chen, L.; Li, D.; Chen, X.; Xu, F.; Zhou, C.; et al. An integrated pan-cancer analysis of PSAT1: A potential biomarker for survival and immunotherapy. Front Genet 2022, 13, 975381. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Zhang, L.; Chen, H.; Lei, Y.; Zhang, T.; Wang, Y.; Jin, P.; Lan, J.; Zhou, L.; Huang, Z.; et al. Regorafenib induces lethal autophagy arrest by stabilizing PSAT1 in glioblastoma. Autophagy 2020, 16, 106–122. [Google Scholar] [CrossRef]
- De Marchi, T.; Timmermans, M.A.; Sieuwerts, A.M.; Smid, M.; Look, M.P.; Grebenchtchikov, N.; Sweep, F.; Smits, J.G.; Magdolen, V.; van Deurzen, C.H.M.; et al. Phosphoserine aminotransferase 1 is associated to poor outcome on tamoxifen therapy in recurrent breast cancer. Sci Rep 2017, 7, 2099. [Google Scholar] [CrossRef]
- Liao, L.; Yu, H.; Ge, M.; Zhan, Q.; Huang, R.; Ji, X.; Liang, X.; Zhou, X. Upregulation of phosphoserine phosphatase contributes to tumor progression and predicts poor prognosis in non-small cell lung cancer patients. Thorac Cancer 2019, 10, 1203–1212. [Google Scholar] [CrossRef]
- Chiang, I.T.; Wang, W.S.; Liu, H.C.; Yang, S.T.; Tang, N.Y.; Chung, J.G. Curcumin alters gene expression-associated DNA damage, cell cycle, cell survival and cell migration and invasion in NCI-H460 human lung cancer cells in vitro. Oncol Rep 2015, 34, 1853–1874. [Google Scholar] [CrossRef]
- Vinod, B.S.; Maliekal, T.T.; Anto, R.J. Phytochemicals as chemosensitizers: from molecular mechanism to clinical significance. Antioxid Redox Signal 2013, 18, 1307–1348. [Google Scholar] [CrossRef]
- Rayan, A.; Raiyn, J.; Falah, M. Nature is the best source of anticancer drugs: Indexing natural products for their anticancer bioactivity. PLoS One 2017, 12, e0187925. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.R.; Chang, C.H.; Hsu, C.F.; Tsai, M.J.; Cheng, H.; Leong, M.K.; Sung, P.J.; Chen, J.C.; Weng, C.F. Natural compounds as potential adjuvants to cancer therapy: Preclinical evidence. Br J Pharmacol 2020, 177, 1409–1423. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Huang, P.; Law, S.; Tian, H.; Leung, W.; Xu, C. Preventive Effect of Curcumin Against Chemotherapy-Induced Side-Effects. Front Pharmacol 2018, 9, 1374. [Google Scholar] [CrossRef] [PubMed]
- Cermak, R.; Wolffram, S. The potential of flavonoids to influence drug metabolism and pharmacokinetics by local gastrointestinal mechanisms. Curr Drug Metab 2006, 7, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Karaboga Arslan, A.K.; Uzunhisarcıklı, E.; Yerer, M.B.; Bishayee, A. The golden spice curcumin in cancer: A perspective on finalized clinical trials during the last 10 years. J Cancer Res Ther 2022, 18, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, K.; Rasoulpoor, S.; Daneshkhah, A.; Abolfathi, S.; Salari, N.; Mohammadi, M.; Rasoulpoor, S.; Shabani, S. Clinical effects of curcumin in enhancing cancer therapy: A systematic review. BMC Cancer 2020, 20, 791. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Tommonaro, G. Curcumin and Cancer. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Ojo, O.A.; Adeyemo, T.R.; Rotimi, D.; Batiha, G.E.; Mostafa-Hedeab, G.; Iyobhebhe, M.E.; Elebiyo, T.C.; Atunwa, B.; Ojo, A.B.; Lima, C.M.G.; et al. Anticancer Properties of Curcumin Against Colorectal Cancer: A Review. Front Oncol 2022, 12, 881641. [Google Scholar] [CrossRef]
- Hafez Ghoran, S.; Calcaterra, A.; Abbasi, M.; Taktaz, F.; Nieselt, K.; Babaei, E. Curcumin-Based Nanoformulations: A Promising Adjuvant towards Cancer Treatment. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Hesari, A.; Azizian, M.; Sheikhi, A.; Nesaei, A.; Sanaei, S.; Mahinparvar, N.; Derakhshani, M.; Hedayt, P.; Ghasemi, F.; Mirzaei, H. Chemopreventive and therapeutic potential of curcumin in esophageal cancer: Current and future status. Int J Cancer 2019, 144, 1215–1226. [Google Scholar] [CrossRef]
- Puliyappadamba, V.T.; Thulasidasan, A.K.; Vijayakurup, V.; Antony, J.; Bava, S.V.; Anwar, S.; Sundaram, S.; Anto, R.J. Curcumin inhibits B[a]PDE-induced procarcinogenic signals in lung cancer cells, and curbs B[a]P-induced mutagenesis and lung carcinogenesis. Biofactors 2015, 41, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Parsai, S.; Keck, R.; Skrzypczak-Jankun, E.; Jankun, J. Analysis of the anticancer activity of curcuminoids, thiotryptophan and 4-phenoxyphenol derivatives. Oncol Lett 2014, 7, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Shehzad, A.; Wahid, F.; Lee, Y.S. Curcumin in cancer chemoprevention: molecular targets, pharmacokinetics, bioavailability, and clinical trials. Arch Pharm (Weinheim) 2010, 343, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Bachmeier, B.E.; Killian, P.; Pfeffer, U.; Nerlich, A.G. Novel aspects for the application of Curcumin in chemoprevention of various cancers. Front Biosci (Schol Ed) 2010, 2, 697–717. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Wu, J.; Han, S.; Liu, Y.; Su, Z.; Zhu, H.L.; Liu, H.K.; Qian, Y. Biotinylated curcumin as a novel chemosensitizer enhances naphthalimide-induced autophagic cell death in breast cancer cells. Eur J Med Chem 2022, 228, 114029. [Google Scholar] [CrossRef]
- Farghadani, R.; Naidu, R. Curcumin as an Enhancer of Therapeutic Efficiency of Chemotherapy Drugs in Breast Cancer. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Haritha, N.H.; Nawab, A.; Vijayakurup, V.; Anto, N.P.; Liju, V.B.; Alex, V.V.; Amrutha, A.N.; Aiswarya, S.U.; Swetha, M.; Vinod, B.S.; et al. Targeting Thymidylate Synthase Enhances the Chemosensitivity of Triple-Negative Breast Cancer Towards 5-FU-Based Combinatorial Therapy. Front Oncol 2021, 11, 656804. [Google Scholar] [CrossRef]
- Vijayakurup, V.; Thulasidasan, A.T.; Shankar, G.M.; Retnakumari, A.P.; Nandan, C.D.; Somaraj, J.; Antony, J.; Alex, V.V.; Vinod, B.S.; Liju, V.B.; et al. Chitosan Encapsulation Enhances the Bioavailability and Tissue Retention of Curcumin and Improves its Efficacy in Preventing B[a]P-induced Lung Carcinogenesis. Cancer Prev Res (Phila) 2019, 12, 225–236. [Google Scholar] [CrossRef]
- Thulasidasan, A.K.T.; Retnakumari, A.P.; Shankar, M.; Vijayakurup, V.; Anwar, S.; Thankachan, S.; Pillai, K.S.; Pillai, J.J.; Nandan, C.D.; Alex, V.V.; et al. Folic acid conjugation improves the bioavailability and chemosensitizing efficacy of curcumin-encapsulated PLGA-PEG nanoparticles towards paclitaxel chemotherapy. Oncotarget 2017, 8, 107374–107389. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, C.; Bao, J.; Jia, X.; Liang, Y.; Wang, X.; Chen, M.; Su, H.; Li, P.; Wan, J.B.; et al. Synergistic chemopreventive effects of curcumin and berberine on human breast cancer cells through induction of apoptosis and autophagic cell death. Sci Rep 2016, 6, 26064. [Google Scholar] [CrossRef]
- Bava, S.V.; Sreekanth, C.N.; Thulasidasan, A.K.; Anto, N.P.; Cheriyan, V.T.; Puliyappadamba, V.T.; Menon, S.G.; Ravichandran, S.D.; Anto, R.J. Akt is upstream and MAPKs are downstream of NF-κB in paclitaxel-induced survival signaling events, which are down-regulated by curcumin contributing to their synergism. Int J Biochem Cell Biol 2011, 43, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Sreekanth, C.N.; Bava, S.V.; Sreekumar, E.; Anto, R.J. Molecular evidences for the chemosensitizing efficacy of liposomal curcumin in paclitaxel chemotherapy in mouse models of cervical cancer. Oncogene 2011, 30, 3139–3152. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.H.; Alsahli, M.A.; Aly, S.M.; Khan, M.A.; Aldebasi, Y.H. Role of Curcumin in Disease Prevention and Treatment. Adv Biomed Res 2018, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Sumathi, C.S.; Rajesh, P.; Kannan, V.R. The Biological Potentialsof Indian Traditional Medicine, Curcumin for Treating Human Diseases. Cardiovasc Hematol Agents Med Chem 2017, 10.2174/1871525715666170830130555. [CrossRef]
- Banik, U.; Parasuraman, S.; Adhikary, A.K.; Othman, N.H. Curcumin: the spicy modulator of breast carcinogenesis. J Exp Clin Cancer Res 2017, 36, 98. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, F.L.; Chen, W.D. Modulation of apoptosis-related cell signalling pathways by curcumin as a strategy to inhibit tumor progression. Mol Biol Rep 2014, 41, 4583–4594. [Google Scholar] [CrossRef]
- Lev-Ari, S.; Vexler, A.; Starr, A.; Ashkenazy-Voghera, M.; Greif, J.; Aderka, D.; Ben-Yosef, R. Curcumin augments gemcitabine cytotoxic effect on pancreatic adenocarcinoma cell lines. Cancer Invest 2007, 25, 411–418. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Guha, S.; Krishnan, S.; Diagaradjane, P.; Gelovani, J.; Aggarwal, B.B. Curcumin potentiates antitumor activity of gemcitabine in an orthotopic model of pancreatic cancer through suppression of proliferation, angiogenesis, and inhibition of nuclear factor-kappaB-regulated gene products. Cancer Res 2007, 67, 3853–3861. [Google Scholar] [CrossRef]
- Kanai, M.; Yoshimura, K.; Asada, M.; Imaizumi, A.; Suzuki, C.; Matsumoto, S.; Nishimura, T.; Mori, Y.; Masui, T.; Kawaguchi, Y.; et al. A phase I/II study of gemcitabine-based chemotherapy plus curcumin for patients with gemcitabine-resistant pancreatic cancer. Cancer Chemother Pharmacol 2011, 68, 157–164. [Google Scholar] [CrossRef]
- Vinod, B.S.; Antony, J.; Nair, H.H.; Puliyappadamba, V.T.; Saikia, M.; Narayanan, S.S.; Bevin, A.; Anto, R.J. Mechanistic evaluation of the signaling events regulating curcumin-mediated chemosensitization of breast cancer cells to 5-fluorouracil. Cell Death Dis 2013, 4, e505. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Diagaradjane, P.; Anand, P.; Harikumar, K.B.; Deorukhkar, A.; Gelovani, J.; Guha, S.; Krishnan, S.; Aggarwal, B.B. Curcumin sensitizes human colorectal cancer to capecitabine by modulation of cyclin D1, COX-2, MMP-9, VEGF and CXCR4 expression in an orthotopic mouse model. Int J Cancer 2009, 125, 2187–2197. [Google Scholar] [CrossRef]
- Hour, T.C.; Chen, J.; Huang, C.Y.; Guan, J.Y.; Lu, S.H.; Pu, Y.S. Curcumin enhances cytotoxicity of chemotherapeutic agents in prostate cancer cells by inducing p21(WAF1/CIP1) and C/EBPbeta expressions and suppressing NF-kappaB activation. Prostate 2002, 51, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.B.; Sengupta, R.; Qazi, S.; Vachhani, H.; Yu, Y.; Rishi, A.K.; Majumdar, A.P. Curcumin enhances the effects of 5-fluorouracil and oxaliplatin in mediating growth inhibition of colon cancer cells by modulating EGFR and IGF-1R. Int J Cancer 2008, 122, 267–273. [Google Scholar] [CrossRef]
- Deeb, D.; Jiang, H.; Gao, X.; Hafner, M.S.; Wong, H.; Divine, G.; Chapman, R.A.; Dulchavsky, S.A.; Gautam, S.C. Curcumin sensitizes prostate cancer cells to tumor necrosis factor-related apoptosis-inducing ligand/Apo2L by inhibiting nuclear factor-kappaB through suppression of IkappaBalpha phosphorylation. Mol Cancer Ther 2004, 3, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.; Ganapathy, S.; Chen, Q.; Srivastava, R.K. Curcumin sensitizes TRAIL-resistant xenografts: molecular mechanisms of apoptosis, metastasis and angiogenesis. Mol Cancer 2008, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Fu, L.; Huang, J.; Dai, Y.; Wang, B.; Xu, G.; Wu, L.; Zhou, H. Curcumin reverses doxorubicin resistance via inhibition the efflux function of ABCB4 in doxorubicin-resistant breast cancer cells. Mol Med Rep 2019, 19, 5162–5168. [Google Scholar] [CrossRef]
- Notarbartolo, M.; Poma, P.; Perri, D.; Dusonchet, L.; Cervello, M.; D’Alessandro, N. Antitumor effects of curcumin, alone or in combination with cisplatin or doxorubicin, on human hepatic cancer cells. Analysis of their possible relationship to changes in NF-kB activation levels and in IAP gene expression. Cancer Lett 2005, 224, 53–65. [Google Scholar] [CrossRef]
- Attia, Y.M.; El-Kersh, D.M.; Ammar, R.A.; Adel, A.; Khalil, A.; Walid, H.; Eskander, K.; Hamdy, M.; Reda, N.; Mohsen, N.E.; et al. Inhibition of aldehyde dehydrogenase-1 and p-glycoprotein-mediated multidrug resistance by curcumin and vitamin D3 increases sensitivity to paclitaxel in breast cancer. Chem Biol Interact 2020, 315, 108865. [Google Scholar] [CrossRef]
- Chirnomas, D.; Taniguchi, T.; de la Vega, M.; Vaidya, A.P.; Vasserman, M.; Hartman, A.R.; Kennedy, R.; Foster, R.; Mahoney, J.; Seiden, M.V.; et al. Chemosensitization to cisplatin by inhibitors of the Fanconi anemia/BRCA pathway. Mol Cancer Ther 2006, 5, 952–961. [Google Scholar] [CrossRef]
- Meiyanto, E.; Putri, D.D.; Susidarti, R.A.; Murwanti, R.; Sardjiman; Fitriasari, A.; Husnaa, U.; Purnomo, H.; Kawaichi, M. Curcumin and its analogues (PGV-0 and PGV-1) enhance sensitivity of resistant MCF-7 cells to doxorubicin through inhibition of HER2 and NF-kB activation. Asian Pac J Cancer Prev 2014, 15, 179–184. [CrossRef]
- Xiao-ai, L.; Bei, W.; Xiao-hong, X.; Lei, P.; Bin, W.; Xiao-xue, D.; Chen-hui, Z.; Qi-wei, D. Curcumin re-sensitizes multidrug resistant (MDR) breast cancer to cisplatin through inducing autophagy by decreasing CCAT1 expression. RSC Advances 2017, 7, 33572–33579. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Curcumin Combination Chemotherapy: The Implication and Efficacy in Cancer. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Fang, Z.; Meng, Q.; Xu, J.; Wang, W.; Zhang, B.; Liu, J.; Liang, C.; Hua, J.; Zhao, Y.; Yu, X.; et al. Signaling pathways in cancer-associated fibroblasts: recent advances and future perspectives. Cancer Commun (Lond) 2023, 43, 3–41. [Google Scholar] [CrossRef] [PubMed]
- Ball, P. Water is an active matrix of life for cell and molecular biology. Proc Natl Acad Sci U S A 2017, 114, 13327–13335. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Bagchi, B. From structure and dynamics to biomolecular functions: The ubiquitous role of solvent in biology. Curr Opin Struct Biol 2022, 77, 102462. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Bolzan, I.; Dal Zilio, S.; Parisse, P.; Andolfi, L.; Lazzarino, M. Water-Air Interface to Mimic In Vitro Tumoral Cell Migration in Complex Micro-Environments. Biosensors (Basel) 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.L.; Bordallo, H.N.; Mamontov, E. Water Dynamics in Cancer Cells: Lessons from Quasielastic Neutron Scattering. Medicina (Kaunas) 2022, 58. [Google Scholar] [CrossRef] [PubMed]
- Amelio, I.; Lisitsa, A.; Knight, R.A.; Melino, G.; Antonov, A.V. Polypharmacology of Approved Anticancer Drugs. Curr Drug Targets 2017, 18, 534–543. [Google Scholar] [CrossRef]
- Bao, L.; Wang, Z.; Wu, Z.; Luo, H.; Yu, J.; Kang, Y.; Cao, D.; Hou, T. Kinome-wide polypharmacology profiling of small molecules by multi-task graph isomorphism network approach. Acta Pharm Sin B 2023, 13, 54–67. [Google Scholar] [CrossRef]
- Lamens, A.; Bajorath, J. Explaining Accurate Predictions of Multitarget Compounds with Machine Learning Models Derived for Individual Targets. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Mitchell, D.C.; Kuljanin, M.; Li, J.; Van Vranken, J.G.; Bulloch, N.; Schweppe, D.K.; Huttlin, E.L.; Gygi, S.P. A proteome-wide atlas of drug mechanism of action. Nat Biotechnol 2023, 10.1038/s41587-022-01539-0. [CrossRef]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat Rev Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef]
- de Visser, K.E.; Joyce, J.A. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell 2023, 41, 374–403. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, Q.; Li, M.; Guo, H.; Liu, W.; Wang, F.; Tian, X.; Yang, Y. Single-cell RNA-seq reveals dynamic change in tumor microenvironment during pancreatic ductal adenocarcinoma malignant progression. EBioMedicine 2021, 66, 103315. [Google Scholar] [CrossRef] [PubMed]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit Rev Food Sci Nutr 2020, 60, 626–659. [Google Scholar] [CrossRef] [PubMed]
- Pluta, R.; Januszewski, S.; Ułamek-Kozioł, M. Mutual Two-Way Interactions of Curcumin and Gut Microbiota. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Luo, W.; Chen, Q.; Chen, X.; Zhou, G.; Sun, H. Curcumin sensitizes response to cytarabine in acute myeloid leukemia by regulating intestinal microbiota. Cancer Chemother Pharmacol 2022, 89, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.M.; Jennings, L.; Thomas, O.P. Marine Biodiscovery in a Changing World. Prog Chem Org Nat Prod 2021, 116, 1–36. [Google Scholar] [CrossRef]
- Nohara, L.; Ellis, S.; Dada, S.; Saranchova, I.; Munro, L.; Pfeifer, C.; Coyle, K.; Dreier, C.; Morrice, J.R.; Shim, D.J.; et al. A NOVEL cell-based screen identifies entities from sea sponges that reverse the immune-escape phenotype of metastatic tumours. Frontiers in Pharmacology 2023, 14. [Google Scholar] [CrossRef]
- Huo, C.; Han, F.; Xiao, Y.; Kim, H.J.; Lee, I.S. Microbial Transformation of Yakuchinone A and Cytotoxicity Evaluation of Its Metabolites. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Saeed, M.E.M.; Yücer, R.; Dawood, M.; Hegazy, M.F.; Drif, A.; Ooko, E.; Kadioglu, O.; Seo, E.J.; Kamounah, F.S.; Titinchi, S.J.; et al. In Silico and In Vitro Screening of 50 Curcumin Compounds as EGFR and NF-κB Inhibitors. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Pandya, N.; Khan, E.; Jain, N.; Satham, L.; Singh, R.; Makde, R.D.; Mishra, A.; Kumar, A. Curcumin analogs exhibit anti-cancer activity by selectively targeting G-quadruplex forming c-myc promoter sequence. Biochimie 2021, 180, 205–221. [Google Scholar] [CrossRef]
- Wang, T.; Wang, J.; Jiang, H.; Ni, M.; Zou, Y.; Chen, Y.; Wu, T.; Ding, D.; Xu, H.; Li, X. Targeted regulation of tumor microenvironment through the inhibition of MDSCs by curcumin loaded self-assembled nano-filaments. Mater Today Bio 2022, 15, 100304. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, Z.; Wang, C.; Su, Q.; Song, H.; Zhang, C.; Huang, P.; Liang, X.J.; Dong, A.; Kong, D.; et al. Co-localized delivery of nanomedicine and nanovaccine augments the postoperative cancer immunotherapy by amplifying T-cell responses. Biomaterials 2020, 230, 119649. [Google Scholar] [CrossRef] [PubMed]
- Das, A.P.; Agarwal, S.M. Recent advances in the area of plant-based anti-cancer drug discovery using computational approaches. Mol Divers 2023, 10.1007/s11030-022-10590-7, 1-25. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



