
Submitted:
03 May 2023
Posted:
04 May 2023
You are already at the latest version
Abstract
Biological invasions are a human-induced environmental disturbance that can cause major changes in ecosystem structure and functioning. Located in the northeastern Mediterranean basin, the Aegean Sea is a hotspot of biological invasions. Although the presence of alien species in the Aegean has been studied and monitored, no assessment has been conducted on their cumulative impacts on native biodiversity. To address this gap and identify the most highly impacted areas and habitats and the most impactful invasive species in the Aegean, we applied the CIMPAL index, a framework developed for mapping the cumulative impacts of invasive species. Coastal habitats showed stronger impacts than the open sea. The highest CIMPAL scores were four times more frequent in the South than in the North Aegean. Shallow (0-60 m) hard substrate was the most heavily impacted habitat type, followed by soft substrates and seagrass meadows. We identified Caulerpa cylindracea, Lophocladia lallemandii, Siganus spp. and Womersleyella setacea as the most im-pactful species across their range of occurrence in the Aegean but, ranking varied depending on the habitat and impact indicator applied. Our assessment can support marine managers in prioritizing decisions and actions to control biological invasions and mitigate their impacts in the Aegean Sea.

Keywords:
Invasive species
; alien
; cryptogenic
; impacts
; CIMPAL
; biodiversity
; mapping
; Aegean
; management
1. Introduction
Human activities are causing significant changes in the oceans, leading to unprecedented disruptions in natural processes [1,2,3]. Biological invasions are regarded as one of the most severe human-induced disturbances as they reshape the structure and functioning of ecosystems worldwide [4,5]. The rate of alien species’ introductions has accelerated since the 19th century [6] and is projected to intensify further in the 21st century, particularly in Europe [7]. Even under optimistic scenarios, the impacts of alien species on biodiversity will continue to grow, with significant consequences for communities and ecosystems [8].
Biological invasions are an integral part of the Anthropocene and contribute to global change [9]. There are ways to prevent new biological invasions by managing introduction pathways and mitigate the impacts of established species through targeted actions focusing on problematic species and priority areas [5,8]. However, managing marine alien species poses several challenges, including accurately assessing their negative impacts and their magnitude [10,11]. Not all alien species are invasive (i.e., have negative impacts on biodiversity, ecosystem services, or human health). Several established alien species have positive impacts, such as providing food for native species or humans, creating new habitats, and controlling other alien species or population explosions [10,12]. The impacts of alien species can vary across space [13] or time [14] and are greatly affected by the level of co-occurrence and interaction with other human disturbances [15,16,17]. Uncertainties, such as in predicting the distribution of alien species and the factors that define habitat invasibility, vulnerability, and resilience, reduce the reliability of bioinvasion studies and obscure the clarity needed to choose specific management actions [18]. Additionally, research on biological invasions is limited by weak evidence, frail correlations, and an overall lack of experimental studies [12,19].
Despite these limitations, the utilization of impact mapping techniques offers a promising tool to tackle the crisis of biodiversity loss and prioritize relevant management actions [20,21,22]. Although these methodologies are demanding, they provide a useful way to identify areas with the highest impacts or pressures, enabling marine managers to define priorities [23,24]. Impact mapping has been successfully applied at different spatial scales, including the Mediterranean Sea, and has even been used to map the impacts of invasive species [13,25].
The Mediterranean Sea has the highest number of alien species globally [26], which has caused significant adverse impacts on native marine biodiversity [12,13]. Over 1000 marine alien species have been introduced in the Mediterranean Sea, with more than 750 having established populations [27,28]. Assessing the impacts of alien and cryptogenic species in the Mediterranean, Tsirintanis et al. [12] assessed 76 species as invasive with significant negative effects on native biodiversity, ecosystem services, and human health. Moreover, the rate of establishment of alien species in the Mediterranean basin is accelerating, with no signs of levelling off [27,29]. The eastern Mediterranean Sea is particularly susceptible to biological invasions, with most new introductions occurring there [29,30]. The region’s overall warmer climatic conditions, faster pace of sea-water temperature rise, the opening of the Suez Canal, and other human-induced disturbances enhance the introduction, establishment, and impacts of thermophilous Lessepsian immigrants on native ecosystems [31,32,33].
Located in the northeastern part of the Mediterranean Sea, the Aegean Sea is an ecoregion [34] that supports rich biodiversity but lacks effective conservation management of its marine resources [35]. As of 2019, 209 alien species have been recorded in the Aegean, of which 149 are established [36]. The Aegean also hosts 48 out of the 76 invasive species that negatively impact Mediterranean ecosystems [12], and the impact of some of these on local biodiversity has been well documented [12,19]. Accumulating evidence over the last 20 years in the area indicates an alarming increase in the abundance of records and the number of reported impactful cryptogenic and alien species (ICAS) [37].
Local climatic conditions, along with other human-activities that enhance the introduction of alien species, their natural expansion and their impacts, render the Aegean an ecoregion under siege that urgently needs effective management measures to eliminate local biodiversity loss within a proactive and adaptive systematic conservation planning approach. Regarding alien species in particular, such an approach requires continuous monitoring of alien species distribution, introduction pathways, and assessment of their impacts (negative or positive) [36,38]. Although the presence of alien species in the Aegean Sea has been continuously monitored over the past two decades [39,40] through scientific networks [41,42], citizen science initiatives [43], collections of unpublished records [44,45], and their introduction pathways have been assessed [31,46], there are no studies that address their cumulative impacts under a systematic framework.
To address this gap and assess the impacts of marine ICAS in the Aegean Sea, we used the CIMPAL index (Cumulative IMPacts of invasive ALien species), a framework developed for mapping the cumulative negative impacts of marine alien species on marine habitats [13]. Our goal was to identify highly impacted areas and habitats, rank ICAS by their impacts, and assist the prioritization of future management decisions and actions to control and mitigate marine bioinvasions in the region.
2. Materials and Methods
We used CIMPAL to evaluate and map the impact of 26 ICAS (Table 1) present in the Aegean Sea, selected based on their negative effects on marine biodiversity, as reported by Tsirintanis et al. [12], and their abundance in the study area, as documented by Ragkousis et al. [37,45]. For the purpose of the analysis, targeted ICAS were classified into three groups: invertebrates, macrophytes, and fish. The foraminifer Amphistegina lobifera and the tunicate Styela plicata were included in invertebrates (Table 1).
To assess the impact of ICAS in the Aegean Sea, we partitioned the area into 1x1 km cells and estimated the percent cover of ten broad habitat types in each cell based on habitat maps produced by Sini et al. [35] and Topouzelis et al. [47]. The ten habitat types were: (1) seagrass meadows, (2) shallow soft substrates (0-60 m depth), (3) deep soft substrates (60-200 m depth), (4) soft substrates of the dysphotic zone (deeper than 200 m), (5) shallow hard substrates (0-60 m depth), (6) deep hard substrates (60-200 m), (7) hard substrates of the dysphotic zone (deeper than 200m), (8) submarine caves, (9) coralligenous formations, and (10) pelagic habitat. For the analysis, we classified habitats with depths of 0-200 m as coastal and those deeper than 200 m as open sea habitats. The pelagic habitat was considered part of both coastal and open sea habitats and covered the entire study area.
The CIMPAL index was calculated for each 1x1 km cell of the study area using the formula . In this formula, Ai refers to the population state of species i in a given cell of the study area, standardized to range between 0 and 1. For each targeted ICAS, we estimated Ai using two approaches: (a) species distribution models (SDMs) (also known as ecological niche models or habitat suitability models) within the Aegean, as derived by Ragkousis et al. [48], and (b) ICAS presence/absence data acquired from Ragkousis et al. [37], updated with additional data from Ragkousis et al. [45]. Hj is the percent cover of habitat j in a given cell of the study area. The impact weight wi,j of ICAS i at habitat type j was estimated using an uncertainty-averse approach [13,49] (Figure 1), based on both the impact magnitude (Figure 2) according to [11,50] and the strength of the reported evidence (Figure 2) according to [19]. The impact magnitude and strength of evidence for every combination of ICAS and habitat type (Table S1) were retrieved from [12].
To rank ICAS based on their negative impacts, we estimated four different indicators: (D1) the total area of an ICAS occurrence, which was calculated as the total number of 1 x 1 km cells with at least one record of the specific ICAS; (D2) the number of cells with estimated impact score >0 for a given ICAS; (D3) the sum of impact score values of a specific ICAS across the entire study area; (D4) the average impact score across the ICAS’s range of occurrence, excluding cells with score values lower than 0.1.
3. Results
The use of CIMPAL to map the cumulative impacts of ICAS in the Aegean Sea revealed interesting spatial patterns of negative effects (Figure 3). The study area was made up of 321,346 grid cells, with a range of cumulative CIMPAL impact scores from 0 to 49.2 and a mean value of 1.35. The North Aegean covered 33% of the study area’s grid cell surface, whereas the South Aegean covered the remaining 67% (Figure 3). Most grid cells (69%) corresponded to the open sea, while coastal habitats accounted for 31% of the Aegean Sea’s surface. Coastal habitats at depths of up to 60 m displayed higher CIMPAL impact scores than deeper habitats or the open sea (Figure 3). Cumulative impacts on the open sea decreased from the north to the south (Figure 3 and Figure 4). However, high CIMPAL scores (>20) only occurred in shallow coastal ecosystems (up to 60 m depth), appeared only locally without covering extensive surface areas, and were four times more frequent (79%) in the South Aegean (Figure 3 and Figure 4).
The pelagic habitat contributed the most to the overall sum of CIMPAL scores for all grid cells in the Aegean Sea, among all impacted habitat types (Figure 5a). This was primarily due to the extensive coverage of this habitat type rather than because of the per-cell values of the CIMPAL index. The CIMPAL score for this habitat type ranged between 0 and 3.9, with a mean habitat impact value at 1.0 across the study area, ranking 4th in terms of average CIMPAL score (Figure 5b). The invasive ctenophore Mnemiopsis leidyi was the only invasive species that impacted the pelagic habitat.
Shallow hard substrates (0-60 m depth) accumulated 5% of the CIMPAL scores (Figure 5a). This habitat covered 0.1% of the study area, exhibited the highest disaggregated CIMPAL impact scores, ranging from 0 to 47.5 and had an average CIMPAL score of 10.6 ranking as the most impacted habitat type (Figure 5b). High scores appeared more frequently in the South than North Aegean (Figure 6a). In total, 17 invasive alien species impacted this habitat, with Siganus luridus, Siganus rivulatus, and C. cylindracea being the most impactful (Figure 6b).
Shallow soft substrates (0-60 m depth) covered 6.5% of the Aegean Sea’s surface and accounted for 27% of the cumulative CIMPAL impact scores (Figure 5a). The impact values for this habitat type ranged from 0 to 11.1, with a mean CIMPAL score of 4.9 ranking as the 2nd most impacted habitat type on average (Figure 5b). Scores were higher in the South than North Aegean (Figure 7a). This habitat type was impacted by eight invasive species, with the invasive green alga Caulerpa cylindracea having the highest impact score, followed by the invasive fish Lagocephalus sceleratus and the foraminifer Amphistegina lobifera (Figure 7b).
Seagrass meadows accounted for 5% of CIMPAL scores (Figure 5a) and covered 0.8% of the study area. The average CIMPAL score for seagrass meadows was 4.2 (Figure 5b), with values ranging from 0 to 11.5. The CIMPAL score exhibited increased heterogeneity across the study area, without clear geographical patterns (Figure 8a). The red algae Lophocladia lallemandii and Womersleyella setacea were the ICAS with the highest CIMPAL scores on seagrass meadows (Figure 8b).
Impacts on caves, coralligenous, and hard substrates between 60-200 m depth accounted for less than 1% (Figure 5a) of the cumulative impact score. No species impacting the rest habitat types were documented.
Invertebrates were the biotic group with the highest contribution to the cumulative ICAS impacts in the Aegean Sea, accounting for 67% of the overall CIMPAL score (Figure 9a). Nevertheless, the invasive ctenophore Mnemiopsis leidyi, which only affects the pelagic habitat, was responsible for 92% of the invertebrate CIMPAL impact score in the Aegean. Invertebrate impacts showed a decreasing CIMPAL score gradient in the open sea, from the North Aegean to the South (Figure 9b). Macrophytes accounted for 20% of the overall CIMPAL score in the Aegean (Figure 9a). Their impact did not show any strong spatial pattern across the study area, except for lower CIMPAL scores in the northeastern part of the Aegean Sea (Figure 9c). Higher CIMPAL scores (>10) were more frequent (78%) in the South than in the North Aegean. Fish accounted for 13% of the cumulative impacts (Figure 9a) and had a stronger impact on the coastal habitats of the South Aegean (Figure 9d); high CIMPAL scores (>10) were much more frequent in the South Aegean (88%).
The impact ranking of ICAS varied depending on the indicator used (Figure 10). Excluding M. leidyi from all rankings, the herbivorous fishes Siganus luridus and S. rivulatus and the lionfish Pterois miles were the top three ranking species according to D1. According to D2, the fishes Lagocephalus sceleratus, Fistularia commersonii, and Parupeneus forsskali were the top three ranking species. D3 identified the green alga Caulerpa cylindracea as the most impactful ICAS in the Aegean, with an impact score approximately three times higher than the second-ranking species, Lagocephalus sceleratus. The top 10 most impactful invasive species, according to D3, included five fish, three macrophytes, and two invertebrates (Figure 10). C. cylindracea was also ranked as the most impactful invasive species in the Aegean by D4, with a significant difference from the second-ranking Lophocladia lallemandii. D2 and D3 exhibited high similarity, with nine common species in their top ten (Figure 10) although with different rankings.
Moreover, indicator D3 was also estimated based on actual ICAS records – D3b – (Figure 11) instead of their modelled distribution – D3a – (Figure 10). D3b ranked six fish, two macrophytes, and two invertebrates in its top 10. Although both methods had seven species in common in their top 10, there were some differences in the rankings. For instance, the two invasive siganids ranked 10th and 11th in D3a, but ranked 2nd and 3rd in D3b. The invasive foraminiferan Amphistegina lobifera ranked 3rd in D3a and 12th in D3b. The lionfish P. miles ranked 19th in D3a and 10th in D3b (Figures 10 & 11).
4. Discussion
According to CIMPAL, the impact scores of ICAS in the Aegean Sea are higher in coastal habitats than in the open sea. The invasive ctenophore Mnemiopsis leidyi is the only ICAS that extends its distribution to the open sea, whereas the impacts of the other ICAS appear to be confined to coastal habitats. Even within coastal waters (0-200 m depth), ICAS impacts were higher in habitats up to 60 meters deep. In a pan-Mediterranean biodiversity assessment, Coll et al. [51] indicated that shallow coastal waters have higher biodiversity values and a higher concentration of alien species. In the application of CIMPAL at Mediterranean scale, Katsanevakis et al. [13] also found that ICAS mainly impacted coastal habitats. These results may be attributed to the fact that most alien species introduced in the Mediterranean Sea are demersal or benthic species that prefer shallower and warmer waters [31].
However, the observed patterns of some ICAS impacts may partly reflect the spatial distribution of research effort, which is primarily conducted in shallow coastal ecosystems, in habitats that are easily accessible by scientific divers or citizen scientists (e.g. rocky reefs and seagrass meadows in waters <60 m), and in areas that represent important introduction pathways (e.g., large ports) or that are systematically monitored by scientific institutes or other marine conservation agencies [13,31,37]. This is a commonly reported problem in studies assessing biodiversity in the Mediterranean Sea. Marine research in the region has traditionally been restricted to shallow waters, whereas large parts of the deep sea remain understudied [35,51]. Research effort bias may have also affected the current mapping of ICAS in the open sea. For example, the spatial distribution of Mnemiopsis leydi was mainly based on sampled data gathered during early summers from 2004-2006 and 2008. However, the sampling was only conducted in open-sea surveys in the North Aegean [52]. Consequently, the resulting habitat suitability modelling depicted its occurrence throughout the North Aegean, but its presence in the South Aegean is largely unknown. Since the species has been reported as far south as Syria and Israel [53,54], its presence in the South Aegean is possible. Furthermore, for several planktonic or hard-to-identify ICAS that are underrepresented in the respective datasets, there is high uncertainty regarding their origin, distribution, and impacts, and as a result, they often cannot even be included as alien species in official lists [18,55,56,57].
Τhe CIMPAL results revealed cumulative impacts of ICAS on coastal habitats throughout the Aegean Sea, with a higher frequency of stronger impacts in the south Aegean. Upon examining the impact maps of the distinct biotic ICAS groups (Figure 9), it became apparent that invasive fish impacts were the primary cause of the higher coastal impact scores in the south. Recent studies on the spatiotemporal distribution patterns of marine ICAS throughout the Greek Seas reported similar patterns, with a higher ICAS richness and abundance of records observed in the South Aegean areas and a corresponding decrease towards the north [31,37]. These distribution trends are believed to be related to specific species traits and introduction pathways [31,58]. For example, the majority of alien fish in the Mediterranean are thermophilous species of Indo-Pacific origin, introduced through the Suez Canal in the southeastern part of the Mediterranean [31], and thus are favored by the higher mean sea water temperatures of the southern Aegean [59].
Macrophytes impacts did not show specific, large-scale spatial patterns across the study area, possibly due to the species’ contrasting spatial distribution. Specifically, certain species, such as Codium fragile, are primarily recorded in the North Aegean, while Halophila stipulacea and Stypopodium schimperi are mainly found in the South Aegean. Other species, such as Asparagopsis spp., Caulerpa cylindracea, and Lophocladia lallemandii, are observed throughout the study area [37,48]. Katsanevakis et al. [31] also depicted a heterogenous spatial distribution of macrophyte alien species in the Aegean Sea. In that study, aquaculture and shipping were reported as the two main pathways for macrophyte introduction in the Mediterranean Sea, indicating no common thermal affinity contrary to the case of Lessepsian species. The varying patterns of macrophyte distribution and their associated impacts are more likely related to their different ecological traits and niches, such as their varying thermal preferences [60,61].
Invertebrate ICAS impacts were generally higher in coastal habitats compared to the open sea, particularly in South Aegean. Katsanevakis et al. [31] identified the Suez Canal and shipping as the two main pathways for invertebrate introductions in the Mediterranean Sea. Therefore, the pool of alien invertebrate species in the region includes both thermophilic species from the Red Sea that tend to be more abundant in the South Aegean and vessel-introduced species with various thermal preferences. The latter are often observed in or near ports [37]. Shipping and ports have been globally associated with bioinvasions [62,63], and shipping is regarded as one of the primary introduction pathways in the Mediterranean Sea [64,65,66]. Invertebrates account for a significant portion of the alien diversity within Mediterranean ports [67]. In the South Aegean Sea, the Saronikos Gulf is considered a biological invasions hotspot due to Piraeus, the largest Greek port. Several of the assessed ICAS, such as Styela plicata, Brachidontes pharaonis, Hydroides elegans, Pinctada radiata, Amathia verticillata, are found in high abundance within Mediterranean ports [64], and some of the depicted CIMPAL impacts are related to them.
The coastal habitats with the highest average cumulative impact were shallow hard and soft substrates between 0-60 m depth and seagrass meadows. Shallow hard substrates between 0-60 m depth were the most impacted habitat type. The main species responsible for the impacts on this habitat type were the invasive herbivorous fish Siganus luridus and Siganus rivulatus. Several studies in the Aegean Sea have linked the presence of Siganus spp. with adverse grazing effects on macroalgal communities [68,69,70,71] and a degraded rocky reef ecological state [72,73]. Three macroalgal ICAS follow in the list of the most impactful species on shallow hard substrates: the green alga C. cylindracea, and the red algae Lophocladia lallemandii and Asparagopsis spp. The invasive green alga impacts biodiversity in these habitats through various mechanisms such as competition, overgrowth, increased sedimentation, and ecosystem engineering [12]. The invasive red algae can adversely impact native species through competition for resources [74,75,76] or through the creation of novel habitats that alter benthic community composition to a more impoverished state [77,78]. The invasive ascidian Styela plicata is the invertebrate ICAS with the most negative impacts on this habitat type. It forms dense aggregations that can cover the available hard substrate, outcompeting native species [79].
Soft substrates between 0-60 m depth exhibited the highest sum of cumulative impacts on coastal ecosystems. Caulerpa cylindracea, the sole macrophyte among the eight ICAS impacting this habitat type, ranked first as the most impactful. This invasive green alga can form dense mats on shallow sediments, leading to major structural changes to the resident soft-bottom faunal communities and ecosystem functioning [80,81,82]. Half of the species impacting soft substrates between 0-60 m depth were fish. Lagocephalus sceleratus and Fistularia commersonnii are considered among the most impactful alien Mediterranean fish, being voracious predators that prey upon several species of the resident biota [83,84,85,86]. Ecological modelling approaches have also indicated that L. sceleratus is an important competitor of native carnivores with high retention rates [87]. Amphistegina lobifera ranked 3rd among species impacting shallow soft substrates. This invasive foraminifer can dominate sediment communities and introduce significant shifts in community composition [88,89] or even alter habitat structure by creating a novel habitat with its accumulating shells [90].
Seagrass meadows in the Aegean Sea were found to be impacted by six ICAS. Among these, the red algae L. lallemandii and Womersleyella setacea have been identified as the most impactful, forming dense epiphytic mats on Posidonia oceanica meadows and impacting their fitness [91,92,93] and the associated communities [94,95]. The pelagic habitat is exclusively impacted by Mnemiopsis leidyi. This invasive ctenophore preys opportunistically on planktivorous organisms [96,97] and can adversely impact the structure of zooplanktonic communities [98].
CIMPAL has been effectively used as an impact assessment tool for alien species in European Seas [13,99] and inland waters [100]. One of the concerns in applying the CIMPAL index is related to the quality of habitat mapping and assessments of population state [13]. In its present application for the Aegean Sea, using a 1x1 km grid and with high-resolution shallow water habitat maps was sufficient to depict spatial patterns. The impact of each studied species was assessed based on its recorded impacts within the Mediterranean [12]. The impact weights used were based on the worst documented impacts for each species, irrespective of the location and timing of the report. However, ICAS impacts vary in space and time, and the fact that no spatiotemporal impact variance has been considered may have led to an overestimation of ICAS impacts. Finally, the current impact assessment utilizes a list of impactful ICAS derived from a recent Mediterranean review [12], which refers to documented impacts in the literature; unpublished impacts may have been missed.
This study is the first comprehensive evaluation of the cumulative adverse impacts of ICAS on the marine habitats of the Aegean Sea. The EU Regulation 1143/2014 stipulates specific measures “to prevent, minimize and mitigate the adverse impact on biodiversity of the introduction and spread within the Union, both intentional and unintentional, of invasive alien species”. Article 19 of the Regulation pertains to managing widely spread invasive alien species of Union concern, and it mandates that the management measures should be proportional to the severity of the environmental impact. Greece enforced the Regulation in February 2021. It was soon realized that a national list of IAS (HELLAS-ALIENS) should be supported by a database with all important information about species origin, traits, status, habitats, pathways of introduction, potential impacts, and geographical distribution. All marine species included in HELLAS-ALIENS [101] were considered in this study. Therefore, this study provides an essential foundation for future management and mitigation actions concerning ICAS. It identifies the most heavily impacted areas, the most affected habitats, and the most impactful ICAS in the region, through the application of CIMPAL and its associated indicators.
5. Conclusions
According to the CIMPAL index, the Aegean Sea’s ecosystems are impacted by ICAS in various spatial patterns. Coastal habitats have higher cumulative impacts compared to the open sea, and the South Aegean has four times more frequent high cumulative impact scores compared to the North. The most heavily impacted habitat is shallow hard substrate (0-60 m), followed by shallow soft substrate (0-60 m) and seagrass meadows. The study identified the most impactful species both overall and by habitat.
Biological invasions have become a significant challenge for the Aegean Sea, substantially affecting its marine ecosystems. As an eastern Mediterranean ecoregion, the Aegean Sea is a biological invasions and climate change hotspot but also has invaluable biodiversity that requires conservation. This study provides an essential tool aligned with relevant European guidelines that can contribute to the prioritization of future management actions and impact mitigation measures for biological invasions in the Aegean Sea.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Matrix of impact weights wi,j for all combinations of the targeted 26 invasive alien and cryptogenic species and the 10 studied habitat types.
Author Contributions
Conceptualization: S.K.; Methodology: S.K., K.T, and M.R. Formal Analysis: K.T., M.R. and S.K.; Investigation: K.T. and S.K.; Resources: all authors; Data Curation: all authors; Writing—Original Draft Preparation: K.T., and M.S.; Writing—Review and Editing: S.K. and A.Z.; Visualization: K.T., M.R. and S.K.; Supervision: S.K.; Project Administration: S.K.; Funding Acquisition: S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Hellenic Foundation for Research and Innovation (H.F.R.I.) under the “First Call for H.F.R.I. Research Projects to support Faculty members and Researchers and the procurement of high-cost research equipment grant”, Project ALAS—“Aliens in the Aegean—a Sea under siege”; project number: HFRI-FM17-1597 [38]. Konstantinos Tsirintanis was co-financed by Greece and the European Union (European Social Fund-ESF) through the Operational Programme “Human Resources Development, Education and Lifelong Learning”, 2014-2020, in the context of the Act “Enhancing Human Resources Research Potential by undertaking a Doctoral Research” Sub-action 2: IKY Scholarship Programme for PhD candidates in the Greek Universities.
Institutional Review Board Statement
Not applicable
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank N. Koukourouvli and T. Chalazas for their assistance in GIS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Solomon, S.; Plattner, G.K.; Knutti, R.; Friedlingstein, P. Irreversible climate change due to carbon dioxide emissions. PNAS 2009, 106, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of Earth’s nitrogen cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for Policymakers. In: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.O., Roberts, D.C. Eds.; Cambridge University Press, Cambridge, UK and New York, NY, USA, 2019; pp. 3–35. [CrossRef]
- Murphy, G.E.; Romanuk, T.N. A meta-analysis of declines in local species richness from human disturbances. Ecol. Evol. 2014, 4, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; Kleunen, M.; Kühn, I.; et al. Projecting the continental accumulation of alien species through to 2050. Global Change Biol. 2021, 27, 970–982. [Google Scholar] [CrossRef]
- Essl, F.; Lenzner, B.; Bacher, S.; Bailey, S.; Capinha, C.; Daehler, C.; Dullinger, S.; Genovesi, P.; Hui, C.; Hulme, P.E; et al. Drivers of future alien species impacts: An expert-based assessment. Global Change Biol. 2020, 26, 4880–4893. [Google Scholar] [CrossRef]
- Lewis, S.L.; Maslin, M.A. Defining the Anthropocene. Nature 2015, 519, 171–180. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; Pyšek, P.; Sousa, R.; Tabacchi, E.; Vilà, M. Impacts of biological invasions: what’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Marková, Z.; Mrugała, A.; Nentwig, W. et al. A Unified Classification of Alien Species Based on the Magnitude of their Environmental Impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef]
- Tsirintanis, K.; Azzurro, E.; Crocetta, F.; Dimiza, M.; Froglia, C.; Gerovasileiou, V.; Langeneck, J.; Mancinelli, G.; Rosso, A.; Stern, N.; et al. Bioinvasion impacts on biodiversity, ecosystem services, and human health in the Mediterranean Sea. Aquat. Invasions 2022, 17, 308–352. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Tempera, F.; Teixeira, H. Mapping the impact of alien species on marine ecosystems: the Mediterranean Sea case study. Divers. Distrib. 2016, 22, 694–707. [Google Scholar] [CrossRef]
- Strayer, D.L.; Eviner, V.T.; Jeschke, J.M.; Pace, M.L. Understanding the long-term effects of species invasions. Trends Ecol. Evol. 2006, 21, 645–651. [Google Scholar] [CrossRef]
- Vergés, A.; Tomas, F.; Cebrian, E.; Ballesteros, E.; Kizilkaya, Z.; Dendrinos, P.; Karamanlidis, A.A.; Spiegel, D.; Sala, E. Tropical rabbitfish and the deforestation of a warming temperate sea. J. Ecol. 2014, 102, 1518–1527. [Google Scholar] [CrossRef]
- Hulme, P.E. Climate change and biological invasions: evidence, expectations, and response options. Biol. Rev. 2017, 92, 1297–1313. [Google Scholar] [CrossRef]
- Gissi, E.; Manea, E.; Mazaris, A.D.; Fraschetti, S.; Almpanidou, V.; Bevilacqua, S.; Coll, M.; Guarnieri, G.; Lloret-Lloret, E.; Pascual, M.; et al. A review of the combined effects of climate change and other human stressors on the marine environment. Sci Total Environ. 2021, 755, 142564. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Moustakas, A. Uncertainty in Marine Invasion Science. Front. Mar. Sci. 2018, 5, 38. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: a pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef]
- Halpern, B.; Frazier, M.; Potapenko, J.; Casey, K.S.; Koenig, K.; Longo, C.; Lowndes, J.S.; Rockwood, R.S.; Selig, E.R.; Selkoe, K.A.; et al. Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat. Commun. 2015, 6, 7615. [Google Scholar] [CrossRef] [PubMed]
- Ban, N.C.; Alidina, H.M.; Ardron, J.A. Cumulative impact mapping: advances, relevance and limitations to marine management and conservation, using Canada's Pacific waters as a case study. Mar. Policy 2010, 34, 876–886. [Google Scholar] [CrossRef]
- Halpern, B.S.; McLeod, K.L.; Rosenberg, A.A.; Crowder, L.B. Managing for cumulative impacts in ecosystem- based management through ocean zoning. Ocean Coast. Manag. 2008, 51, 203–211. [Google Scholar] [CrossRef]
- Halpern, B.S.; Fujita, R. Assumptions, challenges, and future directions in cumulative impact analysis. Ecosphere 2013, 4, 131. [Google Scholar] [CrossRef]
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; Nykjaer, L.; Rosenberg, A.A. Cumulative human impacts on Mediterranean and Black Sea marine ecosystems: assessing current pressures and opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef]
- Costello, M.J.; Dekeyzer, D.; Galil, B.S.; Hutchings, P.; Katsanevakis, S.; Pagad, S.; Robinson, T.B.; Turon, X.; Vandepitte, L.; Vanhoorne, B.; et al. Introducing the World Register of Introduced Marine Species (WRiMS). Manag. Biol. Invasions 2021, 12, 792–811. [Google Scholar] [CrossRef]
- Zenetos, A.; Albano, P.G.; López Garcia, E.; Stern, N.; Tsiamis, K.; Galanidi, M. Established non-indigenous species increased by 40% in 11 years in the Mediterranean Sea. Mediterr. Mar. Sci. 2022, 23, 196–212. [Google Scholar] [CrossRef]
- Zenetos, A.; Albano, P.G.; López Garcia, E.; Stern, N.; Tsiamis, K.; Galanidi, M. Corrigendum to the Review Article (Medit. Mar. Sci. 23/1 2022, 196-212). Mediterr. Mar. Sci. 2022, 23, 876–878. [Google Scholar] [CrossRef]
- Galanidi, M.; Zenetos, A. Data-Driven Recommendations for Establishing Threshold Values for the NIS Trend Indicator in the Mediterranean Sea. Diversity 2022, 14, 57. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Bancila, R.I.; Katsanevakis, S.; Zenetos, A. Introduced species in Mediterranean marine caves: an increasing but neglected threat. Mediterr. Mar. Sci. 2022, 23, 995–1005. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Coll, M.; Piroddi, C.; Steenbeek, J.; Ben Rais Lasram, F.; Zenetos, A.; Cardoso, A.C. Invading the Mediterranean Sea: biodiversity patterns shaped by human activities. Front. Mar. Sci. 2014, 1, 32. [Google Scholar] [CrossRef]
- Rilov, G.; Peleg, O.; Yeruham, E.; Garval, T.; Vichik, A.; Raveh, O. Alien turf: Overfishing, overgrazing and invader domination in south-eastern Levant reef ecosystems. Aquat. Conserv. Mar. Freshwater Ecosyst. 2018, 28, 351–369. [Google Scholar] [CrossRef]
- Yeruham, E.; Shpigel, M.; Abelson, A.; Rilov, G. Ocean warming and tropical invaders erode the performance of a key herbivore. Ecology 2020, 101, e02925. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef]
- Sini, M.; Katsanevakis, S.; Koukourouvli, N.; Gerovasileiou, V.; Dailianis, T.; Buhl-Mortensen, L.; Damalas, D.; Dendrinos, P.; Dimas, X.; Frantzis, A.; et al. Assembling Ecological Pieces to Reconstruct the Conservation Puzzle of the Aegean Sea. Front. Mar. Sci. 2017, 4, 347. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Zenetos, A.; Corsini-Foka, M.; Tsiamis, K. Biological Invasions in the Aegean Sea: Temporal Trends, Pathways, and Impacts. In The Handbook of Environmental Chemistry. Anagnostou, C.L., Kostianoy, A.G., Eds.; Publisher: Springer, Berlin, Heidelberg, Germany, 2020; pp. 1–34. [Google Scholar] [CrossRef]
- Ragkousis, M.; Sini, M.; Koukourouvli, N.; Zenetos, A.; Katsanevakis, S. Invading the Greek Seas: Spatiotemporal Patterns of Marine Impactful Alien and Cryptogenic Species. Diversity 2023, 15, 353. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Tsirintanis, K.; Sini, M.; Gerovasileiou, V.; Koukourouvli, N. Aliens in the Aegean - A sea under siege (ALAS). RIO 2020, 6, e53057. [Google Scholar] [CrossRef]
- Pancucci-Papadopoulou, M.; Zenetos, A.; Corsini-Foka, M.; Politou, C. Update of marine alien species in Hellenic waters. Mediterr. Mar. Sci. 2005, 6, 147–158. [Google Scholar] [CrossRef]
- Zenetos, A.; Corsini-Foka, M.; Crocetta, F.; Gerovasileiou, V.; Karachle, V.; Simboura, M.; Tsiamis, K.; Pancucci-Papadopoulou, M. Deep cleaning of alien and cryptogenic species records in the Greek Seas (2018 update). Manag. Biol. Inv. 2018, 9, 209–226. [Google Scholar] [CrossRef]
- Zenetos, A.; Arianoutsou, M.; Bazos, I.; Balopoulou, S.; Corsini-Foka, M.; Dimiza, M.; Drakopoulou, P.; Katsanevakis, S.; Kondylatos, G.; Koutsikos, N.; Kytinou, E. ELNAIS: a collaborative network on aquatic alien species in Hellas (Greece). Manag. Biol. Inv. 2015, 6, 185–196. [Google Scholar] [CrossRef]
- Karachle, P.; Corsini Foka, M.; Crocetta, F.; Dulcic, J.; Dzhembekova, N.; Galanidi, M.; Ivanova, P.; Shenkar, N.; Skolka, M.; Stefanova, E.; et al. Setting-up a billboard of marine invasive species in the ESENIAS area: current situation and future expectancies. Acta Adriat. 2017, 58, 429–458. [Google Scholar] [CrossRef]
- Giovos, I.; Kleitou, P.; Poursanidis, D.; Batjakas, I.; Bernardi, G.; Crocetta, F.; Doumpas, N.; Kalogirou, S.; Kampouris, T.E.; Keramidas, I.; et al. Citizen-science for monitoring marine invasions and stimulating public engagement: a case project from the eastern Mediterranean. Biol. Invasions 2019, 21, 3707–3721. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Poursanidis, D.; Hoffman, R.; Rizgalla, J.; Rothman, S.B.-S.; Levitt-Barmats, Y.; Hadjioannou, L.; Trkov, D.; Garmendia, J.M.; Rizzo, M.; et al. Unpublished Mediterranean records of marine alien and cryptogenic species. Bioinvasions Rec. 2020, 9, 165–182. [Google Scholar] [CrossRef]
- Ragkousis, M.; Zenetos, A.; Ben Souissi, J.; Hoffman, R.; Ghanem, R.; Taşkın, E.; Muresan, M.; Karpova, E.; Slynko, E.; Dağlı, E.; et al. Unpublished Mediterranean and Black Sea records of marine alien, cryptogenic, and neonative species. BioInvasions Rec. 2023, in press. [Google Scholar] [CrossRef]
- Zenetos, A.; Karachle, P.K.; Corsini-Foka, M.; Gerovasileiou, V.; Simboura, N.; Xentidis, N. J.; Tsiamis, K. Is the trend in new introductions of marine non-indigenous species a reliable criterion for assessing good environmental status? Τhe case study of Greece. Mediterr. Mar. Sci. 2020, 21, 775–793. [Google Scholar] [CrossRef]
- Topouzelis, K.; Makri, D.; Stoupas, N.; Papakonstantinou, A.; Katsanevakis, S. Seagrass mapping in Greek territorial waters using Landsat-8 satellite images. Int. J. Appl. Earth Obs. Geoinf. 2018, 67, 98–113. [Google Scholar] [CrossRef]
- Ragkousis, Μ.; Papazekou, M.; Sini, M.; Zenetos, A.; Mazaris, A.D.; Katsanevakis, S. Modelling the distribution of impactful alien and cryptogenic species in the Aegean Sea. Mediterr. Mar. Sci. 2023, to be submitted.
- Yemshanov, D.; Koch, F.H.; Ducey, M.; Koehler, K. Mapping ecological risks with a portfolio-based technique: incorporating uncertainty and decision-making preferences. Divers. Distrib. 2013, 19, 567–579. [Google Scholar] [CrossRef]
- Volery, L.; Blackburn, T.M.; Bertolino, S.; Evans, T.; Genovesi, P.; Kumschick, S.; Roy, H.E.; Smith, K.G.; Bacher, S. Improving the Environmental Impact Classification for Alien Taxa (EICAT): a summary of revisions to the framework and guidelines. In Frameworks used in Invasion Science. Wilson, J.R., Bacher, S., Eds.; NeoBiota 2020, 62, 547–567. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rias Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef]
- Siapatis, A.; Giannoulaki, M.; Valavanis, V.D.; Palialexis, A.; Schismenou, E.; Machias, A.; Somarakis, S. Modelling potential habitat of the invasive ctenophore Mnemiopsis leidyi in Aegean Sea. In Essential Fish Habitat Mapping in the Mediterranean. Developments in Hydrobiology. Valavanis, V.D. Eds.;, Publisher: Springer, Dordrecht, Germany, 2008, 203, pp. 281–295. [CrossRef]
- Ikhtiyar, S.; Durgham, H.; The biochemical composition of two marine gelatin species Marivagia stellata and Mnemiopsis leidyi in coastal waters of Latakia city. Al-Baath University Journal of Medical, Engineering, Basic and Applied Sciences 2020, 42.
- Galil, B.S.; Kress, N.; Shiganova, T.A. First record of Mnemiopsis leidyi A. Agassiz, 1865 (Ctenophora; Lobata; Mnemiidae) off the Mediterranean coast of Israel. Aquat. Invasions 2009, 4, 357–360. [Google Scholar] [CrossRef]
- Gómez, F. Phytoplankton invasions: Comments on the validity of categorizing the nonindigenous dinoflagellates and diatoms in European Seas. Mar. Pollut. Bull. 2008, 56, 620–628. [Google Scholar] [CrossRef]
- Gómez, F. Comments on the non-indigenous microalgae in the European seas. Mar. Pollut. Bull. 2019, 148, 1–2. [Google Scholar] [CrossRef]
- Tsiamis, K.; Azzurro, E.; Bariche, M.; Çinar, M.E.; Crocetta, F.; De Clerck, O.; Galil, B.; Gomez, F.; Hoffman, R.; Jensen, K. Prioritizing marine invasive alien species in the EU through Horizon Scanning. Aquat. Conserv. Mar. Freshwater Ecosyst. 2020, 30, 794–845. [Google Scholar] [CrossRef]
- Karachle, P.; Oikonomou, A.; Pantazi, M.; Stergiou, K.I.; Zenetos, A. Can biological traits serve as predictors for fishes’ introductions, establishment, and interactions? The Mediterranean Sea as a case study. Biology 2022, 11, 1625. [Google Scholar] [CrossRef] [PubMed]
- Androulidakis, Y.S.; Krestenitis, Y.N. Sea Surface Temperature Variability and Marine Heat Waves over the Aegean, Ionian, and Cretan Seas from 2008–2021. J. Mar. Sci. Eng. 2022, 10, 42. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Yadav, N.S.; Barak, S.; Lima, F.P.; Sapir, Y.; Winters, G. Responses of Invasive and Native Populations of the Seagrass Halophila stipulacea to Simulated Climate Change. Front. Mar. Sci. 2020, 6, 812. [Google Scholar] [CrossRef]
- Bernal-Ibáñez, A.; Gestoso, I.; Ramalhosa, P.; Campanati, C.; Cacabelos, E. Interaction of marine heatwaves and grazing on two canopy-forming algae. J. Exp. Mar. Biol. Ecol. 2022, 556, 151795. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Costello, K.E.; Lynch, S.A.; McAllen, R.; O'Riordan, R.M.; Culloty, S.C. Assessing the potential for invasive species introductions and secondary spread using vessel movements in maritime ports. Mar. Pollut. Bull. 2022, 177, 113496. [Google Scholar] [CrossRef]
- Tempesti, J.; Mangano, M.C.; Langeneck, J.; Lardicci, C.; Maltagliati, F.; Castelli, A. Nonindigenous species in Mediterranean ports: A knowledge baseline. Mar. Environ. Res. 2020, 161, 105056. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; García Raso, J.E.; Çinar, M.E.; Almogi Labin, A.; Ates, A.S.; Azzuro, E.; et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef]
- Zenetos, A.; Ovalis, P.; Giakoumi, S.; Kontadakis, C.; Lefkaditou, E.; Mpazios, G.; Simboura, N.; Tsiamis, K. Saronikos Gulf: a hotspot area for alien species in the Mediterranean Sea. BioInvasions Rec. 2020, 9, 873–889. [Google Scholar] [CrossRef]
- Zenetos, A.; Tsiamis, K.; Galanidi, M.; Carvalho, N.; Bartilotti, C.; Canning Clode, J.; Castriota, L.; Chainho, P.; Comas-González, R.; Costa, A.C.; et al. Status and Trends in the Rate of Introduction of Marine Non-Indigenous Species in European Seas. Diversity 2022, 14, 1077. [Google Scholar] [CrossRef]
- Sala, E.; Kizilkaya, Z.; Yildirim, D.; Ballesteros, E. Alien marine fishes deplete algal biomass in the eastern Mediterranean. PLoS ONE 2011, 6, e17356. [Google Scholar] [CrossRef]
- Giakoumi, S. Distribution patterns of the invasive herbivore Siganus luridus (Rüppell, 1829) and its relation to native benthic communities in the central Aegean Sea, Northeastern Mediterranean. Mar. Ecol. 2013, 35, 96–105. [Google Scholar] [CrossRef]
- Vergés, A.; Tomas, F.; Cebrian, E.; Ballesteros, E.; Kizilkaya, Z.; Dendrinos, P.; Karamanlidis, A.A.; Spiegel, D.; Sala, E. Tropical rabbitfish and the deforestation of a warming temperate sea. J. Ecol. 2014, 102, 1518–1527. [Google Scholar] [CrossRef]
- Salomidi, M.; Katsanevakis, S.; Borja, A.; Braeckman, U.; Damalas, D.; Galparsoro, I.; Mifsud, R.; Mirto, S.; Pascual, M.; Pipitone, C.; et al. Assessment of goods and services, vulnerability, and conservation status of European seabed biotopes: a stepping stone towards ecosystem-based marine spatial management. Mediterr. Mar. Sci., 2012, 13, 49–88. [Google Scholar] [CrossRef]
- Kytinou, E.; Issaris, Y.; Sini, M.; Salomidi, M.; Katsanevakis, S. ECOfast - an integrative ecological evaluation index for an ecosystem-based assessment of shallow rocky reefs. J. Environ. Manage. 2023. submitted . [Google Scholar] [CrossRef]
- Savin, A.; Sini, M.; Xynogala, I.; Vougioukalou, V.; Stamatis, K.; Noe, S.; Ragkousis, M.; Gerovasileiou, V.; Dailianis, T.; Katsanevakis, S Assessment of macroalgal communities on shallow rocky reefs in the Aegean Sea indicates an impoverished ecological status. Mediterr. Mar. Sci. 2023, in press.
- Lodola, A. Distribution and abundance of the tropical macroalgae Caulerpa racemosa var. cylindracea (Chlorophyta: Caulerpaceae) and Asparagopsis taxiformis (Rhodophyta: Bonnemaisoniaceae) in the upper infralittoral fringe of Linosa island (Pelagian Islands, Italy). Scientifica Acta 2013, 7, 3–11.
- Cebrian, E.; Tomas, F.; López-Sendino, P.; Vilà, M.; Ballesteros, E. Biodiversity influences invasion success of a facultative epiphytic seaweed in a marine forest. Biol. Invasions 2018, 20, 2839–2848. [Google Scholar] [CrossRef]
- Rueda, J.L.; Gofas, S.; Aguilar, R.; de la Torriente, A.; García Raso, J.E.; Lo Iacono, C.; Luque, Á.A.; Marina, P.; Mateo-Ramírez, Á.; Moya-Urbano, E. et al. Benthic Fauna of Littoral and Deep-Sea Habitats of the Alboran Sea: A Hotspot of Biodiversity. In: Alboran Sea - Ecosystems and Marine Resources. Báez, J.C., Vázquez, J.T., Eds.; Publisher: Springer, Cham, Germany, 2021, pp. 285–358. [CrossRef]
- Bedini, R.; Bedini, M.; Bonechi, L.; Piazzi, L. Effects of non-native turf-forming Rhodophyta on mobile macro-invertebrate assemblages in the north-western Mediterranean Sea. Mar. Biol. Res. 2015, 11, 430–437. [Google Scholar] [CrossRef]
- Mancuso, F.P.; D'Agostaro, R.; Milazzo, M.; Badalamenti, F.; Musco, L.; Mikac, B.; Brutto, S.L.; Chemello, R. The invasive seaweed Asparagopsis taxiformis erodes the habitat structure and biodiversity of native algal forests in the Mediterranean Sea. Mar. Environ. Res. 2022, 173, 105515. [Google Scholar] [CrossRef]
- Pica, D.; Bloecher, N.; Dell’Anno, A.; Bellucci, A.; Pinto, T.; Pola, L.; Puce, S. Dynamics of a biofouling community in finfish aquaculture: a case study from the South Adriatic Sea. Biofouling 2019, 35, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Argyrou, M.; Demetropoulos, A.; Hadjichristophorou, M. Expansion of the macroalga Caulerpa racemosa and changes in softbottom macrofaunal assemblages in Moni Bay, Cyprus. Oceanolog. Acta 1999, 22, 517–528. [Google Scholar] [CrossRef]
- Rizzo, L.; Pusceddu, A.; Stabili, L.; Alifano, P.; Fraschetti, S. Potential effects of an invasive seaweed (Caulerpa cylindracea, Sonder) on sedimentary organic matter and microbial metabolic activities. Sci. Rep. 2017, 7, 12113. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.; Pusceddu, A.; Bianchelli, S.; Fraschetti, S. Potentially combined effect of the invasive seaweed Caulerpa cylindracea (Sonder) and sediment deposition rates on organic matter and meiofaunal assemblages. Mar. Environ. Res. 2020, 159, 104966. [Google Scholar] [CrossRef] [PubMed]
- Kalogirou, S. Ecological characteristics of the invasive pufferfish Lagocephalus sceleratus (Gmelin, 1789) in the eastern Mediterranean Sea – a case study from Rhodes. Mediterr. Mar. Sci. 2013, 14, 251–260. [Google Scholar] [CrossRef]
- Kalogirou, S.; Corsini, M.; Kondilatos, G.; Wennhage, H. (2007) Diet of the invasive piscivorous fish Fistularia commersonii in a recently colonized area of the eastern Mediterranean. Biol. Invasions 2007, 9, 887–896. [Google Scholar] [CrossRef]
- Bariche, M.; Alwan, N.; El-Assi, H.; Zurayk, R. Diet composition of the Lessepsian bluespotted cornetfish Fistularia commersonii in the eastern Mediterranean. J. Appl. Ichthyol. 2009, 25, 460–465. [Google Scholar] [CrossRef]
- Pancucci-Papadopoulou, M.A.; Raitsos, D.E.; Corsini-Foka, M. Biological invasions and climatic warming: implications for south-eastern Aegean ecosystem functioning. J. Mar. Biol. Assoc. U. K. 2012, 92, 777–789. [Google Scholar] [CrossRef]
- Saygu, İ.; Heymans, J.J.; Fox, C.J.; Özbilgin, H.; Eryaşar, A.R.; Gökçe, G. The importance of alien species to the food web and bottom trawl fisheries of the Northeastern Mediterranean, a modelling approach. J. Mar. Syst. 2020, 202, 103253. [Google Scholar] [CrossRef]
- Caruso, A.; Cosentino, C. The first colonization of the Genus Amphistegina and other exotic benthic foraminifera of the Pelagian Islands and south-eastern Sicily (central Mediterranean Sea). Mar. Micropaleontol. 2014, 111, 38–52. [Google Scholar] [CrossRef]
- Mouanga, G.H. Impact and Range Extension of Invasive Foraminifera in the NW Mediterranean Sea. PhD Thesis, University of Bonn, Germany, 2018. [Google Scholar]
- Streftaris, N.; Zenetos, A. Alien Marine Species in the Mediterranean - the 100 ‘Worst Invasives’ and their Impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef]
- Piazzi, L.; Cinelli, F. Distribution and dominance of two introduced turf-forming macroalgae on the coast of Tuscany, Italy, Northwestern Mediterranean sea in relation to different habitats and sedimentation. Bot. Mar. 2001, 44, 509–520. [Google Scholar] [CrossRef]
- Ballesteros, E.; Cebrian, E.; Alcoverro, T. Mortality of shoots of Posidonia oceanica following meadow invasion by the red alga Lophocladia lallemandii. Bot. Mar. 2007, 50, 8–13. [Google Scholar] [CrossRef]
- Sureda, A.; Box, A.; Terrados, J.; Deudero, S.; Pons, A. Antioxidant response of the seagrass Posidonia oceanica when epiphytized by the invasive macroalgae Lophocladia lallemandii. Mar. Environ. Res. 2008, 66, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Piazzi, L.; Cinelli, F. ; Effets de l’expansion des Rhodophyceae introduites Acrothamnion preissii et Womersleyella setacea sur les communautés algales des rhizomes de Posidonia oceanica de Méditerranée occidentale. Cryptogamie Algol. 2000, 21, 291–300. [Google Scholar] [CrossRef]
- Deudero, S.; Blanco, A.; Box, A.; Mateu-Vicens, G.; Cabanellas-Reboredo, M.; Sureda, A. Interaction between the invasive macroalga Lophocladia lallemandii and the bryozoan Reteporella grimaldii at seagrass meadows: density and physiological responses. Biol. Invasions 2010, 12, 41–52. [Google Scholar] [CrossRef]
- Marchessaux, G.; Faure, V.; Chevalier, C.; Thibault, D. Refugia area for the ctenophore Mnemiopsis leidyi A. Agassiz 1865 in the Berre Lagoon (southeast France): The key to its persistence. Reg. Stud. Mar. Sci. 2020, 39, 101409. [Google Scholar] [CrossRef]
- Marchessaux, G.; Belloni, B.; Gadreaud, J.; Thibault, D. Predation assessment of the invasive ctenophore Mnemiopsis leidyi in a French Mediterranean lagoon. J. Plankton Res. 2021, 43, 161–179. [Google Scholar] [CrossRef]
- Fiori, E.; Benzi, M.; Ferrari, C.R.; Mazziotti, C. Zooplankton community structure before and after Mnemiopsis leidyi arrival. J. Plankton Res. 2019, 41, 803–820. [Google Scholar] [CrossRef]
- Korpinen, S.; Klančnik, K.; Peterlin, M.; Nurmi, M.; Laamanen, L.; Zupančič, G.; Popit, A.; Murray, C.; Harvey, T.; Andersen, J.H.; et al. Multiple pressures and their combined effects in Europe’s seas. ETC/ICM Technical Report 4/2019: European Topic Centre on Inland, Coastal and Marine waters, 2019, 164 pp.
- Magliozzi, C.; Tsiamis, K.; Vigiak, O.; Deriu, I.; Gervasini, E.; Cardoso, A.C. Assessing invasive alien species in European catchments: Distribution and impacts. Sci. Total Environ. 2020, 732, 138677. [Google Scholar] [CrossRef]
- Arianoutsou, M.; Adamopoulou, C.; Andriopoulos, P.; Bazos, I.; Christopoulou, A.; Galanidis, A.; Kalogianni, E.; Karachle, P.K.; Kokkoris, Y.; Martinou, A.F.; et al. HELLAS-ALIENS. The Invasive Alien Species of Greece: time trends, origin and pathways. NEOBIOTA, 2023, accepted.
Figure 1.
The estimation of impact weights wi,j for species i at habitat j based on impact magnitude and strength of evidence, as defined in Fig. 2 (based on Katsanevakis et al. [13,19]).
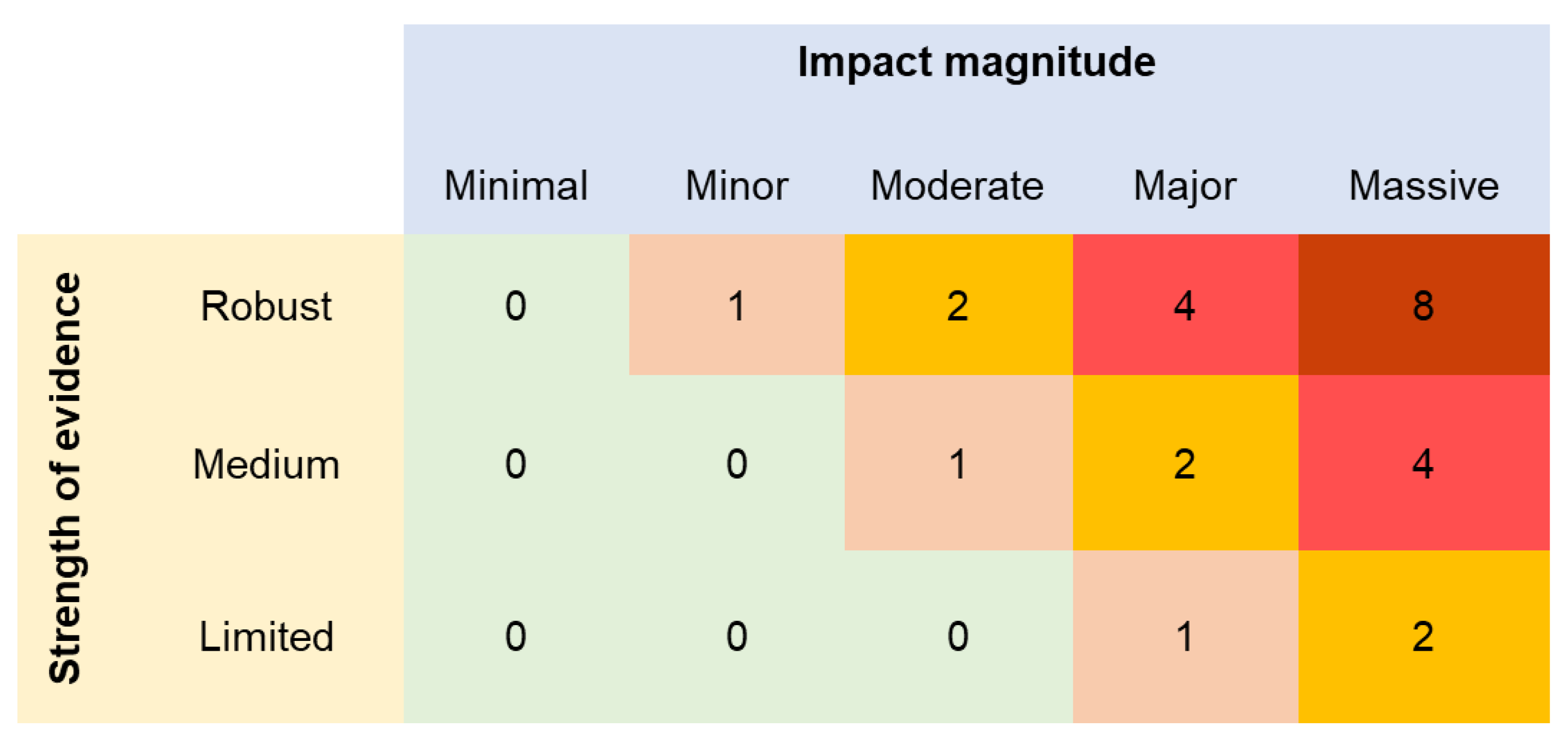
Figure 2.
Strength of evidence types (based on Katsanevakis et al., 2014) and impact magnitude categories (based on [11,50]).
Figure 3.
Mapping of ICAS cumulative impacts in the North and South Aegean Sea habitats using the CIMPAL index. Red rectangles highlight areas of the North and South Aegean illustrated in Figure 4.
Figure 3.
Mapping of ICAS cumulative impacts in the North and South Aegean Sea habitats using the CIMPAL index. Red rectangles highlight areas of the North and South Aegean illustrated in Figure 4.
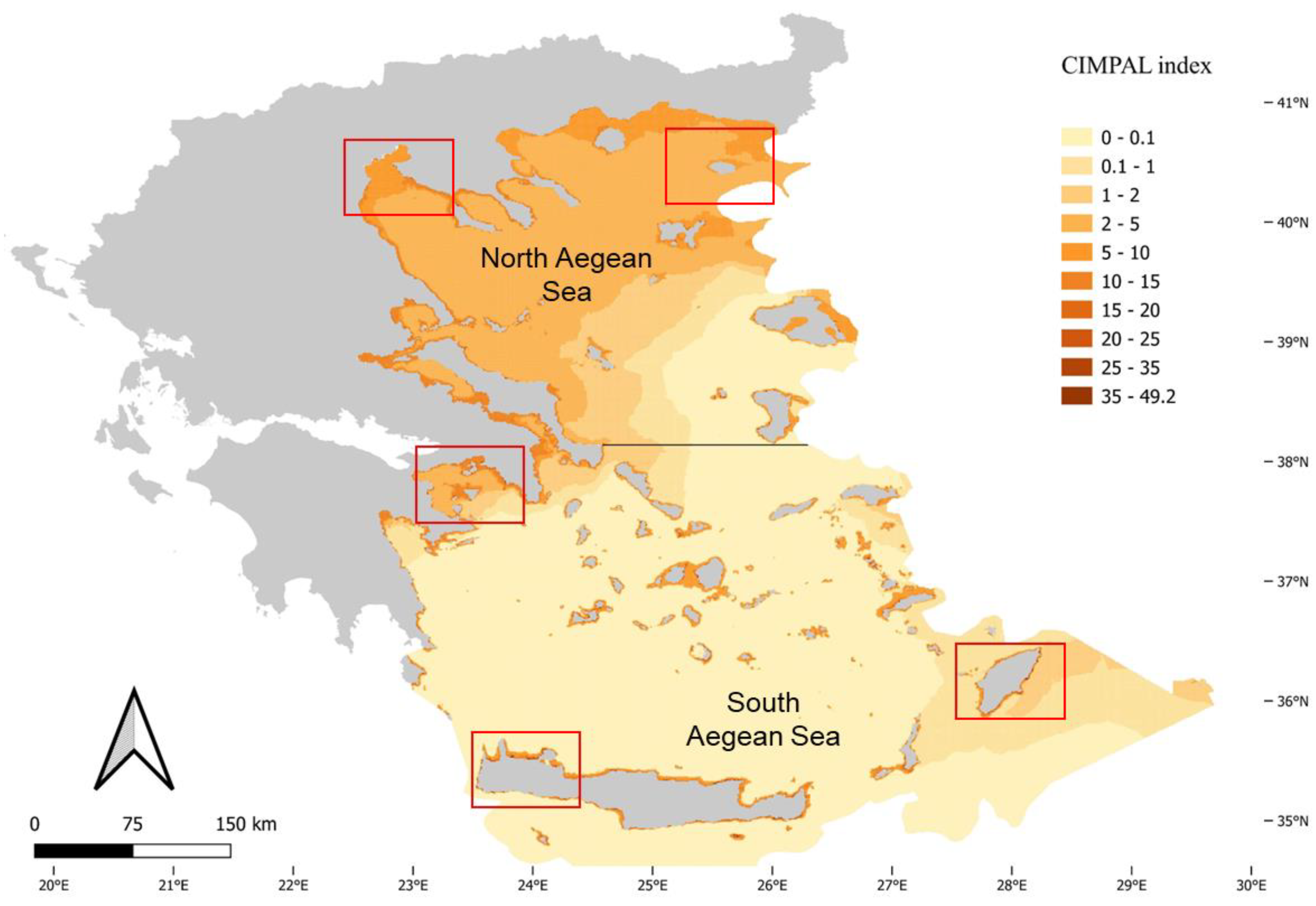
Figure 4.
Cumulative impacts on areas of the South (a, b, c) and North Aegean (d, e). These are a) Chania, Crete, b) Rhodes Island, c) Saronikos Gulf, d) Thermaikos Gulf, e) Samothrace Island.
Figure 4.
Cumulative impacts on areas of the South (a, b, c) and North Aegean (d, e). These are a) Chania, Crete, b) Rhodes Island, c) Saronikos Gulf, d) Thermaikos Gulf, e) Samothrace Island.
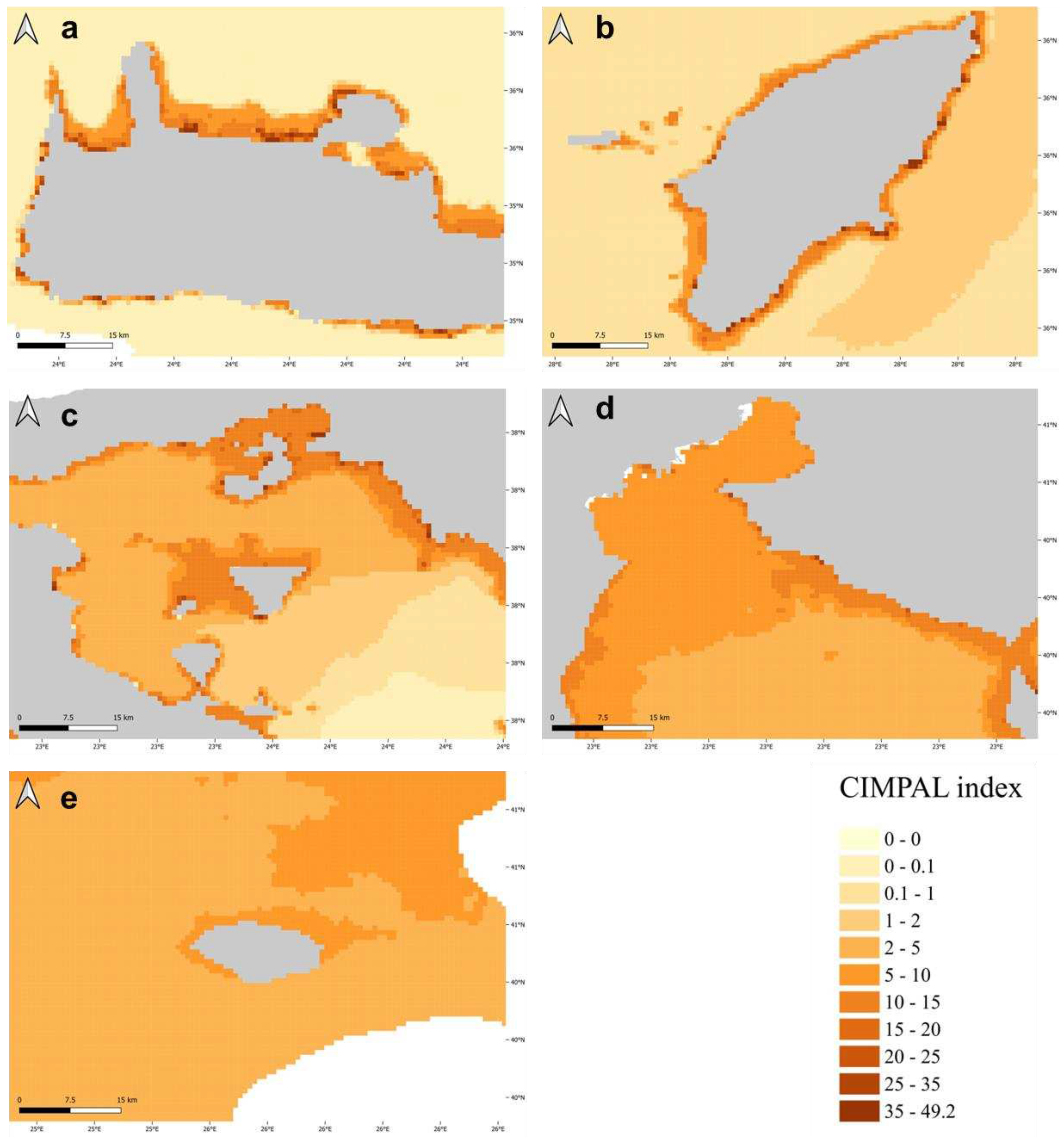
Figure 5.
a) Disaggregation of the total CIMPAL score for all cells in the study area by habitat type, b) average CIMPAL scores per habitat type.
Figure 5.
a) Disaggregation of the total CIMPAL score for all cells in the study area by habitat type, b) average CIMPAL scores per habitat type.
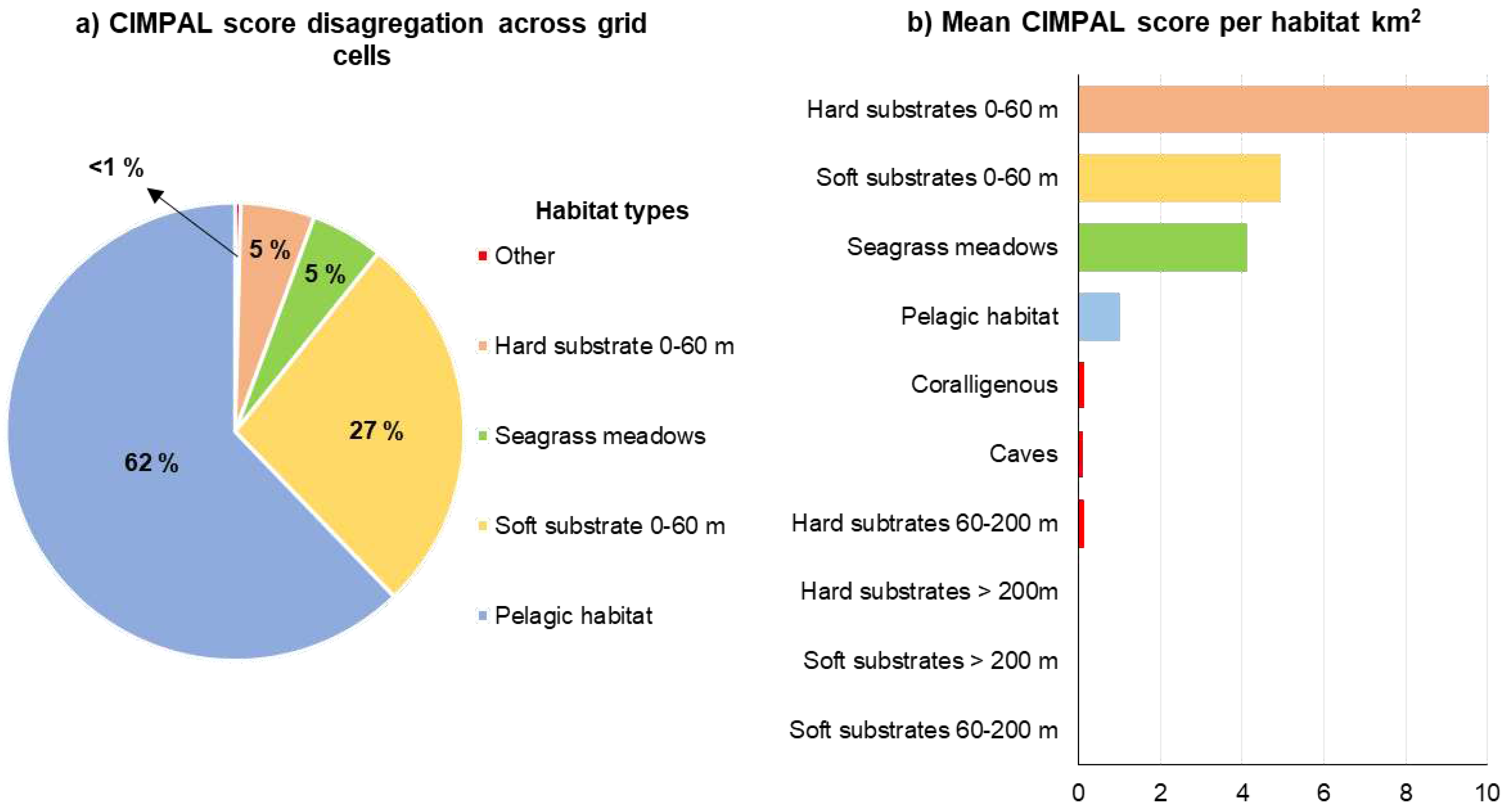
Figure 6.
a) Estimation of CIMPAL index on shallow hard substrates between 0-60 m depth in the Aegean Sea. b) Impacting ICAS on this habitat type, ranked by the estimated total impact score (summed over all grid cells). Green color for macrophytes, blue for fish and purple for invertebrates.
Figure 6.
a) Estimation of CIMPAL index on shallow hard substrates between 0-60 m depth in the Aegean Sea. b) Impacting ICAS on this habitat type, ranked by the estimated total impact score (summed over all grid cells). Green color for macrophytes, blue for fish and purple for invertebrates.
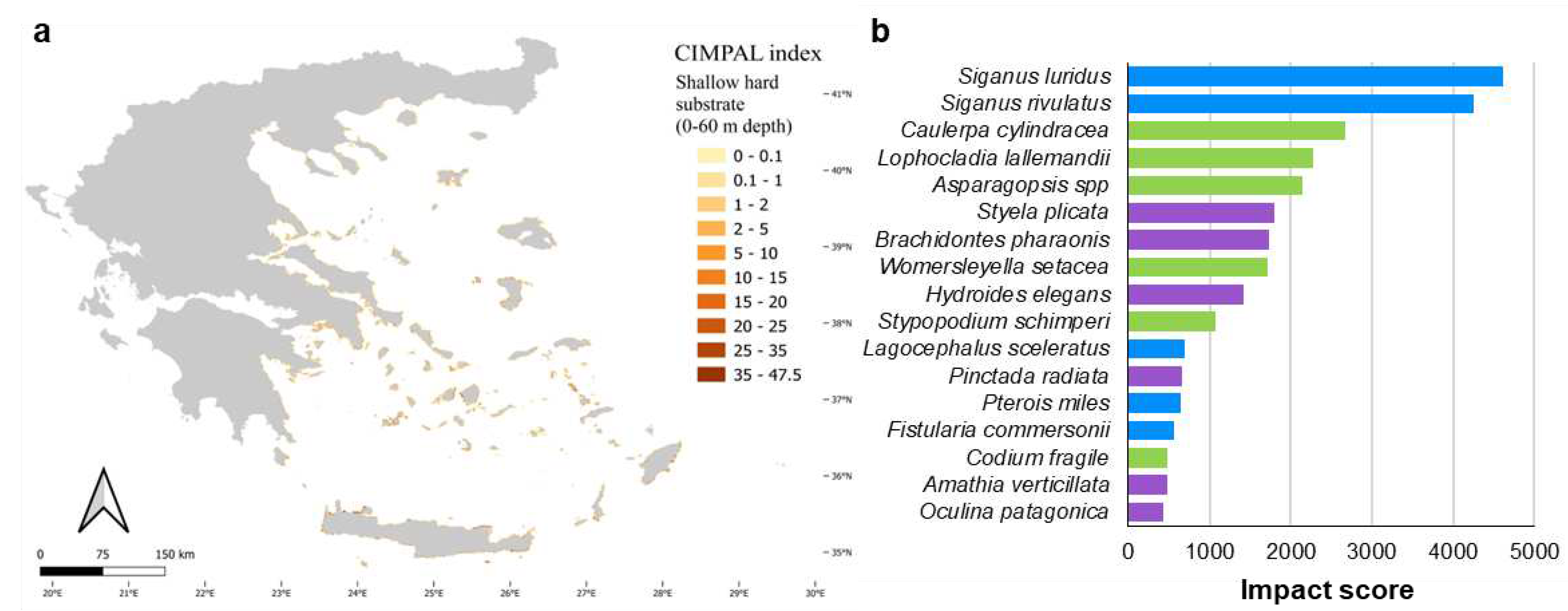
Figure 7.
a) Estimation of CIMPAL index on shallow soft substrates between 0-60 m depth in the Aegean Sea. b) Impacting ICAS on this habitat type, ranked by the estimated total impact score (summed over all grid cells). Green color for macrophytes, blue for fish and purple for invertebrates.
Figure 7.
a) Estimation of CIMPAL index on shallow soft substrates between 0-60 m depth in the Aegean Sea. b) Impacting ICAS on this habitat type, ranked by the estimated total impact score (summed over all grid cells). Green color for macrophytes, blue for fish and purple for invertebrates.
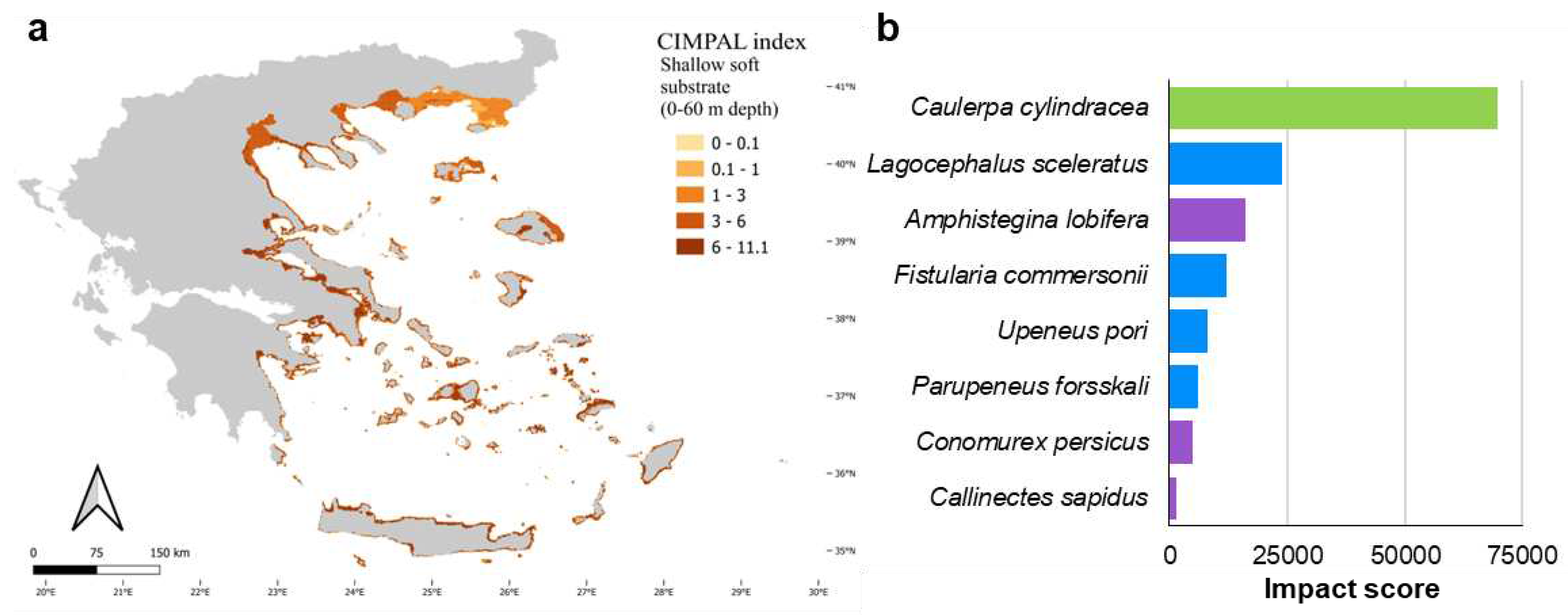
Figure 8.
a) Estimation of CIMPAL index on seagrass meadows in the Aegean Sea. b) Impacting ICAS on this habitat type, ranked by the estimated total impact score (summed over all grid cells). Green color for macrophytes, blue for fish and purple for invertebrates.
Figure 8.
a) Estimation of CIMPAL index on seagrass meadows in the Aegean Sea. b) Impacting ICAS on this habitat type, ranked by the estimated total impact score (summed over all grid cells). Green color for macrophytes, blue for fish and purple for invertebrates.
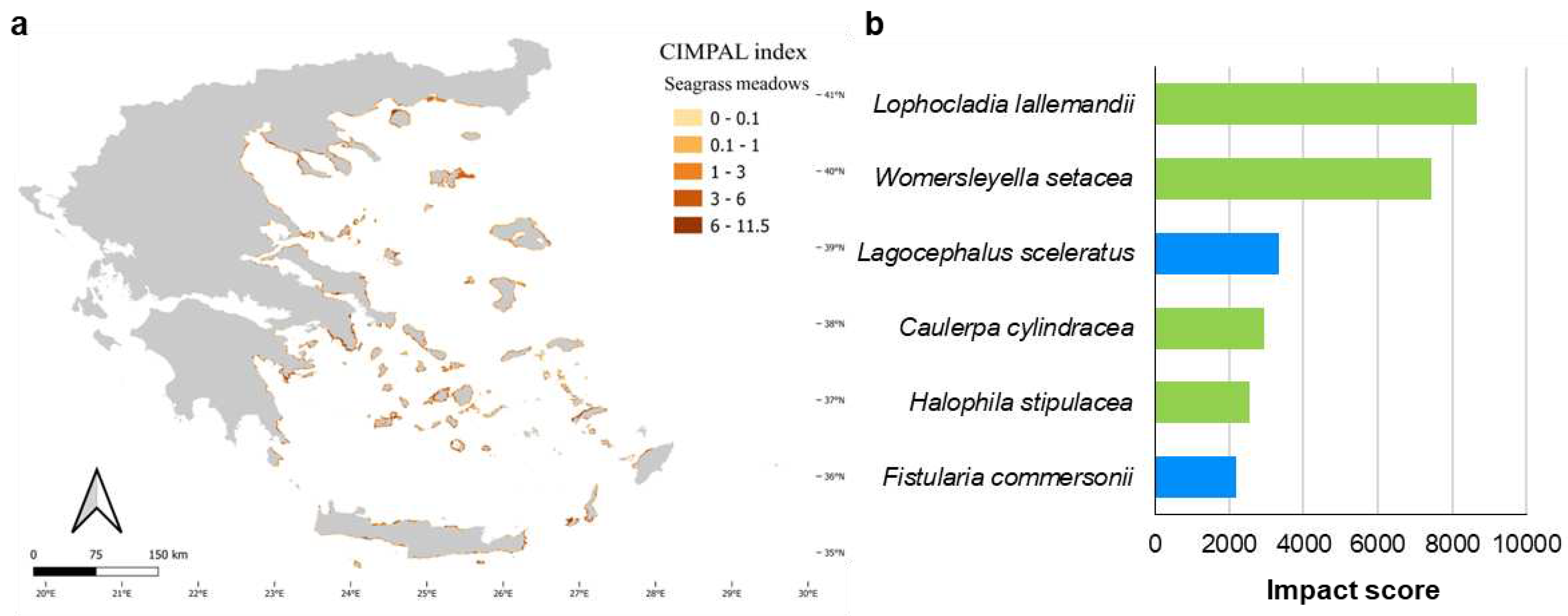
Figure 9.
(a) Disaggregation of the total CIMPAL score for all cells in the study area by biotic group (fish: blue color, macrophytes: green, invertebrates: purple), and maps of the impacts of ICAS in the Aegean Sea for (b) invertebrates, (c) macrophytes, and (d) fish.
Figure 9.
(a) Disaggregation of the total CIMPAL score for all cells in the study area by biotic group (fish: blue color, macrophytes: green, invertebrates: purple), and maps of the impacts of ICAS in the Aegean Sea for (b) invertebrates, (c) macrophytes, and (d) fish.
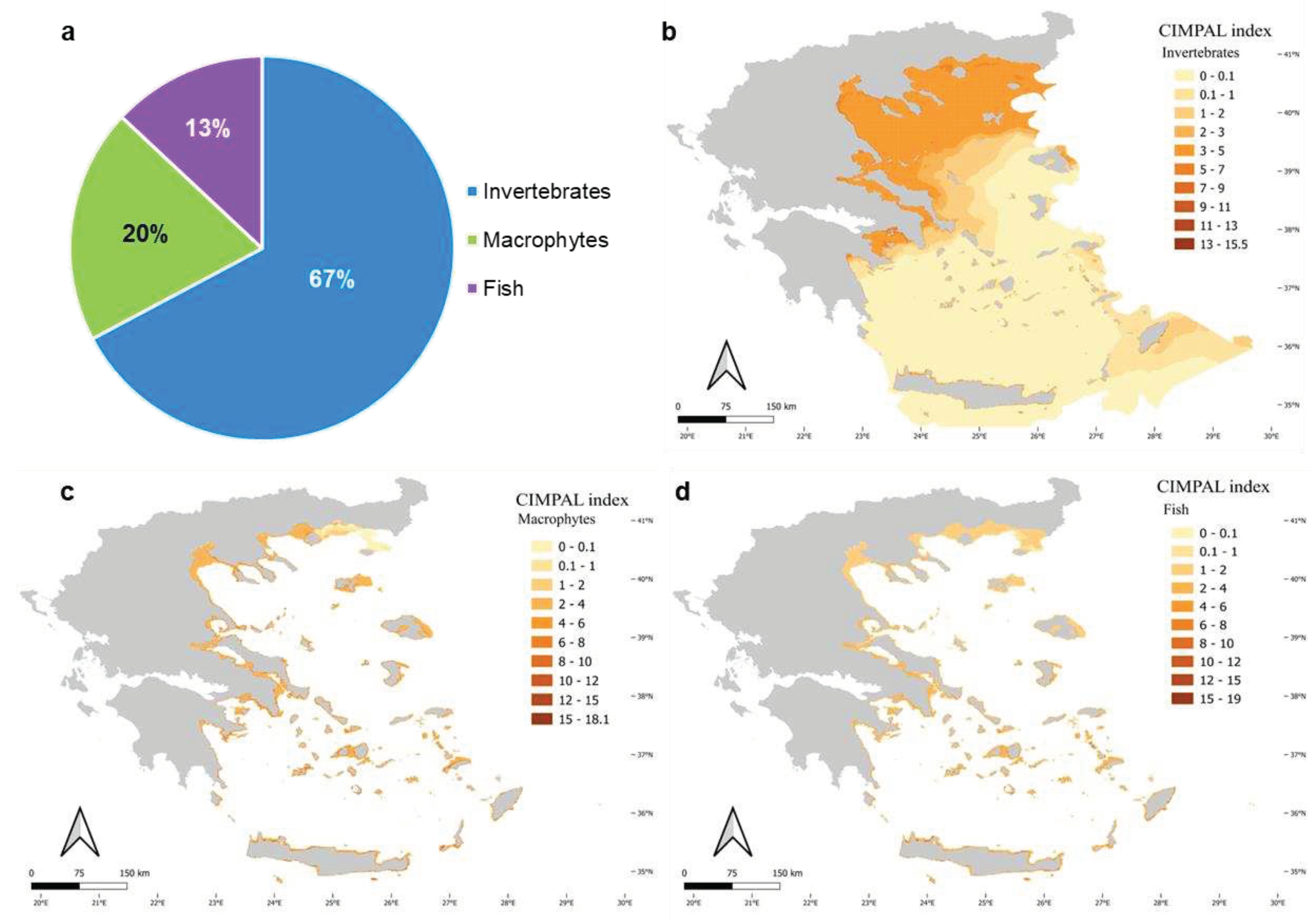
Figure 10.
Ranking of ICAS impact in the Aegean Sea according to the four CIMPAL indicators D1-D4 (excluding M. leidyi). Macrophytes: green color, fish: blue, and invertebrates: purple.
Figure 10.
Ranking of ICAS impact in the Aegean Sea according to the four CIMPAL indicators D1-D4 (excluding M. leidyi). Macrophytes: green color, fish: blue, and invertebrates: purple.
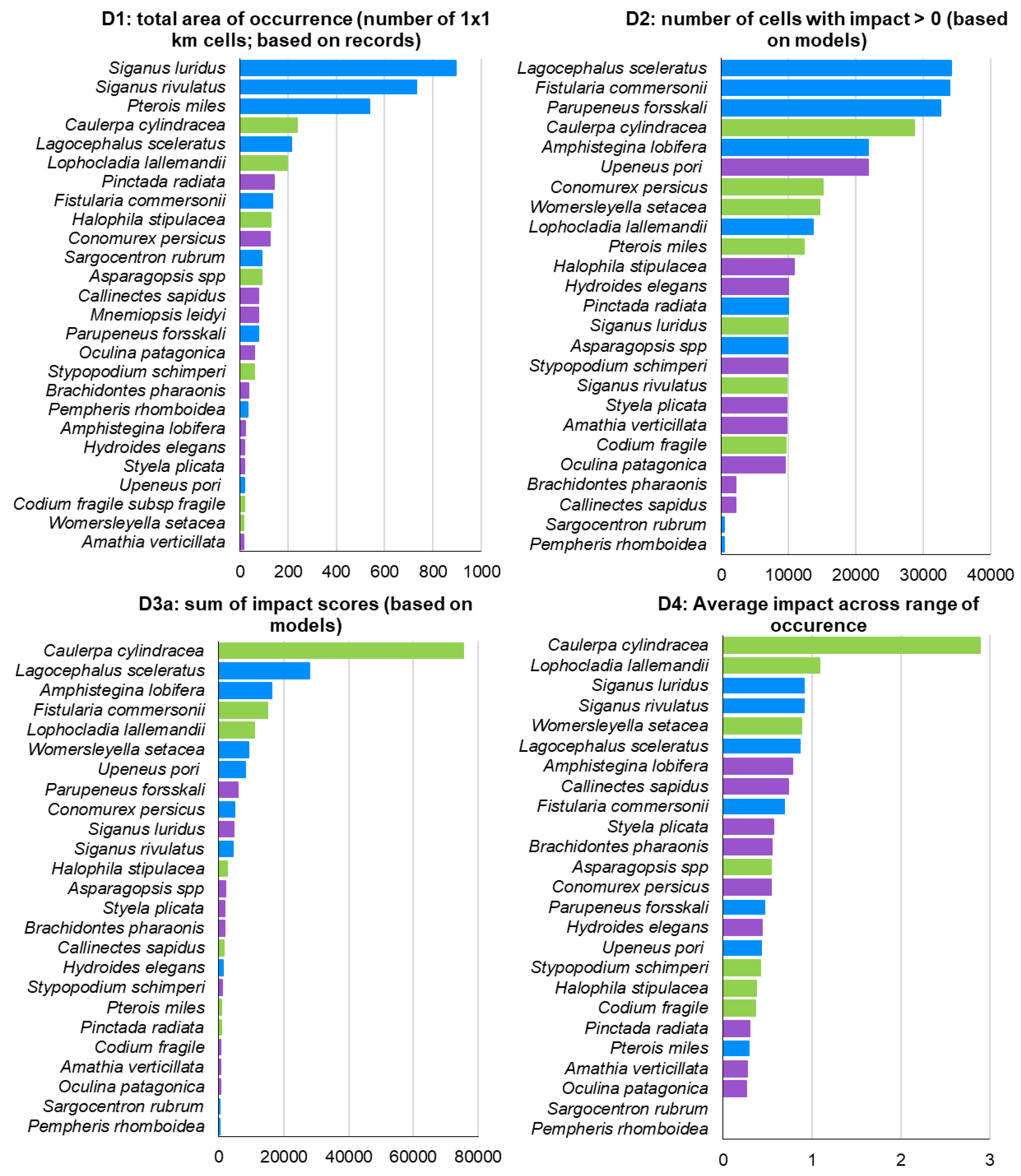
Figure 11.
ICAS impacts ranking using D3 indicator but with presence/absence data derived from field observations instead of modelled data.
Figure 11.
ICAS impacts ranking using D3 indicator but with presence/absence data derived from field observations instead of modelled data.
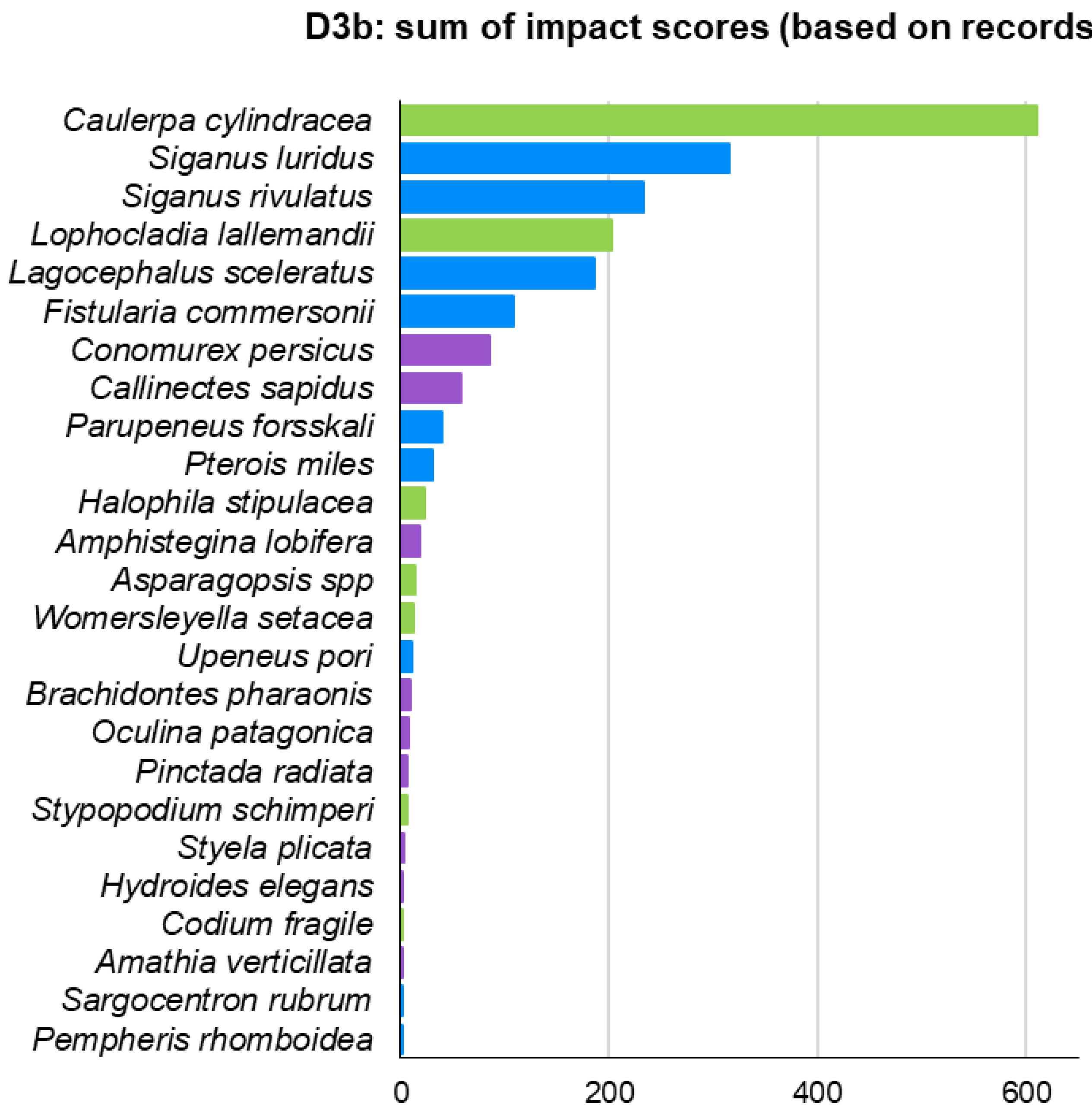

Table 1.
List of the targeted invasive cryptogenic and alien species (ICAS) species and their records in the Aegean Sea, for the application of the CIMPAL index.
Table 1.
List of the targeted invasive cryptogenic and alien species (ICAS) species and their records in the Aegean Sea, for the application of the CIMPAL index.
| Phylum, Class | Species | Status | Biotic group | Number of records |
|---|---|---|---|---|
| Arthropoda: Crustacea | Callinectes sapidus | Alien | Invertebrates | 115 |
| Bryozoa: Gymnolaemata | Amathia verticillata | Cryptogenic | Invertebrates | 19 |
| Chlorophyta: Ulvophyceae | Caulerpa cylindracea | Alien | Macrophytes | 283 |
| Chlorophyta: Ulvophyceae | Codium fragile | Alien | Macrophytes | 22 |
| Chordata: Ascidiacea | Styela plicata | Alien | Invertebrates | 21 |
| Chordata: Teleostei | Fistularia commersonii | Alien | Fish | 171 |
| Chordata: Teleostei | Lagocephalus sceleratus | Alien | Fish | 254 |
| Chordata: Teleostei | Parupeneus forsskali | Alien | Fish | 70 |
| Chordata: Teleostei | Pempheris rhomboidea | Alien | Fish | 31 |
| Chordata: Teleostei | Pterois miles | Alien | Fish | 574 |
| Chordata: Teleostei | Sargocentron rubrum | Alien | Fish | 88 |
| Chordata: Teleostei | Siganus luridus | Alien | Fish | 919 |
| Chordata: Teleostei | Siganus rivulatus | Alien | Fish | 748 |
| Chordata: Teleostei | Upeneus pori | Alien | Fish | 23 |
| Cnidaria: Anthozoa | Oculina patagonica | Cryptogenic | Invertebrates | 51 |
| Ctenophora: Tentaculata | Mnemiopsis leidyi | Alien | Invertebrates | 77 |
| Foraminifera: Globothalamea | Amphistegina lobifera | Alien | Invertebrates | 26 |
| Mollusca: Bivalvia | Brachidontes pharaonis | Alien | Invertebrates | 35 |
| Mollusca: Gastropoda | Conomurex persicus | Alien | Invertebrates | 129 |
| Mollusca: Bivalvia | Pinctada radiata | Alien | Invertebrates | 159 |
| Ochrophyta: Phaeophyceae | Stypopodium schimperi | Alien | Macrophytes | 67 |
| Annelida: Polychaeta | Hydroides elegans | Alien | Invertebrates | 24 |
| Rhodophyta: Florideophyceae | Asparagopsis spp. | Alien | Macrophytes | 117 |
| Rhodophyta: Florideophyceae | Lophocladia lallemandii | Alien | Macrophytes | 224 |
| Rhodophyta: Florideophyceae | Womersleyella setacea | Alien | Macrophytes | 24 |
| Tracheophyta: Magnoliopsida | Halophila stipulacea | Alien | Macrophytes | 132 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



