
Submitted:
04 May 2023
Posted:
04 May 2023
You are already at the latest version
Abstract
Equipped with an early social predisposition immediately post-birth, humans typically form associations with mothers and other family members through exposure learning, canalized by a spontaneous predisposition to biological motion, face configuration, and other cues of animacy. If impaired, reduced social preferences can lead to social interaction impairments such as autism spectrum disorder (ASD) via misguided canalization. Despite being taxonomically distant, domestic chicks also follow a homologous developmental trajectory toward adaptive socialization through imprinting, which is guided via predisposed preferences that are homologous to those of humans, thereby suggesting that chicks are valid animal models of ASD. In addition to the convergent similarities in predisposition with human newborns, accumulating evidence suggests the construct validity of the chick model. Considering the recent progress in the evolutionary neurobiology of vertebrates, we reviewed the advantages and limitations of the chick model of developmental mental diseases in humans.
Keywords:
autism spectrum disorder
; chick
; valproic acid
; nicotinic acetylcholine receptors
; biological motion
; face
; animacy
; thyroid hormone
; bumetanide
1. In search of valid animal models of developmental psychiatric disorders
Biological psychiatry and comparative psychology share the common scientific issue of neurocognitive homology among animals under study. Since the Cambrian period, the nervous system of vertebrates has held a highly conserved “Bauplan” [1,2,3]. Accordingly, homologies are widely found from the level of composite molecules (transmitters, hormones, receptors, and second messengers) to neural pathways and cytoarchitectonic organizations at the macroscopic level [4,5] (specifically for the mammal-bird homologies [5]). Despite its conservative nature, huge phenotypic diversification occurs in the behavioral and cognitive aspects of vertebrates through differentiated development [6], complicating the untangling of homology issues.
In the present review, encouraged by recent progresses of avian model studies [7,8], we argue that chicks and humans exhibit an evolutionary convergence in neurobehavioral traits and examine the validity of domestic chicks as autism spectrum disorder (ASD) animal models on the surface and construct validity (as Willner [9] proposed for the depression model). The former (surface validity) includes an examination of the similarity of behaviors and developmental processes, particularly the perceptual predisposition to faces and biological motion. The latter (construct validity) includes shared neural substrates and molecular cascades, particularly the potential roles of nicotinic acetylcholine (nAChR) transmission in the fetal brain in controlling excitation-inhibition balance in neonates.
2. Diverse environmental risk factors of ASD remain to be specified
ASD is the most prevalent developmental disorder primarily characterized by underdeveloped social interactions and communication [10,11]. It is speculated that the heterogeneous diagnostic phenotypes, as well as the dimensional nature of this disorder, could be associated with a wide range of underlying genetic and environmental factors [12]. In addition to apparent genetic risks [13] (for an exhaustive hereditability study in Sweden); also see [14] for de novo mutations and [15] for common variations), exposure to environmental toxicants during pregnancy and the neonatal period remains a major social and scientific concern [16]. For example, a large-scale twin study [17] revealed high concordance rates among siblings, indicative of the role of genetic factors; however, the authors also reported that common environmental factors shared by these twins could substantially contribute to autism/ASD liability. Complex interactions are thought to occur between the genetic background of ASD susceptibility and the chemical agents acting during pregnancy and the early post-natal period.
However, this hypothesis does not imply that these chemicals need to be eliminated. For example, valproic acid (VPA) has been identified as a risk factor for ASD (see below for details); however, it remains an indispensable antiepileptic medication [18]. The associated risks must be precisely evaluated in close consideration with known benefits. But, a lack of reliable measures often obscures environmental risk management. Studies using appropriate animal models are critical [19]. Rodents (mice and rats) are the most popular model animals [20,21] but cannot be deemed identical to humans. Rodents and primates have undergone distinct evolution since their separation during the late Cretaceous period over 65 million years ago.
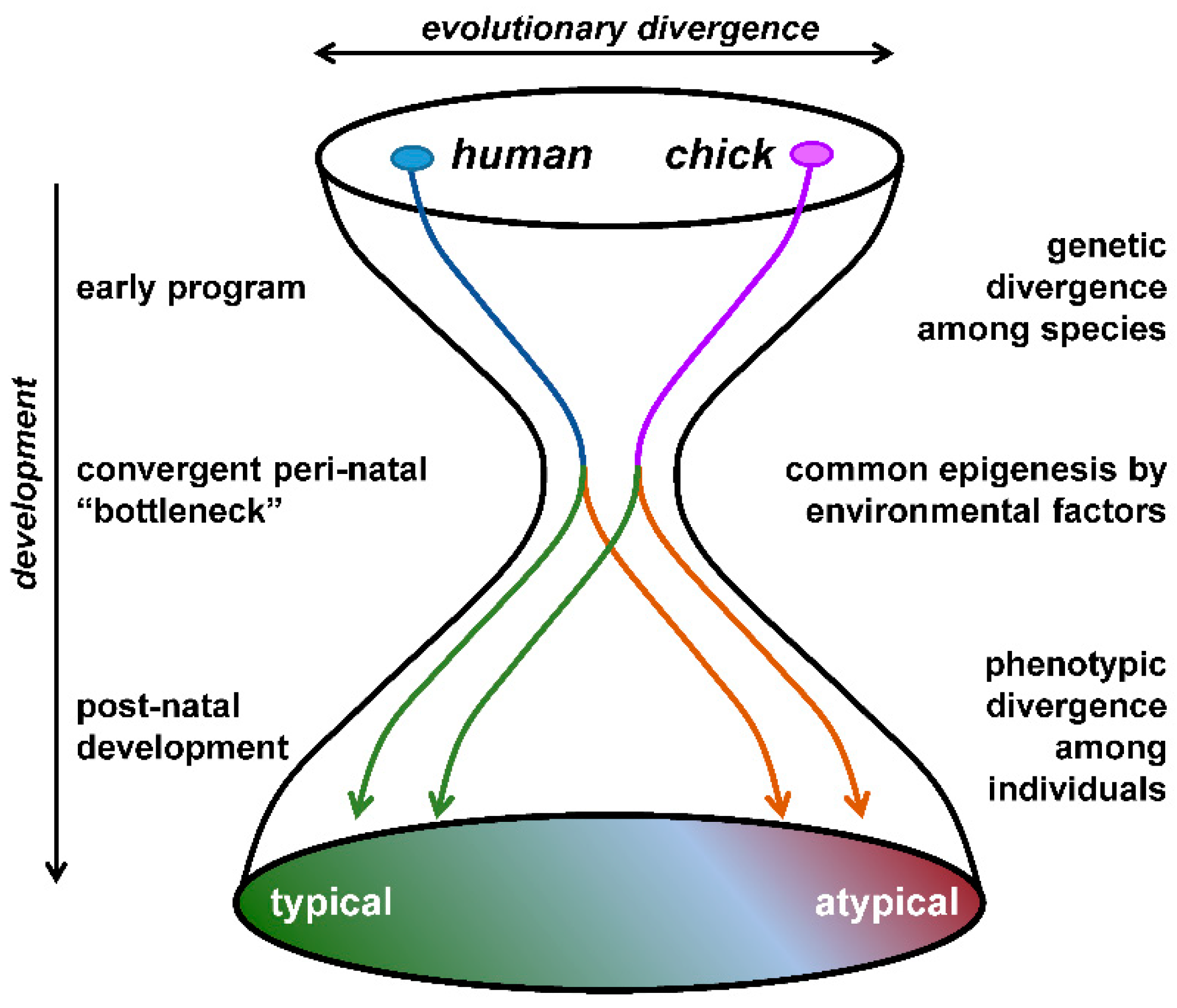
As an alternative animal model, we have focused on newly hatched domesticated chicks (Gallus gallus domesticus) [22,23,24,25]. Birds are descendants of theropod dinosaurs, the major group of sauropsids, whereas primitive mammals diversified as a minor group of synapsids during the Carboniferous Period, ~ 300 million years ago. Considering such a taxonomically distinct animal as a valid model for human psychiatric disorders may sound unrealistic. If the adult phenotypes are solely compared, humans can never be chickens. However, as the hourglass bottleneck theory suggests [26], humans and chickens achieve developmental convergence during the prenatal/early neonatal periods in terms of specific neurocognitive aspects (Figure 1). Determining whether certain animals are human-like demands the elucidation of how phenotypic similarities arise through their respective ontogenies.
3. Visual predispositions canalize the development of social behaviors: common developmental features for the surface validity.
It is speculated that the development of socialized behaviors during the early neonatal period depends on a predisposed visual preference. Human babies exhibit biased visual attention, which subsequently forms social attachments with siblings and parents. The predisposed “Conspec” [27] or “core knowledge” [28] could include a variety of visual features (for reviews see [29,30,31,32,33,34]), however, most studies have focused on face configuration, biological motion, and other cues of animacy.
Face Differential tuning to face configuration has been demonstrated in adolescents with ASD when compared with typically developing controls (for one of the earliest reviews, see [35,36]; also see [37] for a recent study using pictures resembling the food-face paintings by Giuseppe Arcimboldo). Based on the finding that infants prefer faces at 2–4 months old [38,39], a predisposed face preference has been suggested as a reliable diagnostic indicator for ASD. Typically, a definitive ASD diagnosis can be achieved in toddlers aged ~3 years; reliable biomarkers at earlier ages, if available, could facilitate treatment at the early stages [40]. In addition to behavioral measures, physiological measures, such as electroencephalography (EEG), can be employed in newborn babies [41].
Based on data from brain studies, it can be suggested that the subcortical visual pathway (superior colliculus/pulvinar to the amygdala) is responsible for the proto-face configuration (three blobs configured at a low spatial frequency) in human neonates [36]; see [42] for the most reliable account of this pathway in rhesus monkeys. Visually naïve monkeys deprived of social stimuli (including human faces) preferred faces [43], indicating robust development of face perception. Even human fetuses in the third trimester of pregnancy reportedly exhibit head-turn responses to upright face configurations [44]. Consistently, damage to the amygdala has been shown to cause some ASD-like deficiencies according to the “theory of minds” (ToM [45]), although the causal link between the ToM and ASD remains controversial [46].
Considering that the subcortical face pathway is functional in the early prenatal period and critical for the typical development of social behavior [47], we would expect to detect differentiated visual attention to the face in infants with familial risk of ASD and more specifically, in those who are later diagnosed with ASD. However, the use of the “face pop-up” task in these infants revealed clear attention to face similar to that observed in control individuals (or even higher) at 7 and 14 months of age [48]. Conversely, another longitudinal developmental study has reported a steady decline in selective eye-fixating behavior in infants who were subsequently diagnosed with ASD; the finding was observed in typically developed individuals [49]. Therefore, developmental changes need to be carefully examined [50]. More recently, it has been reported that newborns with a high familial risk of ASD (6–10 days of age) show a reduced face preference when compared with low-risk controls [51]; also see [52] for the follow-up longitudinal study on 4-month-old infants).
In addition, visually naïve, newly hatched chicks exhibit an evident inborn preference for faces [53,54,55]. Fetal exposure to VPA substantially reduced the face preference score [56], possibly paralleling that observed in humans. The subcortical visual pathway of birds (optic tectum and arcopallium/nucleus taeniae amygdala) is known to functionally mature early, and newly hatched chicks start actively pecking small conspicuous objects at 1–2 days old. Furthermore, immediate early gene imaging studies have revealed that telencephalic limbic nuclei are involved in the predisposition preference of chicks [57,58,59]. Moreover, traditional rodent models fail to spontaneously exhibit a comparable visual predisposition.
Biological motion Biological motion (BM) preference comprises another aspect of the “Conspec” process; it is easily tested using highly reduced animations composed of relatively few light points (~ a dozen). Both human and chick neonates exhibit BM preference without visual experience ([60,61] in human infants; [62] in chicks; also see [63] for the association of the BM preference with brain asymmetry). In addition to the commonality in their appearance at the early neonatal stage, both human infants and chicks show a clear inversion effect, that is, a preference for the upright walking motion over the inverted upside-down display ([64] in human infants; [65] in chicks).
The motion characteristics of both local movements (such as the movements of the lower limbs) and global features (shape of the body) are critical in humans [66,67]. However, it remains unclear which of these visual components is associated with the development of social cognition. [68] reported that infants depend more on local cues, whereas [69] highlighted the importance of translational displacement of the body. [70] have proposed an integrative hypothesis of the two-process theory, wherein the “step detector” responsible for the local motions of feet below the body precedes the “bodily action evaluator” that processes the global processing of action types and styles. Several studies in both human adults and children have suggested the association between reduced sensitivity to BM and ASD [71,72,73,74] (also see the recent meta-analysis [75]). Importantly, it should be noted that, in our current context, visual preferences for social stimuli (face inversion, averted eye gaze, and BM) markedly differ between the two groups of infants with high and low familial risk of autism [51]. Further longitudinal studies are needed to determine the association between visual preference and the subsequent development of social interactions.
Animacy In addition to the face and BM, both chicks and humans have a visual predisposition to other cues of animacy ([25,76,77] for reviews). For example, living organisms are characterized by self-propelling animacy, a well-established preference in both human babies [78] and visually naïve chicks [79,80]. Unpredictable speed changes in motion are a critical feature of animacy in newborn human babies [81], as observed in newly hatched chicks with distinct inherited variability [82,83] (for involvement of septal and hypothalamic nuclei, see [84]). Both variability in body orientation and the unpredictable temporal contingency of motion are critical in chicks [85,86]. It should also be noted that avoidance of looking at (threatening) objects is also considered to be innately predisposed [87].
4. Imprinting and the early process of attachment formation
In chicks, the BM preference is functionally linked to filial imprinting. Imprinting is a complex process that involves predisposition and experience-based learning; thus, it may be homologous to the processes of attachment formation in human babies. Newly hatched domestic chicks and ducklings form lasting attachments, even when the first object seen is a non-biological artifact rather than a conspecific animal [88,89]. Artifacts such as rotating blue boxes were actually effective as imprinting objects [90]. However, contrary to this popularly accepted idea, memorized preference for artifacts is short-lived and is gradually replaced by more naturalistic stimuli such as stuffed hens [91,92,93]. Accordingly, an innate predisposition gradually emerges after learned attachment fades. In contrast, BM preference emerges first and subsequently guides learning.
Perfectly naïve chicks show an apparent preference for BM, although with a relatively small effect size [62]. When imprinted by motion pictures, the BM preference is enhanced or “permissively induced” [94]. Induction is nonspecific to the exposed stimulus, and any motion (even randomized point-light animation) is a similarly effective inducer. Furthermore, the BM preference facilitates imprinting [95]. BM animations were more effective than non-BM animations, and chicks with a higher BM preference exhibited higher imprinting scores.
Imprinting memory is coupled with BM induction through enhanced thyroid hormone activity. Exposure to motion increases the expression of Dio2, which is responsible for the conversion of circulating thyroid hormone (T4) to its active form (T3) in the epithelial cells of telencephalic capillaries [96]. The enhanced T3 influx into the dorsal pallium (intermediate medial mesopallium, IMM, an avian homolog of the mammalian neocortex, including the association areas) reopens the sensitive period and acutely strengthens learning and BM scores [97,98]. In aged chicks, T3 can reactivate the preference for animate objects [99]. Accordingly, imprinting allows chicks to remain imprintable for a prolonged period, guiding subsequent learning during the extended sensitive period to objects bearing BM features. Notably, these two aspects of imprinting (i.e., memory formation and induced predisposition) appear tightly coupled and not dissociable [100].
Newly hatched domestic chicks could serve as a valid animal model for studying environmental risk factors for ASD, at least at the surface phenomena level. The following section examines how ASD-like deficiencies could arise in the chick model and whether the underlying mechanisms are shared.
5. VPA, an anticonvulsant drug, mediates ASD-like impairment of social behavior development and acute suppression of spontaneous fetal movements
Given that VPA was identified as an environmental risk factor for ASD during pregnancy [101,102,103], studies have attempted to clarify the underlying mechanisms using rodent models (see reviews [20,21,104]). Fetal exposure to VPA impairs social behavior also in chicks [105,106,107,108,109]. These studies have consistently reported impaired social behavior in newborns and hatchlings after fetal VPA exposure, although drug-induced phenotype disorders were not necessarily identical, probably due to task-dependent variation among individuals. For example, [109] reported low imprinting scores exclusively in individuals with a low BM preference,.
VPA has a wide spectrum of pharmacological effects, including actions on N-methyl-D-aspartate type glutamate receptors (NMDA-R [110]) and inhibitory GABA transmission [111]. Moreover, VPA is well-accepted as a potent inhibitor of histone deacetylases (HDACs [112]). Acute anticonvulsant action on the fetus may induce ASD-like phenotypes, as VPA effectively suppresses fetal motion [109]. Spontaneous motion is ubiquitous among fetuses of vertebrates [113] (see also [114,115] for more recent reviews), although its functional roles remain unclear. Nevertheless, suppression of fetal movements fails to account for ASD-like deficiencies, since similarly effective suppressers (e.g., selective blockers of NMDA-R MK-801) failed to cause ASD-like symptoms.
The brain regions and molecular events that are responsible for the VPA-mediated ASD phenotypes remain elusive. In a rat model, VPA enhanced NMDA-R expression and synaptic potentiation in the hippocampus [116], thereby causing an imbalance between excitatory and inhibitory transmission (E-I imbalance; see [117,118] for comprehensive reviews). In support of the hyperexcitation hypothesis, post-natal blockade of NMDA-R by memantine (a drug prescribed for Alzheimer’s disease) rescued social interaction impairment [119]. Consistent with studies performed in rodent models, administering bumetanide (a selective blocker of NKCC1 co-transporter) immediately before training could rescue chicks with VPA-induced impaired imprinting [109]; the impact of bumetanide will be discussed subsequently.
6. Selective impairment of BM predisposition via fetal interference with nAChR receptors, including neonicotinoid insecticides
Pesticide chemicals, particularly considering the rapidly increasing consumption of neonicotinoid insecticides (NNs [120]), are another serious concern in the etiology of ASD. NNs were designed to selectively block cholinergic neurotransmission in insects with low toxicity in vertebrates. Early ecological reports have highlighted the population decline of insectivorous birds [121]. Following concerns regarding the high persistence of NNs in plants and soil, NNs were found to impair the migratory ability of granivorous birds [122,123]. Several recent epidemiological studies have reported the risk of maternal exposure to environmental NNs [124,125,126,127]. An early study [124] estimated the association between the indoor usage of imidacloprid (IMI; one of the most heavily used NNs for flea and tick treatment for pet animals) and ASD, detecting an alarming odds ratio of ~2.0. Prenatal exposure to agricultural pesticides was found to be associated with low intelligence quotient and verbal comprehension [125]. A large-scale study on the association between ambient pesticide usage (NNs included) and ASD in California’s agricultural region [126] detected considerable odd ratios for various pesticide chemicals; the effects of prenatal exposure were boosted by additonal exposure in neonatal infants. A rodent model study assessing acetamiprid (ACE; another NNs) has reported the abnormal development of social and anxiety-related behaviors in males after prenatal and lactational exposure [128] (see [127] for a recent systemic review).
Our study using a chick model [109] revealed high concordance with these reports in humans and rodents. Selective and non-selective blockade of nAChR (using tubocurarine and selective α7 subtype inhibitor), as well as impaired nAChR transmission mediated by IMI, could suppress fetal movements and impair the BM preference of hatchlings. Notably, nAChR blockade did not impair imprinting memory formation, thus revealing distinct dimensions of social behavior malformation from those induced by VPA.
7. Thyroid hormone, E-I imbalance in humans and chicks
Maternal hypothyroidism (gestational hypothyroxinemia) is another risk factor for ASD [129,130,131]. Circulating levels of thyroid hormones (THs, T4 in particular) could be a potential early biological marker for ASD. To date, no consensus has been reached regarding the role of THs in the development of ASD, given that both positive [132] and negative [133] results have been reported. In addition to being a critical determinant of imprinting in chicks [96], TH plays critical roles in diverse neurodevelopmental processes [134], particularly in the maturation of GABAergic transmission via the rapamycin (mTOR; mechanistic target of rapamycin) cascade [135]. The mTOR-GABA cascade may mediate the acute facilitatory effects of THs in chicks [96,136,137]. T3 acutely enhances GABAergic transmission in slice preparations of the chick pallium [138], although its functional link to the behavioral effects remains elusive.
Interestingly, in humans, symptomatic autism comorbid with fragile X syndrome and tuberous sclerosis complex (TSC) is accompanied by mutations in the mTOR signaling pathway. Rapamycin was shown to rescue social impairment in a mouse model of TSC [139]. A more recent study addressing macrocephaly in infants with ASD has suggested that synaptic pathology related to the mTOR pathway is responsible for hyperconnectivity [140].
In the central nervous system, the metabolic control responsible for the balanced management of energy income and growth may underlie appropriate socialization during the early stages of life in humans and chicks. In addition, premature E-I balance (or delayed GABA switch from depolarizing to hyperpolarizing response) could be a key event in neural maturation, affording a potential target for developing effective pharmacotherapies for ASD.
8. GABA switch, nicotinic transmission, and treatment using bumetanide and oxytocin
GABA exerts depolarizing transmission via GABA-A receptors in the embryonic stage, and excitatory GABA exerts trophic functions for the functional maturation of the brain. During the perinatal period, excitation is converted to adult-type inhibitory neurotransmission ([141,142,143] for comprehensive reviews; [144] for the hippocampal development; also see [145] specifically for the schizophrenia etiology); this conversion (referred to as the GABA switch) is mediated by a reduction in the intracellular chloride ion ([Cl-]i), which, in turn, can be attributed to the enhanced expression of cation-linked co-transporters responsible for the efflux of Cl- (KCC2 over NKCC1). To identify regulatory mechanisms underlying the GABA switch, nicotinic cholinergic transmission was found to be critical in the chick ciliary ganglion [146]. Further analyzing the phosphorylation of KCC2 molecules revealed protein kinase C-mediated modulation by glutamatergic and serotonergic actions, as well as the activation of muscarinic acetylcholine receptors [147] (for more recent comprehensive reviews see [148,149]).
The association between GABA switch retardation and ASD was based on the finding that diazepam (an anxiolytic drug, a positive modulator of GABA-mediated inhibition used to relieve anxiety) could paradoxically increase aggressive behaviors in autistic children [150]. Subsequent studies have shown that dysfunctional GABA inhibition could play a pivotal role in ASD etiology [151,152]. Bumetanide has attracted attention as a potent candidate for ameliorating ASD symptoms, given that this agent selectively blocks the NKCC1 co-transporter responsible for Cl- influx [153]. Although initial open-label small-sized trials appeared positive and promising [154,155], recent phase-2 trials failed to afford positive outcomes [156]. A detailed follow-up analysis has identified heterogeneous phenotypes of neurocognitive impairment in patients with ASD, some of which were unaffected by bumetanide [157]. In a chick model, bumetanide treatment immediately before training rescued impairments in both imprinting (by VPA) and BM preference (by nAChR blockade) [109]. Further studies are required to identify underlying targets and pharmacology.
Oxytocin and related nonapeptides comprise another group of candidate drugs for ASD. In humans, intranasal application of oxytocin can acutely ameliorate social deficiencies such as BM perception and social communication in cases of relatively low severity [158,159]. In typically developing individuals, oxytocin receptors peak during early childhood, whereas this peak is absent in those with ASD [160]. Similar facilitatory effects of oxytocin on prosocial behaviors have been observed in dogs [161,162], birds [163,164,165], and fish [166]; also see [167] for the zebrafish model of developmental disorders), suggesting its ubiquitous role in vertebrates. Considering the underlying mechanisms, a study using oxytocin-receptor knock-out mice has suggested that KCC2 is regulated by oxytocin [168]. A more recent study in mice identified a link between the genetic risk of ASD and the oxytocinergic signaling pathway [169]. In chicks, intracranially administered mesotocin (an avian counterpart of oxytocin) enhanced the preference of naïve chicks [165].
Although disadvantageous as a genetic model of ASD owing to the limited availability of powerful tools for gene manipulation, the chick model could substitute rodent models in several technical aspects. Firstly, maternal complications are disregarded in chicks as direct fetal administration is possible in eggshells. Secondly, the time course and dosage of chemicals can be precisely controlled. The short incubation time and quick testing (usually 8 days after drug application) enable the rapid screening of harmful chemicals. Third, the use of chicks satisfies the 3Rs of animal experiments (https://nc3rs.org.uk/who-we-are/3rs): replacement with mammals, reduction in number due to controlled chemical treatments, and refinement by sophisticated visual predispositions like humans.
Chicks are unique models for studying neurocognitive disorders in humans. The fundamental question is, why? Why did chicks converge to human neonates despite distinct evolutionary separations over 300 million years? This remains an unresolved puzzle, which could be addressed by analyzing behavioral phenotypes and disorders (surface validity) and the underlying mechanisms (construct validity). Biological psychiatry and comparative psychology ask the same question: What are we?
Conflicts of Interest
Both authors decleare no conflict of interests.
References
- Grillner, S.; Robertson, B. The basal ganglia over 500 million years. Curr Biol, 2016, 26, R1088–R1100. [Google Scholar] [CrossRef] [PubMed]
- Suryanarayana, S.M.; Pérez-Fernández, J.; Brita Robertson, B.; Grillner, S. The lamprey forebrain – evolutionary implications. Brain Behav Evol. 2021. [CrossRef] [PubMed]
- Suryanarayana, S.M.; Brita Robertson, B.; Grillner, S. The neural bases of vertebrate motor behaviour through the lens of evolution. Philos Trans B, 2021, 377, 20200521. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, G.M.; Grillner, S. Handbook of brain microcircuit. Oxford University Press, Oxford, 2018.
- Güntürkün, O.; Bugnyar, T. Cognition without cortex. Trend Cog Sci, 2016, 20, 291–303. [Google Scholar] [CrossRef]
- Bateson, P.; Gluckman, P. Plasticity, robustness, development and evolution. Cambridge University Press, 2011.
- Csillag, A.; Ádám A, Zacher G. Avian models for mechanisms underlying altered social behavior in autism. Avian models for mechanisms underlying altered social behavior in autism. Frontiers Physiol. 2022. [CrossRef]
- Huang, X.; Cheng, H.-W. Perspective: chicken models for studying the ontogenetic origin of neuropsychiatric disorders. Biomedicines, 2022, 10, 1155. [Google Scholar] [CrossRef]
- Willner, P. The validity of animal models of depression. Psychopharmacol, 1984, 83, 1–16. [Google Scholar] [CrossRef]
- Wing, L.; Gould, J. Severe impairments of social interaction and associated abnormalities in children: epidemiology and classification. J Autism Develop Disord, 1979, 9, 11–29. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 5th ed. Washington: American Psychiatric Association. 2013.
- Tchaconas, A.; Adesman, A. Autism spectrum disorders: a pediatric overview. Curr Opin Pediatr, 2013, 25, 130–143. [Google Scholar] [CrossRef]
- Sandin, S.; Lichtenstein, P.; Kuja-Halkola, R.; Larsson, H.; Hultman, C.M.; Reichenberg, A. The familial risk of autism. JAMA, 2014, 311, 1770–1777. [Google Scholar] [CrossRef]
- Iossifov, I.; O'Roak, B.J.; Sanders, S.J.; et al. The contribution of de novo coding mutations to autism spectrum disorder. Nature, 2014, 515, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Gaugler, T.; Klei, L.; Sanders, S.J. Most genetic risk for autism resides with common variation. Nature Genetics, 2014, 46, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, D.A.; Frye, R.E. Environmental toxicants and autism spectrum disorders: a systematic review. Transl Psychiatry. 2014, 4, e360. [Google Scholar] [CrossRef] [PubMed]
- Hallmayer, J.; Cleveland, S.; Torres, A.; et al. Genetic heritability and shared environmental factors among twin pairs with autism. Ach Gen Psychiatry, 2011, 68, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Meador, K.J.; Loring, D.W. Risks of in utero exposure to valproate. JAMA, 2013, 309, 1730–1731. [Google Scholar] [CrossRef]
- Ergaz Z, Weinstein-Fudim L. ; Ornoy A. Genetic and non-genetic animal models for autism spectrum disorders (ASD). Reprod Toxicol, 2016, 64, 116–140. [Google Scholar] [CrossRef]
- Mabunga, D.F.N.; Gonzales, E.L.T.; Kim, J.-W.; Kim, K.C.; Shin, C.Y. Exploring the validity of valproic acid animal model of autism. Exp Neurobiol, 2015, 24, 285–300. [Google Scholar] [CrossRef]
- Chaliha, D.; Albrecht, M.; Vaccarezza, M.; Takechi, R.; Lam, V.; Al-Salami, H.; Mamo, J. A systematic review of the valproic acid-induced rodent model of autism. Develop Neurosci, 2020, 42, 12–48. [Google Scholar] [CrossRef]
- Vallortigara, G. Core knowledge of object, number, and geometry: a comparative and neural approach. Cog Neuropsychol, 2012, 29, 213–236. [Google Scholar] [CrossRef]
- Rosa-Salva, O.; Farroni, T.; Regolin, L.; Vallortigara, G.; Johnson, M.H. The evolution of social orienting: evidence from chicks (Gallus gallus) and human newborns. Plos One, 2011, 6, e18802. [Google Scholar] [CrossRef]
- Rosa-Salva, O.; Mayer, U.; Vallortigara, G. Roots of a social brain: developmental models of emerging animacy-detecting mechanisms. Neurosci Biobehav Rev, 2015, 50, 150–168. [Google Scholar] [CrossRef]
- Versace, E.; Martinho-Truswell, A.; Kacelnik, A.; Vallortigara, G. Priors in animal and artificial intelligence: where does learning begin? Trends Cog Sci. 2018. [CrossRef]
- Uesaka, M.; Kuratani, S.; Irie, N. The developmental hourglass model and recapitulation: an attempt to integrate the two models. J Exp Zool B (Mol Dev Evol), 2020, 338, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.; Johnson, M.H. CONSPEC and CONLERN: a two-process theory of infant face recognition. Psychol Rev, 1991, 98, 164–181. [Google Scholar] [CrossRef] [PubMed]
- Spelke, E.S.; Kinzler, K.D. Core knowledge. Develop Sci, 2007, 10, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, E.; Vallortigara, G. Evolutionary and Neural Bases of the Sense of Animacy. In: The Cambridge Handbook of Animal Cognition (Eds. Allison Kaufman, Josep Call, James Kaufman), 2021. pp. 295-321, Cambridge University Press, New York.
- Vallortigara, G. Born Knowing. Imprinting and the Origins of Knowledge. MIT Press, Cambridge, Massachusetts, London, England, 2021.
- Versace, E.; Vallortigara, G. Origins of knowledge: Insights from precocial species. Frontiers Behav Neurosci, 2015, 9, 338. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, E.; Loveland, J.L.; Mayer, U.; Rosa-Salva, O.; Versace, E.; Vallortigara, G. Filial responses as predisposed and learned preferences: Early attachment in chicks and babies. Behav Brain Res. 2017, 325, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Salva, O.; Mayer, U.; Versace, E.; Hebert, M.; Lemaire, B.S.; Vallortigara, G. Sensitive periods for social development: Interactions between predisposed and learned mechanisms. Cognition. 2021. [CrossRef]
- Lemaire, B.; Vallortigara, G. Life is in motion (through a chick's eye). Animal Cog. 2022. [CrossRef]
- Johnson, M.H. Imprinting and development of face recognition: from chick to man. Curr Direct Psychol Sci, 1992, 1, 52–55 https://wwwjstororg/stable/20182129. [Google Scholar] [CrossRef]
- Johnson, M.H. Subcortical face processing. Nature Rev Neurosci, 2005, 6, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, M.A.; Guerreschi, M.; Tagliavento, L.; Gitti, F.; Sokolov, A.N.; Fallgatter, A.J.; Fazzi, E. Social cognition in autism: face tuning. Sci Rep, 2017, 7, 2734. [Google Scholar] [CrossRef]
- Mauer, D.; Barrera, M. Infant's perception of natural and distorted arrangemetns of a schematic face. Child Development, 1981, 52, 196–202. [Google Scholar] [CrossRef]
- Mondloch, C.J.; Lewis, T.L.; Budreau, D.R. Face perception during early infancy. Psychol Sci, 1999, 10, 419–422. [Google Scholar] [CrossRef]
- Elsabbagh, M.; Johnson, M.H. Trends Cog Sci, 2009, 14, 81-87. [CrossRef]
- Buiatti, M.; Di Giorgio, E.; Piazza, M.; Polloni, C.; Menna, G.; Taddei, F.; Baldo, E.; Vallortigara, G. Cortical route for face like pattern processing in human newborns. PNAS. 2019, 116, 4625–4630. [Google Scholar] [CrossRef] [PubMed]
- Inagaki M, Inoue KI, Tanabe S, Kimura K, Takada M, Fujita I. Rapid processing of threatening faces in the amygdala of non-human primates: subcortical inputs and dual roles. Cerebral Cortex, 2022, 33, 895–915. [Google Scholar] [CrossRef]
- Sugita, Y. Face perception in monkeys reared with no exposure to faces. PNAS, 2008, 105, 394–398. [Google Scholar] [CrossRef]
- Reido, V.M.; Dunn, K.; Young, R.J.; Amu, J.; Donovan, J.; Reissland, N. The human fetus preferentially engages with face-like visual stimuli. Curr Biol, 2017, 27, 1825–1828. [Google Scholar] [CrossRef]
- Stone, V.E.; Baron-Cohen, S.; Calder, A.; Keane, J.; Young, A. Acquired theory of mind impairments in individuals with bilateral amygdala lesions. Neuropsychologia, 2003, 41, 209–220. [Google Scholar] [CrossRef]
- Gernsbacher, M.A.; Yergeau, M. Empirical failures of the claim that autistic people lack a Theory of Mind. Archiv Scientific Psychol, 2019, 7, 102–118. [Google Scholar] [CrossRef]
- Baron-Cohen, S.; Ring, H.A.; Bullmore, E.T.; Ashwin, C.; Williams, S.C.R. The amygdala theory of autism. Neurosci Biobehav Rev, 2000, 24, 355–364. [Google Scholar] [CrossRef]
- Elsabbagh, M.; Gliga, T.; Pickles, A.; Hudry, K.; Charman, T.; Johnson, M.H.; BASIS Team. The development of face orienting mechanisms in infants at-risk of autism. Behav Brain Res, 2013, 251, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.; Klin, A. Attention to eye is present but in decline in 2-6-month-old infants later diagnosed with autism. Nature, 2013, 504, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Simion, F.; Di Giorgio, E. Face perception and processing in early infancy: inborn predispositions and developmental changes. Frontiers Psychol, 2015, 6, 969. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, E.; Frasnelli, E.; Rosa-Salva, O.; Scattoni, M.L.; Puopolo, M.; Tosoni, D.; Simion, F.; Vallortigara, G. Difference in Visual Social Predispositions Between Newborns at Low- and High-risk for Autism. Sci Rep, 2016, 6, 26395. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, E.; Rosa-Salva, O.; Frasnelli, E.; Calcagni, A.; Lunghi, M.; Scattoni, M.L.; Simion, F.; Vallortigara, G. Abnormal visual attention to simple social stimuli in 4-month-old infants at high risk for Autism. Sci Rep. 2021, 11, 15785. [Google Scholar] [CrossRef]
- Rosa-Salva, O.; Regolin, L.; Vallortigara, G. Faces are special for newly hatched chicks: evidence for inborn domain-specific mechanisms underlying spontaneous preferences for face-like stimuli. Develop Sci, 2010, 13, 565–577. [Google Scholar] [CrossRef]
- Rosa-Salva, O.; Farroni, T.; Regolin, L.; Vallortigara, G.; Johnson, M.H. The evolution of social orienting: evidence from chicks (Gallus gallus) and human newborns. Plos One, 2011, 6, e18802. [Google Scholar] [CrossRef]
- Rosa-Salva, O.; Mayer, U.; Vallortigara, G. Unlearned visual preferences for the head region in domestic chicks. PLoS One, 2019, 14, e0222079. [Google Scholar] [CrossRef]
- Adiletta A, Pedrana S, Rosa-Salva O, Sgadò P. Spontaneous visual preference for face-like stimuli is impaired in newly-hatched domestic chicks exposed to valproic acid during embryogenesis. Frontiers Behav Neurosci, 2021, 15, 733140. [Google Scholar] [CrossRef]
- Mayer, U.; Rosa-Salva, O.; Lorenzi, E.; Vallortigara, G. Social predisposition dependent neuronal activity in the intermediate medial mesopallium of domestic chicks (Gallus gallus domesticus). Behav Brain Res, 2016, 310, 93–102. [Google Scholar] [CrossRef]
- Mayer, U.; Rosa-Salva, O.; Morbioli, F.; Vallortigara, G. The motion of a living conspecific activates septal and preoptic areas in naive domestic chicks (Gallus gallus). Eur J Neurosci, 2017, 45, 423–432. [Google Scholar] [CrossRef]
- Mayer, U.; Rosa-Salva, O.; Loveland, J.L.; Vallortigara, G. Selective response of the nucleus taeniae of the amygdala to a naturalistic social stimulus in visually naive domestic chicks. Sci Rep, 2019, 9, 9849. [Google Scholar] [CrossRef]
- Simion, F.; Regolin, L.; Bulf, H. A predisposition for biological motion in the newborn baby. PNAS, 2008, 105, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, M.A. Biological motion processing as a hallmark of social cognition. Cerebral Cortex, 2012, 22, 981–995. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Regolin, L.; Marconato, F. Visually inexperienced chicks exhibit spontaneous preference for biological motion patterns. Plos Biology, 2005, 3, e208. [Google Scholar] [CrossRef]
- Rugani, R.; Rosa-Salva, O.; Regolin, L.; Vallortigara, G. Brain asymmetry modulates perception of biological motion in newborn chicks (Gallus gallus). Behav Brain Res, 2015, 290, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Troje, N.F.; Westhoff, C. The inversion effect in biological motion perception: evidence for a “life detector”? Curr Biol, 2006, 16, 821–824. [Google Scholar] [CrossRef]
- Vallortigara, G.; Regolin, L. Gravity bias in the interpretation of biological motion by inexperienced chicks. Curr Biol, 2006, 16, R279–R280. [Google Scholar] [CrossRef]
- Chang, D.H.F.; Troje, N.F. Characterizing global and local mechanisms in biological motion perception. J Vision, 2009, 9, 1–10. [Google Scholar] [CrossRef]
- Hirai, M.; Saunders, D.R.; Troje, N.F. Allocation of attention to biological motion: local motion dominates global shape. J Vision, 2011, 11, 1–11. [Google Scholar] [CrossRef]
- Bardi, L.; Regolin, L.; Simion, F. Biological motion preference in humans at birth: role of dynamic and configural properties. Develop Sci, 2011, 14, 353–359. [Google Scholar] [CrossRef]
- Bidet-Ildei, C.; Kitromilides, E.; Orliaguet, J.-P.; Pavlova, M.; Gentaz, E. Preference for point-light biological motion in newborns: contribution of translational displacement. Develop Psychol, 2014, 50, 113–120. [Google Scholar] [CrossRef]
- Hirai, M.; Senju, A. The two-process theory of biological motion processing. Neurosci Biobehav Rev, 2020, 111, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Blake, R.; Turner, L.M.; Smoski, M.J.; Pozdol, S.L.; Stone, W.L. Visual recognition of biological motion is impaired in children with autism. Psychiol Sci, 2003, 14, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, M.D.; Pennington, B.F.; Rogers, S.J. The perception of animacy in young children with autism. J Autism Dev Disord, 2006, 36, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Xu, Q.; Liu, D.; Chen, L. Troje N.F.; He S.; Jiang Y. PNAS, 2018, 115, 1937–1942. [Google Scholar] [CrossRef]
- Kaliukhovich, D.A.; Manyakov, N.V.; Bangerter, A.; Ness, S.; Skalkin, A.; Boice, M.; Goodwin, M.S.; Dawson, G.; Hendren, R.; Leventhal, B.; Shic, F.; Pandina, G. J Autism Devlop Disord, 2021, 51, 2369-2380. [CrossRef]
- Federici, A.; Parma, V.; Vicovaro, M.; Radassao, L.; Casartelli, L.; Ronconi, L. Anomalous perception of biological motion in autism: a conceptual review and meta-analysis. Sci Rep, 2020, 10, 4576. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Rosa-Salva, O. Toolkits for cognition: From core-knowledge to genes. In Tucci, V. (Ed.) Neurophenome: Cutting-edge Approaches and Technologies in Neurobehavioral Genetics, 2017, 229-245, New York, Wiley-Blackwell. ISBN 978-1-118-54071-8.
- Vallortigara, G. 2021. Animacy detection and its role on our attitudes towards other species. In "Human/Animal Relationships in Transformation – Scientific, Moral and Legal Perspectives" (S. Pollo, A. Vitale, eds.), Palgrave.
- Di Giorgio, E.; Lunghi, M.; Simion, F.; Vallortigara, G. Visual cues of motion that trigger animacy perception at birth: the case of self-propulsion. Develop Sci, 2016, 20, e12394. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Salva, O.; Grassi, M.; Lorenzi, E.; Regolin, L.; Vallortigara, G. Spontaneous preference for visual cues of animacy in naïve domestic chicks: the case of speed changes. Cognition, 2016, 157, 49–60. [Google Scholar] [CrossRef]
- Rosa-Salva, O.; Hernik, M.; Broseghini, A.; Vallortigara, G. Visually-naïve chicks prefer agents that move as if constrained by a bilateral body-plan. Cognition, 2018, 173, 106–114. [Google Scholar] [CrossRef]
- Di Giorgio, E.; Lunghi, M.; Vallortigara, G.; Simion, F. Newborns’ sensitivity to speed changes as a building block for animacy perception. Sci Rep, 2021, 11, 542. [Google Scholar] [CrossRef] [PubMed]
- Versace, E.; Fracasso, I.; Baldan, G.; Dalle Zotte, A.; Vallortigara, G. Newborn chicks show inherited variability in early social predispositions for hen-like stimuli. Sci Rep, 2017, 7, 40296. [Google Scholar] [CrossRef] [PubMed]
- Versace, E.; Ragusa, M.; Vallortigara, G. A transient time window for early predispositions in newborn chicks. Sci Rep, 2019, 9, 18767. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, E.; Mayer, U.; Rosa-Salva, O.; Vallortigara, G. Dynamic features of animate motion activate septal and preoptic areas in visually naïve chicks (Gallus gallus). Neuroscience, 2017, 354, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, B.; Rosa-Salva, O.; Fraja, M.; Lorenzi, E.; Vallortigara, G. Spontaneous preference for unpredictability in the temporal contingencies between agents’ motion in naïve domestic chicks. Proc Royal Soc B, 2022, 289, 20221622. [Google Scholar] [CrossRef]
- Rosa-Salva, O.; Hernik, M.; Fabbroni, M.; Lorenzi, E.; Vallortigara, G. Naïve chicks do not prefer objects with stable body orientation, though they may prefer behavioural variability. Animal Cog. 2023. [CrossRef] [PubMed]
- Hebert, M.; Versace, E.; Vallortigara, G. Inexperienced preys know when to flee or to freeze in front of a threat. PNAS, 2019, 116, 22918–22920. [Google Scholar] [CrossRef]
- Spalding, D. Instinct with original observations on young animals. Macmillan’s Mag 1873, 27, 282–293 (Reprinted in British Journal of Animal Behaviour 2:2. [Google Scholar]
- Lorenz, K. The companion in the bird’s world. Auk, 1937, 54, 245–273. [Google Scholar] [CrossRef]
- Horn, G. Pathway of the past: the imprint of memory. Nature Rev Neurosci, 2004, 5, 108–120. [Google Scholar] [CrossRef]
- Bolhuis, J.J.; Johnson, M.H.; Horn, G. Effects of early experience on the development of filial preferences in the domestic chick. Dev Psychobiol, 1985, 18, 299–308 https ://doiorg/101002/dev420180403". [Google Scholar] [CrossRef]
- Johnson, M.H.; Bolhuis, J.J.; Horn, G. Interaction between acquired preferences and developing predispositions during imprinting. Animal Behav, 1985, 33, 1000–1006 https ://doiorg/101016/S0003. [Google Scholar] [CrossRef]
- Johnson, M.H.; Horn, G. Development of filial preferences in dark-reared chicks. Animal Behav, 1988, 36, 675–683 https ://doiorg/101016/S0003. [Google Scholar] [CrossRef]
- Miura, M.; Matsushima, T. Preference for biological motion in domestic chicks: sex-dependent effect of early visual experience. Animal Cog, 2012, 15, 871–879. [Google Scholar] [CrossRef]
- Miura, M.; Matsushima, T. Biological motion facilitates imprinting. Animal Behav, 2016, 116, 171–180. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Aoki, N.; Kitajima, T.; Iikubo, E.; Katagiri, S.; Matsushima, T.; Homma, K.J. Thyroid hormone determines the start of the sensitive period of imprinting and primes later learning. Nature Comm, 2012, 3, 1081. [Google Scholar] [CrossRef]
- Miura, M.; Aoki, N.; Yamaguchi, S.; Homma, K.-J.; Matsushima, T. Thyroid hormone sensitizes the imprinting-associated induction of biological motion preference in domestic chicks. Frontiers Physiol, 2018, 9, 1740. [Google Scholar] [CrossRef]
- Takemura, Y.; Yamaguchi, S.; Aoki, N.; Miura, M.; Homma, K.-J.; Matsushima, T. Gene expression of Dio2 (thyroid hormone converting enzyme) in telencephalon is linked with predisposed biological motion preference in domestic chicks. Behav Brain Res, 2018, 349, 25–30. [Google Scholar] [CrossRef]
- Lorenzi, E.; MLemaire, B.S.; Versace, E.; Matsushima, T.; Vallortigara, G. Resurgence of an inborn attraction for animate objects via thyroid hormone T3. Frontiers Behav Neurosci. 2021. [CrossRef]
- Miura, M.; Nishi, D.; Matsushima, T. Combined predisposed preferences for colour and biological motion make robust development of social attachment through imprinting. Animal Cog, 2020, 23, 169–188. [Google Scholar] [CrossRef]
- Moore, S.J.; Turnpenny, P.; Quinn, A.; Glover, S.; Lloyd, D.J. Montogomery T.; Dean J.C.S. A clinical study of 57 children with fetal anticonvulsant syndrome. J Med Genet, 2000, 37, 489–497. [Google Scholar] [CrossRef]
- Rasalam, A.D.; Hailey, H.; Moore, S.J.; Turnpenny, P.D.; Lloyd, D.J.; Dean, J.C.S. Characteristics of fetal anticonvulsant syndrome associated autistic disorder. Develop Med Child Neurol, 2005, 47, 551–555. [Google Scholar] [CrossRef]
- Christensen, J.; Grønborg, T.K.; Sørensen, M.J.; Schendel, D.; Parner, E.T.; Pedersen, L.H. Vestergaard M. Prenatal valproate exposure and risk of autism spectrum disorders and childhood autism. JAMA, 2013, 309, 1696–1703. [Google Scholar] [CrossRef]
- Nicolini, C.; Fahnestock, M. The valproic acid-induced rodent model of autism. Exp Neurol, 2018, 299, 217–227. [Google Scholar] [CrossRef]
- Nishigori, H.; Kagami, K.; Takahashi, A.; Tezuka, Y.; Sanbe, A.; Nishigori, H. Impaired social behavior in chicks exposed to sodium valproate during the last week of embryogenesis. Psychopharmacol, 2013, 227, 393–402. [Google Scholar] [CrossRef]
- Sgadò, P.; Rosa-Salva, O.; Versace, E.; Vallortigara, G. Embryonic exposure to valproic acid impairs social predispositions of newly-hatched chicks. Sci Rep, 2018, 8, 5919. [Google Scholar] [CrossRef]
- Lorenzi, E.; Pross, A.; Rosa-Salva, O.; Versace, E.; Sgadò, P.; Vallortigara, G. Embryonic exposure to valproic acid affects social predispositions for dynamic cues of animate motion in newly-hatched chicks. Frontiers Physiol, 2019, 10, 501. [Google Scholar] [CrossRef]
- Adiletta, A.; Pedrana, S.; Rosa-Salva, O.; Sgadò, P. Spontaneous visual preference for face-like stimuli is impaired in newly-hatched domestic chicks exposed to valproic acid during embryogenesis. Frontiers Behav Neurosci, 2021, 15, 733140. [Google Scholar] [CrossRef]
- Matsushima, T.; Miura, M.; Patzke, N.; Toji, N.; Wada, K.; Ogura, Y.; Homma, K.J. Sgadò P., Vallortigara G. Fetal blockade of nicotinic acetylcholine transmission causes autism-like impairment of biological motion preference in the neonatal chick. Cereb Cortex Comm. 2022. [CrossRef]
- Gean, P.-W.; Huang, C.-C.; Hung, C.-R.; Tsai, J.-J. Valproic acid suppresses the synaptic response mediated by the NMDA receptors in rat amygdalar slices. Brain Res Bull, 1994, 33, 333–336. [Google Scholar] [CrossRef]
- Winterer, G. Valproate and GABAergic system effects. Neuropsychopharmacol, 2003, 28, 2050–2051. [Google Scholar] [CrossRef] [PubMed]
- Phiel, C.J.; Zhang, F.; Huang, E.Y.; Guenther, M.G.; Lazar, M.A.; Klein, P.S. Histone deacetylase is a direct target of valproic acid, a potent anticonvulsant, mood stabilizer, and teratogen. J Biol Chem, 2001, 28, 36734–36741. [Google Scholar] [CrossRef] [PubMed]
- Bekoff, A.; Stein, P.S.G.; Hamburger, V. Coordinated motor output in the hindlimb of the 7-day chick embryo. PNAS, 1975, 72, 1245–1248. [Google Scholar] [CrossRef] [PubMed]
- Bekoff, A. Spontaneous embryonic motility: an enduring legacy. Internat J Develop Neurosci, 2001, 19, 155–160. [Google Scholar] [CrossRef]
- Blankenship, A.G.; Feller, M.B. Mechanisms underlying spontaneous patterned activity in developing neural circuits. Nature Rev Neurosci, 2010, 11, 18–29. [Google Scholar] [CrossRef]
- Rinaldi, T.; Kulangara, K.; Antoniello, K.; Markram, H. Elevated NMDA receptor levels and enhanced post-synaptic long-term potentiation induced by prenatal exposure to valproic acid. PNAS, 2007, 104, 13501–13506. [Google Scholar] [CrossRef]
- Uzunova, G.; Pallanti, S.; Hollander, E. Excitatory/inhibitory imbalance in autism spectrum disorders: implications for interventions and therapeutics. World J Biol Psychiatry, 2016, 17, 174–186. [Google Scholar] [CrossRef]
- Lee, E.; Lee, J.; Kim, E. Excitation/inhibition imbalance in animal models of autism spectrum disorders. Biol Psychiatry, 2017, 81, 838–847. [Google Scholar] [CrossRef]
- Kang, J.; Kim, E. Suppression of NMDA receptor function in mice prenatally exposed to valproic acid improves social deficits and repetitive behaviors. Frontiers Mol Neurosci. 2016. [CrossRef]
- Costas-Ferreira, C.; Faro, L.R.F. Neurotoxic effects of neonicotinoids on mammals: what is there beyond the activation of nicotinic acetylcholine receptors? - a systematic review. Int J Mol Sci, 2021, 22, 8413. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Foppen, R.P.B.; van Turnhaut, C.A.M.; de Kroon, H.; Jongejans, E. Declines in insectivorous birds are associated with high neonicotinoid concentration. Nature, 2014, 511, 341–343. [Google Scholar] [CrossRef]
- Eng, M.L.; Stutchbury, B.J.M.; Morrissey, C.A. Imidacloprid and chlorypyrifos insecticides impair migratory ability in a seed-eating songbird. Sci Rep, 2017, 7, 15176. [Google Scholar] [CrossRef]
- Eng, M.L.; Stutchbury, B.J.M.; Morrissey, C.A. A neonicotinoid insecticide reduces fueling and delays migration in songbirds. Science, 2019, 365, 1177–1180. [Google Scholar] [CrossRef]
- Keil, A.P.; Daniels, J.L.; Hertz-Picciotto, I. Autism spectrum disorder, flea and tick medication, and adjustments for exposure misclassification: the CHARGE (Childhood Autism Risks from Genetics and Envinronment) case-control study. Environ Health, 2014, 13, 3. [Google Scholar] [CrossRef]
- Gunier, R.B.; Bradman, A.; Harley, K.G.; Kogut, K.; Eskenazi, B. Prenatal residential proximity to agricultural pesticide use and IQ in 7-year-old children. Environ Health Perspect. 2017. [CrossRef]
- von Ehrenstein, O.S.; Ling, C.; Cui, X.; Cockburn, M.; Park, A.S.; Yu, F.; Wu, J.; Ritz, B. Prenatal and infant exposure to ambient pesticides and autism spectrum disorder in children: population based case-control study. the BMJ, 2019, 364, l962. [Google Scholar] [CrossRef]
- Ongono, J.S.; Béranger, R.; Baghdadli, A.; Mortamais, M. Pesticides used in Europe and autism spectrum disorder risk: can novel exposure hypotheses be formulated beyond organophosphates, organochlorines, organochlorines, pyrethroids and carbamates? – A systematic review. Environ Res, 2020, 187, 209646. [Google Scholar] [CrossRef]
- Sano, K.; Isobe, T.; Yang, J.; Win-Shwe, T.; Yoshikane, M.; Nakayama, S.F.; Kawashima, T.; Suzuki, G.; Hashimoto, S.; Nohara, K.; Tohyama, C.; Maekawa, F. In utero and lactational exposure to acetamiprid induces abnormalities in socio-sexual and anxiety-related behaviors of male mice. Frontiers Neurosci, 2016, 10, 228. [Google Scholar] [CrossRef] [PubMed]
- Román, G.C.; Ghassabian, A.; Bongers-Schokking, J.J.; Jaddoe, V.W.V.; Hofman, A.; de Rijke, Y.B.; Verhulst, F.C.; Tiemeier, H. Association of gestational maternal hypothyroxinemia and increased autism risk. Annals Neurol, 2013, 74, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Berbel, P.; Navarro, D.; Román, G.C. An evo-devo approach to thyroid hormones in cerebral and cerebellar cortical development: etiological implications for autism. Frontiers Endocrinol. 2014. [CrossRef] [PubMed]
- Getahun, D.; Jacobsen, S.J.; Fassett, M.J.; Wing, D.A.; Xiang, A.H.; Chiu, V.Y.; Peltier, M.R. 2018. Association between maternal hypothyroidism and autism spectrum disorders in children. Pediatrics Res. [CrossRef]
- Hoshiko, S.; Grether, J.K.; Windham, G.C.; Smith, D.; Fessel, K. Are thyroid hormone concentrations at birth associated with subsequent autism diagnosis? Autism Res, 2011, 4, 456–463. [Google Scholar] [CrossRef]
- Ames, J.L.; Windham, G.C.; Lyall, K.; Pearl, M.; Kharrazi, M.; Yoshida, C.K.; de Water, J.V.; Croen, L.A. Neonatal thyroid stimulating hormone and subsequent diagnosis of autism spectrum disorders and intellectual disability. Autism Res, 2020, 13, 444–455. [Google Scholar] [CrossRef]
- Batista, G.; Hensch, T.K. Critical period regulation by thyroid hormones: potential mechanisms and sex-specific aspects. Frontiers Mol Neurosci. 2019. [CrossRef]
- Westerholz, S.; de Lima, A.D.; Voigt, T. 2013. Thyroid hormone-dependent development of early cortical networks: temporal specificity and the contribution of trkB and mTOR pathways. Frontiers Cell Neurosci. [CrossRef]
- Batista, G.; Johnson, J.L.; Dominguez, E.; Costa-Mattioli, M.; Pena, J.L. Regulation of filial imprinting and structural plasticity by mTORC1 in newborn chickens. Sci Rep, 2018, 8, 8044. [Google Scholar] [CrossRef]
- Lorenzi, E.; Lemaire, B.S.; Versace, E.; Matsushima, T.; Vallortigara, G. Resurgence of an inborn attraction for animate objects via thyroid hormone T3. Frontiers Behav Neurosci. 2021. [CrossRef]
- Saheki, Y.; Aoki, N.; Homma, K.J.; Matsushima, T. Suppressive modulation of the chick forebrain network for imprinting by thyroid hormone: an in vitro study. Frontiers Physiol. 2022. [CrossRef]
- Sato, A.; Kasai, S.; Kobayashi, T.; Takamatsu, Y.; Hino, O.; Ikeda, K.; Mizuguchi, M. Rapamycin reverses impaired social interaction in mouse models of tuberous sclerosis complex. Nature Comm, 2012, 3, 1292. [Google Scholar] [CrossRef] [PubMed]
- Pagani, M.; Barsotti, N.; Bertero, A.; Trakoshis, S.; Ulysse, L. Locarno A. et al. Nature Comm, 2021, 12, 6084. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y. Excitatory actions of GABA during development: the nature of the nurture. Nature Rev Neurosci, 2002, 3, 728–739. [Google Scholar] [CrossRef]
- Represa, A.; Ben-Ari, Y. Tropic actions of GABA on neuronal development. Trends Neurosci, 2005, 28, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y.; Gaiarsa, J.L.; Tyzio, R.; Khazipov, R. GABA: A pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol Rev, 2007, 87, 1215–1284. [Google Scholar] [CrossRef]
- Pfeffer, C.K.; Stein, V.; Keating, D.J.; Maier, H.; Rinke I, Rudhard Y. et al. NKCC1-dependent GABAergic excitation drives synaptic network maturation during early hippocampal development. J Neurosci, 2009, 29, 3419–3430. [Google Scholar] [CrossRef]
- Hyde, T.M.; Lipska, B.K.; Ali, T.; Mathew, S.V.; Law, A.J.; Metitiri, O.E. Expression of GABA signaling molecules KCC2, NKCC1, and GAD1 in cortical development and schizophrenia. J Neurosci, 2011, 31, 11088–11095. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Neff, R.A.; Berg, D.K. Sequential interplay of nicotinic and GABAergic signaling guides neuronal development. Science, 2006, 314, 1610–1613. [Google Scholar] [CrossRef] [PubMed]
- Kahle, K.T.; Deeb, T.Z.; Puskarjov, M.; Silayeva, L.; Liang, B.; Kaila, K.; Moss, S.J. Modulation of neuronal activity by phosphorylation of the K-Cl cotransporter KCC2. Trends Neurosci, 2013, 36, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Kaila, K.; Price, T.J.; Payne, J.A.; Puskarjov, M.; Voipio, J. Cation-chloride cotransporters in neuronal development, plasticity and disease. Nature Rev Neurosci, 2014, 15, 637–654. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhou, J.; Zhang, Y.; Liu, T.; Friedel, P.; Zhu, W. The structural basis of function and regulation of neuronal cotransporters NKCC1 and KCC2. Comm Biol, 2021, 4, 226. [Google Scholar] [CrossRef]
- Marrosu, F.; Marrosu, G.; Rachel, M.G.; Biggio, G. Paradoxical reactions elicited by diazepam in children with classical autism. Funct Neurol, 1987, 2, 355–361. [Google Scholar] [PubMed]
- Nelson, S.B.; Valakh, V. Excitatory/inhibitory balance and circuit homeostasis in autism spectrum disorders. Neuron, 2015, 87, 684–698. [Google Scholar] [CrossRef]
- Delpire, E.; Ben-Ari, Y. A wholistic view of how bumetanide attenuates autism spectrum disorders. Cells, 2022, 11, 2419. [Google Scholar] [CrossRef]
- Hadjikhani, N.; Johnels, J.Å.; Lassalle, A.; Zürcher, N.R.; Hippolyte, L.; Gillberg, C.; Lemonnier, E.; Ben-Ari, Y. Bumetanide for autism: more eye contact, less amygdala activation. Sci Rep, 2018, 8, 3602. [Google Scholar] [CrossRef]
- Fernell, E.; Gustafsson, P.; Gillberg, C. Bumetanide for autism: open-label trial in six children. Acta Paediatrica, 2021, 110, 1548–1553. [Google Scholar] [CrossRef]
- Wang, T.; Shan, L.; Miao, C.; Zhida, X.; Jia, F. Treatment effect of bumetanide in children with autism spectrum disorder: a systematic review and meta-analysis. Frontiers Psychiat. 2021. [CrossRef] [PubMed]
- Sprengers, J.J.; van Andel, D.M.; Zuithoff, N.P.A.; Keijzer-Veen, M.G.; Schulp, A.J.A.; Scheepers, F.E.; Lilien, M.R.; Oranje, B.; Bruining, H. Bumetanide for core symptoms of autism spectrum disorder (BAMBI): a single center, double-blinded, participant-randomized, placebo-controlled, phase-2 superiority trial. J Am Acad Child Adolesc Psychiatry, 2021, 60, 865–876. [Google Scholar] [CrossRef] [PubMed]
- van Andel, D.M.; Sprengers, J.J.; Königs, M.; de Jonge, M.V.; Bruining, H. Effects of bumetanide on neurocognitive functioning in children with autism spectrum disorder: secondary analysis of a randomized placebo-controlled trial. J Autism Develop Disord. 2023. [CrossRef] [PubMed]
- Kéri, S.; Benedek, G. Oxytocin enhances the perception of biological motion in humans. Cog Affect Behav Neurosci, 2009, 9, 237–241. [Google Scholar] [CrossRef]
- Parker, K.J.; Oztan, O.; Libove, R.A.; Harden, A.Y. Intranasal oxytocin treatment for social deficits and biomarkers of response in children with autism. PNAS. 114, 8119-8124. [CrossRef]
- Freeman, S.M.; Palumbo, M.C.; Lawrence, R.H.; Smith, A.L.; Goodman, M.M.; Bales, K.L. Effect of age and autsm spectrum disorder on oxytocin receptor density in the human basal forebrain and midbrain. Transl Psychiat, 2018, 8, 257. [Google Scholar] [CrossRef]
- Nagasawa, M.; Mitsui, S.; En, S.; Ohtani, N.; Ohta, M.; Sakuma, Y.; Onaka, T.; Mogi, K.; Kikusui, T. Oxytocin-gaze positive loop and the coevolution of human-dog bonds. Science, 2015, 348, 333–336. [Google Scholar] [CrossRef]
- Kovács, K.; Kis, A.; Kanizsár, O.; Hernádi, A.; Gácsi, M.; Topál, J. The effect of oxytocin on biological motion perception in dogs (Canis familiaris). Anim Cog, 2016, 19, 513–522. [Google Scholar] [CrossRef]
- Duque, J.F.; Leichner, W.; Ahmann, H.; Stevens, J.R. Mesotocin influences pinyon jay prosociality. Biol Lett, 2018, 14, 20180105. [Google Scholar] [CrossRef]
- Seguchi, A.; Mogi, K.; Izawa, E.-I. Measurement of urinary mesotocin in large-billed crows by enzyme-linked immunosorbent assay. J Vet Med Sci, 2022, 84, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Loveland, J.; Stewart, M.; Vallortigara, G. Effects of oxytocin-family peptides and substance P on locomotor activity and filial preferences in visually naïve chicks. Eur J Neurosci, 2019, 50, 3674–3687 https ://doiorg/101111/ejn14520. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.R.; Carreira, L. Anbalagan S.; Blechman J.; Levkowitz G.; Oliveira R.F. Perceptual mechanisms of social affiliation in zebrafish. Sci Rep, 2020, 10, 3642. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Peterson, R.T. The zebrafish subcortical social brain as a model for studying social behavior disorders. Disease Models Mechanisms, 2019, 12, dmm039446. [Google Scholar] [CrossRef]
- Leonzino, M.; Busnelli, M.; Antonucci, F.; Verderio, C.; Mazzanti, M.; Chini, B. The timing of the excitatory-to-inhibitory GABA switch is regulated by the oxytocin receptor via KCC2. Cell Reports, 2016, 15, 96–103. [Google Scholar] [CrossRef]
- Hörnberg, H.; Pérez-Garci, E.; Schreiner, D.; Hatstatt-Burklé, L.; Magara, F.; Baudouin, S.; Matter, A.; Nacro, K.; Pecho-Vrieseling, E.; Scheiffele, P. Rescue of oxytocin response and social behaviour in a mouse model of autism. Nature, 2020, 584, 252–256. [Google Scholar] [CrossRef]
Figure 1.
Hourglass “bottleneck” model of the convergent evolution of peri-natal epigenetic control of social behaviors.
Figure 1.
Hourglass “bottleneck” model of the convergent evolution of peri-natal epigenetic control of social behaviors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



