
Submitted:
04 May 2023
Posted:
05 May 2023
You are already at the latest version
Abstract
Boron (B) toxicity is an important stress that negatively affects maize yield and quality production. The excessive B content in agricultural lands is a growing problem due to the increase in arid and semi-arid areas because of climate change. Recently, two Peruvian maize landraces, Sama and Pachía, were physiologically characterized based on their tolerance to B toxicity, the former being more tolerant to B excess than Pachía. However, many aspects regarding the molecular mechanisms of these two maize landraces against B toxicity are still unknown. In this study, a leaf proteomic analysis of Sama and Pachía was performed. Out of a total of 2793 proteins identified only 303 proteins were differentially accumulated. Functional analysis indicated that many of these proteins are involved in transcription and translation processes, amino acids metabolism, photosynthesis, carbohydrate metabolism, protein degradation, and protein stabilization and folding. Compared to Sama, Pachía had a higher number of differentially expressed proteins related to protein degradation, and transcription and translation processes under B toxicity conditions, which might reflect the greater protein damage caused by B toxicity in Pachía. Our results suggest that higher tolerance to B toxicity of Sama can be attributed to more stable photosynthesis that would avoid damage caused by stromal over-reduction under this stress condition.
Keywords:
boron toxicity
; proteomic analysis
; maize landrace
; Zea mays
1. Introduction
Boron (B) is an essential element for plants being well known its structural role in both cell walls and membranes [1,2,3,4,5]. Actually, B establishes diester bonds between apiose residues of two rhamnogalacturonan-II (RGII) molecules forming RGII-B complexes that stabilize the pectin network of the cell wall [6,7,8]. Furthermore, B contributes to the preservation of plasmalemma integrity and function [9], likely through the formation of B complexes with membrane components that contain cis-diol groups [10,11]. Thereby, B forms complexes with major constituents of membrane lipid rafts, such a as glycosyl inositol phosphoryl ceramides (GIPCs) [12]. Moreover, B participates in the formation of GIPCs-B-RGII complexes, which connect the plasmalemma to the cell wall [13]. Besides these structural roles, B is also involved in plant development participating in root and shoot elongation, pollen-tube growth, flowering, and fruiting [14,15,16]. In addition, B has been reported to participate in several physiological processes, such as photosynthesis, nucleic acid synthesis, phenolic, nitrogen and polyamines metabolisms, proteins stabilization and biosynthesis, and gene expression, among others [16,17,18,19,20,21,22].
Since B is a micronutrient, the range between its deficient, optimal, and toxic concentrations for plants is very narrow [23]. Therefore, it is common to find soils with inadequate B content for optimal plant development. Soils with high B contents predominantly occurs in arid and semi-arid countries, where this micronutrient accumulates in the topsoil mainly owing to high evapotranspiration and tiny leaching caused by low rainfall, a situation that is often aggravated by irrigation with B-enriched water [22,23]. Additionally, excess B is also found in lands close to coastal areas due to the hydraulic connection between their coastal aquifers and seawater [24] or in regions with recurrent geothermal activities [2]. Climate change is another factor that is contributing to the B increase in soils. Increasing temperatures and decreasing rainfall are predicted in the coming years, which will lead to an increase in agricultural areas with excessive B levels [3,25].
Excessive B contents in soils cause adverse effects such as chlorosis and necrosis in leaves, damages to stems and buds, and misshapen fruits [17,22]. Furthermore, an excess of B provokes DNA damages, inhibition of protein folding, impairment of protein functions and activities, and alterations in photosynthesis and nitrogen and carbon metabolisms, among other processes [2,22,26]. In fact, several photosynthetic parameters, such as CO2 assimilation (PN), photosynthetic electron transport rate (ETR), maximum quantum yield of chlorophyll fluorescence (Fv/Fm), and CO2 use efficiency decreased under B toxicity conditions [22,27]. Because of the aforementioned effects of B toxicity in plant physiology, elevated B contents in agricultural lands reduce crop growth, yield, and quality [22,28]. In fact, a noteworthy decrease in the yield of several main crops subjected to B toxicity has been reported [28]. Despite the large number of effects caused by B toxicity in plants, it is not well known how B produces these alterations. However, it has been suggested that the ability of B to form bonds with molecules containing mono-, di- and poly-hydroxyl groups could be the chemical basis by which B toxicity could trigger morphophysiological alterations [29].
Maize is an important crop that provides approximately half of the calories consumed worldwide being, in addition, one of the principal genetic model plants for crop improvement and food security [30,31,32]. However, maize production is seriously constrained by abiotic and biotic stresses [33]. In particular, B toxicity causes a decrease in maize production as well as in other cereals [28,34,35]. Therefore, the search and molecular characterization of new maize varieties with improved tolerance to B toxicity has become an interesting research topic. In a recent work, two Peruvian maize landraces (Pachía and Sama) were tested for tolerance to high B. The Sama landrace had greater tolerance to B excess than Pachía [27]. In this work, a comparative proteomic characterization of these two maize landraces with different tolerance to B toxicity was performed to improve our molecular knowledge about which proteins are involved in B-toxicity tolerance.
2. Results
A total of 2793 proteins were identified in at least one of the biological replicates of a landrace (Sama or Pachía) and a B treatment analyzed (Table S1a). In addition, the number of proteins detected in both Pachía and Sama in each of the B treatments studied was similar being close to 1100 proteins (Table 1).
Table S1a shows the dataset of the identified proteins indicating their gene ontology (GO) biological processes (GOBP), GO molecular functions (GOMF), and GO cellular compartments (GOCC), and Table S1b summarizes the statistical analysis and fold changes of the proteins. To study the differentially accumulated proteins in Pachía and Sama in both B treatments, four comparison groups were established: 1) Sama and Pachía seedlings subjected to the control B condition (S0.05/P0.05), 2) Sama and Pachía treated with 10 mM B (S10/P10), 3) Sama subjected to 10 mM and 0.05 mM B (S10/S0.05), and 4) Pachía treated with 10 mM and 0.05 mM B (P10/P0.05). A total of 303 proteins had statistically significant differential expression (P ≤ 0.05) in the above groups (Table S2). The S0.05/P0.05 and S10/P10 groups contain those proteins that were differentially expressed between Sama and Pachía in 0.05 mM or 10 mM B, respectively. In media with 0.05 mM B, more proteins were up- and down-accumulated between Sama and Pachía than in 10 mM B (Figure 1 and Table 1). In addition, the S10/S0.05 and P10/P0.05 comparison groups included proteins that were differentially expressed in response to B toxicity in Sama or Pachía, respectively. Pachía had a higher number of proteins induced and repressed by B toxicity than Sama, thus 98 proteins were up-expressed in Pachía in 10 mM B while only 38 in Sama and 51 proteins were down-expressed in Pachía under B toxicity versus 28 in Sama (Figure 1).
2.1. Classification into several functional categories of differentially accumulated proteins in both maize landraces and B treatments
All significant differentially expressed proteins in the four comparison groups described above were functionally classified into 26 categories using several databases (Table S2). The functional categories that included the largest number of differentially accumulated proteins were transcription and translation processes (57), photosynthesis (25), amino acid metabolism (24), protein degradation (23), carbohydrate metabolism (20), and protein stabilization and folding (18) (Figure 2 and Table S2). These main categories together contained more than 50% of the total differentially expressed proteins.
2.2. Differentially expressed proteins in Sama and Pachía in response to B toxicity
Considering that the major aim of this work was to analyze the changes provoked by B toxicity on protein expressions in Pachía and Sama, we will now focus on the proteins that were differentially expressed by B toxicity in these landraces. Thus, 66 and 149 proteins were differentially expressed in response to B toxicity in Sama and Pachía, respectively (Table 1). The main functional categories containing the highest number of differentially expressed proteins under B toxicity in both Sama and Pachía were transcription and translation, photosynthesis, amino acid metabolism, protein degradation, protein stabilization and folding, and reactive oxygen species (ROS) (Figure 3 and Figure 4). Interestingly, most of the proteins belonging to the transcription and translation category were induced in response to B toxicity in both Sama and Pachía, the number of differentially induced proteins being remarkably higher in Pachía (Figure 3 and Figure 4). However, almost all proteins included in the photosynthesis category were repressed in 10 mM B, the number of down-accumulated proteins being also higher in Pachía than in Sama (Figure 4 and Table S2). Regarding protein degradation, and protein stabilization and folding, most of the differentially expressed proteins in 10 mM B were found in Pachía, suggesting that B toxicity would alter the structure and folding of proteins in this landrace. In addition, many of the proteins in the ROS category were induced by B toxicity in both landraces (Figure 4 and Table S2). Although the groups of carbon assimilation and metabolism, lipid metabolism, and respiration included a smaller number of proteins that those mentioned above, nevertheless, a larger number of differentially expressed proteins were found in Pachía under B toxicity (Figure 4 and Table S2). Other interesting categories were cell death, cell division, cell wall, ribosome biogenesis, and RNA binding and processing which, despite having a very small number of proteins regulated by B toxicity, had an interesting distribution in both landraces and B treatments. In fact, in the cell death and cell wall categories, only proteins whose expressions were induced by B toxicity were found in Pachía, however, the cell division, ribosome biogenesis, and RNA binding and processing categories also contained proteins with higher accumulation in 10 mM B but in both landraces (Figure 4 and Table S2).
A total of 18 proteins were commonly expressed (repressed or induced) in both landraces in response to B toxicity, with the amino acid metabolism and photosynthesis categories having the highest number of proteins (Table 2). All proteins of the amino acid metabolism group were up-accumulated under B toxicity conditions, with these inductions being slightly greater in Pachía than in Sama. Interestingly, however, all commonly expressed proteins from the photosynthesis category were repressed by B toxicity, these repressions being remarkably higher in Pachía than in Sama (Table 2).
Table 3 and Table 4 list the most strongly differentially expressed proteins that were up- or down-regulated more than twofold by B toxicity in Pachía and Sama, respectively. In Pachía, 105 proteins had strong differential expression under B toxicity, while only 27 were found in Sama. Photosynthesis was the functional category containing the highest number of proteins whose expressions were strongly down-accumulated in response to B toxicity in both Pachía and Sama, however, interestingly, both minor number of repressed and very strongly repressed (FC <0.33) proteins were observed in Sama (Tables 3,4, and S2). Different subunits of the NDH complex (NDHS, B1, B2, J, and H) were strongly repressed by B toxicity in Pachía but not in Sama (Tables 3, 4, and S2). In addition, only in Pachía were detected proteins related to protein degradation processes whose expressions were mainly induced by B toxicity suggesting that enhanced damage would be provoked by 10 mM B in Pachía proteins (Table 3). Furthermore, B toxicity markedly induced a larger number (15) of proteins in Pachía belonging to the transcription and translation category (Table 3).
Table 5 shows the proteins that were strongly up- or down-accumulated when protein expressions of Sama were compared to those of Pachía in media with 10 mM B. Sama had a remarkable up-accumulation of four proteins involved in photosynthesis (ZmPIFI and OEE2-1), chlorophyll biosynthesis (ChlH1), and secondary metabolism (PAO1) being, in addition, this last protein strongly induced in response to B toxicity (Table 4 and Table 5). However, in Pachía several proteins were detected with a strong accumulation in 10 mM B when compared with Sama (shown in Table 5 as strongly down-accumulated proteins in Sama) highlighting, among them, histone H1 and ribosomal protein S7 which, besides, were strongly induced by B toxicity (Table 3 and Table 5).
Finally, in both Pachía and Sama, proteins exclusively detected in one of these landraces were found, among them, Nfc103a and eIF3a, which were only identified in Pachía in 10 mM B (Table 6).
3. Discussion
Although 2793 proteins were detected in this proteomic analysis, only 303 proteins were differentially accumulated (Tables S1a and S2), which were classified into 26 functional categories. Functional analysis indicated that pathways involved in transcription and translation processes, amino acids metabolism, photosynthesis, carbohydrate metabolism, protein degradation, and protein stabilization and folding were highly enriched categories in both landraces (Figure 2). Remarkably, the expression levels of proteins related to these enriched processes were significantly different between Pachía and Sama.
3.1. Several proteases and translation-related proteins would allow Pachía to survive in media with B excess
Pachía is a B-sensitive maize cultivar described by Mamani-Huarcaya et al. [27]. Interestingly, the highest number of differentially accumulated proteins (DAPs) was found in the comparison group P10/P0.05 (Figure 1) suggesting that the B toxicity damage caused in Pachía could be partially relieved by these proteins. A remarkable number of these DAPs included in the categories of protein degradation (11), and transcription and translation (15) were strongly overexpressed in Pachía (Table S2 and 3). However, only four proteins of the transcription and translation group were markedly induced by 10 mM B in Sama (Table 4). The B-sensitive Citrus grandis had a higher number of proteins involved in protein degradation that was also overexpressed under B toxicity conditions in comparison with B-tolerant Citrus sinensis [36]. These authors concluded that B toxicity caused greater protein damage and proteolysis in C. grandis. Therefore, the high number of protein degradation-related proteins that were overexpressed in Pachía in 10 mM B would suggest that B toxicity would cause greater damage in Pachía proteins than in those of Sama leading to increased proteolysis in B-sensitive Pachía. Proteins related to protein degradation strongly overexpressed in Pachía included, among others, cysteine protease14 and four serine proteases (Table 3). Proteases have been implicated in plant acclimation to abiotic stress, playing a major role in the degradation of damaged and misfolded proteins, thus contributing to cell survival. In fact, cysteine and serine proteases are involved in degradation of misfolded proteins and protection against abiotic stresses [37,38,39,40]. Hence, these five proteases could have a main role in the degradation of damaged and misfolded proteins in Pachía under excess B, contributing to maintaining the correct conformation of Pachía proteins and, therefore, to the survival of this landrace under this stressful condition. In addition, a noteworthy number of proteins involved in transcription and translation processes were overexpressed at 10 mM B in Pachía, namely, 30 in contrast to only nine of Sama (Table S2). Proteomic analysis performed with dehydration, salt, and temperature stresses in cereals also displayed alterations in the levels of translation-related proteins, such as initiation factors and the ribosome constituent proteins ([41] and references therein). Furthermore, it has been suggested that a B excess provokes inhibition of RNA-dependent processes, such as transcription and translation, owing to the ability of B to form complexes with ribose molecules [42]. In this regard, Tanaka et al. [43] have suggested that B or boric acid acts on the translation machinery likely forming complexes with cis-diol groups of rRNA and tRNA. In addition, it has currently been proposed that high-B stress enhances ribosome frequency on stop codons leading to a global ribosome stalling [44]. Consequently, the high contents of leaf-soluble B in Pachía seedlings subjected to 10 mM B reported by Mamani-Huarcaya et al. [27] would generate an increased formation of B complexes with cis-diol groups of RNA that would damage ribosomes leading to a drop in protein synthesis likely through a global ribosome stalling. The strong overexpression of several ribosomal proteins would maintain the Pachía ribosome stability in B toxicity (Tables S2 and 3). These results are consistent with those reported for rice, where several ribosomal protein large subunit genes were upregulated under temperature stress, suggesting that their encoded proteins might be involved in stress amelioration, likely maintaining the proper functioning of ribosomes [41]. Interestingly, the eukaryotic translation initiation factor 3 subunit A (eIF3a) was exclusively detected in B toxicity in Pachía (Table 6). These factors are one of the most significant components involved in plant protein synthesis and, specifically, rice eIF3A has been proposed to play an important role in different stresses [45]. Therefore, eIF3a would also help to alleviate the drop in protein synthesis in Pachía. Thereby, Pachía would partly ameliorate injuries caused by B toxicity on protein synthesis and ribosome by overexpressing a high number of transcription- and translation-related proteins, abolishing a non-viable reduction of transcription and translation processes.
3.2. Proteins that would confer Sama more B toxicity tolerance
Polyamine oxidase 1 (PAO1) is an interesting protein that was clearly up-accumulated in Sama when compared to Pachía at 10 mM B and was also strongly induced in Sama by B toxicity (Table 4 and Table 5). This enzyme catalyzes the back conversion of spermine (Spm) to spermidine (Spd), and Spd to putrescine (Put) [46]. Maize polyamines play a crucial role in abiotic stress response [33]. In fact, it has been reported that Put protects the plant photosynthetic apparatus against several abiotic stresses [47]. Moreover, the conjugation of Put to PSII proteins may lead to the structural and functional stability of PSII [46,48]. Therefore, the over-accumulation of PAO1 in Sama plants subjected to B toxicity would generate an increase in Put levels that would protect their photosynthetic apparatus resulting in the higher PN observed in Sama under this stress, as described by Mamani-Huarcaya et al. [27]. This finding is consistent with results reported for Karoon, a drought-tolerant maize cultivar. Pakdel et al. [46] proposed that higher expression of PAO genes and enzymatic polyamine oxidation activity would protect the photosynthetic apparatus of Karoon under water stress.
3.2.1. Lower repression of photosynthesis-related proteins would enhance the B-toxicity tolerance of Sama
Photosynthesis is one of the essential physiological processes affected by B toxicity [2,22]. Photosynthetic efficiency could be achieved in Sama under B toxicity conditions increasing the synthesis of photosynthetic pigments, since chlorophyll content is a major limiting component of the photosynthetic efficiency [49]. Interestingly, Sama had a strong over-accumulation of magnesium-chelatase subunit H1 chloroplastic (ChlH1) at 10 mM B in comparison with those from Pachía (Table 5). ChlH binds to porphyrin and catalyzes the insertion of Mg2+ into protoporphyrin IX [50]. Accordingly, the over-accumulation of ChlH1 in Sama would explain its higher contents of chlorophyll a in B toxicity and the higher PN described by Mamani-Huarcaya et al. [27].
In this study, 25 proteins related to photosynthetic light reactions were differentially accumulated, most of them involved in electron transport, light harvesting, and oxygen evolving processes (Table S2). Pachía and Sama presented several photosynthesis-related proteins that were repressed by B toxicity when their expressions were compared with those of Pachía and Sama, respectively, in media with 0.05 mM B (Table S2). However, the number of these DAPs was lower in Sama than in Pachía (11 versus 16, respectively; Table S2) and, besides, those proteins commonly down-accumulated in both landraces had a weaker decrease in Sama (Table 2). In addition, only two photosynthetic proteins were strongly down-expressed 3-fold or more (corresponding to FC≤ 0.33) by B toxicity in Sama in contrast to ten proteins found in Pachía (Table 3 and Table 4). This decreased accumulation of photosynthesis related-proteins may cause lower photosynthetic performance in B-toxicity-treated Pachía plants than in Sama plants, as described by Mamani-Huarcaya et al. [27]. Therefore, Sama would retain sufficient levels of photosynthesis-related proteins in 10 mM B, which would allow it to maintain photosynthetic parameters at similar levels to those of the control conditions, as reported by Mamani-Huarcaya et al. [27]. Furthermore, three photosynthesis-related proteins were up-accumulated in Sama when their expressions were compared with those of Pachía in 10 mM B, namely, oxygen-evolving enhancer protein 2-1 chloroplastic (OEE2-1), post-illumination chlorophyll fluorescence increase (ZmPIFI), and NAD(P)H-quinone oxidoreductase subunit S chloroplastic (NDHS) (Tables 5 and S2). OEE2-1 is likely an extrinsic protein of the oxygen-evolving complex (OEC) (UniProt; https://www.uniprot.org/). The OEC is stabilized and protected by extrinsic polypeptides [51]. The strong OEE2-1 over-accumulation in 10 mM B in Sama could facilitate the stability and protection of the OEC leading to the higher photosynthetic electron transporter rate (ETR) observed in this landrace [27]. Regarding ZmPIFI, it is homologous to the PIFI protein of Arabidopsis thaliana (AtPIFI), an essential component of the NAD(P)H dehydrogenase (NDH) complex involved in chlororespiratory electron transport around PSI [52]. The Atpifi mutant had a lower nonphotochemical quenching (NPQ) than wild type under high light irradiances, suggesting that AtPIFI would protect plants from photooxidative stress triggered by excessive light [52]. Consequently, both ZmPIFI over-accumulation and the higher NPQ values that Sama showed in 10 mM B, unlike those from Pachía (Table 5; [27]), suggest that ZmPIFI would also be a component of the maize NDH complex playing a role in oxidative photoprotection of this landrace under B-toxicity conditions. Furthermore, unlike Sama, several subunits of the NDH complex were markedly repressed in Pachía by B toxicity (Tables S2, 3, and 4). The NDH complex mediates cyclic electron transport around PSI playing a crucial role in C4 photosynthesis [53,54]. NDH-mediated cycle electron transport (NDH-CET) performs two functions: 1) maintaining photosynthetic redox balance in the electron transfer avoiding stromal overreduction and functioning as a safety valve for excess electrons under stress, and 2) supplying ATP for efficient carbon assimilation, especially under stressful conditions [53,54,55,56]. The finding that none of the above components of the NDH complex was significantly repressed by B-toxicity in Sama suggests that its NDH-CET would prevent stromal overreduction and would protect against photooxidation. This fact would explain the high values of net photosynthetic CO2 assimilation (PN), maximum photochemical efficiency (Fv’/Fm’), and quantum yield efficiency of PSII electron transport (ΦPSII) reported in Sama at 10 mM B, which were similar to those of control conditions [27]. Consistent with our data, Zhu et al. [56] have suggested that an increased abundance of NDH subunits in salt-stressed wheat would enhance NDH-CET alleviating the accumulation of excess electrons and maintaining energy homeostasis. Moreover, the subunit S of the NDH complex was over-accumulated in Sama under B toxicity when compared to those from Pachía, leading to a likely higher amount of NDH-complex that would provide extra ATP to achieve better PN and growth at this landrace in media with 10 mM B as, in fact, was observed by Mamani-Huarcaya et al. [27]. In addition, a higher supply of ATP could be obtained in Sama in comparison to Pachía under B toxicity from a weaker decrease of the α- and β-chloroplastic subunits of ATP synthase in Sama (Table 2). Although B excess causes photosynthetic damage [2,22], plants have evolved mechanisms to repair these injuries that require a high amount of ATP from chloroplastic ATP synthase [57,58]. In Sama, B toxicity barely affected photosynthetic parameters [27]. This finding points out that this landrace would own mechanisms to repair its photosynthetic machinery. Likely, one of these mechanisms would be to provide greater ATP availability, which would be achieved by maintaining sufficient levels of NDH and ATP synthase complexes that would synthesize the amounts of ATP needed to repair its photosynthetic machinery and, therefore, to maintain its photosynthetic values at levels similar to those of control conditions.
4. Materials and Methods
4.1. Plant materials and growth conditions
Sama and Pachía, two Peruvian maize landraces from the Sama valley and the Pachía district (to the east of Tacna), were used in this study. Seeds were surface-sterilized as described by Mamani-Huarcaya et al. [27]. Afterwards, the seeds were placed in seedbeds filled with a perlite/vermiculite mixture (1/1, v/v) and watered with deionized H2O. After seven days, seedlings were transplanted to 30-L plastic containers with a nutrient solution (NS) that was identical to the one used by Mamani-Huarcaya et al. [27]. After two days of acclimation to hydroponic medium, the seedlings were divided into groups and transferred to fresh NS supplemented with 10 mM H3BO3 (B toxicity conditions) or 0.05 mM H3BO3 (control conditions). This medium was aerated by air pumps and renewed twice a week. The seedlings were germinated and grown hydroponically in a growth chamber under a 12 h light/12 h dark regime (215 µmol m–2 s–1 of photosynthetically active radiation at plant height), at 22ºC and 50% relative humidity. The plants were randomly harvested 10 days after the onset of the B treatments and their leaves were quickly separated with a scalpel, frozen in liquid nitrogen and stored at –80°C until further analysis.
4.2. Protein extraction and digestion
Maize leaves (200-250 mg fresh weight) from four separate seedlings per condition (B treatment and maize landrace) were ground to a fine powder in a mortar precooled with liquid nitrogen. Proteins were extracted with trichloroacetic acid (TCA)/acetone-phenol [59], solubilized in a solution containing 7 M urea, 2 M thiourea and 2% (w/v) CHAPS (3 [(3-cholamidopropyl) dimethylammonium]-1-propanesulfonate), and quantified by the Bradford method using bovine serum albumin (BSA) as a standard [60].
The cleaning of maize protein extract, protein digestion, and mass spectrometry determinations were carried out at the Proteomics Facility for Research Support Central Service (SCAI) of the University of Córdoba (Spain) as follows.
Biological quadruplicate samples were separated and cleaned as described. Leaf protein extracts (50 µg of BSA protein equivalents per sample) were electrophoretically pre-concentrated in a centimeter band of 10% (w/v) SDS-PAGE gel. Protein bands were excised from the gels and, afterwards, the gel pieces were distained in 200 mM ammonium bicarbonate/50% acetonitrile for 15 min, followed by 5 min in 100% acetonitrile. Proteins were reduced by addition of 20 mM dithiothreitol in 25 mM ammonium bicarbonate and incubated for 20 min at 55 °C. The mixture was cooled to room temperature and then free thiols were alkylated by adding 40 mM iodoacetamide in 25 mM ammonium bicarbonate for 20 min in the dark. Finally, the gel pieces were washed twice in 25 mM ammonium bicarbonate.
Proteolytic digestion was performed by addition of trypsin to a final concentration of 12.5 ng/µL in 25 mM ammonium bicarbonate at 37 ºC overnight. Protein digestion was stopped by adding trifluoroacetic acid at a final concentration of 1% (v/v). Finally, the digested samples were vacuum-dried and dissolved in a mixture of 2% (v/v) acetonitrile and 0.05% (v/v) trifluoroacetic acid.
4.3. Shotgun-DDA-LC-MS/MS analysis
Peptide separations were performed on a nano-LC using Dionex Ultimate 3000 nano UPLC (Thermo Scientific, San Jose, CA, USA), equipped with a C18 75 μm × 50 cm Acclaim Pepmap column (Thermo Scientific, San Jose, CA, USA), at 40 °C at a flow rate of 300 nL/min. Peptide mixtures were previously concentrated and cleaned up on a 300 µm x 5 mm Acclaim Pepmap precolumn (Thermo Scientific, San Jose, CA, USA) using 2% acetonitrile/0.05% trifluoroacetic acid at 5 µL/min for 5 min. Peptides were eluted with a gradient of 60 min ranging from 96% solvent A (0.1% formic acid) to 90% solvent B (80% acetonitrile and 0.1% formic acid), followed by an 8 min wash at 90% solvent B and a 12 min re-equilibration at 4% solvent B. Eluted peptides were converted to gas-phase ions by nanoelectrospray ionization and analyzed on a Thermo Orbitrap Fusion mass spectrometer (Thermo Scientific, San Jose, CA, USA) operated in the positive mode. Survey scans of peptide precursors were acquired over the m/z range 400−1500 at 120K resolution (at 200 m/z) with a 4 × 105 ion count target. Tandem MS was performed by isolation at 1.2 Da with the quadrupole. Monoisotopic precursor ions were fragmented by CID (Chemically Induced Dimerization) in the ion trap, which was set up as follows: automatic gain control, 2 × 103; maximum injection time, 50 ms; and normalized collision energy of 35%. Only those precursors with charge state 2–5 were sampled for MS2. A dynamic exclusion time of 15 s and a tolerance of 10 ppm around the selected precursor and its isotopes were used to avoid redundant fragmentations. The instrument was run in top 30 mode with 3-s cycles, meaning the instrument would continuously perform MS2 events until a maximum of top 30 non-excluded precursors or 3 s, whichever was shorter.
4.4. Protein quantification
Charge state deconvolution and deisotoping were not performed. MS2 spectra were searched using MaxQuant software v. 1.5.7.4 [61]. MS2 spectra were searched with Andromeda engines against a database of Uniprot Zea mays_Jun19. Peptides generated from tryptic digestion were searched employing the following parameters: up to one missed cleavage, carbamidomethylation of cysteines as fixed modifications, and oxidation of methionine as variable modifications. The precursor mass tolerance was 10 ppm and product ions were searched at 0.6 Da tolerances. A target-decoy search strategy was applied, which integrates multiple peptide parameters such as length, charge, number of modifications, and identification score into a single quality that acts as statistical evidence on the quality of each single peptide spectrum match. The identified peptides were grouped into proteins according to the law of parsimony and filtered to 1% false discovery rate (FDR). Peptide quantification was carried out using MaxQuant software, in a MaxLFQ label-free quantification method [62]. In the MaxLFQ label-free quantification method, a retention time alignment and identification transfer protocol (“match-between runs” feature inMaxQuant) was applied. Proteins identified from only one peptide were not taken into account in this analysis. Peak intensities across the whole set of quantitative data for all peptides in the samples were imported from the LFQ intensities of proteins from the MaxQuant analysis and normalized according to Cox et al. [62]. LFQ normalized intensity values were transformed to a logarithmic scale with base two. Protein quantification and calculation of statistical significance were carried out using Student-t test and error correction (P-value ≤ 0.05). The criteria used to consider a protein as differentially expressed were as follows: (a) the protein was consistently present in at least three biological replicates per condition; (b) it had statistically significant differences (Student-t test, P ≤ 0.05) between genotypes or B treatments; and (c) a fold change ≥ 1.5 or ≤ 0.66667. The differentially accumulated proteins were manually categorized by function using different databases (Uniprot, https://www.uniprot.org/; Maize Genetics and Genomics, https://www.maizegdb.org/; ExplorEnz, https://www.enzyme-database.org/; BRENDA, https://www.brenda-enzymes.org/; KEGG: Kyoto Encyclopedia of Genes and Genomes, https://www.genome.jp/kegg/; and PANTHER: Protein ANalysis THrough Evolutionary Relationships, http://pantherdb.org/).
5. Conclusions
Overexpression of several proteases and transcription- and translation-related proteins would allow Pachía to degrade and replace partially the proteins damaged by B toxicity achieving survival under this stress condition.
In Sama, PAO1 over-accumulation and weaker knockdown of several subunits of NDH and ATP synthase complexes under B excess would confer a greater B toxicity tolerance to this landrace by: 1) acting as an electron safety valve that would avoid stromal overreduction, and thus decrease photosynthetic damage and, 2) providing an additional supply of ATP that would contribute to repair the photosynthetic system of Sama.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1a: Dataset of proteins identified by shotgun-DDA analysis; Table S1b: Dataset, statistical analysis and fold change of proteins identified by shotgun-DDA analysis; Table S2: Fold change ratios, P-values and statistical significances of all significantly accumulated proteins classified by functional categories.
Author Contributions
Plant growth and harvesting, and protein extraction and quantification, B.M.M.-H. with substantial contribution of M.T.N.-G. and J.R.; research planning and design, J.R. with the help of all authors; analysis and interpretation of data, J.R.; writing - first draft, J.R.; editing and review - second draft, J.R. and A.G.-F. All authors critically revised the manuscript and made significant contributions. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Agencia Andaluza de Cooperación Internacional para el Desarrollo (Consejería de Igualdad y Políticas Sociales) from Junta de Andalucía (2016SEC014), and by the Consejería de Economía, Innovación, Ciencia y Empleo from Junta de Andalucía (BIO-266).
Data Availability Statement
The data presented in this study are available in the text and supplemental data.
Acknowledgments
The authors would like to thank SCAI of the University of Córdoba (Spain) for protein identification and, especially, to Carlos Fuentes Almagro his skillful technical assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Warington, K. The effect of boric acid and borax on the broad bean and certain other plants. Ann. Bot. 1923, 37, 629–672. [Google Scholar] [CrossRef]
- Princi, M.P.; Lupini, A.; Araniti, F.; Longo, C.; Mauceri, A.; Sunseri, F.; Abenavoli, M.R. Boron toxicity and tolerance in plants: recent advances and future perspectives. In Plant Metal Interaction: Emerging Remediation Techniques; Ahmad, P., Ed.; Elsevier Inc: Amsterdam, Netherlands, 2016; pp. 115–147. [Google Scholar]
- González-Fontes, A.; Fujiwara, T. Advances in plant boron. Int. J. Mol. Sci. 2020, 21, 4107. [Google Scholar] [CrossRef]
- González-Fontes, A. Why boron is an essential element for vascular plants. New Phytol. 2020, 226, 1228–1230. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Abreu, I.; Bell, R.W.; Bienert, M.D.; Brown, P.H.; Dell, B.; Fujiwara, T.; Goldbach, H.E.; Lehto, T.; Mock, H.-P.; et al. Boron: an essential element for vascular plants. New Phytol. 2020, 226, 1232–1237. [Google Scholar] [CrossRef]
- Ishii, T.; Matsunaga, T. Isolation and characterization of a boron-rhamnogalacturonan-II complex from cell walls of sugar beet pulp. Carbohydr. Res. 1996, 284, 1–9. [Google Scholar] [CrossRef]
- Kobayashi, M.; Matoh, T.; Azuma, J. Two chains of rhamnogalacturonan II are cross-linked by borate-diol ester bonds in higher plant cell walls. Plant Physiol. 1996, 110, 1017–1020. [Google Scholar] [CrossRef]
- O’Neill, M.A.; Warrenfeltz, D.; Kates, K.; Pellerin, P.; Doco, T.; Darvill, A.G.; Albersheim, P. Rhamnogalacturonan-II, a pectic polysaccharide in the walls of growing plant cell, forms a dimer that is covalently cross-linked by a borate ester. J. Biol. Chem. 1996, 271, 22923–22930. [Google Scholar] [CrossRef]
- Cakmak, I.; Römheld, V. Boron deficiency-induced impairments of cellular functions in plants. Plant Soil 1997, 193, 71–83. [Google Scholar] [CrossRef]
- Goldbach, H.E.; Yu, Q.; Wingender, R.; Schulz, M.; Wimmer, M.; Findeklee, P.; Baluška, F. Rapid response reactions of roots to boron deprivation. J. Plant Nutr. Soil Sci. 2001, 164, 173–181. [Google Scholar] [CrossRef]
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Voxeur, A.; Fry, S.C. Glycosylinositol phosphorylceramides from Rosa cell cultures are boron-bridged in the plasma membrane and form complexes with rhamnogalacturonan II. Plant J. 2014, 79, 139–149. [Google Scholar] [CrossRef]
- Wang, N.; Yang, C.; Pan, Z.; Liu, Y.; Peng, S. Boron deficiency in woody plants: various responses and tolerance mechanisms. Front. Plant Sci. 2015, 6, 916. [Google Scholar] [CrossRef] [PubMed]
- Blevins, D.G.; Lukaszewski, K.M. Boron in plant structure and function. Annu. Rev. Plant Physiol. Plant Molec. Biol. 1998, 49, 481–500. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Cristóbal, J.J.; Navarro-Gochicoa, M.T.; Rexach, J.; González-Fontes, A.; Herrera-Rodríguez, M.B. Plant response to boron deficiency and boron use efficiency in crop plants. In Plant Micronutrient Use Efficiency. Molecular and Genomic Perspectives in Crop Plants; Hossain, M.A., Kamiya, T., Burritt, D.J., Phan Tran, L.-S., Fujiwara., T., Eds.; Elsevier Inc., 2018; pp. 109–121. [Google Scholar]
- Shireen, F.; Nawaz, M.A.; Chen, C.; Zhang, Q.; Zheng, Z.; Sohail, H.; Sun, J.; Cao, H.; Huang, Y.; Bie, Z. Boron: functions and approaches to enhance its availability in plants for sustainable agriculture. Int. J. Mol. Sci. 2018, 19, 1856. [Google Scholar] [CrossRef] [PubMed]
- Reid, R.J.; Hayes, J.E.; Post, A.; Stangoulis, J.C.R.; Graham, R.D. A critical analysis of the causes of boron toxicity in plants. Plant Cell Environ. 2004, 27, 1405–1414. [Google Scholar] [CrossRef]
- Camacho-Cristóbal, J.J.; Rexach, J.; González-Fontes, A. Boron in plants: deficiency and toxicity. J. Integr. Plant Biol. 2008, 50, 1247–1255. [Google Scholar] [CrossRef]
- Beato, V.M.; Rexach, J.; Navarro-Gochicoa, M.T.; Camacho-Cristóbal, J.J.; Herrera-Rodríguez, M.B.; Maldonado, J.M.; González-Fontes, A. A tobacco asparagine synthetase gene responds to carbon and nitrogen status and its root expression is affected under boron stress. Plant Sci. 2010, 178, 289–298. [Google Scholar] [CrossRef]
- Beato, V.M.; Navarro-Gochicoa, M.T.; Rexach, J.; Herrera-Rodríguez, M.B.; Camacho-Cristóbal, J.J.; Kempa, S.; Weckwerth, W.; González-Fontes, A. Expression of root glutamate dehydrogenase genes in tobacco plants subjected to boron deprivation. Plant Physiol. Biochem. 2011, 49, 1350–1354. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.-W.; Guo, L.-X.; Liu, Y.-Z.; Jin, L.-F.; Hussain, S.B.; Du, W.; Deng, Z.; Peng, S.-A. Transcriptome changes associated with boron deficiency in leaves of two citrus scion-rootstock combinations. Front. Plant Sci. 2017, 8, 317. [Google Scholar] [CrossRef]
- Landi, M.; Margaritopoulou, T.; Papadakis, I.E.; Araniti, F. Boron toxicity in higher plants: an update. Planta 2019, 250, 1011–1032. [Google Scholar] [CrossRef]
- Brdar-Jokanovi´c, M. Boron toxicity and deficiency in agricultural plants. Int. J. Mol. Sci. 2020, 21, 1424. [Google Scholar] [CrossRef]
- Kabay, N.; Güler, E.; Bryjak, M. Boron in seawater and methods for its separation—a review. Desalination 2010, 261, 212–217. [Google Scholar] [CrossRef]
- Chatzistathis, T.; Fanourakis, D.; Aliniaeifard, S.; Kotsiras, A.; Delis, C.; Tsaniklidis, G. Leaf age-dependent effects of boron toxicity in two Cucumis melo varieties. Agronomy. 2021, 11, 759. [Google Scholar] [CrossRef]
- Uluisik, I.; Karakaya, H.C.; Koc, A. The importance of boron in biological systems. J. Trace Elem. Med. Biol. 2018, 45, 156–162. [Google Scholar] [CrossRef]
- Mamani-Huarcaya, B.M.; González-Fontes, A.; Navarro-Gochicoa, M.T.; Camacho-Cristóbal, J.J.; Ceacero, C.J.; Herrera-Rodríguez, M.B.; Fernández Cutire, Ó.; Rexach, J. Characterization of two Peruvian maize landraces differing in boron toxicity tolerance. Plant Physiol. Biochem. 2022, 185, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.; Kancheti, M.; Raza, M.B.; Shiv, A.; Mangal, V.; Rathod, G.; Altaf, M.A.; Kumar, A.; Aftab, T.; Kumar, R.; et al. Mechanistic insight on boron-mediated toxicity in plant vis-a-vis its mitigation strategies: a review. Int. J. Phytoremediat. 2023, 25, 9–26. [Google Scholar] [CrossRef]
- Ralston, N.V.C.; Hunt, C.D. Diadenosine phosphates and S-adenosylmethionine: novel boron binding biomolecules detected by capillary electrophoresis. Biochim. Biophys. Acta 2001, 1527, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Pandey, P.; Stoerger, V.; Xu, Y.; Qiu, Y.; Ge, Y.; Schnable, J.C. Conventional and hyperspectral timeseries imaging of maize lines widely used in field trials. GigaScience 2018, 72, 1–11. [Google Scholar]
- Andorf, C.; Beavis, W.D.; Hufford, M.; Smith, S.; Suza, W.P.; Wang, K.; Woodhouse, M.; Yu, J.; Lübberstedt, T. Technological advances in maize breeding: past, present, and future. Theor. Appl. Genet. 2019, 132, 817–849. [Google Scholar] [CrossRef]
- Kausch, A.P.; Wang, K.; Kaeppler, H.F.; Gordon-Kamm, W. Maize transformation: history, progress, and perspectives. Mol. Breed. 2021, 41, 38. [Google Scholar] [CrossRef]
- Xi, Y.; Hu, W.; Zhou, Y.; Liu, X.; Qian, Y. Genome-wide identification and functional analysis of polyamine oxidase genes in maize reveal essential roles in abiotic stress tolerance. Front. Plant Sci. 2022, 13, 950064. [Google Scholar] [CrossRef]
- Ogunwole, A.A.; Otusanya, O.O.; Oloyede, F.A.; Olabamiji, T.M. Comparative effects of boron toxicity and deficiency on the growth, chlorophyll, protein and some cations accumulation in Zea mays seedlings. Int. J. Sci. Res. Innov. 2015, 17, 316–335. [Google Scholar]
- Gotz, L.F.; Silvestrin, F.; Motta, A.C.V.; Pauletti, V. Response to application and tissue diagnosis of boron deficiency and toxicity in maize. Commun. Soil Sci. Plant Anal. 2021, 52, 2898–2911. [Google Scholar] [CrossRef]
- Sang, W.; Huang, Z.-R.; Yang, L.-T.; Guo, P.; Ye, X.; Chen, L.-S. Effects of high toxic boron concentration on protein profiles in roots of two citrus species differing in boron-tolerance revealed by a 2-DE based MS approach. Front. Plant Sci. 2017, 8, 180. [Google Scholar] [CrossRef]
- Fan, T.; Bykova, N.V.; Rampitsch, C.; Xing, T. Identification and characterization of a serine protease from wheat leaves. Eur. J. Plant Pathol. 2016, 146, 293–304. [Google Scholar] [CrossRef]
- Malefo, M.B.; Mathibela, E.O.; Crampton, B.G.; Makgopa, M.E. Investigating the role of Bowman-Birk serine protease inhibitor in Arabidopsis plants under drought stress. Plant Physiol. Biochem. 2020, 149, 286–293. [Google Scholar] [CrossRef]
- D’Ippolito, S.; Rey-Burusco, M.F.; Feingold, S.E.; Guevara, M.G. Role of proteases in the response of plants to drought. Plant Physiol. Biochem. 2021, 168, 1–9. [Google Scholar] [CrossRef]
- Sharma, P.; Gayen, D. Plant protease as regulator and signaling molecule for enhancing environmental stress-tolerance. Plant Cell Reports. 2021, 40, 2081–2095. [Google Scholar] [CrossRef]
- Moin, M.; Bakshi, A.; Saha, A.; Dutta, M.; Madhav, S.M.; Kirti, P.B. Rice ribosomal protein large subunit genes and their spatio-temporal and stress regulation. Front. Plant Sci. 2016, 7, 1284. [Google Scholar] [CrossRef]
- Nozawa, A.; Miwa, K.; Kobayashi, M.; Fujiwara, T. Isolation of Arabidopsis thaliana cDNAs that confer yeast boric acid tolerance. Biosci. Biotechnol. Biochem. 2006, 70, 1724–1730. [Google Scholar] [CrossRef]
- Tanaka, M.; Sotta, N.; Yamazumi, Y.; Yamashita, Y.; Miwa, K.; Murota, K.; Chiba, Y.; Hirai, M.Y.; Akiyama, T.; Onouchi, H.; et al. The minimum open reading frame, AUG-stop, induces boron-dependent ribosome stalling and mRNA degradation. Plant Cell 2016, 28, 2830–2849. [Google Scholar] [CrossRef] [PubMed]
- Sotta, N.; Chiba, Y.; Miwa, K.; Takamatsu, S.; Tanaka, M.; Yamashita, Y.; Naito, S.; Fujiwara, T. Global analysis of boron-induced ribosome stalling reveals its effects on translation termination and unique regulation by AUG-stops in Arabidopsis shoots. Plant J. 2021, 106, 1455–1467. [Google Scholar] [CrossRef]
- Saidi, A.; Hajibarat, Z. In-silico analysis of eukaryotic translation initiation factors (eIFs) in response to environmental stresses in rice (Oryza sativa). Biologia 2020, 75, 1731–1738. [Google Scholar] [CrossRef]
- Pakdel, H.; Hassani, S.B.; Ghotbi-Ravandi, A.A.; Bernard, F. Contrasting the expression pattern change of polyamine oxidase genes and photosynthetic efficiency of maize (Zea mays L.) genotypes under drought stress. J. Biosci. 2020, 45, 73. [Google Scholar] [CrossRef]
- Shu, S.; Guo, S.-R.; Sun, J.; Yuan, L.-Y. Effects of salt stress on the structure and function of the photosynthetic apparatus in Cucumis sativus and its protection by exogenous putrescine. Physiol. Plant. 2012, 146, 285–296. [Google Scholar] [CrossRef]
- Hamdani, S.; Yaakoubi, H.; Carpentier, R. Polyamines interaction with thylakoid proteins during stress. J. Photochem. Photobiol. B-Biol. 2011, 104, 314–331. [Google Scholar] [CrossRef]
- Chen, M.; Blankenship, R.E. Expanding the solar spectrum used by photosynthesis. Trends Plant Sci. 2011, 16, 427–431. [Google Scholar] [CrossRef]
- Zhang, D.; Chang, E.; Yu, X.; Chen, Y.; Yang, Q.; Cao, Y.; Li, X.; Wang, Y.; Fu, A.; Xu, M. Molecular characterization of magnesium chelatase in soybean [Glycine max (L.) Merr.]. Front. Plant Sci. 2018, 9, 720. [Google Scholar] [CrossRef]
- Sasi, S.; Venkatesh, J.; Daneshi, R.F.; Gururani, M.A. Photosystem II extrinsic proteins and their putative role in abiotic stress tolerance in higher plants. Plants, 2018, 7, 100. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Portis, A.R. A novel nucleus-encoded chloroplast protein, PIFI, is involved in NAD(P)H dehydrogenase complex-mediated chlororespiratory electron transport in Arabidopsis. Plant Physiol. 2007, 144, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, N.; Takabayashi, A.; Noguchi, K.; Tazoe, Y.; Yamamoto, H.; von Caemmerer, S.; Sato, F.; Endo, T. NDH-mediated cyclic electron flow around photosystem I is crucial for C4 photosynthesis. Plant Cell Physiol. 2016, 57, 2020–2028. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Liu, Y.; Bai, C.; Yong, J.W.H. The significance of chloroplast NAD(P)H dehydrogenase complex and its dependent cyclic electron transport in photosynthesis. Front. Plant Sci. 2021, 12, 661863. [Google Scholar] [CrossRef]
- Yamori, W.; Shikanai, T. Physiological functions of cyclic electron transport around photosystem I in sustaining photosynthesis and plant growth. Annu. Rev. Plant Biol. 2016, 67, 81–106. [Google Scholar] [CrossRef]
- Zhu, D.; Luo, F.; Zou, R.; Liu, J.; Yan, Y. Integrated physiological and chloroplast proteome analysis of wheat seedling leaves under salt and osmotic stresses. J. Proteomics 2021, 234, 104097. [Google Scholar] [CrossRef] [PubMed]
- Murata, N.; Nishiyama, Y. ATP is a driving force in the repair of photosystem II during photoinhibition. Plant Cell Environ. 2018, 41, 285–299. [Google Scholar] [CrossRef]
- Araniti, F.; Miras-Moreno, B.; Lucini, L.; Landi, M.; Abenavoli, M.R. Metabolomic, proteomic and physiological insights into the potential mode of action of thymol, a phytotoxic natural monoterpenoid phenol. Plant Physiol. Biochem. 2020, 153, 141–153. [Google Scholar] [CrossRef]
- Wang, W.; Vignani, R.; Scali, M.; Cresti, M. A universal and rapid protocol for protein extraction from recalcitrant plant tissues for proteomic analysis. Electrophoresis 2006, 27, 2782–2786. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of proteins utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteomics 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Number of significantly (P ≤ 0.05) accumulated proteins up or down, represented as positive and negative, respectively, comparing maize landraces and B treatments. Seedlings were subjected to 0.05 and 10 mM B for 10 days. Results were obtained from 3-4 separate plants of each landrace and B treatments. For more details, see Materials and Methods. S: Sama landrace; P: Pachía landrace; 0.05: 0.05 mM B (B control treatment); 10: 10 mM B (B toxicity treatment). The numbers above the columns represent the numbers of proteins accumulated up (green) or down (red).
Figure 1.
Number of significantly (P ≤ 0.05) accumulated proteins up or down, represented as positive and negative, respectively, comparing maize landraces and B treatments. Seedlings were subjected to 0.05 and 10 mM B for 10 days. Results were obtained from 3-4 separate plants of each landrace and B treatments. For more details, see Materials and Methods. S: Sama landrace; P: Pachía landrace; 0.05: 0.05 mM B (B control treatment); 10: 10 mM B (B toxicity treatment). The numbers above the columns represent the numbers of proteins accumulated up (green) or down (red).
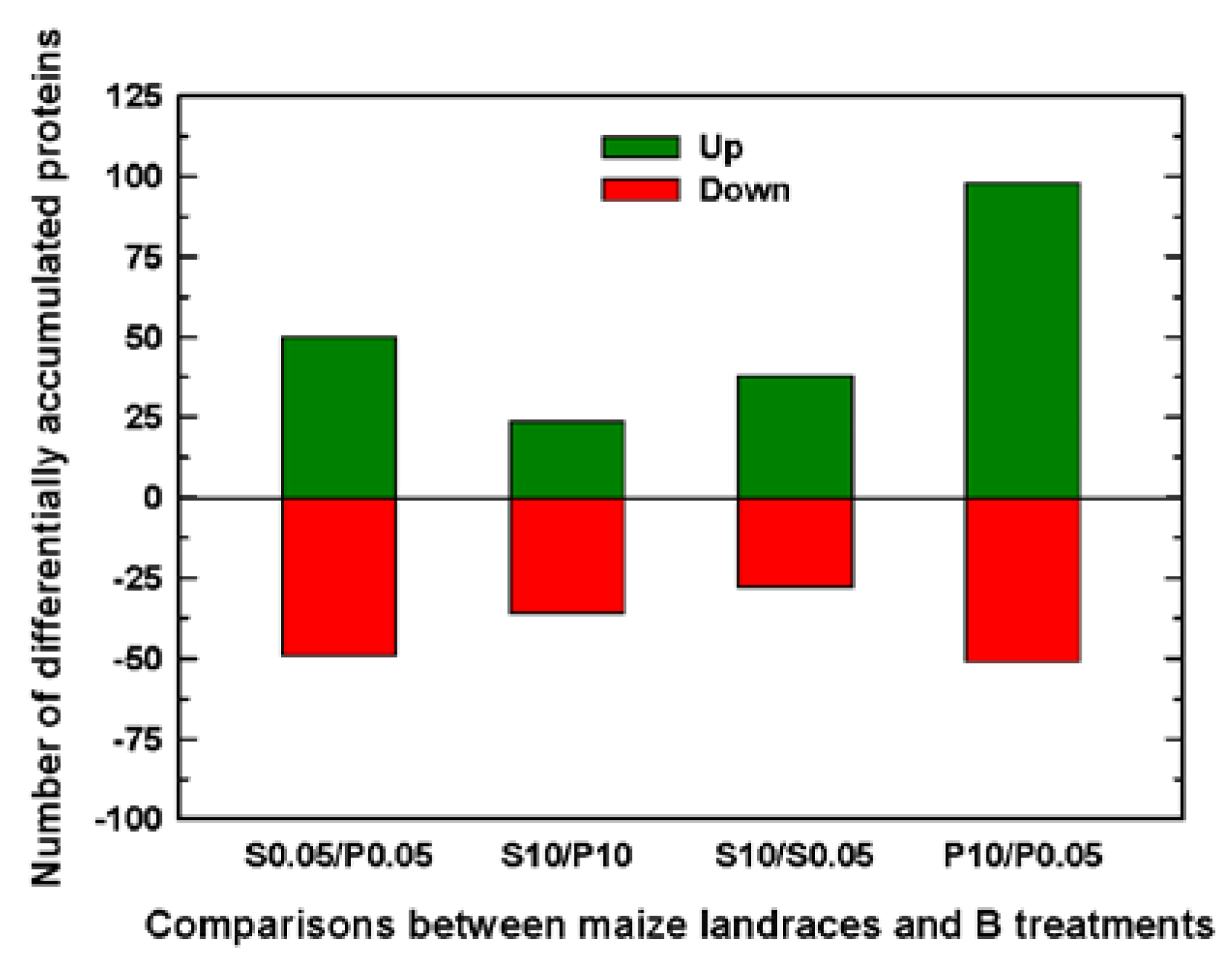
Figure 2.
Number of differentially accumulated proteins (DAPs) in the different functional categories obtained from the four comparisons shown in Figure 1 and Table S2. Seedlings of Sama and Pachía landraces were subjected to 0.05 and 10 mM B for 10 days. Results were obtained by addition of the DAPs in the four comparisons. For more details, see Materials and Methods. Aa: amino acid metabolism; CA: carbon assimilation and Calvin cycle; CM: carbohydrate metabolism; CDe: cell death; CDi: cell division; CW: cell wall; DNA: DNA and chromatin organization and DNA repair; LM: lipid metabolism; NM: nitrogen metabolism; NPM: nucleotide, purine, and pyrimidine metabolism; OT: others; ORP: oxidation and reduction processes; PLR: photosynthetic light reactions; PB: pigment biosynthesis; PD: protein degradation; PSF: protein stabilization and folding; ROS: reactive oxygen species scavenging pathways/response to oxidative stress; R: respiration metabolism (glycolysis, TCA cycle, and mitochondrial electron transfer); RB: ribosome biogenesis; RBP: RNA binding and processing; SM: secondary metabolism; SG: signaling; ST: stress; TT: transcription and translation processes; TP: transporters and transport processes; NWK: not well-known proteins.
Figure 2.
Number of differentially accumulated proteins (DAPs) in the different functional categories obtained from the four comparisons shown in Figure 1 and Table S2. Seedlings of Sama and Pachía landraces were subjected to 0.05 and 10 mM B for 10 days. Results were obtained by addition of the DAPs in the four comparisons. For more details, see Materials and Methods. Aa: amino acid metabolism; CA: carbon assimilation and Calvin cycle; CM: carbohydrate metabolism; CDe: cell death; CDi: cell division; CW: cell wall; DNA: DNA and chromatin organization and DNA repair; LM: lipid metabolism; NM: nitrogen metabolism; NPM: nucleotide, purine, and pyrimidine metabolism; OT: others; ORP: oxidation and reduction processes; PLR: photosynthetic light reactions; PB: pigment biosynthesis; PD: protein degradation; PSF: protein stabilization and folding; ROS: reactive oxygen species scavenging pathways/response to oxidative stress; R: respiration metabolism (glycolysis, TCA cycle, and mitochondrial electron transfer); RB: ribosome biogenesis; RBP: RNA binding and processing; SM: secondary metabolism; SG: signaling; ST: stress; TT: transcription and translation processes; TP: transporters and transport processes; NWK: not well-known proteins.
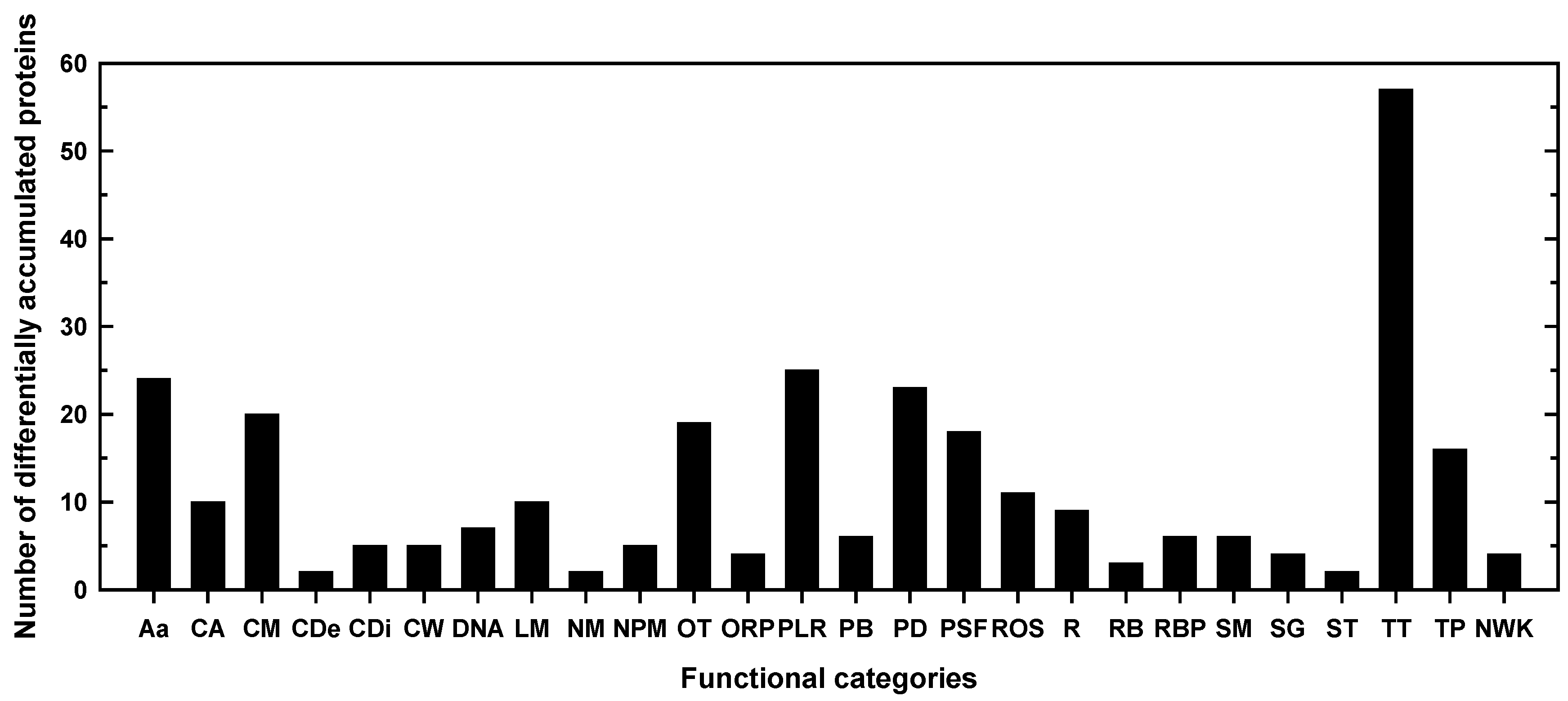
Figure 3.
Functional categories of 303 maize proteins given as the number of those significantly expressed, represented as positive (up-accumulated) and negative (down-accumulated). Seedlings of Sama (S) and Pachía (P) landraces were subjected to 0.05 and 10 mM B for 10 days. Results were obtained from 3-4 separate plants of each landrace and B treatments. For more details, see Materials and Methods. 0.05: 0.05 mM B (B control treatment); 10: 10 mM B (B toxicity treatment).
Figure 3.
Functional categories of 303 maize proteins given as the number of those significantly expressed, represented as positive (up-accumulated) and negative (down-accumulated). Seedlings of Sama (S) and Pachía (P) landraces were subjected to 0.05 and 10 mM B for 10 days. Results were obtained from 3-4 separate plants of each landrace and B treatments. For more details, see Materials and Methods. 0.05: 0.05 mM B (B control treatment); 10: 10 mM B (B toxicity treatment).
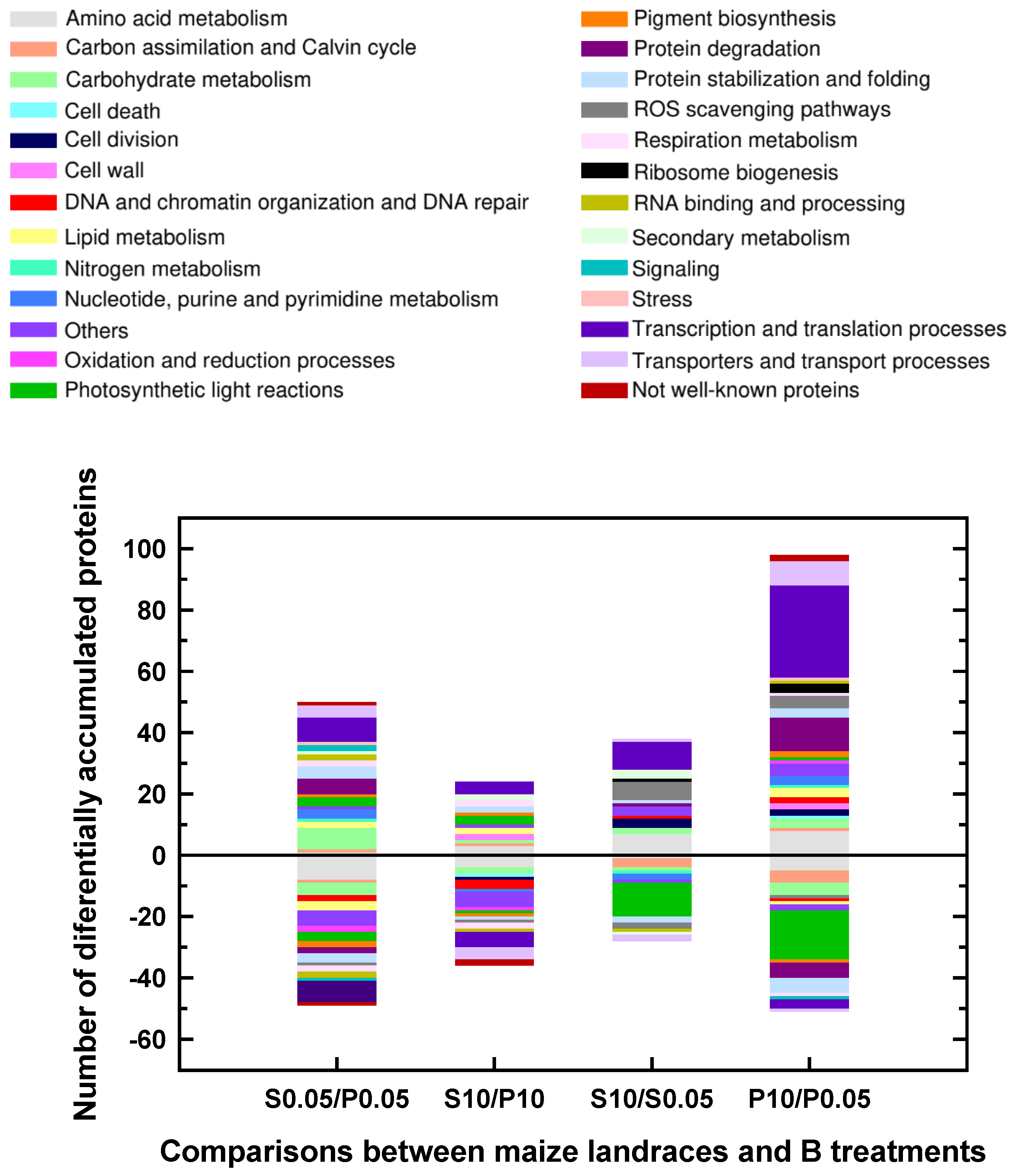
Figure 4.
Number of induced or repressed proteins in Sama and Pachía landraces in the different functional categories obtained from the comparisons between B toxicity and B control conditions shown in Table S2. Seedlings of Sama and Pachía landraces were subjected to 0.05 (control) and 10 mM (toxicity) B for 10 days. Results were obtained by addition of the induced or repressed proteins in Sama and Pachía. For more details, see Materials and Methods. Aa: amino acid metabolism; CA: carbon assimilation and Calvin cycle; CM: carbohydrate metabolism; CDe: cell death; CDi: cell division; CW: cell wall; DNA: DNA and chromatin organization and DNA repair; LM: lipid metabolism; NM: nitrogen metabolism; NPM: nucleotide, purine, and pyrimidine metabolism; OT: others; ORP: oxidation and reduction processes; PLR: photosynthetic light reactions; PB: pigment biosynthesis; PD: protein degradation; PSF: protein stabilization and folding; ROS: reactive oxygen species scavenging pathways/response to oxidative stress; R: respiration metabolism (glycolysis, TCA cycle, and mitochondrial electron transfer); RB: ribosome biogenesis; RBP: RNA binding and processing; SM: secondary metabolism; SG: signaling; ST: stress; TT: transcription and translation processes; TP: transporters and transport processes; NWK: not well-known proteins.
Figure 4.
Number of induced or repressed proteins in Sama and Pachía landraces in the different functional categories obtained from the comparisons between B toxicity and B control conditions shown in Table S2. Seedlings of Sama and Pachía landraces were subjected to 0.05 (control) and 10 mM (toxicity) B for 10 days. Results were obtained by addition of the induced or repressed proteins in Sama and Pachía. For more details, see Materials and Methods. Aa: amino acid metabolism; CA: carbon assimilation and Calvin cycle; CM: carbohydrate metabolism; CDe: cell death; CDi: cell division; CW: cell wall; DNA: DNA and chromatin organization and DNA repair; LM: lipid metabolism; NM: nitrogen metabolism; NPM: nucleotide, purine, and pyrimidine metabolism; OT: others; ORP: oxidation and reduction processes; PLR: photosynthetic light reactions; PB: pigment biosynthesis; PD: protein degradation; PSF: protein stabilization and folding; ROS: reactive oxygen species scavenging pathways/response to oxidative stress; R: respiration metabolism (glycolysis, TCA cycle, and mitochondrial electron transfer); RB: ribosome biogenesis; RBP: RNA binding and processing; SM: secondary metabolism; SG: signaling; ST: stress; TT: transcription and translation processes; TP: transporters and transport processes; NWK: not well-known proteins.
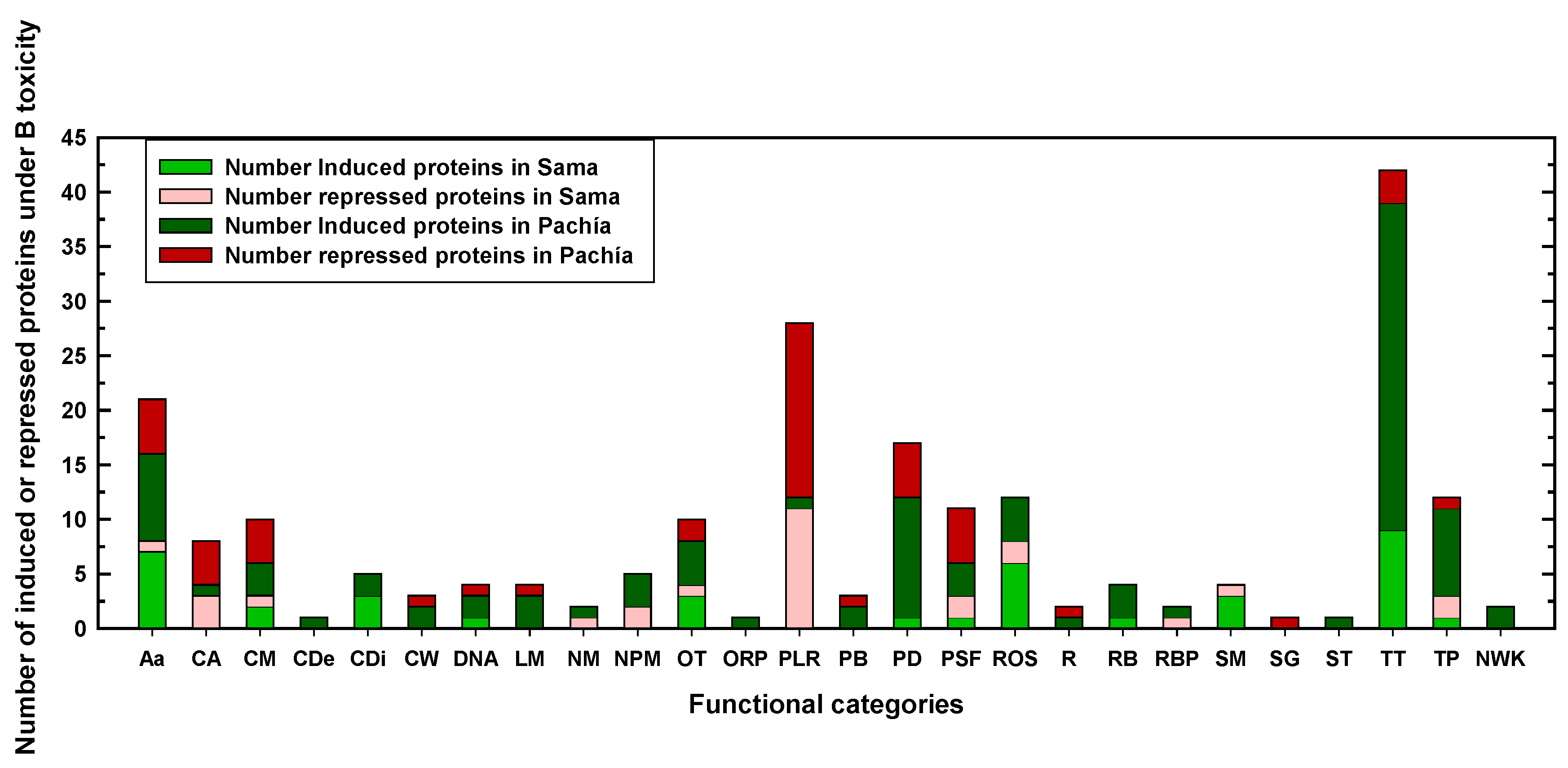

Table 1.
Number of proteins detected in leaves of Pachía (P) and Sama (S) landraces under different boron (B) treatments and number of significant differentially accumulated proteins (DAPs) in Pachía and Sama landraces under different B treatments.
Table 1.
Number of proteins detected in leaves of Pachía (P) and Sama (S) landraces under different boron (B) treatments and number of significant differentially accumulated proteins (DAPs) in Pachía and Sama landraces under different B treatments.
| P0.05 mM (Control) | P10 mM B (B toxicity) | S0.05 mM (control) | S10 mM (B toxicity) | |||
|---|---|---|---|---|---|---|
| Number of detected proteins1 | 1100 | 1040 | 1111 | 1145 | ||
|
S0.05 versus P0.05 (Control conditions) |
S10 versus P10 (B toxicity conditions) |
|||||
| Number of significant DAPs between Sama and Pachía | 99 | 60 | ||||
|
Sama S10 versus S0.05 |
Pachía P10 versus P0.05 |
|||||
| Number of significant DAPs by B toxicity | 66 | 149 | ||||
1 Numbers of proteins that were detected in at least one landrace (Sama or Pachía) and one B treatment analyzed.
Table 2.
Commonly expressed proteins in both Pachía and Sama landraces in response to boron (B) toxicity.
Table 2.
Commonly expressed proteins in both Pachía and Sama landraces in response to boron (B) toxicity.
| Pachía | Sama | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein ID1 | Gene Name/ID2 | Protein name / Annotation |
FC3 | P-value4 | FC3 |
P- value4 |
FCSA/ FCPA5 |
Function/Biological process6 | |||||||
| Amino acid metabolism | |||||||||||||||
| B6SKB7 | Zm00001d031013 | Methylcrotonoyl-CoA carboxylase subunit α | 4.44 | 0.0022 | 3.56 | 0.0049 | 0.80 | Leucine degradation | |||||||
| A0A1D6K836 | Zm00001d029848 | Branched-chain-amino-acid aminotransferase | 2.35 | 0.0272 | 1.65 | 0.0241 | 0.70 | Branched-chain amino acid biosynthesis | |||||||
| B4G011 | Zm00001d046923 | D-3-phosphoglycerate dehydrogenase chloroplastic | 2.31 | 0.0154 | 1.52 | 0.0202 | 0.66 | Serine biosynthesis | |||||||
| A0A1D6DW07 | Zm00001d002051 | D-3-phosphoglycerate dehydrogenase | 1.78 | 0.0494 | 1.69 | 0.0175 | 0.95 | Serine biosynthesis | |||||||
| Carbon assimilation / Calvin cycle | |||||||||||||||
| O24574 | Zm00001d004894 | Ribulose bisphosphate carboxylase small chain | 0.38 | 0.0113 | 0.33 | 0.0466 | 0.87 | Carbon dioxide fixation | |||||||
| Carbohydrate metabolism | |||||||||||||||
| Q9FQ11 | Zm00001d010523 | Sucrose-phosphatase 1 | 1.50 | 0.0154 | 1.58 | 0.0420 | 1.05 | Sucrose biosynthesis | |||||||
| A0A1D6IJ76 | Zm00001d022107 | Glyceraldehyde-3-phosphate dehydrogenase A | 0.34 | 0.0319 | 0.51 | 0.0019 | 1.52 | Carbon metabolism | |||||||
| Cell division | |||||||||||||||
| A0A1D6FRI4 | Zm00001d010500 | ERBB-3 binding protein 1 | 1.89 | 0.0387 | 1.58 | 0.0266 | 0.84 | Cell division and cell growth regulation | |||||||
| Photosynthetic light reactions | |||||||||||||||
| A0A1D6HS38 | Zm00001d018779 | Oxygen-evolving enhancer protein 2-1 chloroplastic (OEE2-1) | 0.27 | 0.0110 | 0.48 | 0.0354 | 1.78 | Photosynthesis. Photosystem II oxygen evolving complex | |||||||
| B4FWG2 | Zm00001d048422 | Photosynthetic NDH subunit of subcomplex B 2 chloroplastic | 0.25 | 0.0047 | 0.41 | 0.0200 | 1.62 | Photosynthetic electron transport flow around photosystem I to produce ATP | |||||||
| A0A1X7YHG9 | AtpA | ATP synthase subunit α chloroplastic (ATPα) | 0.20 | 0.0166 | 0.61 | 0.0163 | 2.99 | Chloroplast ATP synthesis coupled proton transport | |||||||
| P46617 | PetA | Cytochrome f | 0.18 | 0.0193 | 0.29 | 0.0161 | 1.59 | Photosynthetic electron transport activity | |||||||
| P00827 | Zm00001d006403 | ATP synthase subunit β chloroplastic (ATPβ) | 0.15 | 0.0076 | 0.52 | 0.0274 | 3.45 | Chloroplast ATP synthesis coupled proton transport | |||||||
| Reactive Oxygen Species (ROS) Scavenging Pathways / Response to oxidative stress | |||||||||||||||
| A0A1D6MSE3 | Zm00001d040721 | Dihydrolipoyl dehydrogenase | 2.30 | 0.0273 | 1.80 | 0.0205 | 0.78 | Cell redox homeostasis | |||||||
| A0A1D6JPH3 | Zm00001d027769 | Glutathione reductase | 2.21 | 0.0053 | 1.71 | 0.0436 | 0.77 | Cell redox homeostasis. Glutathione metabolic process. Cellular oxidant detoxification | |||||||
| Ribosome biogenesis | |||||||||||||||
| K7UTH7 | Zm00001d009596 | GTPase ERA1 chloroplastic | 2.61 | 0.0108 | 1.81 | 0.0126 | 0.69 | Ribosome biogenesis. Ribosomal small subunit assembly. rRNA processing | |||||||
| Transcription and translation processes | |||||||||||||||
| A0A1D6LIV5 | Zm00001d035802 | Phenylalanine--tRNA ligase beta subunit cytoplasmic | 2.56 | 0.0314 | 2.23 | 0.0093 | 0.87 | Translation. Phenylalanyl-tRNA aminoacylation | |||||||
| Transporters and transport processes | |||||||||||||||
| B6SP43 | Zm00001d007597 | ABC family1 | 4.54 | 0.0103 | 2.69 | 0.0125 | 0.59 | ATPase-coupled transmembrane transporter activity | |||||||
1Proteins ID, Protein identification (ID) number in the UniProt database; 2Gene Name, name or ID number of the corresponding gene of the differentially expressed protein as searched in the Maize Genetics and Genomics Database (MaizeGDB; https://www.maizegdb.org/; 3Fold Change, is expressed as the ratio of LFQ intensities (on a logarithmic scale) of proteins between 10 and 0.05 mM B treatments; 4P-value, statistical level (using Student’s t-test) ≤ 0.05, at which differential protein expression was accepted as significant; 5FCSA/FCPA, is the ratio between fold change of Sama and Pachía. 6Function/Biological process, annotated biological functions or biological process based on different databases. Induced proteins are highlighted with light green rows and repressed proteins with light red rows. For more details, see Materials and Methods. Results were obtained from 3-4 separate plants of each landrace.
Table 3.
Proteins with higher differential expression in Pachía leaves in response to boron (B) toxicity. This table shows the proteins strongly induced or repressed by B toxicity in Pachía by comparing their expressions with those of Pachía in medium with 0.05 mM B.
Table 3.
Proteins with higher differential expression in Pachía leaves in response to boron (B) toxicity. This table shows the proteins strongly induced or repressed by B toxicity in Pachía by comparing their expressions with those of Pachía in medium with 0.05 mM B.
| Protein ID1 | Gene Name/ID2 | Protein name/Annotation | FC3 | P-Value4 | Function/Biological process5 |
|---|---|---|---|---|---|
| AMINO ACID AND PEPTIDE METABOLISMS | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| B6SKB7 | Zm00001d031013 | Methylcrotonoyl-CoA carboxylase subunit α | 4.44 | 0.0022 | Leucine degradation |
| B6SWZ4 | Zm00001d050336 | Methylcrotonoyl-CoA carboxylase β chain mitochondrial | 2.85 | 0.0154 | Leucine degradation |
| A0A1D6K836 | Zm00001d029848 | Branched-chain-amino-acid amino-transferase | 2.35 | 0.0272 | Branched-chain amino acid biosynthesis |
| B4G011 | Zm00001d046923 | D-3-phosphoglycerate dehydroge-nase chloroplastic | 2.31 | 0.0154 | Serine biosynthesis |
| C4J411 | Zm00001d028464 | Imidazole glycerol phosphate synthase hisHF | 2.17 | 0.0017 | Histidine biosynthesis |
| C4JBG7 | Zm00001d015088 | 3-isopropylmalate dehydratase large subunit | 2.14 | 0.0320 | Leucine biosynthesis |
| Strongly repressed proteins by B toxicity in Pachía | |||||
| B4FUH2 | Zm00001d043382 | Aspartate aminotransferase | 0.48 | 0.0195 | Amino acid metabolic process |
| B4FU01 | Zm00001d045153 | Cystathionine β-lyase chloroplas-tic | 0.44 | 0.0235 | Methionine biosynthetic. Cysteine biosynthetic process via cystathionine |
| A0A1D6ICL3 | Zm00001d021596 | Adenosine 5-phosphosulfate reductase-like1 | 0.29 | 0.0140 | Cysteine biosynthetic process. Sulfate reduction |
| B6TZD1 | Zm00001eb168430 | Methylthioribose-1-phosphate isomerase | 0.24 | 0.0461 | Methionine biosynthesis |
| CARBON ASSIMILATION AND CALVIN CYCLE | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6FQE4 | Zm00001d010321 | Pyruvate phosphate dikinase | 2.31 | 0.0449 | C4 photosynthetic carbon assimilation cycle |
| Strongly repressed proteins by B toxicity in Pachía | |||||
| O24574 | Zm00001d004894 | Ribulose bisphosphate carboxylase small chain | 0.38 | 0.0113 | Carbon dioxide fixation |
| B4FQ59 | Zm00001d017711 | Phosphoribulokinase | 0.33 | 0.0004 | Calvin- Benson cycle |
| Q9ZT00 | Zm00001eb164390 | Ribulose bisphosphate carboxylase/oxygenase activase chloroplastic | 0.26 | 0.0090 | Carbon dioxide fixation. Rubisco activator activity |
| CARBOHYDRATE METABOLISM | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6NE29 | Zm00001d043662 | α-amylase 3 chloroplastic | 2.05 | 0.0460 | Starch degradation |
| Strongly repressed proteins by B toxicity in Pachía | |||||
| A0A1D6M7C2 | Zm00001d038579 | Phosphoglycerate kinase cytosolic | 0.49 | 0.0136 | Glycolysis and gluconeogenesis |
| B4FRC9 | Zm00001d011965 | Transaldolase | 0.41 | 0.0407 | Pentose-phosphate shunt |
| A0A1D6IJ76 | Zm00001d022107 | Glyceraldehyde-3-phosphate dehydrogenase A | 0.34 | 0.0319 | Carbon metabolism |
| CELL DEATH | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| B4F8B9 | Zm00001d018468 | S-(hydroxymethyl)glutathione dehydrogenase | 2.81 | 0.0027 | Cell death. Formaldehyde oxidation (glutathione-dependent) |
| CELL WALL | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| B4F9J1 | Zm00001d046357 | β-galactosidase | 3.17 | 0.0092 | Xyloglucan degradation |
| DNA AND CHROMATIN ORGANIZATION AND DNA REPAIR | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| B6TGH8 | Zm00001d034479 | Histone H1 | 3.60 | 0.0349 | Chromosome condensation. Nucleosome assembly. Nucleosome positioning |
| C0P6Q6 | Zm00001d040416 | DNA gyrase subunit B | 3.48 | 0.0007 | DNA topological change |
| Strongly repressed proteins by B toxicity in Pachía | |||||
| B6SK03 | Zm00001d053295 | Ubiquitin-conjugating enzyme E2 variant 1C | 0.39 | 0.0409 | DNA postreplication repair. Protein polyubiquitination |
| LIPID METABOLISM | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| K7VQG5 | Zm00001d008727 | Phospholipase D | 2.30 | 0.0244 | Phospholipid degradation |
| A0A1D6NE81 | Zm00001d043680 | Phospholipase A1-IIδ | 2.02 | 0.0390 | Lipid degradation |
| Strongly repressed proteins by B toxicity | |||||
| B4FLS8 | Zm00001d003584 | 12-oxo-phytodienoic acid reductase 5 | 0.33 | 0.0436 | Fatty acid and oxylipin biosynthesis |
| NITROGEN METABOLISM | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6PZA5 | Zm00001d049995 | Nitrate reductase | 2.19 | 0.0077 | Nitrate reductase (NADH) activity. Nitrate assimilation |
| OTHERS | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6JGY3 | Zm00001d026515 | Molybdopterin molybdenum-transferase | 2.92 | 0.0023 | Molybdenum cofactor biosynthesis |
| A0A1D6HUN3 | Zm00001d019040 | D-2-hydroxyglutarate dehydrogenase mitochondrial | 2.09 | 0.0380 | Lysine degradation |
| Strongly repressed proteins by B toxicity in Pachía | |||||
| C0PDB6 | Zm00001d039535 | HXXXD-type acyl-transferase family protein | 0.40 | 0.0112 | N-acyltransferase activity |
| C0PE12 | Zm00001d009877 | Protein plastid transcriptionally active 16 chloroplastic | 0.24 | 0.0121 | Circadian rhythm |
| OXIDATION AND REDUCTION PROCESSES | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6M498 | Zm00001d038189 | FAD/NAD(P)-binding oxidoreductase family protein | 2.04 | 0.0101 | Oxidoreductase activity |
| PHOTOSYNTHETIC LIGHT REACTIONS | |||||
| Strongly repressed proteins by B toxicity in Pachía | |||||
| B6SSB9 | Zm00001d035859 | Plastocyanin | 0.50 | 0.0300 | Photosynthetic electron transport |
| A0A1D6GU53 | Zm00001d014564 | Oxygen-evolving enhancer protein 1-1 chloroplastic | 0.47 | 0.0268 | Photosynthesis. Oxygen evolving activity. Photosystem II assembly and stabilization |
| B6SUC4 | Zm00001d046786 | Chlorophyll a-b binding protein, chloroplastic | 0.41 | 0.0086 | Photosynthesis. Light harvesting in photosystem I |
| B6T927 | Zm00001d014349 | NAD(P)H-quinone oxidoreductase subunit S chloroplastic (NDHS) | 0.39 | 0.0095 | Photosynthetic electron transport chain |
| P25709 | NdhH | NAD(P)H-quinone oxidoreductase subunit H, chloroplastic | 0.37 | 0.0022 | Photosynthesis, light reaction. Photosynthetic electron transport chain. Couples the photosynthetic redox reaction to proton translocation |
| B6SP99 | Zm00001d024148 | Photosynthetic NDH subunit of subcomplex B 1 chloroplastic | 0.33 | 0.0137 | Photosynthetic electron transport in photosystem I |
| B4FJP7 | Zm00001d027729 | Photosynthetic NDH subunit of subcomplex B 2 chloroplastic | 0.32 | 0.0169 | Photosynthetic electron transport in photosystem I |
| B4FR80 | Zm00001d033098 | Post-illumination chlorophyll fluorescence increase (ZmPIFI) | 0.28 | 0.0270 | Chlororespiration |
| A0A1D6HS38 | Zm00001d018779 | Oxygen-evolving enhancer protein 2-1 chloroplastic (OEE2-1) | 0.27 | 0.0110 | Photosynthesis. Photosystem II oxygen evolving complex |
| B4FWG2 | Zm00001d048422 | Photosynthetic NDH subunit of subcomplex B 2 chloroplastic | 0.25 | 0.0047 | Photosynthetic electron transport flow around photosystem I to produce ATP |
| P19124 | NdhJ | NAD(P)H-quinone oxidoreductase subunit J, chloroplastic | 0.22 | 0.0147 | Photosynthesis, light reaction, photosynthetic electron transport chain. Couples the photosynthetic redox reaction to proton translocation |
| A0A1X7YHG9 | AtpA | ATP synthase subunit α (ATPα) | 0.20 | 0.0166 | Chloroplast ATP synthesis coupled proton transport |
| P46617 | PetA | Cytochrome f | 0.18 | 0.0193 | Photosynthetic electron transport chain |
| P00827 | Zm00001d009488 | ATP synthase subunit β, chloroplastic (ATPβ) | 0.15 | 0.0076 | Chloroplast ATP synthesis coupled proton transport |
| A0A1D6JYG6 | Zm00001d028670 | Photosynthetic NDH subunit of lumenal location 1 chloroplastic | 0.13 | 0.0134 | Part of photosystem II oxygen evolving complex |
| PIGMENT BIOSYNTHESIS | |||||
| Strongly repressed proteins by B toxicity in Pachía | |||||
| A0A1D6FAV8 | Zm00001d008203 | Protoporphyrinogen oxidase | 0.38 | 0.0173 | 3,8-divinyl-chlorophyllide a and protoporphyrinogen IX biosynthesis |
| PROTEIN DEGRADATION | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| B4FS65 | Zm00001d005391 | Cysteine protease 14 | 4.38 | 0.0146 | Proteolysis. Proteolysis involved in protein catabolic process |
| A0A1D6HM49 | Zm00001d018282 | Subtilisin-like protease SBT1.4 | 3.70 | 0.0399 | Serine protease. Serine-type endopeptidase activity. Proteolysis |
| A0A1D6H4R4 | Zm00001d015962 | Prolyl oligopeptidase family protein | 3.58 | 0.0080 | Proteolysis. Serine protease. Serine-type peptidase activity |
| Q84TL7 | Zm00001d011036 | Legumin-like protein | 2.86 | 0.0453 | Protein ubiquitination. Nutrient reservoir activity. Storage protein |
| A0A1D6KWW2 | Zm00001d033194 | Subtilisin-like protease | 2.85 | 0.0403 | Proteolysis. Serine protease. Serine-type endopeptidase activity |
| A0A1D6KV27 | Zm00001d032956 | Acylamino-acid-releasing enzyme | 2.54 | 0.0086 | Proteolysis. Serine protease. Serine-type endopeptidase activity |
| C0HI51 | Zm00001d044102 | Zn-dependent exopeptidase superfamily protein | 2.53 | 0.0131 | Proteolysis. Aminopeptidase. Metalloaminopeptidase activity |
| Q84TL6 | Zm00001d035597 | Legumin-like protein | 2.36 | 0.0386 | Protein ubiquitination. Storage protein. Nutrient reservoir activity |
| A0A1D6HL34 | Zm00001d018145 | Presequence protease 2 chloroplastic/mitochondrial | 2.22 | 0.0180 | Proteolysis. Metalloendopeptidase activity. Protein processing |
| K7VGG8 | Zm00001d010522 | ATP-dependent zinc metalloprotease FTSH 10 mitochondrial | 2.07 | 0.0359 | Proteolysis. Metalloprotease mitochondrial |
| C4JC43 | Zm00001d049100 | Target of Myb protein 1 | 2.04 | 0.0450 | Proteolysis. Protein transport to vacuole involved in ubiquitin-dependent protein catabolic process via the multivesicular body sorting pathway |
| Strongly repressed proteins by B toxicity in Pachía | |||||
| A0A1D6H558 | Zm00001d016036 | Chloroplast processing peptidase | 0.47 | 0.0438 | Protease. Serine-type endopeptidase activity |
| B4FQJ6 | Zm00001d018309 | 26S protease regulatory subunit 7 homolog A | 0.46 | 0.0249 | Proteolysis. Protein catabolic process. Peptidase activity |
| A0A1D6FKP2 | Zm00001d009613 | Protease Do-like 1 chloroplastic | 0.45 | 0.0496 | Proteolysis. Serine-type endopeptidase activity |
| K7TTX0 | Zm00001d025628 | Plant UBX domain-containing protein 4 | 0.44 | 0.0107 | Proteasome-mediated ubiquitin-dependent protein catabolic process |
| PROTEIN STABILIZATION AND FOLDING | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6FN98 | Zm00001d009948 | Heat shock 70 kDa protein 14 | 2.28 | 0.0487 | Protein folding. Stress response |
| B6SZ69 | Zm00001d028630 | Heat shock cognate 70 kDa protein 2 | 2.02 | 0.0398 | Protein refolding. Stress response |
| Strongly repressed proteins by B toxicity in Pachía | |||||
| A0A1D6KC46 | Zm00001d030346 | Hsp20/alpha crystallin family protein | 0.49 | 0.0499 | Chaperone. Response to heat |
| C0PKD9 | Zm00001d052101 | Chaperonin10 | 0.42 | 0.0428 | Chaperone cofactor-dependent protein refolding Protein folding. Chaperone |
| G2XK63 | Zm00001d040257 | T-complex protein 1 subunit beta | 0.27 | 0.0065 | Protein folding. Chaperone |
| B4FR04 | Zm00001d019052 | Peptidylprolyl isomerase | 0.23 | 0.0205 | Protein folding. Rotamase |
| REACTIVE OXYGEN SPECIES (ROS) SCAVENGING PATHWAYS / RESPONSE TO OXIDATIVE STRESS | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6K5D2 | Zm00001d029457 | Nucleoredoxin1 | 2.91 | 0.0117 | Protection against oxidative stress. Cellular oxidant detoxification |
| A0A1D6MSE3 | Zm00001d040721 | Dihydrolipoyl dehydrogenase | 2.30 | 0.0273 | Cell redox homeostasis |
| A0A1D6JPH3 | Zm00001d027769 | Glutathione reductase | 2.21 | 0.0053 | Cell redox homeostasis. Cellular oxidant detoxification. Glutathione metabolic process. |
| K7US39 | Zm00001d009163 | Dihydrolipoyl dehydrogenase | 2.19 | 0.0088 | Cell redox homeostasis |
| RIBOSOME BIOGENESIS | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| B4FPB7 | Zm00001d006100 | 60S ribosomal protein L7a | 2.63 | 0.0051 | Ribosome biogenesis. Maturation of LSU-rRNA |
| K7UTH7 | Zm00001d009596 | GTPase ERA1 chloroplastic | 2.61 | 0.0108 | Ribosome biogenesis. Ribosomal small subunit assembly. rRNA processing |
| B4F7Y1 | Zm00001d031640 | 60S ribosomal protein L7a-1 | 2.39 | 0.0448 | Ribosomal protein. Maturation of LSU-rRNA |
| RNA BINDING AND PROCESSING | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6HT50 | Zm00001d018891 | Chloroplast RNA processing 4 | 2.60 | 0.0142 | mRNA catabolic process |
| SIGNALING | |||||
| Strongly repressed proteins by B toxicity in Pachía | |||||
| P49235 | Zm00001eb411380 | 4-hydroxy-7-methoxy-3-oxo-3,4-dihydro-2H-1,4-benzoxazin-2-yl glucoside beta-D-glucosidase 1, chloroplastic | 0.19 | 0.0090 | Cytokinin signaling pathway |
| STRESS | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| B4F9K2 | Zm00001d005315 | Calcium-dependent lipid-binding (CaLB domain) family protein | 2.11 | 0.0402 | Defense response. Response to stress |
| TRANSCRIPTION AND TRANSLATION PROCESSES | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6LEN8 | Zm00001d035139 | MA3 domain-containing protein | 4.95 | 0.0073 | Negative regulation of transcription, DNA-templated. Regulation of translation |
| Q6R9D1 | GRMZM5G806488 | Ribosomal protein S7 | 3.89 | 0.0202 | Translation. Ribosomal small subunit assembly. Structural constituent of ribosome |
| A0A1D6IAN8 | Zm00001d021400 | Octicosapeptide/Phox/Bem1p (PB1) domain-containing protein / tetratricopeptide repeat (TPR)-containing protein | 3.47 | 0.0323 | RNA processing |
| C0P456 | Zm00001d002789 | Pentatricopeptide repeat-containing protein | 3.26 | 0.0259 | Likely involved in posttranscriptional control of gene expression in organelles |
| A0A1D6NR59 | Zm00001d044745 | Probable alanine--tRNA ligase, chloroplastic | 2.74 | 0.0097 | Translation. Alanyl-tRNA aminoacylation |
| A0A1D6LIV5 | Zm00001d035802 | Phenylalanine--tRNA ligase beta subunit cytoplasmic | 2.56 | 0.0314 | Translation. Phenylalanyl-tRNA aminoacylation |
| B6T5F2 | Zm00001d011992 | 60S ribosomal protein L13 | 2.48 | 0.0387 | Translation. Structural constituent of ribosome |
| A0A1D6HM03 | Zm00001d018274 | Isoleucine--tRNA ligase chloroplastic/mitochondrial | 2.29 | 0.0087 | Translation. Isoleucyl-tRNA aminoacylation |
| A0A1D6QAN9 | Zm00001d051885 | ATG8-interacting protein 1 | 2.28 | 0.0047 | Box C/D RNA 3'-end processing. rRNA processing |
| A0A1D6FRP3 | Zm00001d010530 | Cysteine--tRNA ligase 1 cytoplasmic | 2.26 | 0.0470 | Translation. Cysteinyl-tRNA aminoacylation |
| B4FMD3 | Zm00001d012978 | 40S ribosomal protein S23-2 | 2.13 | 0.0451 | Translation. Structural constituent of ribosome |
| K7UTZ2 | Zm00001d009761 | Spliceosome RNA helicase BAT1 isoform 1 | 2.10 | 0.0455 | RNA splicing. RNA helicase activity |
| K7TY03 | Zm00001d023741 | Alanine--tRNA ligase | 2.07 | 0.0144 | Translation. Alanyl-tRNA aminoacylation |
| B4FYR2 | Zm00001d038865 | 60S ribosomal protein L28 | 2.05 | 0.0275 | Translation. Structural constituent of ribosome |
| B6U151 | Zm00001d002104 | Glutamyl-tRNA(Gln) amidotransferase subunit A, chloroplastic/mitochondrial | 2.02 | 0.0464 | Mitochondrial translation |
| Strongly repressed proteins by B toxicity in Pachía | |||||
| C0P7X7 | Zm00001d034808 | 30S ribosomal protein S6 alpha chloroplastic | 0.50 | 0.0059 | Translation. Structural constituent of ribosome |
| B4FUZ5 | Zm00001d047581 | 30S ribosomal protein S1 | 0.46 | 0.0055 | Translation. Ribosomal protein |
| O50018 | Zm00001d046449 | Elongation factor 1-α | 0.29 | 0.0269 | Translation. Translation elongation factor activity |
| TRANSPORTERS AND TRANSPORT PROCESSES | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| B6SP43 | Zm00001d007597 | ABC family1 | 4.54 | 0.0103 | ATPase-coupled transmembrane transporter activity |
| A0A1D6H2R4 | Zm00001d015569 | H+-exporting diphosphatase | 4.34 | 0.0050 | Ion transport. Pyrophosphate hydrolysis-driven proton transmembrane transporter activity |
| A0A1D6MS70 | Zm00001d040686 | Protein translocase subunit SECA1 chloroplastic | 4.12 | 0.0173 | Protein transport |
| A0A1D6DSW6 | Zm00001d001788 | K+ efflux antiporter 2 chloroplastic | 3.79 | 0.0414 | Chloroplast potassium ion trans-port |
| B6T5R1 | Zm00001d010504 | Ran-binding protein 1 | 3.16 | 0.0492 | Intracellular transport. Protein and mRNA transport. Nucleocytoplasmic transport |
| A0A1D6KSB0 | Zm00001d032615 | Protein TIC110 chloroplastic | 2.35 | 0.0118 | Protein import into chloroplast stroma |
| NOT WELL-KNOWN PROTEINS | |||||
| Strongly induced proteins by B toxicity in Pachía | |||||
| A0A1D6KKK1 | Zm00001d031677 | MtN19-like protein | 2.62 | 0.0464 | Not well determined |
| A0A1D6JI62 | Zm00001d026632 | Stem-specific protein TSJT1 | 2.43 | 0.0283 | Not well determined |
Only proteins considered differentially expressed, namely those with fold-changes ≥2.0 or ≤ 0.5 and P-values ≤ 0.05, are shown in this table. Induced proteins are highlighted with light green rows and repressed proteins with light red rows. 1Proteins ID, Protein identification number in the UniProt database. 2Gene Name, name or ID number of the corresponding gene of the differentially expressed protein as searched in the Maize Genetics and Genomics Database (MaizeGDB; https://www.maizegdb.org/). 3Fold Change, is expressed as the ratio of LFQ intensities (on a logarithmic scale) of proteins between 10 and 0.05 mM B treatments in Pachía. Results were obtained from 3-4 separate plants. 4P-value, statistical level (using Student’s t-test) below ≤0.05, at which differential protein expression was accepted as significant. 5Function/Biological process, annotated biological functions or biological process based on different databases. For more details, see Materials and Methods.
Table 4.
Proteins with higher differential expression in Sama leaves in response to boron (B) toxicity. This table shows the proteins strongly induced or repressed by B toxicity in Sama by comparing their expressions with those of Sama in medium with 0.05 mM B.
Table 4.
Proteins with higher differential expression in Sama leaves in response to boron (B) toxicity. This table shows the proteins strongly induced or repressed by B toxicity in Sama by comparing their expressions with those of Sama in medium with 0.05 mM B.
| Protein ID1 | Gene Name/ID2 | Protein name/Annotation | FC3 | P-value4 | Function/Biological process5 |
|---|---|---|---|---|---|
| AMINO ACID AND PEPTIDE METABOLISMS | |||||
| Strongly induced proteins by B toxicity in Sama | |||||
| B6SKB7 | Zm00001d031013 | Methylcrotonoyl-CoA carboxylase subunit alpha | 3.56 | 0.0049 | Leucine degradation |
| C4J3S6 | Zm00001d004960 | 2-isopropylmalate synthase 1 chloroplastic | 2.17 | 0.0025 | Leucine biosynthesis |
| CARBON ASSIMILATION AND CALVIN CYCLE | |||||
| Strongly repressed proteins by B toxicity in Sama | |||||
| O24574 | Zm00001d004894 | Ribulose bisphosphate carboxylase small chain | 0.33 | 0.0466 | Carbon dioxide fixation |
| P05348 | Rbcs | Ribulose bisphosphate carboxylase small chain, chloroplastic | 0.13 | 0.0096 | Carbon dioxide fixation |
| CELL DIVISION | |||||
| Strongly induced proteins by B toxicity in Sama | |||||
| C0P4T2 | Zm00001d042664 | Patellin-1 | 3.05 | 0.0149 | Cell division and cell cycle |
| NUCLEOTIDE, PURINE AND PYRIMIDINE METABOLISM | |||||
| Strongly repressed proteins by B toxicity in Sama | |||||
| A0A1D6P7V2 | Zm00001d047217 | 5-hydroxyisourate hydrolase | 0.50 | 0.0136 | Purine metabolism |
| PHOTOSYNTHETIC LIGHT REACTIONS | |||||
| Strongly repressed proteins by B toxicity in Sama | |||||
| A0A1D6HS38 | Zm00001d018779 | Oxygen-evolving enhancer protein 2-1 chloroplastic (OEE2-1) | 0.48 | 0.0354 | Photosynthesis. Photosystem II oxygen evolving complex |
| B6SSN3 | Zm00001d015385 | Chlorophyll a-b binding protein, chloroplastic | 0.43 | 0.0435 | Light harvesting in photosystem I |
| B4FWG2 | Zm00001d048422 | Photosynthetic NDH subunit of subcomplex B 2 chloroplastic | 0.41 | 0.0200 | Photosynthetic electron transport flow around photosystem I to produce ATP |
| P06670 | NdhK | NAD(P)H-quinone oxidoreductase subunit K, chloroplastic | 0.38 | 0.0149 | Photosynthetic electron transport coupled photosynthetic proton transport |
| A0A1X7YHF7 | PsbD | Photosystem II D2 protein | 0.35 | 0.0347 | Photosynthetic electron transport in photosystem II |
| P46617 | PetA | Cytochrome f | 0.29 | 0.0161 | Photosynthetic electron transport chain |
| B6SQV5 | Zm00001d049387 | Photosystem II 10 kDa polypeptide | 0.14 | 0.0438 | Photosynthesis. Photosystem II oxygen evolving complex |
| PROTEIN STABILIZATION AND FOLDING | |||||
| Strongly repressed proteins by B toxicity in Sama | |||||
| A0A1D6GJM6 | Zm00001d013455 | Peptidylprolyl isomerase | 0.40 | 0.0275 | Protein folding. Rotamase |
| C4J6Y2 | Zm00001d018077 | Peptidylprolyl isomerase | 0.18 | 0.0422 | Protein folding. Rotamase |
| REACTIVE OXYGEN SPECIES (ROS) SCAVENGING PATHWAYS / RESPONSE TO OXIDATIVE STRESS | |||||
| Strongly induced proteins by B toxicity in Sama | |||||
| B4FSM5 | Zm00001d040341 | Peroxiredoxin | 2.68 | 0.0112 | Cellular response to oxidative stress. Hydrogen peroxide catabolic process. Cell redox homeostasis |
| Strongly repressed proteins by B toxicity in Sama | |||||
| B6U038 | Zm00001d005482 | Thiol-disulfide isomerase and thioredoxins | 0.44 | 0.0223 | Antioxidant activity. Cellular oxidant detoxification. Thioredoxin-dependent peroxiredoxin activity |
| B4FZ35 | Zm00001d002240 | CHL-Zea mays chloroplastic li-pocalin | 0.31 | 0.0272 | Response to oxidative stress. Violaxanthin, antheraxanthin and zeaxanthin interconversion |
| SECONDARY METABOLISM | |||||
| Strongly induced proteins by B toxicity in Sama | |||||
| O64411 | Zm00001d024281 | Polyamine oxidase 1 (PAO1) | 3.34 | 0.0108 | Spermine degradation. Amine and polyamine degradation |
| Strongly repressed proteins by B toxicity in Sama | |||||
| B6TAE7 | Zm00001d028575 | Tropinone reductase | 0.44 | 0.0313 | Tropane alkaloid biosynthesis |
| STRESS | |||||
| Strongly induced proteins by B toxicity in Sama | |||||
| A0A1D6NJS4 | Zm00001d044222 | Tetratricopeptide repeat (TPR)-containing protein | 2.12 | 0.0428 | N-terminal peptidyl-methionine acetylation. Protein maturation |
| TRANSCRIPTION AND TRANSLATION PROCESSES | |||||
| Strongly induced proteins by B toxicity in Sama | |||||
| A0A1D6IBP5 | Zm00001d021507 | Asparagine--tRNA ligase chloroplastic/mitochondrial | 2.66 | 0.0491 | Translation. Asparaginyl-tRNA aminoacylation |
| B4FSE0 | Zm00001d033913 | Alba DNA/RNA-binding protein | 2.48 | 0.0244 | Translational initiation. RNA binding |
| B6T872 | Zm00001d021020 | 60S ribosomal protein L32 | 2.28 | 0.0415 | Translation. Structural constituent of ribosome |
| A0A1D6LIV5 | Zm00001d035802 | Phenylalanine--tRNA ligase beta subunit cytoplasmic | 2.23 | 0.0093 | Translation. Phenylalanyl-tRNA aminoacylation |
| B4FJ27 | Zm00001d011741 | 40S ribosomal protein S24 | 2.13 | 0.0363 | Translation. Structural constituent of ribosome |
| TRANSPORTERS AND TRANSPORT PROCESSES | |||||
| Strongly induced proteins by B toxicity in Sama | |||||
| B6SP43 | Zm00001d007597 | ABC family1 | 2.69 | 0.0125 | ATPase-coupled transmembrane transporter activity |
Only proteins considered differentially expressed namely those with fold-changes ≥2.0 or ≤ 0.5 and P-values ≤ 0.05, are shown in this table. Induced proteins are highlighted with light green rows and repressed proteins with light red rows. 1Proteins ID, Protein identification number in the UniProt database. 2Gene Name, name or ID number of the corresponding gene of the differentially expressed protein as searched in the Maize Genetics and Genomics Database (MaizeGDB; https://www.maizegdb.org/). 3Fold Change, is expressed as the ratio of LFQ intensities (on a logarithmic scale) of proteins between 10 and 0.05 mM B treatments in Sama. Results were obtained from 3-4 separate plants. 4P-value, statistical level (using Student’s t-test), at which differential protein expression was accepted as significant (≤0.05). 5Function/Biological process, annotated biological functions or biological process based on different databases. For more details, see Materials and Methods.
Table 5.
Proteins with higher differential expression between Sama and Pachía leaves under boron (B) toxicity condition. This table shows the strongly up- or down-accumulated proteins in Sama in media with 10 mM B compared to those of Pachía in 10 mM B.
Table 5.
Proteins with higher differential expression between Sama and Pachía leaves under boron (B) toxicity condition. This table shows the strongly up- or down-accumulated proteins in Sama in media with 10 mM B compared to those of Pachía in 10 mM B.
| Protein ID1 | Gene Name/ID2 | Protein name/Annotation | FC3 | P-value4 | Function/Biological process5 | ||
|---|---|---|---|---|---|---|---|
| AMINO ACID AND PEPTIDE METABOLISMS | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6ICL3 | Zm00001d021596 | Adenosine 5-phosphosulfate reductase-like1 | 2.33 | 0.0417 | Cysteine biosynthetic process. Sulfate reduction | ||
| CARBON ASSIMILATION AND CALVIN CYCLE | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6EXF1 | Zm00001d006520 | PDK regulatory protein1 | 2.16 | 0.0167 | Regulation of C4 photosynthetic carbon assimilation cycle | ||
| CARBOHYDRATE METABOLISM | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| Q9SYS1 | Zm00001d021702 | β-amylase | 2.63 | 0.0499 | β-amylase activity. Starch degradation | ||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6K5L6 | Zm00001d029502 | Glucose-6-phosphate 1-dehydrogenase | 0.36 | 0.0411 | Pentose phosphate pathway | ||
| A0A1D6LY56 | Zm00001d037480 | Alkaline α galactosidase 2 | 0.33 | 0.0438 | Carbohydrate metabolic process | ||
| CELL DEATH | |||||||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6JNJ8 | Zm00001d027656 | Lethal leaf-spot 1 | 0.32 | 0.0016 | Cell death. Chlorophyll catabolic process | ||
| CELL DIVISION | |||||||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6JH24 | Zm00001d026532 | Protein RCC2 | 0.42 | 0.0214 | Cell division | ||
| CELL WALL | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6MWZ7 | Zm00001d041578 | Glossy6 | 3.27 | 0.0403 | Epicuticular wax accumulation. Intracellular trafficking of cuticular waxes | ||
| DNA AND CHROMATIN ORGANIZATION AND DNA REPAIR | |||||||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| B4FQA5 | Zm00001d018981 | Histone1a | 0.35 | 0.0318 | Chromosome condensation. Nucleosome assembly | ||
| B6TGH8 | Zm00001d034479 | Histone H1 | 0.31 | 0.0138 | Chromosome condensation. Nucleosome assembly. Nucleosome positioning | ||
| LIPID METABOLISM | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| Q8W0V2 | Zm00001d033623 | Lipoxygenase 3 | 5.06 | 0.0455 | Fatty acid and oxylipin biosynthesis | ||
| Q06XS3 | Zm00001d053675 | Lipoxygenase 10 | 3.44 | 0.0247 | Fatty acid and oxylipin biosynthesis | ||
| OTHERS | |||||||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| B6TY16 | Zm00001d040331 | SUN domain protein2 | 0.41 | 0.0262 | Nuclear envelope organization | ||
| B4F7V3 | Zm00001d021582 | Protein phosphatase 2C isoform ε | 0.39 | 0.0214 | Protein dephosphorylation | ||
| A0A1D6HUN3 | Zm00001d019040 | D-2-hydroxyglutarate dehydrogenase mitochondrial | 0.33 | 0.0024 | Photorespiration | ||
| OXIDATION AND REDUCTION PROCESSES | |||||||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| B4F987 | Zm00001d020984 | Putative sarcosine oxidase | 0.23 | 0.0321 | Sarcosine oxidase activity | ||
| PHOTOSYNTHETIC LIGHT REACTIONS | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| B4FR80 | Zm00001d033098 | Post-illumination chlorophyll fluorescence increase (ZmPIFI) | 2.52 | 0.0097 | Chlororespiration | ||
| A0A1D6HS38 | Zm00001d018779 | Oxygen-evolving enhancer protein 2-1 chloroplastic (OEE2-1) | 2.31 | 0.0325 | Photosynthesis. Photosystem II oxygen evolving complex | ||
| PIGMENT BIOSYNTHESIS | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6JHX0 | Zm00001d026603 | Magnesium-chelatase subunit ChlH1 chloroplastic (ChlH1) | 2.90 | 0.0484 | Chlorophyll biosynthetic process | ||
| PROTEIN STABILIZATION AND FOLDING | |||||||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6KE29 | Zm00001d030725 | Heat shock protein 70 | 0.43 | 0.0406 | Protein refolding. Protein folding chaperone. Cellular response to unfolded protein | ||
| RESPIRATION (GLYCOLISIS, TCA CYCLE AND MITOCHONDRIAL ELECTRON TRANSFER) | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| B4G1C9 | Zm00001d023606 | Dihydrolipoamide acetyltransferase component of pyruvate dehydrogenase complex | 2.04 | 0.0332 | Acetyl-CoA biosynthetic process from pyruvate | ||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6MAK9 | Zm00001d038792 | Phosphotransferase | 0.49 | 0.0331 | Glycolysis | ||
| SECONDARY METABOLISM | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| O64411 | Zm00001d024281 | Polyamine oxidase 1 (PAO1) | 5.15 | 0.0007 | Spermine degradation. Amine and polyamine degradation | ||
| TRANSCRIPTION AND TRANSLATION | |||||||
| Strongly up-accumulated proteins in Sama in media with 10 mM B | |||||||
| B4FP25 | Zm00001d047296 | 40S ribosomal protein S19 | 6.38 | 0.0289 | Translation. Structural constituent of ribosome. Ribosomal small subunit assembly | ||
| B6TDF7 | Zm00001d019898 | Plastid-specific 30S ribosomal protein 2 | 2.31 | 0.0243 | Ribosomal protein. Ribonucleoprotein complex. RNA-binding | ||
| C0PEC4 | Zm00001d032420 | 30S ribosomal protein S5 chloroplastic | 2.12 | 0.0487 | Translation. Structural constituent of ribosome | ||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| B6SX73 | Zm00001d016549 | 60S ribosomal protein L35 | 0.42 | 0.0284 | Translation. Structural constituent of ribosome | ||
| Q6R9D1 | GRMZM5G806488 | Ribosomal protein S7 | 0.35 | 0.0426 | Translation. Structural constituent of ribosome. Ribosomal small subunit assembly | ||
| Transporter and transport processes | |||||||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6H2R4 | Zm00001d015569 | H+-exporting diphosphatase | 0.33 | 0.0169 | Ion transport. Pyrophosphate hydrolysis-driven proton transmembrane transporter activity | ||
| A0A1D6K7N5 | Zm00001d029762 | Hexose transporter | 0.20 | 0.0439 | Hexose transporter | ||
| Unknown or not well determined | |||||||
| Strongly down-accumulated proteins in Sama in media with 10 mM B | |||||||
| A0A1D6KKK1 | Zm00001d031677 | MtN19-like protein | 0.23 | 0.0121 | Not well determined | ||
Only proteins considered differentially expressed namely those with fold-changes ≥2.0 or ≤ 0.5 and P-values ≤ 0.05, are shown in this table. Induced proteins are highlighted with light green rows and repressed proteins with light red rows. 1Proteins ID, Protein identifying number in the UniProt database. 2Gene Name, name or ID number of the corresponding gene of the differentially expressed protein as searched in the Maize Genetics and Genomics Database (MaizeGDB; https://www.maizegdb.org/). 3Fold Change, is expressed as the ratio of LFQ intensities (on a logarithmic scale) of proteins between Sama and Pachía in media with 10 mM B. Results were obtained from 3-4 separate plants. 4P-value, statistical level (using Student’s t-test) below ≤0.05, at which protein differential expression was accepted as significant. 5Function/Biological process, annotated biological functions or biological process based on different databases. For more details, see Materials and Methods. .
Table 6.
Proteins exclusively detected in Pachía or Sama leaves in at least one B treatment.
| Protein ID1 | Gene Name/ID2 | Protein name/Annotation | Function/Biological process3 |
|---|---|---|---|
| DNA AND CHROMATIN ORGANIZATION AND DNA REPAIR | |||
| Protein exclusively detected in Pachía in 10 mM B | |||
| A0A1D6KX75 | Zm00001d033247 | Nfc103a - nucleosome/chromatin assembly factor C | Nucleosome/chromatin assembly. DNA repair. Chromatin remodeling, regulation of DNA-templated transcription |
| OTHERS | |||
| Protein exclusively detected in Sama in both B treatments | |||
| K7VAT7 | Zm00001d046569 | Protein kinase superfamily protein with octicosapeptide/Phox/Bem1p domain | Protein serine/threonine kinase activity. Protein phosphorylation |
| REACTIVE OXYGEN SPECIES (ROS) SCAVENGING PATHWAYS / RESPONSE TO OXIDATIVE STRESS | |||
| Protein exclusively detected in Pachía in both B treatments | |||
| B4FKV6 | Zm00001d014341 | Peroxidase 54 | Response to oxidative stress. Peroxidase activity |
| TRANSCRIPTION AND TRANSLATION | |||
| Protein exclusively detected in Pachía in 10 mM B | |||
| A0A096RFR6 | Zm00001d039518 | Eukaryotic translation initiation factor 3 subunit A (eIF3a) | Translation initiation factor activity. Protein synthesis. Formation of cytoplasmic translation initiation complex |
| Transporter and transport processes | |||
| Proteins exclusively detected in Pachía in both B treatments | |||
| A0A1D6EU13 | Zm00001d006238 | Calcium lipid binding protein-like | Lipid transport |
| A0A1D6JN64 | Zm00001d027580 | Outer mitochondrial membrane porin1 (ommp1) | Voltage-gated anion channel activity. Inorganic anion transport, transmembrane transport, anion transmembrane transport |
| Protein exclusively detected proteins in Sama in both B treatments | |||
| Q7Y1W6 | Zm00001d018693 | Pentatricopeptide repeat 2 (PPR2) | Chloroplast translation |
| Unknown or not well determined | |||
| Protein exclusively detected proteins in Sama in both B treatments | |||
| A0A1D6DWG9 | Zm00001d002089 | Tetratricopeptide repeat (TPR)-like superfamily protein | Unknown |
1Proteins ID, Protein identification number in the UniProt database. 2Gene Name, name or ID number of the corresponding gene of the identified protein as searched in the Maize Genetics and Genomics Database (MaizeGDB; https://www.maizegdb.org/). 3Function/Biological process, annotated biological functions or biological process based on different databases. For more details, see Materials and Methods. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



