
Submitted:
05 May 2023
Posted:
05 May 2023
You are already at the latest version
Abstract
Kombucha is a popular sparkling sugared tea, fermented by a symbiotic culture of acetic acid bacteria (AAB) and yeast. The demand for Kombucha is increasing worldwide mainly due to its perceived health benefits and appealing sensory properties. This study isolated and characterised the dominant AAB and yeast from starter culture and Kombucha broth after 0, 1, 3, 5, 7, 9, 11, and 14 days of fermentation at ambient temperature (~22°C). Changes in the microbial composition were associated with variations in the physico-chemical characteristics of Kombucha tea, such as pH, titratable acidity, and total soluble solids (TSS). During fermentation, the acidity increased, and TSS decreased. The yield, moisture content and water activity of the cellulosic pellicles which had developed by the end of fermentation were attributed to the presence of AAB. Yeast and AAB were isolated from the Kombucha samples using glucose yeast extract mannitol ethanol acetic acid (GYMEA) and yeast extract glucose chloramphenicol (YGC) media respectively, using serial dilutions. Phenotypic and taxonomic identification of AAB and yeast were determined by morphological and biochemical characterisation, followed by sequence analysis of the ribosomal RNA gene (16S rRNA for AAB and ITS for yeast). The dominant AAB species in the cellulosic pellicle and Kombucha broth was identified as Komagataeibacter rhaeticus. The yeast isolates belonged to Debaryomyces prosopidis and Zygosaccharomyces lentus.
Keywords:
Kombucha
; Acetic acid bacteria
; yeast
; Komagataeibacter rhaeticus
; Zygosaccharomyces lentus
; Debaryomyces prosopidis
1. Introduction
Kombucha is a popular functional sparkling fermented tea beverage consumed for more than 5000 years [1]. The taste of Kombucha varies from slightly fruity, sour and fizzy to vinegar-like depending on the fermentation conditions [2]. Regular consumption of Kombucha is perceived to confer numerous health-promoting benefits including antioxidants, antimicrobial, anti-carcinogenic, detoxification, antitumor and antihypertensive activities [2,3,4,5]. However, most of the perceived health benefits are based on in vitro models. Therefore, in vivo clinical trials are necessary to confirm the functional activities.
Kombucha is usually fermented from sucrose-sweetened black or green tea with an undefined symbiotic culture of bacteria and yeast (SCOBY) embedded in a cellulosic pellicle at ambient temperature for 7-14 days [6,7,8]. Other substrates such as oolong tea, thyme, lemon balm, peppermint, rosemary, wheat grass, guava and oak leaves, fruit juices, milk and laver have also been used to make Kombucha [5,9,10,11]. During fermentation, sucrose is firstly hydrolysed by yeast to glucose and fructose. The glucose produced is then metabolised into gluconic and glucuronic acids by the AAB, whereas fructose is metabolised by yeast to produce ethanol and carbon dioxide. The resulting alcohol is further metabolised by the AAB into acetic acid. After fermentation, Kombucha has a complex chemical composition and favourable bioactive components including organic acids (acetic, gluconic, lactic and glucuronic acids), polyphenols (epicatechin, epigallocatechin, epicatechin gallate, and epigallocatechin gallate), vitamins (B1, B2, B6, B12, and C), minerals (Cu, Fe, Mn, Ni, Zn), ethanol and amino acids [3,9,12]. Variation in the chemical and nutritional metabolites present in Kombucha are attributed to the diverse microbial composition of the SCOBY, fermentation conditions, and types of tea (substrate) used [2,13].
The microbial composition of Kombucha is diverse and varies between fermentations, but is mainly composed of AAB and yeast [14]. Previous studies indicate that gluconic acid-producing AAB are the dominant bacteria in the Kombucha culture [15]. The most common AAB include Komagataibacter (K.) kombuchae, K. saccharivorans, K. rhaeticus, K. intermedius, K. europaeus, K. hansenii, and K. xylinus; Gluconobacter (G.) oxydans, G. potus; Gluconacetobacter (G.) sacchari, G. sp A4; Acetobacter (A.) musti, A. pasteurianus, A. aceti, and A. nitrogenifigens sp. nov. [4,12,16,17]. Small amounts of lactic acid bacteria (LAB) have also been reported in Kombucha [6], including Lacticaseibacillus casei, Lactiplantibacillus plantarum, Lactobacillus rhamnosus, Lactobacillus mali, Pediococcus pentosaceus, P. acidiliactici, Liqourilactobacillus nagelii, Oenococcus oeni and Bifidobacterium [18,19,20,21,22,23]. The most commonly reported yeast species in Kombucha are Brettanomyces (B.) bruxellensis, B. lambicus, B. intermedius; Candida (C.) albican, C. kefir, C. zemplinina; Dekkera (D.) bruxelensis, D. anomala; Hanseniaspora (H.) valbyensis; Pichia (P.) fermentans, P. occidentalis, P. kudriavzevii; Saccharomyces (S.) cerevisiae; Schizosaccharomyces (S.) pombe; Zygosaccharomyces (Z.) bailii and Z. rouxii [13,24,25,26,27,28]. Interactions between the bacteria and yeast produces Kombucha with different chemical compositions [29].
The unique cellulose forming characteristics of Kombucha may be useful for a variety of new applications in medicine and food production. The SCOBY has been used as artificial skin to treat wounds or burnt skin [30]. Dehydrated SCOBY may be used to produce Kombucha berry gummies, and SCOBY hamburger due to their unique texture and flavour [12]. Therefore, it is important to determine and optimise the yield of the cellulosic pellicle from Kombucha fermentation for these innovative applications.
Current challenges for industrial production of Kombucha include the limited knowledge about the dynamics of its microbial community such as the distribution of the microbial composition and lack of standardised fermentation process [6,31]. The diversity in the microbial composition of Kombucha may be due to differences in temperature, tea types, climate and also environmental conditions [7]. An improved understanding of the microbial composition of the SCOBY and changes in the levels of microorganisms during fermentation will contribute to better control of Kombucha production.
This study determined the dominant microbial composition, and physico-chemical characteristics of the Kombucha tea broth during fermentation and the cellulosic pellicle. The microbial communities of Kombucha and cellulosic pellicle during fermentation were examined by culture-dependent methods and ribosomal genotyping.
2. Materials and Methods
2.1. Preparation of Fermented Kombucha
The Kombucha starter culture (cellulosic pellicle and liquid broth) was obtained from commercial Kombucha available in local supermarkets in Auckland, New Zealand in 2021. The samples were transported to the Food Technology Department at Massey University, Auckland, under a cold chain maintained at 4 ℃ after which they were stored under refrigeration until required for analysis (Figure 1).
2.2. Kombucha Fermentation Progress
Laboratory scale fermented Kombucha was prepared using the modified methods of Greenwalt, Steinkraus and Ledford [14] and Jayabalan, Malbasa, Loncar, Vitas and Sathishkumar [15]. Sucrose (50 g/L) (white sugar, Chelsea, New Zealand) was added into boiling water and mixed until completely dissolved. Black tea bags (Bell®, New Zealand) (5 g/L) from a local supermarket were added to the mixture and allowed to infuse for 10 min then removed. The sugared tea was cooled to ambient temperature (~22 °C). The entire SCOBY (25 g/L) was gently placed on the surface of the sugared tea and previous fermented Kombucha tea broth (20 %, v/v) was added to reduce the pH to prevent growth of spoilage microorganisms. The container was covered with a clean disposable cloth to keep out foreign matter. Fermentation was carried out at 22 °C for 14 days in a water bath (T100, Grant Optima, UK). Kombucha tea broth samples were collected for immediate physico-chemical and microbiological analyses at day 0, then every 2 days until the end of fermentation. The newly formed cellulosic pellicle (25 g) and samples (20 mL) of the fermented Kombucha broth were collected at the end of fermentation. The two components were maintained in fresh sugared tea broth at 4 °C for the next fermentation.
2.3. Physico-Chemical Characteristics of Kombucha Broth Samples During Fermentation
2.3.1. Determination of pH, Titratable Acidity (TA), and Total Soluble Solids (TSS) of Kombucha Tea Broth during Fermentation
The pH of the Kombucha tea broth collected at different time points was measured using a Sartorius glass electrode pH meter (Model PH-11, Germany). Titratable acidity was determined by acid-base titration. Briefly, Kombucha tea broth samples (20 mL) from different stages of fermentation were titrated against standardised 0.1 M sodium hydroxide to pH 8.2. The calculated TA was expressed as a percentage (%) of acetic acid per gramme of sample, as it is the major organic acid in Kombucha (Equation (1)) [32]. The TSS of Kombucha tea broth samples were determined using a refractometer (Atago, pr-32 alpha, UK), according to Amarasinghe, et al. [33].
VNaOH = volume of NaOH (mL)
MNaOH = molarity of NaOH (M)
Vsample = volume of sample (mL)
2.3.2. Determination of Wet Yield, Water Holding Capacity (WHC), Moisture Content and Water Activity of Cellulosic Pellicle at the End of the Fermentation
The wet yield of the fresh pellicle produced during fermentation was determined using the method of Aung and Eun [34] with slight modifications. The weight of the initial mother pellicle used in the fermentation was measured at day 0 and at the end of fermentation (day 14). The wet yield of tea fungus was calculated using Equation (2).
The water holding capacity (WHC) and moisture content were determined using the methods of Avcioglu, et al. [35] with slight modifications. The fresh cellulosic pellicle was cut into small pieces and mixed by blending (BB25EP, Waring, USA) for one minute. About 5-10 g of the pellicle were weighed into an aluminium pan and dried at 70 °C overnight. The WHC and moisture content were calculated using Equations (3) and (4), respectively.
W1: weight of empty container (g)
W2: weight of container and sample before drying (g)
W3: weight of container and sample after drying (g)
The water activity of the pellicle was measured using a water activity meter (AQUALAB 4TE, USA).
2.3. Isolation and Enumeration of AAB, LAB and Yeast from Kombucha
Samples of both the cellulosic pellicle and Kombucha tea collected separately during fermentation were used for the isolation and identification of AAB, LAB and yeast. Serial dilutions (10-6) of the Kombucha tea broth and pellicle were plated on appropriate microbiological media. Isolation of AAB from the Kombucha tea/pellicle was performed on modified glucose yeast extract mannitol ethanol acetic acid agar (GYMEA) containing 5% D-glucose (w/v) (ThermoFisher, Waltham, Massachusetts, USA), 2.5% mannitol (w/v) (ThermoFisher, Waltham, Massachusetts, USA), 1% yeast extract (w/v) (Sigma-Aldrich, St. Louis, Missouri, USA), bacteriological agar (ThermoFisher, Waltham, Massachusetts, USA) using a slightly modified method of Wang, Rutherfurd-Markwick, Zhang and Mutukumira [2]. Absolute ethanol (2% v/v) (Sigma-Aldrich, St. Louis, Missouri, USA), glacial acetic acid (0.5% v/v) were added after autoclaving. The addition of cycloheximide (100 mg/L) and penicillin (1 mL/L) were intended to inhibit the growth of yeast and LAB respectively. Samples for isolation of LAB and yeast were plated on De Man, Rogosa and Sharpe (MRS) agar plates (ThermoFisher, Waltham, Massachusetts, USA) and yeast extract chloramphenicol (YGC) plates, respectively. The AAB were cultivated at 30 ℃ for 5-7 days; yeast was incubated at 25 ℃ for 5 days aerobically. The potential LAB were incubated anaerobically at 37 ℃ for 72 h. Colonies of the grown microorganisms were enumerated, and results were expressed as colony forming unit (CFU)/mL or CFU/g. Representative AAB, LAB and yeast isolates were purified by successive streaking on glucose yeast extract (GY), MRS and potato dextrose agar (PDA), respectively. The purified cultures were stored in 20% glycerol (w/w) at -80 °C for long term preservation, and on GY, MRS or PDA agar plates at 4 °C with monthly sub-culturing for routine use [36].
2.4. Phenotypic Characterisation of AAB and Yeast Isolated from Kombucha Tea Broth and Cellulosic Pellicle during Fermentation
2.4.1. Phenotypic Characterisation of AAB
Three to five presumptive AAB isolates were randomly selected from the highest dilution (10-6) of Kombucha tea broth at each time interval during fermentation [2]. A loopful of freshly purified AAB culture was used to determine phenotypic characterisation. The purified AAB isolates were incubated on GY agar plates at 30 °C for 5-7 days and GY broth overnight with shaking at 170 rpm (Orbital Shakers, Ohaus, Australia). The AAB culture was Gram-stained to examine cell morphology under oil immersion using the Carl Zeiss Transmission light microscope (Model HBO 50/AC, Germany), cell length of AAB isolates was measured using the AvixVision microscope software 4.8.1. Biochemical tests were then conducted on the gram-negative isolates. Catalase positive tests were determined by formation of air bubbles on young purified colonies after the addition of 1-2 drops of 6% H2O2 (v/v) (Sigma-Aldrich, St, Louis, Missouri, USA) [2,37]. Growth at different temperatures (25, 30, and 37 ℃) was conducted on GY gar plates and incubated aerobically for 5-7 days. Growth at low pH was determined in GY broth at pH 2 and pH 3 and incubated aerobically at 30 ℃ for 24 h. The oxidase test was conducted with oxidase strips (Oxoid, UK), and colour change was observed for 30 s [38]. The oxidation of ethanol to acetic acid and then oxidation acetic acid to water and CO2 by AAB was carried out using Carr medium [39]. The ethanol tolerance test was performed on yeast extract agar plates containing 2%, 4%, 6%, 8% and 10% (v/v) absolute ethanol [40]. Acid produced from sucrose, D-glucose, trehalose and lactose was determined using the method of Arifuzzaman, et al. [41] with slight modifications. Test of the formation of cellulose by the AAB isolates was determined using the modified methods of previous reports [2,42,43]. The AAB isolates were inoculated in Hestrin-Schramm (HS) broth for 5-7 days at 30 ℃. A small portion of the cellulose developed on the surface of the broth was carefully transferred into a 2-mL micro centrifuge tube mixed with 1 mL of 0.1 M NaOH solution and heated at 90 °C on a hotplate (Benchmark Scientific, USA) for 45 min. Oxidation of lactate and acetate of AAB isolates were determined using published methods [2,44]. Ketogenesis of glycerol to dihydroxyacetone (DHA) was performed with the modified method of Swings [45]. The AAB isolates were inoculated in glycerol yeast extract agar plates for 5-7 days at 30 °C, the surface of the agar plates was then flooded with Benedict’s reagent and incubated at room temperature for another 3 h to observe any colour change.
2.4.2. Phenotypic Characteristics of Yeast
Three to five presumptive yeast isolates were randomly collected at each time interval during fermentation. The purified yeast isolates from Kombucha tea broth and cellulosic pellicle were plated on PDA plates and incubated at 25 °C for 5 days. The cell morphology of yeast isolates was observed using the methylene blue staining method and the cell length was determined using the Carl Zeiss Transmission light microscope (Model HBO 50/AC, Germany) [2,46,47]. The young, fresh and representative budding yeast isolates were then characterised using biochemical tests. The cycloheximide resistance test was conducted using the modified method of Kurtzman, et al. [48]. The yeast isolates were streaked on the PDA plates containing 0.01% and 0.1% cycloheximide (w/v) respectively and incubated at 25 °C for 5 days. The formation of colonies indicated resistance to the antibiotic. Growth at different temperature was assessed on PDA plates and yeast isolates were incubated at 25, 30, and 37 °C for 5 days, respectively. Resistance to high osmotic pressure was determined using the modified method of Kurtzman, et al. [49]. Yeast isolates were inoculated on agar containing sodium chloride (10% w/v) and D-glucose (5% w/v) and incubated at 25 ℃ for 5 days. The growth of colonies suggested that the isolates had resistance to high osmotic pressure. The glacial acetic acid tolerance test was conducted on media containing (5% w/v) D-glucose, (1% w/v) tryptone, (1% w/v) yeast extract, (2% w/v) agar and (1% v/v) glacial acetic acid. Growth at low pH (2 and 3) and acid produced from sucrose, D-glucose, trehalose and lactose were previously described in Section 2.4.1.
2.5. Sequence Analysis of Ribosomal RNA Genes
Total DNA from representative AAB and yeast isolates were extracted using the Promega Wizard Genomic Purification Kit (ThermoFisher, Waltham, Massachusetts, USA) following the manufacturers’ instructions. The full-length 16S rRNA gene of AAB isolates was amplified by the polymerase chain reaction (PCR) using universal primer 27F and 1492R [2,50] and Taq DNA polymerase from Invitrogen (ThermoFisher, New Zealand).
For yeast isolates, the internal transcribed spacer (ITS) regions were amplified by PCR using primers ITS1 and ITS4 [51]. The PCR products were purified with E.Z.N.A Cycle Pure Kit (Omega Bio-Tek InC., Norcross), and subsequent sequencing of the amplicons was performed by Massey Genome Service (Palmerston North, New Zealand), using primers 27F and 1492R, plus an internal primer 533F for 16S rRNA genes [52] and primers ITS1 and ITS4 for the ITS region. DNA sequences were processed using Geneious 9.0.5 (Biomatters Ltd., Auckland, New Zealand).
2.6. Statistical Analysis
The physicochemical and phenotypical characteristics experiments were performed in triplicate. Data on pH, Titratable acidity, total soluble solids, yield, water activity, and moisture content were analysed by Microsoft Excel 2016 (Microsoft, USA) and represented as mean ± standard deviation. Data on pH, titratable acidity, total soluble solids, viable cell counts were analysed by one-way analysis of variance (ONEWAY-ANOVA) using the IBM SPSS version 26 (IBM, USA) to determine significant differences between the means (p<0.05). The AAB and yeast DNA were analysed by Geneious 9.1.8 (Biomatters, Ltd., Auckland). The closest homologues or best hits were identified by searching the EzBioCloud 16S rRNA gene database for AAB and ITS database of NCBI using Basic Local Alignment Tool (BLAST) [53].
3. Results and Discussion
3.1. Physico-chemical Characteristics of Kombucha Samples during Fermentation
3.1.1. Acidity and TSS of Kombucha during Fermentation
Changes in acidity of Kombucha tea broth during fermentation are shown in Figure 2. The pH of Kombucha tea broth decreased from 3.94±0.04 to 3.16±0.01 and the TA increased from 0.02±0.00 to 0.14±0.13% during fermentation for 14 days at 22 °C (p<0.05). A sharp decrease in pH was observed during the first three days of fermentation and then it decreased slowly to the lowest pH observed at the end of fermentation (3.16±0.01). Meanwhile, TA increased steadily throughout fermentation. Such changes in acidity are expected. During fermentation, sucrose is hydrolysed by the yeast into glucose and fructose. The AAB then convert the two simple sugars into various organic acids such as acetic, gluconic, glucuronic acids which contribute to the acidity and functional activities such as the antimicrobial activity of Kombucha [29]. Changes in the hydrogen ion concentration (pH) and TA during fermentation can affect the growth of fermenting microorganisms like AAB and yeast in Kombucha [3]. Low acidic beverages (<pH 4) are considered as microbiologically safe as it can prevent the growth of pathogenic microorganisms [14]. Currently, there is no standardised pH range for Kombucha products. However, a very low pH may affect the overall sensory characteristics of Kombucha conferring a very sour flavour [54]. It is not recommended to produce Kombucha with a pH < 2.5 as it may be harmful to some consumers due to the high acidity [55]. A pH range between 2.5 and 4.2 is considered desirable and safe for Kombucha beverages [56].
3.2. Wet Yield, WHC, Moisture Content and Water Activity of Cellulosic Pellicle at the End of Fermentation.
An approximately 2-cm thick transparent cellulosic layer was observed on the surface of the mother pellicle during Kombucha fermentation. The AAB which produce the cellulosic pellicle are aerobic and therefore the pellicle floats on top of the broth [35]. The size of the cellulose pellicle gives an indication of the success of the fermentation process [5]. In this study, the wet yield of SCOBY was 27.75±0.05 % by the end of fermentation (Table 1). Bacterial cellulose has been widely used as an environmentally friendly nanomaterial in the cosmetic and food industries due to its high mechanical strength, high elasticity, high water holding capacity, and chemical stability [35,57]. The WHC was 17.82±0.16 g water/g dry pellicle which was lower than cellulose produced by Gluconacetobacter xylinus (98.5g water/g dry pellicle) in a previous study [58]. WHC indicates the water retained by the cellulosic pellicle, a higher WHC results in a loose cellulosic structure which is desirable for nanomaterial applications [59].
3.3. Isolation and Enumeration of AAB and Yeast during Fermentation of Kombucha
Changes (p<0.05) in the microbial population during fermentation of the Kombucha tea broth are shown in Figure 4. Yeast and AAB were found in the Kombucha tea broth during fermentation and the pellicle. No LAB were present in the Kombucha in this study. Kombucha was prepared using back-slopping fermentation, with the initial concentrations of yeast and AAB being 3.08±0.42 log CFU/mL and 3.08±0.48 log CFU/mL respectively in the Kombucha tea broth. The microbial population of yeast and AAB in the Kombucha tea broth increased by nearly 2 log CFU/mL during fermentation for 14 days (Figure 4). Although the initial cell counts of AAB were lower, the final population was slightly higher than the yeast count. Both AAB and yeast counts increased markedly from day 0 to day 5 and then slowly to the end of fermentation; yeast and AAB counts remained relatively similar from day 5 to day 11 with small fluctuations visible between day 9 and day 11. The cell counts from the cellulosic pellicle were 7.44±0.31 log CFU/mL for AAB and 7.30±0.06 log CFU/mL for yeast, which were higher than the number of microorganisms present in the tea broth (p<0.05).
3.4. Phenotypic Characterisation of AAB and Yeast Isolated from the Kombucha Broth and Cellulosic Pellicle
3.4.1. Morphology of AAB and Yeast Isolated from Kombucha Broth and Cellulosic Pellicle
Sixty-eight strains (n=29 AAB, n=39 yeast) were isolated from the black tea Kombucha during 14 days of fermentation. Based on colony morphology there was one group of AAB and two distinguishable groups of yeast isolated from the Kombucha tea broth and pellicle. The AAB colonies had a circular shape, smooth surfaces of brownish-red colour with an entire margin. The diameter of the AAB colonies ranged from 1.0-1.5 mm; the morphology of yeast grown on YGC plates is shown in Table 2. Different morphologies of the yeast colonies indicated that the fungal isolates could belong to different species.
Based on Gram staining reactions and cell morphology, 28 of the 29 AAB isolates from Kombucha tea broth and pellicle were Gram-negative [39]. All the presumptive AAB cells were rod-shaped with a similar cell length (~1.50 µm), and most occurred as singles. The cell morphology of AAB isolates in this study were similar to Glucoacetobacter or Komagataeibacter [60,61]. The formation of multipolar budding was observed from both groups (I and II) of yeast isolates. The average length of yeast cells ranged from 5.00 to 11.00 µm for the two groups of fungi, and were present in single, pairs or in groups. The yeast cells were ovoidal to ellipsoidal shape in group I and spherical shape in group II The presumptive AAB isolates (n=28) and yeast (n=39) from Kombucha tea broth and pellicle were subjected to further characterisation.
3.4.2. Phenotypic Characteristics of AAB and Yeast Isolated from Kombucha Tea Broth and Pellicle
The phenotypic and differential biochemical characteristics of the AAB isolates are shown in Table 3. Based on their biochemical profiles, the 28 AAB isolates were divided into six groups, and based on the data presented in Table 3, they were presumed to belong to the genus Komagataeibacter. All six groups of AAB isolates were oxidase-negative and catalase positive and were able to grow at 25, 30, and 37 ℃ after 5-7 days incubation. The bacteria were able to grow at pH 3 but not pH 2. All the AAB isolates produced cellulose after 5-7 days incubation at 30 ℃ without shaking. The transparent cellulose was not soluble after heating in aqueous NaOH. The production of cellulose is a typical characteristic of the species of Gluconacectobacter and Komagataeibacter [60]. All six groups of AAB isolates oxidised both lactate and acetate as the agar plates turned to blue.
All six groups of AAB isolates were able to grow in agar plates containing 2% (v/v) ethanol (alcoholic tolerance test). However, the six groups of AAB exhibited different growth profiles in media containing 4% to 10% (v/v) ethanol. Group IV isolates were able to grow in the presence of 10% ethanol, while group II and III were not able to grow in the presence of 4% (v/v) ethanol. The ethanol concentration of non-alcoholic Kombucha from previous studies was reported to between 0.2% to 1.1% (v/v) at the end of fermentation [3,6]. However, higher alcohol content above the tolerance level of AAB has been reported post-fermentation of Kombucha [2]. Therefore, it is important to monitor the fermentation process of Kombucha as high levels of ethanol are undesirable for the growth of AAB.
All six groups of isolates oxidised ethanol to acetic acid as the agar turned to yellow after three days incubation at 30 ℃. Results suggest that groups II, IV, and VI oxidised acetic acid to water and CO2 after incubation for >5 days as the yellow became purple. The other three groups were not able to oxidise acetic acid probably because they were deficient in succinate dehydrogenase and α-ketoglutarate dehydrogenase [62]. All six groups were also able to oxidise glycerol to dihydroxyacetone (DHA). In the food industry, AAB convert glycerol to produce DHA, producing a crust-like taste in winemaking [63]. The presence of DHA in Kombucha may confer a sweet and pleasant aroma and enhance the production of cellulose that can positively impact the sensory characteristics of Kombucha [2].
Table 4 shows the phenotypic characteristics of yeast isolated from Kombucha tea broth and cellulosic pellicle with four isolates belonging to group II and 34 isolates to group I. Group II yeast isolates were presumed to be the dominant yeast during Kombucha fermentation. Group I isolates were not able to grow at 30 ℃ and 37 ℃ while group II isolates exhibited good growth at 30 ºC and weak growth at 37 °C. Both group I and group II isolates were able to grow on agar containing 0.01% cycloheximide. At higher cycloheximide concentrations, group I isolates showed weak growth. Group I isolates had better growth on agar containing 1% glacial acetic acid than group II. The two groups of yeast isolates were able to produce acid from D-glucose and sucrose, but only group II isolates produced acid from trehalose. These biochemical tests enable differentiation of the yeast isolates into further groups [2]. However, it is difficult to identify yeast isolates to species or genus level based only on biochemical and physiological tests. Therefore, molecular methods were conducted for further identification.
3.5. Genetic Identification of Representative AAB and Yeast Isolates
Six representative AAB isolates from each group were subjected to sequence analysis of the full-length 16S rRNA gene. Similarly, three representative yeast isolates were subjected to ITS sequencing after biochemical testing. All six AAB isolates belonged to Komagataeibacter rhaeticus, showing 99.92% sequence similarity with the reference strain (Table 5). Only one nucleotide (nt) mismatch was found within the 1287 nt aligned sequence region. Therefore, in this study the dominant species present in both tea broth and SCOBY was identified as K. rhaeticus.
All 28 isolates obtained from different fermentation times in this study were isolated from New Zealand SCOBY in 2021 and there were minor differences in phenotypic characteristics compared with K. rhzeticus DST GL02T [64]. This cellulose-producing strain was reported in Italian apple fruit and its basonym was Gluconacetobacter rhaeticus [64]. It was then re-classified into the genus Komagataibacter in 2012 [65]. Strains AF-1, P1463 and K3 were isolated from Kombucha on solid Hestrin-Schramm medium [43,57,66]. For example, Groups I, III, IV were not able to oxidise ethanol to acetic acid while reference strains can. Additionally, all six groups from our study exhibited different alcoholic tolerance in contrast to the reference strain [64,65]. Although these six group isolates belong to the same species, they were different strains and exhibited different biochemical profiles. The presence of K. rhaeticus contributes to the development of the cellulosic structure of Kombucha as shown by the results from the cellulose formation test (Table 3).
The demand for bacterial cellulose (BC) has been rapidly increasing for applications in food, biomedicine and cosmetics [43,57]. However, low yield and high operational costs limit the production of BC [57]. Extensive research has been conducted to overcome these difficulties including isolating more productive strains, using lower cost carbon substrates, and optimising fermentation conditions [67]. Various strains from the genus Komagataibacter have been reported to be potential producers of BC including K. xylinus, K. saccharivorans, K. hansenii and K. rhaeticus [67,68,69]. K. rhaeticus is a promising cellulose producing resource as it may have similar physicochemical characteristics to the BC produced by K. xylinus but with a higher yield under similar cultivation conditions [66]. Therefore, more research should be done to optimise the yield of BC synthesised by K. rhaeticus isolated from this study and their impact on Kombucha fermentation.
Based on the ITS sequence analysis, group I yeast isolates belonged to Zygosaccharomyces lentus and showed 99.84% sequence identity with the type strain CBS 8574. Only one mismatch was found with an ITS region of 634 nt in length. Group I isolates were obtained from both the cellulosic pellicle and Kombucha tea broth at each sampling point during fermentation (Figure 5). Z. lentus was firstly isolated from spoiled whole-orange drink and tentatively identified as Zygosaccharomyces bailli based on its carbohydrate assimilation profiles analysed by API ID 32C system [70]. This species has been isolated from Kombucha obtained in Germany and Ireland [71,72]. Strains of Z. lentus were distinguished from other strains of Zygosaccharomyces by their inability to grow above 25 ℃ but they could grow at low temperatures (4 ℃) and low pH [73]. The inability of Z. lentus to grow under aerobic conditions was due to their sensitivity to oxygen [70]. Similarly, our yeast strains could grow at 25 ℃ but not 30 ℃ (Table 4). The higher cell counts of Z. lentus may be due to the favourable fermentation conditions used in this study (22 ℃ and low pH) under static conditions. As fermentation temperature can affect the balance of microorganisms in Kombucha [74], fermentation above 25 ℃ with the SCOBY used in this study is not recommended as the dominant yeast species Z. lentus is not able to grow at that temperature. Strains isolated in this study were most similar in phenotypic characteristics to reference strains IGC and 2406, especially their ability to grow in medium containing 1% (w/v) acetic acid at low pH (pH 2 and 3) [73].
Group II yeast isolates were present at the beginning (days 3, 5 and 7) of the fermentation in the Kombucha tea broth only (Figure 5). Previous research also found that yeast was more diverse in the broth around day 7 of fermentation [75]. Although the culture-based methods used in this study may provide a good understanding of the physiological characteristics of the isolates, they are only effective in identifying the culturable microorganisms [76]. There may be unculturable yeast present in the SCOBY that cannot be detected using culture-based methods.
Group II yeast belonged to Debaryomyces prosopidis with 100% identity to the type strain D. prosopidis JCM 9913. This species was considered a minor microbe from the Kombucha yeast community in this study due its low proportion (10.3%). The phenotypic characteristics of the four strains isolated from this study agreed with the reference strains. The presence of D. prosopidis contributes positively to the texture and flavour characteristics of fermented meat products possibly due to their high lipolytic activity and production of hexanal [77]. The presence of this species in Kombucha may therefore contribute to its sensory profile. Recently, this species has been used to produce arabitol, a potential polyols sweetener- from glycerol [78]. As glycerol is an intermediate metabolite during Kombucha fermentation, the reaction between glycerol and D. prosopidis to produce arabitol may impact the sensory characteristics of Kombucha to some extent. Z. lentus and D. prosopidis are rarely isolated from Kombucha samples and therefore there is scanty information about their role in the fermentation. Hence, more research should be conducted to investigate the interactions between the two yeast species found in this SCOBY and their impact on Kombucha fermentation and metabolites.
4. Conclusions
The results of this study identified the microbial composition of a Kombucha SCOBY produced in New Zealand. All the AAB strains isolated from black tea Kombucha broth and pellicle during fermentation were identified as Komagataeibacter rhaeticus, which is known for cellulose production. The yeast species belonged to Zygosacharomyces lentus and Debaryomyces prosopidis, with the latter being dominant in the Kombucha tea broth and pellicle. This study contributes important information on the microbiological composition of the cellulosic pellicle and physico-chemical characteristics of Kombucha. Based on the findings of this study, it is recommended to ferment Kombucha using the SCOBY from this study at <25 ℃, as the dominant yeast species may be compromised. Improved knowledge of the microorganisms involved in Kombucha fermentation can help manufacturers to better monitor the progress of fermentation and aid in industrial scale-up to produce consistent high-quality products.
Author Contributions
Boying Wang: Conceptualisation, investigation, Data curation, writing, original draft, editing, writing, -review & editing. Kay Rutherfurd-Markwick: Conceptualization, writing-review & editing. Naran Naren: investigation. Xue-Xian Zhang: Conceptualization, writing-review & editing. Anthony N. Mutukumira: Funding acquisition, Project leader and overall management of the study, Conceptualization, writing-review & editing.
Funding
This research received funding from Massey University through College of Sciences REaDI Funding #SFA009.
Data Availability Statement
Not applicable.
Acknowledgments
This research was supported by Massey University through a scholarship for the first author. Equipment and other resources were provided by the School of Food and Advanced Technology of Massey University, Auckland, New Zealand. The authors thanks Rachel Liu for technical assistance in the microbiological analysis and Nuri Begum for physicochemical analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dufresne, C.; Farnworth, E. Tea, Kombucha, and health: A review. Food Res. Int. 2000, 33, 409–421. [Google Scholar] [CrossRef]
- Wang, B.; Rutherfurd-Markwick, K.; Zhang, X.-X.; Mutukumira, A.N. Isolation and characterisation of dominant acetic acid bacteria and yeast isolated from Kombucha samples at point of sale in New Zealand. Curr. Res. Food Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Neffe-Skocińska, K.; Sionek, B.; Ścibisz, I.; Kołożyn-Krajewska, D. Acid Contents and the Effect of Fermentation Condition of Kombucha Tea Beverages on Physicochemical, Microbiological and Sensory Properties. CyTA-J. Food 2017, 15, 601–607. [Google Scholar] [CrossRef]
- Wang, B.; Rutherfurd-Markwick, K.; Zhang, X.-X.; Mutukumira, A.N. Kombucha: Production and Microbiological Research. Foods 2022, 11, 3456. [Google Scholar] [CrossRef] [PubMed]
- Aung, T.; Eun, J.-B. Impact of time and temperature on the physicochemical, microbiological, and nutraceutical properties of laver kombucha (Porphyra dentata) during fermentation. LWT 2022, 154, 112643. [Google Scholar] [CrossRef]
- Laureys, D.; Britton, S.J.; De Clippeleer, J. Kombucha tea fermentation: A review. J. Am. Soc. Brew. Chem. 2020, 78, 165–174. [Google Scholar] [CrossRef]
- Bishop, P.; Pitts, E.R.; Budner, D.; Thompson-Witrick, K.A. Kombucha: Biochemical and microbiological impacts on the chemical and flavor profile. Food Chem. Adv. 2022, 100025. [Google Scholar] [CrossRef]
- Nyiew, K.Y.; Kwong, P.J.; Yow, Y.Y. An overview of antimicrobial properties of kombucha. Compre. Rev. Food Sci. Food Saf. 2022, 21, 1024–1053. [Google Scholar] [CrossRef]
- Emiljanowicz, K.E.; Malinowska-Pańczyk, E. Kombucha from alternative raw materials–The review. Crit. Rev. Food Sci. Nutri. 2020, 60, 3185–3194. [Google Scholar] [CrossRef]
- Velićanski, A.; Cvetković, D.; Markov, S. Characteristics of Kombucha fermentation on medicinal herbs from Lamiaceae family. Romanian Biotechnol. Lett. 2013, 18, 8034–8042. [Google Scholar]
- Dutta, H.; Paul, S.K. Kombucha drink: Production, quality, and safety aspects. In Product. Manag. Beverages; Elsevier: 2019; pp. 259–288.
- Coelho, R.M.D.; de Almeida, A.L.; do Amaral, R.Q.G.; da Mota, R.N.; de Sousa, P.H.M. Kombucha. Int.l J.Gastron. Food Sci. 2020, 22, 100272. [Google Scholar] [CrossRef]
- Savary, O.; Mounier, J.; Thierry, A.; Poirier, E.; Jourdren, J.; Maillard, M.-B.; Penland, M.; Decamps, C.; Coton, E.; Coton, M. Tailor-made microbial consortium for Kombucha fermentation: Microbiota-induced biochemical changes and biofilm formation. Food Res. Int. 2021, 147, 110549. [Google Scholar] [CrossRef]
- Greenwalt, C.J.; Steinkraus, K.H.; Ledford, R.A. Kombucha, the fermented tea: Microbiology, composition, and claimed health effects. J. Food Prot. 2000, 63, 976–981. [Google Scholar] [CrossRef]
- Jayabalan, R.; Malbasa, R.V.; Loncar, E.S.; Vitas, J.S.; Sathishkumar, M. A Review on Kombucha Tea-Microbiology, Composition, Fermentation, Beneficial Effects, Toxicity, and Tea Fungus. Compr Rev Food Sci Food Saf 2014, 13, 538–550. [Google Scholar] [CrossRef]
- Jayabalan, R.; Chen, P.-N.; Hsieh, Y.-S.; Prabhakaran, K.; Pitchai, P.; Marimuthu, S.; Thangaraj, P.; Swaminathan, K.; Yun, S.E. Effect of solvent fractions of kombucha tea on viability and invasiveness of cancer cells—Characterization of dimethyl 2-(2-hydroxy-2-methoxypropylidine) malonate and vitexin. Indian J. Biotechnol. 2011, 10, 62–75. [Google Scholar]
- Mukadam, T.A.; Punjabi, K.; Deshpande, S.D.; Vaidya, S.P.; Chowdhary, A.S. Isolation and characterization of bacteria and yeast from Kombucha tea. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 32–41. [Google Scholar] [CrossRef]
- Nguyen, N.K.; Dong, N.T.; Nguyen, H.T.; Le, P.H. Lactic acid bacteria: Promising supplements for enhancing the biological activities of kombucha. Springerplus 2015, 4, 91. [Google Scholar] [CrossRef]
- Pei, J.; Jin, W.; Abd El-Aty, A.; Baranenko, D.A.; Gou, X.; Zhang, H.; Geng, J.; Jiang, L.; Chen, D.; Yue, T. Isolation, purification, and structural identification of a new bacteriocin made by Lactobacillus plantarum found in conventional kombucha. Food Control 2020, 110, 106923. [Google Scholar] [CrossRef]
- Diguta, C.F.; Nitoi, G.D.; Matei, F.; Luta, G.; Cornea, C.P. The Biotechnological Potential of Pediococcus spp. Isolated from Kombucha Microbial Consortium. Foods 2020, 9. [Google Scholar] [CrossRef]
- Yang, J.; Lagishetty, V.; Kurnia, P.; Henning, S.M.; Ahdoot, A.I.; Jacobs, J.P. Microbial and chemical profiles of commercial Kombucha products. Nutrients 2022, 14, 670. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, M.; Justine, S.; Filofteia, D.C.; Petruta, C.C.; Gabriela, L.; Roxana, U.E.; Florentina, M.; Camelia Filofteia, D.; Călina Petruța, C.; Gabriela, L. Lactic acid bacteria strains isolated from Kombucha with potential probiotic effect. Roman. Biotechnol. Lett. 2018, 23, 13592–13598. [Google Scholar]
- Coton, M.; Pawtowski, A.; Taminiau, B.; Burgaud, G.; Deniel, F.; Coulloumme-Labarthe, L.; Fall, A.; Daube, G.; Coton, E. Unraveling microbial ecology of industrial-scale Kombucha fermentations by metabarcoding and culture-based methods. FEMS Microbiol Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Mayser, P.; Fromme, S.; Leitzmann, C.; Grunder, K. The yeast spectrum of the 'tea fungus Kombucha'. Mycoses 1995, 38, 289–295. [Google Scholar] [CrossRef]
- Ramadani, A.; Abulreesh, H. Isolation and Identification of Yeast Flora in Local Kombucha Sample: Al Nabtah. Umm Al Qura Univ. J App. Sci. 2010, 2, 42–51. [Google Scholar]
- Jayabalan, R.; Malbaśa, R.V.; Sathishkumar, M. Kombucha Tea: Metabolites. Fungal Metabolites 2017, 965–978. [Google Scholar]
- Teoh, A.L.; Heard, G.; Cox, J. Yeast ecology of Kombucha fermentation. Int. J. Food Microbiol. 2004, 95, 119–126. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; Bouajila, J.; Pace, M.; Leech, J.; Cotter, P.D.; Souchard, J.P.; Taillandier, P.; Beaufort, S. Metabolome-microbiome signatures in the fermented beverage, Kombucha. Int J Food Microbiol. 2020, 333, 108778. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.P.; Taillandier, P. Understanding Kombucha Tea Fermentation: A Review. J. Food Sci. 2018, 83, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Czaja, W.K.; Young, D.J.; Kawecki, M.; Brown, R.M. The future prospects of microbial cellulose in biomedical applications. Biomacromolecules 2007, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, Í.A.C.L.d.; Rolim, V.A.d.O.; Gaspar, R.P.L.; Rossini, D.Q.; de Souza, R.; Bogsan, C.S.B. The Technological Perspectives of Kombucha and Its Implications for Production. Fermentation 2022, 8, 185. [Google Scholar] [CrossRef]
- Waisundara, V.Y. Usage of Kombucha ‘Tea Fungus’ for Enhancement of Functional Properties of Herbal Beverages. In Frontiers New Trd. Sci.Fermented Food Beverages; IntechOpen: 2018.
- Amarasinghe, H.; Weerakkody, N.S.; Waisundara, V.Y. Evaluation of physicochemical properties and antioxidant activities of kombucha "Tea Fungus" during extended periods of fermentation. Food Sci Nutr 2018, 6, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Aung, T.; Eun, J.B. Production and characterization of a novel beverage from laver (Porphyra dentata) through fermentation with kombucha consortium. Food Chem. 2021, 350, 129274. [Google Scholar] [CrossRef] [PubMed]
- Avcioglu, N.H.; Birben, M.; Bilkay, I.S. Optimization and physicochemical characterization of enhanced microbial cellulose production with a new Kombucha consortium. Process Biochem. 2021, 108, 60–68. [Google Scholar] [CrossRef]
- Du Toit, W.J.; Lambrechts, M.G. The enumeration and identification of acetic acid bacteria from South African red wine fermentations. Int. J Food Microbiol. 2002, 74, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Reiner, K. Catalase Test Protocol. Am. Soc. Micorbiol. 2010, 1–6. [Google Scholar]
- Shields, P.; Cathcart, L. Oxidase Test Protocol. Am. Soc., Microbiol, 2010; 1–10. [Google Scholar]
- Gomes, R.J.; Borges, M.F.; Rosa, M.F.; Castro-Gomez, R.J.H.; Spinosa, W.A. Acetic Acid Bacteria in the Food Industry: Systematics, Characteristics and Applications. Food Technol. Biotechnol. 2018, 56, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Gullo, M.; Giudici, P. Acetic acid bacteria in traditional balsamic vinegar: Phenotypic traits relevant for starter cultures selection. Int. J. Food Microbiol. 2008, 125, 46–53. [Google Scholar] [CrossRef]
- Arifuzzaman, M.; Hasan, M.Z.; Rahman, S.B.; Pramanik, M.K. Isolation and Characterization of Acetobacter and Gluconobacter Spp. from Sugarcane and Rotten Fruits. Res. Revs. Biosciences 2014, 8, 359–365. [Google Scholar]
- Lavasani, P.S.; Motevaseli, E.; Shirzad, M.; Modarressi, M.H. Isolation and identification of Komagataeibacter xylinus from Iranian traditional vinegars and molecular analyses. Iran. J. Microbiol. 2017, 9, 338–347. [Google Scholar]
- Semjonovs, P.; Ruklisha, M.; Paegle, L.; Saka, M.; Treimane, R.; Skute, M.; Rozenberga, L.; Vikele, L.; Sabovics, M.; Cleenwerck, I. Cellulose synthesis by Komagataeibacter rhaeticus strain P 1463 isolated from Kombucha. Appl. Microbiol. Biotechnol. 2017, 101, 1003–1012. [Google Scholar] [CrossRef]
- Asai, T.; Iizuka, H.; Komagata, K. The flagellation and taxonomy of genera Gluconobacter and Acetobacter with reference to the existence of intermediate strains. J. Gen. Appl. Microbiol. 1964, 10, 95–126. [Google Scholar] [CrossRef]
- Swings, J. Phenotypic Identification of Acetic Acid Bacteria. Identification Methods in Applied and Environmental Microbiology (The Society for Applied Bacteriology Technical Series No. 29). 1992, 103-110.
- Matthews, C. On the Staining of Yeast Cells by Methylene Blue, etc. J. Inst. Brewing 1914, 20, 488–496. [Google Scholar] [CrossRef]
- Painting, K.; Kirsop, B. A quick method for estimating the percentage of viable cells in a yeast population, using methylene blue staining. World J. Microbiol. Biotechnol. 1990, 6, 346–347. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Methods for isolation, phenotypic characterization and maintenance of yeasts. In The Yeasts; Elsevier: Amsterdam, The Netherlands, 2011; pp. 87–110. [Google Scholar]
- Kurtzman, C.P.; Boekhout, T.; Robert, V.; Fell, J.W.; Deak, T. Methods to identify yeasts. Yeasts food: Benefi Detrimental Asp. 2003, 69-121. [CrossRef]
- Yuan, Y.; Feng, F.; Chen, L.; Yao, Q.; Chen, K. Directional Isolation of ethanol-tolerant Acetic Acid Bacteria from Industrial Fermented Vinegar. Eur. Food Res. and Technol. 2013, 236, 573–578. [Google Scholar] [CrossRef]
- Matei, B.; Diguță, C.F.; Popa, O.; Cornea, C.P.; Matei, F. Molecular identification of yeast isolated from different kombucha sources. The Annals of the University Dunarea de Jos of Galati. Fascicle VI-Food Technol. 2018, 42, 17–25. [Google Scholar]
- Reller, L.B.; Weinstein, M.P.; Petti, C.A. Detection and identification of microorganisms by gene amplification and sequencing. Clinic. Infectious Dis. 2007, 44, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S rRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Systematic Evolut. Microbiol. 2017, 67, 1613. [Google Scholar] [CrossRef]
- Chu, S.-C.; Chen, C. Effects of origins and fermentation time on the antioxidant activities of kombucha. Food Chem. 2006, 98, 502–507. [Google Scholar] [CrossRef]
- Nummer, B.A. Kombucha brewing under the Food and Drug Administration model Food Code: Risk analysis and processing guidance. J. Environ. Health 2013, 76, 8–11. [Google Scholar]
- de Miranda, J.F.; Ruiz, L.F.; Silva, C.B.; Uekane, T.M.; Silva, K.A.; Gonzalez, A.G.M.; Fernandes, F.F.; Lima, A.R. Kombucha: A review of substrates, regulations, composition, and biological properties. J. Food Sci. 2022, 87, 503–527. [Google Scholar] [CrossRef]
- Jacek, P.; da Silva, F.A.S.; Dourado, F.; Bielecki, S.; Gama, M. Optimization and characterization of bacterial nanocellulose produced by Komagataeibacter rhaeticus K3. Carbohydr. Polym. Technol. Appl. 2021, 2, 100022. [Google Scholar] [CrossRef]
- Wu, J.-M.; Liu, R.-H. Cost-effective production of bacterial cellulose in static cultures using distillery wastewater. J. Biosci. Bioengi. 2013, 115, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Lu, S.; Yang, Y. Production of bacterial cellulose from byproduct of citrus juice processing (citrus pulp) by Gluconacetobacter hansenii. Cellulose 2018, 25, 6977–6988. [Google Scholar] [CrossRef]
- Yamada, Y.; Yukphan, P. Genera and Species in Acetic Acid Bacteria. Int. J. Food Microbiol. 2008, 125, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Mamlouk, D.; Gullo, M. Acetic Acid Bacteria: Physiology and Carbon Sources Oxidation. Indian J. Microbiol. 2013, 53, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Raspor, P.; Goranovic, D. Biotechnological Applications of Acetic Acid Bacteria. Crit. Rev. Biotechnol. 2008, 28, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Drysdale, G.; Fleet, G. Acetic Acid Bacteria in Winemaking: A Review. Am. J. Enology and Viticulture 1988, 39, 143–154. [Google Scholar] [CrossRef]
- Dellaglio, F.; Cleenwerck, I.; Felis, G.E.; Engelbeen, K.; Janssens, D.; Marzotto, M. Description of Gluconacetobacter swingsii sp. nov. and Gluconacetobacter rhaeticus sp. nov., isolated from Italian apple fruit. Int. J. Systematic Evolut. Microbiol. 2005, 55, 2365–2370. [Google Scholar] [CrossRef]
- Yamada, Y.; Yukphan, P.; Vu, H.T.L.; Muramatsu, Y.; Ochaikul, D.; Tanasupawat, S.; Nakagawa, Y. Description of Komagataeibacter gen. nov., with proposals of new combinations (Acetobacteraceae). J. Gen. Applied Microbiol. 2012, 58, 397–404. [Google Scholar] [CrossRef]
- Machado, R.T.; Gutierrez, J.; Tercjak, A.; Trovatti, E.; Uahib, F.G.; de Padua Moreno, G.; Nascimento, A.P.; Berreta, A.A.; Ribeiro, S.J.; Barud, H.S. Komagataeibacter rhaeticus as an alternative bacteria for cellulose production. Carbohydr. Polym. 2016, 152, 841–849. [Google Scholar] [CrossRef]
- Thorat, M.N.; Dastager, S.G. High yield production of cellulose by a Komagataeibacter rhaeticus PG2 strain isolated from pomegranate as a new host. RSC Advances 2018, 8, 29797–29805. [Google Scholar] [CrossRef]
- Gupte, Y.; Kulkarni, A.; Raut, B.; Sarkar, P.; Choudhury, R.; Chawande, A.; Kumar, G.R.K.; Bhadra, B.; Satapathy, A.; Das, G.; et al. Characterization of nanocellulose production by strains of Komagataeibacter sp. isolated from organic waste and Kombucha. Carbohydr. Polym. 2021, 266, 118176. [Google Scholar] [CrossRef]
- Gopu, G.; Govindan, S. Production of bacterial cellulose from Komagataeibacter saccharivorans strain BC1 isolated from rotten green grapes. Preparative Biochem. Biotechnol. 2018, 48, 842–852. [Google Scholar] [CrossRef]
- Steels, H.; Bond, C.J.; Collins, M.D.; Roberts, I.N.; Stratford, M.; James, S.A. Zygosaccharomyces lentus sp. nov., a new member of the yeast genus Zygosaccharomyces Barker. Int. J.Systematic Evolut. Microbiol. 1999, 49, 319–327. [Google Scholar] [CrossRef]
- Hopfe, S.; Flemming, K.; Lehmann, F.; Mockel, R.; Kutschke, S.; Pollmann, K. Leaching of rare earth elements from fluorescent powder using the tea fungus Kombucha. Waste Manag 2017, 62, 211–221. [Google Scholar] [CrossRef]
- Marsh, A.J.; O'Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-based analysis of the bacterial and fungal compositions of multiple kombucha (tea fungus) samples. Food Microbiol. 2014, 38, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Steels, H.; James, S.; Roberts, I.; Stratford, M. Zygosaccharomyces lentus: A significant new osmophilic, preservative-resistant spoilage yeast, capable of growth at low temperature. J.Appl.Microbiol. 1999, 87, 520–527. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Troise, A.D.; Vitaglione, P.; Ercolini, D. Different temperatures select distinctive acetic acid bacteria species and promotes organic acids production during Kombucha tea fermentation. Food Microbiol. 2018, 73, 11–16. [Google Scholar] [CrossRef]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha tea fermentation: Microbial and biochemical dynamics. Int. J. Food Microbiol. 2016, 220, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Moyer, C.L.; Dobbs, F.C.; Karl, D.M. Estimation of diversity and community structure through restriction fragment length polymorphism distribution analysis of bacterial 16S rRNA genes from a microbial mat at an active, hydrothermal vent system, Loihi Seamount, Hawaii. Appl. Environ. Microbiol. 1994, 60, 871–879. [Google Scholar] [CrossRef]
- Song, Z.; Cao, Y.; Zhang, Y.; Zhang, Z.; Shi, X.; Zhang, W.; Wen, P. Effects of storage methods on the microbial community and quality of Sichuan smoked bacon. LWT 2022, 158, 113115. [Google Scholar] [CrossRef]
- Filippousi, R.; Tsouko, E.; Mordini, K.; Ladakis, D.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Sustainable arabitol production by a newly isolated Debaryomyces prosopidis strain cultivated on biodiesel-derived glycerol. Carbon Resour. Convers. 2022, 5, 92–99. [Google Scholar] [CrossRef]
Figure 1.
Microbial analysis of Kombucha tea broth and cellulosic pellicle. Adapted from Ref. [2].
Figure 1.
Microbial analysis of Kombucha tea broth and cellulosic pellicle. Adapted from Ref. [2].
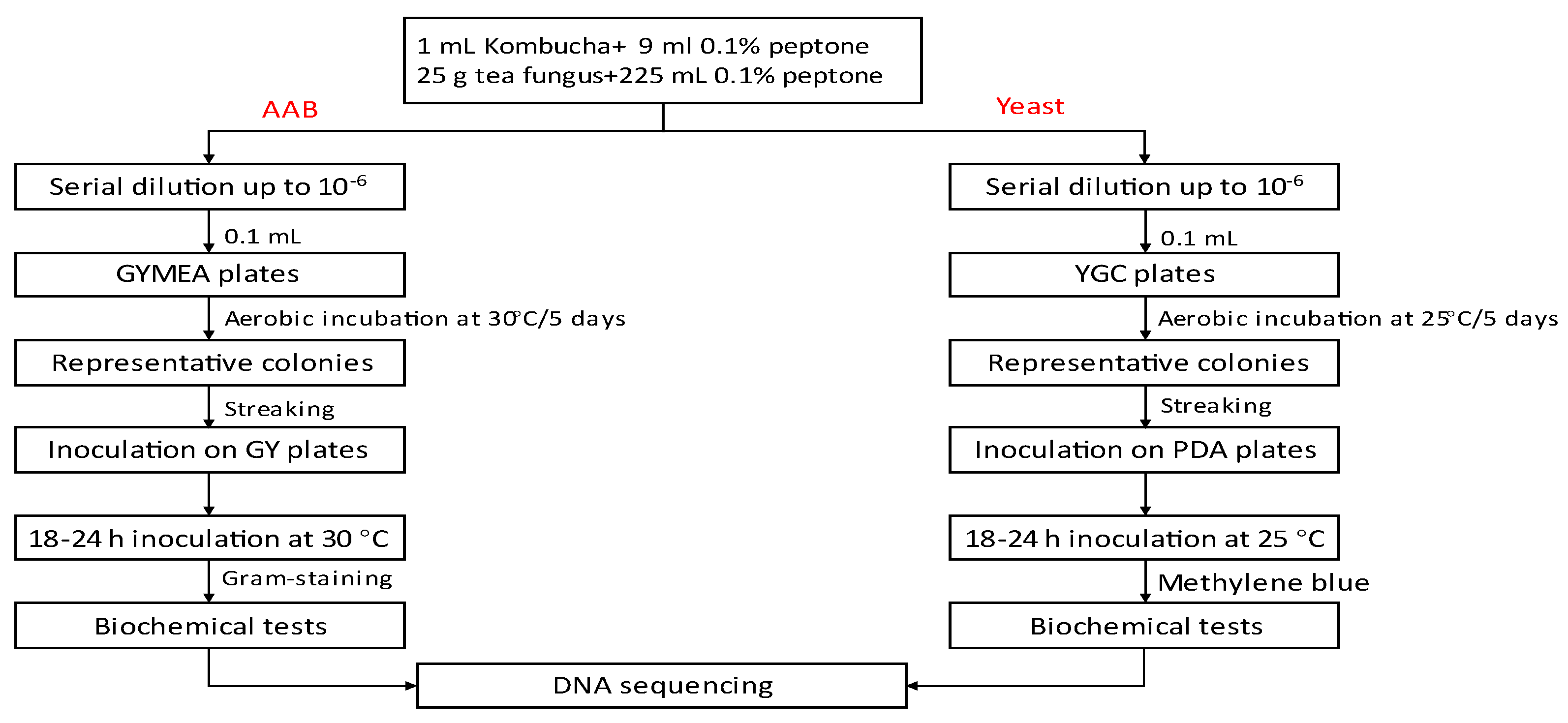
Figure 2.
pH and titratable acidity of Kombucha tea broth during fermentation for 14 days at 22℃ (n=3).
Figure 2.
pH and titratable acidity of Kombucha tea broth during fermentation for 14 days at 22℃ (n=3).
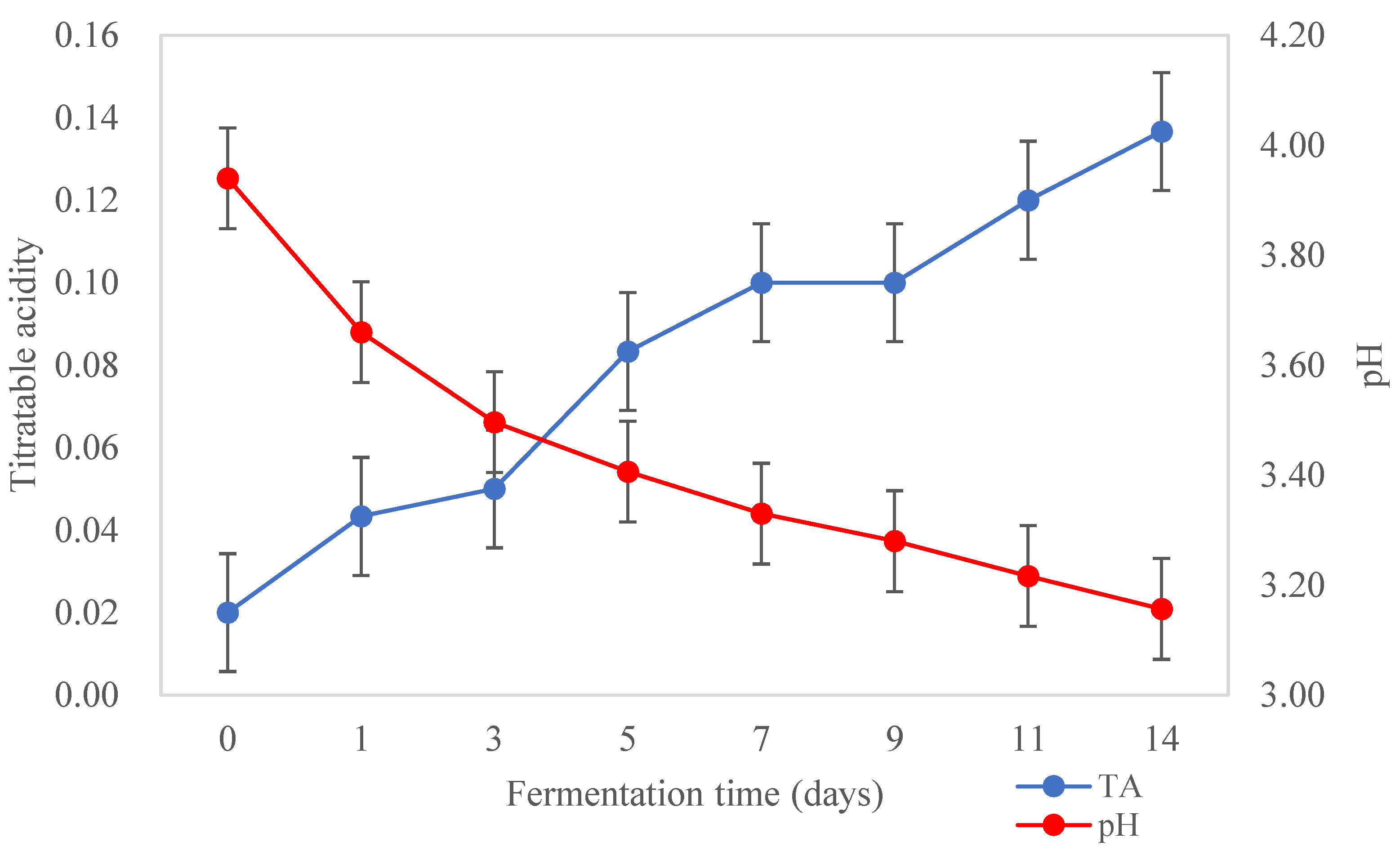
Figure 3.
Total soluble solid (TSS) of Kombucha tea broth during fermentation for 14 days at 22 ℃ (n=3).
Figure 3.
Total soluble solid (TSS) of Kombucha tea broth during fermentation for 14 days at 22 ℃ (n=3).
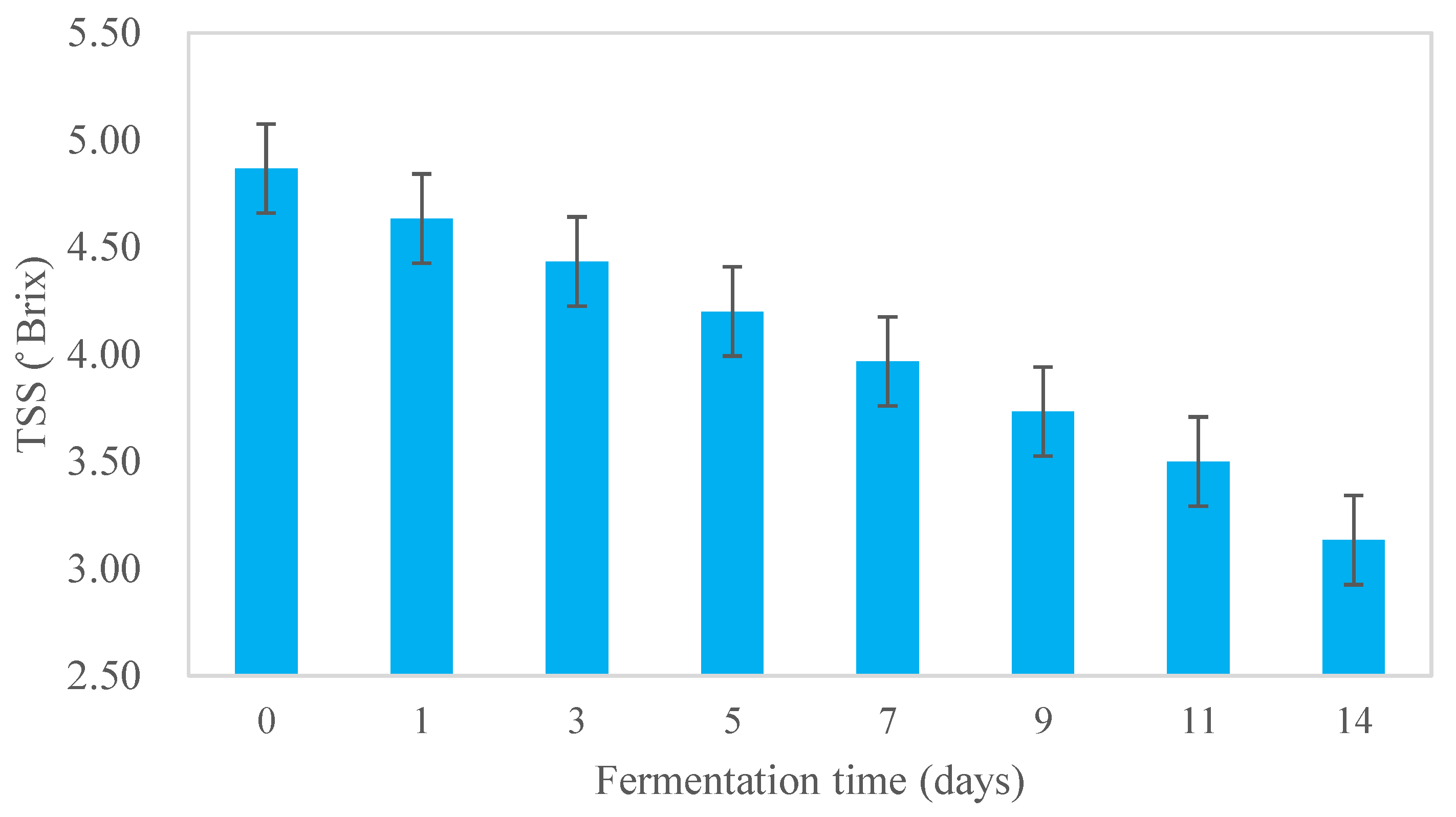
Figure 4.
AAB and yeast count in Kombucha tea broth during fermentation.
Figure 5.
Distribution of yeast isolates in Kombucha tea broth during fermentation for 14 days.
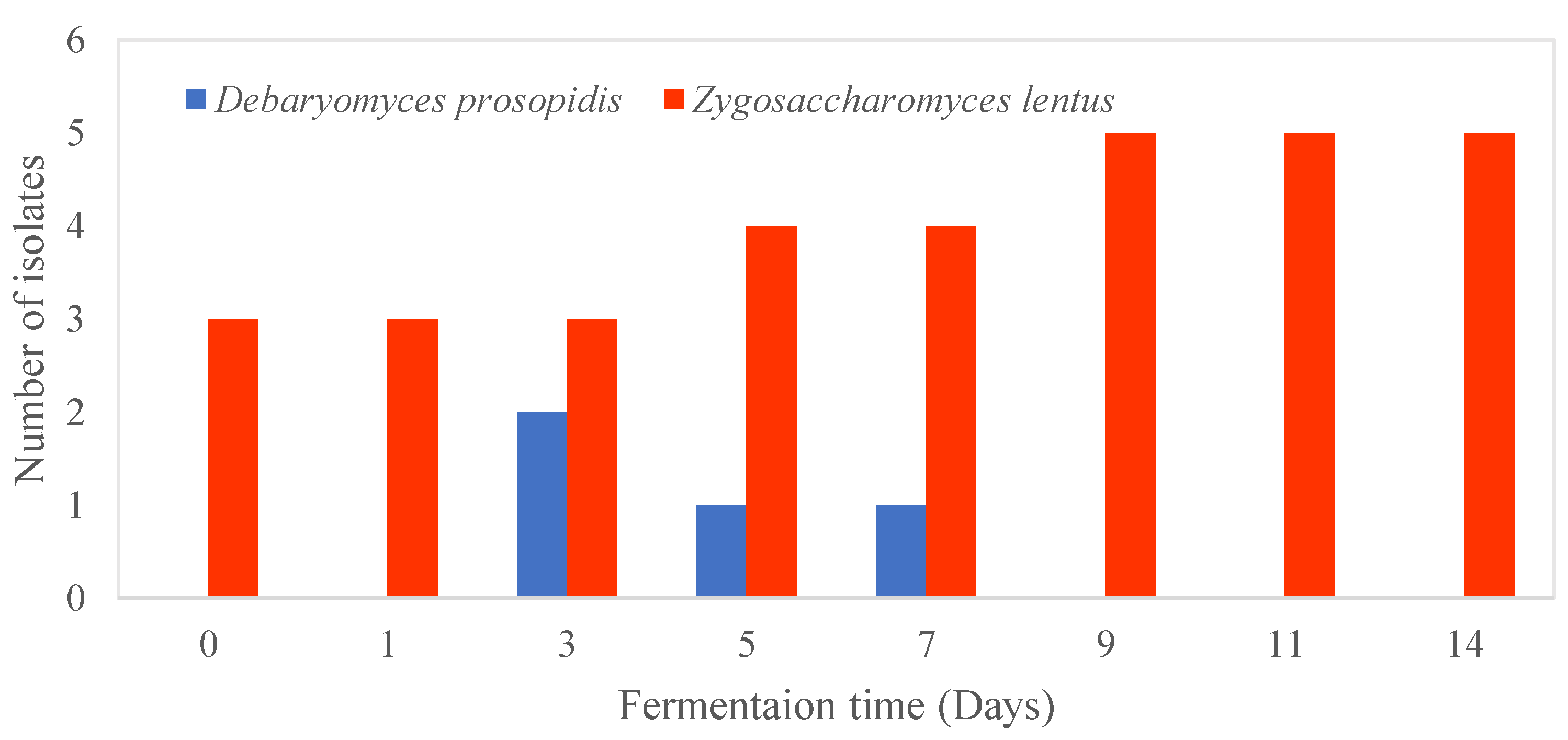

Table 1.
Wet yield, water holding capacity (WHC) moisture content and water activity of cellulosic pellicle.
Table 1.
Wet yield, water holding capacity (WHC) moisture content and water activity of cellulosic pellicle.
| Characteristics | Cellulosic Pellicle |
|---|---|
| Wet yield (%) | 27.75±0.08 |
| WHC (g water/g dry pellicle) | 17.82±0.16 |
| Moisture content | 94.68±0.05 |
| Water activity at 21.45℃ | 0.996±0.002 |
Table 2.
Appearance of yeast colonies grown on YGC agar plates.
| Colony Group | Description of Appearance |
|---|---|
| Group I | Circular and slightly umbonate in the centre, white colour, smooth surface and entire margin |
| Group II | Circular, brownish cream colour, flat and smooth surface |
Table 3.
Phenotypic characteristics of AAB isolated from Kombucha broth and cellulosic pellicle.
| Tests | Group I | Group II | Group III | Group IV | Group V | Group VI |
|---|---|---|---|---|---|---|
| Oxidase | - | - | - | - | - | - |
| Catalase | + | + | + | + | + | + |
| Growth at different temperature | ||||||
| Growth at 25℃ | + | + | + | + | + | + |
| Growth at 30℃ | + | + | + | + | + | + |
| Growth at 37℃ | + | + | + | + | + | + |
| Growth at different pH | ||||||
| pH 2 | - | - | - | - | - | - |
| pH 3 | + | + | + | + | + | + |
| Cellulose formation | + | + | + | + | + | + |
| Growth without acetic acid | + | + | + | + | + | + |
| Oxidation of acetate | + | + | + | + | + | + |
| Oxidation of lactate | + | + | + | + | + | + |
| Alcoholic tolerance (v/v) | ||||||
| 2% | + | + | + | + | + | + |
| 4% | + | - | - | + | + | + |
| 6% | - | - | - | + | + | - |
| 8% | - | - | - | + | - | - |
| 10% | - | - | - | + | - | - |
| Acid produced from: | ||||||
| D-glucose | + | + | + | + | + | + |
| Sucrose | - | - | - | - | - | - |
| Lactose | - | - | - | - | - | - |
| Trehalose | - | - | - | - | - | - |
| Oxidation of ethanol | - | + | - | + | - | - |
| Oxidation of ethanol to water and CO2 | - | + | - | + | - | - |
| Ketogenesis of glycerol to DHA | + | + | + | + | + | + |
Note: (+) indicated positive reaction (-) indicated negative reaction. Experiments were replicated 3 times. Group 1 (n=4), Group 2 (n=8), Group 3(n=6), Group 4 (n=2), Group 5 (n=5), Group 6 (n=3).
Table 4.
Phenotypic characteristics of yeast isolated from Kombucha broth and cellulosic pellicles.
| Tests | Group I | Group II |
|---|---|---|
| Growth in broth at 37 ℃ | - | w |
| Growth in broth at 30 ℃ | - | + |
| Growth in broth at 25 ℃ | + | + |
| Growth in medium containing 0.01% cycloheximide | + | + |
| Growth in medium containing 0.1% cycloheximide | w | + |
| Growth in medium containing 1% glacial acetic acid | + | w |
| Growth in medium containing 5% glucose and 10% NaCl | - | w |
| Growth in broth at pH 2 | + | + |
| Growth in broth at pH 3 | + | + |
| Acid produced from: | ||
| D-glucose | + | + |
| Sucrose | + | + |
| Trehalose | - | + |
| Lactose | - | - |
Note: (+) indicated positive reaction (-) indicated negative reaction, (w) indicated weakly growth. Experiments were replicated 3 times. Group I (n=35) and Group II (n=4).
Table 5.
Genetic identification of representative AAB and yeast isolates.
| Type Strain Codes | Top Hit Taxon | Top Hit Strains | GenBank Accession | % Similarity | Variation |
|---|---|---|---|---|---|
| AAB | |||||
| TFT1AAB26 | Komagataibacter rhaeticus | DST GL02 | AY180961 | 99.92 | 1/1292 |
| Yeast | |||||
| GI: D3T3Y9 | Zygosaccharomyces lentus | CBS 8574 | NR_156001.1 | 99.84 | 1/634 |
| GII: TFT1Y39 | Debaryomyces prosopidis | JCM 9913 | NR_077067.1 | 100.00 | 0/600 |
Note: Other AAB strain codes associated with identification of D5T1AAB1, D7T2AAB10, D11T1AAB16, D11T2AAB17, TFT3AAB29. GI: yeast group I, GII: yeast group II.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



