
Submitted:
06 May 2023
Posted:
08 May 2023
You are already at the latest version
Abstract
The aim of this pilot study was to investigate whether single nucleotide polymorphisms (SNP) in the gene encoding the catalytic subunit of glutamate cysteine ligase (GCLC) are associated with the risk and clinical features of psoriasis. A total of 944 unrelated individuals, including 474 patients with a diagnosis of psoriasis and 470 healthy controls, were recruited for the study. Six common SNPs in the GCLC gene were genotyped using the MassArray-4 system. Polymorphisms rs648595 (OR=0.56 95%CI 0.35-0.90, Pperm=0.017) and rs2397147 (OR=0.54 95%CI 0.30-0.98, Pperm=0.05) were associated with susceptibility to psoriasis in males. In the male group, diplotype rs2397147-C/C×rs17883901-G/G was associated with decreased risk of psoriasis (FDR-adjusted P=0.014), whereas diplotype rs6933870-G/G×rs17883901-G/G (FDR-adjusted P=0.045) showed association with increased disease risk in females. Joint effects of SNPs with tobacco smoking (rs648595 and rs17883901) and alcohol abuse (rs648595 and rs542914) on the risk of psoriasis were observed (Pperm≤0.05). Furthermore, we found multiple sex-independent associations between GCLC gene polymorphisms and various clinical features such as earlier disease onset, the psoriatic triad, and specific localizations of skin lesions. The present study is the first to show that polymorphisms of the GCLC gene are significantly associated with the risk of psoriasis and related to its clinical features.
Keywords:
Psoriasis
; genetic susceptibility
; oxidative stress
; glutathione
; glutamate cysteine ligase
; GCLC
; single nucleotide polymorphism
; cigarette smoking
; alcohol abuse
; gene–environment interactions
1. Introduction
Psoriasis is a chronic immune-inflammatory-mediated dermatosis characterized by thickened, scaly erythema or plaques [1,2]. Psoriasis is recognized by the World Health Organization as a serious non-communicable disease [3]. Clinical variants of the disease include psoriasis vulgaris, arthritis, pustular, and erythrodermic types; however, psoriasis vulgaris is the most common form, accounting for about 90% of cases and affecting 3% of Caucasians [4]. A study by Kubanov and co-workers demonstrated a substantial disease burden on psoriasis patients in Russia [5].
The etiology and pathogenesis of psoriasis remain mysteries, making the disease’s management more challenging [6]. Psoriasis is characterized by sustained inflammation, which results in uncontrolled keratinocyte proliferation and defective differentiation [7]. Psoriatic inflammation is caused and maintained by disruptions in the innate and adaptive cutaneous immune responses [6,8], which coexist with autoinflammatory perpetuation or T cell-driven autoimmune reactions [7]. The overlap of autoimmune and autoinflammatory mechanisms in the pathogenesis of psoriasis has led to the development of biological therapy for the disease. However, despite the fact that targeted therapies focusing on the inhibition of cytokines such as IL-23 and IL-17 showed high clinical efficacy, psoriasis remains so far not curable disease [7].
Psoriasis is known as a complex multifactorial disease whose development is determined by the interaction between genetic and environmental factors [9,10,11]. Linkage analysis, an effective method to identify the chromosomal location of disease genes, has discovered nine separate genomic regions known as Psoriasis Susceptibility regions (PSORS1-9), comprising many genetic variants, a part of which has been fine-mapped as disease-linked loci [11,12]. Progress in the development of high-throughput genotyping technologies enabled the implementation of genome-wide association studies (GWAS), a research approach where large case-control cohorts were genotyped for tens of thousands of single nucleotide polymorphisms (SNPs) across the genome [11]. According to the GWAS catalog (https://www.ebi.ac.uk/gwas/home, accessed on April 29, 2023), 57 GWASs have been conducted so far to unravel the genetic background of psoriasis in different populations around the world, and 946 SNPs have been identified as loci associated with disease susceptibility or severity and those influencing the efficacy of anti-psoriatic therapy. Nevertheless, despite considerable genetic research and achievements, the etiology of psoriasis and its primary molecular mechanisms remain elusive.
It has been argued that increased production of reactive oxygen species (ROS) and a decreased antioxidant defense leading to the activation of oxidative stress are involved in the pathogenesis of psoriasis and influence disease duration and severity [13,14,15,16]. Despite the fact that the important role of oxidative stress in the etiopathogenesis of psoriasis remains undisputable after decades of research, a limited number of studies have been undertaken so far to assess whether genetic variation of antioxidant defense enzymes contributes to psoriasis susceptibility. A larger portion of the studies looked for the link between psoriasis risk and genetic polymorphisms of glutathione-S-transferases [17,18,19,20], enzymes catalyzing the conjugation of reduced glutathione (GSH) to xenobiotic compounds for their detoxification.
Glutathione is a low-molecular-weight thiol, a tripeptide consisting of glutamate, cysteine, and glycine, which plays a major role in maintaining intracellular redox balance and antioxidant defense [21]. It is involved in many crucial biological functions, such as xenobiotic detoxification, maintaining mitochondrial function, modulation of cell proliferation, wound healing, and inhibition of apoptosis [21,22]. Furthermore, glutathione is utilized as a cofactor by glutathione peroxidases and glutathione S-transferases for glutathionylation of selected proteins and toxic substance conjugation. GSH is also required for the maturation of cytosolic iron-sulfur proteins, which are essential for cell viability and involved in the maintenance of DNA metabolism, genome integrity, protein translation, and other critical biological functions [22,23]. It is important to note that glutathione is involved in the skin metabolic clearance system [24], protects DNA and mitochondria from oxidative damage, and ensures the survival of keratinocytes in normal and wounded skin [25]. Glutathione deficiency is well known to be associated with increased susceptibility to oxidative stress, a pathological condition implicated in the pathogenesis of psoriasis [26], and therefore we can suggest that oxidative stress may be responsible for the modulation of inflammatory and autoimmune mechanisms underlying the diseases [27,28]. Despite the obvious importance of glutathione in skin metabolism, existing research data in psoriasis on the roles of genes encoding enzymes involved in glutathione metabolism, primarily glutamate cysteine ligase, an enzyme catalyzing the initial rate-limiting step of GSH biosynthesis [29], is surprisingly absent. We propose that genetic polymorphisms of glutamate cysteine ligase may explain inter-individual differences in glutathione biosynthesis and influence the risk of psoriasis, making SNPs attractive markers for testing disease susceptibility. Therefore, the purpose of our pilot study was to investigate whether common polymorphisms at the gene encoding the catalytic subunit of glutamate cysteine ligase (GCLC) are associated with the risk and clinical features of psoriasis.
2. Materials and Methods
2.1. Study Participants and Clinical Examination
Informed consent was signed by all subjects involved in this study. The protocol of the present study was approved by the Ethical Review Committee of Kursk State Medical University (protocol No. 8, 13.11.2017). A total of 944 unrelated individuals of European descent (predominantly Russians), including 474 patients with a diagnosis of psoriasis and 470 healthy controls, were used for this study. The enrollment of patients with psoriasis was done in Medvenka Central District Hospital (Kursk region), the Center for Medical Examinations and Prevention (Kursk), and Kursk Regional Multidisciplinary Clinical Hospital in a period between September 2018 and December 2021. The control group of subjects without chronic diseases was recruited from our previous studies [30,31,32]. The diagnosis of psoriasis was verified by qualified dermatologists based on the typical clinical picture of skin rashes and their localization [6]. The study included patients with classic plaque psoriasis, palmoplantar, seborrheic, and scalp psoriasis, von Zumbusch type of generalized pustular psoriasis, inverse psoriasis, guttate psoriasis, and erythrodermic psoriasis, as well as psoriasis comorbidities such as psoriatic arthritis and onychodystrophy [33]. The Psoriasis Area and Severity Index (PASI) was used for the clinical assessment of the severity of the course of psoriasis [34]. Enrolled patients did not suffer from chronic infectious diseases, including HIV and hepatitis, and did not have severe chronic conditions that manifested before psoriasis. Patients who were receiving biologic therapy at the time of the recruitment and pregnant women were not included in the study. Study participants completed a validated doctor-administered questionnaire [35] to assess risk factors for psoriasis, such as cigarette smoking [36] and alcohol consumption [37]. Information on smoking status (ever/never) was available from all psoriatic patients and healthy subjects. Data on alcohol intake were available from all patients with psoriasis and only 220 individuals from the control group. Alcohol intake habits were assessed through the number of drinks consumed per week, as described previously [38,39]. Briefly, according to the reported frequency of alcohol intake, study individuals were categorized into two groups: (1) subjects who consumed alcohol 1 to 2 days a month or less often and (2) those drinking alcohol 1 or more days a week. The second group was considered as alcohol abusers.
2.2. Selection of single nucleotide polymorphisms (SNPs)
GCLC is a catalytic subunit of glutamate-cysteine ligase and is the first rate-limiting enzyme of glutathione synthesis [29]. Six common (minor allele frequency ≥ 5%) SNPs such as rs524553, rs542914, rs648595, rs6933870, rs2397147 and rs17883901 of the GCLC gene were selected for the study according to the functional properties of the polymorphisms (the presence of eQTL, Expression Quantitative Trait Loci, in the skin from GTEx portal, https://gtexportal.org) and linkage disequilibrium (r2 ≥ 0.8) between them (HapMap data, European population). Candidate Gene SNP Selection (GenePipe) at the SNPinfo Web Server (https://snpinfo.niehs.nih.gov/snpinfo/selegene.html (accessed on April 25, 2021)) was used for SNP selection.
2.3. Genetic Analysis
Venous blood samples were collected from the cubital vein of study subjects into EDTA-coated tubes and immediately frozen and stored at −20°C until processed. Total DNA was purified by the standard phenol/chloroform extraction and ethanol precipitation. Genotyping of the SNPs was performed with the MALDI-TOF mass spectrometry iPLEX platform on the MassArray-4 system (Agena Bioscience, Inc., San Diego, CA, USA). Primer sequences used for genotyping are available upon request. To guarantee quality control, 5% of DNA samples were genotyped in duplicates while being blind to the case-control status. The concordance rate of the control genotyping was >99%. Genetic investigations were carried out at the Research Institute for Genetic and Molecular Epidemiology of Kursk State Medical University (Kursk, Russia).
2.4. Statistical Analysis
Statistical power was estimated using the GAS power calculator (https://csg.sph.umich.edu/abecasis/gas_power_calculator/, accessed on May 21, 2022). It has been estimated that we could detect a genotype relative risk (GRR) of 1.30–1.45 with 82–98% power in the overall analysis (474 cases and 470 controls) and a GRR of 1.40–1.5 with 76–83% power in the analysis of groups stratified by sex/risk factors at α = 0.05. Fisher's exact test was used to assess the distribution of genotype frequencies according to the Hardy-Weinberg equilibrium (HWE). Allele and genotype frequencies in the study groups and their associations with the risk of psoriasis were analyzed using the PLINK software v.1.9 [40]. Logistic regression analysis was used to evaluate the associations of GCLC gene polymorphisms with the risk of psoriasis and binary clinical phenotypes. The crude odds ratio (OR) and 95 percent confidence intervals (95% CI) were calculated to assess SNP-binary phenotype associations. Associations of SNPs with continuous phenotypes were evaluated with linear regression analysis, with estimation of differences in mean between genotypes and 95% CI using the SNPstats software (https://www.snpstats.net/start.htm, accessed on April 12, 2023). For SNP-disease associations, allelic, recessive, dominant, and log-additive genetic models were evaluated. Haplotype analysis and visualization of the haplotypic structure of the GCLC gene were performed by the Haploview software, v.4.2 [41]. P-values (Pperm) for allele/genotype/haplotype associations were estimated through adaptive permutations using PLINK and Haploview. Gene-environment interactions were analyzed in groups stratified by risk factors such as cigarette smoking and alcohol abuse. Replication for associations between GCLC gene polymorphisms and psoriasis was performed using the Gene ATLAS database of the UK Biobank (http://geneatlas.roslin.ed.ac.uk (accessed on January 17, 2023)). Associations of pairwise genotype combinations (diplotypes) with the risk of psoriasis were estimated by the chi-squared test and adjusted for multiple comparisons by the false discovery rate (FDR) procedure (False Discovery Rate Online Calculator, https://tools.carbocation.com/FDR, accessed on April 9, 2023).
3. Results
3.1. Baseline and clinical characteristics of the study patients
Baseline and clinical characteristics of the study patients are listed in Table 1. The group of patients with psoriasis was matched to the control group for sex (P = 0.30). The psoriasis patients were more than ten years younger than the healthy subjects. The duration of psoriasis was 10 (4–21) years. The mean age of disease onset was 27 (18–40) years old. The number of smokers in each group was about equal. However, the number of subjects abusing alcohol in the patient group was seven times higher than in the control group (P<0.0001). The psoriatic triad was diagnosed in 54.4% of patients. Most often, psoriatic rashes in patients were observed in the upper (80.0%) and lower (57.4%) extremities, head (47.9%), and trunk (33.08%), which is typical for psoriasis. The most prevalent comorbidities among psoriasis patients were hypertension (22.6%), chronic renal (6.4%), and gastrointestinal (7.0%) diseases.
3.2. Association of GCLC Gene Polymorphisms with the Risk of Psoriasis
Genotype frequencies for five polymorphisms were GCLC in Hardy-Weinberg equilibrium in both cases and controls. Only one SNP, rs17883901, showed a deviation from HWE in both groups (P=0.001). We analyzed associations between the GCLC gene polymorphisms and the risk of psoriasis in entire groups and groups stratified by sex. Table 2 shows a summary of associations between GCLC gene polymorphisms and psoriasis risk in the entire and sex-stratified groups. Allelic, additive, dominant, and recessive genetic models of SNP-disease associations were evaluated, and P-values (Pperm) were assessed through adaptive permutation tests. The most significant Pperm was considered to be the selected genetic model of SNP-disease association. Genotype and allele frequencies of the GCLC gene in healthy controls and patients with psoriasis, along with the most significant Pperm of SNP-disease associations, are reported in Table 3. As can be seen from Table 3, none of the polymorphisms showed an association with the risk of psoriasis as analyzed in the entire group of patients. However, the sex-stratified analysis allowed detecting that SNPs rs648595 (OR=0.56 95%CI 0.35-0.90, Pperm=0.017, recessive model) and rs2397147 (OR=0.54 95%CI 0.30-0.98, Pperm=0.05, recessive model) of the GCLC gene were associated with susceptibility to psoriasis in males.
3.3. Joint Effects of GCLC Gene Polymorphisms on the Risk of Psoriasis
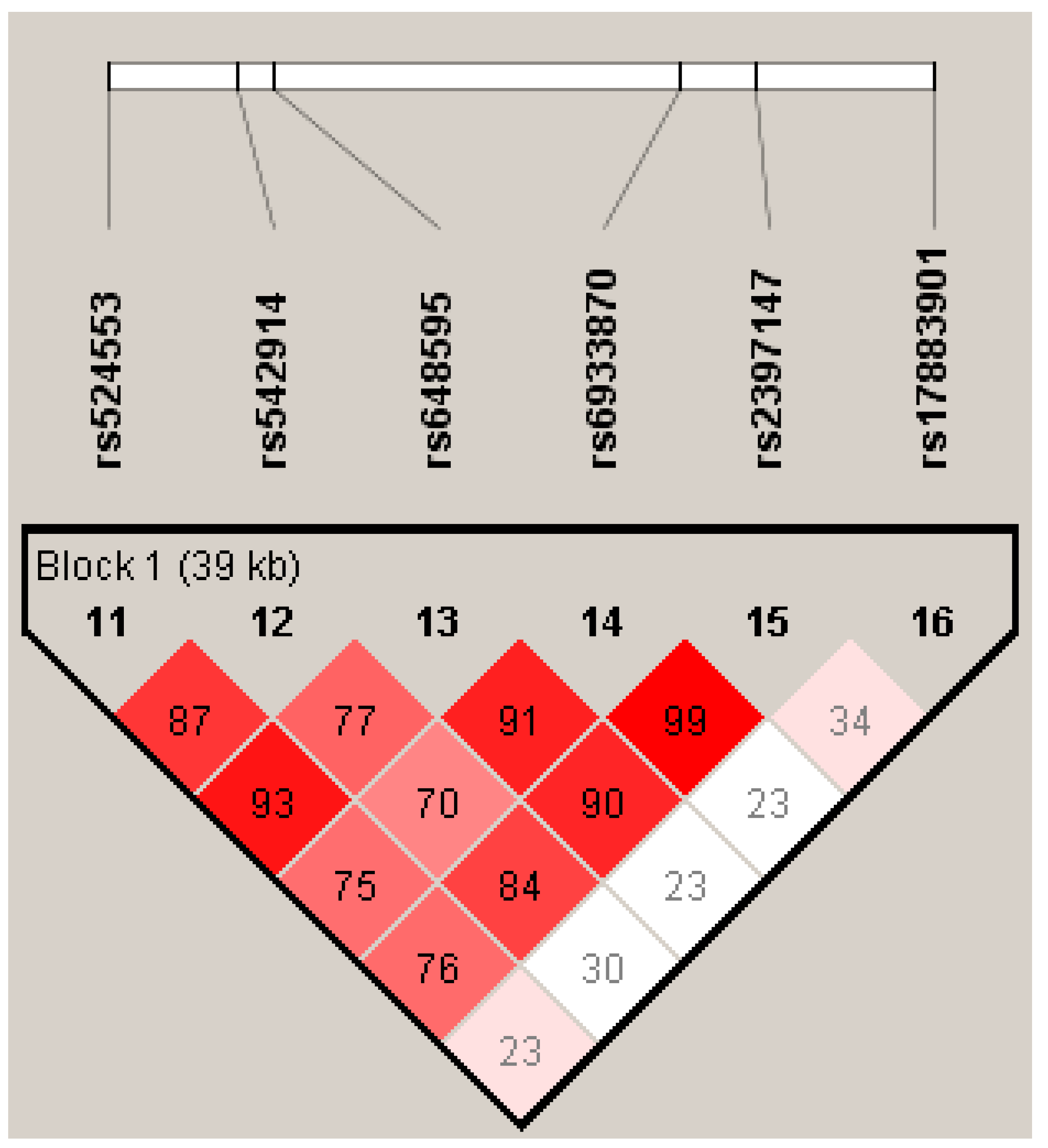
The joint effects of GCLC gene polymorphisms on psoriasis risk were evaluated through haplotype and diplotype analyses. The GCLC haplotypes and their association with psoriasis risk in the entire and sex-stratified groups are shown in Table 4. Four common haplotypes of GCLC (H1–H4) with a frequency of more than 5% were identified in the study groups. The rare haplotype H12, with a frequency of 1%, was detected only in females. Figure 1 shows the linkage disequilibrium plot of the GCLC gene generated by the Haploview software. The polymorphism rs17883901 was not linked to any of the other studied SNPs in the GCLC gene. As can be seen from Table 4, none of the haplotypes was meaningfully associated with the risk of psoriasis, both in the entire and sex-stratified groups (Pperm>0.05).
The results of the diplotype analysis are shown in Table 5. In the entire group, genotype combinations such as rs542914-C/C × rs648595-G/T (FDR-adjusted P=0.03) and rs648595-G/G × rs6933870-C/G (FDR-adjusted P=0.016) of GCLC showed associations with increased and decreased risk of psoriasis, respectively. In the male group, diplotype rs2397147-C/C ×rs17883901-G/G was associated with decreased risk of psoriasis (FDR-adjusted P=0.014), whereas diplotype rs6933870-G/G × rs17883901-G/G (FDR-adjusted P=0.045) showed an association with increased disease risk in females. The remaining six diplotypes associated with disease risk in males did not reach statistical significance after adjusting for multiple tests.
3.4. Gene-Environment Interactions and Psoriasis risk
Since psoriasis is a multifactorial disease, it appears important to investigate the joint influence of environmental risk factors and gene polymorphisms on disease development. Two risk factors, such as cigarette smoking and alcohol abuse, were used for the analysis of gene-environment interactions in psoriasis. Table 6 shows a summary of associations between GCLC gene polymorphisms and psoriasis risk in groups stratified by cigarette smoking and alcohol abuse habits. We found that SNP rs648595 is associated with the risk of psoriasis in cigarette smokers (OR=0.55, 95%CI 0.31-0.99, Pperm=0.049, recessive model), whereas no association of this polymorphism was seen in non-smokers (OR=0.88, 95%CI 0.59-1.31, Pperm=0.52, recessive model). In contrast, SNP rs17883901 showed association with the risk of psoriasis in non-smokers (OR=0.22, 95%CI 0.02-1.97, P=0.14, Pperm=0.002, recessive model), whereas no association with this variant was observed in smoker subjects (OR=0.89, 95%CI 0.11-5.90, P=0.84, Pperm=0.99, recessive model). Notably, polymorphisms rs542914 (OR=0.57, 95%CI 0.36-0.90, Pperm=0.015, recessive model) and rs648595 (OR=0.60, 95%CI 0.39-0.92, Pperm=0.03, recessive model) of GCLC were associated with decreased risk of psoriasis in non-drinkers of alcohol. However, no protective effects of these SNPs against the risk of psoriasis were identified in alcohol abusers (P>0.05).
3.5. Replication of Associations between GCLC Gene Polymorphisms Psoriasis Risk in a population of UK Biobank
It is stated that replication helps ensure that a genotype-phenotype relationship discovered in an original study represents a credible association and is not a chance finding or an artifact due to uncontrolled biases [42,43]. Therefore, we performed a replication analysis of associations between the studied GCLC gene polymorphisms and psoriasis susceptibility in two large populations from the UK Biobank.
Table 7 shows the results of replication analysis to confirm associations between the studied polymorphisms of the GCLC gene and psoriasis risk in a population of the UK Biobank. It has been revealed that two SNPs, such as rs6933870 (P=0.063) and rs2397147 (P=0.057) of GCLC, showed a clear tendency in their association with the risk of psoriasis in one of the UK cohorts. Formally, we cannot conclude that the GCLC gene polymorphisms we studied are replicated in an independent population. Non-replication of SNP-disease associations might be in part explained by inter-population genetic differences, and this issue has been proposed to be readily resolved by the use of a gene-based approach rather than either an SNP-based or a haplotype-based approach [42,44]. Pursuing this proposal, we performed an association analysis of psoriasis with all SNPs of the GCLC gene genotyped in the UK Biobank cohorts. As a result (Table 8), 75 and 21 SNPs of the GCLC gene in the first and second UK Biobank cohorts, respectively, have been found to be associated with the risk of psoriasis at a P-value ≤ 0.05. Two polymorphisms of GCLC, such as rs547541077 (P=0.004) and rs7764361 (P=0.039) were associated with psoriasis risk in both cohorts.
Figure 1.
Linkage disequilibrium (LD) plot of the GCLC gene generated by the Haploview software, v.4.2. Lewontin’s standardized coefficient D' values serve as a means to represent LD. The magnitude and significance of pairwise LD are shown by shading, with a red-to-white gradient showing higher-to-lower LD values.
Figure 1.
Linkage disequilibrium (LD) plot of the GCLC gene generated by the Haploview software, v.4.2. Lewontin’s standardized coefficient D' values serve as a means to represent LD. The magnitude and significance of pairwise LD are shown by shading, with a red-to-white gradient showing higher-to-lower LD values.
3.6. Association of GCLC Gene Polymorphisms with Clinical Features of Psoriasis
Associations of GCLC gene polymorphisms with clinical manifestations of psoriasis were analyzed and adjusted for sex. It has been revealed that a carriage of genotypes rs542914CA and AA of GCLC was positively associated with the psoriatic triad (OR=1.72, 95%CI 1.18-2.51, P=0.005). Earlier onset of psoriasis was associated with the effects of SNPs rs648595 (difference -2.04, 95%CI -3.67 - -0.40, P=0.015) and rs6933870 (difference -1.73, 95%CI -3.36 - -0.10, P=0.038). Carriage of genotype rs524553TT of GCLC was found to be associated with more frequent flare-ups of psoriasis (difference 0.67, 95%CI 0.01 - 1.33, P=0.047). Polymorphisms have been found to be associated with psoriasis localization features. Figure 2 summarizes the findings of the analysis. SNP rs648595 showed association with scalp psoriasis (OR=1.32, 95%CI 1.01-1.74, P=0.04, log-additive genetic model). Polymorphisms rs648595 (difference 0.17, 95%CI 0.00 - 0.35, P=0.048, additive genetic model) and rs2397147 (difference 0.27, 95%CI 0.03 - 0.50, P=0.025, overdominant genetic model) of GCLC were associated with an increased area of skin lesions on the scalp. In addition, genotypes rs2397147TC and C/C were associated with increased infiltration (difference 0.23, 95%CI 0.03 - 0.43, P=0.023) and peeling (difference 0.22, 95%CI 0.03 - 0.42, P=0.026) of psoriatic lesions on the trunk. Genotype rs524553CT was also associated with increased infiltration (difference 0.22, 95%CI 0.01 - 0.42, P=0.037) and peeling (difference 0.22, 95%CI 0.02 - 0.43, P=0.029) of psoriatic lesions on the trunk. The polymorphism rs17883901 of GCLC was found to be associated with psoriasis on the knees (OR=2.34, 95%CI 1.20-4.58, P=0.019, additive genetic model). Moreover, genotype rs17883901AA was associated with psoriasis on the wrist (OR=31.25, 95%CI 2.68-364.40, P=0.007) and fingers (OR=13.99, 95%CI 1.25-157.15, P=0.03, recessive model). Interestingly, genotypes rs648595 GT and GG were also found to be associated with type 2 diabetes in patients with psoriasis (OR=2.80, 95%CI 1.06-7.37, P=0.021). Notably, all the observed associations with clinical features occurred regardless of sex.
4. Discussion
Since the skin is frequently exposed to environmental insults such as ultraviolet irradiation, exposure to toxic chemicals, or mechanical injury causing oxidative or chemical stress, one of the principal physiologic roles of the skin is as a robust barrier against xenobiotics and free radicals for their metabolic elimination and detoxification [25,45,46]. For promoting these functions, human skin possesses a significant potential for phase II metabolism through reactions of glucuronidation, sulfation, N-acetylation, and glutathione conjugation [45] and therefore the cytoprotective effects of GSH are likely to be of importance in this tissue. Experimental studies by Telorack and co-workers [25] have revealed that knockout mice with keratinocyte-specific deficiency in glutamate cysteine ligase showed a strong reduction in the viability of cell culture in vitro and in the skin in vivo. Furthermore, the authors observed that keratinocytes in glutathione-deficient mice died by apoptosis, ferroptosis, and necroptosis, and the increased cell death was attributed to increased levels of reactive oxygen and nitrogen species, causing DNA and mitochondrial damage [25]. This important research demonstrates the epidermis's exceptional antioxidant (glutathione) capability, which ensures skin integrity and effective wound healing. A deficiency of skin glutathione may contribute to psoriasis development. Genetic polymorphisms of glutamate cysteine ligase that are correlated with a decrease in GCLC mRNA and protein expression, enzyme activity, and GSH content [47,48,49,50,51] represent attractive markers for studying the molecular mechanisms of psoriasis. It is known from the literature, polymorphisms of the GCLC gene have been found to be associated with the risk of cardiometabolic diseases such as coronary artery disease [52,53], ischemic stroke [54], type 1 [55,56] and type 2 [51] diabetes mellitus, polycystic ovary syndrome [57], nonalcoholic fatty liver disease [58] as well as other multifactorial disorders such as bronchial asthma [59], pulmonary tuberculosis [60] and colorectal cancer [61]. However, no studies have been designed so far to investigate the role of GCLC gene polymorphisms in psoriasis susceptibility.
The present study is the first to show that polymorphisms of the GCLC gene are significantly associated with the risk of psoriasis and related to its clinical features. Despite the fact that none of the studied polymorphisms showed an association with the risk of psoriasis in the overall group, two SNPs, such as rs648595 and rs2397147, were found to be associated with a decreased risk of psoriasis in males, suggesting sexual dimorphism in the relationship between the gene variation and susceptibility to psoriasis. Sexual dimorphism was also seen in associations between GCLC diplotypes and disease risk: rs2397147-C/C × rs17883901-G/G was associated with decreased risk of psoriasis in males, whereas diplotype rs6933870-G/G × rs17883901-G/G showed an association with increased disease risk in females. These findings were not surprising because gender differences in psoriasis risk and severity have become a discussable issue among dermatologists in the last few years [62,63].
Notably, sexual dimorphism has also been demonstrated in some genetic association studies on skin disease such as atopic dermatitis [64]. Environmental risk factors such as cigarette smoking and alcohol abuse may explain the mechanisms by which sexual dimorphism determines susceptibility to psoriasis [37,36,65]. We investigated the effect of GCLC gene polymorphisms on psoriasis risk depending on these environmental risk factors in the studied population. We found that polymorphism rs648595 is associated with the risk of psoriasis in exclusively cigarette smokers. Another SNP, rs17883901 of GCLC, was associated with the risk of psoriasis only in non-smokers. Furthermore, polymorphisms rs542914 and rs648595 were found to be associated with decreased risk of psoriasis in non-drinkers of alcohol, whereas no protective effects of these SNPs against the disease risk were seen in subjects who were alcohol abusers.
In addition, the present study revealed multiple sex-independent associations between GCLC gene polymorphisms and various clinical features in patients. In particular, SNP rs542914 was associated with the psoriatic triad, whereas rs648595 and rs6933870 polymorphisms were associated with an earlier onset of psoriasis. Furthermore, SNP rs524553TT of GCLC was found to be associated with more frequent flare-ups of psoriasis. The polymorphisms rs648595, rs2397147, rs524553, and rs17883901 have been specifically associated with psoriatic lesions of various localizations on the body. These findings suggest area-specific genetic effects of the studied polymorphisms of the GCLC gene that may be attributed to inter-individual differences in gene expression and therefore rates in glutathione biosynthesis by the skin from different body areas, as was demonstrated with regard to the rate of glutathione conjugation in different organs [66]. It is also known that the levels of glutathione may vary in sun-exposed and sun-protected areas [67], suggesting that UV exposure may impact glutathione biosynthesis in the skin.
A replication analysis of associations between the studied GCLC gene polymorphisms and psoriasis susceptibility in two large populations from the UK Biobank allowed the identification of two SNPs, such as rs6933870 and rs2397147, that showed a clear tendency in the association with the risk of psoriasis. We hypothesize that differences in genetic architecture between the studied populations, including changes in minor allele frequencies and linkage disequilibrium between polymorphisms, may explain the non-replication of SNP-disease relationships. In this context, if the GCLC gene is important for psoriasis pathogenesis, this means that other SNPs of this gene may contribute to disease risk in the UK Biobank populations. Pursuing this assumption, we have analyzed all SNPs of the GCLC gene genotyped in the UK Biobank cohorts with regard to their association with psoriasis risk and found that more than 70 of GCLC gene polymorphisms are associated with disease risk, at least at a P-value ≤ 0.05.
The functional implications of GCLC polymorphisms associated with the development of psoriasis were particularly important for the pathophysiological interpretation of the observed SNP-phenotype relationships. Functional annotation of some polymorphisms of the GCLC gene was performed in our previous study [54]. In particular, we found that allele rs648595G (this SNP showed the most significant associations with psoriasis) is associated with decreased expression of GCLC in blood, non-sun-exposed suprapubic skin, and sun-exposed lower leg skin. Meanwhile, this allele is associated with increased levels of GCLC mRNA in the pancreas, where its expression is two-fold lower than in the skin (GTEx portal, https://gtexportal.org, accessed on May 1, 2023). Similar associations were seen for the rs2397147С allele. Polymorphism rs648595 of GCLC has regulatory potential and is located in transcription factor (TF) binding or DNase hypersensitivity sites [54]. Data from the mQTL Database (http://www.mqtldb.org, accessed on May 1, 2023) shows that, at the pregnancy time point, SNP rs648595 is associated with trans-mQTL - hypomethylation of CpG cg06093712 (position 4:13546145) and trans-mQTL - hypomethylation of CpG cg16209652 (position 2: 142888957). Annotation of the rs648595 polymorphism with the bioinformatics resource HaploReg v4.2 (https://pubs.broadinstitute.org/mammals/haploreg/haploreg.php, accessed on May 1, 2023) showed that the SNP is predicted to be located within the TF-binding site for transcription factor AP-1 (activator protein 1) that is known to control gene expression in response to various stimuli such as cytokines, growth hormones, stress, and infections [68]. In the liver, SNP rs648595 is enriched with enhancer (H3K4me1 and H3K27ac) and promotor (H3K4me3 and H3K9ac) histone marks that may regulate the transcriptional activity of genes. In particular, H3K4me1 (monomethylation on lysine 4 of histone H3) is a dynamic modification that was specifically found to mark both active and primed enhancers [69]. Enhancers bearing the H3K4me1 mark were found to be poised for activation in response to external stimuli [70]. A work by Lauberth and co-workers [71] provided the mechanism by which H3K4me3 promotes rapid gene activation. Finally, H3K9ac co-occurs highly with histone marks such as H3K14ac and H3K4me3 and these three marks are known as the hallmark of active gene promoters [72]. Taken together, the above data from epigenetic studies clearly show that polymorphism rs648595 of the GCLC gene represents an important genetic variant that allows for the activation of gene expression in the liver. Thus, it can be assumed that in the liver, the main organ that synthesizes most of the body's glutathione to provide organs and tissues, this polymorphism, located in the enhancer region, is apparently involved in the mechanisms of activation of the GCLC gene, thereby increasing the rate of glutathione biosynthesis.
Our findings of gene-environment interactions indicate that risk factors such as cigarette smoking and alcohol abuse can modify the associations between GCLC gene polymorphisms and the risk of psoriasis (Table 6). It is known that reduced glutathione plays an important role in ethanol detoxification, and acute ethanol administration was found to deplete GSH in the liver and other organs [73]. The leveling of the protective effects of the rs648595 and rs542914 polymorphisms in chronic alcohol abusers appears to be explained by the fact that persistent ethanol intake may diminish the endogenous pool of glutathione [74,75]. Meanwhile, in vitro study by Kimura and co-workers [76] has revealed that primary human hepatocytes treated with 100 and 200 mM ethanol were found to induce transcriptional activity of the GCLC gene through the activation of the NF-κB pathway. Tobacco smoking is also well known to deplete glutathione [77,78,79]. Thus, our study supports the causative roles of tobacco smoking and alcohol abuse in the development of psoriasis, and the negative effects of these environmental factors eliminate the protective role of polymorphisms of the GCLC gene against disease risk. Sexual dimorphism in the discovered associations of GCLC gene polymorphisms with the risk of psoriasis is apparently associated with differences in environmental risk factors such as smoking and alcohol abuse between males and females. Considering the well-recognized role of oxidative stress in the pathogenesis of psoriasis [13,14,15,16], the mechanisms by which glutathione exerts protective effects against disease development are associated with the key role of glutathione in detoxifying ROS and environmental toxicants, penetrating and generating in the skin. We think that the role of glutathione in the pathogenesis of psoriasis is not limited by the protection of cells from oxidative damage. The involvement of glutathione in the molecular mechanisms of psoriasis may also be explained by the role of GSH in the regulation of cell proliferation and wound healing as well as in the inhibition of apoptotic pathways [21,22]. Furthermore, glutathione play an important role in the regulation of the immune system and inflammation, two faces of the same biological coin [80]. Glutathione has been shown to have a wide range of effects on the immune system, either activating or suppressing the immune response to control inflammation. In particular, reduced glutathione is required for the control of innate and adaptive immunological processes such as T-lymphocyte proliferation, phagocytic activity of polymorphonuclear neutrophils, and dendritic cell functions, as well as antigen presentation by antigen-presenting cells [80,81,82]. Changes in glutathione concentrations may be critical in many autoimmune disease disorders, including psoriasis, that are commonly induced, detrimented, and perpetuated by inflammatory or immune responses mediated by ROS [83]. For instance, glutathione was found to suppress the immune reaction in mice with allergic contact dermatitis [84]. GSH is known to inhibit the production of most inflammatory cytokines and maintain adequate production of interferon-gamma by dendritic cells, a process that is essential for the host defense against intracellular pathogens [80].
Our study has several limitations. Since our study was the first to investigate the contribution of GCLC gene polymorphisms to psoriasis risk in relatively small groups of patients, further studies in independent populations with a larger sample size are required to replicate the observed associations. However, such studies are recommended to follow the gene-based approach to look for associations between psoriasis and a wider spectrum of polymorphisms in the GCLC gene. Following this approach, nevertheless, it should be taken into account that the studied polymorphisms might be characterized by weak or moderate phenotypic effects that cannot be reproduced in independent populations given their genetic heterogeneity in minor allele frequencies and linkage disequilibrium between the loci [85,86]. Some studies have recently reported that genetic differences between races or ethnicities may account for inter-population differences in glutathione metabolism [47,87]. A relatively small number of study subjects in the case and control groups did not allow analyzing the joint effects of GCLC gene polymorphisms and environmental risk factors (smoking and alcohol abuse) separately in males and females in order to obtain estimates of sex-specific gene-environmental interactions contributing to psoriasis susceptibility. Since the studied polymorphisms of the GCLC gene are located in noncoding regions, their phenotypic effects should be interpreted with caution because no investigations were done to assess gene expression in skin biopsies of study patients.
5. Conclusions
The present study demonstrated for the first time that polymorphisms in the gene encoding the catalytic subunit of glutamate cysteine ligase represent novel genetic markers for susceptibility to psoriasis. The phenotypic effects of GCLC gene polymorphisms on the development of psoriasis are modified by tobacco smoking and alcohol abuse, known environmental factors increasing disease risk. We suppose that the GCLC gene may contribute to the risk of psoriasis through a diminished biosynthesis of glutathione in the skin, where GSH regulates a plethora of cellular processes such as redox homeostasis, detoxification of xenobiotics, innate and adaptive immune functions, inflammation, cell proliferation and differentiation, and apoptosis that have been implicated in disease pathogenesis. Better understanding the relation between the GCLC gene polymorphisms and glutathione biosynthesis in the skin and the molecular mechanisms by which this gene contributes to psoriasis will open new scientifically based options for disease therapy and prevention targeted on glutathione metabolism. The use of pharmacogenetics and precision medicine approaches [88,89] will make it possible to subclassify patient groups based on environmental risk factors and clinically significant genetic variants that affect glutathione metabolism and thus personalize treatment and prevention of psoriasis.
Author Contributions
Conceptualization, A.P.; methodology, A.P., E.E., O.B.,R.S. and M.C.; software, A.P., M.S.; validation, A.P., M.C.; formal analysis, A.P.,E.E., O.B.; investigation, A.P., E.E, R.S., O.B.,A.S.; resources, E.E, R.S.; data curation, A.P., E.E, M.S.; writing—original draft preparation, A.P., E.E., M.S.; writing—review and editing, A.P.,M.S., M.C.; visualization, A.P.; supervision, A.P.; project administration, A.P.. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of Kursk State Medical University (protocol No. 8, 13.11.2017).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data supporting reported results are available upon request.
Acknowledgments
We are grateful to all patients with [psoriasis, healthy volunteers, and the staff of hospitals of the Kursk region who participated in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gudjonsson JE, Elder JT. Psoriasis: epidemiology. Clin Dermatol. 2007 Nov-Dec;25(6):535-46. [CrossRef]
- Campanati A, Marani A, Martina E, Diotallevi F, Radi G, Offidani A. Psoriasis as an Immune-Mediated and Inflammatory Systemic Disease: From Pathophysiology to Novel Therapeutic Approaches. Biomedicines. 2021 Oct 21;9(11):1511. [CrossRef]
- World Health Organization. Global report on psoriasis. World Health Organization. 2016.
- Lebwohl M. Psoriasis. Lancet. 2003 Apr 5;361(9364):1197-204. [CrossRef]
- Kubanov AA, Bakulev AL, Fitileva TV, Novoderezhkina E, Gilloteau I, Tian H, Howe T, Pietri G. Disease Burden and Treatment Patterns of Psoriasis in Russia: A Real-World Patient and Dermatologist Survey. Dermatol Ther (Heidelb). 2018 Dec;8(4):581-592. [CrossRef]
- Di Meglio P, Villanova F, Nestle FO. Psoriasis. Cold Spring Harb Perspect Med. 2014 Aug 1;4(8):a015354. [CrossRef]
- Rendon A, Schäkel K. Psoriasis Pathogenesis and Treatment. Int J Mol Sci. 2019 Mar 23;20(6):1475. [CrossRef]
- Harden JL, Krueger JG, Bowcock AM. The immunogenetics of Psoriasis: A comprehensive review. J Autoimmun. 2015 Nov;64:66-73. [CrossRef]
- Grjibovski AM, Olsen AO, Magnus P, Harris JR. Psoriasis in Norwegian twins: contribution of genetic and environmental effects. J Eur Acad Dermatol Venereol. 2007 Nov;21(10):1337-43. [CrossRef]
- Gupta R, Debbaneh MG, Liao W. Genetic Epidemiology of Psoriasis. Curr Dermatol Rep. 2014 Mar;3(1):61-78. [CrossRef]
- Capon F. The Genetic Basis of Psoriasis. Int J Mol Sci. 2017 Nov 25;18(12):2526. [CrossRef]
- Fan X, Yang S, Huang W, Wang ZM, Sun LD, Liang YH, Gao M, Ren YQ, Zhang KY, Du WH, Shen YJ, Liu JJ, Zhang XJ. Fine mapping of the psoriasis susceptibility locus PSORS1 supports HLA-C as the susceptibility gene in the Han Chinese population. PLoS Genet. 2008 Mar 21;4(3):e1000038. [CrossRef]
- Trouba KJ, Hamadeh HK, Amin RP, Germolec DR. Oxidative stress and its role in skin disease. Antioxid Redox Signal. 2002 Aug;4(4):665-73. [CrossRef]
- Cannavò SP, Riso G, Casciaro M, Di Salvo E, Gangemi S. Oxidative stress involvement in psoriasis: a systematic review. Free Radic Res. 2019 Aug;53(8):829-840. [CrossRef]
- Pleńkowska J, Gabig-Cimińska M, Mozolewski P. Oxidative Stress as an Important Contributor to the Pathogenesis of Psoriasis. Int J Mol Sci. 2020 Aug 27;21(17):6206. [CrossRef]
- Dobrică EC, Cozma MA, Găman MA, Voiculescu VM, Găman AM. The Involvement of Oxidative Stress in Psoriasis: A Systematic Review. Antioxidants (Basel). 2022 Jan 29;11(2):282. [CrossRef]
- Yang S, Yan KL, Zhang XJ, Xiao FL, Fan X, Gao M, Cui Y, Wang PG, Zhang GL, Sun LD, Wang ZM, Wang DZ, Zhang KY, Huang W, Liu JJ. Systematic evaluation of association between the microsomal glutathione S-transferase 2 common variation and psoriasis vulgaris in Chinese population. Arch Dermatol Res. 2006 Aug;298(3):107-12. [CrossRef]
- Solak B, Karkucak M, Turan H, Ocakoğlu G, Özemri Sağ Ş, Uslu E, Yakut T, Erdem T. Glutathione S-Transferase M1 and T1 Gene Polymorphisms in Patients with Chronic Plaque-Type Psoriasis: A Case-Control Study. Med Princ Pract. 2016;25(2):155-8. [CrossRef]
- Srivastava DSL, Jain VK, Verma P, Yadav JP. Polymorphism of glutathione S-transferase M1 and T1 genes and susceptibility to psoriasis disease: A study from North India. Indian J Dermatol Venereol Leprol. 2018 Jan-Feb;84(1):39-44. [CrossRef]
- Guarneri F, Sapienza D, Papaianni V, Marafioti I, Guarneri C, Mondello C, Roccuzzo S, Asmundo A, Cannavò SP. Association between genetic polymorphisms of glutathione S-transferase M1/T1 and psoriasis in a population from the area of the strict of messina (Southern Italy). Free Radic Res. 2020 Jan;54(1):57-63. [CrossRef]
- Wu G, Fang YZ, Yang S, Lupton JR, Turner ND. Glutathione metabolism and its implications for health. J Nutr 2004; 134(3):489–492. [CrossRef]
- Sies H. Glutathione and its role in cellular functions. Free Radic Biol Med. 1999 Nov;27(9-10):916-21. [CrossRef]
- Sipos K, Lange H, Fekete Z, Ullmann P, Lill R, Kispal G. Maturation of cytosolic iron-sulfur proteins requires glutathione. J Biol Chem. 2002 Jul 26;277(30):26944-9. [CrossRef]
- Jacquoilleot S, Sheffield D, Olayanju A, Sison-Young R, Kitteringham NR, Naisbitt DJ, Aleksic M. Glutathione metabolism in the HaCaT cell line as a model for the detoxification of the model sensitisers 2,4-dinitrohalobenzenes in human skin. Toxicol Lett. 2015 Aug 19;237(1):11-20. [CrossRef]
- Telorack M, Meyer M, Ingold I, Conrad M, Bloch W, Werner S. A Glutathione-Nrf2-Thioredoxin Cross-Talk Ensures Keratinocyte Survival and Efficient Wound Repair. PLoS Genet. 2016 Jan 25;12(1):e1005800. [CrossRef]
- Prussick R, Prussick L, Gutman J. Psoriasis Improvement in Patients Using Glutathione-enhancing, Nondenatured Whey Protein Isolate: A Pilot Study. J Clin Aesthet Dermatol. 2013 Oct;6(10):23-6.
- Zhou Q, Mrowietz U, Rostami-Yazdi M. Oxidative stress in the pathogenesis of psoriasis. Free Radic Biol Med. 2009 Oct 1;47(7):891-905. [CrossRef]
- Medovic MV, Jakovljevic VL, Zivkovic VI, Jeremic NS, Jeremic JN, Bolevich SB, Ravic Nikolic AB, Milicic VM, Srejovic IM. Psoriasis between Autoimmunity and Oxidative Stress: Changes Induced by Different Therapeutic Approaches. Oxid Med Cell Longev. 2022 Mar 12;2022:2249834. [CrossRef]
- Franklin CC, Backos DS, Mohar I, White CC, Forman HJ, Kavanagh TJ. Structure, function, and post-translational regulation of the catalytic and modifier subunits of glutamate cysteine ligase. Mol Aspects Med. 2009 Feb-Apr;30(1-2):86-98. [CrossRef]
- Klyosova E, Azarova I, Polonikov A. A Polymorphism in the Gene Encoding Heat Shock Factor 1 (HSF1) Increases the Risk of Type 2 Diabetes: A Pilot Study Supports a Role for Impaired Protein Folding in Disease Pathogenesis. Life (Basel). 2022 Nov 20;12(11):1936. [CrossRef]
- Lazarenko V, Churilin M, Azarova I, Klyosova E, Bykanova M, Ob'edkova N, Churnosov M, Bushueva O, Mal G, Povetkin S, Kononov S, Luneva Y, Zhabin S, Polonikova A, Gavrilenko A, Saraev I, Solodilova M, Polonikov A. Comprehensive Statistical and Bioinformatics Analysis in the Deciphering of Putative Mechanisms by Which Lipid-Associated GWAS Loci Contribute to Coronary Artery Disease. Biomedicines. 2022 Jan 25;10(2):259. [CrossRef]
- Kobzeva KA, Shilenok IV, Belykh AE, et al. C9orf16 (BBLN) gene, encoding a member of Hero proteins, is a novel marker in ischemic stroke risk. Research Results in Biomedicine. 2022;8(3):278-292. [CrossRef]
- Griffiths CE, Barker JN. Pathogenesis and clinical features of psoriasis. Lancet. 2007 Jul 21;370(9583):263-271. [CrossRef]
- Fredriksson T, Pettersson U. Severe psoriasis--oral therapy with a new retinoid. Dermatologica. 1978;157(4):238-44. [CrossRef]
- Klyosova, E.Y.; Azarova, I.E.; Sunyaykina, O.A.; Polonikov, A.V. Validity of a brief screener for environmental risk factors of age-related diseases using type 2 diabetes and coronary artery disease as examples. Res. Results Biomed. 2022, 8, 130–137.
- Armstrong AW, Harskamp CT, Dhillon JS, Armstrong EJ. Psoriasis and smoking: a systematic review and meta-analysis. Br J Dermatol. 2014 Feb;170(2):304-14. [CrossRef]
- Brenaut E, Horreau C, Pouplard C, Barnetche T, Paul C, Richard MA, Joly P, Le Maître M, Aractingi S, Aubin F, Cribier B, Jullien D, Ortonne JP, Misery L. Alcohol consumption and psoriasis: a systematic literature review. J Eur Acad Dermatol Venereol. 2013 Aug;27 Suppl 3:30-5. [CrossRef]
- Polonikov AV, Ivanov VP, Solodilova MA. CYP2E1 gene promoter polymorphism -1293G>C increases the risk of essential hypertension in men with alcohol abuse. Bull Exp Biol Med. 2013 Oct;155(6):734-7. [CrossRef]
- Polonikov AV, Samgina TA, Nazarenko PM, Bushueva OY, Ivanov VP. Alcohol Consumption and Cigarette Smoking are Important Modifiers of the Association Between Acute Pancreatitis and the PRSS1-PRSS2 Locus in Men. Pancreas. 2017 Feb;46(2):230-236. [CrossRef]
- Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D, Maller J, Sklar P, de Bakker PI, Daly MJ, Sham PC. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007 Sep;81(3):559-75. [CrossRef]
- Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005 Jan 15;21(2):263-5. [CrossRef]
- Neale BM, Sham PC. The future of association studies: gene-based analysis and replication. Am J Hum Genet. 2004 Sep;75(3):353-62. [CrossRef]
- Kraft P, Zeggini E, Ioannidis JP. Replication in genome-wide association studies. Stat Sci. 2009 Nov 1;24(4):561-573. [CrossRef]
- Belonogova NM, Zorkoltseva IV, Tsepilov YA, Axenovich TI. Gene-based association analysis identifies 190 genes affecting neuroticism. Sci Rep. 2021 Jan 28;11(1):2484. [CrossRef]
- Manevski N, Swart P, Balavenkatraman KK, Bertschi B, Camenisch G, Kretz O, Schiller H, Walles M, Ling B, Wettstein R, Schaefer DJ, Itin P, Ashton-Chess J, Pognan F, Wolf A, Litherland K. Phase II metabolism in human skin: skin explants show full coverage for glucuronidation, sulfation, N-acetylation, catechol methylation, and glutathione conjugation. Drug Metab Dispos. 2015 Jan;43(1):126-39. [CrossRef]
- Azevedo Martins TE, Sales de Oliveira Pinto CA, Costa de Oliveira A, Robles Velasco MV, Gorriti Guitiérrez AR, Cosquillo Rafael MF, Tarazona JPH, Retuerto-Figueroa MG. Contribution of Topical Antioxidants to Maintain Healthy Skin—A Review. Scientia Pharmaceutica. 2020; 88(2):27. https://doi.org/10.3390/scipharm88020027.
- Le TM, Willis AS, Barr FE, Cunningham GR, Canter JA, Owens SE, Apple RK, Ayodo G, Reich D, Summar ML. An ethnic-specific polymorphism in the catalytic subunit of glutamate-cysteine ligase impairs the production of glutathione intermediates in vitro. Mol Genet Metab. 2010 Sep;101(1):55-61. [CrossRef]
- Nichenametla SN, Lazarus P, Richie JP Jr. A GAG trinucleotide-repeat polymorphism in the gene for glutathione biosynthetic enzyme, GCLC, affects gene expression through translation. FASEB J. 2011 Jul;25(7):2180-7. [CrossRef]
- Butticaz C, Gysin R, Cuénod M, Do KQ. Interaction of GAG trinucleotide repeat and C-129T polymorphisms impairs expression of the glutamate-cysteine ligase catalytic subunit gene. Free Radic Biol Med. 2011 Mar 1;50(5):617-23. [CrossRef]
- Wang D, Curtis A, Papp AC, Koletar SL, Para MF. Polymorphism in glutamate cysteine ligase catalytic subunit (GCLC) is associated with sulfamethoxazole-induced hypersensitivity in HIV/AIDS patients. BMC Med Genomics. 2012 Jul 23;5:32. [CrossRef]
- Azarova I, Klyosova E, Lazarenko V, Konoplya A, Polonikov A. Genetic variants in glutamate cysteine ligase confer protection against type 2 diabetes. Mol Biol Rep. 2020 Aug;47(8):5793-5805. [CrossRef]
- Koide S, Kugiyama K, Sugiyama S, Nakamura S, Fukushima H, Honda O, Yoshimura M, Ogawa H. Association of polymorphism in glutamate-cysteine ligase catalytic subunit gene with coronary vasomotor dysfunction and myocardial infarction. J Am Coll Cardiol. 2003 Feb 19;41(4):539-45. [CrossRef]
- Skvortsova L, Perfelyeva A, Khussainova E, Mansharipova A, Forman HJ, Djansugurova L. Association of GCLM -588C/T and GCLC -129T/C Promoter Polymorphisms of Genes Coding the Subunits of Glutamate Cysteine Ligase with Ischemic Heart Disease Development in Kazakhstan Population. Dis Markers. 2017;2017:4209257. [CrossRef]
- Polonikov A, Bocharova I, Azarova I, Klyosova E, Bykanova M, Bushueva O, Polonikova A, Churnosov M, Solodilova M. The Impact of Genetic Polymorphisms in Glutamate-Cysteine Ligase, a Key Enzyme of Glutathione Biosynthesis, on Ischemic Stroke Risk and Brain Infarct Size. Life (Basel). 2022 Apr 18;12(4):602. [CrossRef]
- Bekris LM, Shephard C, Janer M, Graham J, McNeney B, Shin J, Zarghami M, Griffith W, Farin F, Kavanagh TJ, Lernmark A. Glutamate cysteine ligase catalytic subunit promoter polymorphisms and associations with type 1 diabetes age-at-onset and GAD65 autoantibody levels. Exp Clin Endocrinol Diabetes. 2007 Apr;115(4):221-8. [CrossRef]
- Vieira SM, Monteiro MB, Marques T, Luna AM, Fortes MA, Nery M, Queiroz M, Dib SA, Vendramini MF, Azevedo MJ, Canani LH, Parisi MC, Pavin EJ, Giannella-Neto D, Corrêa-Giannella ML. Association of genetic variants in the promoter region of genes encoding p22phox (CYBA) and glutamate cysteine ligase catalytic subunit (GCLC) and renal disease in patients with type 1 diabetes mellitus. BMC Med Genet. 2011 Sep 30;12:129. [CrossRef]
- Yang C, Xi M, Liu H, Bai H, Jiang C, Liu Q, Fan P. Association of Polymorphisms of Glutamate Cysteine Ligase Genes GCLC C-129 T and GCLM C-588 T with Risk of Polycystic Ovary Syndrome in Chinese Women. Reprod Sci. 2022 Jun;29(6):1790-1800. [CrossRef]
- Macaluso FS, Maida M, Petta S. Genetic background in nonalcoholic fatty liver disease: A comprehensive review. World J Gastroenterol. 2015 Oct 21;21(39):11088-111. [CrossRef]
- Polonikov AV, Ivanov VP, Solodilova MA, Khoroshaya IV, Kozhuhov MA, Panfilov VI. The relationship between polymorphisms in the glutamate cysteine ligase gene and asthma susceptibility. Respir Med. 2007 Nov;101(11):2422-4. [CrossRef]
- Yuniastuti A, Susanti R, Mustikaningtyas D. Polymorphism of Glutamate-Cysteine Ligase Subunit Catalytic (GCLC) Gene in Pulmonary Tuberculosis Patients. Pak J Biol Sci. 2017;20(8):397-402. [CrossRef]
- Bykanova MA, Solodilova MA, Azarova IE, Klyosova EY, Bushueva OY, Polonikova AA, Churnosov MI, Polonikov AV. Genetic variation at the catalytic subunit of glutamate cysteine ligase contributes to the susceptibility to sporadic colorectal cancer: a pilot study. Mol Biol Rep. 2022 Jul;49(7):6145-6154. [CrossRef]
- Hägg D, Sundström A, Eriksson M, Schmitt-Egenolf M. Severity of Psoriasis Differs Between Men and Women: A Study of the Clinical Outcome Measure Psoriasis Area and Severity Index (PASI) in 5438 Swedish Register Patients. Am J Clin Dermatol. 2017 Aug;18(4):583-590. [CrossRef]
- Murer C, Sgier D, Mettler SK, Guillet C, Maul JT, Djamei V, Navarini AA, Anzengruber F. Gender differences in psoriasis: a Swiss online psoriasis survey. Arch Dermatol Res. 2021 Mar;313(2):89-94. [CrossRef]
- Dvornyk V, Ponomarenko I, Belyaeva T, Reshetnikov E, Churnosov M. Filaggrin gene polymorphisms are associated with atopic dermatitis in women but not in men in the Caucasian population of Central Russia. PLoS One. 2021 Dec 9;16(12):e0261026. [CrossRef]
- Bayaraa B, Imafuku S. Relationship between environmental factors, age of onset and familial history in Japanese patients with psoriasis. J Dermatol. 2018 Jun;45(6):715-718. [CrossRef]
- Temellini A, Castiglioni M, Giuliani L, Mussi A, Giulianotti PC, Pietrabissa A, Angeletti CA, Mosca F, Pacifici GM. Glutathione conjugation with 1-chloro-2,4-dinitrobenzene (CDNB): interindividual variability in human liver, lung, kidney and intestine. Int J Clin Pharmacol Ther. 1995 Sep;33(9):498-503.
- Dilokthornsakul W, Dhippayom T, Dilokthornsakul P. The clinical effect of glutathione on skin color and other related skin conditions: A systematic review. J Cosmet Dermatol. 2019 Jun;18(3):728-737. [CrossRef]
- Hess J, Angel P, Schorpp-Kistner M. AP-1 subunits: quarrel and harmony among siblings. J Cell Sci. 2004 Dec 1;117(Pt 25):5965-73. [CrossRef]
- Local A, Huang H, Albuquerque CP, Singh N, Lee AY, Wang W, Wang C, Hsia JE, Shiau AK, Ge K, Corbett KD, Wang D, Zhou H, Ren B. Identification of H3K4me1-associated proteins at mammalian enhancers. Nat Genet. 2018 Jan;50(1):73-82. [CrossRef]
- Creyghton MP, Cheng AW, Welstead GG, Kooistra T, Carey BW, Steine EJ, Hanna J, Lodato MA, Frampton GM, Sharp PA, Boyer LA, Young RA, Jaenisch R. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc Natl Acad Sci U S A. 2010 Dec 14;107(50):21931-6. [CrossRef]
- Lauberth SM, Nakayama T, Wu X, Ferris AL, Tang Z, Hughes SH, Roeder RG. H3K4me3 interactions with TAF3 regulate preinitiation complex assembly and selective gene activation. Cell. 2013 Feb 28;152(5):1021-36. [CrossRef]
- Karmodiya K, Krebs AR, Oulad-Abdelghani M, Kimura H, Tora L. H3K9 and H3K14 acetylation co-occur at many gene regulatory elements, while H3K14ac marks a subset of inactive inducible promoters in mouse embryonic stem cells. BMC Genomics. 2012 Aug 24;13:424. [CrossRef]
- Vogt BL, Richie JP Jr. Glutathione depletion and recovery after acute ethanol administration in the aging mouse. Biochem Pharmacol. 2007 May 15;73(10):1613-21. [CrossRef]
- Guerri C, Grisolía S. Changes in glutathione in acute and chronic alcohol intoxication. Pharmacol Biochem Behav. 1980;13 Suppl 1:53-61. [CrossRef]
- Lauterburg BH, Velez ME. Glutathione deficiency in alcoholics: risk factor for paracetamol hepatotoxicity. Gut. 1988 Sep;29(9):1153-7. [CrossRef]
- Kimura T, Kawasaki Y, Okumura F, Sone T, Natsuki R, Isobe M. Ethanol-induced expression of glutamate-cysteine ligase catalytic subunit gene is mediated by NF-kappaB. Toxicol Lett. 2009 Mar 10;185(2):110-5. [CrossRef]
- Haramaki N, Ikeda H, Takajo Y, Katoh A, Kanaya S, Shintani S, Haramaki R, Murohara T, Imaizumi T. Long-term smoking causes nitroglycerin resistance in platelets by depletion of intraplatelet glutathione. Arterioscler Thromb Vasc Biol. 2001 Nov;21(11):1852-6. [CrossRef]
- van der Toorn M, Smit-de Vries MP, Slebos DJ, de Bruin HG, Abello N, van Oosterhout AJ, Bischoff R, Kauffman HF. Cigarette smoke irreversibly modifies glutathione in airway epithelial cells. Am J Physiol Lung Cell Mol Physiol. 2007 Nov;293(5):L1156-62. [CrossRef]
- Bazzini C, Rossetti V, Civello DA, Sassone F, Vezzoli V, Persani L, Tiberio L, Lanata L, Bagnasco M, Paulmichl M, Meyer G, Garavaglia ML. Short- and long- term effects of cigarette smoke exposure on glutathione homeostasis in human bronchial epithelial cells. Cell Physiol Biochem. 2013;32(7):129-45. [CrossRef]
- Ghezzi P. Role of glutathione in immunity and inflammation in the lung. Int J Gen Med. 2011 Jan 25;4:105-13. [CrossRef]
- Dröge W, Breitkreutz R. Glutathione and immune function. Proc Nutr Soc. 2000 Nov;59(4):595-600. [CrossRef]
- Diotallevi M, Checconi P, Palamara AT, Celestino I, Coppo L, Holmgren A, Abbas K, Peyrot F, Mengozzi M, Ghezzi P. Glutathione Fine-Tunes the Innate Immune Response toward Antiviral Pathways in a Macrophage Cell Line Independently of Its Antioxidant Properties. Front Immunol. 2017 Sep 29;8:1239. [CrossRef]
- Perricone C, De Carolis C, Perricone R. Glutathione: a key player in autoimmunity. Autoimmun Rev. 2009 Jul;8(8):697-701. [CrossRef]
- Hirai A, Minamiyama Y, Hamada T, Ishii M, Inoue M. Glutathione metabolism in mice is enhanced more with hapten-induced allergic contact dermatitis than with irritant contact dermatitis. J Invest Dermatol. 1997 Sep;109(3):314-8. [CrossRef]
- Azarova I, Klyosova E, Polonikov A. The Link between Type 2 Diabetes Mellitus and the Polymorphisms of Glutathione-Metabolizing Genes Suggests a New Hypothesis Explaining Disease Initiation and Progression. Life (Basel). 2021 Aug 28;11(9):886. [CrossRef]
- International HapMap 3 Consortium; Altshuler DM, Gibbs RA, Peltonen L, Altshuler DM, Gibbs RA, Peltonen L, Dermitzakis E, Schaffner SF, Yu F, Peltonen L, Dermitzakis E, Bonnen PE, Altshuler DM, Gibbs RA, de Bakker PI, Deloukas P, Gabriel SB, Gwilliam R, Hunt S, Inouye M, Jia X, Palotie A, Parkin M, Whittaker P, Yu F, Chang K, Hawes A, Lewis LR, Ren Y, Wheeler D, Gibbs RA, Muzny DM, Barnes C, Darvishi K, Hurles M, Korn JM, Kristiansson K, Lee C, McCarrol SA, Nemesh J, Dermitzakis E, Keinan A, Montgomery SB, Pollack S, Price AL, Soranzo N, Bonnen PE, Gibbs RA, Gonzaga-Jauregui C, Keinan A, Price AL, Yu F, Anttila V, Brodeur W, Daly MJ, Leslie S, McVean G, Moutsianas L, Nguyen H, Schaffner SF, Zhang Q, Ghori MJ, McGinnis R, McLaren W, Pollack S, Price AL, Schaffner SF, Takeuchi F, Grossman SR, Shlyakhter I, Hostetter EB, Sabeti PC, Adebamowo CA, Foster MW, Gordon DR, Licinio J, Manca MC, Marshall PA, Matsuda I, Ngare D, Wang VO, Reddy D, Rotimi CN, Royal CD, Sharp RR, Zeng C, Brooks LD, McEwen JE. Integrating common and rare genetic variation in diverse human populations. Nature. 2010 Sep 2;467(7311):52-8. [CrossRef]
- Kinney N, Kang L, Bains H, Lawson E, Husain M, Husain K, Sandhu I, Shin Y, Carter JK, Anandakrishnan R, Michalak P, Garner H. Ethnically biased microsatellites contribute to differential gene expression and glutathione metabolism in Africans and Europeans. PLoS One. 2021 Mar 25;16(3):e0249148. [CrossRef]
- Sychev DA, Malova EU. Evidence-based pharmacogenetics: Is it possible? Int J Risk Saf Med. 2015;27 Suppl 1:S97-8. [CrossRef]
- Johansson Å, Andreassen OA, Brunak S, Franks PW, Hedman H, Loos RJF, Meder B, Melén E, Wheelock CE, Jacobsson B. Precision medicine in complex diseases-Molecular subgrouping for improved prediction and treatment stratification. J Intern Med. 2023 Apr 24. [CrossRef]
Figure 2.
Associations of GCLC gene polymorphisms with psoriatic lesions on the body.
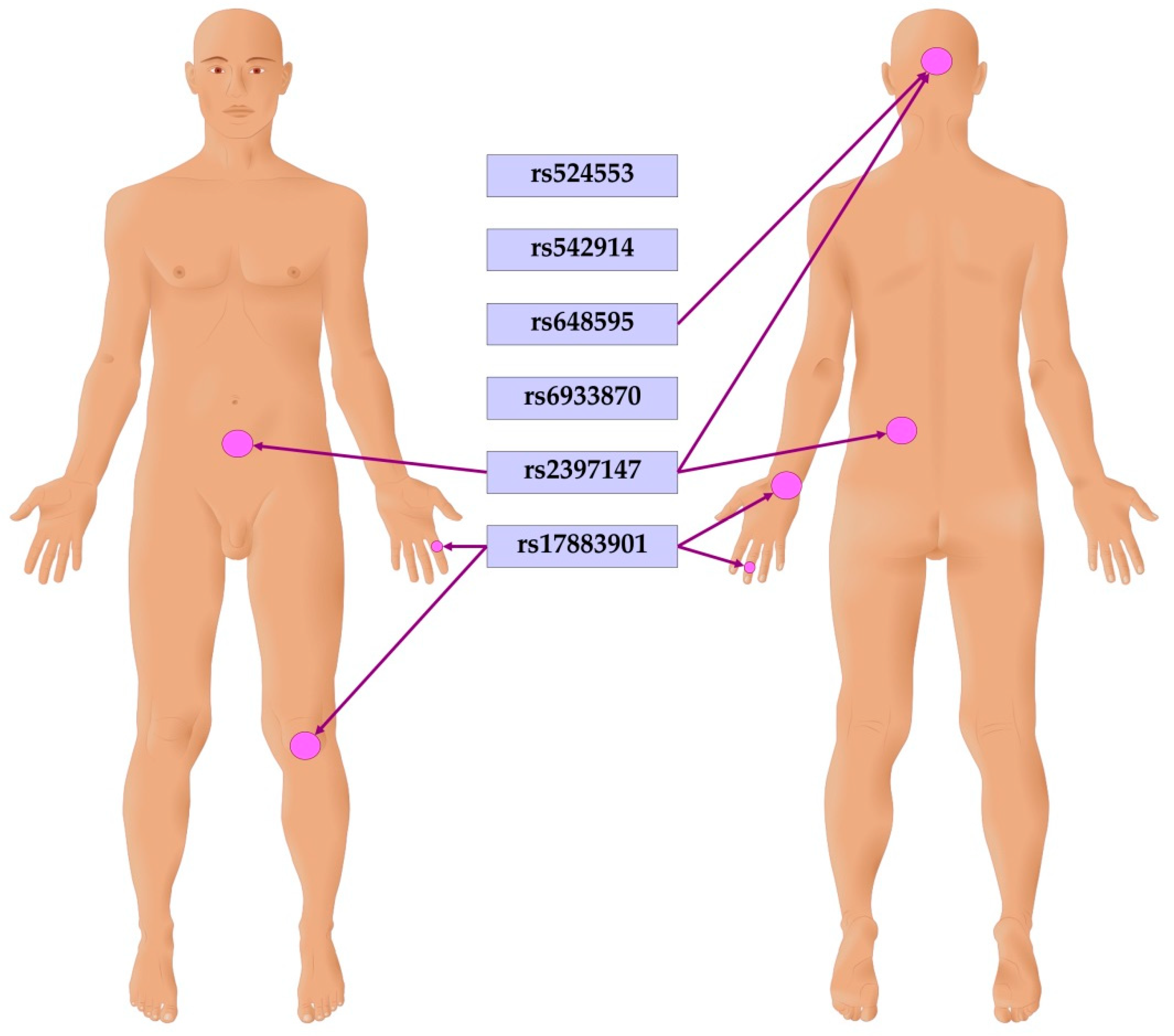

Table 1.
Baseline and clinical characteristics of the study patients.
| Characteristics | Patients with psoriasis n=474 |
Healthy controls n=470 |
P-value* |
| Baseline characteristics | |||
| Age, mean ± standard deviation | 44.3 ± 13.6 | 55.3 ± 6.7 | <0.0001 |
| Males, n (%) | 252 (53.2) | 234 (49.8) | 0.30 |
| Females, n (%) | 222 (46.8) | 236 (50.2) | |
| Risk factors | |||
| Smokers, (ever/never), n (%) | 168 (35.4) | 148 (31.5) | 0.20 |
| Alcohol abusers1, n (%) | 105 (21.2) | 7 (3.2) | <0.0001 |
| Locations of psoriatic lesions | |||
| Psoriatic triad | 256 (54.0) | - | - |
| Scalp | 227 (47.9) | - | |
| Trunk | 160 (33.08) | - | - |
| Hands | 379 (80.0) | - | - |
| Legs | 272 (57.4) | - | - |
| Joints | 128 (27.0) | - | - |
| Low back | 24 (5.1) | - | - |
| Knees | 59 (12.4) | - | - |
| Hips | 21 (4.4) | - | - |
| Elbows | 33 (7.0) | - | - |
| Fingers | 60 (12.6) | - | - |
| Ankles | 24 (5.1) | - | - |
| Feet/toes | 23 (4.9) | - | - |
| Thumbs | 18 (3.8) | - | - |
| Shoulders | 11 (2.3) | - | - |
| Wrists | 33 (7.0) | - | - |
| Nails | 123 (25.9) | - | - |
| Comorbidities | |||
| Type 2 diabetes, n (%) | 15 (3.2) | - | - |
| Arterial hypertension, n (%) | 106 (22.6) | - | - |
| Coronary artery disease, n (%) | 27 (5.7) | - | - |
| Cerebral stroke, n (%) | 9 (1.9) | - | - |
| Chronic thyroid disease, n (%) | 7 (1.5) | - | - |
| Chronic renal disease, n (%) | 30 (6.4) | - | - |
| Chronic gastric disease, n (%) | 33 (7.0) | - | - |
| Chronic pulmonary disease, n (%) | 7 (1.5) | - | - |
| Oncological disease, n (%) | 8 (1.7) | - | - |
|
1 Data on alcohol intake were available from 220 subjects of the control group. *Bold is statistically significant P-value. | |||
Table 2.
A summary of associations between GCLC gene polymorphisms and psoriasis risk in the entire and sex-stratified groups.
Table 2.
A summary of associations between GCLC gene polymorphisms and psoriasis risk in the entire and sex-stratified groups.
| SNP ID | Minor allele | N | Permutation P-values (Pperm) estimated for genetic models of SNP-disease associations* | |||
| Allelic | Additive | Dominant | Recessive | |||
| Entire groups | ||||||
| rs524553 | T | 939 | 0.36 | 0.28 | 0.42 | 0.20 |
| rs542914 | A | 941 | 0.18 | 0.23 | 0.67 | 0.11 |
| rs648595 | G | 941 | 0.21 | 0.58 | 1.00 | 0.13 |
| rs6933870 | G | 942 | 1.00 | 1.00 | 1.00 | 0.86 |
| rs2397147 | C | 940 | 0.48 | 0.29 | 0.86 | 0.40 |
| rs17883901 | A | 810 | 0.63 | 0.78 | 1.00 | 0.15 |
| Males | ||||||
| rs524553 | T | 485 | 0.38 | 0.43 | 0.50 | 0.20 |
| rs542914 | A | 485 | 0.55 | 0.41 | 1.00 | 0.28 |
| rs648595 | G | 484 | 0.048 | 0.23 | 0.86 | 0.017 |
| rs6933870 | G | 485 | 0.25 | 0.13 | 0.32 | 0.09 |
| rs2397147 | C | 484 | 0.11 | 0.11 | 0.31 | 0.05 |
| rs17883901 | A | 418 | 1.00 | 1.00 | 1.00 | 0.33 |
| Females | ||||||
| rs524553 | T | 454 | 0.78 | 0.67 | 0.58 | 0.78 |
| rs542914 | A | 456 | 0.59 | 0.32 | 0.59 | 0.48 |
| rs648595 | G | 457 | 1.00 | 0.64 | 0.52 | 0.78 |
| rs6933870 | G | 457 | 0.32 | 0.45 | 0.55 | 0.22 |
| rs2397147 | C | 456 | 1.00 | 1.00 | 0.59 | 0.43 |
| rs17883901 | A | 392 | 0.58 | 0.78 | 0.67 | 0.06 |
| Significance of SNP-disease associations was assessed by adaptive permutations using the PLINK software, v.1.9. | ||||||
Table 3.
Genotype and allele frequencies of the GCLC gene in healthy controls and patients with psoriasis*.
Table 3.
Genotype and allele frequencies of the GCLC gene in healthy controls and patients with psoriasis*.
| SNP | Genotype/ allele |
Healthy Controls n (%)1 |
Patients with psoriasis n (%)1 |
OR2 (95% CI) | Pperm3 |
| Entire groups | |||||
| rs524553 | C/C | 273 (58.3) | 285 (60.5) | 0.67 (0.34-1.30) | 0.20R |
| C/T | 173 (37.0) | 171 (36.3) | |||
| T/T | 22 (4.7) | 15 (3.2) | |||
| T | 217 (23.2) | 201 (21.3) | 0.90 (0.72-1.12) | 0.36 | |
| rs542914 | C/C | 168 (35.8) | 174 (36.9) | 0.75 (0.52-1.08) | 0.11R |
| C/A | 227 (48.4) | 240 (50.9) | |||
| A/A | 74 (15.8) | 58 (12.3) | |||
| A | 375 (40.0) | 356 (37.7) | 0.91 (0.76-1.09) | 0.18 | |
| rs648595 | T/T | 147 (31.4) | 144 (30.4) | 0.75 (0.54-1.05) | 0.13R |
| T/G | 225 (48.1) | 252 (53.3) | |||
| G/G | 96 (20.5) | 77 (16.3) | |||
| G | 417 (44.6) | 406 (42.9) | 0.94 (0.78-1.12) | 0.21 | |
| rs6933870 | C/C | 160 (34.0) | 163 (34.5) | 0.93 (0.65-1.33) | 0.86R |
| C/G | 237 (50.4) | 240 (50.9) | |||
| G/G | 73 (15.5) | 69 (14.6) | |||
| G | 383 (40.7) | 378 (40.0) | 0.97 (0.81-1.17) | 0.99 | |
| rs2397147 | T/T | 183 (39.2) | 198 (41.9) | 0.90 (0.74-1.09) | 0.29A |
| T/C | 231 (49.5) | 230 (48.6) | |||
| C/C | 53 (11.3) | 45 (9.5) | |||
| C | 337 (36.1) | 320 (33.8) | 0.91 (0.75-1.09) | 0.48 | |
| rs17883901 | G/G | 334 (89.1) | 388 (89.2) | 0.43 (0.11-1.72) | 0.15R |
| G/A | 35 (9.3) | 44 (10.1) | |||
| A/A | 6 (1.6) | 3 (0.7) | |||
| A | 47 (6.3) | 50 (5.7) | 0.91 (0.60-1.38) | 0.63 | |
| Males | |||||
| rs524553 | C/C | 137 (58.5) | 152 (60.6) | 0.56 (0.23-1.38) | 0.20R |
| C/T | 84 (35.9) | 91 (36.2) | |||
| T/T | 13 (5.6) | 8 (3.2) | |||
| T | 110 (23.5) | 107 (21.3) | 0.88 (0.65-1.19) | 0.38 | |
| rs542914 | C/C | 81 (34.6) | 87 (34.7) | 0.75 (0.44-1.26) | 0.28R |
| C/A | 117 (50.0) | 134 (53.4) | |||
| A/A | 36 (15.4) | 30 (11.9) | |||
| A | 189 (40.4) | 194 (38.6) | 0.93 (0.72-1.20) | 0.55 | |
| rs648595 | T/T | 71 (30.5) | 78 (31.1) | 0.56 (0.35-0.90) | 0.017R |
| T/G | 110 (47.2) | 138 (55.0) | |||
| G/G | 52 (22.3) | 35 (13.9) | |||
| G | 214 (45.9) | 208 (41.4) | 0.83 (0.65-1.07) | 0.048 | |
| rs6933870 | C/C | 73 (31.2) | 87 (34.7) | 0.64 (0.38-1.06) | 0.09R |
| C/G | 120 (51.3) | 134 (53.4) | |||
| G/G | 41 (17.5) | 30 (11.9) | |||
| G | 202 (43.2) | 194 (38.6) | 0.83 (0.64-1.07) | 0.25 | |
| rs2397147 | T/T | 85 (36.5) | 101 (40.2) | 0.54 (0.30-0.98) | 0.05R |
| T/C | 116 (49.8) | 130 (51.8) | |||
| C/C | 32 (13.7) | 20 (8.0) | |||
| C | 180 (38.6) | 170 (33.9) | 0.81 (0.63-1.06) | 0.11 | |
| rs17883901 | G/G | 167 (89.3) | 204 (88.3) | 0.54 (0.09-3.24) | 0.33R |
| G/A | 17 (9.1) | 25 (10.8) | |||
| A/A | 3 (1.6) | 2 (0.9) | |||
| A | 23 (6.1) | 29 (6.3) | 1.02 (0.58-1.80 | 0.99 | |
| Females | |||||
| rs524553 | C/C | 136 (58.1) | 133 (60.5) | 0.91 (0.62-1.32) | 0.58D |
| C/T | 89 (38.0) | 80 (36.4) | |||
| T/T | 9 (3.8) | 7 (3.2) | |||
| T | 107 (22.9) | 94 (21.4) | 0.92 (0.67-1.25) | 0.78 | |
| rs542914 | C/C | 87 (37.0) | 87 (39.4) | 0.88 (0.68-1.15) | 0.32A |
| C/A | 110 (46.8) | 106 (48) | |||
| A/A | 38 (16.2) | 28 (12.7) | |||
| A | 186 (39.6) | 162 (36.7) | 0.88 (0.68-1.15) | 0.59 | |
| rs648595 | T/T | 76 (32.3) | 66 (29.7) | 1.13 (0.76-1.68) | 0.52D |
| T/G | 115 (48.9) | 114 (51.4) | |||
| G/G | 44 (18.7) | 42 (18.9) | |||
| G | 203 (43.2) | 198 (44.6) | 1.06 (0.82-1.37) | 0.99 | |
| rs6933870 | C/C | 87 (36.9) | 76 (34.4) | 1.37 (0.82-2.27) | 0.22R |
| C/G | 117 (49.6) | 106 (48.0) | |||
| G/G | 32 (13.6) | 39 (17.6) | |||
| G | 181 (38.3) | 184 (41.6) | 1.15 (0.88-1.49) | 0.32 | |
| rs2397147 | T/T | 98 (41.9) | 97 (43.7) | 1.29 (0.70-2.37) | 0.43R |
| T/C | 115 (49.1) | 100 (45) | |||
| C/C | 21 (9.0) | 25 (11.3) | |||
| C | 157 (33.5) | 150 (33.8) | 1.01 (0.77-1.33) | 0.99 | |
| rs17883901 | G/G | 167 (88.8) | 184 (90.2) | 0.30 (0.03-2.95) | 0.06R |
| G/A | 18 (9.6) | 19 (9.3) | |||
| A/A | 3 (1.6) | 1 (0.5) | |||
| A | 24 (6.4) | 21 (5.1) | 0.80 (0.44-1.45) | 0.58 | |
|
* The table shows the best genetic models for SNP-disease associations. 1 Absolute number and percentage of individuals/chromosomes with a particular genotype/allele. 2 Odds ratio with 95% confidence intervals (crude analysis) estimated for the best association model. 3 P-value estimated for the best association model through adaptive permutations. Superscripts denote SNP association models: R, recessive; D, dominant; A, additive. Bold depicts statistically significant P-values and odds ratios. | |||||
None of the polymorphisms was significantly associated with the risk of psoriasis in females.
Table 4.
Haplotypes of the GCLC gene and their association with psoriasis risk in the entire and sex-stratified groups
Table 4.
Haplotypes of the GCLC gene and their association with psoriasis risk in the entire and sex-stratified groups
| Haplotypes | SNP | Patients with psoriasis | Healthy Controls | Chi Square | P-value | |||||
| rs524553 | rs542914 | rs648595 | rs6933870 | rs2397147 | rs17883901 | |||||
| Entire groups | ||||||||||
| H1 | C | C | T | C | T | G | 0.482 | 0.463 | 0.635 | 0.426 |
| H2 | T | A | G | G | C | G | 0.154 | 0.162 | 0.192 | 0.661 |
| H3 | C | A | G | G | C | G | 0.121 | 0.128 | 0.186 | 0.666 |
| H4 | C | C | G | G | T | G | 0.056 | 0.043 | 1.681 | 0.195 |
| H5 | C | A | T | C | T | G | 0.043 | 0.042 | 0.017 | 0.898 |
| H6 | C | C | G | C | T | G | 0.032 | 0.027 | 0.422 | 0.516 |
| H7 | C | C | T | C | T | A | 0.019 | 0.026 | 0.940 | 0.332 |
| H8 | T | A | G | C | T | G | 0.017 | 0.027 | 2.247 | 0.134 |
| H9 | T | A | G | G | C | A | 0.023 | 0.020 | 0.133 | 0.715 |
| H10 | C | C | T | G | C | G | 0.018 | 0.018 | 0.016 | 0.900 |
| H11 | C | A | G | G | C | A | 0.010 | 0.013 | 0.301 | 0.583 |
| H12 | - | - | - | - | - | - | - | - | - | - |
| Males | ||||||||||
| H1 | C | C | T | C | T | G | 0.495 | 0.457 | 1.407 | 0.236 |
| H2 | T | A | G | G | C | G | 0.160 | 0.175 | 0.357 | 0.550 |
| H3 | C | A | G | G | C | G | 0.115 | 0.136 | 0.989 | 0.320 |
| H4 | C | C | G | G | T | G | 0.045 | 0.043 | 0.017 | 0.896 |
| H5 | C | A | T | C | T | G | 0.048 | 0.030 | 2.040 | 0.153 |
| H6 | C | C | G | C | T | G | 0.026 | 0.024 | 0.040 | 0.842 |
| H7 | C | C | T | C | T | A | 0.020 | 0.024 | 0.223 | 0.637 |
| H8 | T | A | G | C | T | G | 0.016 | 0.022 | 0.456 | 0.499 |
| H9 | T | A | G | G | C | A | 0.025 | 0.018 | 0.604 | 0.437 |
| H10 | C | C | T | G | C | G | 0.015 | 0.023 | 0.839 | 0.359 |
| H11 | C | A | G | G | C | A | 0.013 | 0.014 | 0.030 | 0.863 |
| H12 | - | - | - | - | - | - | - | - | - | - |
| Females | ||||||||||
| H1 | C | C | T | C | T | G | 0.463 | 0.464 | 0.001 | 0.981 |
| H2 | T | A | G | G | C | G | 0.150 | 0.158 | 0.109 | 0.741 |
| H3 | C | A | G | G | C | G | 0.130 | 0.119 | 0.243 | 0.622 |
| H4 | C | C | G | G | T | G | 0.069 | 0.045 | 2.445 | 0.118 |
| H5 | C | A | T | C | T | G | 0.037 | 0.052 | 1.155 | 0.283 |
| H6 | C | C | G | C | T | G | 0.034 | 0.027 | 0.418 | 0.518 |
| H7 | C | C | T | C | T | A | 0.023 | 0.028 | 0.321 | 0.571 |
| H8 | T | A | G | C | T | G | 0.018 | 0.031 | 1.511 | 0.219 |
| H9 | T | A | G | G | C | A | 0.017 | 0.017 | 0.010 | 0.919 |
| H10 | C | C | T | G | C | G | 0.021 | 0.013 | 0.761 | 0.383 |
| H11 | C | A | G | G | C | A | - | - | - | - |
| H12 | T | C | G | G | C | G | 0.010 | 0.010 | 0.001 | 0.983 |
| Estimation of haplotype frequencies and significance of haplotype-disease associations was done using the Haploview software, v.4.2. | ||||||||||
Table 5.
GCLC genotype combinations showed associations with psoriasis risk.
| Genotype combination | Patients | Controls | P-value | OR (95% CI)3 | ||
| n1 | %2 | n1 | %2 | |||
| Entire groups | ||||||
| rs542914-C/C × rs648595-G/T | 55 | 11.7 | 35 | 7.5 | 0.03 | 1.63 (1.04-2.54) |
| rs648595-G/G × rs6933870-C/G | 13 | 2.8 | 28 | 6.0 | 0.016 | 0.45 (0.23-0.87) |
| Males | ||||||
| rs524553-C/C ×rs648595-G/G | 7 | 2.8 | 18 | 7.7 | 0.025 | 0.36 (0.15-0.85) |
| rs524553-C/C× rs6933870-G/G | 6 | 2.4 | 15 | 6.4 | 0.05 | 0.37 (0.15-0.95) |
| rs542914-A/A × rs648595-G/G | 19 | 7.6 | 31 | 13.3 | 0.038 | 0.54 (0.30-0.98) |
| rs648595-G/G × rs17883901-G/G | 21 | 9.1 | 33 | 17.7 | 0.009 | 0.47 (0.26-0.84) |
| rs6933870-G/G × rs2397147-C/C | 20 | 8.0 | 32 | 13.7 | 0.042 | 0.55 (0.30-0.99) |
| rs6933870-G/G × rs17883901-G/G | 19 | 8.2 | 27 | 14.4 | 0.044 | 0.53 (0.29-0.99) |
| rs2397147-C/C ×rs17883901-G/G | 11 | 4.8 | 21 | 11.2 | 0.014 | 0.40 (0.19-0.85) |
| Females | ||||||
| rs6933870-G/G × rs17883901-G/G | 32 | 15.8 | 17 | 9.0 | 0.045 | 1.88 (1.01-3.52) |
|
1 Absolute number of individuals with particular genotype combination (minor alleles in genotypes are underlined). 2 Percentage of individuals with particular genotype combination. 3 OR, odds ratio; CI, confidence interval. Bold is statistically significant P-value after an adjustment for FDR of 0.05 (https://tools.carbocation.com/FDR). | ||||||
Table 6.
A summary of associations between GCLC gene polymorphisms and psoriasis risk in cigarette smoking- and alcohol abuse-stratified groups.
Table 6.
A summary of associations between GCLC gene polymorphisms and psoriasis risk in cigarette smoking- and alcohol abuse-stratified groups.
| SNP ID | Minor allele | Permutation P-values (Pperm) estimated for genetic models of SNP-disease associations | |||||||||
| N | Genetic models | N | Genetic models | ||||||||
| Allelic | Additive | Dominant | Recessive | Allelic | Additive | Dominant | Recessive | ||||
| Smokers | Non-smokers | ||||||||||
| rs524553 | T | 315 | 1.00 | 0.52 | 0.63 | 0.64 | 624 | 0.46 | 0.43 | 0.86 | 0.34 |
| rs542914 | A | 315 | 0.86 | 0.55 | 0.67 | 0.67 | 626 | 0.21 | 0.59 | 0.86 | 0.10 |
| rs648595 | G | 316 | 0.12 | 0.44 | 0.52 | 0.049 | 625 | 0.86 | 0.78 | 1.00 | 0.52 |
| rs6933870 | G | 315 | 0.65 | 0.52 | 0.86 | 0.59 | 627 | 1.00 | 1.00 | 0.86 | 0.67 |
| rs2397147 | C | 315 | 0.67 | 0.33 | 0.48 | 0.25 | 625 | 0.86 | 0.46 | 0.39 | 0.73 |
| rs17883901 | A | 275 | 0.24 | 0.16 | 0.09 | 1.00 | 535 | 0.18 | 0.11 | 0.18 | 0.002 |
| Alcohol abusers | Non-drinkers | ||||||||||
| rs524553 | T | 110 | 0.26 | 0.09 | 0.10 | NA | 580 | 0.24 | 0.08 | 0.20 | 0.15 |
| rs542914 | A | 112 | 0.11 | 0.053 | 0.06 | NA | 579 | 0.034 | 0.026 | 0.16 | 0.015 |
| rs648595 | G | 112 | 0.33 | 0.19 | 0.58 | NA | 580 | 0.05 | 0.04 | 0.26 | 0.03 |
| rs6933870 | G | 111 | 0.18 | 0.11 | 0.23 | NA | 581 | 0.29 | 0.14 | 0.18 | 0.27 |
| rs2397147 | C | 112 | 0.19 | 0.22 | 0.14 | NA | 579 | 0.20 | 0.09 | 0.14 | 0.25 |
| rs17883901 | A | 98 | 0.79 | NA | NA | NA | 498 | 0.55 | 0.48 | 0.67 | 0.09 |
| Significance of SNP-disease associations was assessed by adaptive permutations using the PLINK software, v.1.9. NA, not available. | |||||||||||
Table 7.
Replication of associations between the studied polymorphisms of the GCLC gene and psoriasis risk in a population of the UK Biobank1.
Table 7.
Replication of associations between the studied polymorphisms of the GCLC gene and psoriasis risk in a population of the UK Biobank1.
| Psoriasis phenotype2 | Variant | Eff, allele | beta | OR beta | P-value | MAF | HWE |
| psoriasis | rs524553 | T | 0.00030445 | 1.03 | 0.24054 | 0.248703 | 0.8257 |
| L40 Psoriasis | rs524553 | T | 0.00014144 | 1.03 | 0.43139 | 0.248703 | 0.8257 |
| psoriasis | rs542914 | A | 0.00031437 | 1.03 | 0.16739 | 0.409665 | 0.7591 |
| L40 Psoriasis | rs542914 | A | 0.00018358 | 1.03 | 0.24466 | 0.409665 | 0.7591 |
| psoriasis | rs648595 | G | 0.00034186 | 1.03 | 0.12131 | 0.485677 | 0.2804 |
| L40 Psoriasis | rs648595 | G | 0.00019125 | 1.04 | 0.21101 | 0.485677 | 0.2804 |
| psoriasis | rs6933870 | G | 0.00041555 | 1.04 | 0.062535 | 0.478105 | 0.1793 |
| L40 Psoriasis | rs6933870 | G | 0.00015391 | 1.03 | 0.3195 | 0.478105 | 0.1793 |
| psoriasis | rs2397147 | C | 0.00043391 | 1.04 | 0.057101 | 0.407803 | 0.6808 |
| L40 Psoriasis | rs2397147 | C | 0.000164 | 1.03 | 0.29943 | 0.407803 | 0.6808 |
| psoriasis | rs17883901 | G | -0.0001631 | 0.986 | 0.68281 | 0.0837 | 0.05018 |
| L40 Psoriasis | rs17883901 | G | -0.0002265 | 0.959 | 0.4129 | 0.0837 | 0.05018 |
|
1 The calculations were obtained from the Gene ATLAS web site (http://geneatlas.roslin.ed.ac.uk/). accessed by 28.04.2023 2 “Psoriasis” phenotype investigated in a cohort of 5175 cases and 447089 controls).: “L40 Psoriasis” phenotype investigated in a cohort of 2437 cases and 449827 controls; MAF. minor allele frequency; HWE. Hardy-Weinberg equilibrium P-value. | |||||||
Table 8.
Polymorphisms the GCLC gene showed significant associations (P≤0.05) with the risk of psoriasis in a population of the UK Biobank.
Table 8.
Polymorphisms the GCLC gene showed significant associations (P≤0.05) with the risk of psoriasis in a population of the UK Biobank.
| N | Variant | Position | Eff, Allele | Trait | Beta | P-value | MAF |
| Psoriasis phenotype: “psoriasis” (5175 cases and 447089 controls) | |||||||
| 1 | rs183555084 | 53463377 | A | psoriasis | 0.0054328 | 0.00048311 | 0.005415 |
| 2 | rs536001584 | 53491157 | A | psoriasis | 0.0069889 | 0.0037087 | 0.002296 |
| 3 | rs78863400 | 53507843 | G | psoriasis | 0.0020045 | 0.0049101 | 0.0245 |
| 4 | rs114919458 | 53478492 | A | psoriasis | 0.0020638 | 0.0077221 | 0.020874 |
| 5 | rs77162334 | 53473387 | A | psoriasis | 0.0015616 | 0.0084049 | 0.036438 |
| 6 | rs547541077 | 53524639 | A | psoriasis | 0.0083841 | 0.011327 | 0.001201 |
| 7 | rs55661362 | 53463674 | G | psoriasis | 0.0026909 | 0.012278 | 0.011041 |
| 8 | rs78331008 | 53489705 | G | psoriasis | 0.0014586 | 0.014341 | 0.036042 |
| 9 | rs115558853 | 53325654 | C | psoriasis | -0.0019831 | 0.016188 | 0.018765 |
| 10 | rs6902510 | 53493460 | T | psoriasis | -0.00052994 | 0.019054 | 0.405458 |
| 11 | rs62398116 | 53405203 | G | psoriasis | -0.0008575 | 0.019471 | 0.110262 |
| 12 | rs189491343 | 53341496 | G | psoriasis | -0.0019418 | 0.020189 | 0.01847 |
| 13 | rs7762921 | 53319569 | T | psoriasis | -0.00065758 | 0.021591 | 0.1828 |
| 14 | rs62398159 | 53490625 | A | psoriasis | -0.00051684 | 0.022213 | 0.406839 |
| 15 | rs56013020 | 53390696 | A | psoriasis | 0.00082793 | 0.022789 | 0.103296 |
| 16 | rs7739121 | 53510423 | C | psoriasis | -0.00049679 | 0.024997 | 0.467815 |
| 17 | rs72944719 | 53358473 | G | psoriasis | -0.0010805 | 0.025673 | 0.05524 |
| 18 | rs7761225 | 53315323 | C | psoriasis | -0.00064261 | 0.025687 | 0.179565 |
| 19 | rs6458936 | 53314296 | G | psoriasis | -0.00064298 | 0.025697 | 0.179334 |
| 20 | rs1914707 | 53311047 | G | psoriasis | -0.00063643 | 0.026486 | 0.181629 |
| 21 | rs563831 | 53327107 | G | psoriasis | 0.00063512 | 0.026486 | 0.183707 |
| 22 | rs4715409 | 53511015 | T | psoriasis | -0.00049168 | 0.02667 | 0.467022 |
| 23 | rs1518511 | 53313237 | C | psoriasis | -0.00063702 | 0.027149 | 0.179343 |
| 24 | rs6908614 | 53501678 | T | psoriasis | -0.00048943 | 0.027196 | 0.462606 |
| 25 | rs642103 | 53323152 | G | psoriasis | -0.00062689 | 0.028507 | 0.18174 |
| 26 | rs1914706 | 53311463 | T | psoriasis | -0.00062752 | 0.028627 | 0.181766 |
| 27 | rs72943672 | 53399516 | T | psoriasis | -0.00074715 | 0.028945 | 0.1182 |
| 28 | rs6933919 | 53313748 | G | psoriasis | -0.00062901 | 0.029059 | 0.179555 |
| 29 | rs4712030 | 53317469 | A | psoriasis | -0.00062437 | 0.029102 | 0.181758 |
| 30 | rs1467408 | 53351289 | A | psoriasis | -0.00052426 | 0.029222 | 0.361091 |
| 31 | rs9382209 | 53311804 | G | psoriasis | -0.00062389 | 0.02952 | 0.18191 |
| 32 | rs149644917 | 53519358 | A | psoriasis | -0.010751 | 0.029585 | 0.000499 |
| 33 | rs1401155 | 53312629 | C | psoriasis | -0.00062709 | 0.029593 | 0.17955 |
| 34 | rs9357769 | 53508264 | C | psoriasis | 0.00048131 | 0.029829 | 0.4664 |
| 35 | rs6908786 | 53494357 | A | psoriasis | -0.00047818 | 0.03092 | 0.466556 |
| 36 | rs587178 | 53325255 | T | psoriasis | 0.00061535 | 0.031491 | 0.182191 |
| 37 | rs6901352 | 53500138 | C | psoriasis | -0.0004754 | 0.031615 | 0.466514 |
| 38 | rs6908860 | 53494615 | T | psoriasis | -0.00047638 | 0.031652 | 0.464814 |
| 39 | rs681682 | 53440021 | C | psoriasis | -0.0072738 | 0.032871 | 0.001361 |
| 40 | rs543473 | 53439524 | T | psoriasis | -0.0072796 | 0.032941 | 0.001359 |
| 41 | rs681585 | 53439958 | G | psoriasis | -0.0072742 | 0.033023 | 0.00136 |
| 42 | rs9474608 | 53505134 | A | psoriasis | -0.00047139 | 0.033072 | 0.466612 |
| 43 | rs681635 | 53439987 | A | psoriasis | -0.0072632 | 0.033272 | 0.001359 |
| 44 | rs2397146 | 53360119 | A | psoriasis | -0.00053256 | 0.033642 | 0.273716 |
| 45 | rs607285 | 53326491 | T | psoriasis | 0.00060766 | 0.033745 | 0.182155 |
| 46 | rs62416866 | 53398370 | A | psoriasis | -0.00077679 | 0.033936 | 0.100838 |
| 47 | rs742528 | 53360191 | A | psoriasis | -0.00052981 | 0.034548 | 0.273993 |
| 48 | rs623928 | 53335695 | T | psoriasis | 0.00061135 | 0.034551 | 0.180506 |
| 49 | rs629162 | 53326283 | G | psoriasis | 0.00060422 | 0.034685 | 0.182369 |
| 50 | rs676637 | 53335353 | C | psoriasis | 0.00061072 | 0.03473 | 0.180538 |
| 51 | rs624432 | 53335555 | G | psoriasis | 0.00061046 | 0.034804 | 0.180555 |
| 52 | rs642625 | 53333732 | T | psoriasis | 0.00061027 | 0.034833 | 0.180511 |
| 53 | rs618033 | 53339289 | T | psoriasis | 0.00061046 | 0.034957 | 0.180357 |
| 54 | rs600722 | 53332887 | T | psoriasis | 0.00060973 | 0.034961 | 0.180513 |
| 55 | rs631783 | 53338531 | A | psoriasis | 0.00060876 | 0.035396 | 0.180454 |
| 56 | rs619955 | 53338845 | T | psoriasis | 0.00060877 | 0.035396 | 0.180457 |
| 57 | rs485371 | 53341627 | T | psoriasis | 0.00060874 | 0.035527 | 0.180356 |
| 58 | rs12196344 | 53457292 | A | psoriasis | -0.00048763 | 0.036061 | 0.404087 |
| 59 | rs9367538 | 53506487 | G | psoriasis | -0.00046273 | 0.036479 | 0.466245 |
| 60 | rs7764361 | 53492467 | C | psoriasis | 0.00046427 | 0.037421 | 0.456163 |
| 61 | rs663087 | 53342704 | T | psoriasis | 0.00060223 | 0.037659 | 0.180217 |
| 62 | rs646403 | 53347484 | T | psoriasis | 0.00059431 | 0.040381 | 0.180136 |
| 63 | rs12194171 | 53464937 | C | psoriasis | 0.00046011 | 0.041523 | 0.3968 |
| 64 | rs11756739 | 53316777 | A | psoriasis | 0.0029885 | 0.04429 | 0.006094 |
| 65 | rs4712031 | 53320273 | G | psoriasis | -0.00056517 | 0.04448 | 0.190022 |
| 66 | rs2092421 | 53473076 | A | psoriasis | -0.00045072 | 0.045589 | 0.398208 |
| 67 | rs4269374 | 53461179 | G | psoriasis | -0.00044872 | 0.04647 | 0.397012 |
| 68 | rs9349679 | 53470507 | A | psoriasis | -0.00044669 | 0.047497 | 0.39642 |
| 69 | rs34997452 | 53518439 | T | psoriasis | -0.0027868 | 0.047543 | 0.006643 |
| 70 | rs10807461 | 53472150 | T | psoriasis | -0.00044608 | 0.047762 | 0.398057 |
| 71 | rs738472 | 53477038 | C | psoriasis | -0.00045789 | 0.048043 | 0.353182 |
| 72 | rs6458946 | 53472830 | T | psoriasis | -0.00044442 | 0.048672 | 0.397982 |
| 73 | rs114749455 | 53489865 | G | psoriasis | 0.0022206 | 0.048751 | 0.0103 |
| 74 | rs2143399 | 53461749 | A | psoriasis | -0.00044292 | 0.049341 | 0.397029 |
| 75 | rs74357476 | 53476523 | T | psoriasis | 0.0014009 | 0.050596 | 0.025311 |
| Psoriasis phenotype: “L40 Psoriasis” (2437 cases and 449827 controls) | |||||||
| 1 | rs185956124 | 53496212 | C | L40 Psoriasis | 0.0026856 | 0.0036274 | 0.00747649 |
| 2 | rs547541077 | 53524639 | A | L40 Psoriasis | 0.0065265 | 0.0044446 | 0.00120121 |
| 3 | rs189622943 | 53509408 | T | L40 Psoriasis | 0.0035101 | 0.0095446 | 0.00341133 |
| 4 | rs183043870 | 53509634 | G | L40 Psoriasis | 0.0035128 | 0.0095673 | 0.00341141 |
| 5 | rs78735978 | 53360036 | C | L40 Psoriasis | 0.0012576 | 0.015913 | 0.0231714 |
| 6 | rs41271287 | 53370147 | T | L40 Psoriasis | 0.0011902 | 0.018795 | 0.0236652 |
| 7 | rs17215384 | 53510321 | T | L40 Psoriasis | 0.00039365 | 0.02118 | 0.28084 |
| 8 | rs77516417 | 53373662 | A | L40 Psoriasis | -0.001175 | 0.021204 | 0.02313 |
| 9 | rs574202 | 53481989 | G | L40 Psoriasis | 0.00035417 | 0.021427 | 0.489829 |
| 10 | rs12661112 | 53486714 | A | L40 Psoriasis | 0.00037194 | 0.021838 | 0.343991 |
| 11 | rs563699 | 53479410 | C | L40 Psoriasis | 0.00035124 | 0.022359 | 0.490659 |
| 12 | rs558026 | 53478773 | A | L40 Psoriasis | 0.00035803 | 0.022979 | 0.392597 |
| 13 | rs583513 | 53477688 | T | L40 Psoriasis | 0.00034605 | 0.024098 | 0.491525 |
| 14 | rs7759126 | 53484485 | C | L40 Psoriasis | 0.00035645 | 0.028105 | 0.343339 |
| 15 | rs12665537 | 53509452 | G | L40 Psoriasis | 0.00035343 | 0.030008 | 0.33107 |
| 16 | rs67228890 | 53511814 | G | L40 Psoriasis | 0.00034794 | 0.034841 | 0.327456 |
| 17 | rs74449072 | 53521238 | G | L40 Psoriasis | 0.00061918 | 0.039019 | 0.0749875 |
| 18 | rs7764361 | 53492467 | C | L40 Psoriasis | 0.00031881 | 0.039179 | 0.456163 |
| 19 | rs9382225 | 53511696 | T | L40 Psoriasis | -0.00033714 | 0.039962 | 0.328914 |
| 20 | rs5020412 | 53349885 | C | L40 Psoriasis | 0.00084548 | 0.041197 | 0.0354 |
| 21 | rs4715412 | 53511836 | T | L40 Psoriasis | -0.00033138 | 0.044253 | 0.328611 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



