
Submitted:
08 May 2023
Posted:
09 May 2023
You are already at the latest version
Abstract
Foxtail millet (Setaria italica (L.) P. Beauv) is an important food and forage crop that is well adapted to nutrient-poor soils. However, little is known about how foxtail millet adapts to low nitrogen (LN) at the physiological and molecular levels. In the present study, two foxtail millet varieties with contrasting low nitrogen (LN) tolerance properties were investigated through integrative analyses of physiological featureparameters and transcriptomics. The physiological results showed that JG20 (high tolerance to LN) had higher biomass accumulation, nitrogen content and nitrogen use efficiency compared with JG22 (low tolerancesensitive to LN) under LN. JG20 had higher soluble sugar and soluble protein concentration in shoots compared with JG22 under LN, while contcentrations of soluble sugar and soluble protein contents were higher in the roots of JG22. Higher levels of CTK contentzeatin concentration were found in the shoots of JG20 com-pared with JG22, and a higher ABA contcentration was found in both the shoots and roots of JG22 compared with JG20 under LN. In the transcriptomics results, JG20 had more differentially ex-pressed genes (DEGs) than JG22 both in shoots and roots in response to LN. These LN-responsive genes were enriched in glycolysis, starch and sucrose metabolism, photosynthesis, biosynthesis of amino acids, hormone metabolism and nitrogen metabolism. Furthermore, the high-affinity nitrate transporter gene, SiNRT2.1, and glutamine synthetase gene, SiGS5, chlorophyll apoprotein of photosystem II gene, SiPsbQ, ATP synthase subunit gene, b, auxin-responsive protein gene, SiIAA25, and aldose 1-epimerase gene, SiAEP, in shoots, and high-affinity nitrate transporter gene, SiNRT2.3, SiNRT2.4, and glutamate synthase gene, SiGOGAT2, auxin-responsive protein gene, SiIAA4, fructose-bisphosphate aldolase gene, SiAEP5, in root, were important genes in-volved in the LN tolerance of foxtail millet. These results provide a detailed description of the physiological and transcriptome response of foxtail millet under LN condition.
Keywords:
foxtail millet 1
; nitrogen use efficiency 2
; varieties3
; physiological parameters 4
; transcriptomics 5.
1. Introduction
Nitrogen is an important and essential macronutrient for plant growth and development [1, 2], and is part of all building blocks of life, including nucleic acids, amino acids, proteins, lipids and metabolic products [3, 4]. Nitrogen fertilizer, as important input factor for crop production, has played an important role in increasing crop yield in recent decades [5, 6]. However, the cost of nitrogen fertilizer accounts for a large proportion of agricultural input [7]. It has been reported that no more than 40% of the applied N is taken up by crops, and most of the applied N is retained in the soil and lost to the atmosphere, groundwater and rivers through volatilization, leaching, and surface runoff [8]. The adverse side effects of nitrogen application has seriously damaged the environment [8-10]. Therefore, decreasing the nitrogen application and increasing nitrogen use efficiency (NUE) is necessary for agricultural sustainability [11]. NUE comprises two components: nitrogen uptake efficiency (NUpE) and nitrogen utilization efficiency (NUtE) [12]. Many studies have shown that there exists significant genetic difference in NUE and its two components [13-16]. NUE refers to dry matter produced by plants absorbed per unit mass of nitrogen and results from the coordination of carbon and nitrogen metabolism [17]. Nitrogen metabolism includes N absorption [6, 18], mainly including nitrate (NO3−) and ammonium (NH4+) under different soil conditions [6]; then, the absorbed nitrogen is assimilated through nitrate reductase (NR), nitrite reduction (NiR), glutamine synthetase (GS) and glutamine-2-oxoglutarate aminotransferase (GOGAT) cycle processes [6]. The metabolic products are then converted into different nitrogen-containing compounds for carbon metabolism, including photosynthesis, phytohormones and fatty acid biosynthesis [19-21]. These processes can be controlled by numerous genes. Therefore, a better understanding of the underlying mechanism behind nitrogen uptake, transport, and assimilation would be helpful in providing a theoretical basis for improving NUE [22].
Foxtail millet [Setaria italica (L.) P. Beauv] is one of the oldest crops in the world [23], and has high drought tolerance and resistance to infertility, and has been widely cultivated in Asia for food and fodder [24, 25]. It is now used as a model species for genomics and basic biological process due to its attractive qualities, including small diploid genome (2n = 18, ~ 420 Mb), short life cycle, prolific seed production, and C4 photosynthesis [24, 26]. It was reported that foxtail millet can perform root thickening to improve N uptake under low-nitrogen conditions [27]. Transcription profiling by RNA-Seq is a successful and widely used approach to explore molecular aspects of nutrient stress [22]. Several studies have reported that this method is widely used for investigating the mechanism of low N stress tolerance on wheat [28], rice [29], rapeseed [22], barley [30]. A previous study also explored potential regulatory factors and functional key genes in foxtail millet in response to low nitrogen [31]. However, our previous study had shown that there existed significant genotypic variations among different foxtail millet varieties for NUE [32]. The molecular mechanism controlling genotypic variations between different foxtail millet varieties with contrasting low-nitrogen tolerance are still unknown. Meanwhile, understanding physiological and molecular mechanism of foxtail millet in response to long-term low nitrogen stress should be close to actual production and provide a more meaningful theoretical basis. Therefore, two foxtail millet varieties—Jigu20 (JG20) and Jigu22 (JG22)—with contrasting NUE that were identified in our previous research were used to compare the differences between these two varieties in terms of physiological and transcriptomic changes in response to long-term nitrogen stress. Our results reveal the molecular mechanism of foxtail millet in response to chronic low nitrogen and identifies genes associated with high low nitrogen (LN) tolerance, which could assist in the genetic improvement of NUE in foxtail millet.
2. Results
2.1. Phenotypic Characteristics and N accumulation of Foxtail Millet
Significant differences between JG20 and JG22 were detected with respect to the growth and biomass of seedlings under low-nitrogen conditions (Figure 1A,B). The shoot and root biomass of both varieties were significantly decreased under low-nitrogen conditions compared with control (Figure 1A,B). The LN-sensitive variety JG22 had higher shoot and root biomass than JG20 under a sufficient nitrogen supply, whereas the shoot and root biomass of LN-tolerant JG20 were significantly higher than that of JG22 under low-nitrogen conditions (Figure 1A,B). The N concentration and N content of the two varieties decreased significantly under low-nitrogen conditions compared with control (Figure 2A). The N concentrations of shoots in the LN-sensitive variety JG22 were higher than in the LN-tolerant variety JG20 under low-nitrogen conditions, yet N content showed the inverse effect, where the LN-tolerant variety JG20 had higher N content in the shoots than the LN-sensitive variety JG22 under LN conditions (Figure 2B), which was attributed to it having a high biomass under LN (Figure 1B). Nitrogen use efficiency (NUE) varied significantly between the two varieties (Figure 2C). The NUE of shoots and roots were equal in the two varieties under normal N supply, whereas, the LN-tolerant variety JG20 had higher NUE both in shoots and roots compared with JG22 under low-nitrogen conditions (Figure 2C).
2.2. Physiological Characteristics and Response of Foxtail Millet to Low Nitrogen
Low nitrogen had no significant effects on the concentration of soluble protein except that in the shoots of the LN-tolerant variety JG20, which was significantly increased by LN compared with control (Figure 3A). The soluble protein concentration of shoots in the LN-tolerant variety JG20 was higher than that in the LN-sensitive variety JG22, whereas the soluble protein concentration of the roots in JG20 was lower than in JG22 under low-nitrogen conditions (Figure 3A). The free amino acid concentrations of shoots and roots in JG20 and JG22 were both significantly reduced under low nitrogen compared with control (Figure 3B), this is caused by the synthesis of protein from amino acid to keep normal protein concentration to maintain metabolism. Comparison of free amino acid concentration between the two varieties was consistent with the soluble protein concentration under low-nitrogen conditions; the LN-tolerant variety JG20 had higher free amino acid concentration in shoots and lower free amino acid concentration in roots compared with the LN-sensitive variety JG22 under low-nitrogen conditions (Figure 3B). LN had different effects on the nitrate nitrogen concentration in the shoots and roots of the two varieties; the nitrate nitrogen concentration in the shoots of the two varieties was significantly increased, while in roots, it was significantly reduced under LN conditions compared with control (Figure 3C).
Carbon metabolism is significantly affected by LN. The response of soluble sugar concentration to LN varied between the two varieties, LN had significantly increased soluble sugar concentration in shoots and had no effect on soluble sugar concentration in the roots of JG20 compared with control (Figure 4A), whereas the soluble sugar concentration in the shoots was significantly reduced, and in the roots significantly increased in JG22 under LN conditions compared with control (Figure 4A). Sucrose concentrations in shoots and roots in the two varieties were increased by LN, except that in the shoots of JG22, which was invariable between control and LN (Figure 4B). Sucrose concentration in the shoots of JG20 was higher than that in the shoots of JG22 under LN, while sucrose concentration in the roots of JG20 was lower than that in the roots of JG22 under LN (Figure 4B).
Hormones including indole-3-acetic acid (IAA), cytokinin (zeatin) and abscisic acid (ABA) varied significantly under LN and between varieties. Indole-3-acetic acid contents of the shoots and roots in the two varieties were both significantly reduced under LN compared with control (Figure 4C). There was no significant difference in the IAA content in the shoots between the two varieties under LN. IAA concentration in the roots in JG22 was higher than that in the roots of JG20 under LN (Figure 4C). LN had significantly reduced zeatin concentration in the shoots and roots of the two varieties, except zeatin concentration in the shoots of JG22, which was invariable between control and LN (Figure 4D). Zeatin concentration in the shoots of JG20 was higher than that in the shoots of JG22 under LN, while zeatin concentration in the roots was undifferentiated between JG20 and JG22 under LN (Figure 4D). ABA concentration was reduced in both the shoots and roots of the two varieties under LN, except ABA concentration in the shoots of JG20, which was significantly increased under LN compared with control (Figure 4E). JG22 had higher ABA concentration in both the shoots and roots than the concentration in JG20 under LN (Figure 4E).
2.3. Differentially Expressed Genes (DEGs) between JG20 and JG22 under Low Nitrogen.
The RNA-Seq was performed using the shoot and root samples of JG20 and JG22 seedlings, which were named LJG20-CK, RJG20-CK, LJG22-CK, and RJG22-CK for under control, and LJG20-LN, RJG20-LN, LJG22-LN and RJG22-LN under low-nitrogen conditions, respectively (Table 1). Using three independent biological replicates of each sample, a total twenty-four samples were constructed and sequenced using the Solexa/Illumina platform. A total of 161 GB of raw bases were generated from the transcriptome libraries of the 24 samples, and after filtering out adapter and low-quality sequences, approximately 150GB (95.39%) of clean reads, representing an average of 43.6 million clean reads per sample, were obtained (Table 1). On average, over 86.5% reads were mapped to the Setaria italic reference genome and over 84.5% reads were mapped to unique regions (Table 1).
To verify the RNA-Seq data, 15 DEGs regulated in response to low-nitrogen conditions were randomly selected for RT-qPCR validation in LN-tolerant variety JG20(Figure 5A) and LN-sensitive variety JG22 (Figure 5B). The result showed that they had similar expression patterns and there was a high correlation coefficient of 0.936 (R2) between RNA-Seq and RT-qPCR (Figure 5C), which demonstrated that the RNA-Seq data were reliable and repeatable.
A comparison of DEGs among the varieties in response to LN conditions facilitates an understanding of the regulatory mechanisms associated with LN and helps to identify the roles of independent varieties in the regulation. Of these DEGs, a total of 4,256 (JG20LN vs. JG20CK) and 1,291 (JG22LN vs. JG22CK) genes in shoots and 3,191 (JG20LN vs. JG20CK) and 1,881 (JG22LN vs. JG22CK) genes in roots were differentially expressed in LN-tolerant JG20 and LN-sensitive JG22 foxtail millet varieties, respectively, in response to LN conditions, which showed that the LN-tolerant variety JG20 had more DEGs than the LN-sensitive variety JG22, in both shoots and roots, in response to LN conditions (Figure 6A,C, Supplementary Table S2). Meanwhile, 3,581 (JG22CK vs. JG20CK) and 1,132 (JG22LN vs. JG20LN) genes in shoots and 3,852 (JG22CK vs. JG20CK) and 2,028 (JG22LN vs. JG20LN) genes in roots were differentially expressed between the two different varieties under control and LN conditions (Figure 6A,C, Supplementary Table S2). In the combined LN-responsive and genotype-specific DEGs datasets, more than 60.7% (4,362) of DEGs in the shoots and 64.8% (5,133) in the roots were unique to one dataset, 35.7% (2,560) of DEGs in the shoots and 32.3% (2,556) in the roots were shared by two datasets, 3.4% (242) of DEGs in the shoots and 2.6% (209) in the roots were shared by three datasets, and only 0.2% (13) of DEGs in the shoots and 0.3% (20) in the roots were expressed in all four datasets (Figure 6B,D). The intersecting DEGs in JG20LN vs. JG20CK and JG22LN vs. JG22CK were 662 and 694 in the shoots and roots, respectively, among which the number of DEGs in response to LN without considering variance in genotype was only 494 in the shoots and 557 in the roots (Figure 6B,D).
2.4. Functional Categorization of DEGs in Response to Low Nitrogen in Two Foxtail Millet Varieties
The DEGs in response to low nitrogen were mainly enriched in terms of biological process (BP) and molecular function (MF) through gene ontology (GO) enrichment analysis in the two foxtail millet varieties, except for the cellular components in the shoots of the LN-tolerant variety JG20 (Figure 7). The biosynthetic and metabolic processes of carbohydrates, including the metabolic and biosynthetic processes of cellulose, polysaccharide, and glucan, were the top GO biological process (GOBP) subcategories in the shoots of JG20LN vs. JG20CK (Figure 7A). The GO subcategories of the cellular component (GOCC) were mainly focused on the extracellular region and the photosystem Ⅰ, while the GO subcategories of molecular function (GOMF) mainly included transferase activity, cellulose synthase activity, and hydrolase activity in the shoots of JG20LN vs. JG20CK (Figure 7A). All GO terms contained more down- than up-regulated genes in the shoots of LN-sensitive JG22 in response to low nitrogen, where nucleoside metabolic and carbohydrate catabolic processes were the top GO subcategories for biological processes (GOBP), while the main GO subcategories of molecular function (GOMF) were phosphofructokinase activity, kinase activity, and binding in the shoots of JG22LN vs. JG22CK (Figure 7B). The gene ontology (GO) enrichment analysis showed different GO terms in the root of two foxtail millet varieties in response to low nitrogen. The more down-regulated genes were enriched in the GO subcategories of biological process (GOBP), mainly focusing on nucleotide biosynthetic and metabolic processes, while transmembrane transporter activity, binding and inhibitor activity were the main GO subcategories of molecular function (GOMF) in the roots of JG20 in response to low nitrogen (Figure 7C). The biosynthetic process and transmembrane transport of trehalose were the top of GO subcategories of biological process (GOBP), while binding and transport activity were the main GO subcategories of molecular function (GOMF) in the roots of JG22LN vs. JG22CK (Figure 7D).
In shoots, a total of 680 and 236 DEGs were assigned to 119 and 94 KEGG pathways in JG20LN vs. JG20CK and JG22LN vs. JG22CK, respectively, while 529 and 397 DEGs were assigned to 111 and 107 KEGG pathways in the roots of JG20LN vs. JG20CK and JG22LN vs. JG22CK, respectively. The top 20 KEGG pathways in the four comparisons are shown in Figure 8. The significant KEGG pathways in the shoots of JG20 were related to starch and sucrose metabolism, phenylpropanoid biosynthesis, and amino acid biosynthesis and metabolism (Figure 8A), while biosynthesis of amino acids and saccharide metabolism, including glycolysis, gluconeogenesis, and fructose and mannose metabolism were the main KEGG pathways in the shoots of JG22 (Figure 8B), which showed that DEGs of the LN-tolerant variety JG20 were not only enriched in amino acid metabolism, but also in starch and sucrose metabolism, in order to improve nitrogen use in response to low nitrogen. The KEGG pathways in roots were different from those in shoots, and varied between varieties. The DEGs in the roots of LN-tolerant JG20 were significantly assigned to carbon metabolism, transporters, and amino acid metabolism (Figure 8C) in response to LN, and plant–pathogen interaction, MAPK signaling pathway, and hormone metabolism were the main KEGG pathways in the roots of LN-sensitive JG22 under LN conditions (Figure 8D).
2.5. Gene Expression Profiling of Genes Involved in N Source Transport and Assimilation in Response to Low Nitrogen
The expression patterns of genes involved in N transport and assimilation in response to LN are profiled in Figure 9 (A,B,C). The nitrogen metabolism process is shown in the flowchart, and contains two parts: nitrogen uptake and transport; and nitrogen assimilation (Figure 9A). The transcript levels of genes involved in N uptake and transport varied more drastically in roots than in shoots suffering from low nitrogen in both varieties (Figure 9B). The transcription of genes NRT2.1d, NRT2.1e, NRT2.3 and NRT2.4 increased significantly in the shoots of the LN-tolerant variety JG20, whereas that in the LN-sensitive variety JG22 was invariable or slightly decreased in response to low-nitrogen conditions (Figure 9B). Compared with NRT genes in the shoots, the transcription of NRT genes was more up-regulated in roots, and varied among varieties; for example, NRT2.1a, NRT2.3 and NRT2.4 were significantly up-regulated in JG20, while NRT2.1d and NRT2.1e were more significantly up-regulated in JG22 in response to low nitrogen (Figure 9B). The expression of AMT genes varied more significantly between the two varieties than NRT genes, and the expression of AMT1.1 and AMT1.2 were both significantly increased in the shoots of the two varieties in response to LN, while AMT3.2 was uniquely up-regulated in the shoots of the LN-tolerant variety JG20 in response to LN (Figure 9B). The AMT3.1 and AMT3.2 genes showed significant increases in expression in the roots of both foxtail millet varieties under N-starved conditions, while AMT1.1 and AMT2.1 showed significantly increased expression only in the roots of JG22 in response to LN (Figure 9B).
Nitrogen assimilation was coordinated by expression of genes involved in NR, NiR, GS, GOGAT and GDH. The expression of NR and NiR varied significantly between the two foxtail millet varieties, which were up-regulated in both the shoots and roots of JG22, except for NiR in shoots, which was significantly increased in the roots of JG22. Menwhile, the expressions of NR and NiR were down-regulated in the shoots of JG20, especially for NR (NADH) (Figure 9C). The expression of genes involved in GS-GOGAT (GDH) differed among different parts and varieties. GS5, GOGAT (X3) and GOGAT2 (NADH) were significantly up-regulated, while GOGAT1 (NADH) and GDH were significantly down-regulated in the shoots of JG20 in response to LN (Figure 9C). GS1.1 showed increased expression, and GS1.3 exhibited reduced expression in the roots of both foxtail millet varieties in response to LN, which indicated that they were independent of the genotype. The expression of gene GOGAT1 (NADH) was significantly decreased in the roots of both foxtail millet varieties, while those of GOGAT2 (NADH) and GDH (X2) were significantly up-regulated in the roots of JG20 in response to LN (Figure 9C).
2.6. Gene Expression Profiling of Genes Involved in Photosynthesis, Hormone Signal Transduction and Glycolysis in Response to Low Nitrogen
Photosynthesis and glycolysis are two important pathways participating in carbon metabolism. The overall gene expression of the photosystem I (PSI), photosystem II (PS II), F-type ATPase and light-harvesting chlorophyll protein complex (LHC) are suppressed under the N limitation in shoots of the two varieties, with the exception of genes of PsbQ and b which are up-regulated in the shoot of JG20 (Figure 10A, Supplementary Table S5).
Plant hormones play an important role in regulating the growth and development. In the current study, the expression of genes involved in signal transduction pathways associated with several plant hormones are altered in response to LN (Figure 10B), suggesting that hormone pathways play critical roles in the shoot and root of foxtail millet in response to nitrogen availability. Genes associated with auxin-responsive protein SAUR and indole-3-acetic acid-amido synthetase GH3 are mostly down-regulated in two varieties, especially in shoot of two varieties (Figure 10B, Supplementary Table S6), which could explain the decreased IAA content in two varieties under LN condition. Downregulation of cytokinine histidine-containing phosphotransfer protein AHP under LN is detected in roots, while it is up-regulated in shoots of two varieties (Figure 10B, Supplementary Table S6), which could be key gene regulating the decreased cytokinine content in shoot. Abscisic acid receptor gene PYR/PYL is up-regulated in shoots of two varieties under LN (Figure 10B, Supplementary Table S6), this may cause increased ABA content in shoots of two varieties under LN (Figure 10B). Other genes involved in ethylene synthesis are both down-regulated in shoots and roots of two varieties under LN (Figure 10B, Supplementary Table S6).
Glycolysis is one of important primary metabolic pathways in plants, which can supply energy and carbon skeletons for other metabolism pathways. In shoot, downregulation of most DEGs under LN are detected, while TIM, PGK, PCK and ALDH are up-regulated in two varieties, especially in JG20 (Figure 10C, Supplementary Table S7). AEP, ATP-PFK, PDH-E2 and DLD are up-regulated under LN in roots of two varieties (Figure 10C, Supplementary Table S7).
2.7. Schematic Representation of the Main Processes and Genes Involved in Foxtail Millet in Response to Low Nitrogen
The most relevant processes and important genes related to NUE were obtained in two foxtail millet varieties (Figure 11). Nitrogen transfer genes ammonium transporters (SiAMTs) and nitrate transporters (SiNRTs) are both upregulated in root of two varieties, while they are only upregulated in shoot of LN-tolerant variety JG20, and downregulated in LN-sensitive variety JG22. Nitrogen assimilation genes glutamine synthetase (SiGS) and glutamate synthase (SiGOGAT) are upregulated in shoot of two varieties, especially in JG20 in response to LN. Nitrogen uptake and utilization efficiency are strongly associated with hormone signal, upregulated hormone signal transduction-related genes including auxin, cytokinine and abscisic acid, such as AUX/IAA, AFR, AHP, B-ARR, A-ARR, PYR/ PYL in shoot and AUX/IAA, GH3, SAUR, AHP, A-ARR, PP2C, SnRK2, ABF in root are found in LN-tolerant variety JG20 in response to LN. Carbon metabolism including photosynthesis and glycolysis significantly affect nitrogen use efficiency, which provides a carbon source and energy for nitrogen metabolism. Chlorophyll apoprotein of photosystem II and F-type ATPase genes such as PsbQ and b are upregulated in LN-tolerant variety JG20 in response to LN. In the pathway of glycolysis, AEP, ATP-PFK, FBA, TIM, PGK, ALDH, ENO and PGMP are upregulated in shoot of LN-tolerant variety JG20 and downregulated in shoot of LN-sensitive variety JG22 in response to LN. In root of two varieties, ATP-PFK are up-regulated, while PGMP, FBA and PCK genes are downregulated.
2.8. Expression Inventories of LN-Responsive TFs in Different Foxtail Millet Varieties
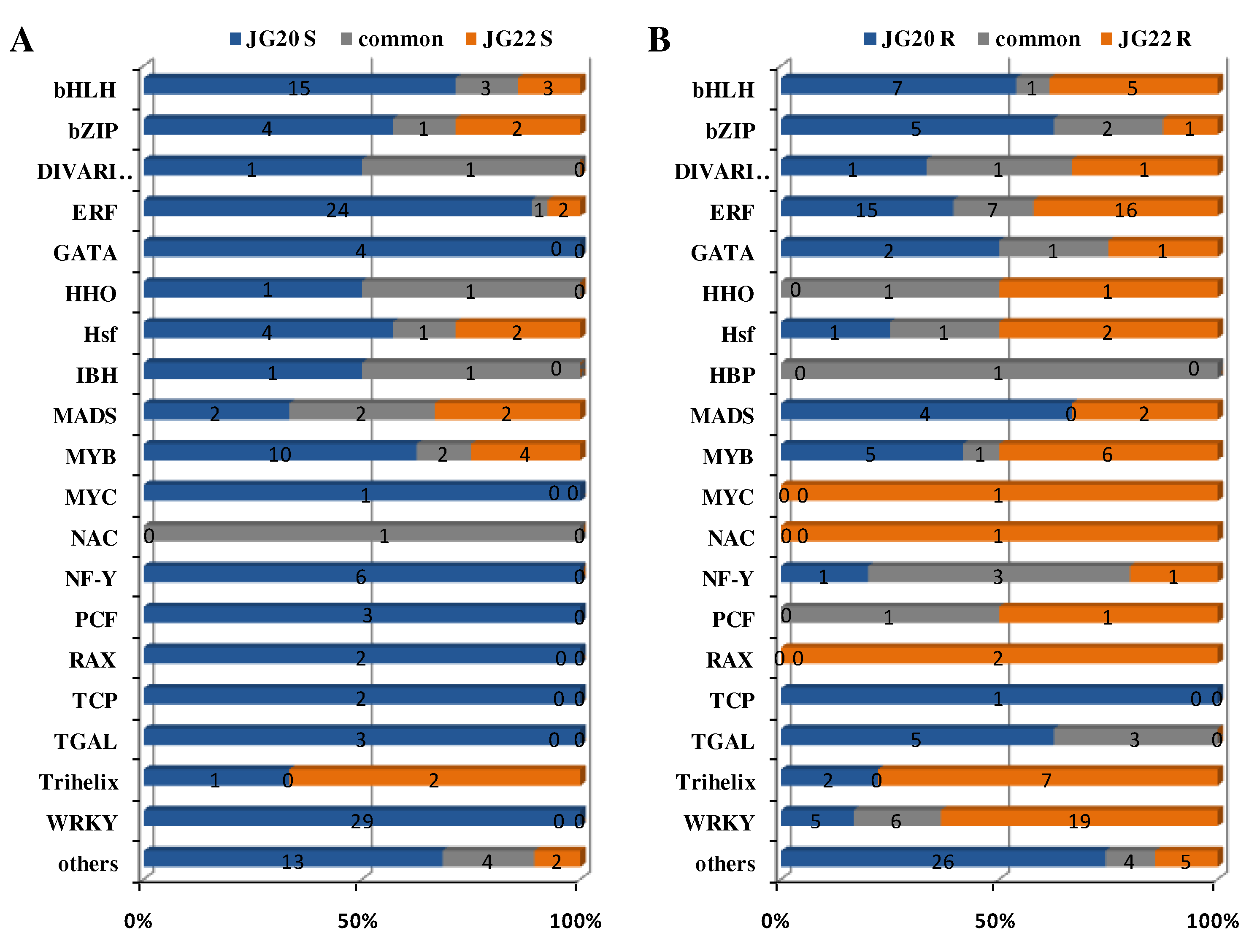
TFs play an important role in mediating plant growth by regulating various stress-inducible genes. In total, 144 and 37 differentially expressed TFs were detected in the shoots of JG20 and JG22, and 113 and 105 differentially expressed TFs were detected in the roots of JG20 and JG22 under low-nitrogen conditions, and these were mainly distributed among 20 TFs families (Figure 10). The number of differentially expressed TFs in the shoots of the LN-tolerant variety JG20 was higher than that in the shoots of LN-sensitive variety JG22 (Figure 10A), and TFs belonging to the WRKY (20.1%), ethylene-responsive transcription factor (EFR; 17.4%), bHLH (12.5%) and MYB (8.3%) families were the most abundant in the shoots of the LN-tolerant variety JG20, while the MYB (16.2%), bHLH (16.2%) and MADS (10.8%) families were the top TFs in the shoots of the LN-sensitive variety JG22, while the most common TFs in the shoots of the two foxtail millet varieties belonged predominantly to the bHLH (16.7%), MYB (11.1%) and MADS (11.1%) families (Figure 10A, Supplementary Table S3). The EFR and WRKY families accounted for the main differentially expressed TFs in the roots of both foxtail millet varieties that were common between them, with proportions corresponding to 19.5%, 21.9%, 21.2% and 9.7%, 18.2%, 23.8% in JG20, JG22 and common between them, respectively (Figure 10B, Supplementary Table S3).
Figure 10.
Changes in transcription factor (TF) expression in different foxtail millet varieties. (A) Distribution of TF families in the shoots of JG20 and JG22 in response to low nitrogen. (B) Distribution of TF families in the roots of JG20 and JG22 in response to low nitrogen.
Figure 10.
Changes in transcription factor (TF) expression in different foxtail millet varieties. (A) Distribution of TF families in the shoots of JG20 and JG22 in response to low nitrogen. (B) Distribution of TF families in the roots of JG20 and JG22 in response to low nitrogen.
3. Discussion
Nitrogen, as a key nutrient element for plant growth and development, plays a vital role in agricultural production. A lot of studies have shown that large genotypic variation of NUE exists in different crops [13-15, 44]. Significant variation of NUE was also shown among different foxtail millet varieties in our previous research [32, 45]. Foxtail millet has high infertility tolerance, especially under low-nitrogen conditions, and is considered to be an environmentally friendly crop [24]. Therefore, it is important to understand the underlying regulatory mechanism of NUE in foxtail millet. In the present study, two previously selected foxtail millet varieties with contrasting NUE were used to identify the physiological and molecular mechanisms when responding to long-term low-nitrogen conditions, with the aim of providing more practical and meaningful theories for improving the NUE of foxtail millet.
Consistent with studies in other crops inhibited by long-term LN [46-48], both the shoot and root growths of foxtail millet were restrained under long-term LN conditions in the present study (Figure 1A,B), but this varied between varieties. The LN-tolerant variety JG20 had a superior shoot and root system compared to the LN-sensitive variety JG22 under long-term LN, indicating that JG20 had higher LN tolerance under long-term conditions. It has been reported that LN reduces foxtail millet growth, resulting in a considerably shorter root system and an increased root/shoot ratio [27], which was verified in the LN-sensitive variety JG22 under LN conditions in the present study (Figure 1A, Supplementary Figure S1); however, the LN-tolerant variety JG20 had a longer root system and a decreased root/shoot ratio (Figure 1A, Supplementary Figure S1), indicating that the foxtail millet variety with higher nitrogen use had developed a longer root system as an obvious morphological response to N deficiency and increased the ratio of dry matter accumulation in the shoots under long-term LN.
The N concentration and N content of two foxtail millet varieties were both significantly decreased in response to long-term low nitrogen (Figure 2A,B), which was consistent with previous research. The N concentrations in the shoots and roots of the LN-sensitive variety JG22 was higher than in those of the LN-tolerant variety JG20 (Figure 2A), while the N contents in the shoots and roots of the LN-sensitive variety JG22 were lower than in those of the LN-tolerant variety JG20 under LN conditions (Figure 2B). This indicated that the LN-tolerant variety could use low N nitrogen concentrations to accumulate a large amount of dry matter in both shoots and roots, as is reflected in the high NPFP (Figure 2C). As unexpected, the soluble protein of shoots and roots in the two varieties under LN conditions was equivalent to the control, and even the soluble protein of shoots in the LN-tolerant variety JG20 increased under LN conditions (Figure 3A), which has also been found in early stages in maize ear and in foxtail millet root [27, 49]. This may indicate that foxtail millet could maintain normal soluble protein level to sustain various metabolic activities in response to LN, especially for the LN-tolerant variety. The free amino acid concentrations of the two varieties were both decreased in shoots and roots under LN (Figure 3B), which was consistent with the previous findings in foxtail millet. The decrease of free amino acid can explain the equivalent soluble protein being synthesized by amino acid under LN condition. The LN-tolerant variety JG20 had higher free amino acid concentration in shoots, while it had lower free amino acid concentration in roots under LN conditions compared with control, indicating that the high-N-use foxtail millet variety can allocate more free amino acids to the shoot to ensure the N demand of photosynthetic apparatuses and other processes under low-nitrogen conditions, which was consistent, and was verified by the fact that the nitrate concentration as a nitrogen source was significantly increased in the shoots and decreased in the roots in both foxtail millet varieties in response to long-term low nitrogen (Figure 3C).
Hormones play an important role in regulating plant development, growth, and adaptation to environmental stress [50]. Auxin regulates cell division, elongation, and differentiation during plant development and growth, and is also important in N signaling [51, 52]. In the present study, the auxin concentration (IAA) and zeatin concentrations were significantly lower in the shoots and roots under LN (Figure 4C,D), leading to smaller shoot and root systems under LN (Figure 1A,B). This is consistent with findings in maize [49] and in foxtail millet [27]. ABA accumulation could be an essential hormonal regulatory mechanism for N limitation, and can restrain growth in foxtail millet [27]. ABA concentration increased in the shoots of the LN-tolerant variety JG20 and decreased in the roots of both varieties (Figure 4E). When comparing the two varieties, the LN-sensitive variety JG22 had higher ABA concentrations in both shoots and roots compared with the LN-tolerant variety JG20, which could explain why the LN-tolerant variety was able to develop well under LN conditions.
To advance our understanding of the molecular mechanisms responding to long-term LN in foxtail millet, we performed a comparative transcriptional analysis between the two varieties with contrasting NUE using RNA-Seq. More than 10,400 DEGs were identified that responded to LN for the two varieties. The analysis of expression patterns during long-term N-deficient conditions between the two varieties revealed an abundant diversity in gene expression, indicating that these stress-responsive genes may play functional roles in the N stress tolerance of foxtail millet. The LN-tolerant variety had a greater number of DEGs than the LN-sensitive variety in response to long-term LN, indicating that the LN-tolerant variety was able to activate more responding genes, thus having a relatively stronger ability to adapt to N deficiency than the LN-sensitive variety. Among these DEGs, only a small number of DEGs (515 DEGs in the shoots and 602 DEGs in the roots) were represented in both varieties, without considering the variance in genotype with respect to response to LN, which showed that there existed large differences in molecular genotypic variation in response to LN.
The interaction between the metabolisms of C and N determines plant growth and development [53], and soluble sugars such as sucrose, glucose and fructose, which function as an energy source in plants, ply an important role in various physiological and biological processes, including growth, development and stress response [54]. In this research, genes encoding several enzymes in the metabolism of starch, sucrose and glycolysis were significantly induced in response to LN in the two foxtail millet varieties. A large number of genes encoding soluble sugars such as beta-glucosidase, glucanotransferase, and sucrose-phosphatase were up-regulated in both the shoots and roots of the two varieties (Supplementary Table S4), and the numbers of up-regulated genes in the shoots of the LN-tolerant variety JG20 were significantly higher than in the LN-sensitive variety JG22 (Supplementary Table S4), which is consistent with the increased soluble sugar and sucrose concentration in the shoots of JG20 under LN (Figure 4A,B). The ratio of up-regulated genes to total DEGs in the roots of JG22 was higher than that in the roots of JG20, which was verified by the higher soluble sugar and sucrose concentrations in the roots of JG22 compared with in JG20 (Figure 4A,B). In shoot, downregulation of most DEGs under LN are detected, while TIM, PGK, PCK and ALDH are up-regulated in two varieties, especially in JG20 (Figure 10C, Supplementary Table S7). AEP, ATP-PFK, PDH-E2 and DLD are up-regulated under LN in roots of two varieties (Figure 10C, Supplementary Table S7). This indicated that enhancing the synthesis and accumulation of soluble sugars, and glycolysis metabolism in shoots under LN could be an important trait for high-nitrogen-use foxtail millet, and an important pathway for improving LN tolerance in foxtail millet.
A large number of studies have shown that the synthesis of plant hormones is tightly regulated in response to stresses [50, 55], and genes encoding hormones have been reported to be regulated by LN stress [56]. Genes encoding indole-3-acetic acid synthesis and zeatin, such as indole-3-acetic acid-amido synthetase, auxin-responsive protein, and histidine-containing phosphotransfer protein, were down-regulated in both varieties in response to N deficiency (Supplementary Table S6), which was consistent with the decrease in indole-3-acetic acid and zeatin concentration under LN (Figure 4C,D). The ABA concentrations in both varieties were increased in response to LN (Figure 4E), which could be explained by the up-regulated genes of abscisic acid receptor in foxtail millet in response to LN (Supplementary Table S6).
Nitrogen metabolism, comprising both N acquisition and transport, and N assimilation, was more directly affected by LN. Nitrate transporters and ammonium transporters are two important transport systems for nitrogen uptake [57]. In the current research, high-affinity nitrate transporters genes NRT2.3 and NRT2.4 were only up-regulated in the shoots and roots of the LN-tolerant variety JG20 in response to long-term LN, and ammonium transporter gene AMT3.2 was only up-regulated in the shoots of the LN-tolerant variety JG20 under N deficiency, whereas that in the LN-sensitive variety JG22 was consistent or slightly decreased in response to low-nitrogen conditions (Figure 9B). This indicated that high-affinity nitrate transporter genes NRT2.3 and NRT2.4, as well as ammonium transporter gene AMT3.2, may play important roles in high nitrogen transport in the LN-tolerant foxtail millet variety in response to N deficiency. The GS/GOGAT cycle, as the main pathway of nitrogen assimilation [58, 59], is considered to be an important potential maker for selecting high-NUE genotypes in wheat [60]. In the present result, GS5, GOGAT (X3) and GOGAT2 (NADH) were significantly up-regulated in the shoots of JG20 in response to LN (Figure 9C), which showed that these genes can play a vital role in nitrogen assimilation in LN-tolerant foxtail millet in response to LN. Glutamate dehydrogenase (GDH) can catalyze the exchange between NH4+ and glutamate, and play a small role in nitrogen assimilation [11, 61] (Figure 9A). The GDH genes in the shoots of the LN-tolerant variety JG20 was significantly down-regulated under LN, which is consistent with the findings in high-NUE wheat [11]. This showed that nitrogen assimilation in the high-NUE genotype may be more dependent on the GS/GOGAT cycle.
TFs regulating the expression of other genes in metabolic pathways [62] have been reported to be involved in the regulation of nitrogen metabolism [22,63]. It has been reported that MYB-like TFs play an important role in response to low-nitrogen stress in foxtail millet [31]. In the present study, lots of TFs were involved in regulating metabolism in response to LN, and there existed differentially expressed patterns of TFs in both varieties (Figure 10A). TFs belonging to the WRKY (20.1%), ethylene-responsive transcription factor (EFR; 17.4%), bHLH (12.5%) and MYB (8.3%) families were the most abundant in the shoots of the LN-tolerant variety JG20, while those belonging to the MYB (16.2%), bHLH (16.2%) and MADS (10.8%) families were the top TFs in the shoots of the LN-sensitive variety JG22 (Figure 10A, Supplementary Table S3), which indicated that WRKY and EFR TFs may play vital roles in regulating the responses of foxtail millet to N deficiency for the LN-tolerant foxtail millet variety. bHLH (16.7%), MYB (11.1%) and MADS (11.1%) families in shoots and EFR (21.2%) and WRKY (23.8%) families in roots were the most commonly expressed TFs in both varieties, showing that these TFs play roles in roots that are independent of foxtail millet genotype.
4. Materials and Methods
4.1. Materials and Experimental Design
Two foxtail millet varieties that were identified in our previous research were used in this experiment. Jigu20 (JG20), a low nitrogen (LN) tolerant variety, and Jigu22 (JG22), an LN sensitive variety, have contrasting biomass and NUE under LN conditions. They were both bred by the crop research institute, Shandong Academy of Agricultural Science, have different parents and belong to different genotypes. The experiment was conducted in a rain-proof shed in the crop research institute, Shandong Academy of Agricultural Science, in Ji’nan city, in the province of Shandong, China.
The experiment was designed with a completely randomized design, with two nitrogen levels—0.2 mmol L-1 (low nitrogen, LN) and 6 mmol L-1 (normal nitrogen, CK)—and a total of four treatments: Jigu20 under normal nitrogen condition (JG20-CK), Jigu20 under low-nitrogen conditions (JG20-LN), Jigu22 under normal nitrogen condition (JG22-CK), Jigu22 under low-nitrogen conditions (JG22-LN). Seeds of two foxtail millet varieties were sown on 20 June 2020 in a plastic pot with the following dimensions: 150mm diameter × 130mm height, with a permeable bottom. The pots were filled with 3kg purified sand substrate of 1.28 g∙cm-3. Each treatment has three replicate pots, each pot was sown with 100 seed density and the seedlings were thinned to a density of 10 plants per pot according to the normal foxtail millet production density at five days after emergence. Plants were watered once with modified Hoagland nutrition [32] at 7 days after seeding with two N levels (0.2 and 6 mmol L-1) every two days. Before spraying the nutrient solution, the pots were always watered enough with purified water to wash off the residual nutrient element.
The plants were harvested at 15 days after the treatment; half of the plants in each pot were grouped in dry samples, which were separated into shoots and roots, and were dried at 105℃ for 30 min and then to constant weight at 75℃. The dry samples were used for weighing the biomass and determining the nitrogen content. Fresh samples of shoots and roots were immediately frozen in liquid nitrogen and then stored at -80℃, before being taken for determination of the soluble protein concentration, free amino acid content, nitrate nitrogen concentration, soluble sugar concentration, sucrose concentration, hormone content, gene expression level and RNA-seq analysis.
4.2. Measurement of NUE
The biomass of the shoots and roots was weighed to calculate the NUE, using the following equation [33]: NUE =
4.3. Physiological Measurements
The N concentration of the plants was determined using the Kjeldahl method. The N content of plants was calculated using the dry matter content multiplied by the N concentration. Soluble proteins were extracted and analyzed using a standard kit with bovine serum albumin [34]. Total free amino acid concentration was extracted using 10% (v/v) acetic acid and was measured with the ninhydrin colorimetric method at 750nm using leucine as standard [27]. Nitrate nitrogen was extracted with boiling water for 30 min, and nitrate concentration was determined using the nitrosalicylic acid colorimetric method at 410nm [35]. Hormones including endogenous IAA, ZA and ABA were quantified using a High-Performance liquid Chromatography (HPLC) system [36, 37].
4.4. cDNA Library Contruction and Sequencing for RNA-Seq
cDNA library construction and sequencing for RNA-seq were based on methods described in our previous research [38]. The total RNA was extracted from samples using Trizol® (Invitrogen, Carlsbad, CA, USA). The RNA Nano 6000 Assay Kit for the Bioanalyzer 2100 system (Agilent Technologies, Santa Clara, CA, USA) and a NanoPhotometer spectrophotometer (IMPLEN, Westlake Village, CA, USA) were used to assess the integrity and purity of RNA, respectively. A total amount of 1 μg high-quality RNA per sample was used for the RNA sample preparations. Sequencing libraries were generated using NEBNext® UltraTM RNA Library Prep Kit for Illumina® (NEB, Ipswich, MA, USA), following the manufacturer’s recommendations. Sequencing of the constructed cDNA libraries was carried out at Novogene Bioinformatics Technology Co. Ltd. (Beijing, China).
Raw data of fastq format were initially processed to remove reads containing adapters, reads containing poly N, and low-quality sequence reads (>50% bases with Q-values ≤ 20), and the Q20, Q30, GC contents, and sequence duplication of the clean data were analyzed. Then, the clean reads were aligned with the foxtail millet genome (https://www.ncbi.nlm.nih.gov/genome/?term= Setaria italica) using HISAT2 v2.0.5 to identify relevant sequences. The mapped reads of each sample were assembled by StringTie (v1.3.3b) using a reference-based approach [39].
Gene expression levels were calculated based on FRKM (Fragments Per Kilobase of transcript sequence per Millions base pairs sequenced) value [40]. Differentially expressed genes (DEGs) between the two treatments were identified using DESeq2 R package (1.20.0). Threshold values of FDR (the false discovery rate) ≤0.05 and absolute log2fold-change ≥1 were applied to judge the significance of differences in gene expression levels [41, 42].
Gene ontology (GO) and Kyoto encyclopedia of genes and genomes (KEGG) analysis for the DEGs were conducted using cluster Profiler R package. GO terms with corrected p-value < 0.05 were considered significantly enriched by differentially expressed genes and the top 20 KEGG pathways were selected.
4.5. Validation of DEGs Using qRT-PCR
To indentify the reliability of the RNA-Seq results, the expression of 15 randomly selected DEGs was examined by qRT-PCR analysis using SYBR Premix Ex Taq (Clontech Takara, Shiga, Japan) on a 7500 Real-Time PCR System machine (Applied Biosystems, Foster City, CA, USA) [38]. qRT-PCR data were standardized with SiActin (gene ID: 101779009) as an internal reference. The gene-specific primes are presented in Supplementary Table S1. The gene expression of qRT-PCR was calculated using relative expression to the transcription level of SiActin in each sample using the 2-△△CT method [43].
4.6. Statistical Analysis
Means and standard errors were calculated using the data from three independent samples. Analyses of variance were carried out using SPSS18.0 (SPSS 18.0, SPSS Inc., USA) and the least significant difference method (LSD) was used to test the differences for biomass, N content, NUE, and physiological parameters between control and LN treatments of two foxtail millet varieties. Significance was specified at the level of P<0.05.
5. Conclusions
In summary, using our integrative analyses of the physiological and transcriptomics datasets, on the basis of the current results, it was found that the LN-tolerant foxtail millet variety had greater biomass accumulation, nitrogen content, and NUE, which were attributed to its having a high metabolism capacity as a result of having high soluble sugar, soluble protein and CTK concentrations, as well as low ABA concentrations, in the shoots under LN. This was related to a vast number of genes associated with high NUE in LN-tolerant foxtail millet. These genes are involved in nitrogen uptake and assimilation, photosynthesis, starch and sucrose metabolism, glycolysis, hormone metabolism, and TFs. In addition, some new candidate genes related to LN tolerance were identified, which could be useful for developing foxtail millet varieties with enhanced NUE.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: The root/shoot ratio in two foxtail millet varieties under control and low-nitrogen conditions. ; Table S1: The gene-specific primes for selected DEGs; Table S2: Differentially expressed genes (DEGs) of two foxtail millet varieties under control and low-nitrogen conditions. ; Table S3: Number and percentage of differentially expressed TFs between two different foxtail millet varieties in response to LN condition. ; Table S4: The DEGs related to starch and sucrose metabolism in two varieties. ; Table S5: The DEGs related with Plant hormone signal transduction in two varieties.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, E.C. and Y.G.; methodology, E.C. and Y.G.; software, E.C.; validation, E.C. and Y.G.; formal analysis, E.C.; investigation, E.C., B.Y., Z.L., X.Y. and J.N.; resources, E.C. and Y.G.; data curation, E.C., L.Q., F.L. and R.W.; writing—original draft preparation, E.C.; writing—review and editing, E.C., L.Q., F.L., and Y.G.; visualization, H.Z., H.W. and B.L.; supervision, Y.G.; project administration, E.C. and Y.G.; funding acquisition, E.C. and Y.G.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (32272226); Shandong Provincial Natural Science Foundation (ZR2022MC052); China Agriculture Research System of MOF and MARA (CARS-06-14.5-A19); Key R&D Program of Shandong Province, China (2022TZXD0029); the Agricultural Fine Seed Project of Shandong Province (2021LZGC006).
Data Availability Statement
The datasets generated and analyzed during the current study are available in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) database (http://www.ncbi.nlm.nih.gov/sra/) (Accession number: PRJNA838221). Additional supporting tables are included as Supplementary Data. All plant materials are available from the Crop Research Institute, Shandong Academy of Agricultural Sciences, P. R. China.
Acknowledgments
We are thankful for funding from National Natural Science Foundation of China (32272226); Shandong Provincial Natural Science Foundation (ZR2022MC052); China Agriculture Research System of MOF and MARA (CARS-06-14.5-A19); Key R&D Program of Shandong Province, China (2022TZXD0029); the Agricultural Fine Seed Project of Shandong Province (2021LZGC006).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study.
References
- Weber:, K.; Burow, M. Nitrogen – essential macronutrient and signal controlling flowering time. Physiol. Plant. 2018, 162, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, B.; Chu, C. Nitrogen use efficiency in crops: lessons from Arabidopsis and rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Cramer, M.D. Root Nitrogen Acquisition and Assimilation. Plant Soil. 2005, 274, 1–36. [Google Scholar] [CrossRef]
- Meharg, A. Marschner's Mineral Nutrition of Higher Plants., 3rd ed.; Elsevier/Academic: Amsterdam, Netherlands, 2011; p. 684. [Google Scholar]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in Agriculture: Balancing the Cost of an Essential Resource. Annu. Rev. Env Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.; Alvarez, J.; Araus, V.; Riveras, E.; Brooks, M.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.; Coruzzi, G.; Gutierrez, R. Nitrate 2020: Thirty years from transport to signaling networks. Plant Cell. 2020, 32, 2094–2119. [Google Scholar] [CrossRef] [PubMed]
- Sebilo, M.; Mayer, B.; Nicolardot, B.; Pinay, G.; Mariotti, A. Long-term fate of nitrate fertilizer in agricultural soils. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 18185–18189. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; de Vries, W.; Liu, X.; Hao, T.; Zeng, M.; Shen, J.; Zhang, F. Enhanced acidification in Chinese croplands as derived from element budgets in the period 1980–2010. Sci. Total Environ. 2018, 618, 1497–1505. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, X.; Vitousek, P. Chinese agriculture: An experiment for the world. Nature 2013, 497, 33–35. [Google Scholar] [CrossRef]
- Zhang, X.; Li, F.; Ding, Y.; Ma, Q.; Yi, Y.; Zhu, M.; Ding, J.; Li, C.; Guo, W.; Zhu, X. Transcriptome Analysis of Two Near-Isogenic Lines with Different NUE under Normal Nitrogen Conditions in Wheat. Biology. 2021, 10, 1126. [Google Scholar] [CrossRef]
- Han, M.; Okamoto, M.; Beatty, P.H.; Rothstein, S.J.; Good, A.G. The Genetics of Nitrogen Use Efficiency in Crop Plants. Annu. Rev. Genet. 2015, 49, 269–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Ding, G.; Li, L.; Cai, H.; Ye, X.; Zou, J.; Xu, F. Identification and characterization of improved nitrogen efficiency in interspecific hybridized new-type Brassica napus. Ann. Bot. 2014, 114, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wang, W.; Ou, S.; Tang, J.; Li, H.; Che, R.; Zhang, Z.; Chai, X.; Wang, H.; Wang, Y.; Liang, C.; Liu, L.; Piao, Z.; Deng, Q.; Deng, K.; Xu, C.; Liang, Y.; Zhang, L.; Li, L.; Chu, C. Variation in NRT1. 1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar] [PubMed]
- Stahl, A.; Vollrath, P.; Samans, B.; Frisch, M.; Wittkop, B.; Snowdon, R.J. Effect of breeding on nitrogen use efficiency-associated traits in oilseed rape. J. Exp. Bot. 2019, 70, 1969–1986. [Google Scholar] [CrossRef] [PubMed]
- Bingham, I.J.; Karley, A.J.; White, P.J.; Thomas, W.T.B.; Russell, J.R. Analysis of improvements in nitrogen use efficiency associated with 75 years of spring barley breeding. Eur. J. Agron. 2012, 42, 49–58. [Google Scholar] [CrossRef]
- Palenchar, P.M.; Kouranov, A.; Lejay, L.V.; Coruzzi, G.M. Genome-wide patterns of carbon and nitrogen regulation of gene expression validate the combined carbon and nitrogen (CN)-signaling hypothesis in plants. Genome Biol. 2004, 5, R91. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Beeckman, T.; Xu, G. Plant nitrogen nutrition: sensing and signaling. Curr. Opin. Plant Biol. 2017, 39, 57–65. [Google Scholar] [CrossRef]
- Baslam, M.; Mitsui, T.; Sueyoshi, K.; Ohyama, T. Recent Advances in Carbon and Nitrogen Metabolism in C3 Plants. Int. J. Mol. Sci. 2021, 22, 318. [Google Scholar] [CrossRef]
- Shen, T.; Xiong, Q.; Zhong, L.; Shi, X.; Cao, C.; He, H.; Chen, X. Analysis of main metabolisms during nitrogen deficiency and compensation in rice. Acta Physiol. Plant. 2019, 41, 68. [Google Scholar] [CrossRef]
- Krouk, G. Hormones and nitrate: a two-way connection. Plant Mol. Biol. 2016, 91, 599–606. [Google Scholar] [CrossRef]
- Yang, N.; Li, S.; Wang, S.; Li, Q.; Xu, F.; Shi, L.; Wang, C.; Ye, X.; Cai, H.; Ding, G. Dynamic transcriptome analysis indicates extensive and discrepant transcriptomic reprogramming of two rapeseed genotypes with contrasting NUE in response to nitrogen deficiency. Plant Soil. 2020, 456, 369–390. [Google Scholar] [CrossRef]
- Yang, X.; Wan, Z.; Perry, L.; Lu, H.; Wang, Q.; Zhao, C.; Li, J.; Xie, F.; Yu, J.; Cui, T.; Wang, T.; Li, M.; Ge, Q. Early millet use in northern China. Proc. Natl. Acad. Sci. U.S.A. 2012, 109, 3726–3730. [Google Scholar] [CrossRef] [PubMed]
- Xianmin, D.; Schnable, J.; L, B.; Li, J. Initiation of Setaria as a model plant. Front. Agric. Sci. Eng. 2014, 1, 16. [Google Scholar] [CrossRef]
- Tang, S.; Li, L.; Wang, Y.; Chen, Q.; Zhang, W.; Jia, G.; Zhi, H.; Zhao, B.; Diao, X. Genotype-specific physiological and transcriptomic responses to drought stress in Setaria italica (an emerging model for Panicoideae grasses). Sci. Rep. 2017, 7, 10009. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, H.; Li, X.; Shen, H.; Gao, J.; Hou, S.; Zhang, B.; Mayes, S.; Bennett, M.; Ma, J.; Wu, C.; Sui, Y.; Han, Y.; Wang, X. A mini foxtail millet with an Arabidopsis-like life cycle as a C4 model system. Nat. Plants. 2020, 6, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, F.; Ahmad, Z.; Wang, R.; Han, J.; Shen, Q.; Chang, F.; Diao, X.; Zhang, F.; Li, X. Foxtail Millet [Setaria italica (L.) Beauv.] Grown under Low Nitrogen Shows a Smaller Root System, Enhanced Biomass Accumulation, and Nitrate Transporter Expression. Front. Plant. Sci. 2018, 9, 205. [Google Scholar]
- Zhang, X.; Ma, Q.; Li, F.; Ding, Y.; Yi, Y.; Zhu, M.; Ding, J.; Li, C.; Guo, W.; Zhu, X. Transcriptome Analysis Reveals Different Responsive Patterns to Nitrogen Deficiency in Two Wheat Near-Isogenic Lines Contrasting for Nitrogen Use Efficiency. Biology. 2021, 10, 1126. [Google Scholar] [CrossRef]
- Wang, R.; Qian, J.; Fang, Z.; Tang, J. Transcriptomic and physiological analyses of rice seedlings under different nitrogen supplies provide insight into the regulation involved in axillary bud outgrowth. BMC Plant Biol. 2020, 20, 197. [Google Scholar] [CrossRef]
- Quan, X.; Zeng, J.; Chen, G.; Zhang, G. Transcriptomic analysis reveals adaptive strategies to chronic low nitrogen in Tibetan wild barley. BMC Plant Biol. 2019, 19, 68. [Google Scholar] [CrossRef]
- Ge, L.; Dou, Y.; Li, M.; Qu, P.; He, Z.; Liu, Y.; Xu, Z.; Chen, J.; Chen, M.; Ma, Y. SiMYB3 in Foxtail Millet (Setaria italica) Confers Tolerance to Low-Nitrogen Stress by Regulating Root Growth in Transgenic Plants. Int. J. Mol. Sci. 2019, 20, 5741. [Google Scholar] [CrossRef]
- Erying, C.; Ling, Q.; Yanbing, Y.; Huawen, Z.; Hailian, W.; Bin, L.; Shuting, Y.; Runfeng, W.; Yanan, G. Variability of nitrogen use efficiency by foxtail millet cultivars at the seedling stage. Pesqui. Agropecu. Bras. 2020, 55, 00832. [Google Scholar] [CrossRef]
- Kant, S.; Bi, Y.-M.; Rothstein, S.J. Understanding plant response to nitrogen limitation for the improvement of crop nitrogen use efficiency. J. Exp. Bot. 2011, 62, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Sprent, J.I.; Raven, J.A.; Eady, P.E. Relationships between shoot to root ratio, growth and leaf soluble protein concentration of Pisum sativum, Phaseolus vulgaris and Triticum aestivum under different nutrient deficiencies. Plant, Cell Environ. 2010, 22, 949–958. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid Colorimetric Determination of Nitrate in Plant-Tissue by Nitration of Salicylic-Acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Luo, Y.; Li, W.; Huang, C.; Yang, J.; Jin, M.; Chen, J.; Pang, D.; Chang, Y.; Li, Y.; Wang, Z. Exogenous abscisic acid coordinating leaf senescence and transport of assimilates into wheat grains under drought stress by regulating hormones homeostasis. Crop J. 2021, 9, 901–914. [Google Scholar] [CrossRef]
- Li, W.; Zhang, M.; Qiao, L.; Chen, Y.; Zhang, D.; Jing, X.; Gan, P.; Huang, Y.; Gao, J.; Liu, W.; Shi, C.; Cui, H.; Li, H.; Chen, K. Characterization of wavy root 1, an agravitropism allele, reveals the functions of OsPIN2 in fine regulation of auxin transport and distribution and in ABA biosynthesis and response in rice (Oryza sativa L.). Crop J. 2022, 12, 004. [Google Scholar] [CrossRef]
- Qin, L.; Chen, E.; Li, F.; Yu, X.; Ying, R. Genome-Wide Gene Expression Profiles Analysis Reveal Novel Insights into Drought Stress in Foxtail Millet (Setaria italica L.). Int. J. Mol. Sci. 2020, 21, 1–21. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods. 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Robinson, M.; Oshlack, A. Robinson MD, Oshlack A. . A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol 11: R25. Genome Biol. 2010, 11, R25. [Google Scholar]
- Yoav, B.; Daniel, Y. The control of the false discovery rate in multiple testing under dependency. Ann Stat. 2001, 29, 1165–1188. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Riche, A.B. Impacts of G x E x M on Nitrogen Use Efficiency in Wheat and Future Prospects. Front. Plant. Sci. 2020, 11, 1157. [Google Scholar] [CrossRef] [PubMed]
- Erying, C.; Yanbing, Y.; Bingwen, C.; Ling, Q.; Huawen, Z.; Bin, L.; Hailian, W.; Guiling, C.; Yanan, G. Yield and nitrogen use efficiency in summer foxtail millet. Soil Fert. Sci. China. 2015, 93–97. [Google Scholar]
- Chun, L.; Mi, G.; Li, J.; Chen, F.; Zhang, F. Genetic Analysis of Maize Root Characteristics in Response to Low Nitrogen Stress. Plant Soil. 2005, 276, 369–382. [Google Scholar] [CrossRef]
- Tian, Q.; Chen, F.; Liu, J.; Zhang, F.; Mi, G. Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots. J. Plant Physiol. 2008, 165, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.U.N.; Chen, F.; Yuan, L.; Zhang, F.; Mi, G. A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress. Plant, Cell Environ. 2015, 38, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Han, J.; Wang, R.; Li, X. Down-regulation of nitrogen/carbon metabolism coupled with coordinative hormone modulation contributes to developmental inhibition of the maize ear under nitrogen limitation. Planta. 2016, 244, 111–124. [Google Scholar] [CrossRef]
- Wolters, H.; Jürgens, G. Survival of the flexible: hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 2009, 10, 305–317. [Google Scholar] [CrossRef]
- Jones, B.; Gunnerås, S.A.; Petersson, S.V.; Tarkowski, P.; Graham, N.; May, S.; Dolezal, K.; Sandberg, G.; Ljung, K. Cytokinin Regulation of Auxin Synthesis in Arabidopsis Involves a Homeostatic Feedback Loop Regulated via Auxin and Cytokinin Signal Transduction. Plant Cell. 2010, 22, 2956–2969. [Google Scholar] [CrossRef]
- Sakakibara, H. CYTOKININS: Activity, Biosynthesis, and Translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and Signaling Aspects Underpinning the Regulation of Plant Carbon Nitrogen Interactions. Mol. Plant. 2010, 3, 973–996. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.-L. Sucrose Metabolism: Gateway to Diverse Carbon Use and Sugar Signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Merchante, C.; Alonso, J.M.; Stepanova, A.N. Ethylene signaling: simple ligand, complex regulation. Curr. Opin. Plant Biol. 2013, 16, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.-M.; Wang, R.-L.; Zhu, T.; Rothstein, S.J. Global transcription profiling reveals differential responses to chronic nitrogen stress and putative nitrogen regulatory components in Arabidopsis. BMC Genomics. 2007, 8, 281. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Cheng, Y.-H.; Chen, K.-E.; Tsay, Y.-F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef]
- Zhong, C.; Bai, Z.-G.; Zhu, L.-F.; Zhang, J.-H.; Zhu, C.-Q.; Huang, J.-L.; Jin, Q.-Y.; Cao, X.-C. Nitrogen-mediated alleviation of photosynthetic inhibition under moderate water deficit stress in rice (Oryza sativa L.). Environ. Exp. Bot. 2019, 157, 269–282. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, Z.; Wang, T.; Fu, Y.; Chen, Y.; Hu, B.; Yamagishi, J.; Ren, W. Environmental Compensation Effect and Synergistic Mechanism of Optimized Nitrogen Management Increasing Nitrogen Use Efficiency in Indica Hybrid Rice. Front. Plant. Sci. 2019, 10, 245. [Google Scholar] [CrossRef]
- Zhang, Z.; Xiong, S.; Wei, Y.; Meng, X.; Wang, X.; Ma, X. The role of glutamine synthetase isozymes in enhancing nitrogen use efficiency of N-efficient winter wheat. Sci. Rep. 2017, 7, 1000. [Google Scholar] [CrossRef]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops. J. Exp. Bot. 2002, 53, 979–987. [Google Scholar] [CrossRef]
- Ng, D.W.; Abeysinghe, J.K.; Kamali, M. Regulating the Regulators: The Control of Transcription Factors in Plant Defense Signaling. Int. J. Mol. Sci.. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Zhang, Y.; Du, W.; Xia, H.; Fan, S.; Zhang, B. Signaling Responses to N Starvation: Focusing on Wheat and Filling the Putative Gaps With Findings Obtained in Other Plants. A Review. Front. Plant. Sci. 2021, 12, 656696. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of low nitrogen on phenotypic characteristics and biomass of two foxtail millet varieties with different low-nitrogen tolerance. JG20-CK, JG20 under control; JG20-LN, JG20 under low nitrogen; JG22-CK, JG22 under control; JG22-LN, JG22 under low nitrogen. (A) Phenotypic characteristics of JG20 and JG22 under control and low-nitrogen conditions; Bar = 10cm. (B) Biomass of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. Error bars represent standard error of three biological replicates; different lowercase letters indicate significance at the level of P<0.05 between different treatments and varieties inshoots and roots.
Figure 1.
Effect of low nitrogen on phenotypic characteristics and biomass of two foxtail millet varieties with different low-nitrogen tolerance. JG20-CK, JG20 under control; JG20-LN, JG20 under low nitrogen; JG22-CK, JG22 under control; JG22-LN, JG22 under low nitrogen. (A) Phenotypic characteristics of JG20 and JG22 under control and low-nitrogen conditions; Bar = 10cm. (B) Biomass of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. Error bars represent standard error of three biological replicates; different lowercase letters indicate significance at the level of P<0.05 between different treatments and varieties inshoots and roots.
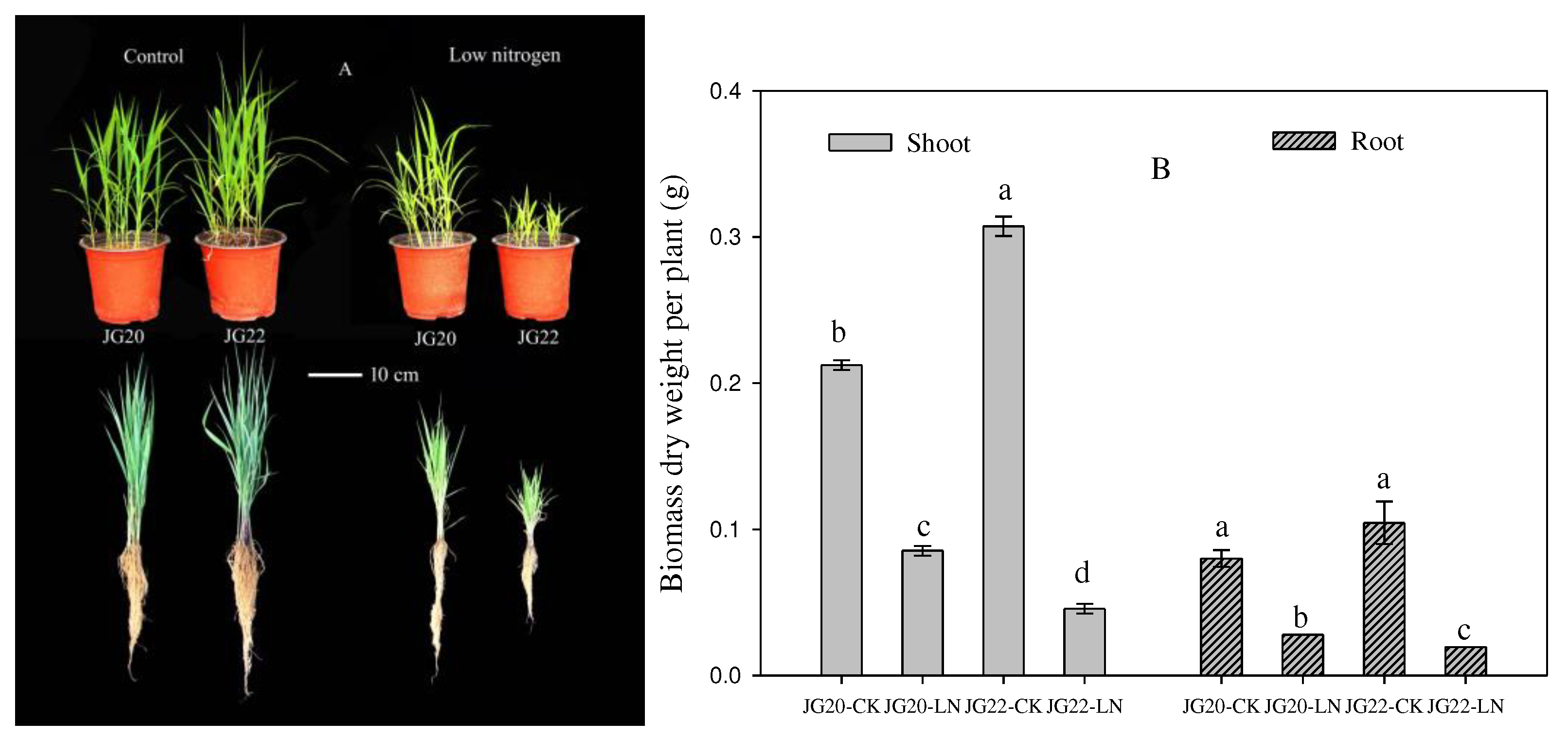
Figure 2.
Effect of low nitrogen on N concentration, N content, and N use efficiencyof two foxtail millet varieties with different low-nitrogen tolerances. JG20-CK, JG20 under control; JG20-LN, JG20 under low nitrogen; JG22-CK, JG22 under control; JG22-LN, JG22 under low nitrogen. (A) Nitrogen concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (B) Nitrogen content of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (C) Nitrogen partial productivity of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. Error bars represent standard error of three biological replicates; different lowercase letters indicate significance at the level of P<0.05 between different treatments and varieties in shoots and roots.
Figure 2.
Effect of low nitrogen on N concentration, N content, and N use efficiencyof two foxtail millet varieties with different low-nitrogen tolerances. JG20-CK, JG20 under control; JG20-LN, JG20 under low nitrogen; JG22-CK, JG22 under control; JG22-LN, JG22 under low nitrogen. (A) Nitrogen concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (B) Nitrogen content of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (C) Nitrogen partial productivity of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. Error bars represent standard error of three biological replicates; different lowercase letters indicate significance at the level of P<0.05 between different treatments and varieties in shoots and roots.
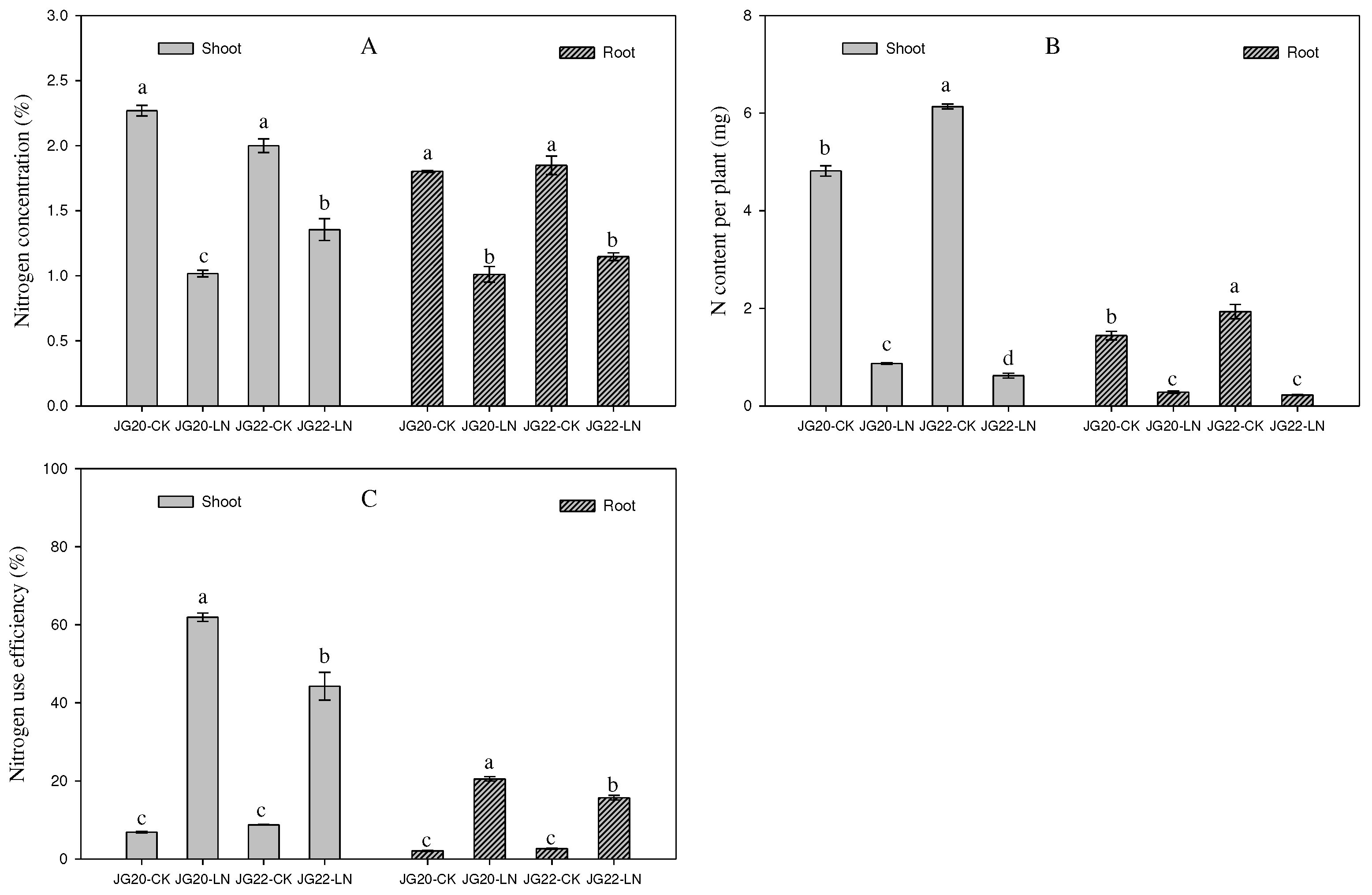
Figure 3.
Effect of low nitrogen on soluble protein concentration, free amino acid concentration and nitrate nitrogen concentration of two foxtail millet varieties. JG20-CK, JG20 under control; JG20-LN, JG20 under low nitrogen; JG22-CK, JG22 under control; JG22-LN, JG22 under low nitrogen. (A) Soluble protein concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (B) Free amino acid concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (C) Nitrate nitrogen concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. Error bars represented standard error of three biological replicates; different lowercase letters indicate significance at the level of P<0.05 between different treatments and varieties in shoots and roots.
Figure 3.
Effect of low nitrogen on soluble protein concentration, free amino acid concentration and nitrate nitrogen concentration of two foxtail millet varieties. JG20-CK, JG20 under control; JG20-LN, JG20 under low nitrogen; JG22-CK, JG22 under control; JG22-LN, JG22 under low nitrogen. (A) Soluble protein concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (B) Free amino acid concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (C) Nitrate nitrogen concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. Error bars represented standard error of three biological replicates; different lowercase letters indicate significance at the level of P<0.05 between different treatments and varieties in shoots and roots.
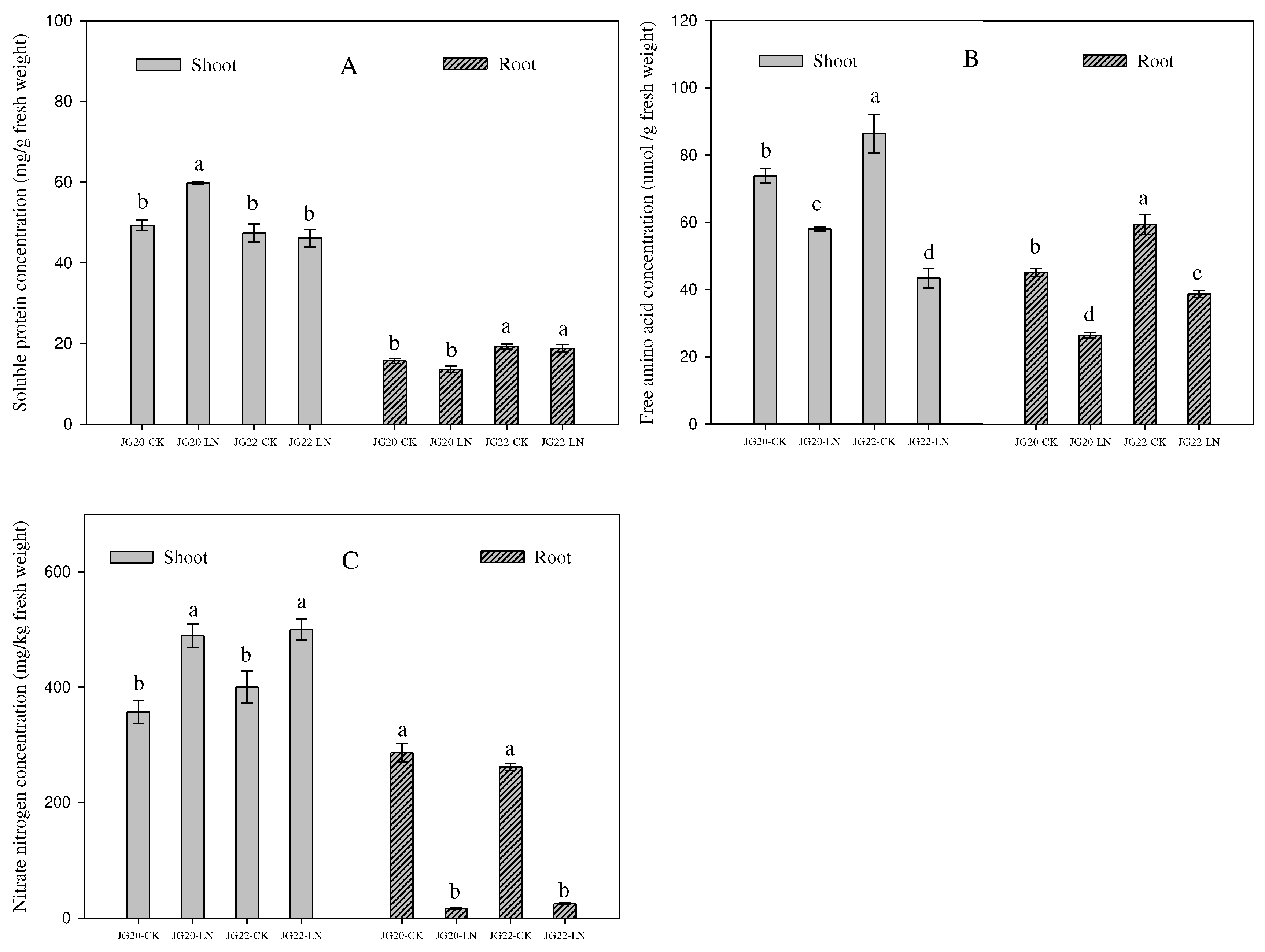
Figure 4.
Effect of low nitrogen on soluble sugar concentration, sucrose concentration, indole-3-acetic acid concentration, zeatin concentration and abscisic acid concentration of two foxtail millet varieties with different low-nitrogen tolerance. JG20-CK, JG20 under control; JG20-LN, JG20 under low nitrogen; JG22-CK, JG22 under control; JG22-LN, JG22 under low nitrogen. (A) Soluble sugar concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (B) Sucrose concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (C) Indole-3-acetic acid concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (D) Zeatin concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (E) Abscisic acid concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. Error bars represented standard error of three biological replicates; different lowercase letters indicate significance at the level of P<0.05 between different treatments and varieties in shoots and roots.
Figure 4.
Effect of low nitrogen on soluble sugar concentration, sucrose concentration, indole-3-acetic acid concentration, zeatin concentration and abscisic acid concentration of two foxtail millet varieties with different low-nitrogen tolerance. JG20-CK, JG20 under control; JG20-LN, JG20 under low nitrogen; JG22-CK, JG22 under control; JG22-LN, JG22 under low nitrogen. (A) Soluble sugar concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (B) Sucrose concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (C) Indole-3-acetic acid concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (D) Zeatin concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. (E) Abscisic acid concentration of shoots and roots of JG20 and JG22 under control and low-nitrogen conditions. Error bars represented standard error of three biological replicates; different lowercase letters indicate significance at the level of P<0.05 between different treatments and varieties in shoots and roots.
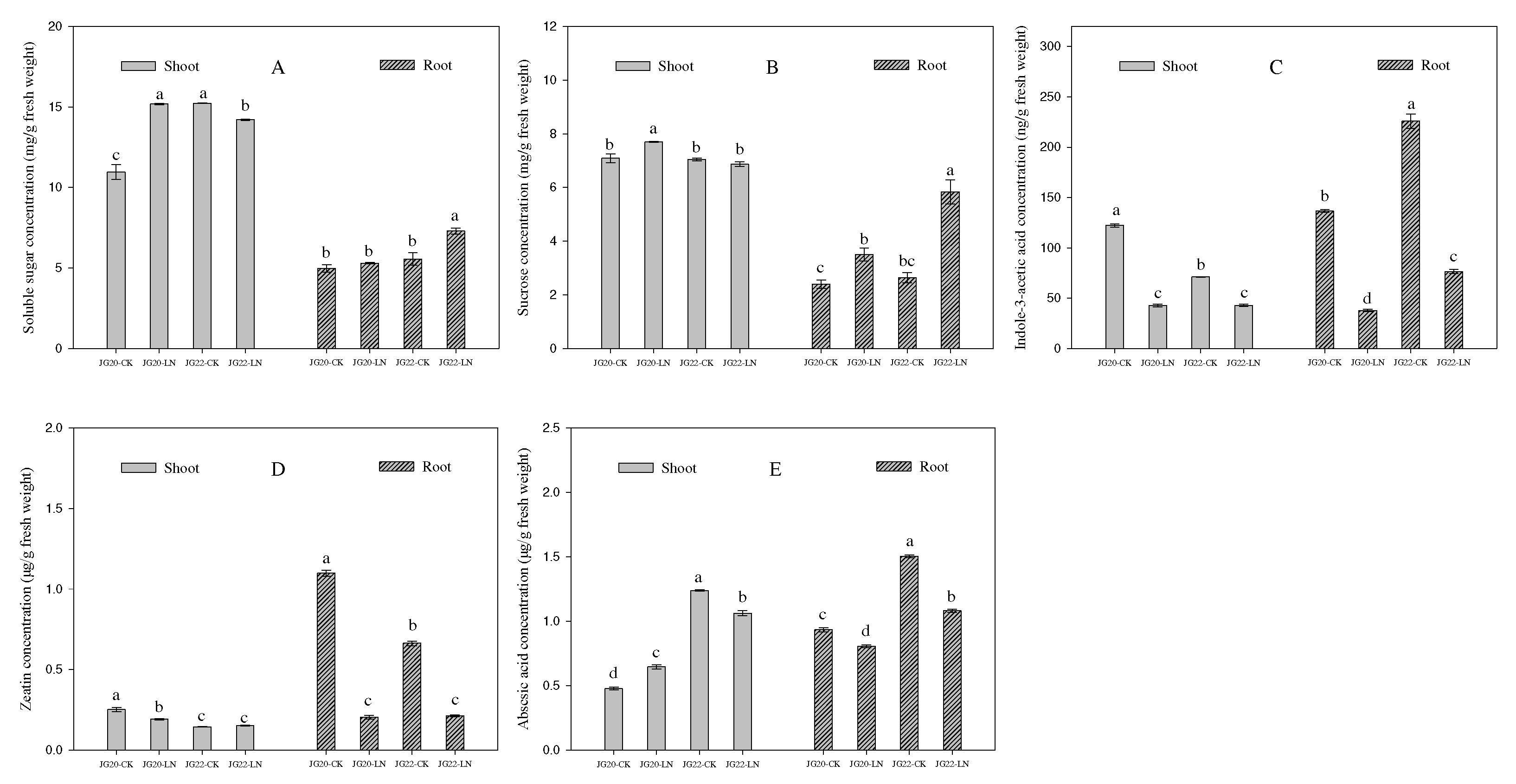
Figure 5.
Comparison of the expression profiles of selected DEGs determined by RT-qPCR and RNA-Seq analyses. Expression levels of 15 DEGs in the low-nitrogen tolerant variety JG20 (A) and the low-nitrogen sensitive variety JG22 (B). Values are presented as log2 (fold-change). The X-axis represents gene ID, according to the NCBI database. (C) Scatter plots of the expression levels of 15 DEGs in control and low-nitrogen conditions. X and Y axes represent log2 (fold-change) determined by RAN-Seq and RT-qPCR, respectively (R2=0.936). Error bars represent the standard error of three biological replicates.
Figure 5.
Comparison of the expression profiles of selected DEGs determined by RT-qPCR and RNA-Seq analyses. Expression levels of 15 DEGs in the low-nitrogen tolerant variety JG20 (A) and the low-nitrogen sensitive variety JG22 (B). Values are presented as log2 (fold-change). The X-axis represents gene ID, according to the NCBI database. (C) Scatter plots of the expression levels of 15 DEGs in control and low-nitrogen conditions. X and Y axes represent log2 (fold-change) determined by RAN-Seq and RT-qPCR, respectively (R2=0.936). Error bars represent the standard error of three biological replicates.
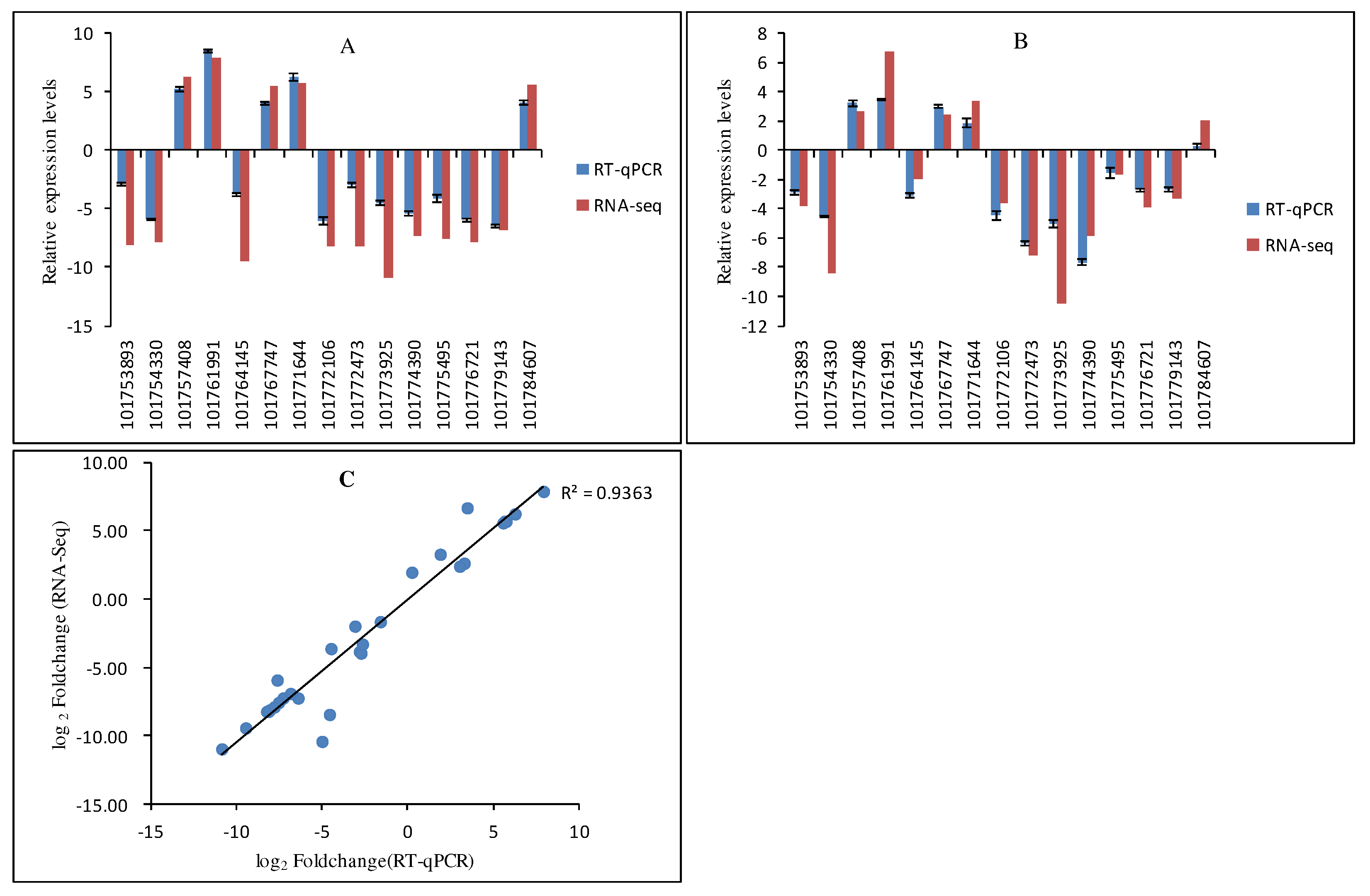
Figure 6.
Gene expression analyses of JG20 and JG22 subjected to low-nitrogen conditions. (A) The numbers of DEGs in shoots in different comparison groups. (B) Venn diagram of the numbers of DEGs in shoots in different comparisons among groups. (C) The numbers of DEGs in roots in different comparison groups. (D) Venn diagram of the numbers of DEGs in roots in different comparisons among groups. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
Figure 6.
Gene expression analyses of JG20 and JG22 subjected to low-nitrogen conditions. (A) The numbers of DEGs in shoots in different comparison groups. (B) Venn diagram of the numbers of DEGs in shoots in different comparisons among groups. (C) The numbers of DEGs in roots in different comparison groups. (D) Venn diagram of the numbers of DEGs in roots in different comparisons among groups. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
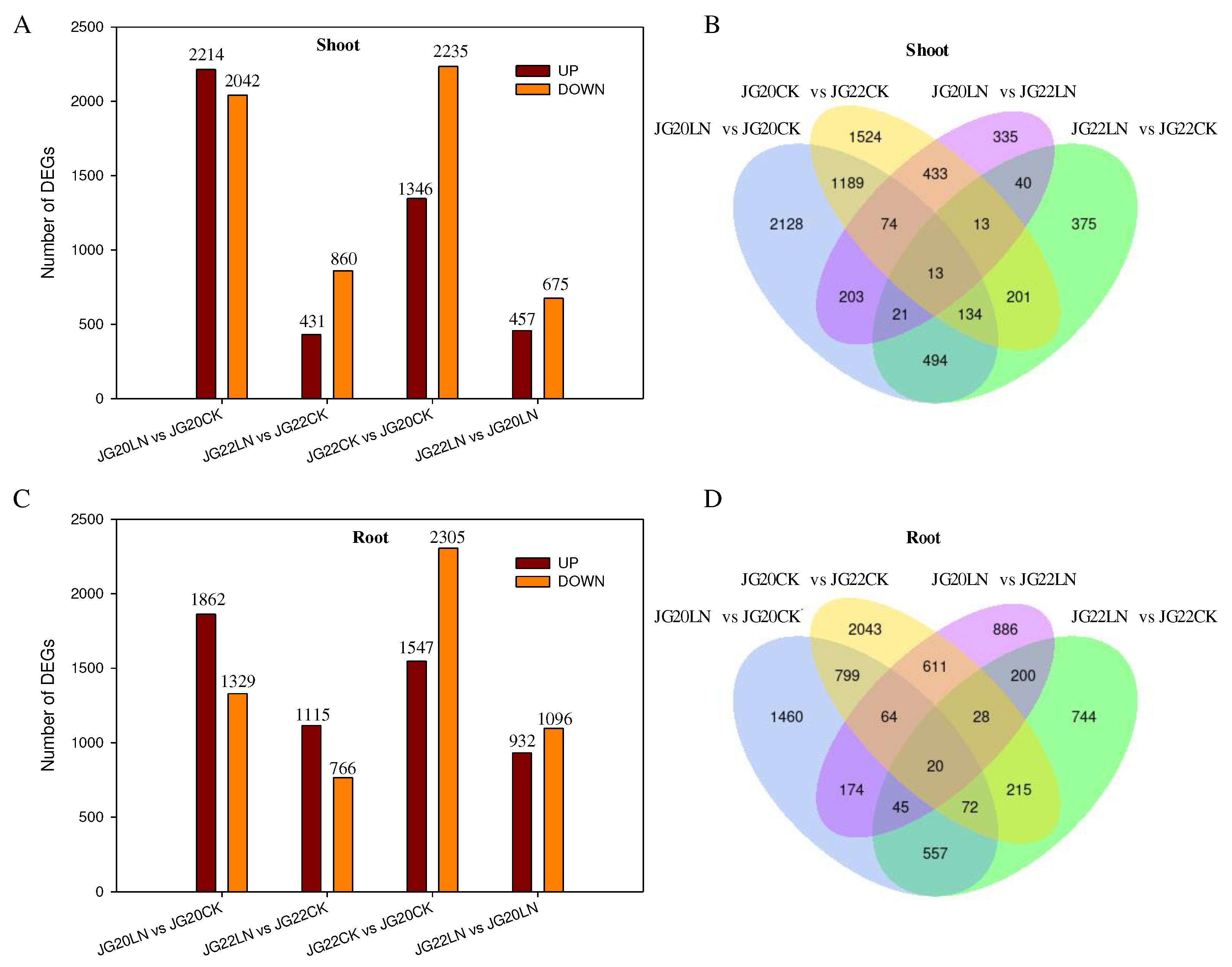
Figure 7.
Gene ontology (GO) enrichment of DEGs in response to low nitrogen (LN) in LN-tolerant JG20 and LN-sensitive JG22 foxtail millet varieties. (A) GO enrichment of DEGs in the shoots between LN and CK in JG20. (B) GO enrichment of DEGs in the shoots between LN and CK in JG22. (C) GO enrichment of DEGs in roots between LN and CK in JG20. (D) GO enrichment of DEGs in roots between LN and CK in JG22. Blue columns indicate the numbers of up-regulated genes, while red columns indicate numbers of down-regulated genes. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
Figure 7.
Gene ontology (GO) enrichment of DEGs in response to low nitrogen (LN) in LN-tolerant JG20 and LN-sensitive JG22 foxtail millet varieties. (A) GO enrichment of DEGs in the shoots between LN and CK in JG20. (B) GO enrichment of DEGs in the shoots between LN and CK in JG22. (C) GO enrichment of DEGs in roots between LN and CK in JG20. (D) GO enrichment of DEGs in roots between LN and CK in JG22. Blue columns indicate the numbers of up-regulated genes, while red columns indicate numbers of down-regulated genes. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.

Figure 8.
Scatter plots of top 20 KEGG pathways in two varieties in response to low nitrogen. (A) KEGG analysis of DEGs identified in the shoots of JG20 between CK and LN. (B) KEGG analysis of DEGs identified in the shoots of JG22 between CK and LN. (C) KEGG analysis of DEGs identified in the roots of JG22 between CK and low nitrogen LN. (D) KEGG analysis of DEGs identified in the roots of JG22 between CK and low nitrogen LN. Gene Ratio shows the ratio of the number of DEGs in a specific pathway to the total number of DEGs in KEGG. Pathways are listed along the y-axis. The circle area indicates the number of DEGs, and the circle color represents the ranges of the corrected P-values.
Figure 8.
Scatter plots of top 20 KEGG pathways in two varieties in response to low nitrogen. (A) KEGG analysis of DEGs identified in the shoots of JG20 between CK and LN. (B) KEGG analysis of DEGs identified in the shoots of JG22 between CK and LN. (C) KEGG analysis of DEGs identified in the roots of JG22 between CK and low nitrogen LN. (D) KEGG analysis of DEGs identified in the roots of JG22 between CK and low nitrogen LN. Gene Ratio shows the ratio of the number of DEGs in a specific pathway to the total number of DEGs in KEGG. Pathways are listed along the y-axis. The circle area indicates the number of DEGs, and the circle color represents the ranges of the corrected P-values.
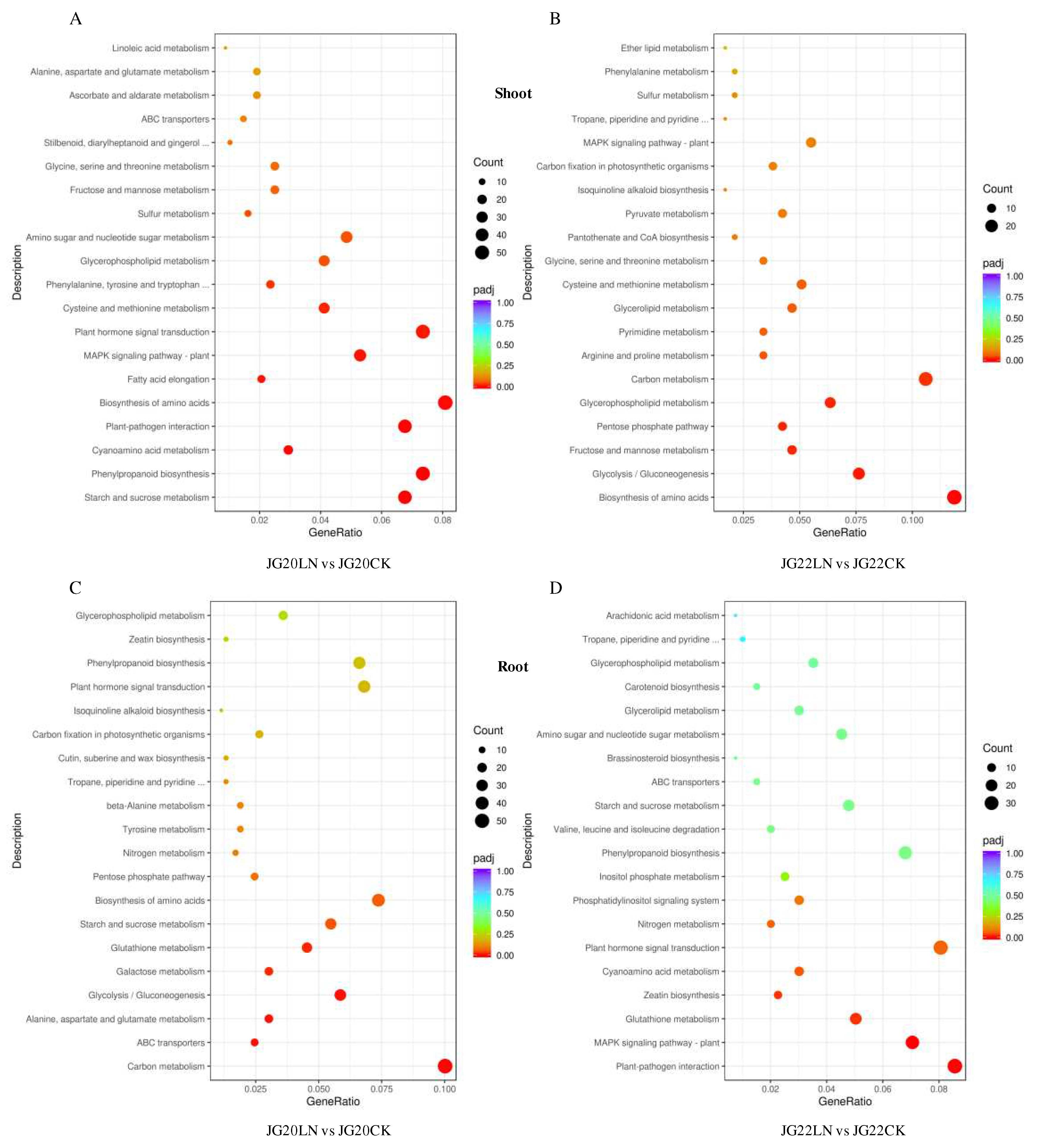
Figure 9.
Expression profiles of genes involved in nitrogen (N) source uptake, transport, and assimilation in the shoots and roots of two foxtail millet varieties in response to low nitrogen. (A) Outline of N uptake, transport, and assimilation. (B) Heatmap visualization of expression profiles of inorganic N source transporters (NRT and AMT) in N-starved foxtail millet shoots and roots. (C) Heatmap visualization of expression profiles of inorganic N source assimilation (NR, NiR, GS, GOGAT, and GDH) in N-starved foxtail millet shoots and roots. NRT (nitrate transporter); AMT (ammonium transporter); NR (nitrate reductase); NiR (nitrite reductase); GS (glutamine synthetase); GOGAT (glutamate synthetase); GDH (glutamate dehydrogenase); Glu, glutamate; Gln, glutamine; 2-OG, 2-oxoglutarate; AA, amino acid. The bar on the right side of the heatmap represents relative expression level of DEGs. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
Figure 9.
Expression profiles of genes involved in nitrogen (N) source uptake, transport, and assimilation in the shoots and roots of two foxtail millet varieties in response to low nitrogen. (A) Outline of N uptake, transport, and assimilation. (B) Heatmap visualization of expression profiles of inorganic N source transporters (NRT and AMT) in N-starved foxtail millet shoots and roots. (C) Heatmap visualization of expression profiles of inorganic N source assimilation (NR, NiR, GS, GOGAT, and GDH) in N-starved foxtail millet shoots and roots. NRT (nitrate transporter); AMT (ammonium transporter); NR (nitrate reductase); NiR (nitrite reductase); GS (glutamine synthetase); GOGAT (glutamate synthetase); GDH (glutamate dehydrogenase); Glu, glutamate; Gln, glutamine; 2-OG, 2-oxoglutarate; AA, amino acid. The bar on the right side of the heatmap represents relative expression level of DEGs. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
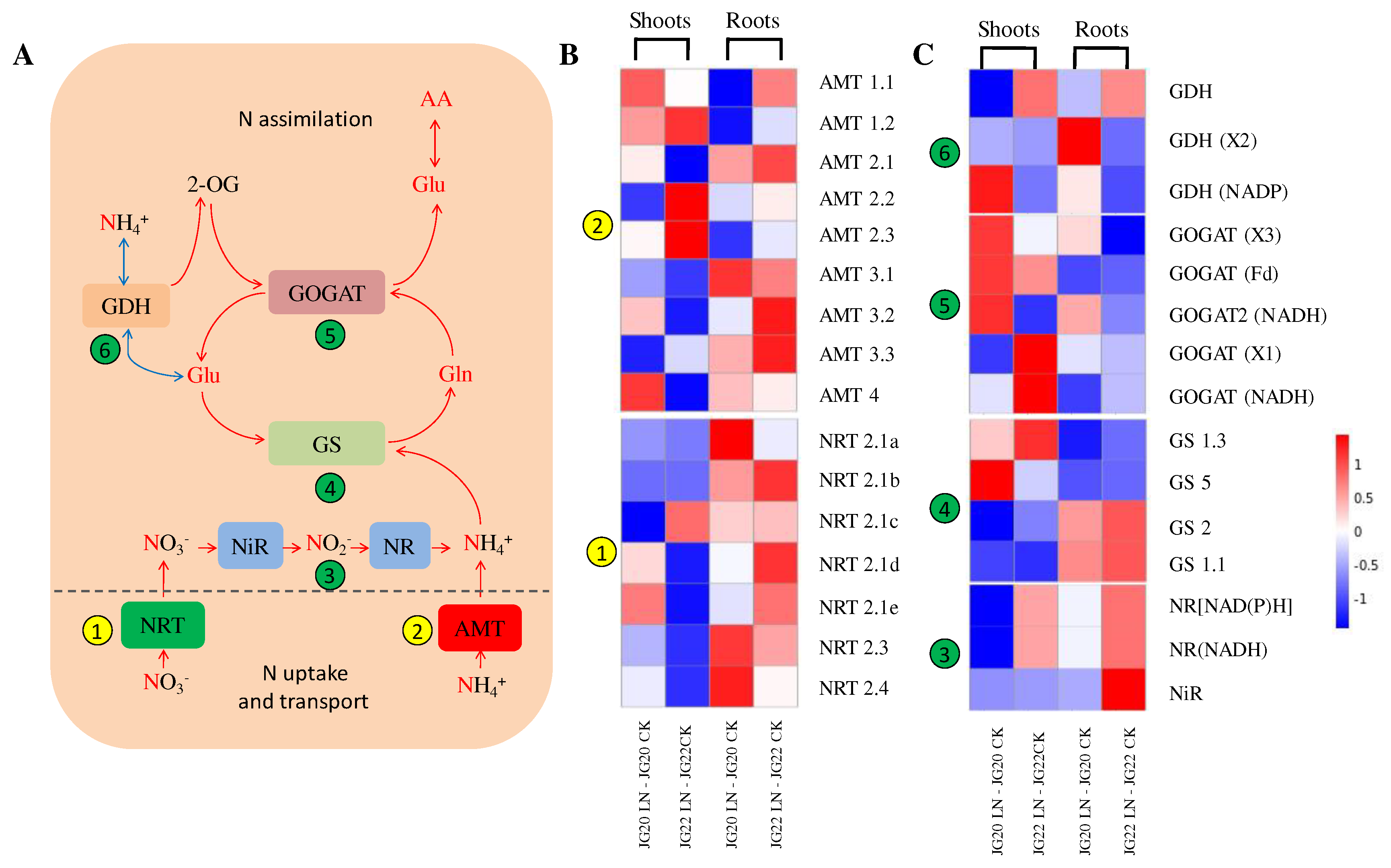
Figure 10.
Heatmap visualization of expression profiles of genes involved in photosynthesis (A), hormone signal transduction (B) and glycolysis (C) in two foxtail millet varieties in response to low nitrogen. Psa, chlorophyll apoprotein of photosystem I; Psb, chlorophyll apoprotein of photosystem II; b, ATP synthase subunit b; LHC, light-harvesting chlorophyll protein complex; Lhcb1, light-harvesting complex II chlorophyll b binding protein; AUX, Auxin; IAA, Indole-3-acetic acid; AFR, Auxin response factor; GH3, indole-3-acetic acid-amido synthetase GH3; SAUR, auxin-responsive protein SAUR; AHP, histidine-containing phosphotransfer protein; A-ARR/B-ARR, two-component response regulator ORR; TF, transcription factor; EBF, EIN3-binding F-box protein; EIN3, ethylene-insensitive protein 3; PYR/PYL, abscisic acid receptor; PP2C, probable protein phosphatase 2C; SnRK2, serine/threonine-protein kinase; ABF, G-box-binding factor; HXK, hexokinase; AEP, aldose 1-epimerase; GPI, glucose-6-phosphate isomerase; ATP-PFK, ATP-dependent 6-phosphofructokinase; FBA, fructose-bisphosphate aldolase; TIM, triosephosphate isomerase; PGK, phosphoglycerate kinase; PGMP, phosphoglycerate mutase-like protein; ENO, enolase; PCK, phosphoenolpyruvate carboxykinase; PK, pyruvate kinase; PDH-E1α, pyruvate dehydrogenase E1 component subunit alpha; PDC, pyruvate decarboxylase; PDH-E2, pyruvate dehydrogenase E2 component; DLD, dihydrolipoamide dehydrogenase; ALDH, aldehyde dehydrogenase. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
Figure 10.
Heatmap visualization of expression profiles of genes involved in photosynthesis (A), hormone signal transduction (B) and glycolysis (C) in two foxtail millet varieties in response to low nitrogen. Psa, chlorophyll apoprotein of photosystem I; Psb, chlorophyll apoprotein of photosystem II; b, ATP synthase subunit b; LHC, light-harvesting chlorophyll protein complex; Lhcb1, light-harvesting complex II chlorophyll b binding protein; AUX, Auxin; IAA, Indole-3-acetic acid; AFR, Auxin response factor; GH3, indole-3-acetic acid-amido synthetase GH3; SAUR, auxin-responsive protein SAUR; AHP, histidine-containing phosphotransfer protein; A-ARR/B-ARR, two-component response regulator ORR; TF, transcription factor; EBF, EIN3-binding F-box protein; EIN3, ethylene-insensitive protein 3; PYR/PYL, abscisic acid receptor; PP2C, probable protein phosphatase 2C; SnRK2, serine/threonine-protein kinase; ABF, G-box-binding factor; HXK, hexokinase; AEP, aldose 1-epimerase; GPI, glucose-6-phosphate isomerase; ATP-PFK, ATP-dependent 6-phosphofructokinase; FBA, fructose-bisphosphate aldolase; TIM, triosephosphate isomerase; PGK, phosphoglycerate kinase; PGMP, phosphoglycerate mutase-like protein; ENO, enolase; PCK, phosphoenolpyruvate carboxykinase; PK, pyruvate kinase; PDH-E1α, pyruvate dehydrogenase E1 component subunit alpha; PDC, pyruvate decarboxylase; PDH-E2, pyruvate dehydrogenase E2 component; DLD, dihydrolipoamide dehydrogenase; ALDH, aldehyde dehydrogenase. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
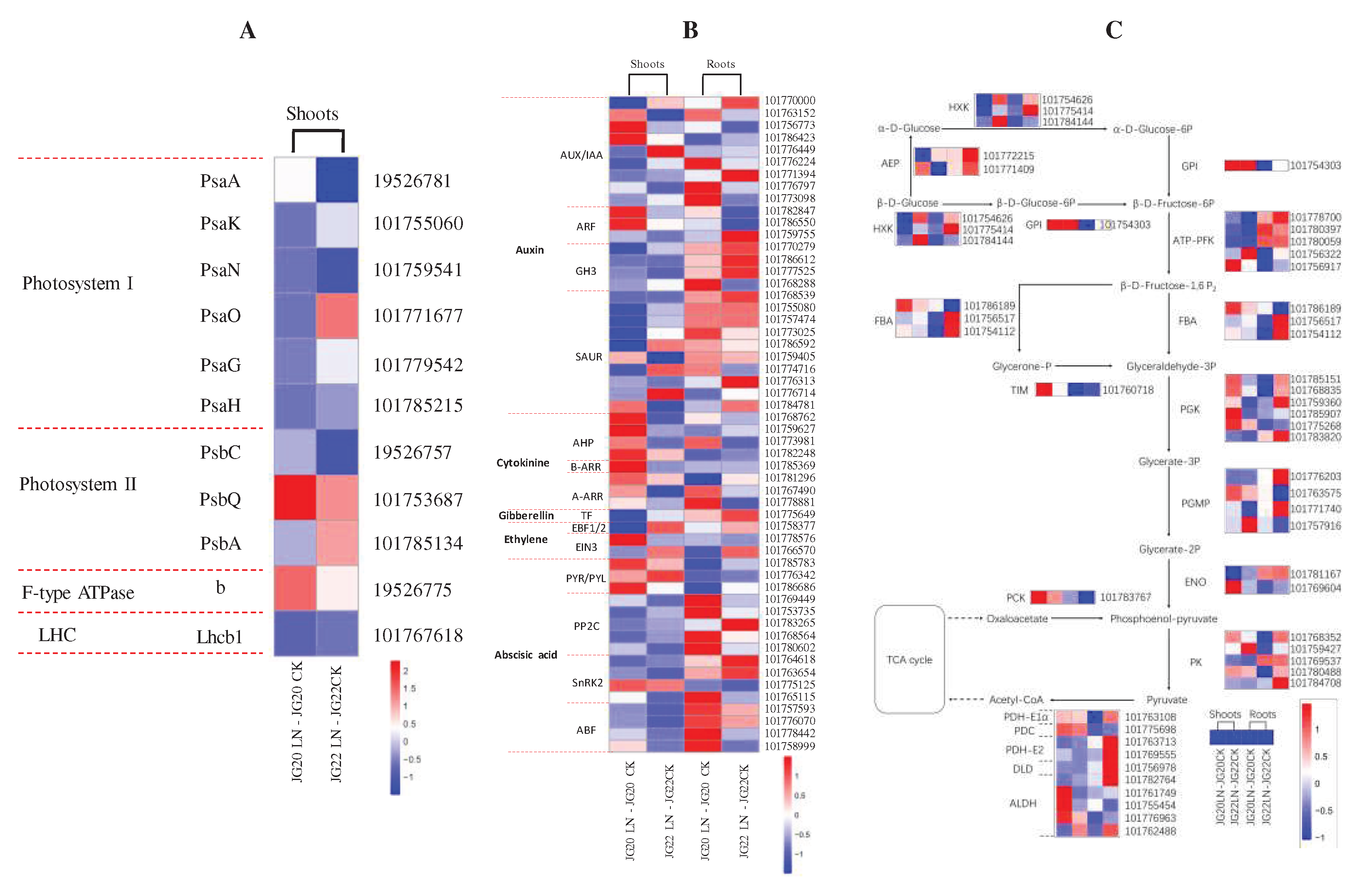
Figure 11.
Schematic representation of the main processes and important genes involved in low nitrogen response in two foxtail millet varieties with different nitrogen use efficiency. The color scale represents increased (red) or decreased (blue) fold-change expression of DEGs in samples exposed to low nitrogen. NRT, nitrate transporter; AMT, ammonium transporter; GS, glutamine synthetase; GOGAT, glutamate synthetase; AUX, Auxin; IAA, Indole-3-acetic acid; AFR, Auxin response factor; GH3, indole-3-acetic acid-amido synthetase GH3; SAUR, aux-in-responsive protein SAUR; AHP, histidine-containing phosphotransfer protein; A-ARR/B-ARR, two-component response regulator ORR; PYR/PYL, abscisic acid receptor; PP2C, probable pro-tein phosphatase 2C; SnRK2, serine/threonine-protein kinase; ABF, G-box-binding factor; Psb, chlorophyll apoprotein of photosystem II; b, ATP synthase subunit b; AEP, aldose 1-epimerase; GPI, glucose-6-phosphate isomerase; ATP-PFK, ATP-dependent 6-phosphofructokinase; FBA, fructose-bisphosphate aldolase; TIM, tri-osephosphate isomerase; PGK, phosphoglycerate kinase; PGMP, phosphoglycerate mutase-like protein; ENO, enolase; PCK, phosphoenolpyruvate carboxykinase; ALDH, aldehyde dehydrogenase. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
Figure 11.
Schematic representation of the main processes and important genes involved in low nitrogen response in two foxtail millet varieties with different nitrogen use efficiency. The color scale represents increased (red) or decreased (blue) fold-change expression of DEGs in samples exposed to low nitrogen. NRT, nitrate transporter; AMT, ammonium transporter; GS, glutamine synthetase; GOGAT, glutamate synthetase; AUX, Auxin; IAA, Indole-3-acetic acid; AFR, Auxin response factor; GH3, indole-3-acetic acid-amido synthetase GH3; SAUR, aux-in-responsive protein SAUR; AHP, histidine-containing phosphotransfer protein; A-ARR/B-ARR, two-component response regulator ORR; PYR/PYL, abscisic acid receptor; PP2C, probable pro-tein phosphatase 2C; SnRK2, serine/threonine-protein kinase; ABF, G-box-binding factor; Psb, chlorophyll apoprotein of photosystem II; b, ATP synthase subunit b; AEP, aldose 1-epimerase; GPI, glucose-6-phosphate isomerase; ATP-PFK, ATP-dependent 6-phosphofructokinase; FBA, fructose-bisphosphate aldolase; TIM, tri-osephosphate isomerase; PGK, phosphoglycerate kinase; PGMP, phosphoglycerate mutase-like protein; ENO, enolase; PCK, phosphoenolpyruvate carboxykinase; ALDH, aldehyde dehydrogenase. The threshold for differential expression was set at log2 fold-change > 1 and padj < 0.05.
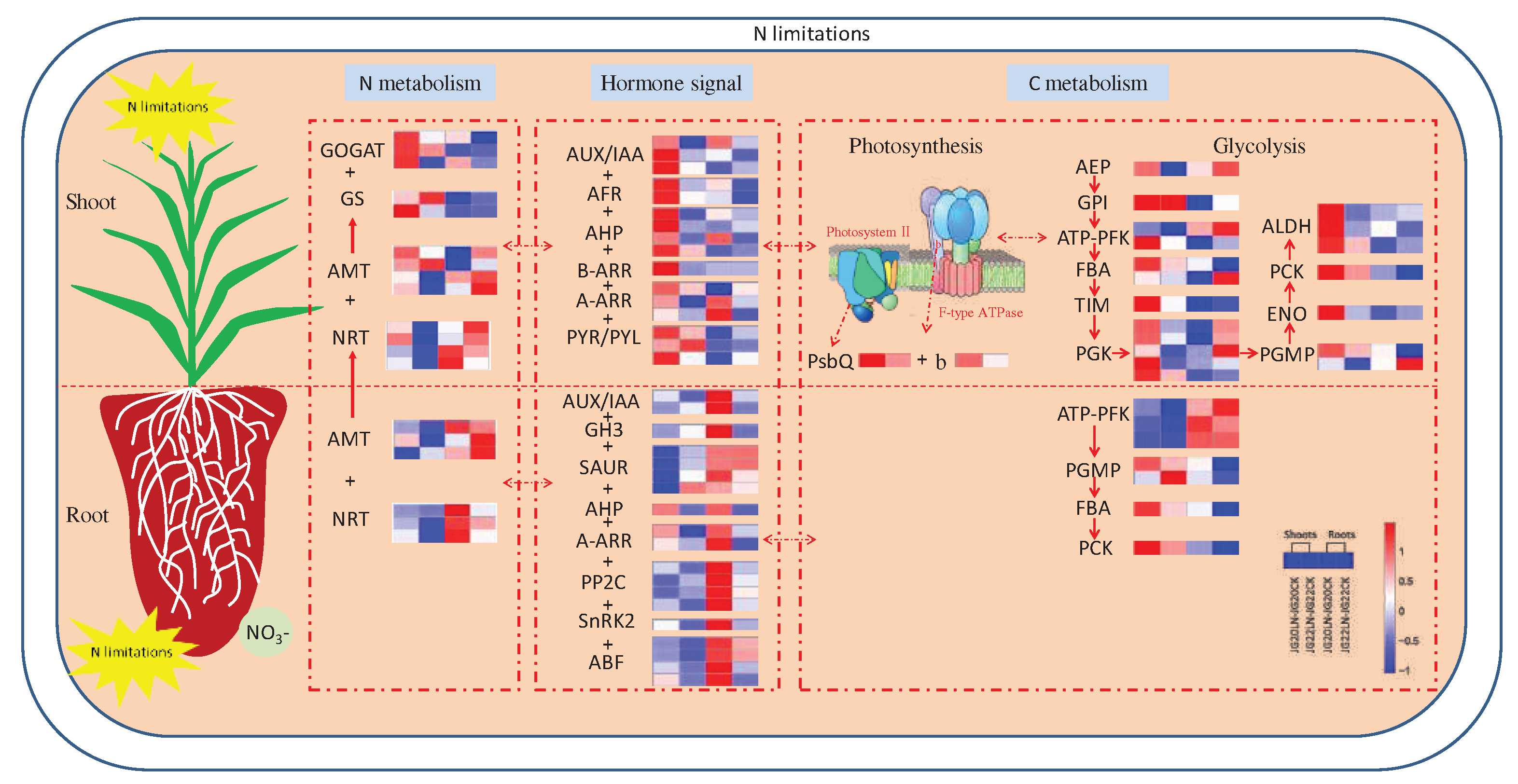

Table 1.
Quality statistics of read numbers of foxtail millet transcriptomics.
| Sample name | Raw reads | Clean reads | Clean bases | Error rate (20%) |
Q20 (%) |
Q30 (%) |
GC content (%) |
Total mapped | Uniquely mapped | Multiple mapped |
|---|---|---|---|---|---|---|---|---|---|---|
| SJG20-CK | 45434210 | 44021351 | 6.60G | 0.02 | 98.18 | 94.78 | 56.54 | 42466340(96.47%) | 40838306(92.77%) | 1628034(3.70%) |
| RJG20-CK | 46926017 | 44908641 | 6.73G | 0.02 | 98.32 | 95.10 | 53.21 | 38087242(84.81%) | 37391122(83.26) | 696119(1.55%) |
| SJG20-LN | 46568083 | 45266295 | 6.79G | 0.02 | 98.15 | 94.69 | 56.37 | 43654878(96.44%) | 42879943(94.73%) | 774935(1.71%) |
| RJG20-LN | 45728815 | 44559130 | 6.68G | 0.02 | 98.11 | 94.52 | 52.95 | 33272477(74.67%) | 32684085(73.35%) | 588392(1.32%) |
| SJG22-CK | 42193254 | 41110815 | 6.17G | 0.02 | 98.36 | 95.08 | 53.98 | 39846088(96.92%) | 38835094(94.46%) | 1010994(2.46%) |
| RJG22-CK | 43185923 | 42196855 | 6.33G | 0.02 | 98.33 | 94.95 | 50.22 | 30066139(71.25%) | 29460001(69.82%) | 606138(1.44%) |
| SJG22-LN | 44891345 | 43960698 | 6.59G | 0.02 | 98.13 | 94.55 | 54.73 | 42392060(96.43%) | 41541056(94.50%) | 851004(1.94%) |
| RJG22-LN | 43616102 | 42607394 | 6.39G | 0.02 | 98.24 | 94.77 | 52.21 | 31884267(74.83%) | 31199409(73.23%) | 684858(1.61%) |
Notes: SJG20-CK, Shoot of JG20 under control; RJG20-CK, Root of JG20 under control; SJG20-LN, Shoot of JG20 under low nitrogen; RJG20-LN, Root of JG20 under low nitrogen; SJG22-CK, Shoot of JG22 under control; RJG22-CK, Root of JG22 under control; SJG22-LN, Shoot of JG22 under low nitrogen; RJG22-LN, Root of JG22 under low nitrogen; Raw Reads: The number of reads before filtering; Clean Reads: Filtered reads; Total Clean Bases (Gb): Total number of bases after filtration; Q20 (%): Proportion of nucleotides with a quality value larger than 20 in the filtered reads; Q30 (%): Proportion of nucleotides with a quality value larger than 30 in the filtered reads; GC content: Percent of GC bases on total bases.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



