
Submitted:
08 May 2023
Posted:
09 May 2023
You are already at the latest version
Abstract
Because rice is native to tropical and subtropical regions, it is generally more sensitive to cold stress compared to other cereals. Low temperature stress has become one of the most important conditions that affects the growth, development, and yield of rice. Plant NAC (NAM, ATAF1/2, and CUC) transcription factors are known to play significant regulatory roles in the stress response. In our research, we found that OsNAC050 (LOC_Os03g60080) is mainly expressed in leaves, and low temperature can further up-regulate its expression. OsNAC050 mutants created using CRISPR-Cas9 gene editing technology showed significantly enhanced tolerance to low temperature treatment. Detection of enzyme activities related to the redox pathway also showed that the mutants had stronger viability under low temperature stress. Comparative transcriptome analysis showed that photosynthesis and soluble sugar metabolism were significantly affected in the osnac050 mutant lines, suggesting that OsNAC050 may participate in the above molecular pathways in the response to low temperature stress. The results will enhance our understanding of molecular mechanisms underlying the responses to cold stress in rice and can provide new strategies for engineering cold-tolerance in high-yielding rice varieties.
Keywords:
OsNAC050
; CRISPR-Cas9
; cold stress
; RNA-seq
; photosynthesis
; soluble sugar
1. Introduction
Low temperature is one type of environmental stress that has a strong effect on plant growth (Zhang et al. 2022). When the ambient temperatures are continuously lower than the lowest temperature at which plants can grow normally for a period of time, low temperature stress will occur (Yu, Liu, et al. 2022). The abnormal growth of plants caused by low temperatures above 0°C is called chilling injury. Many plants prefer warm temperatures; examples are rice, maize (Zea mays), kidney beans (Phaseolus vulgaris), and tropical and subtropical fruit crops such as bananas and pineapples, which are highly susceptible to cold damage(Guo et al. 2022; Yang et al. 2021). Under normal conditions, when the temperature is below 10°C, chilling damage will occur, which mainly causes discoloration of the plants, necrosis, and formation of lesions on the surfaces of leaves. In the early growth stage of rice seedlings, low temperatures and cold air currents are likely to increase the incidence of bacterial wilt that result in seedling mortality(Wang et al. 2021). However, in the late stage of rice panicle differentiation at the flowering stage, low temperature exposure for an extended period of time will also affect grain yield due to the abnormal development of pollen grains(Liu et al. 2021).
The NAC TF family are known to be related to abiotic stress (Sun et al. 2022). Transgenic rice lines overexpressing SNAC1, SNAC2, and SNAC3 show improved tolerance to salt, while RNAi lines in which these genes are silenced display increased plant susceptibility to these stresses(Zhang et al. 2021; Liu et al. 2014). The NAC-type transcription factor OsNAC2 regulates the expression of ABA-dependent genes and abiotic stress tolerance in rice(Shen et al. 2017). It has been reported that the suppressor line (ONAC095-SRDX) was reduced than wild type (Huang et al. 2016). Overexpressing the NAC transcription factor gene OsY37 promotes leaf senescence and causes early heading in rice (El Mannai et al. 2017). Knockdown of OsNAC5 renders RNAi plants more susceptible to oxidative damage(Takasaki et al. 2010).
With the maturity of gene knockout technology, CRISPR-Cas9 genome editing has rapidly become an important tool in genomic research due to its advantages of conceptually straightforward operation, precise targeting, and relatively high editing efficiency. Therefore, in this study, a member of the NAC transcription factor family, OsNAC050, was identified and knockout mutants were created using CRISPR-Cas9 gene editing. Our results will provide additional information on the functions of a member of the rice NAC transcription factor family.
2. Materials and Methods
2.1. Plant Growth Conditions and Stress Treatments
Plump seeds were chosen from ‘Nipponbare’ (Oryza sativa L. ssp. japonica) and the mutants. The hulls were removed and the seeds were surface-sterilized in 1% NaClO (bleach) for 15 min, followed by washing six times in sterilized distilled water. Fifty seeds were sterilized and placed on MS medium for germination in a light incubator at 28oC under a 16 h light (3,000 Lux)/8 h dark photoperiod. WT and osnac050 mutant rice plants grown for four weeks (28 d) were selected for low temperature stress treatment, and the seedlings were transferred to a refrigerator at 4°C on day 29. Physiological indicators of the treated plants were assayed at the same times. The plants were sampled at the designated time points with three biological replicates.
2.2. Quantification of relative gene expression by using qRT-PCR
Total RNA was isolated from these samples using the modified Trizol Reagent (Invitrogen, Waltham, MA, USA) following the manufacturer’s protocol. The RNA samples were then subjected to qRT-PCR to quantify OsNAC050 expression in WT plants, using the primers shown in Supplemental Table 1. The rice Actin gene was used as the internal control for normalization of gene expression. The 2–DDCt method was used to calculate relative expression levels. To study the expression rule of OsNAC050 gene expression under different treatments, carry out abiotic stress on it, and take samples at the 0, 1, 3, 6, 9, 12 hour.
2.3. Subcellular localization of OsNAC050
The full length of the gene was cloned into the positioning vector by means of enzyme digestion and connection, and transformed into the rice protoplast(Yoo, Cho and Sheen 2007).:The subcellular localization vector: pBWA(V)HS-ccdb-GLosgfp. Nikon C2-ER laser confocal microscope was used for observation, in which the information of thew2 nuclear marker: Fusion fluorescent protein: mkate, excitation light 561nm, emission light 580nm. Nuclear localization signal protein: nls Amino acid sequence: MDPKKKRKV Fusion fluorescent protein: mKATE excited light at 561nm and emitted light at 580nm(Zhao et al. 2017).
2.3. Targeted Mutagenesis of OsNAC050
OsNAC050 mutants were generated using the CRISPR-Cas9 knock out vector pZHY988. CRISPR-P (http://crispr.hzau.edu.cn/CRISPR2) was used to design the sgRNA (small guide RNA) and to predict potential off-target sites. After the sgRNA was synthesized, the vector was constructed using recombinant DNA methods. Ten positive clones subjected to Sanger sequencing. After detection and DNA sequencing identified the desired vector, rice callus was transformed using Agrobacterium tumefaciens EHA105(Wang et al. 2020). The sequences of primers used to detect off-targets are given in Supplementary Table 1 (Tang et al. 2016; Zhong et al. 2018). Double chain insertion T named osnac050 (T/T). Double chain deletion 5-bp deletion (ATGAG) named osnac050 (-5/-5).
2.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
qRT-PCR assays were used to quantify OsNAC050 expression profile analysis and ten pairs of gene-specific primers (Supplementary Table 1) were designed to generate the amplicons used to verify gene expression levels determined using RNA-seq data. We sampled roots, stems, and leaves at the seedling stage (4 weeks), and roots, stems, sheaths, leaves, and ears at the heading stage (10 weeks). After the seeds are vernalized at 4℃, they were planted in soil and grown under specified greenhouse conditions and watered regularly. During sampling, we selected three plants per line at the same growth stage for RNA extraction. To examine OsNAC050 expression levels under various stress and hormone treatments, we subjected four-week-old ‘Nipponbare’ seedlings to cold (4°C), heat (42°C), PEG 6000 (20%, w/v), NaCl (200 mM), H2O2 (1%), IAA (100 μM), ABA (100 μM), and GA3 (100 μM) treatments. Also, qRT-PCR was used to validate the accuracy of the RNA-seq data using 10 randomly selected genes with significantly different expression profiles.Plants were sampled at different time periods, with three replicates for each sampling, and then RNA was extracted. Error bars show the standard deviation based on three independent biological replicates. We used the SYBR Green qPCR kit (Takara) according to the manufacturer's instructions to perform quantitative real-time PCR (qRT-PCR) on an iQ5.0 instrument (Bio Rad, USA). The relative gene expression levels were calculated using the 2–ΔΔCt method. Quantification was achieved by normalizing the expression to that of the actin and GAPDH genes.
2.5. Physiological Measurements
Seeds of the WT and OsNAC050 mutants were grown in pots to the 4-week seedling stage, followed by the induction of cold stress. Five days later, we measured the physiological indices using our previously described methods [14].
2.6. Transcriptome analysis and data analysis
WT and osnac050(T/T), osnac050(-5/-5) plants were used for RNA-seq analysis. Plants were grown for 10 weeks under normal conditions or for eight weeks under normal conditions and then cold-treated for two weeks. Mixed samples were made by combining three T2 -generation seedlings. The RNA samples were sequenced and analyzed at the Beijing Institute of Genomics (Shenzhen, China). Genes with p-value < 0.05 and fold change >2 are advised to define differentially expressed gene (DEGs).
3. Results
3.1. OsNAC050 is a cold-inducible transcription factor gene
Our experimental results showed that the expression of the OsNAC050 gene in leaves was significantly higher than in other tissues both in seedlings and heading stage. At heading stage, OsNAC050 showed significant expression in leaves; the relative expression in leaves was 143.95-fold that of roots, 80-fold that of stems, and 77.94-fold that of sheaths (Figure 1A). The subcellular localization results showed that OsNAC050 was mainly expressed in the nucleus (Figure 1B). In response to low temperature (4°C) treatment, the expression of OsNAC050 increased to 3.3-fold the initial expression level after about 9 h, and the expression of OsNAC050 changed significantly under low temperature stress. From the analyses of OsNAC050 expression in the different treatments, we found that OsNAC050 expression was induced by various stimuli, and it was particularly sensitive to PEG, NaCl, low temperature, and GA3. Therefore, we hypothesized that OsNAC050 is involved in the response pathways for drought, high salt, low temperature, and GA3 stresses in rice (Figure 1C).
3.2. Targeted mutagenesis of OsNAC050
The Cas9 sgRNA was designed to target a sequence near the 5’ end of the first exon of OsNAC050. The design site and sequence are shown in Figure 2A. Following transformation of ‘Nipponbare’, a total of 15 T0-generation transgene-positive individual plants were obtained, and amplify the sequence near the mutation site for sequencing (Figure 2B). The results of sequencing and comparison showed that four of the 15 transgene-positive plants assayed carried mutations in OsNAC050; the mutation rate was 26.7% and the biallelic mutation rate was 6.7%. The sequence analysis results for the 15 transgenic positive plants tested are shown in Figure 2C. Among them, only OsNAC050-sgRNA01-09 was a homozygous biallelic mutant, with a single-base T insertion at the same position in both alleles, named osnac050 (T/T). OsNAC050-sgRNA01-01 has a 1bp insertion (A). OsNAC050-sgRNA01-04 has a 1-bp deletion (G) osnac050 and OsNAC050-sgRNA01-11 has a 5-bp deletion (ATGAG) in one allele. After T2 generation separation, screening homozygous mutants named osnac050 (-1/-1) and osnac050 (-5/-5). The mutants were screened and identified, and the non-transgenic and homozygous mutant plants were identified and propagated for subsequent experiments. All the following experiments were performed with T2-generation homozygous mutant plants.
Upstream (5’) and downstream primers (3’) were designed to amplify the 10 predicted most likely off-target sites, and the target fragments were amplified for Sanger sequencing analysis (Supplemental Figure 1). Among the 10 most likely off-target sites, no off-target mutations were identified.
3.3. Loss of function of OsNAC050 increases cold tolerance in rice seedlings
After five days of low temperature treatment, the mutants were showed that grow better than the wild type (Figure 3A). The experiment was repeated several times and the results were consistent, which indicated that knocking out OsNAC050 expression resulted in improved tolerance to low temperature in rice seedlings. The results of NBT and DAB staining for the WT and osnac050 mutant plants in the treatments showed that more superoxide ion (O2-) accumulated in the leaves of the WT lines compared to the mutants in the low temperature treatment, and the amount of blue precipitate was significantly higher in WT than in the mutant lines (Figure 3B). The relative water contents, chlorophyll contents, SOD (superoxide dismutase), POD (peroxidase), and CAT (catalase) activities were higher in the mutants, which also had less accumulation of MDA, compared to WT (Figure 3C-H). The O2- content in the WT plants under low temperature stress was 4-4.5 times that of the mutant plants, and the accumulation of H2O2 in the WT plants was 3.5-4 times that of the mutants, indicating that the mutants experienced less low-temperature-induced oxidative damage (Figure 3I-J). In conclusion, the rice OsNAC050 mutant lines were better at dealing with reactive oxygen species (ROS) caused by low temperature stress, which is consistent with the increased tolerance to low temperature in the mutant plants.
3.4. Knocking out OsNAC050 expression changed global gene expression in rice.
According to the genome-wide expression analysis of osnac050 mutants, the gene expression of the whole genome was greatly changed after knocking out the OsNAC050 gene (Figure 4 A, C). The number of differentially expressed genes (DEGs) in the samples was counted. Under normal conditions, a comparison of WT and osnac050(T/T) identified 6,605 DEGs, of which 4,294 were up-regulated and 2,311 were down-regulated. The WT vs osnac050(-5/-5) comparison had a total of 2,714 DEGs, in which 1,772 showed up-regulated expression and 942 were down-regulated (Figure 4B). In the low temperature treatment, there were 3,851 DEGs between WT and osnac050(T/T), including 1,425 up-regulated genes and 2,426 down-regulated genes. The comparison of WT and osnac050(-5/-5) under low temperature had 7,000 differentially expressed genes, of which 4,200 were up-regulated and 2,800 were down-regulated (Figure 4D). Ten genes were randomly selected to verify the reliability of our transcriptome data (Supplemental figure 2).
Under the same treatment conditions, the DEGs in the intersection of the WT vs osnac050(T/T) vs osnac050(-5/-5) comparison are more representative of the differential gene expression changes resulting from knocking out OsNAC050 (Supplemental figure 3). GO (Gene Ontology) annotation was performed on some of the DEGs. A total of 73 sub-categories were annotated, among which the largest number of DEGs was concentrated in the Molecular Function (MF) category. This provides some evidence that knocking out OsNAC050 expression may affect the association between cells. The terms ‘organic cyclic compound binding’ and ‘heterocyclic compound binding’ were greatly affected, indicating that OsNAC050 may affect the synthesis of organic compounds. The significant changes in the Cellular Component (CC) category are mainly in ‘membrane structure’ (membranes and membrane parts), so we can speculate that knocking out OsNAC050 will lead to changes in the main components of the cell membrane and organelle membranes. In the Biological Process (BP) GO category, the proportion of DEGs in ‘response to stimulus’ was high, which indicates that OsNAC050 could affect the stress response in plants (Figure 4E).
The results of the DEG annotation based on the KEGG (Kyoto Encyclopedia of Genes and Genomes) database are as follows: in the no treatment group, cell cycle, DNA replication and phenylpropanoid biosynthesis were the three most affected signaling pathways. The DEGs are also concentrated in the metabolic processes of ‘mismatch repair’, ‘purine metabolism’, and ‘base excision repair’ (Supplemental Figure 4). In the low temperature group, the DEGs were mainly concentrated in the ‘metabolic pathways’ and the ‘biosynthesis of secondary metabolites. In addition, there are DEGs enriched in ‘drug metabolism cytochrome P450’ and ‘photosynthesis’, both of which can directly affect light reception rice and thus affect the photosynthesis rate. At the same time, the two pathways of ‘starch and sucrose metabolism’ and ‘amino sugar and nucleotide sugar metabolism’ were partially enriched (Supplemental Figure 5). These results may suggest that osnac050 mutant plants can withstand low temperatures by increasing the biological processes involved in soluble sugar content and photosynthetic efficiency.
Analysis of the DEGs identified possible transcription-level responses to low temperature
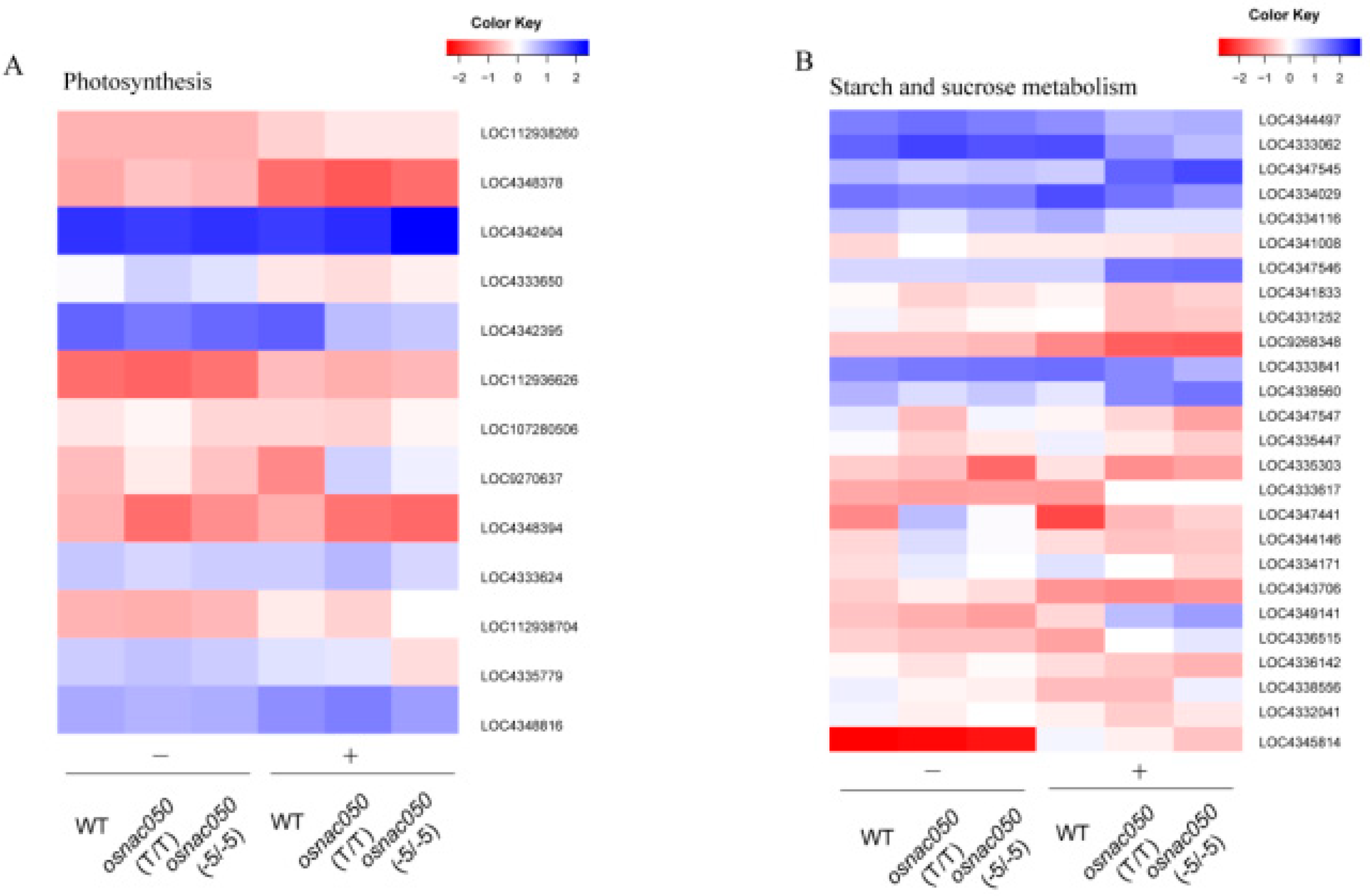
Because the DEGs in the KEGG analysis were annotated to the ‘photosynthesis’ and ‘starch and sucrose anabolic pathways’, and both play an important role in plant growth and development and the low temperature response. The significantly differential gene expression in the two signaling pathways was analyzed (Figure 5). There were 13 main genes in which the expression levels changed in ‘photosynthesis’. LOC4348378 participates in the photosynthetic electron transport process (GO: 0045156), transfers electrons in the cyclic electron transport pathway of photosynthesis, affects the formation of the chloroplast thylakoid membrane (GO: 0009535), and directly affects photosystem II (GO: 0009523). LOC4342404 is the main gene encoding the psaK subunit in the photosystem I reaction center in the chloroplast, and mainly affects photosystem I. LOC4333650, LOC4342395, LOC4333624, and LOC112938704 participate in the photosynthetic electron transport chain (GO: 0009767), affecting the photosystem II reaction center (GO: 0009539). LOC9270637 is involved in chlorophyll synthesis (GO:0009507) (Figure 5A). Taken together, these results suggest that OsNAC050 may regulate the response of genes that encode proteins in the photosystem I and photosystem II reaction centers to low temperature.
The anabolic pathway of starch and sucrose is a signaling mechanism that directly affects the response of plants to low temperature. The higher the soluble sugar content, the stronger the cold resistance of the plant. The expression of 26 genes in ‘starch and sucrose biological processes’ changed significantly in OsNAC05-knockout plants. Among them, the expression of LOC4344497 was up-regulated, so it may promote the fructokinase activity encoded by this gene (GO: 0008865). LOC4333062 regulates synthetic transferase activity (GO: 0016740), sucrose synthase activity (GO: 0016157), and transglycosylation activity (GO: 0016757), which directly affects sucrose metabolism (GO: 0005985); this gene is up-regulated at low temperatures and its expression promotes the above biological processes. LOC4334029 is involved in many pathways, and the protein it encodes regulates both cellulase activity (GO: 0008810) and hydrolase activity (GO: 0004553), which directly affects the process of polysaccharide catabolism (GO: 0000272) and the cellulose catabolism pathway (GO: 0030245). LOC4333841 and LOC4336142 are involved in a variety of carbohydrate metabolism and synthesis pathways, affecting hydrolase activity (GO: 0004553), β-galactosidase activity (GO: 0004565), β-mannosidase activity (GO: 0004567), β-mannosidase activity, β-Glucosidase activity (GO: 0008422), endoglucanase 1,3-β-D-glucosidase activity (GO: 0042973), pectin β-glucosidase activity (GO: 0050224), and cellobiose glucosidase activity (GO: 0080079). LOC4332041 is mainly involved in the starch catabolism pathway (GO: 0005983) and sucrose catabolism process (GO: 0005987). In summary, this study shows that knocking out OsNAC050 significantly changes the sucrose and starch metabolic pathways, and it is very likely that knocking out OsNAC050 expression can promote the accumulation of soluble sugars through the sucrose and starch metabolic synthesis pathway, thereby regulating the low temperature response in rice (Figure 5B).
4. Discussion
NAC TFs have been reported to be involved in the regulation of plant responses to various stresses, including cold, drought, and salinity (Yarra and Wei 2021; Ren et al. 2021). After the genomes of Arabidopsis, rice and other plant species were sequenced, NAC genes were systematically identified and shown to encode an important family of stress-related transcription factors (Mao et al. 2017; Liu et al. 2020). More than 100 genes have been classified as NAC family genes in rice by multiple research groups. Although several OsNAC genes have been functionally identified, the functions of many have yet to be determined. Therefore, why OsNAC TFs have so many functional homologues and how the functions of many OsNAC proteins are regulated through complex networks remains unclear. In addition, chilling injury has always been an environmental factor that limits the growth of rice, and exposure to low temperatures at the seedling stage can significantly inhibit rice development, resulting in a large reduction in grain yield (Chen et al. 2022; Kang et al. 2022). In our study, we showed that osnac050 mutant rice seedlings were tolerant to low temperatures after knocking out OsNAC050 expression. OsNAC050 expression is specifically induced in seedlings in response to cold stress conditions, but showed no obvious response to ABA, IAA, and hydrogen peroxide stresses. Under normal conditions, OsNAC050 is mainly expressed in leaves, and the relative expression is up to 100-fold higher than in other tissues such as roots, stems, and panicles. In addition, we found that the initial expression level decreased between one and three hours after exposure to low temperature, and then increased rapidly between six and twelve hours. Taken together, these results show that OsNAC050 may not be a transcription factor gene that responds initially and rapidly to cold stress, but rather may cause secondary and long-term responses that affect cold stress tolerance in rice (Figure 1).
After transforming ‘Nipponbare’ with the CRISPR/Cas9 vector designed to target a region of the first exon of OsNAC050, we identified only three mutants in the 15 transgenic plants. T2 generation plants were isolated for the three mutations and the homozygous mutant plants were selected for propagation and then used in the experiment. Our results showed that the seeds germination slower after knocking out OsNAC050 expression. Because we found that gibberellic acid (GA3) can inhibit expression of OsNAC050, we speculated that it might affect the gibberellin pathway, which in turn affects germination, making germination of osnac050 mutant seeds slower than in WT (Figure 2D).
Photosynthesis is probably the most basic and complex physiological process that occurs in plants, but it can be severely affected by adverse environmental conditions such as drought, salinity, and high temperature(Joo et al. 2022; Xiong et al. 2021). Photosynthesis involves sensitive pigments and photosystems as well as CO2 reduction pathways, which, along with chloroplast ultrastructure and electron transport systems, may be disrupted by abiotic stresses, which ultimately affect photosynthetic capacity(Gong et al. 2014). The accumulation of photosynthetic pigments such as chlorophyll, which is involved in capturing solar radiation to drive the photosynthetic machinery, is a potential biochemical indicator of drought tolerance in different crops(Liu, Quan and Bartels 2022). In this study, the osnac050 mutant plants exhibited higher chlorophyll contents (Figure 3D), which is very likely the reason for the increased tolerance to low temperatures (Bao et al. 2016). ROS, such as superoxide radicals (O2-), hydrogen peroxide (H2O2), monooxyl radicals (O21), alkoxyl radicals (RO), and hydroxyl radicals (OH-), which are highly reactive towards proteins, lipids, carbohydrates, and DNA, are produced in plants in response to various abiotic stresses including low temperature (Ramanjulu and Bartels 2002). MDA levels are a well-known indicator used to determine the degree of lipid peroxidation caused by ROS (reactive oxygen species) damage. In the present study, a higher MDA content was detected in WT plants after exposure to low temperature stress, indicating that these plants suffered more extensive oxidative damage to cell membranes. In response to ROS toxicity, plants have evolved efficient antioxidant mechanisms to adapt(Mohamadi Esboei et al. 2022). These mechanisms mainly include non-enzymatic scavengers as well as antioxidant enzymes (Jiang et al. 2021). Non-enzymatic antioxidants include ascorbic acid and reduced glutathione, and enzymatic antioxidants include superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), which are important in the drought response. Rapid accumulation under stress conditions can minimize oxidative damage(Toczewska et al. 2022). In the present study, mutant plants exhibited higher SOD, CAT, and POD activities than WT plants after cold stress (Figure 3). OsNAC050 enhances low temperature tolerance by stimulating the antioxidant defense system as measured by MDA content and SOD, CAT, and POD activities.
To investigate the molecular changes in the osnac50 mutant plants in more depth, we sequenced their transcriptomes and compared them to that of the wild-type ‘Nipponbare’ (Figure 4). Identification of differentially expressed genes between the mutant and WT transcriptomes showed that photosynthesis and the synthesis of starch and sucrose were the two metabolic pathways that were significantly altered in the mutants, and these are the main driving force for plant growth and can provide the necessary energy for plants to synthesize organic compounds(Di et al. 2022). The results of this study found that knocking out OsNAC050 mainly affected the synthesis of chloroplast thylakoid membranes and the transport of photosynthetic electron chains in photosystem II (Wang et al. 2022). The chloroplast is the main reaction center of photosynthesis. Chloroplasts have an inner and outer membrane, which provides a place for the attachment of enzymes and other proteins and electron transport(Li et al. 2021). Among the membrane structures, thylakoids are stacked membranous disks found within the chloroplast matrix and are where the light reactions occur, so the thylakoid membranes are essential for photosynthesis(Szechynska-Hebda et al. 2015; Lu et al. 2022). The photosynthetic electron transport chain is a series of systems responsible for connecting electrons on the photosynthetic membrane. This system can carry the main substrates of photosynthesis such as water or oxygen while transferring electrons(Gai et al. 2020; Yamori et al. 2011). Due to the obvious changes in these two signaling pathways, we can speculate that knocking out OsNAC050 expression may promote the light response in photosynthesis, thereby enhancing low temperature tolerance in the osnac050 mutants.
The synthesis of starch and sucrose are also important products of photosynthesis (Lopez-Gonzalez et al. 2022). Low temperature stress will lead to a reduction in enzyme activity, affecting the ability of plant cells to synthesize starch and sucrose (Apriyanto et al. 2022). At the same time, carbohydrate accumulation can help plants resist damage caused by low temperatures, and the sucrose component in the soluble sugars can also increase plant tolerance to low temperature exposure (Yu, Xu, et al. 2022; Xie et al. 2022). Our comparative transcriptomic analysis found that after knocking out OsNAC050, most of the genes encoding enzymes such as sucrose synthase, cellulase, hydrolase, and others were down-regulated, and these genes are mainly involved in various catabolic pathways, such as fructose catabolism, polysaccharide decomposition, cellulose hydrolysis, glucose decomposition, and starch catabolism (Shi et al. 2022; Nilholm et al. 2022). This implies that the osnac050 mutant plants respond to low temperature because the synthesis rate of enzymes in the catabolism processes is reduced, resulting in a decrease in the decomposition rate, thus leading to the accumulation of sucrose and starch, which in turn increases low temperature tolerance in the mutant plants (Figure 5)(Roth et al. 2022). This part of our study analyzed the phenotypic and transcriptome data of the osnac050 mutants in a deep exploration of the possible molecular mechanism of low temperature tolerance in rice, which will have important applications in rice breeding and research.
5. Conclusions
The above results show that the expression of the rice transcription factor gene OsNAC050 is induced by low temperature. After knocking out the expression of OsNAC050, the mutant plants displayed a low temperature-tolerant phenotype when exposed to low temperatures. A comparative analysis of the transcriptomes of ‘Nipponbare’ and the osnac50 mutants by RNA-seq was used to determine the possible mechanisms, and it was found that the ‘photosynthesis’ and ‘sucrose and glucose’ synthesis pathways were affected after knocking out the expression of OsNAC050. The differentially expressed genes in ‘photosynthesis’ included several photosynthetic pathways involving genes of the photosystem I and photosystem II active centers. In the sucrose and glucose anabolic pathways, we focused on genes involved in the decomposition of cellulose and sucrose, and we found that most of the metabolic pathways involved in the processes of decomposition and hydrolysis were changed. In conclusion, we analyzed the expression characteristics and gene function of the transcription factor gene OsNAC050, and determined its possible mechanism of action and regulatory target sites. The results of our study provide an important theoretical basis for breeding for low temperature tolerance in rice.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
This work was funded by Youth Science and Technology Innovation Project of Tianjin Academy of Agricultural Sciences (2022015).
Data Availability Statement
Transcriptome raw data has been uploaded to National Center for Biotechnology Information (NCBI) BioProject database; the accession number was PRJNA892163.
References
- Apriyanto, A., J. Compart, V. Zimmermann, S. Alseekh, A. R. Fernie, and J. Fettke. 2022. 'Indication that starch and sucrose are biomarkers for oil yield in oil palm (Elaeis guineensis Jacq.)', Food Chem, 393: 133361. [CrossRef]
- Bao, G., C. Zhuo, C. Qian, T. Xiao, Z. Guo, and S. Lu. 2016. 'Co-expression of NCED and ALO improves vitamin C level and tolerance to drought and chilling in transgenic tobacco and stylo plants', Plant Biotechnol J, 14: 206-14. [CrossRef]
- Chen, K., Z. Shi, S. Zhang, Y. Wang, X. Xia, Y. Jiang, S. Gull, L. Chen, H. Guo, T. Wu, H. Zhang, J. Liu, and W. Kong. 2022. 'Methylation and expression of rice NLR genes after low temperature stress', Gene, 845: 146830. [CrossRef]
- Di, Q., Y. Li, S. Li, A. Shi, M. Zhou, H. Ren, Y. Yan, C. He, J. Wang, M. Sun, and X. Yu. 2022. 'Photosynthesis Mediated by RBOH-Dependent Signaling Is Essential for Cold Stress Memory', Antioxidants (Basel), 11. [CrossRef]
- El Mannai, Y., K. Akabane, K. Hiratsu, N. Satoh-Nagasawa, and H. Wabiko. 2017. 'The NAC Transcription Factor Gene OsY37 (ONAC011) Promotes Leaf Senescence and Accelerates Heading Time in Rice', Int J Mol Sci, 18. [CrossRef]
- Gai, Z., L. Liu, J. Zhang, J. Liu, and L. Cai. 2020. 'Effects of exogenous alpha-oxoglutarate on proline accumulation, ammonium assimilation and photosynthesis of soybean seedling (Glycine max(L.) Merr.) exposed to cold stress', Sci Rep, 10: 17017. [CrossRef]
- Gong, B., X. Li, K. M. VandenLangenberg, D. Wen, S. Sun, M. Wei, Y. Li, F. Yang, Q. Shi, and X. Wang. 2014. 'Overexpression of S-adenosyl-L-methionine synthetase increased tomato tolerance to alkali stress through polyamine metabolism', Plant Biotechnol J, 12: 694-708. [CrossRef]
- Guo, Z., L. Cai, C. Liu, Z. Chen, S. Guan, W. Ma, and G. Pan. 2022. 'Low-temperature stress affects reactive oxygen species, osmotic adjustment substances, and antioxidants in rice (Oryza sativa L.) at the reproductive stage', Sci Rep, 12: 6224.
- Huang, L., Y. Hong, H. Zhang, D. Li, and F. Song. 2016. 'Rice NAC transcription factor ONAC095 plays opposite roles in drought and cold stress tolerance', BMC Plant Biol, 16: 203. [CrossRef]
- Jiang, Z., Q. Chen, L. Chen, H. Yang, M. Zhu, Y. Ding, W. Li, Z. Liu, Y. Jiang, and G. Li. 2021. 'Efficiency of Sucrose to Starch Metabolism Is Related to the Initiation of Inferior Grain Filling in Large Panicle Rice', Front Plant Sci, 12: 732867. [CrossRef]
- Joo, J. Y., M. S. Kim, Y. G. Cho, A. R. Fernie, and J. Sung. 2022. 'Transcriptional Comparison of Genes Associated with Photosynthesis, Photorespiration, and Photo-Assimilate Allocation and Metabolic Profiling of Rice Species', Int J Mol Sci, 23.
- Kang, M., G. Liu, Y. Zeng, J. Zhou, J. Shi, L. Tang, L. Liu, W. Cao, Y. Zhu, and B. Liu. 2022. 'Extreme Low-Temperature Stress Affects Nutritional Quality of Amino Acids in Rice', Front Plant Sci, 13: 905348. [CrossRef]
- Li, F., H. Deng, Y. Wang, X. Li, X. Chen, L. Liu, and H. Zhang. 2021. 'Potato growth, photosynthesis, yield, and quality response to regulated deficit drip irrigation under film mulching in a cold and arid environment', Sci Rep, 11: 15888.
- Liu, G., X. Li, S. Jin, X. Liu, L. Zhu, Y. Nie, and X. Zhang. 2014. 'Overexpression of rice NAC gene SNAC1 improves drought and salt tolerance by enhancing root development and reducing transpiration rate in transgenic cotton', PLoS One, 9: e86895. [CrossRef]
- Liu, X. H., Y. S. Lyu, W. Yang, Z. T. Yang, S. J. Lu, and J. X. Liu. 2020. 'A membrane-associated NAC transcription factor OsNTL3 is involved in thermotolerance in rice', Plant Biotechnol J, 18: 1317-29. [CrossRef]
- Liu, X., W. Quan, and D. Bartels. 2022. 'Stress memory responses and seed priming correlate with drought tolerance in plants: an overview', Planta, 255: 45. [CrossRef]
- Liu, X., X. Zhang, R. Cao, G. Jiao, S. Hu, G. Shao, Z. Sheng, L. Xie, S. Tang, X. Wei, and P. Hu. 2021. 'CDE4 encodes a pentatricopeptide repeat protein involved in chloroplast RNA splicing and affects chloroplast development under low-temperature conditions in rice', J Integr Plant Biol, 63: 1724-39. [CrossRef]
- Lopez-Gonzalez, C., S. Juarez-Colunga, S. Trachsel, N. Marsch-Martinez, C. S. Gillmor, and A. Tiessen. 2022. 'Analysis of Global Gene Expression in Maize (Zea mays) Vegetative and Reproductive Tissues That Differ in Accumulation of Starch and Sucrose', Plants (Basel), 11. [CrossRef]
- Lu, G., L. Wang, L. Zhou, X. Su, H. Guo, and H. Cheng. 2022. 'Overexpression of AmCBF1 enhances drought and cold stress tolerance, and improves photosynthesis in transgenic cotton', PeerJ, 10: e13422.
- Mao, C., S. Lu, B. Lv, B. Zhang, J. Shen, J. He, L. Luo, D. Xi, X. Chen, and F. Ming. 2017. 'A Rice NAC Transcription Factor Promotes Leaf Senescence via ABA Biosynthesis', Plant Physiol, 174: 1747-63. [CrossRef]
- Mohamadi Esboei, M., A. Ebrahimi, M. R. Amerian, and H. Alipour. 2022. 'Melatonin confers fenugreek tolerance to salinity stress by stimulating the biosynthesis processes of enzymatic, non-enzymatic antioxidants, and diosgenin content', Front Plant Sci, 13: 890613.
- Nilholm, C., L. Manoharan, B. Roth, M. D'Amato, and B. Ohlsson. 2022. 'A starch- and sucrose-reduced dietary intervention in irritable bowel syndrome patients produced a shift in gut microbiota composition along with changes in phylum, genus, and amplicon sequence variant abundances, without affecting the micro-RNA levels', United European Gastroenterol J, 10: 363-75.
- Ramanjulu, S., and D. Bartels. 2002. 'Drought- and desiccation-induced modulation of gene expression in plants', Plant Cell Environ, 25: 141-51.
- Ren, Y., Z. Huang, H. Jiang, Z. Wang, F. Wu, Y. Xiong, and J. Yao. 2021. 'A heat stress responsive NAC transcription factor heterodimer plays key roles in rice grain filling', J Exp Bot, 72: 2947-64. [CrossRef]
- Roth, B., J. Myllyvainio, M. D'Amato, E. Larsson, and B. Ohlsson. 2022. 'A Starch- and Sucrose-Reduced Diet in Irritable Bowel Syndrome Leads to Lower Circulating Levels of PAI-1 and Visfatin: A Randomized Controlled Study', Nutrients, 14. [CrossRef]
- Shen, J., B. Lv, L. Luo, J. He, C. Mao, D. Xi, and F. Ming. 2017. 'The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice', Sci Rep, 7: 40641. [CrossRef]
- Shi, W., Q. Ma, W. Yin, T. Liu, Y. Song, Y. Chen, L. Song, H. Sun, S. Hu, T. Liu, R. Jiang, D. Lv, B. Song, J. Wang, and X. Liu. 2022. 'The transcription factor StTINY3 enhances cold-induced sweetening resistance by coordinating starch resynthesis and sucrose hydrolysis in potato', J Exp Bot, 73: 4968-80. [CrossRef]
- Sun, Y., K. Song, M. Guo, H. Wu, X. Ji, L. Hou, X. Liu, and S. Lu. 2022. 'A NAC Transcription Factor from 'Sea Rice 86' Enhances Salt Tolerance by Promoting Hydrogen Sulfide Production in Rice Seedlings', Int J Mol Sci, 23. [CrossRef]
- Szechynska-Hebda, M., I. Wasek, G. Golebiowska-Pikania, E. Dubas, I. Zur, and M. Wedzony. 2015. 'Photosynthesis-dependent physiological and genetic crosstalk between cold acclimation and cold-induced resistance to fungal pathogens in triticale (Triticosecale Wittm.)', J Plant Physiol, 177: 30-43. [CrossRef]
- Takasaki, H., K. Maruyama, S. Kidokoro, Y. Ito, Y. Fujita, K. Shinozaki, K. Yamaguchi-Shinozaki, and K. Nakashima. 2010. 'The abiotic stress-responsive NAC-type transcription factor OsNAC5 regulates stress-inducible genes and stress tolerance in rice', Mol Genet Genomics, 284: 173-83. [CrossRef]
- Tang, X., X. Zheng, Y. Qi, D. Zhang, Y. Cheng, A. Tang, D. F. Voytas, and Y. Zhang. 2016. 'A Single Transcript CRISPR-Cas9 System for Efficient Genome Editing in Plants', Mol Plant, 9: 1088-91. [CrossRef]
- Toczewska, J., A. Zalewska, T. Konopka, and M. Maciejczyk. 2022. 'Enzymatic antioxidants activity in gingival crevicular fluid and saliva in advanced periodontitis', Oral Dis. [CrossRef]
- Wang, B., Z. Zhong, X. Wang, X. Han, D. Yu, C. Wang, W. Song, X. Zheng, C. Chen, and Y. Zhang. 2020. 'Knockout of the OsNAC006 Transcription Factor Causes Drought and Heat Sensitivity in Rice', Int J Mol Sci, 21. [CrossRef]
- Wang, W., J. Du, L. Chen, Y. Zeng, X. Tan, Q. Shi, X. Pan, Z. Wu, and Y. Zeng. 2021. 'Transcriptomic, proteomic, and physiological comparative analyses of flooding mitigation of the damage induced by low-temperature stress in direct seeded early indica rice at the seedling stage', BMC Genomics, 22: 176. [CrossRef]
- Wang, Z., J. Sun, X. Zu, J. Gong, H. Deng, R. Hang, X. Zhang, C. Liu, X. Deng, L. Luo, X. Wei, X. Song, and X. Cao. 2022. 'Pseudouridylation of chloroplast ribosomal RNA contributes to low temperature acclimation in rice', New Phytol. [CrossRef]
- Xie, F., H. Zhang, Z. Xiong, Y. Wu, and L. Ai. 2022. 'Effects and mechanism of sucrose on retrogradation, freeze-thaw stability, and texture of corn starch-tamarind seed polysaccharide complexes', J Food Sci, 87: 623-35.
- Xiong, H., L. Hua, I. Reyna-Llorens, Y. Shi, K. M. Chen, N. Smirnoff, J. Kromdijk, and J. M. Hibberd. 2021. 'Photosynthesis-independent production of reactive oxygen species in the rice bundle sheath during high light is mediated by NADPH oxidase', Proc Natl Acad Sci U S A, 118.
- Yamori, W., N. Sakata, Y. Suzuki, T. Shikanai, and A. Makino. 2011. 'Cyclic electron flow around photosystem I via chloroplast NAD(P)H dehydrogenase (NDH) complex performs a significant physiological role during photosynthesis and plant growth at low temperature in rice', Plant J, 68: 966-76. [CrossRef]
- Yang, Y., M. Jiang, J. Feng, C. Wu, W. Shan, J. Kuang, J. Chen, Z. Hu, and W. Lu. 2021. 'Transcriptome analysis of low-temperature-affected ripening revealed MYB transcription factors-mediated regulatory network in banana fruit', Food Res Int, 148: 110616. [CrossRef]
- Yarra, R., and W. Wei. 2021. 'The NAC-type transcription factor GmNAC20 improves cold, salinity tolerance, and lateral root formation in transgenic rice plants', Funct Integr Genomics, 21: 473-87.
- Yoo, S. D., Y. H. Cho, and J. Sheen. 2007. 'Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis', Nat Protoc, 2: 1565-72. [CrossRef]
- Yu, J., S. Xu, X. Liu, T. Li, D. Zhang, N. Teng, and Z. Wu. 2022. 'Starch Degradation and Sucrose Accumulation of Lily Bulbs after Cold Storage', Int J Mol Sci, 23. [CrossRef]
- Yu, Z., H. Liu, S. Mao, J. Zhang, J. Zhang, E. Yu, and L. Qu. 2022. 'Low-Temperature Thermal Desorption Effectively Mitigates Accumulation of Total Mercury and Methylmercury in Rice (Oryza sativa L.)', Bull Environ Contam Toxicol. [CrossRef]
- Zhang, J., Y. Sun, Z. Zhou, Y. Zhang, Y. Yang, X. Zan, X. Li, J. Wan, X. Gao, R. Chen, Z. Huang, L. Li, and Z. Xu. 2022. 'OsSCL30 overexpression reduces the tolerance of rice seedlings to low temperature, drought and salt', Sci Rep, 12: 8385.
- Zhang, X., Y. Long, X. Chen, B. Zhang, Y. Xin, L. Li, S. Cao, F. Liu, Z. Wang, H. Huang, D. Zhou, and J. Xia. 2021. 'A NAC transcription factor OsNAC3 positively regulates ABA response and salt tolerance in rice', BMC Plant Biol, 21: 546. [CrossRef]
- Zhao, F., T. Zhao, L. Deng, D. Lv, X. Zhang, X. Pan, J. Xu, and G. Long. 2017. 'Visualizing the Essential Role of Complete Virion Assembly Machinery in Efficient Hepatitis C Virus Cell-to-Cell Transmission by a Viral Infection-Activated Split-Intein-Mediated Reporter System', J Virol, 91. [CrossRef]
- Zhong, Z., Y. Zhang, Q. You, X. Tang, Q. Ren, S. Liu, L. Yang, Y. Wang, X. Liu, B. Liu, T. Zhang, X. Zheng, Y. Le, Y. Zhang, and Y. Qi. 2018. 'Plant Genome Editing Using FnCpf1 and LbCpf1 Nucleases at Redefined and Altered PAM Sites', Mol Plant, 11: 999-1002. [CrossRef]
Figure 1.
OsNAC050 expression profile analysis. (A) The expression of OsNACO50 in different tissues and organs of rice was detected and quantified using qRT-PCR. Samples of roots, sheaths, and leaves were collected from 4-week old seedlings, and roots, stems, sheaths, leaves, and ears were sampled during the reproductive growth period from plants prior to heading. The error bars represents the standard error (SE) based on three biological replicates. (B) Nuclear localization of the OsNAC050 protein in rice protoplasts. NLS, nuclear location signal. Scale bars=10 µm. (C) OsNAC050 expression levels in response to various abiotic stresses and hormone treatments. Four week old seedlings were treated with cold treatment (4 oC), heat treatment (42oC), PEG 6000 (20%, w/v), NaCl (200 mm), H2O2 (1%), IAA (100 µm), ABA (100 μm) And GA3 (100 µm). The relative expression levels of OsNAC050 were determined using qRT-PCR at the specified times. The error bars indicate that the SE is based on three independent biological replicates.
Figure 1.
OsNAC050 expression profile analysis. (A) The expression of OsNACO50 in different tissues and organs of rice was detected and quantified using qRT-PCR. Samples of roots, sheaths, and leaves were collected from 4-week old seedlings, and roots, stems, sheaths, leaves, and ears were sampled during the reproductive growth period from plants prior to heading. The error bars represents the standard error (SE) based on three biological replicates. (B) Nuclear localization of the OsNAC050 protein in rice protoplasts. NLS, nuclear location signal. Scale bars=10 µm. (C) OsNAC050 expression levels in response to various abiotic stresses and hormone treatments. Four week old seedlings were treated with cold treatment (4 oC), heat treatment (42oC), PEG 6000 (20%, w/v), NaCl (200 mm), H2O2 (1%), IAA (100 µm), ABA (100 μm) And GA3 (100 µm). The relative expression levels of OsNAC050 were determined using qRT-PCR at the specified times. The error bars indicate that the SE is based on three independent biological replicates.
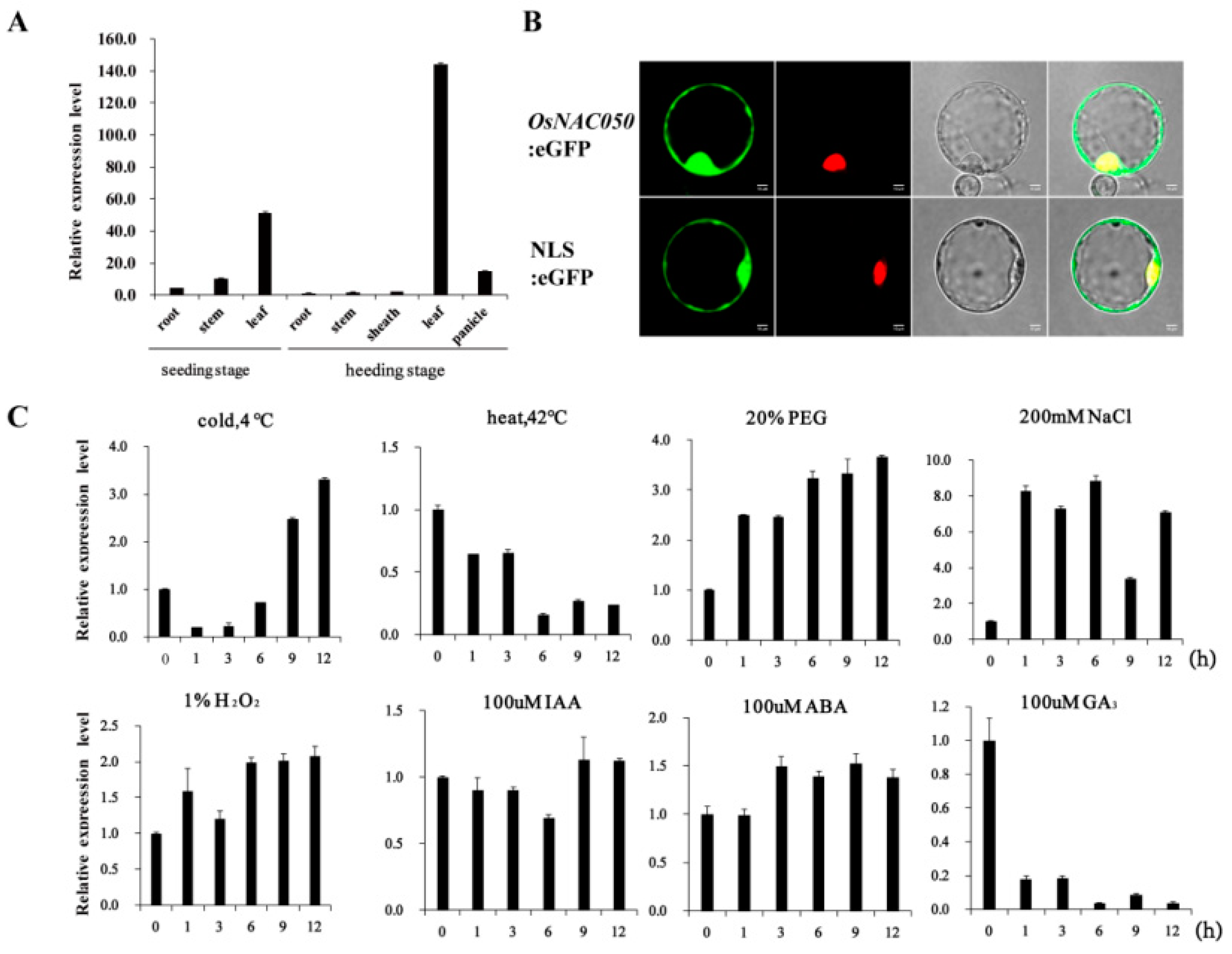
Figure 2.
Targeted mutagenesis using CRISPR-Cas9 gene editing. (A) sgRNA site design in the first exon of OsNAC050. The target sequence is shown in black, and the PAM (protospacer adjacent motif) site is shown in red. (B) Gel electrophoresis of amplified DNA fragments from 15 independent OsNAC050-sgRNA01 T0 -generation lines for sanger sequencing to identify the mutant genotype. WT, wild type ‘Nipponbare’. (C) Sanger sequencing of the targets site of OsNAC050-sgRNA01 T0 -generation lines. (D) Phenotypic analysis of OsNAC050 mutant lines under germinating period. Experiments showthatthegerminationspeedofmutantsdecreases.
Figure 2.
Targeted mutagenesis using CRISPR-Cas9 gene editing. (A) sgRNA site design in the first exon of OsNAC050. The target sequence is shown in black, and the PAM (protospacer adjacent motif) site is shown in red. (B) Gel electrophoresis of amplified DNA fragments from 15 independent OsNAC050-sgRNA01 T0 -generation lines for sanger sequencing to identify the mutant genotype. WT, wild type ‘Nipponbare’. (C) Sanger sequencing of the targets site of OsNAC050-sgRNA01 T0 -generation lines. (D) Phenotypic analysis of OsNAC050 mutant lines under germinating period. Experiments showthatthegerminationspeedofmutantsdecreases.
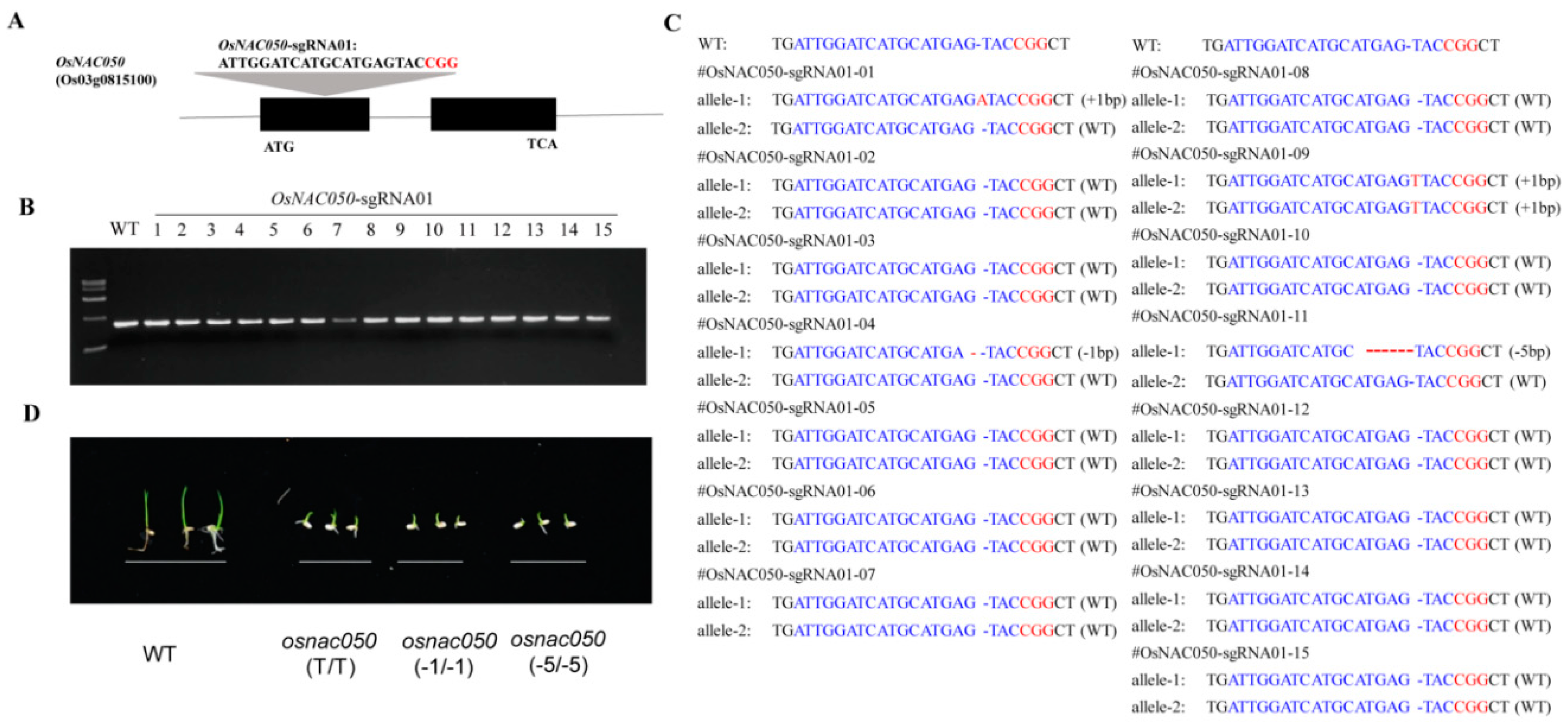
Figure 3.
Cold-tolerance in the osnac050 mutants. (A) Phenotypes of osnac050 T2 -generation mutant plants exposed to cold stress. (B) Levels of O2- and H2O2 in WT and osnac050 T2 -generation mutant lines subjected to cold stress. Rice leaf samples were stained separately with nitroblue tetrazolium (NBT) and diaminobenzidine (DAB). (C) Determination of relative water content and (D) chlorophyll content after 5-days of low temperature stress. (E-H) Malondialdehyde (MDA) content (E), Superoxide dismutase (SOD) activity (F), Peroxidase (POD) activity (G), and Catalase (CAT) activity after low temperature stress (H). O2- production rate and (J) H2O2 content after low temperature stress. Bars represent the mean ± SE of three independent experiments. ∗ and ∗ ∗ represent significant differences at p.
Figure 3.
Cold-tolerance in the osnac050 mutants. (A) Phenotypes of osnac050 T2 -generation mutant plants exposed to cold stress. (B) Levels of O2- and H2O2 in WT and osnac050 T2 -generation mutant lines subjected to cold stress. Rice leaf samples were stained separately with nitroblue tetrazolium (NBT) and diaminobenzidine (DAB). (C) Determination of relative water content and (D) chlorophyll content after 5-days of low temperature stress. (E-H) Malondialdehyde (MDA) content (E), Superoxide dismutase (SOD) activity (F), Peroxidase (POD) activity (G), and Catalase (CAT) activity after low temperature stress (H). O2- production rate and (J) H2O2 content after low temperature stress. Bars represent the mean ± SE of three independent experiments. ∗ and ∗ ∗ represent significant differences at p.
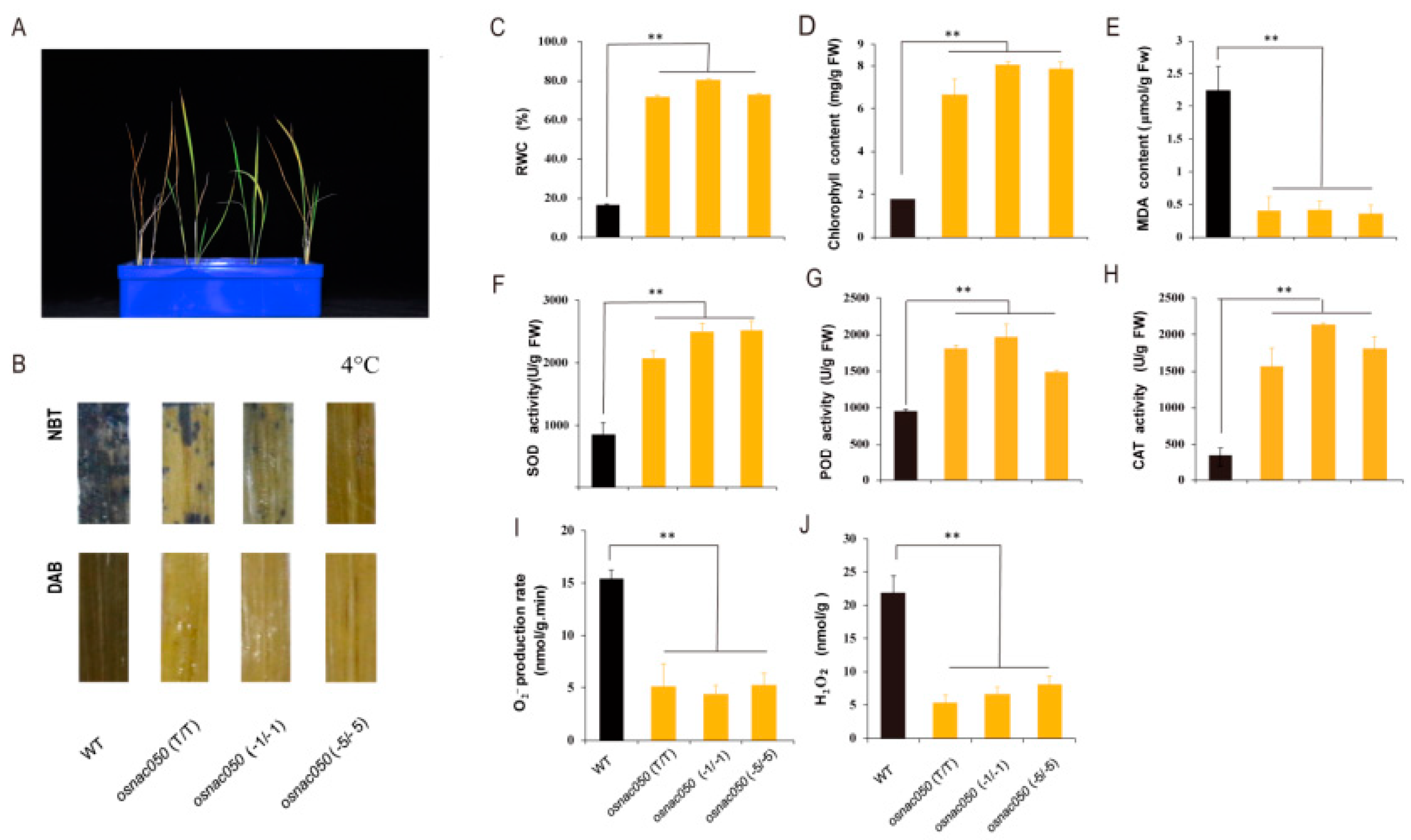
Figure 4.
Global gene expression changes in OsNAC050-knockout rice plants. (A) Cluster analysis of the most important differentially expressed genes (DEGs) between the WT and osnac050 T2 -generation mutant lines. Targeted knockout of OsNAC050 expression resulted in changes in gene expression, physiology, growth, and development compared with ‘Nipponbare’ and the control without low temperature stress treatment. Relative expression corresponds to the log2 (FPKM) value of each gene. WT, osnac050(T/T), and osnac050(-5/-5) T2 mutant line, expression profiles were obtained from RNA-seq data. Total RNA was extracted from a mixture of three different plants. Genes with p-value < 0.05 and foldchange >2 are advised to define differentially expressed genes (DEGs). (B) DEGs in the WT vs. osnac050(T/T) and WT vs. osnac050(-5/-5) comparisons without cold stress. (C) Cluster analysis of the most important DEGs between the WT and osnac050 T2 mutant lines after cold stress. (D) DEGs in the WT vs. osnac050(T/T) and WT vs. osnac050(-5/-5) lines after cold stress. (E) Gene ontology (GO) classification of DEGs between the WT and two osnac050 mutants under normal (red) and cold stress conditions (blue) into the three main GO classes CC (Cellular Component), MF (Molecular Function), and BP (Biological Process). The x-axis displays the GO terms selected by the user, and the y-axis displays the percentage of genes (specific genes divided by total genes.
Figure 4.
Global gene expression changes in OsNAC050-knockout rice plants. (A) Cluster analysis of the most important differentially expressed genes (DEGs) between the WT and osnac050 T2 -generation mutant lines. Targeted knockout of OsNAC050 expression resulted in changes in gene expression, physiology, growth, and development compared with ‘Nipponbare’ and the control without low temperature stress treatment. Relative expression corresponds to the log2 (FPKM) value of each gene. WT, osnac050(T/T), and osnac050(-5/-5) T2 mutant line, expression profiles were obtained from RNA-seq data. Total RNA was extracted from a mixture of three different plants. Genes with p-value < 0.05 and foldchange >2 are advised to define differentially expressed genes (DEGs). (B) DEGs in the WT vs. osnac050(T/T) and WT vs. osnac050(-5/-5) comparisons without cold stress. (C) Cluster analysis of the most important DEGs between the WT and osnac050 T2 mutant lines after cold stress. (D) DEGs in the WT vs. osnac050(T/T) and WT vs. osnac050(-5/-5) lines after cold stress. (E) Gene ontology (GO) classification of DEGs between the WT and two osnac050 mutants under normal (red) and cold stress conditions (blue) into the three main GO classes CC (Cellular Component), MF (Molecular Function), and BP (Biological Process). The x-axis displays the GO terms selected by the user, and the y-axis displays the percentage of genes (specific genes divided by total genes.
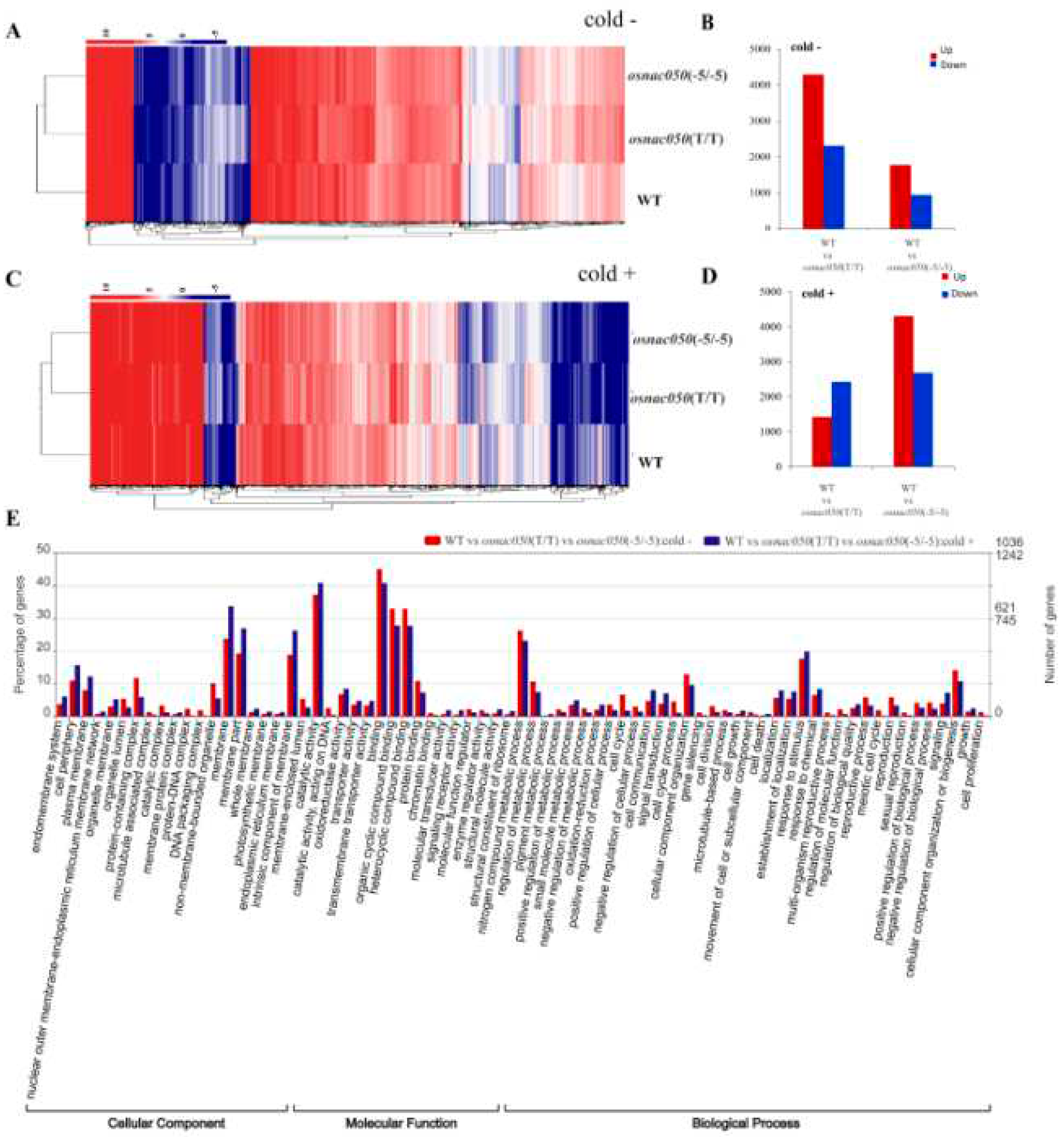
Figure 5.
Transcriptomic analysis of system regulatory genes in the WT and osnac050 mutant lines under low temperature stress. (A and B) Heatmaps showing changes in gene expression in the DEGs in two major pathways between WT and two osnac050 mutants with and without cold treatment. (A) Photosynthesis. (B) Starch and sucrose metabolism. Log2 fold-change (FC) values for DEGs in WT and osnac050(T/T) and osnac050(-5/-5) mutant lines are shown before (cold-) and after (cold+) low temperature treatment. Genes with p-value < 0.05 and fold change >2 are advised to define differentially expressed genes.
Figure 5.
Transcriptomic analysis of system regulatory genes in the WT and osnac050 mutant lines under low temperature stress. (A and B) Heatmaps showing changes in gene expression in the DEGs in two major pathways between WT and two osnac050 mutants with and without cold treatment. (A) Photosynthesis. (B) Starch and sucrose metabolism. Log2 fold-change (FC) values for DEGs in WT and osnac050(T/T) and osnac050(-5/-5) mutant lines are shown before (cold-) and after (cold+) low temperature treatment. Genes with p-value < 0.05 and fold change >2 are advised to define differentially expressed genes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



