
Submitted:
08 May 2023
Posted:
09 May 2023
You are already at the latest version
Abstract
Begomovirus infection in plant causes a great economic loss in many countries every year. Increasing rate of the infectivity of this single stranded DNA virus forces to study of its transmission in detail for the necessary researches regarding the control of this disease. This pathogenic virus is whitefly vector borne and transferred from one plant to another plant during the suckling of phloem sap by that vector. Viral transmissibility through this insect vector may depend on the genetic variations within the cryptic species groups. Two distinct categories of begomoviruses viz., bipartite and monopartite types consist of different genome organization. Virion particles modify the intracellular environment of the host according to their need of replication and survival. Viral replication takes place through RCR as well as RDR methods. Interplay among host, vector and pathogen is crucial for the establishment of infection. Several endosymbiotic organisms living within the insect vector also have vital role here. Being infected with this virus, host plant responds with defensive activities like TGS, PTGS, autophagy, hormonal regulation, metabolic alteration etc. However, virus also counteracts those through the manipulation of several pathways of cellular events. It is necessary to study in different directions and utilize advanced molecular biological techniques to develop begomovirus resistance within plants.
Keywords:
Begomovirus
; Whitefly
; Viral genome
; Viral transmission
; Host-vector-pathogen interaction
; Host defense
1. Introduction
Different types of viral infections in plants are great threat in sustainable agriculture as the worldwide demand for food is increasing day by day. Rapid transmissibility of virion particles, change in climatic condition and global trading cause sudden spreading of diseases within a vast area. Geminiviruses, belongs to the family Geminiviridae, infect a wide range of economically important plants and attribute a major crop loss all over the world. Geminiviruses are single-stranded DNA viruses, having twinned icosahedral structure, cause diseases like bright yellow mosaic, yellow mosaic, yellow mottle, leaf curling etc. Geminivirus interferes host cell machinery with the modification of gene expression, cell signaling, metabolic reprogramming etc. Being transmitted through insect vectors, those spread into different hosts like tomato, cotton, maize, potato, beet, wheat, cassava etc. Based on the genetics, phylogenetic relationships, host interaction and insect vectors, geminiviruses are categorized into Becurtovirus, Begmovirus, Capulavirus, Curtovirus, Eragrovirus, Grablovirus, Masterovirus, Topocuvirus and Turncurtovirus [1,2]. Among these, begomovirus are one of the largest group, transmitted through whiteflies (Bamisia tabaci) which sit abaxial surface of the leaf and suck phloem sap. These tiny Hemipteran insect vectors are the members of Aleyrodidae family. Some considerable genetic and behavioral variations have been found in different haplotypes of whitefly vectors. On the basis of genetic variation in cytochrome oxidase unit, those groups of population have been considered as a cryptic species complex where the members are reproductively isolated. Several cryptic species like Middle East-Asia Minor 1 (MEAM1), Asia-I, Asia I-India, Asia II-1, Asia II-5, Asia II-7, Asia II-8, Asia II-11, and China-3, Mediterranean (MED), NW1, NW2 are reported already. Co-adaptation of those variants may influence on transmission efficiency [3,4]. Begomovirus mainly infects dicot plants bearing edible vegetables, distributed within tropical, sub-tropical and mild temperate climatic zone. The business oriented production of vegetables improves food security, income opportunities, poverty reduction along with regional development but the annual economic loss from this viral disease affects livelihood directly [5]. Additionally, some non-edible economically important plants like tobacco, cotton are also infected with the begomoviruses. Abutilon mosaic disease, cucumber mosaic disease, sweet potato leaf curl disease, cotton leaf curl disease, bean golden mosaic disease, tomato curl disease, chilli leaf curl disease etc. are commonly found in our country [6]. In spite of utilizing the host DNA polymerase having proof-reading activity, begomoviruses exhibit significant level of genetic variation and nucleotide substitution. Frequent recombination and re-assortment of the viral genome, increase their adaptive fitness to invade wide range of hosts in the different ecological niche [7]. All those events compel to control the begomovirus as per the requirement for preventing financial condition of the countries from this viral infection. For this, it is important to understand the biology of this viral transmission critically. Most of the literatures have represented the characterization of begomovirus, their genomic variation and diversification, infectivity in different plants, several gene expression profiles in plants etc. However, very little information is available about the whitefly mediated transmission of begomovirus together with the respective host response. In this review, we have focused on the genomic structure and mechanism of transmission of begomovirus, interaction among plant-vector-pathogen required for viral infection and specific stress responses inside the plant body with some other aspects.
2. Genome Organization and Transmission of Begomovirus
Begomoviruses are categorized into two groups, viz., bipartite begomovirus (e.g. Tomato Leaf Curl New Delhi Virus) and monopartite begomovirus (e.g. Tomato Mottel Leaf Curl Virus). Bipartite begomovirus contains two circular single-stranded DNA molecules (≈2800 nucleotides), known as DNA-A and DNA-B whereas monopartite begmovirus contains a single DNA molecule which is similar to DNA-A of bipartite begomovirus [8]. DNA-A encodes replication associated protein (REP), transcription activator protein (TrAP), replication enhancer protein (REn), coat protein (CP), pre-coat protein, and viral suppressors of RNA silencing (VSRs) protein. On the other hand, DNA-B encodes viral movement protein (MP) and nuclear shuttle protein (NSP). Several genes are present in the two types of DNAs for executing the specific workflow. For example, in case of Tomato Leaf Curl New Delhi virus eight different genes viz., AV1, AV2, AV3, AC1, AC2 , AC3, AC4 and AC5 are found within DNA-A contributing the abovementioned functions whereas DNA-B contains BV1 and BC1 genes to perform the respective tasks. Both the genomic components share a common region (CR) which controls gene expression and replication initiation. Sometimes, extra single-stranded DNA molecules are found in monopartite begmovinuses, known as alpha-satellites and beta-satellites. Satellite DNA molecules often act as VSRs and mediate host DNA suppression. For replication, coating and movement, monopartite begomovirus needs a helper virus in many cases. Genes present in alpha-satellite, are responsible for the synthesis of replication initiator protein and which are in beta-satellite are the pathogenicity determinant [9,10]. Spreading of begomovirus through white flies occurs in a circulative persistent manner. Whitefly inserts its stylets in the vascular tissues of the plant to ingest the phloem sap and inoculate the virion particles into the host. Virions are then translocated through their digestive system, haemolymph and salivary gland, and subsequently, transmitted into the phloem sap of another plants ingested by the insect later. Coat protein located on the virion surface, is essential for the viral invasion into the female reproductive system of the insect vector, and then transovarially transmitted to the progeny, with variable infectivity potential [4,11]. Virion particles modulate the host cell cycle and transcriptional events to make the surrounding microenvironment favorable for its replication. Circular genomic DNA of begomovirus, principally, replicates through the rolling circle replication (RCR) mechanism. When transmitted through the whitefly, the viral genome enters into the host cell nucleus and three stage based replication process takes place. Initially, conjugated with the host cellular factors, the single-stranded viral DNA is converted into supercoiled double-stranded DNA intermediate with the help of host DNA polymerase. This leads to the transcription of several viral factors. After this, double-stranded DNA acts as a template for the synthesis of single-stranded viral DNA through rolling circle replication mode utilizing the both viral and host factors. Synthesized single-stranded DNA then re-enters into the replication pool and assists viral assembly and spreading. Viral DNA molecules are packed into host nucleosome and formation of minichromosomes take place within the host nucleus. Those minichromosomes are used as a template for viral transcription [12,13]. Beside this, begomovirus also replicates through recombination driven replication (RDR) mechanism where recombination is closely associated with the replication. This mechanism is able to recover the viral DNA fragments resulting from incomplete synthesis or nucleolytic attack [14,15]. Viral replication and transcription are encoded by the early activated genes followed by late activated genes encoding the encapsidation and movement. With the alteration of host gene expression and the regulation of cell signaling, viruses increase the disease severity after establishing the successful infection [16]. Infection mechanism of begomovirus in plant cell is diagrammatically represented in Figure 1.
3. Interaction among Plant, Whitefly and Begomovirus
The tripartite interactions among host, vector and pathogen employ significant impacts on the population dynamics of the whitefly and the disease development within host. It seems to be that the relationship between whitefly and begomovirus is mutualistic [17]. After infection, begomovirus alters the nutrition status of plant with the suppression of plant defense against the whitefly in such way that is suitable for that insect vector [18]. From the comparative analysis of honeydew excreted by viruliferous and non-viruliferous whitefly, it was speculated that begomovirus helps to improve the nutritional assimilation (such as essential amino acids and sugars) of the vector and likewise enhances its performance level [19]. In addition, endosymbiotic bacteria are often involved in this interaction. They are ubiquitously found in Hemipteran insects. The endosymbionts grown within Bamisia tabaci, (e.g. Candidatus Portiera aleyrodidarum under Oceanospirillales) and vertically transmitted through reproduction are known as primary endosymbionts. Primary endosymbionts supply essential amino acids and carotenoids to their whitefly host. Secondary endosymbionts are also important for the activities of the vector. They are associated later and often transmitted horizontally. Seven secondary endosymbionts have been found within Bamisia tabaci. These are Candidatus Fritschea bemisiae (Chlamydiales), Candidatus Cardinium hertigii (Bacteroidales), Candidatus Hemipteriphilus asiaticus (Rickettsiales), Candidatus Hamiltonella defense (Enterobacteriales), Arsenophonus spp. (Enterobacteriales) and Wolbachia spp. (Rickettsiales) [17]. Some endosymbionts may regulate defense system of the host plant. This regulation seems to influence the vector-pathogen interaction as well as pathogen transmission. As the required amino acids are provided to the vector by the symbionts, the alteration in the composition of symbiotic association may affect the survival of whitefly and plant mediated whitefly-begomovirus interaction. Apart from these, co-occurrence of other pathogens, herbivores and natural enemies together with whiteflies in begomovirus infected plants may interfere with whitefly–begomovirus interaction. Environmental heterogeneity and several abiotic factors like temperature, humidity, soil chemistry etc. also have influence on this tripartite interaction [17,20]. Several aspects need to be investigated for the better understanding of this interaction. These are the followings: role of insect proteins and endosymbionts for the viral transmission efficiency, manipulation of physiological aspects of plant, metabolic regulation and the role of pathogenicity factors and other proteins which modulate the physiology of plant and whitefly behaviour.
4. Host Plant Responses against the Begomovirus Infection
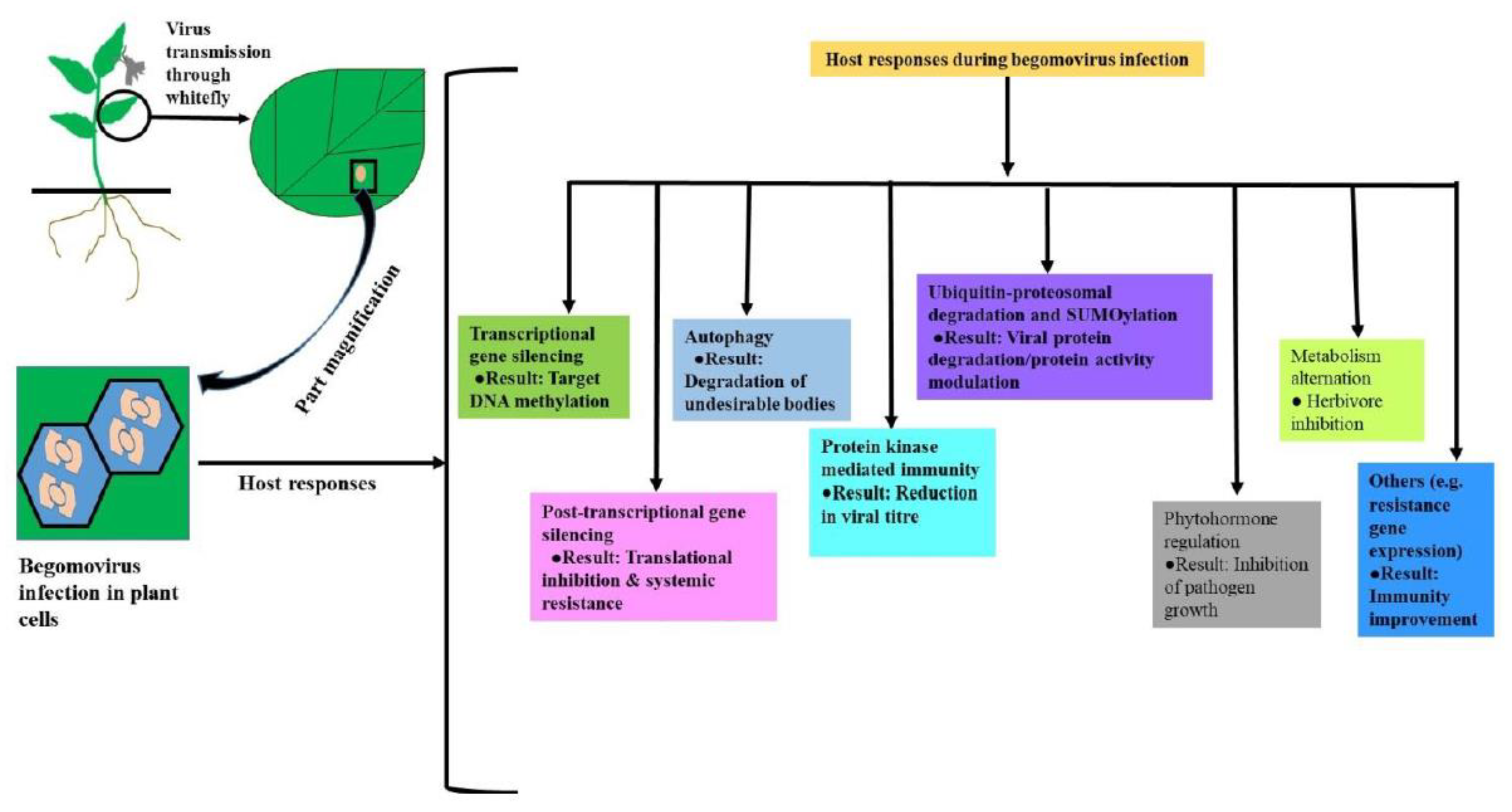
Host plants acquire different strategies to combat pathogenicity after viral invasion. Some of the most important defensive mechanisms like (a) transcriptional gene silencing (TGS), (b) post-transcriptional gene silencing (PTGS), (c) autophagy, (d) protein kinase-mediated immunity, (e) Ubiquitin–proteasomal degradation and SUMOylation, (f) regulation of phytohormones, (g) alternations in primary and secondary metabolism of plant [21] are discussed here.
4.1. Transcriptional Gene Silencing (TGS)
With the help of host cell machinery, the viral single stranded DNA replicates and binds with histones of the host, to form minichromosomes. Plants suppress this minichromosome formation by RNA directed DNA methylation apparatus and ultimately silence the viral gene expression through transcriptional gene silencing (TGS) in response to the viral invasion into the cell. Small non-coding RNAs like siRNA and miRNA play significant role in this type of silencing. RNA directed DNA methylation is mediated by the RNA polymerase IV & V of the host cell. Those RNA polymerases generate siRNA (24 nt) to elevate the target DNA methylation de novo. RNA polymerase IV catalyzes the formation of single stranded non-coding transcripts from viral chromatin. From this, double stranded RNA is formed by RNA dependent RNA polymerase with CLASSY 1 manner. Double stranded RNAs are then diced by DICER-LIKE 3 (DCL-3) ribonucleases and generate 24-nt siRNAs duplexes. Those are stabilized by HUAENHANCER1 (HEN1) and loaded to the RNA-induced silencing complex (RISC) containing ARGONAUTE 4 (AGO4)/AGO-6, later. Transcription of RNA polymerase V occurs with the help of DDR complex where DEFECTIVE IN RNA DIRECTED DNA METHYLATION 1 (DRD1), DEFECTIVE IN MERISTEM SILENCING 3 (DMS3), REQUIRED FOR DNA METHYLATION 1 (RDM1) and DEFECTIVE IN MERISTEM SILENCING 4 (DMS4) are present [22]. Transcripts formed by RNA polymerase V, base pair with siRNA present in AGO4-RISC complex. This is stabilized through the association of AGO4 with carboxy-terminal tail of Nuclear RNA polymerase E (NRPE1) and KOW DOMAIN-CONTAINING TRANSCRIPTION FACTOR 1 (KTF1). AGO4 then binds to RDM1 and involves cytosine methyl transferase like DOMAINS REARRANGED METHYL TRANSFERASE 2 (DRDM2) for de novo methylation of viral DNA. Histone methyltransferase KRYPTONITE (KYP) and DNA methyltransferase CHROMOMETHYLTRANSFERASE 3 have important role in maintaining the process of Transcriptional Gene Silencing [23]. However, begomovirus encodes TGS suppressors to overcome the Transcriptional Gene Silencing. For example, in bipartite begomovirus, transactivator protein (TrAP) encoded by V2 of TYLCV, acts as TGS suppressor and promotes virulence [24]. To carry out TGS, miRNA works through the following steps such as primary transcription of miRNA from MIRNA genes, with the help of RNA polymerase II, followed by its processing with Dicer-like proteins. After this, methylation of miRNA and its association with RISC, occur. Then the miRNA binds to the target mRNA and degrades it [25]. To inhibit this, V2 protein of often directly binds to miRNA and interfere with mRNA homeostasis and creates developmental abnormalities within host plant [26].
4.2. Post-Transcriptional Gene Silencing (PTGS)
The post-transcriptional gene silencing mainly occurs through the targeting of viral RNA transcript by siRNA-meditated silencing pathway. PTGS starts to target the double stranded RNA fragments derived either from bidirectional transcription or from secondary hairpin structures of viral RNA. Those double stranded RNAs are recognized by Dicer-like protein (DCL) and double stranded RNA binding protein (DRB) and processed to form siRNAs (21-24 nt). Methylation of 3’ end of the siRNA by HUA ENHANCER 1 (HEN1) protein, protects it from 3’-5’ exonucleolytic degradation and uridylation. Simultaneously, the siRNA duplexes are recruited onto AGO4 to favour the targeting and the formation of RISC. This results the mRNA degradation with cytoplasmic endonucleases or translational inhibition. Further progress of PTGS occurs at the systemic site by primary siRNAs to develop systemic resistance [21,27]. Several suppressors are developed by viruses for lessening this immunity responses of host through interfering at different level such as PTGS inactivation, interfering siRNA biogenesis plus host defense inhibition [28]. For example, BV1 encodes Nuclear Shuttle Protein (NSP) which induces ASYMMETRIC LEAVES 2 (AS2) expression through which enhances decapping activity of DECAPPING 2 (DCP2), mRNA turnover and disturbing siRNA accumulation and RNA silencing process of host. Upregulation of a calmodulin-like protein (CaM) by βC1, inhibits the degradation of suppressor proteins and reduce RDR6 activity and eventually affects the anti-viral activity of host [29].
4.3. Autophagy
Autophagy is a natural, conserved process for degrading undesirable, dysfunctional bodies present in the cytoplasm. It has significant potentiality to develop innate and adaptive immunity in plant against viral infection. βC1 protein of CLCuMuB, interacts with the autophagy related protein NbATG8 and become degraded as per the reports of in vitro and in vivo studies. Mutation causing the disruption of βC1- NbATG8 interaction promotes susceptibility to the viral infection. Recently, begomoviruses are reported to have the ability to manipulate autophagy mediated host defense with inducing autophagy of host factors related to different defensive pathway [21,30].
4.4. Protein Kinase-Mediated Immunity
Protein kinases are important components for several physiological processes including defensive response. Commonly, SnRK1, MAP kinases and receptor like kinases (RLKs) direct the host responses during virus infection. SUCROSE NON-FERMENTING1-related protein kinase 1 (SnRK1) functions in stress response [21]. During TYLCCNB infection in Nicotiana benthamiana, SnRK1 phosphorylates βC1 protein at serine 33 and threonine 78 residues through which negatively impact on viral titre in plant [31]. To recover this, viral TrAP protein often interacts with SnRK1 and inhibits its antiviral activity [32]. Mitogen-activated protein kinase (MAPKs) are known to be involved in signal transduction and defense against different biotic and abiotic stresses. The role of MAPK in antiviral defense response are reported also beside the evidences showing antibacterial and antifungal immunity development in plant. It was found that MAPK1 upregulated the salicylic acid (SA) mediated expression of pathogenesis-related (PR) genes in Vigna mungo [33]. In MAPK3 silenced tomato plant, poor expression of genes involved SA/JA mediated defensive pathway were found [34]. To survive in this adverse condition, βC1 protein of TYLCCNV interacts with MAPK inhibitors for limiting its antiviral activity [35]. Receptor-like kinases (RLKs) are transmembrane proteins transducing extracellular signals through their specific ligand binding domains, a membrane-spanning region, and cytoplasmic serine-threonine kinase domain, and also regulate the innate immunity against several pathogens [36]. One RLK (namely NIK), targets viral NSP and mediates immune response against begomovirus [37]. However, this type of resistance is limited by the viral NSP as it interacts with NIKs and suppresses its activity [38].
4.5. Ubiquitin–Proteasomal Degradation and SUMOylation
In ubiquitin–proteasomal degradation system, ubiquitin protein is conjugated to the lysine moiety of a target protein and ultimately directs the target protein to 26S proteosomal degradation. Three specialized enzymes are required to carry out the sequential events of this post-transcriptional modification. These are E1 (ubiquitin activating enzyme), E2 (ubiquitin conjugating enzyme), E3 (ubiquitin ligase) [39]. Degradation of viral protein through ubiquitin-proteosomal pathway after begomovirus infection in host cell is reported by authors. A tobacco protein with RING-finger domain, named NtRFP1, functions as E3 ligase, intervenes βC1 ubiquitination and diminishes the β-satellite mediated disease symptom generation. Virus often utilize the ubiquitin-proteosomal degradation to destroy cellular proteins acting as restriction factors for viral replication and survival [40]. SUMOylation is a similar type of post-transcriptional modification involving small ubiquitin like modifier (SUMO) in the ligation to lysine moiety of target protein and thereby modulation of that protein activity. In case of Synedrella yellow vein clearing virus (SyYVCV) infection, C-terminal SUMO interacting motif of βC1, binds to cellular components and protein degradation occurs. To counter this, virus alters the SUMOylation patterns of specific host factors for creating favourable environment for viral replication [41].
4.6. Regulation of Phytohormones
Phytohormones like salicylic acid (SA), jasmonic acid (JA), and ethylene generate significant host defense responses against plant pathogens. Anti-viral activity of auxin, cytokinin, gibberellic acid, brassinosteroids and abscisic acid are yet to be explored [21,42]. In TYLCSV infected tomato plant, upregulation of JA responsive gene like COI1 was reported. Sometimes βC1 interacts with MYC2 to suppress JA-mediated responses [43,44]. CaLCuV infected Arabidopsis thaliana has displayed activation of SA pathway [45,46]. However, the reduction of SA biosynthesis has been reported in during TYLCSV infection in S. lycopersicum. Auxin signalling involves camalexins and glucosinolates that selectively inhibits several pathogenic growth but AC4 protein of ToLCNDV reduces the endogenous auxin level and disrupts signalling cascade by interacting with CYP450 monooxygenase [47,48]. More than one hormone may work in concert in response to viral invasion but viruses try to counter those with the manipulation of respective pathways.
4.7. Alternations in Primary and Secondary Metabolism of Plant
Viral infection causes alteration in metabolism inside the plant body. Beet mild curly top virus (BMCTV) induces a high level of glucose, fructose, galactose, and myoinositol in chilli and utilize those for capsid formation [49]. Alteration in metabolite content favours the abundance, fecundity, and transmission ability of the vector whitefly [50]. Several volatile organic substances, produced from secondary metabolism of plant, have potential role in attracting insects which spread pathogenic organisms. However, several secondary metabolites are produced in host plant to prevent itself from herbivores. Zingiberone, curcumene and P-cymene are reported to have toxicity against whitefly. Utilization of those secondary metabolites in proper way can be a time efficient herbivory controlling approach along with pathogenic attack [51,52,53].
Some other responses of host during viral infection has been found in literatures. As for example, In TYLCV infected plants, expression of several resistance genes like Ty1, Ty6 etc. is induced and resulted lowering of viral titre in host cell. However, the resistance contributed by Ty genes can be held down when beta-satellite is present within virion genome [54,55]. Major types of host responses during begomovirus infection have been shown in (Figure 2).
5. Future Prospect and Conclusion
Plants are capable of developing defense strategies against begomoviral infection. TGS causes viral genome methylation, consequently, leads to the repression of viral pathogenicity whereas PTGS mediates the degradation of the viral mRNAs to inhibit the viral infection. Host defence regulatory mechanisms like autophagy, ubiquitination, hormonal signalling, and protein kinase activities also play a significant role in the protection of host from begomovirus infection [21]. Those can be analysed thoroughly and utilized for controlling that type of viral infection in plant. miRNAs and siRNAs can be used as effective tools for achieving broad spectrum resistance in plants against begomoviruses. However, different types of viral suppressors are evolved also according to the wide range of host defense strategies. Spreading of begomoviral diseases is highly accelerated due to this. Various approaches like conventional cross-breeding, non-coding RNA based molecular methods etc. have been adopted to control this viral infections but emergence of more than one virus infections at a time, is the reason for limiting the success. Recently, extensive efforts are giving by the researchers for standardizing the CRISPR-cas9 system for generating adaptive immunity and resistance against begomoviruses. Selection of targets within viral genome is crucial factor in achieving the desired success regarding this. Identification of the specific host factors involved in the resistance development during plant-virus interaction, is also important. Multi-directional genetic engineering using multiple targets, can aid in the development of disease resistant plants, hopefully. Role of plant defense in vector-virus interaction should be studied also. Resolving the questions behind this, may promote the development of novel strategies for virus control and manage the crop-disease problems easily.
Funding
This work is supported by SERB-DST(SRG/2020/001618), University of Calcutta for providing the basic infrastructure.
Conflicts of Interest
The authors declare there is no conflict of interest.
References
- Fondong, V.N. Geminivirus Protein Structure and Function. Mol. Plant Pathol. 2013, 14, 635–649. [Google Scholar] [CrossRef] [PubMed]
- Beam, K.; Ascencio-Ibáñez, J.T. Geminivirus Resistance: A Minireview. Front. Plant Sci. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Devendran, R.; Kumar, M.; Ghosh, D.; Yogindran, S.; Karim, M.J.; Chakraborty, S. Capsicum-Infecting Begomoviruses as Global Pathogens: Host–Virus Interplay, Pathogenesis, and Management. Trends Microbiol. 2022, 30, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Krishna, R.; Ansari, W.A.; Jaiswal, D.K. Whitefly-Transmitted Plant Viruses and Their Management. Emerg. Trends Plant Pathol. 2020, 175–195. [Google Scholar] [CrossRef]
- Leke, W.N.; Mignouna, D.B.; Brown, J.K.; Kvarnheden, A. Begomovirus Disease Complex : Emerging Threat to Vegetable Production Systems of West and Central Africa. Agric. Food Secur. 2015, 4, 1–14. [Google Scholar] [CrossRef]
- Jones, D.R. Plant Viruses Transmitted by Whiteflies. Eur. J. Plant Pathol. 2003, 109, 195–219. [Google Scholar] [CrossRef]
- Nigam, D. Genomic Variation and Diversification in Begomovirus Genome in Implication to Host and Vector Adaptation. Plants 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A. ICTV Virus Taxonomy Profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [CrossRef]
- Kumar, Y.; Hallan, V.; Zaidi, A.A. Molecular Characterization of a Distinct Bipartite Begomovirus Species Infecting Tomato in India. Virus Genes 2008, 37, 425–431. [Google Scholar] [CrossRef]
- Venkataravanappa, V.; Lakshminarayana Reddy, C.N.; Swaranalatha, P.; Jalali, S.; Briddon, R.W.; Reddy, M.K. Diversity and Phylogeography of Begomovirus-Associated Beta Satellites of Okra in India. Virol. J. 2011, 8, 555. [Google Scholar] [CrossRef]
- Gray, S.M.; Banerjee, N. Mechanisms of Arthropod Transmission of Plant and Animal Viruses. Microbiol. Mol. Biol. Rev. 1999, 63, 128–148. [Google Scholar] [CrossRef] [PubMed]
- Yadava, P.; Suyal, G.; Mukherjee, S.K. Begomovirus DNA Replication and Pathogenicity. Curr. Sci. 2010, 98, 260–268. [Google Scholar]
- Leonard, A. The Different Faces of Rolling-Circle Replication and Its Multifunctional Initiator Proteins. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Roumagnac, P.; Varsani, A. Recombination in Eukaryotic Single Stranded DNA Viruses. Viruses 2011, 3, 1699–1738. [Google Scholar] [CrossRef]
- Rodr, E.; Lozano-dur, R.; Piedra-aguilera, A.; Cruzado, L.; Bejarano, E.R.; Castillo, A.G. Geminivirus Rep Protein Interferes with the Plant DNA Methylation Machinery and Suppresses Transcriptional Gene Silencing. New Phytol. 2013, 199, 464–475. [Google Scholar]
- Mansoor, S.; Zafar, Y.; Briddon, R.W. Geminivirus Disease Complexes : The Threat Is Spreading. Trends Plant Sci. 2006, 11, 209–212. [Google Scholar] [CrossRef]
- Luan, J.; Wang, X.; Colvin, J.; Liu, S. Plant-Mediated Whitefly – Begomovirus Interactions : Research Progress and Future Prospects. Bull. Entomol. Res. 2014, 104, 267–276. [Google Scholar] [CrossRef]
- Zhang, T.; Luan, J.; Qi, J.-F.; Huang, C.-J.; Li, Meng; Zhou, Xueping; Liu, S. Begomovirus – Whitefly Mutualism Is Achieved through Repression of Plant Defences by a Virus Pathogenicity Factor. Mol. Ecol. 2012, 21, 1294–1304. [Google Scholar] [CrossRef]
- Wang, J.; Bing, X.; Li, M.; Ye, G.; Liu, S. Infection of Tobacco Plants by a Begomovirus Improves Nutritional Assimilation by a White Fl Y. Entomol. Exp. Appl. 2012, 144, 191–201. [Google Scholar] [CrossRef]
- Ueda, S.; Onuki, M.; Yamashita, M.; Yamato, Y. Pathogenicity and Insect Transmission of a Begomovirus Complex between Tomato Yellow Leaf Curl Virus and Ageratum Yellow Vein Betasatellite. Virus Genes 2012, 44, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Reddy, K.; Bhattacharyya, D.; Chakraborty, S. Plant Responses to Geminivirus Infection : Guardians of the Plant Immunity. Virol. J. 2021, 18, 1–25. [Google Scholar] [CrossRef]
- Jackel, J.N.; Storer, J.M.; Coursey, T.; Bisaro, D.M. Arabidopsis RNA Polymerases IV and V Are Required To Establish H3K9 Methylation, but Not Cytosine Methylation, on Geminivirus. J. Virol. 2016, 90, 7529–7540. [Google Scholar] [CrossRef]
- He, X.; Hsu, Y.; Zhu, S.; Wierzbicki, A.T.; Pontes, O.; Pikaard, S.; Liu, H.; Wang, C.; Jin, H.; Zhu, J. An Effector of RNA-Directed DNA Methylation in Arabidopsis Is an ARGONAUTE 4- and RNA-Binding Protein. Cell 2010, 137, 498–508. [Google Scholar] [CrossRef]
- Wang, B.; Li, F.; Huang, C.; Yang, X.; Qian, Y.; Xie, Y. V2 of Tomato Yellow Leaf Curl Virus Can Suppress Methylation-Mediated Transcriptional Gene. gene Virol. 2013, 1–18. [Google Scholar] [CrossRef]
- Wang, J.; Mei, J.; Ren, G. Plant MicroRNAs : Biogenesis, Homeostasis, and Degradation. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Chellappan, P.; Vanitharani, R.; Fauquet, C.M. MicroRNA-Binding Viral Protein Interferes with Arabidopsis Development. Proc. Natl. Acad. Sci. 2005, 102, 10381–10386. [Google Scholar] [CrossRef]
- Elvira-matelot, E. Diversity of RNA Silencing Pathways in Plants. Plant Gene Silenc. Mech. Appl. 2022, 1–32. [Google Scholar] [CrossRef]
- Bisaro, D.M. Silencing Suppression by Geminivirus Proteins. Viro 2006, 344, 158–168. [Google Scholar] [CrossRef]
- Li, F.; Huang, C.; Li, Z.; Zhou, X. Suppression of RNA Silencing by a Plant DNA Virus Satellite Requires a Host Calmodulin-Like Protein to Repress RDR6 Expression. PLoS Pathog. 2014, 10, 11–14. [Google Scholar] [CrossRef]
- Haxim, Y.; Ismayil, A.; Jia, Q.; Wang, Y.; Zheng, X.; Chen, T.; Qian, L.; Liu, N.; Wang, Y.; Han, S.; et al. Autophagy Functions as an Antiviral Mechanism against Geminiviruses in Plants. Plant Biol. 2017, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Wang, Q.; Xiao, R.; Cao, L.; Wang, Y. Mimic Phosphorylation of a  C1 Protein Encoded by TYLCCNB Impairs Its Functions as a Viral Suppressor of RNA Silencing and a Symptom Determinant. J. Virol. 2017, 91, 1–18. [Google Scholar] [CrossRef]
- Hao, L.; Wang, H.; Sunter, G.; Bisaro, D.M. Geminivirus AL2 and L2 Proteins Interact with and Inactivate SNF1 Kinase. Plant Cell 2003, 15, 1034–1048. [Google Scholar] [CrossRef]
- Patel, A.; Dey, N.; Chaudhuri, S.; Pal, A. Molecular and Biochemical Characterization of a Vigna Mungo MAP Kinase Associated with Mungbean Yellow Mosaic India Virus Infection and Deciphering Its Role in Restricting the Virus Multiplication. Plant Sci. 2017. [Google Scholar] [CrossRef]
- Li, Y.; Qin, L.; Zhao, J.; Muhammad, T.; Cao, H.; Li, H. SlMAPK3 Enhances Tolerance to Tomato Yellow Leaf Curl Virus ( TYLCV ) by Regulating Salicylic Acid and Jasmonic Acid Signaling in Tomato ( Solanum Lycopersicum ). PLoS One 2017, 1–21. [Google Scholar] [CrossRef]
- Gui, X. β C1 Protein Encoded in Geminivirus Satellite Concertedly Targets MKK2 and MPK4 to Counter Host Defense. PLoS Pathog. 2019, 1–23. [Google Scholar]
- Huang, B.; Du, D.; Zhang, R.; Wu, X.; Xing, Z.; He, Y.; Huang, W. Synthesis, Characterization and Biological Studies of Diosgenyl Analogues. Bioorganic Med. Chem. Lett. 2012, 22, 7330–7334. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Carolino, S.M.B.; Mariano, A.C.; Andrade, M.O.; Oliveira, M.L.; Baracat-pereira, M.C.; Brommonshenkel, S.H.; Fontes, E.P.B. Identification of a Novel Receptor-like Protein Kinase That Interacts with a Geminivirus Nuclear Shuttle Protein. Virology 2004, 318, 24–31. [Google Scholar] [CrossRef]
- Florentino, L.H.; Santos, A.; Fontenelle, M.R.; Pinheiro, G.L.; Zerbini, F.M.; Baracat-pereira, M.C.; Fontes, E.P.B. A PERK-Like Receptor Kinase Interacts with the Geminivirus Nuclear Shuttle Protein and Potentiates Viral Infection. J. Virol. 2006, 80, 6648–6656. [Google Scholar] [CrossRef]
- Adams, E.H.G.; Spoel, S.H. The Ubiquitin-Proteasome System as a Transcriptional Regulator of Plant Immunity. J. Exp. Bot. 2018, 0–3. [Google Scholar] [CrossRef]
- Shen, Q.; Hu, T.; Bao, M.; Cao, L.; Zhang, H.; Song, F.; Xie, Q.; Zhou, X. Tobacco RING E3 Ligase NtRFP1 Mediates Ubiquitination and Proteasomal Degradation of a Geminivirus-Encoded b C1. Mol. Plant 2016, 9, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Chatterjee, K.S.; Jha, V.; Das, R.; Shivaprasad, P. V Stability of Begomoviral Pathogenicity Determinant β C1 Is Modulated by Mutually Antagonistic SUMOylation and SIM Interactions. BMC Biol. 2020, 1–24. [Google Scholar]
- Ghosh, D. Molecular Interplay between Phytohormones and Geminiviruses : A Saga of a Never-Ending Arms Race. J. Exp. Bot. 2021, 72, 2903–2917. [Google Scholar] [CrossRef] [PubMed]
- Miozzi, L.; Napoli, C.; Sardo, L.; Accotto, G.P. Transcriptomics of the Interaction between the Monopartite Phloem-Limited Geminivirus Tomato Yellow Leaf Curl Sardinia Virus and Solanum Lycopersicum Highlights a Role for Plant Hormones, Autophagy and Plant Immune System Fine Tuning during Infection. PLoS One 2014, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Weldegergis, B.T.; Li, J.; Jung, C.; Qu, J.; Sun, Y.; Qian, H. Virulence Factors of Geminivirus Interact with MYC2 to Subvert Plant Resistance and Promote Vector Performance. Plant Cell 2014, 26, 4991–5008. [Google Scholar] [CrossRef] [PubMed]
- Oa, G.I.W.; Sozzani, R.; Lee, T.; Chu, T.; Wolfinger, R.D.; Cella, R.; Hanley-bowdoin, L. Global Analysis of Arabidopsis Gene Expression Uncovers a Complex Array of Changes Impacting Pathogen Response and Cell Cycle During. Plant Physiol. 2008, 148, 436–454. [Google Scholar] [CrossRef]
- Duan, K.; Liu, B.; Li, C.; Zhang, H.; Yu, T.; Qu, J.; Zhou, M.; Chen, L.; Meng, S.; Hu, Y.; et al. Effectiveness of Convalescent Plasma Therapy in Severe COVID-19 Patients. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 9490–9496. [Google Scholar] [CrossRef] [PubMed]
- Naseem, M.; Kaltdorf, M.; Dandekar, T. The Nexus between Growth and Defence Signalling : Auxin and Cytokinin Modulate Plant Immune Response Pathways. J. Exp. Bot. 2015, 1–12. [Google Scholar] [CrossRef]
- Vinutha, T.; Vanchinathan, S.; Bansal, N.; Kumar, G.; Permar, V.; Watts, A.; Ramesh, S. V; Praveen, S. Tomato Auxin Biosynthesis / Signaling Is Reprogrammed by the Geminivirus to Enhance Its Pathogenicity. Planta 2020, 1–14. [Google Scholar] [CrossRef]
- Mahmoudabadi, G.; Milo, R.; Phillips, R. Energetic Cost of Building a Virus. Proc. Natl. Acad. Sci. 2017, E4324–E4333. [Google Scholar] [CrossRef]
- Ghosh, S. Factors Determining Transmission of Persistent Viruses by Bemisia Tabaci and Emergence of New Virus – Vector Relationships. Viruses 2021, 13, 1–13. [Google Scholar] [CrossRef]
- Luan, J.; Yao, D. Suppression of Terpenoid Synthesis in Plants by a Virus Promotes Its Mutualism with Vectors. Ecol. Lett. 2013, 16, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.E.I.; Auslane, H.J.M.C.; Schuster, D.J. Repellency of Ginger Oil to Bemisia Argentifolii ( Homoptera : Aleyrodidae ) on Tomato. Ecotoxicology 2004, 1310–1318. [Google Scholar]
- Oa, T.I.W.; Weidner, M.; Schu, S.; Bleeker, P.M.; Diergaarde, P.J.; Ament, K.; Both, M.T.J. De; Haring, M.A.; Schuurink, R.C.; Wageningen, A.E. The Role of Specific Tomato Volatiles In. Plant Physiol. 2009, 151, 925–935. [Google Scholar] [CrossRef]
- Verlaan, M.G.; Hutton, S.F.; Ibrahem, R.M.; Kormelink, R.; Visser, R.G.F.; Scott, J.W.; Edwards, J.D.; Bai, Y. The Tomato Yellow Leaf Curl Virus Resistance Genes Ty-1 and Ty-3 Are Allelic and Code for DFDGD-Class RNA – Dependent RNA Polymerases. PLoS Genet. 2013, 9, 1–11. [Google Scholar] [CrossRef]
- Yan, Z.H.E.; Elink, R.K. Ty-1, a Universal Resistance Gene against Geminiviruses That Is Compromised by Co-Replication of a Betasatellite. Mol. Plant Pathol. 2018, 1, 160–172. [Google Scholar] [CrossRef]
Figure 1.
Diagrammatic representation of begomovirus infection mechanism.
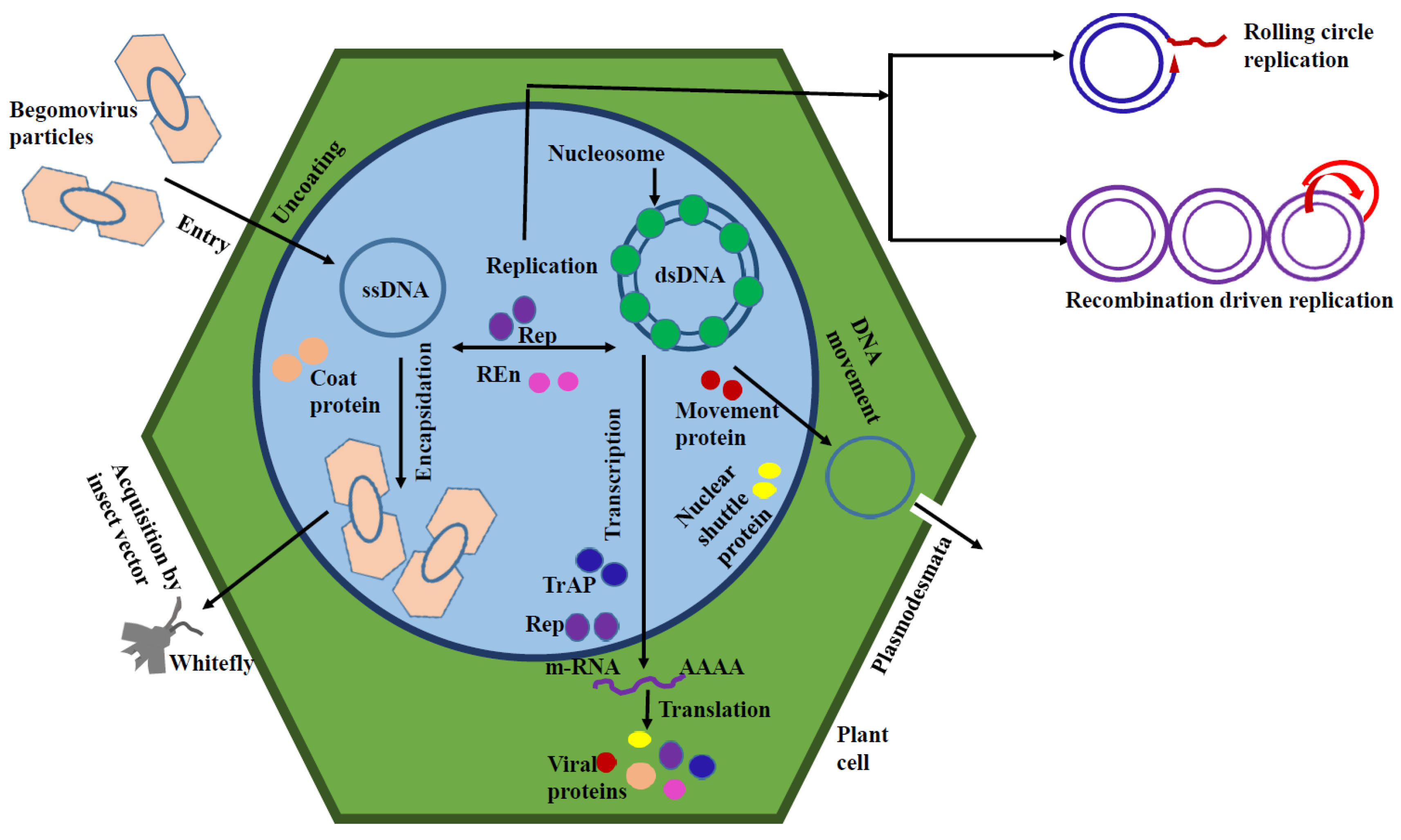
Figure 2.
Schematic representation of vector borne begomovirus infection and responses in host cell.
Figure 2.
Schematic representation of vector borne begomovirus infection and responses in host cell.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



