
Submitted:
05 May 2023
Posted:
09 May 2023
You are already at the latest version
Abstract
Background: Carbapenem-resistant Klebsiella pneumoniae (CRKP) is a serious public health issue. Aim of the study was to identify the antimicrobial resistance and accessory genes, the clonal relatedness and the evolution dynamics of selected CRKP isolates recovered in an adult and a pediatric intensive care unit of a tertiary hospital in Greece. Methods: Twenty-four CRKP isolates recovered during 2018-2022 were included in the study. Next-generation sequencing was performed using the Ion Torrent PGM Platform. The identification of the plasmid content, MLST and antimicrobial resistance genes, as well as the comparison of multiple genome alignments and the identification of core genome single-nucleotide polymorphism sites, were performed applying various bioinformatics software. Results: The isolates belonged to eight sequence types 11, 15, 30, 35, 39, 307, 323, and 512. A variety of carbapenemases (KPC, VIM, NDM and OXA-48) and resistance genes were detected. CRKP strains shared visually common genomic regions with the reference strain (NTUH-K2044). ST15, ST323, ST39, and ST11 CRKP isolates presented on average 17, 6, 16 and 866 recombined SNPs, respectively. All isolates belonging to ST15, ST323 and ST39 were classified in distinct phylogenetic branches, while ST11 isolates were assigned to a two-subclade branch. For large CRKP sets, the phylogeny seems to change approximately every seven SNPs. Conclusion: The current study provides insight into the genetic characterization of CRKP isolates in the ICUs of a tertiary hospital. Our results indicate clonal dispersion of ST15, ST323, and ST39 and highly diverged ST11 isolates.
Keywords:
carbapenem-resistant Klebsiella pneumoniae
; whole genome sequencing
; core genome single-nucleotide polymorphism analysis
; molecular epidemiology
1. Introduction
Klebsiella pneumoniae is recognized as an important Gram-negative opportunistic pathogen that causes community- and hospital-acquired infections globally [1]. It can cause ventilator-associated pneumonia among patients in intensive care units (ICUs), bloodstream infections and urinary tract infections [2,3,4]. The emergence of multidrug-resistant (MDR) K. pneumoniae has become a major public health problem causing a significant increase of morbidity and mortality worldwide [5]. The spread of carbapenem-resistant K. pneumoniae (CRKP) has generally been considered an increasingly serious issue for clinical practice due to the limitation of therapeutic options [6,7,8]. Greece is endemic for CRKP [9,10,11,12,13]; data from the European Antimicrobial Resistance Surveillance Network committee for the year 2020 indicate that the prevalence of CRKP in Greece was 66.3%, the highest among the European countries [14].
The rapid dissemination of CRKP is closely related to the antimicrobial resistance genes carried by plasmids and transferable genetic elements [15,16,17,18]. CRKP strains accumulate antimicrobial resistance genes due to inadequate implementation of infection control measures in healthcare settings and the irrational use of antimicrobials resulting in the emergence of MDR, extremely drug-resistant and pandrug-resistant phenotypes [19]. The pathogenic potential of these strains is determined by virulence factors, such as capsular polysaccharide synthesis, fimbriae, pili, outer membrane protein and iron acquisition systems [20].
The application of next-generation sequencing (NGS) has become a powerful tool for obtaining whole genome sequences (WGS), resulting in genetic characterization of the strains and a better understanding of the genomic diversity providing fast and in-depth information about the bacterial pathogenicity [21]. WGS analysis is recommended for genotyping and determining relatedness between clinical K. pneumoniae strains, as well as for the surveillance of virulence genes [22]. The aim of the present study was to identify the antimicrobial resistance and accessory genes of selected CRKP isolates recovered in an adult intensive care unit (ICU) and a pediatric ICU (PICU) of a tertiary hospital in Greece, to evaluate their clonal relatedness through core genome single-nucleotide polymorphism (cgSNP) analysis and to calculate the evolution dynamics of this bacterial population.
2. Materials and Methods
2.1. Clinical isolates - Setting
The present study included 24 CRKP isolates collected during June 2018 to July 2022 from 18 patients hospitalized in the ICU (8 female and 10 male) and six patients hospitalized in the PICU (4 female and 2 male) of a tertiary university-affiliated general hospital in Greece. The median age of the adult patients was 57.5 years (range 30 - 88), while the median age of the children was 6.2 years (range 0.25 - 17). Τhe selection of the isolates was based on the type of carbapenemase(s) previously detected in order to include all possible combinations, as they were pre-screened as described [11]. Eight isolates were collected from colonization sites (rectal swabs), while 16 isolates were taken from various types of clinical samples (Table 1).
2.2. Ethics approval
The study was approved by the Ethics Committee of Aristotle’s University Medical Faculty (no. of approval 5.160/18-12-19).
2.3. Microbiological methods - Antimicrobial susceptibility testing
CRKP isolates were identified through the automated system VITEK 2 (bioMérieux, Marcy-l’Étoile, France) using the Gram-Negative Identification card (GN ID). Antimicrobial susceptibility testing (AST) of isolates was performed using the AST 318 and XN10 cards, while the use of minimum inhibitory concentration (MIC) test strips (Liofilchem srl, Roseto, Italy) was applied to confirm susceptibility to ceftazidime-avibactam (CAZ-AVI). Identification and susceptibility cards were interpreted according to the manufacturer's instructions. The Clinical and Laboratory Standards Institute (CLSI) susceptibility breakpoints were applied for the interpretation of results [23]. The breakpoints approved by the US Food and Drug Administration (FDA) were used to interpret the MICs of tigecycline [24].
All CRKP isolates were previously screened for carbapenemase production using a lateral flow immunochromatographic assay (LFIA) (NG-Test CARBA 5, NG Biotech, France). The results were confirmed by a multiplex PCR in a single reaction following a modified protocol that was previously described [11,25].
2.4. DNA extraction and whole genome sequencing
Bacterial DNA was extracted using the DNA extraction kit (Qiagen, Hilden, Germany). The DNA concentration (sample starting concentration between 10–100 ng/μl) was measured using the Qubit double-strand DNA HS assay kit (Q32851, Life Technologies Corporation, Grand Island, NY, USA).
NGS was performed using the Ion Torrent PGM Platform (Life Technologies Corporation). All procedures regarding purification, ligation, barcoding, library preparation process, emulsion PCR and enrichment were performed following the manufacturer’s instructions. PCR products were loaded on an Ion-316™ chip. The Ion PGM Hi-Q View (200) chemistry was applied using the Ion PGM Hi-Q View Sequencing kit (A25592).
2.5. Assembly assessment and genome annotation
2.6. MLST and detection of antimicrobial resistance genes and plasmids
Τhe sequence types (ST), plasmid types and antimicrobial resistance genes were identified through the Center for Genomic Epidemiology website using the MLST web server and the related online databases, the PlasmidFinder version 2.0, and the Resfinder version 4.1, respectively [29,30,31]. The Comprehensive Antibiotic Resistance Database was also used with the selection criteria hits set to perfect (100%) and strict (>95%) identity to the reference sequence [32]. The detection of genes related to virulence capsule, efflux and regulator systems was performed according to the protocols available from the Pasteur Institute (https://bigsdb.pasteur.fr/cgi-bin/bigsdb/bigsdb.pl?db=pubmlst_klebsiella_seqdef).
2.7. Genomic comparison – Core genome single-nucleotide polymorphism (cgSNP)-based phylogenetic analysis
The BRIG software was used to compare visually multiple genomes alignment using a default minimum threshold of 50% [27]. The Reference sequence Alignment based Phylogeny builder version 1.12 (REALPHY; https://realphy.unibas.ch/realphy/) was used to identify relevant SNP sites for core genome phylogenetic analysis [33]. Default input parameters were applied for cgSNP identification. The K. pneumoniae NTUH-K2044 sequence was used as a reference for the multiple alignments and the visual comparison of the whole genomes [28]. The produced alignment was used for the creation of an unrooted tree on the entire core genome through the approximate maximum likelihood (ML) method using PhyML [34]. Pairwise analysis between isolates was also performed using REALPHY, as previously described by Dixit et al. [35]. The calculation of SNPs between isolates was conducted through REALPHY, using a Poisson mixture model plus a negative binomial for the recombined regions. REALPHY was also used to assess to which extent mutually consistent SNPs are clustered along the alignment; the lengths of segments along the alignment that are consistent with a single phylogeny were calculated [33]. In addition, the lower bound for the ratio between the total number of phylogeny changes (C) and substitution events (S) (C/S) that occur along the core genome alignment was calculated. The same ratio was calculated for any subset of CRKP isolates by removing 5% of potentially homoplastic sites [36]. Moreover, a rough estimate for the average number of times T that a randomly chosen position in the core genome alignment has been overwritten by recombination in its history was calculated, that is the time since the genetic ancestors of the position in the alignment diverged from a common ancestor.
3. Results
3.1. Antimicrobial susceptibility testing
All CRKP isolates were resistant to β-lactam antibiotics (including penicillins, cephalosporins, monobactams and carbapenems). Nineteen isolates (79.2%) were resistant to at least one aminoglycoside, 23 isolates (95.8%) showed resistance to quinolones, eight isolates (33.3%) were resistant to fosfomycin and tigecycline, and 21 (87.5%) to cotrimoxazole. Four out of five isolates that carried the class A β-lactamase blaKPC showed susceptibility to ceftazidime-avibactam (CAZ-AVI). The phenotypic resistance rates to antimicrobials of CRKP isolates are shown in the heatmap of Figure 1.
3.2. MLST, antimicrobial resistance genes, plasmids
The CRKP isolates belonged to eight different STs: ST11 (7, 29.1%), ST39 (7, 29.1%), ST323 (3, 12.5%), ST15 (7, 29.1%), ST307 (1, 4.2%), ST30 (1, 4.2%), ST512 (1, 4.2%) and ST35 (1, 4.2%).
The resistance genes and the plasmids content per isolate are detailed in Table 2. The prevalence of genes conferring resistance to beta-lactams was 100%, to aminoglycosides 79.2%, to quinolones 95.8%, to fosfomycin and tigecycline 33.3%, and to cotrimoxazole 87.5%. The most prevalent plasmid was IncF which is associated with the spread of several extended-spectrum β-lactamase (ESBL) genes or carbapenemases and was detected in all CRKP strains; additional plasmids were found in fifteen strains, such as Col and IncC.
3.3. Virulence factors and Efflux and Regulator Systems
The virulence factors detected included fimbrial genes, as 23/24 isolates possessed the mrk cluster (type 3 fimbriae); siderophores, as yersiniabactin genes (ybt, irp and fyuA) were present in 17 isolates, while genes encoding aerobactin siderophore (iut) were identified in five isolates (three of them carried also yersiniabactin genes). In addition, the kfu gene (Klebsiella ferric iron uptake which is a regulator of iron transport system) was detected in four isolates.
Efflux and regulator systems genes encoding the AcrAB efflux pump (acr, mar, sox, rob, ram, sdiA, fis and envR) or the OqxAB efflux pump (oqx and rar) were present in all isolates (Table 2).
3.4. Genomic comparison among CRKP strains
Circular genome maps were generated for the four STs with more than one isolate (ST11, ST39, ST323 and ST15). It was found that isolates belonging to the same ST shared many identical regions with the reference strain. CRKP isolates C4112 and C746 of the ST11, D2452 and D6184 of the ST39, Z866 of the ST323, and B2562 of the ST15 displayed visually the highest similarity with the reference strain throughout their whole genome (Figure 2).
3.5. CgSNP-based phylogenetic analysis
The length of the multiple core genome alignment of all CRKP isolates was 1,892,845 base pairs. The core genome length corresponds to about 36.1% of the reference genome length. Nearly 1.9 million columns in the core genome alignment (almost 98.5%) showed identical nucleotides for all isolates, meaning that 1.5% of the genome is polymorphic.
A phylogenetic tree including the cgSNP-based phylogenetic analysis of all isolates, along with the reference strain, is seen in Figure 3.
When performing all possible pairwise analyses among ST15 isolates, an average of 17 recombined SNPs (range 5-26) were detected; the SNP diversity along the core genome was very low, that is 0-3 cgSNPs per kilobase. ST323 CRKP strains presented an average of six recombined SNPs (range 5-6), and the SNP diversity was extremely low, that is 0-2 cgSNPs per kilobase. ST39 CRKP strains displayed an average of 16 recombined SNPs (range 4-43) with the SNP diversity being low, that is 0-5 cgSNPs per kilobase. Finally, ST11 CRKP strains showed an average of 866 recombined SNPs (range 1-1652), with the SNP diversity being clearly higher, that is 0-66 cgSNPs per kilobase.
For pairs of CRKP isolates, for instance C4112 and C746 with relatively low diversity (7.5 x 10-4) (Figure 4G), the impact of recombination is almost directly clear in the pattern of SNP density along the genome. While the SNP density is very low along most of the genome, that is 0-2 SNPs per kilobase, there are a few segments where the SNP density is much higher (Figure 4A) and comparable to the typical SNP density between randomly selected highly diverged CRKP isolates, that is 30-65 SNPs per kilobase (Figure 4C). For some pairs of isolates, for example, A18940 and D2856, with higher genetic diversity (1.5 x 10-3) (Figure 4G), the frequency of these high-spiking recombined regions increases (Figure 4B), until most of the genome is covered by such regions when comparing highly divergent isolates (5.5 x 10-3), for instance A5051 and C2482 (Figure 4G, Figure 4C). The distribution of SNP densities was described through a majority of clonally inherited regions with mostly up to three SNPs per kilobase, and a long tail of recombined regions with up to 50 or 65 SNPs per kilobase (Figure 4D-F).
Figure 5 highlights the distribution of the lengths of tree-compatible stretches. This distribution had a mode at n=2, stretches were normally around 7-12 consecutive SNPs, and very rarely longer than 15-20 consecutive SNPs (Figure 5A). Similarly, tree-compatible segments were typically just a few hundreds of base pairs long (around 1000-1500 base pairs), and very rarely more than 2000 base pairs (Figure 5B).
Figure 6 underscores the ratio C/S for random subsets of all 24 isolates. When comprising all 24 isolates, a ratio C/S=0.15 was obtained for the 5% homoplasy-corrected full alignment. For small subsets of isolates, the C/S ratio displayed significant fluctuations. For instance, for subsets of n=7 isolates, the C/S ratio presented a mean of 0.094 ± 0.046. However, as the number of isolates in the subset increased, the ratios converged to the value C/S=0.15, and for large subsets of isolates there was little variation in this ratio. Consequently, for alignments of large sets of isolates, the phylogeny should change at least approximately every seven SNPs (Figure 6). In addition, on average, each randomly chosen position on the core genome has been overwritten at least T=4344 times (5% homoplasies removed).
4. Discussion
In the current study, the WGSs of 24 CRKP isolates were analysed. It was found that they belonged to eight different STs, with ST11 and ST39 being the most prevalent. Ten out of 24 isolates (41.6%) carried blaNDM-1, and two of them (8.3%) transferred concurrently blaNDM-1 and blaOXA-48. Six out of 10 NDM-producers (60.0%) were assigned to ST11 which is common in NDM-positive CRKP strains, and it is globally distributed, especially in Asia, and particularly in China, but also in Europe [37,38]. In Greece, the first outbreak by NDM-1-producing K. pneumoniae ST11 clonal strain occurred during 2011-2012 [38], while the first report of K. pneumoniae ST11 clinical isolate co-producing NDM-1 and OXA-48 carbapenemases was reported in 2016 [39]. However, three out of 10 NDM-producers (30.0%) were assigned to ST15; this has also been previously described [40].
ST39 was also the most prevalent type encompassing seven isolates (29.1%); two of them carried blaKPC-2, while one of them (14.3%) harboured blaKPC-33 (a KPC-2 variant). CRKP strains belonging to ST39 have been detected only in sporadic cases so far, like ST39 carrying blaKPC-2 [41], or blaVIM-1 [42]. ST39 isolates transferring KPC-33, have been recently detected in Greece [8]. The present study also included the recently reported four ST39 isolates carrying both blaKPC-2 and blaVIM-1 [43]. Three CRKP isolates (12.5%) were assigned to ST323; sporadic cases of ST323 CRKP have been previously reported in Greece and other European countries [44,45]. It is of interest that one isolate, A1746/22, carried the VEB-25 gene (article submitted for publication); VEB-25 is a variant of VEB-1 and confers resistance to CAZ-AVI [46,47].
Twenty of the 24 isolates harboured at least two genes encoding aminoglycoside-modifying enzymes (AMEs), and six of them carried one 16S rRNA methylase gene (armA, rmtB). In addition, 12 of these isolates carried at least one gene of the three AME classes (acetyltransferases-aac, phosphotransferase-aph, nucleotransferases-ant). A disagreement between phenotype and genotype has been detected in five isolates (20.8%) (detection of AME-encoding resistance genes but phenotypically sensitive to both aminoglycosides tested). A disagreement of approximately 10% between genotype and phenotype for aminoglycosides for VITEK 2 automated system has been reported previously [48].
Fluoroquinolone resistance was observed in seven isolates (29.2%), which carried the plasmid-encoded qnr gene, as previously described for CRKP isolates [49]. The multidrug efflux pump oqxAB gene was detected in all strains [50], while the aac(6’)-Ib-cr gene (which encodes an aminoglycoside acetyltransferase that modifies not only aminoglycosides but also fluoroquinolones) has been identified in 16 isolates, as previously described [49].
For fosfomycin resistance, the gene fosA was detected in all isolates, but 16 of them (64.0%) were phenotypically susceptible, evidenced by an increase in the MIC, as previously described [51]. Resistance genes for cotrimoxazole (sul or dfrA) were found in all isolates except one. The Inc-type plasmids detected in all isolates and the Col-type plasmids detected in 15 isolates have been reported in other studies [43,52].
In terms of adhesive structures, almost all isolates possessed the mrk cluster, characteristic of type 3 fimbriae, which promotes bacterial adhesion to both abiotic and biotic surfaces [53]. Regarding siderophores, the majority of isolates carried yersiniabactin genes that are significantly associated with pathogenesis and invasive infections in humans [54], while aerobactin (detected in five isolates) is more specific for hypervirulent K. pneumoniae [55]. The acquisition of a chimeric plasmid including genes for the siderophores yersiniabactin and aerobactin, grants cefiderocol activity against CRKP strains [56].
NGS allows for more accurate analyses regarding the clonal relatedness of strains compared to conventional typing methods [57]. Several studies have described the molecular epidemiology of CRKP isolates through NGS using a gene-by-gene comparison or SNP analysis [58,59].
In pairwise comparisons, isolates of the ST15, ST323 and ST39 presented an average of 17, 6 and 16 recombined SNPs, respectively, implying the clonal relatedness of these isolates in accordance with their ST classification. Similarly, Miro et al. report the clonal relatedness of OXA-48-producing ST405 CRKP strains further studied with core genome MLST, showing between 6 and 17 cgSNPs [57]. Moreover, Onori et al. reveal the clonal relatedness of sporadic KPC-producing ST258 CRKP strains presenting an average of 20 or 27 cgSNPs [60]. ST15 and ST39 strains display a more increased number of cgSNPs compared to ST323 strains, and this is highlighted as a slightly more increased diversity in the corresponding clades of the phylogenetic tree (Figure 3). However, ST15 and ST39 have been isolated over a 33- and 36-month period, respectively, while ST323 have been collected over a 2.5-month period (Table 1). This could indicate a difference in the measured paces of the molecular clock between these clades, as previously proposed [60].
On the contrary, ST11 isolates showed an average of 866 cgSNPs, indicating the polyclonality of these isolates. This indication is further strengthened by the variability in their content of carbapenemase and antimicrobial resistance genes (Table 2) and by their phylogenetic division into two distinct subclades (Figure 3). The high average of cgSNPs occurred between ST11 strains belonging to these two different subclades resulting in SNP patterns (Figure 4A). Most probably, these high SNP density patterns result from horizontal recombination events, e.g. transfer of plasmid or phage, as previously proposed for other bacterial species [36]. These results underscore the higher discriminatory power of cgSNP analysis compared to MLST [57].
In addition, the current study highlighted novel ways in which these cgSNPs could be used to quantify phylogenetic structures and the role of recombination in genome evolution. It was shown that there are pairs of closely related isolates, and that most of their DNA have been clonally inherited, however most of the changes derived from recombination events. When focusing on the core genome alignment, it has been shown that until the current genetic state of CRKP strains each genomic locus has been overwritten thousands of times by recombination in the history of their evolution. Sakoparnig et al. reached the same conclusions when focusing on E. coli isolates and they disclose a similar C/S ratio [36]. Moreover, an attempt was conducted to calculate the probability for a pair of SNPs to be consistent with a common phylogeny, as a function of their genome distance [61]. Several other studies have also attempted to quantify recombination in bacteria and assess the relative rate with which different lineages have recombined, focusing though on different types of bacteria [36,61]. To the best of our knowledge, our study is the first to accomplish such a task studying CRKP strains.
One of the basic challenges among WGS-based typing methods is the definition of a cut-off value for the identification of the clonal relatedness between isolates; however this remains doubtful, as it seems to be not only species-dependent but also population-dependent [57,60,62]. Thus, a threshold range should be evaluated along with clinical epidemiological data.
In conclusion, the current study provides insight into the genetic characterization of CRKP isolates circulating during a 4-year period in the ICUs of a Greek tertiary hospital. Phylogenetic analysis showed that some of them were epidemiologically related. Given that NGS has become more affordable, its use in hospital settings is extremely helpful as the obtained data could lead the infection control and prevention strategies.
Author Contributions
Charalampos Zarras: Investigation, Review and editing. Theodoros Karampatakis: Methodology, Conceptualization, Formal analysis, Writing the initial draft. Styliani Pappa: Investigation. Elias Iosifidis: Provision of study materials, Review and editing. Eleni Vagdatli: Investigation. Emmanuel Roilides: Provision of study materials, Review and editing. Anna Papa: Conceptualization, Methodology, Data curation, Formal analysis, Visualization, Reviewing and editing, Supervision, Funding acquisition.
Funding
The current study was supported by the European Union's Horizon 2020 projects COMPARE (grant number 643476) and VEO (grant number 874735).
Institutional Review Board Statement
The study was approved by the Ethics Committee of Aristotle’s University Medical Faculty (no. of approval 5.160/18-12-19).
Data Availability Statement
The data of this study have been deposited in the European Nucleotide Archive (ENA) at EMBL-EBI under accession numbers PRJEB42192 and PRJEB46676.
Acknowledgments
We thank Prof. Surbhi Malhotra-Kumar and Ms. Minh Nguyen-Ngoc from the University of Antwerp in Belgium for the technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Juan CH, Fang SY, Chou CH, Tsai TY, Lin YT. Clinical characteristics of patients with pneumonia caused by Klebsiella pneumoniae in Taiwan and prevalence of antimicrobial-resistant and hypervirulent strains: a retrospective study. Antimicrob Resist Infect Control 2020; 9: 4.
- Martin RM, Bachman MA. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front Cell Infect Microbiol 2018; 8: 4.
- Shao C, Wang W, Liu S, Zhang Z, Jiang M, Zhang F. Molecular Epidemiology and Drug Resistant Mechanism of Carbapenem-Resistant Klebsiella pneumoniae in Elderly Patients With Lower Respiratory Tract Infection. Front Public Health 2021; 9: 669173.
- Gupta A, Bhatti S, Leytin A, Epelbaum O. Novel complication of an emerging disease: Invasive Klebsiella pneumoniae liver abscess syndrome as a cause of acute respiratory distress syndrome. Clin Pract 2018; 8: 1021.
- Ferri M, Ranucci E, Romagnoli P, Giaccone V. Antimicrobial resistance: A global emerging threat to public health systems. Crit Rev Food Sci Nutr 2017; 57: 2857-2876.
- Nelson K, Hemarajata P, Sun D, Rubio-Aparicio D, Tsivkovski R, Yang S et al. Resistance to Ceftazidime-Avibactam Is Due to Transposition of KPC in a Porin-Deficient Strain of Klebsiella pneumoniae with Increased Efflux Activity. Antimicrob Agents Chemother 2017; 61.
- Wang B, Pan F, Wang C, Zhao W, Sun Y, Zhang T et al. Molecular epidemiology of Carbapenem-resistant Klebsiella pneumoniae in a paediatric hospital in China. Int J Infect Dis 2020; 93: 311-319.
- Galani I, Karaiskos I, Angelidis E, Papoutsaki V, Galani L, Souli M et al. Emergence of ceftazidime-avibactam resistance through distinct genomic adaptations in KPC-2-producing Klebsiella pneumoniae of sequence type 39 during treatment. Eur J Clin Microbiol Infect Dis 2021; 40: 219-224.
- Galani I, Nafplioti K, Adamou P, Karaiskos I, Giamarellou H, Souli M et al. Nationwide epidemiology of carbapenem resistant Klebsiella pneumoniae isolates from Greek hospitals, with regards to plazomicin and aminoglycoside resistance. BMC Infect Dis 2019; 19: 167.
- Karampatakis T, Tsergouli K, Politi L, Diamantopoulou G, Iosifidis E, Antachopoulos C et al. Molecular Epidemiology of Endemic Carbapenem-Resistant Gram-Negative Bacteria in an Intensive Care Unit. Microb Drug Resist 2019; 25: 712-716.
- Zarras C, Pappa S, Zarras K, Karampatakis T, Vagdatli E, Mouloudi E et al. Changes in molecular epidemiology of carbapenem-resistant Klebsiella pneumoniae in the intensive care units of a Greek hospital, 2018-2021. Acta Microbiol Immunol Hung 2022.
- Karampatakis T, Antachopoulos C, Iosifidis E, Tsakris A, Roilides E. Molecular epidemiology of carbapenem-resistant Klebsiella pneumoniae in Greece. Future Microbiol 2016; 11: 809-823.
- Protonotariou E, Meletis G, Pilalas D, Mantzana P, Tychala A, Kotzamanidis C et al. Polyclonal Endemicity of Carbapenemase-Producing Klebsiella pneumoniae in ICUs of a Greek Tertiary Care Hospital. Antibiotics (Basel) 2022; 11.
- Surveillance atlas of infectious diseases.Available from: https://atlas.Ecdc.Europa.Eu/public/index.Aspx. 2020.
- Hatrongjit R, Kerdsin A, Akeda Y, Hamada S. Detection of plasmid-mediated colistin-resistant and carbapenem-resistant genes by multiplex PCR. MethodsX 2018; 5: 532-536.
- Mari-Almirall M, Ferrando N, Fernandez MJ, Cosgaya C, Vines J, Rubio E et al. Clonal Spread and Intra- and Inter-Species Plasmid Dissemination Associated With Klebsiella pneumoniae Carbapenemase-Producing Enterobacterales During a Hospital Outbreak in Barcelona, Spain. Front Microbiol 2021; 12: 781127.
- Yanat B, Rodriguez-Martinez JM, Touati A. Plasmid-mediated quinolone resistance in Enterobacteriaceae: a systematic review with a focus on Mediterranean countries. Eur J Clin Microbiol Infect Dis 2017; 36: 421-435.
- Bosch T, Lutgens SPM, Hermans MHA, Wever PC, Schneeberger PM, Renders NHM et al. Outbreak of NDM-1-Producing Klebsiella pneumoniae in a Dutch Hospital, with Interspecies Transfer of the Resistance Plasmid and Unexpected Occurrence in Unrelated Health Care Centers. J Clin Microbiol 2017; 55: 2380-2390.
- Silva DDC, Rampelotto RF, Lorenzoni VV, Santos SOD, Damer J, Horner M et al. Phenotypic methods for screening carbapenem-resistant Enterobacteriaceae and assessment of their antimicrobial susceptibility profile. Rev Soc Bras Med Trop 2017; 50: 173-178.
- Paczosa MK, Mecsas J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol Mol Biol Rev 2016; 80: 629-661.
- Brhelova E, Antonova M, Pardy F, Kocmanova I, Mayer J, Racil Z et al. Investigation of next-generation sequencing data of Klebsiella pneumoniae using web-based tools. J Med Microbiol 2017; 66: 1673-1683.
- Enany S, Zakeer S, Diab AA, Bakry U, Sayed AA. Whole genome sequencing of Klebsiella pneumoniae clinical isolates sequence type 627 isolated from Egyptian patients. PLoS One 2022; 17: e0265884.
- Performance Standards for Antimicrobial Susceptibility Testing, 32nd Edition,CLSI standard M02. Clinical and Laboratory Standards Institute, Wayne, PA. 2020.
- Administration. USFaD. FDA approves new antibacterial drug Avycaz. FDA news release; U.S. FDA, Silver Spring26 February 2015.
- Tsakris A, Pournaras S, Woodford N, Palepou MF, Babini GS, Douboyas J et al. Outbreak of infections caused by Pseudomonas aeruginosa producing VIM-1 carbapenemase in Greece. J Clin Microbiol 2000; 38: 1290-1292.
- Geneious Prime 2021.2.1 https://www.geneious.com.
- Alikhan NF, Petty NK, Ben Zakour NL, Beatson SA. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 2011; 12: 402.
- Wu KM, Li LH, Yan JJ, Tsao N, Liao TL, Tsai HC et al. Genome sequencing and comparative analysis of Klebsiella pneumoniae NTUH-K2044, a strain causing liver abscess and meningitis. J Bacteriol 2009; 191: 4492-4501.
- Larsen MV, Cosentino S, Rasmussen S, Friis C, Hasman H, Marvig RL et al. Multilocus sequence typing of total-genome-sequenced bacteria. J Clin Microbiol 2012; 50: 1355-1361.
- Carattoli A, Zankari E, Garcia-Fernandez A, Voldby Larsen M, Lund O, Villa L et al. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother 2014; 58: 3895-3903.
- Zankari E, Hasman H, Cosentino S, Vestergaard M, Rasmussen S, Lund O et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother 2012; 67: 2640-2644.
- Jia B, Raphenya AR, Alcock B, Waglechner N, Guo P, Tsang KK et al. CARD 2017: expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res 2017; 45: D566-D573.
- Bertels F, Silander OK, Pachkov M, Rainey PB, van Nimwegen E. Automated reconstruction of whole-genome phylogenies from short-sequence reads. Mol Biol Evol 2014; 31: 1077-1088.
- Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 2010; 59: 307-321.
- Dixit PD, Pang TY, Studier FW, Maslov S. Recombinant transfer in the basic genome of Escherichia coli. Proc Natl Acad Sci U S A 2015; 112: 9070-9075.
- Sakoparnig T, Field C, van Nimwegen E. Whole genome phylogenies reflect the distributions of recombination rates for many bacterial species. Elife 2021; 10.
- Zhang R, Liu L, Zhou H, Chan EW, Li J, Fang Y et al. Nationwide Surveillance of Clinical Carbapenem-resistant Enterobacteriaceae (CRE) Strains in China. EBioMedicine 2017; 19: 98-106.
- Voulgari E, Gartzonika C, Vrioni G, Politi L, Priavali E, Levidiotou-Stefanou S et al. The Balkan region: NDM-1-producing Klebsiella pneumoniae ST11 clonal strain causing outbreaks in Greece. J Antimicrob Chemother 2014; 69: 2091-2097.
- Protonotariou E, Meletis G, Chatzopoulou F, Malousi A, Chatzidimitriou D, Skoura L. Emergence of Klebsiella pneumoniae ST11 co-producing NDM-1 and OXA-48 carbapenemases in Greece. J Glob Antimicrob Resist 2019; 19: 81-82.
- Berglund B, Hoang NTB, Lundberg L, Le NK, Tarnberg M, Nilsson M et al. Clonal spread of carbapenem-resistant Klebsiella pneumoniae among patients at admission and discharge at a Vietnamese neonatal intensive care unit. Antimicrob Resist Infect Control 2021; 10: 162.
- Liu H, Wilksch J, Li B, Du J, Cao J, Zhang X et al. Emergence of ST39 and ST656 extensively drug-resistant Klebsiella pneumoniae isolates in Wenzhou, China. Indian J Med Microbiol 2017; 35: 145-146.
- Cabanel N, Rosinski-Chupin I, Chiarelli A, Botin T, Tato M, Canton R et al. Evolution of VIM-1-Producing Klebsiella pneumoniae Isolates from a Hospital Outbreak Reveals the Genetic Bases of the Loss of the Urease-Positive Identification Character. mSystems 2021; 6: e0024421.
- Karampatakis T, Zarras C, Pappa S, Vagdatli E, Iosifidis E, Roilides E et al. Emergence of ST39 carbapenem-resistant Klebsiella pneumoniae producing VIM-1 and KPC-2. Microb Pathog 2022; 162: 105373.
- Giakkoupi P, Papagiannitsis CC, Miriagou V, Pappa O, Polemis M, Tryfinopoulou K et al. An update of the evolving epidemic of blaKPC-2-carrying Klebsiella pneumoniae in Greece (2009-10). J Antimicrob Chemother 2011; 66: 1510-1513.
- Piekarska K, Zacharczuk K, Wolkowicz T, Wolaniuk N, Rzeczkowska M, Gierczynski R. Emergence of Enterobacteriaceae co-producing CTX-M-15, ArmA and PMQR in Poland. Adv Clin Exp Med 2019; 28: 249-254.
- Galani I, Karaiskos I, Souli M, Papoutsaki V, Galani L, Gkoufa A et al. Outbreak of KPC-2-producing Klebsiella pneumoniae endowed with ceftazidime-avibactam resistance mediated through a VEB-1-mutant (VEB-25), Greece, September to October 2019. Euro surveillance : bulletin Europeen sur les maladies transmissibles = European communicable disease bulletin 2020; 25.
- Voulgari E, Kotsakis SD, Giannopoulou P, Perivolioti E, Tzouvelekis LS, Miriagou V. Detection in two hospitals of transferable ceftazidime-avibactam resistance in Klebsiella pneumoniae due to a novel VEB beta-lactamase variant with a Lys234Arg substitution, Greece, 2019. Euro surveillance : bulletin Europeen sur les maladies transmissibles = European communicable disease bulletin 2020; 25.
- Livermore DM, Struelens M, Amorim J, Baquero F, Bille J, Canton R et al. Multicentre evaluation of the VITEK 2 Advanced Expert System for interpretive reading of antimicrobial resistance tests. J Antimicrob Chemother 2002; 49: 289-300.
- Garcia-Fulgueiras V, Magallanes C, Reyes V, Cayota C, Galiana A, Vieytes M et al. In Vivo High Plasticity of Multi-Drug Resistant ST258 Klebsiella pneumoniae. Microb Drug Resist 2021; 27: 1126-1130.
- Albarri O, AlMatar M, Ocal MM, Koksal F. Overexpression of Efflux Pumps AcrAB and OqxAB Contributes to Ciprofloxacin Resistance in Clinical Isolates of K. pneumonia. Curr Protein Pept Sci 2022; 23: 356-368.
- Diez-Aguilar M, Canton R. New microbiological aspects of fosfomycin. Rev Esp Quimioter 2019; 32 Suppl 1: 8-18.
- Sugita K, Aoki K, Komori K, Nagasawa T, Ishii Y, Iwata S et al. Molecular Analysis of bla(KPC-2)-Harboring Plasmids: Tn4401a Interplasmid Transposition and Tn4401a-Carrying ColRNAI Plasmid Mobilization from Klebsiella pneumoniae to Citrobacter europaeus and Morganella morganii in a Single Patient. mSphere 2021; 6: e0085021.
- Di Martino P, Cafferini N, Joly B, Darfeuille-Michaud A. Klebsiella pneumoniae type 3 pili facilitate adherence and biofilm formation on abiotic surfaces. Res Microbiol 2003; 154: 9-16.
- Lam MMC, Wick RR, Wyres KL, Gorrie CL, Judd LM, Jenney AWJ et al. Genetic diversity, mobilisation and spread of the yersiniabactin-encoding mobile element ICEKp in Klebsiella pneumoniae populations. Microb Genom 2018; 4.
- Zhu J, Wang T, Chen L, Du H. Virulence Factors in Hypervirulent Klebsiella pneumoniae. Front Microbiol 2021; 12: 642484.
- Di Pilato V, Henrici De Angelis L, Aiezza N, Baccani I, Niccolai C, Parisio EM et al. Resistome and virulome accretion in an NDM-1-producing ST147 sublineage of Klebsiella pneumoniae associated with an outbreak in Tuscany, Italy: a genotypic and phenotypic characterisation. Lancet Microbe 2022; 3: e224-e234.
- Miro E, Rossen JWA, Chlebowicz MA, Harmsen D, Brisse S, Passet V et al. Core/Whole Genome Multilocus Sequence Typing and Core Genome SNP-Based Typing of OXA-48-Producing Klebsiella pneumoniae Clinical Isolates From Spain. Front Microbiol 2019; 10: 2961.
- Snitkin ES, Zelazny AM, Thomas PJ, Stock F, Group NCSP, Henderson DK et al. Tracking a hospital outbreak of carbapenem-resistant Klebsiella pneumoniae with whole-genome sequencing. Sci Transl Med 2012; 4: 148ra116.
- Marsh JW, Krauland MG, Nelson JS, Schlackman JL, Brooks AM, Pasculle AW et al. Genomic Epidemiology of an Endoscope-Associated Outbreak of Klebsiella pneumoniae Carbapenemase (KPC)-Producing K. pneumoniae. PLoS One 2015; 10: e0144310.
- Onori R, Gaiarsa S, Comandatore F, Pongolini S, Brisse S, Colombo A et al. Tracking Nosocomial Klebsiella pneumoniae Infections and Outbreaks by Whole-Genome Analysis: Small-Scale Italian Scenario within a Single Hospital. J Clin Microbiol 2015; 53: 2861-2868.
- Arnold BJ, Gutmann MU, Grad YH, Sheppard SK, Corander J, Lipsitch M et al. Weak Epistasis May Drive Adaptation in Recombining Bacteria. Genetics 2018; 208: 1247-1260.
- Dallman T, Ashton P, Schafer U, Jironkin A, Painset A, Shaaban S et al. SnapperDB: a database solution for routine sequencing analysis of bacterial isolates. Bioinformatics 2018; 34: 3028-3029.
Figure 1.
MICs (μg/mL) heatmap of the 24 clinical isolates of carbapenem-resistant K. pneumoniae (CRKP) isolates according to CLSI.
Figure 1.
MICs (μg/mL) heatmap of the 24 clinical isolates of carbapenem-resistant K. pneumoniae (CRKP) isolates according to CLSI.
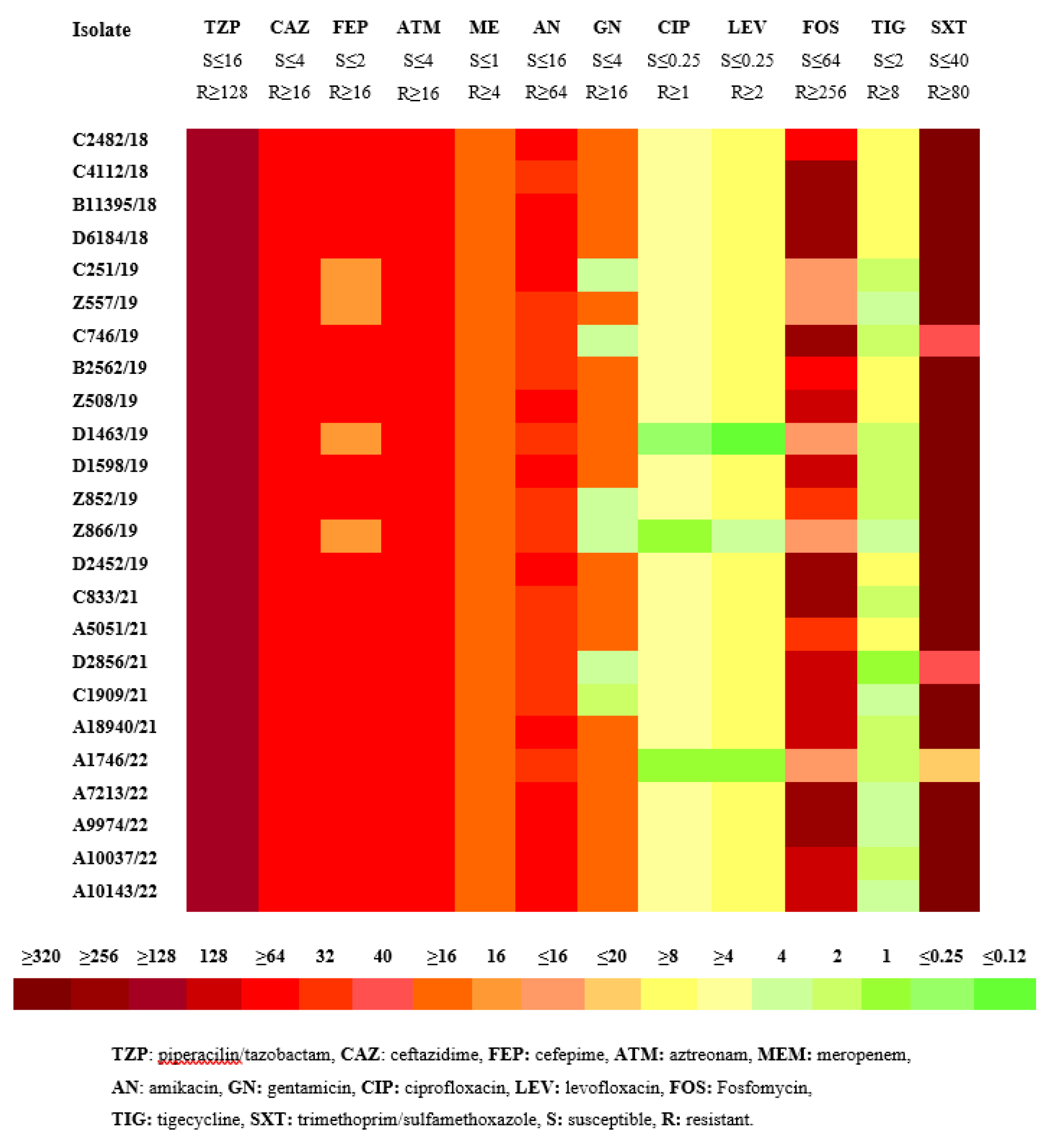
Figure 2.
Circular genome map of (A) seven ST11 CRKP strains, (B) the seven ST39 CRKP strains, (C) the three ST323 CRKP strains and (D) the three ST15 CRKP strains. The genome maps were produced through the BLAST Ring Image Generator (BRIG) software [27]. The genomes of the seven ST11 CRKP isolates (A10143, A7213, A9974, C4112, C746, C833 and D2856), the seven ST39 CRKP isolates (B11395, C2482, D1598, D2452, D6184, Z508 and C1909), the three ST323 CRKP strains (Z557, D1463 and Z866) and the three ST15 CRKP strains (A10037, A5051 and B2562) were mapped separately to the reference genome K. pneumoniae NC_012731. From the inner to the external ring, the innermost ring displays the size of the genome in kbp, followed by GC content (black), GC skew (dark green and dark purple) and K. pneumoniae NC_012731 (red). The white vertical gaps represent sequences of the K. pneumoniae NC_012731 reference strain that are absent in the sequences of the ST11, ST39, ST323 and ST15 CRKP strains or present sequences <50% identical to the reference genome.
Figure 2.
Circular genome map of (A) seven ST11 CRKP strains, (B) the seven ST39 CRKP strains, (C) the three ST323 CRKP strains and (D) the three ST15 CRKP strains. The genome maps were produced through the BLAST Ring Image Generator (BRIG) software [27]. The genomes of the seven ST11 CRKP isolates (A10143, A7213, A9974, C4112, C746, C833 and D2856), the seven ST39 CRKP isolates (B11395, C2482, D1598, D2452, D6184, Z508 and C1909), the three ST323 CRKP strains (Z557, D1463 and Z866) and the three ST15 CRKP strains (A10037, A5051 and B2562) were mapped separately to the reference genome K. pneumoniae NC_012731. From the inner to the external ring, the innermost ring displays the size of the genome in kbp, followed by GC content (black), GC skew (dark green and dark purple) and K. pneumoniae NC_012731 (red). The white vertical gaps represent sequences of the K. pneumoniae NC_012731 reference strain that are absent in the sequences of the ST11, ST39, ST323 and ST15 CRKP strains or present sequences <50% identical to the reference genome.
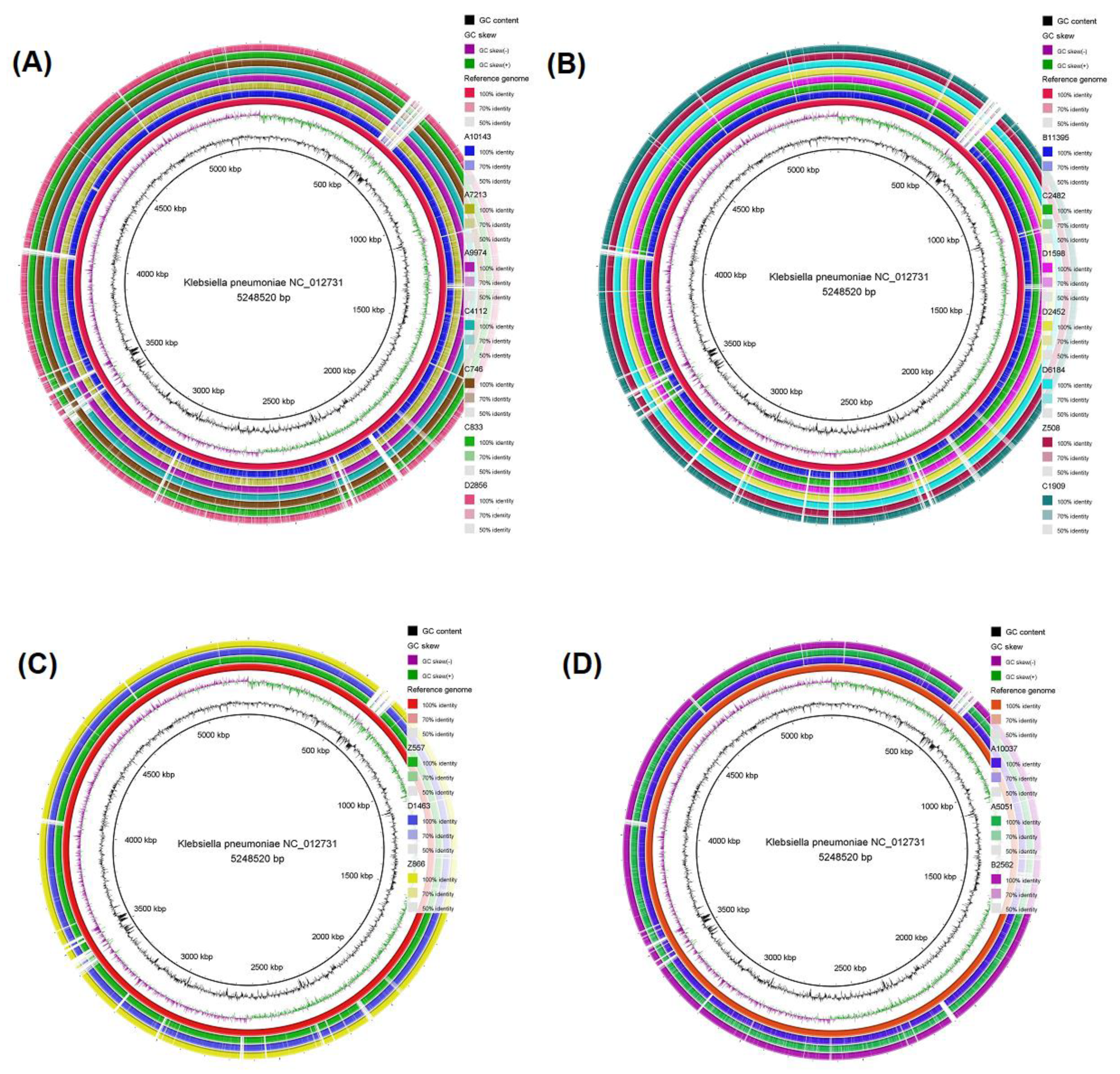
Figure 3.
The tree built on the entire core genome with PhyML. The branch color indicates the fraction of supporting versus supporting plus clashing SNPs for each branch of the core tree, which support the initial bipartition of the strains. MLST types: ST11: D2856, C4112, C833, C746, A10143, A9974, A7213, ST512: A18940, ST30: C251, ST307: Z852, ST323: Z866, D1463, Z557, ST35: A1746, ST15: B2562, A5051, A10037, ST39: D1598, D2452, C2482, Z508, C1909, B11395, D6184.
Figure 3.
The tree built on the entire core genome with PhyML. The branch color indicates the fraction of supporting versus supporting plus clashing SNPs for each branch of the core tree, which support the initial bipartition of the strains. MLST types: ST11: D2856, C4112, C833, C746, A10143, A9974, A7213, ST512: A18940, ST30: C251, ST307: Z852, ST323: Z866, D1463, Z557, ST35: A1746, ST15: B2562, A5051, A10037, ST39: D1598, D2452, C2482, Z508, C1909, B11395, D6184.
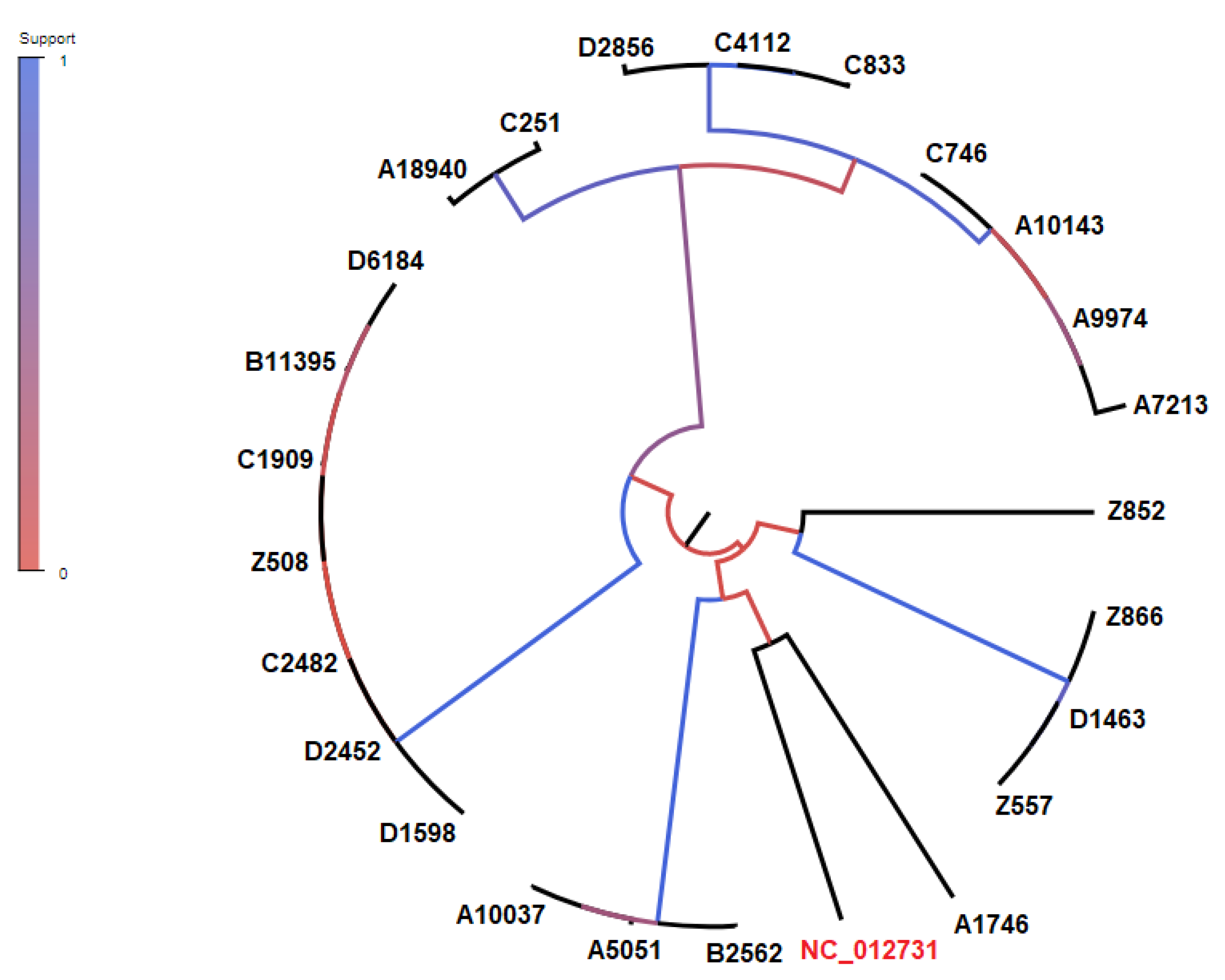
Figure 4.
Pairwise analysis of recombination between carbapenem-resistant K. pneumoniae CRKP strains. (A-C) SNP densities (SNPs per kilobase) along the core genome for three pairs of CRKP strains with various nucleotide diversities. (D-F) Corresponding histograms for the number of SNPs per kilobase (dots) together with fits of the Poisson mixture model for C4112-C746 (green), A18940-D2856 (red), and A5051-C2482 (blue) CRKP strains. The vertical axis is on a logarithmic scale. (G) For each pair of CRKP strains (dots), the fraction of the genome that was inherited clonally is shown as a function of the nucleotide divergence of the pair, shown on a logarithmic scale. The three CRKP pairs that are highlighted in panels (A-F) are depicted as the green, red, and blue dots. (H) Fraction of all SNPs that lie in recombined regions as a function of the clonally inherited fraction of the genome. (I) Cumulative distribution of the clonal fractions of CRKP pairs. (J) Cumulative distribution of the lengths of recombined segments for CRKP pairs that are in the mostly clonal regime.
Figure 4.
Pairwise analysis of recombination between carbapenem-resistant K. pneumoniae CRKP strains. (A-C) SNP densities (SNPs per kilobase) along the core genome for three pairs of CRKP strains with various nucleotide diversities. (D-F) Corresponding histograms for the number of SNPs per kilobase (dots) together with fits of the Poisson mixture model for C4112-C746 (green), A18940-D2856 (red), and A5051-C2482 (blue) CRKP strains. The vertical axis is on a logarithmic scale. (G) For each pair of CRKP strains (dots), the fraction of the genome that was inherited clonally is shown as a function of the nucleotide divergence of the pair, shown on a logarithmic scale. The three CRKP pairs that are highlighted in panels (A-F) are depicted as the green, red, and blue dots. (H) Fraction of all SNPs that lie in recombined regions as a function of the clonally inherited fraction of the genome. (I) Cumulative distribution of the clonal fractions of CRKP pairs. (J) Cumulative distribution of the lengths of recombined segments for CRKP pairs that are in the mostly clonal regime.
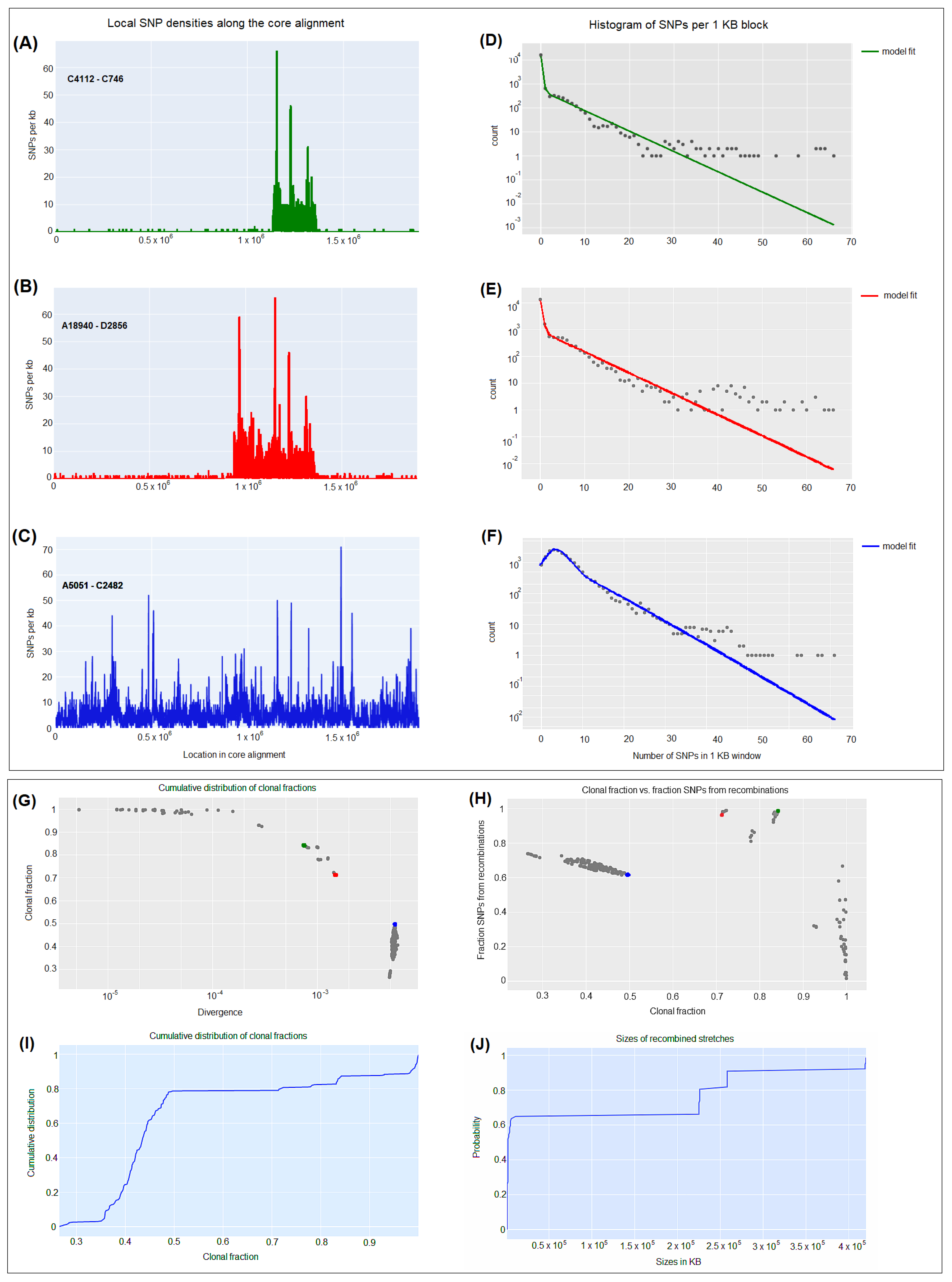
Figure 5.
SNP compatibility along the core genome alignment displays the shortness of tree-compatible segments. (A) Probability distribution of the number of consecutive SNP columns that are consistent with a common phylogeny for the core genome alignment. (B) Probability distribution of the number of consecutive nucleotides consistent with a common phylogeny for the core genome alignment.
Figure 5.
SNP compatibility along the core genome alignment displays the shortness of tree-compatible segments. (A) Probability distribution of the number of consecutive SNP columns that are consistent with a common phylogeny for the core genome alignment. (B) Probability distribution of the number of consecutive nucleotides consistent with a common phylogeny for the core genome alignment.
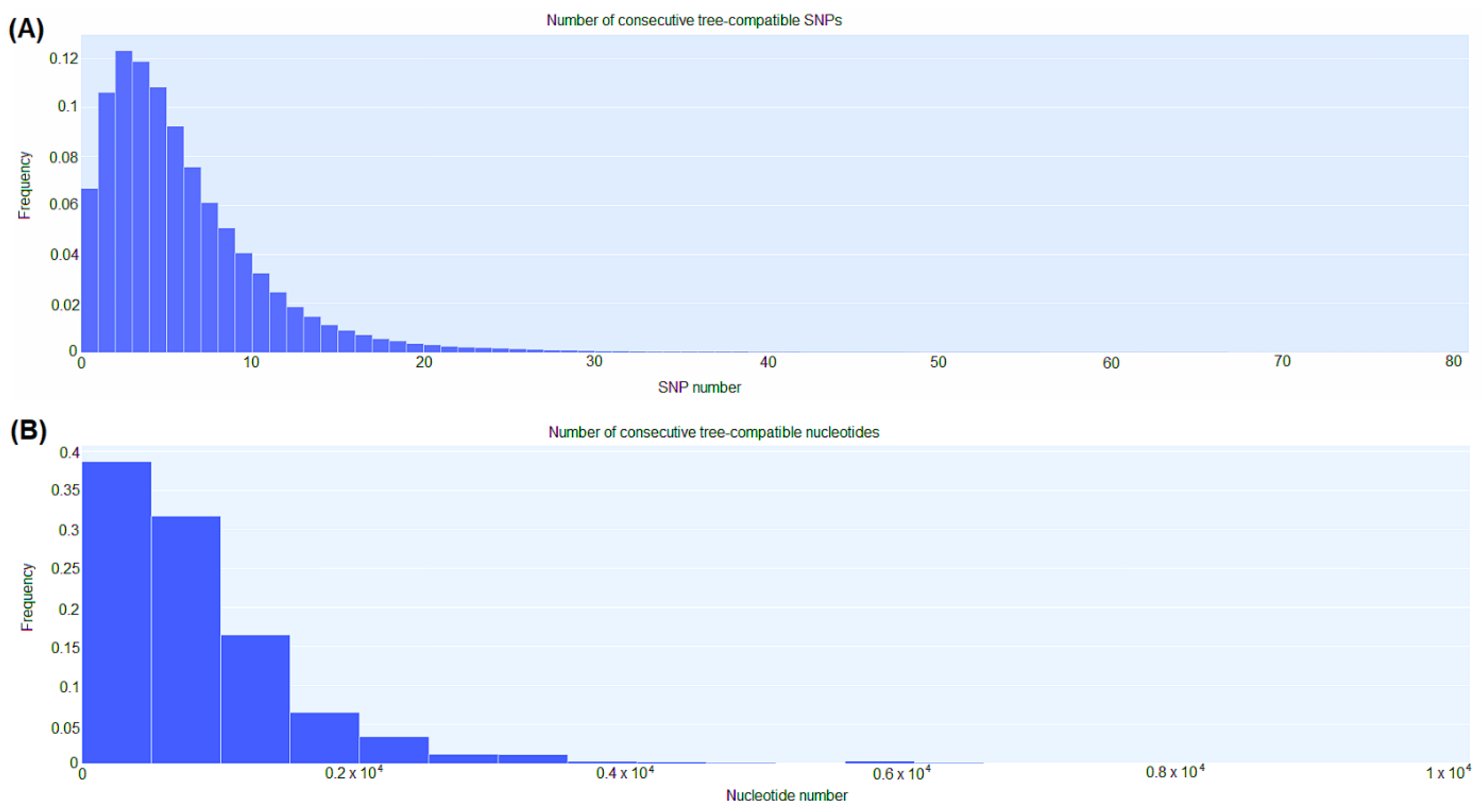
Figure 6.
Ratio C/S of the minimal number of phylogeny changes C to substitutions S for random subsets of carbapenem-resistant K. pneumoniae (CRKP) strains using the alignment from which 5% of potentially homoplastic positions have been removed. For CRKP strain numbers ranging from n=4 to n=24, random subsets of n strains were collected and the ratios C/S of phylogeny changes to SNPs in the alignment were calculated. The figure shows box-whisker plots that indicate, for each CRKP strain number n, means and standard deviations for C/S across subsets. The horizontal red line displays C/S=0.15.
Figure 6.
Ratio C/S of the minimal number of phylogeny changes C to substitutions S for random subsets of carbapenem-resistant K. pneumoniae (CRKP) strains using the alignment from which 5% of potentially homoplastic positions have been removed. For CRKP strain numbers ranging from n=4 to n=24, random subsets of n strains were collected and the ratios C/S of phylogeny changes to SNPs in the alignment were calculated. The figure shows box-whisker plots that indicate, for each CRKP strain number n, means and standard deviations for C/S across subsets. The horizontal red line displays C/S=0.15.
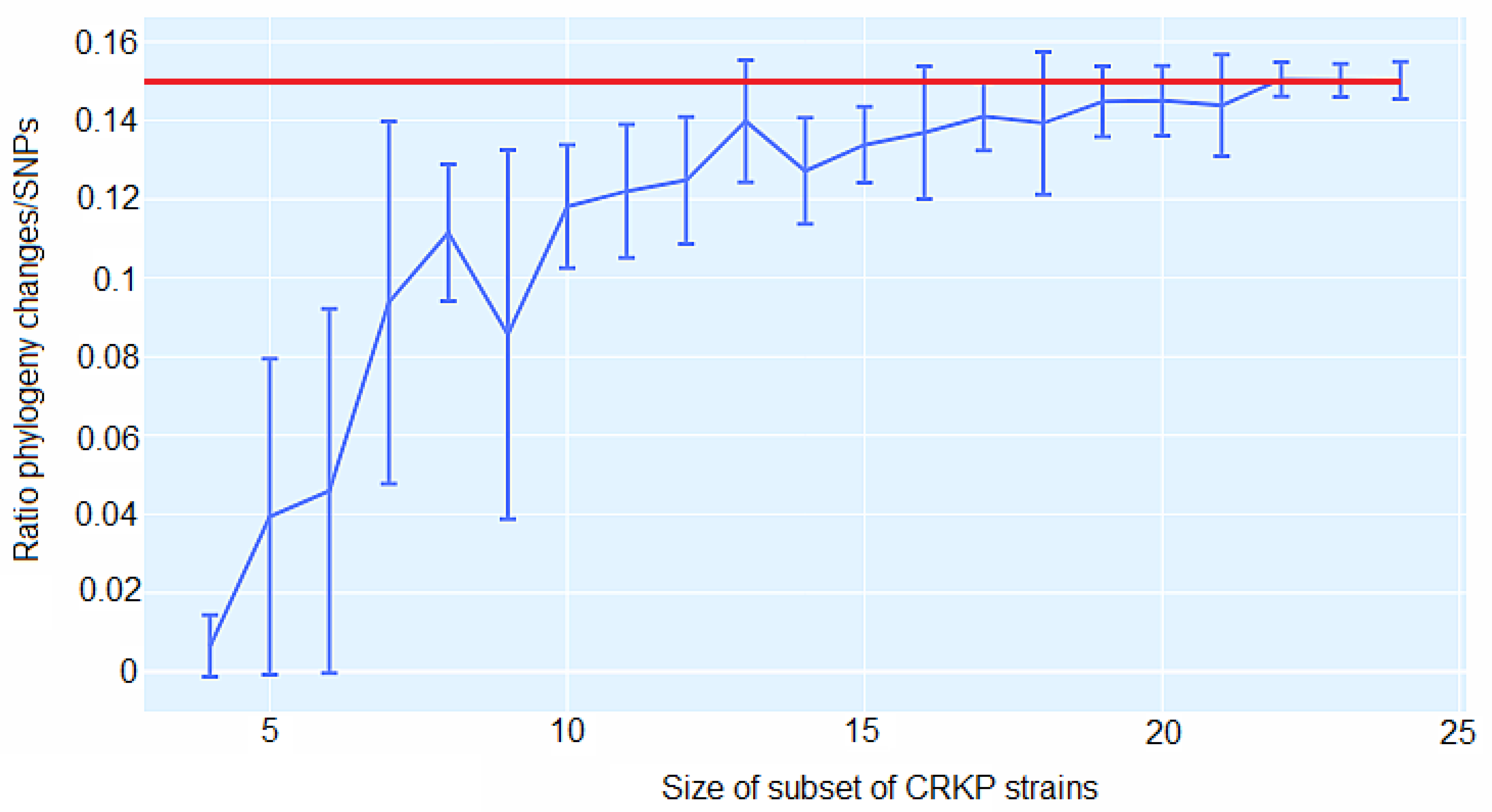

Table 1.
Clinical data of carbapenem-resistant Klebsiella pneumoniae isolates.
| Strain | Date of isolation | Sex | Age (years) |
Ward | Biological sample | Infection / Colonization |
|---|---|---|---|---|---|---|
| C2482/18 | 9/6/2018 | F | 8 | PICU | Rectal swab | Colonization |
| C4112/18 | 7/9/2018 | F | 56 | ICU | Rectal swab | Colonization |
| B11395/18 | 24/10/2018 | F | 1 | PICU | Urine | Infection |
| D6184/18 | 19/11/2018 | F | 8 | PICU | Bronchial aspirate | Infection |
| C251/19 | 17/1/2019 | F | 17 | PICU | Rectal swab | Colonization |
| Z557/19 | 30/1/2019 | M | 57 | ICU | Rectal swab | Colonization |
| C746/19 | 7/2/2019 | M | 73 | ICU | Rectal swab | Colonization |
| B2562/19 | 4/3/2019 | M | 79 | ICU | Urine | Infection |
| Z508/19 | 20/3/2019 | F | 60 | ICU | Rectal swab | Colonization |
| D1463/19 | 22/3/2019 | M | 55 | ICU | Bronchial aspirate | Infection |
| D1598/19 | 1/4/2019 | M | 40 | ICU | Bronchial aspirate | Infection |
| Z852/19 | 12/4/2019 | M | 3 | PICU | Rectal swab | Colonization |
| Z866/19 | 15/4/2019 | M | 49 | ICU | Rectal swab | Colonization |
| D2452/19 | 24/5/2019 | M | 73 | ICU | Bronchial aspirate | Infection |
| C833/21 | 31/3/2021 | F | 57 | ICU | Wound | Infection |
| A5051/21 | 25/4/2021 | F | 69 | ICU | Blood | Infection |
| D2856/21 | 7/7/2021 | F | 54 | ICU | Bronchial aspirate | Infection |
| C1909/21 | 8/7/2021 | M | 85 | ICU | Wound | Infection |
| A18940/21 | 24/12/2021 | F | 58 | ICU | Blood | Infection |
| A1746/22 | 25/2/2022 | M | 0.25 | PICU | Blood | Infection |
| A7213/22 | 3/5/2022 | F | 30 | ICU | Blood | Infection |
| A9974/22 | 16/6/2022 | F | 67 | ICU | Blood | Infection |
| A10037/22 | 16/6/2022 | F | 53 | ICU | Blood | Infection |
| A10143/22 | 19/6/2022 | M | 88 | ICU | Blood | Infection |
ICU: Intensive Care Unit, PICU: Pediatric Intensive Care Unit.
Table 2.
Genetic characteristics of the 24 carbapenem-resistant K. pneumoniae isolates of the study.
Table 2.
Genetic characteristics of the 24 carbapenem-resistant K. pneumoniae isolates of the study.
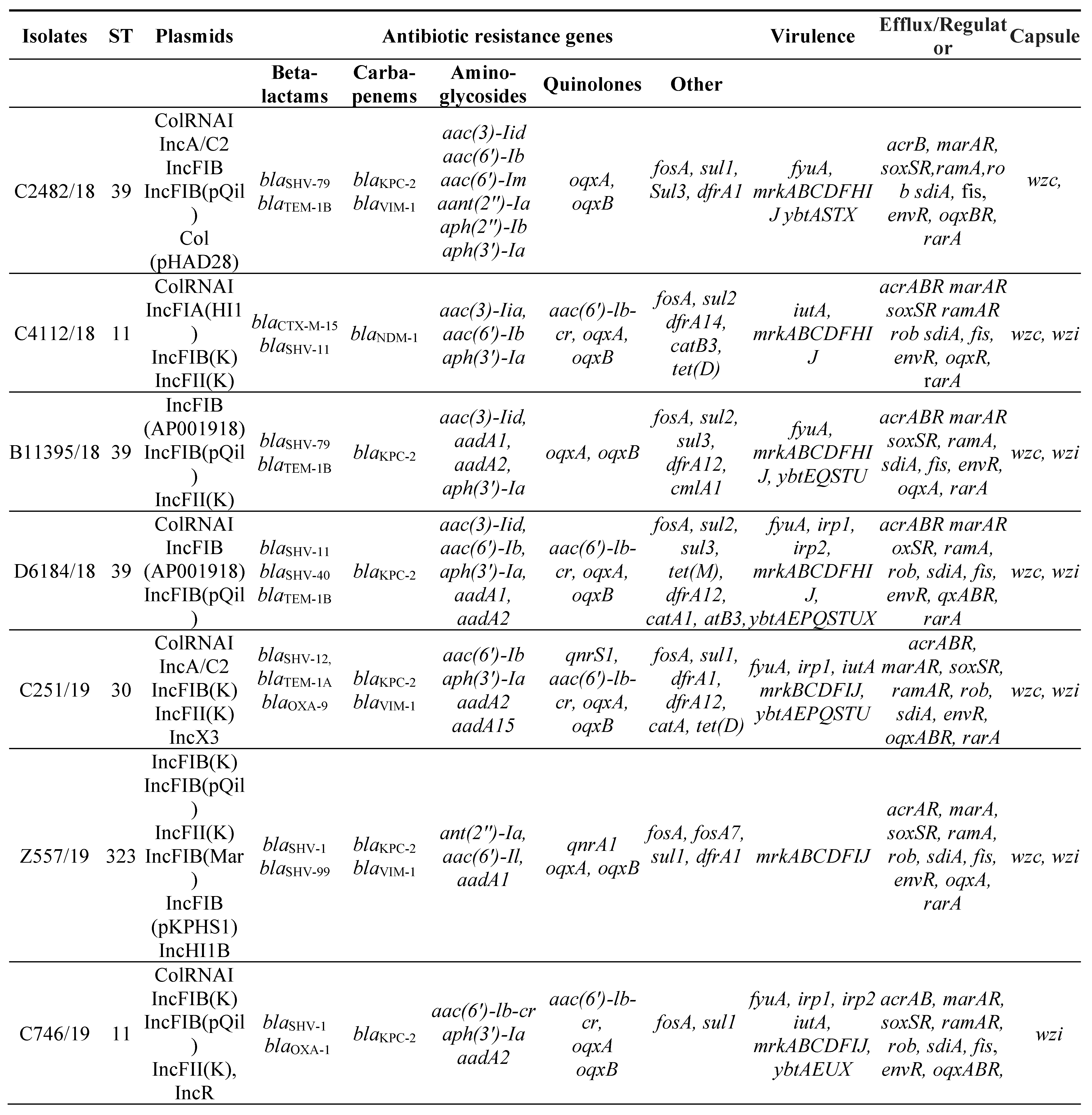
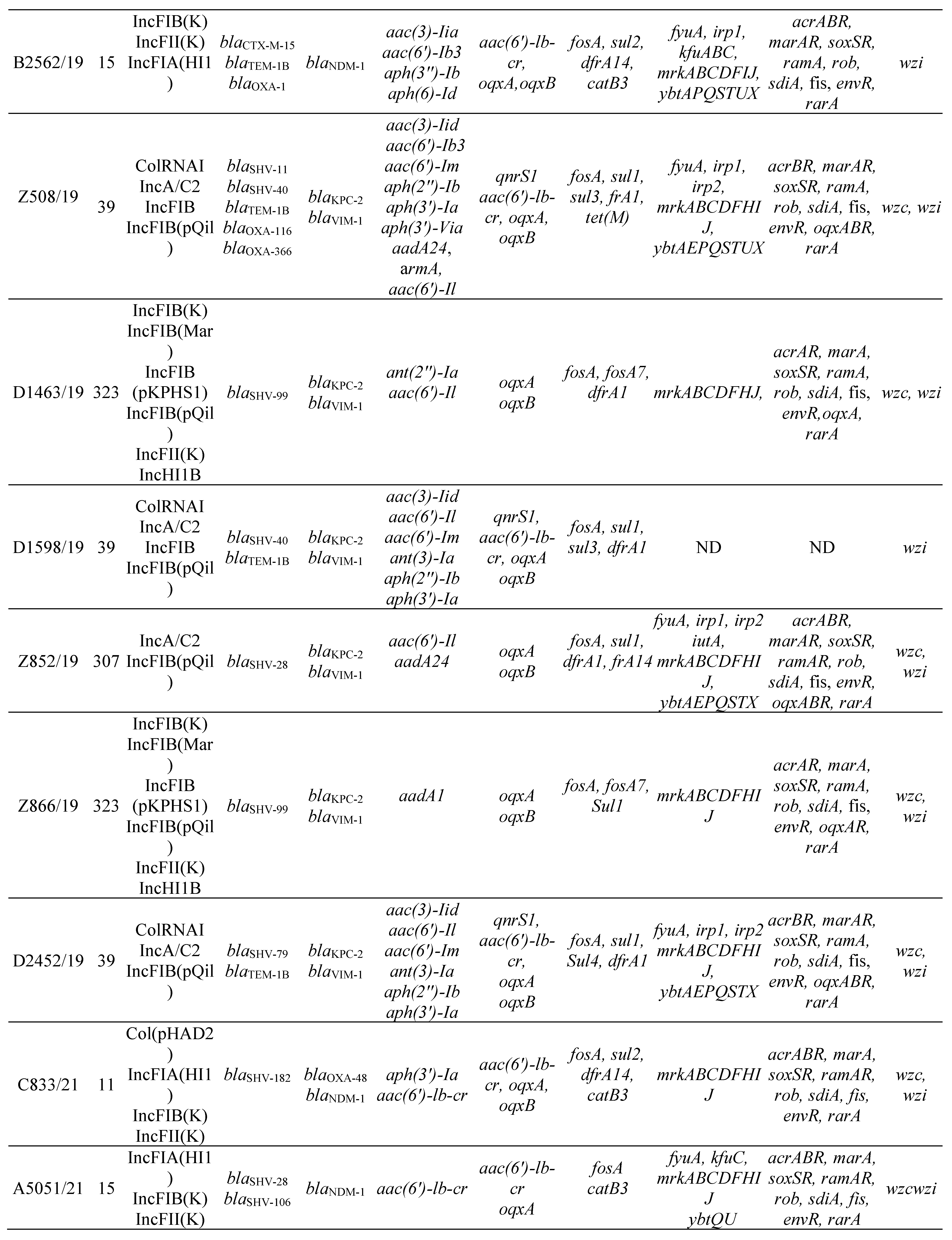
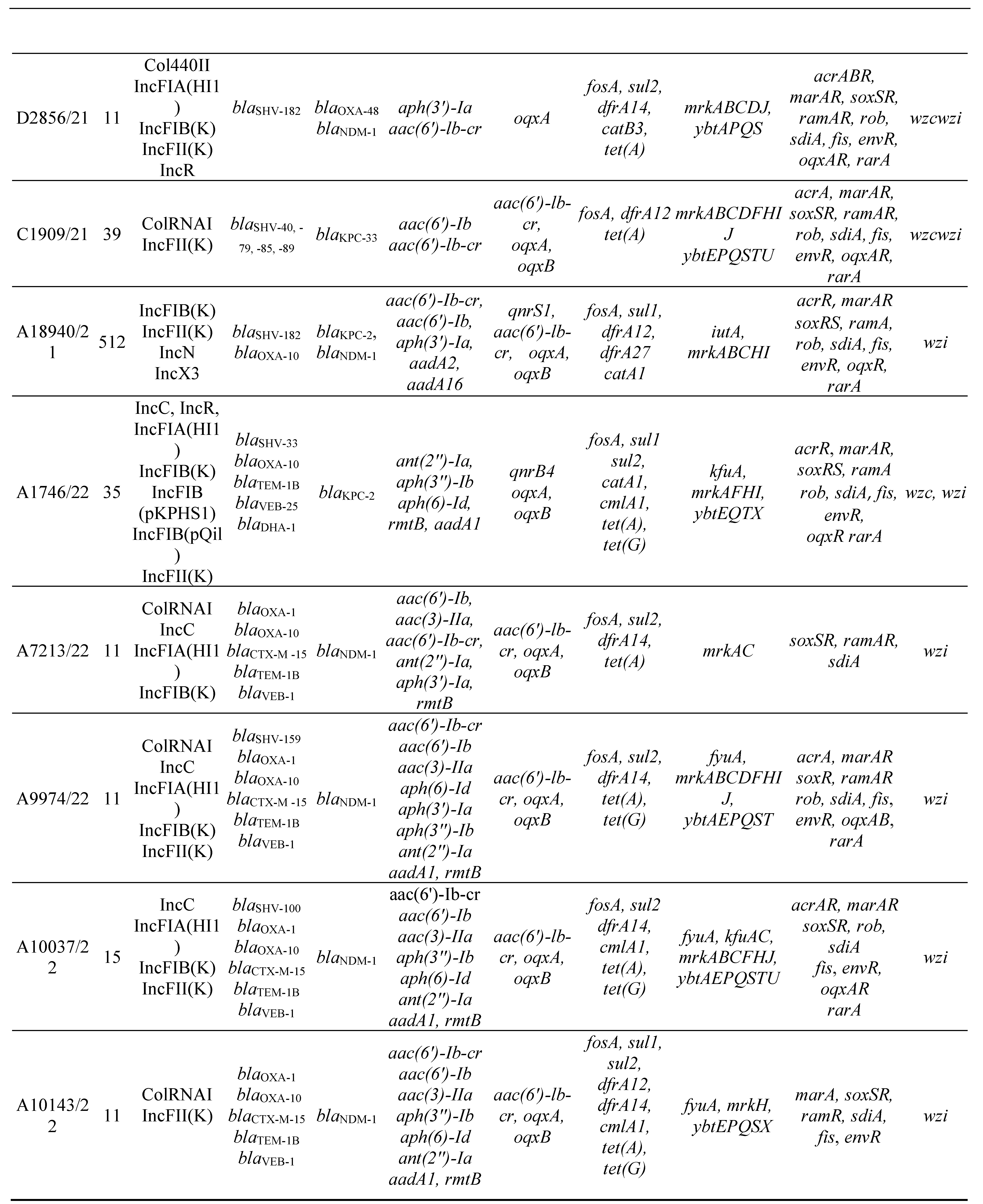
*Yersiniabactin cluster: ybt, irp and fyuA genes; Aerobactin cluster: iuc and iut genes; AcrAB efflux pump: acr, mar, sox, rob, ram sdiA, fis and envR, genes; OqxAB efflux pump: oqx and rar genes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



