
Submitted:
15 May 2023
Posted:
16 May 2023
You are already at the latest version
Abstract
Glutamate NMDA receptor (NMDAR) hypofunction has been proposed to underlie schizophrenia symptoms because administration of NMDAR antagonists reproduces behavioral and molecular schizophrenia-like phenotypes in healthy humans and animal models. However, the role of specific NMDAR subunits in these schizophrenia-relevant characteristics is largely unknown. Mounting evidence implicates the GluN2D subunit of NMDAR in some of these symptoms and pathology. Firstly, genetic and post-mortem studies show changes in the GluN2D subunit in people with schizophrenia. Secondly, the psychosis-inducing effects of NMDAR antagonists are blunted in GluN2D-knockout mice, suggesting that the GluN2D subunit mediates NMDAR antagonist-induced psychotomimetic effects. Thirdly, in the mature brain, the GluN2D subunit is relatively enriched in parvalbumin (PV)-containing interneurons, a cell type hypothesized to underlie the cognitive symptoms of schizophrenia. Lastly, the GluN2D subunit is widely and abundantly expressed early in development which could be of importance considering schizophrenia is a disorder that has its origins in early neurodevelopment. The limitations of currently available therapies warrant further research into novel therapeutic targets such as the GluN2D subunit which may help us better understand underlying disease mechanisms and develop novel and more effective treatment options.
Keywords:
GluN2D
; Schizophrenia
; NMDA receptor
; NMDAR antagonists
1. Schizophrenia – an overview
Schizophrenia is a severe, debilitating, chronic neuropsychiatric disorder with a lifetime prevalence of 0.72% [1]. It has a complex and heterogeneous presentation making it difficult to diagnose and to identify a consistent underlying aetiology or pathology. People with schizophrenia demonstrate high inter-individual variability with respect to symptoms, disease course, outcome and response to treatment. Schizophrenia symptoms are divided into three main classes: positive, negative and cognitive. Of the three, positive symptoms are the most easily identifiable and are defined as psychotic features that are not usually present in healthy people, and include hallucinations, delusions and disorganised speech and behaviour [2,3]. Negative symptoms refer to a reduction or disruption in normal emotions and behaviours manifesting as social and emotional withdrawal, apathy, and avolition [2,3]. Cognitive symptoms vary in severity amongst patients and include elements such as deficits in verbal memory, working memory, executive functioning, cognitive flexibility and processing speed [2,4].
Schizophrenia typically has an early onset, with most patients being diagnosed in their late adolescence - early adulthood years. The onset of the first psychotic episode is usually preceded by a prodromal period during which symptoms gradually emerge and this period can last several years. An early onset combined with long-term deficits in social, educational and occupational function make this disorder one of the leading causes of chronic disability with significant impacts on the quality of life of patients and their families and caregivers [5]. Additionally, this disorder places a substantial economic burden on health care systems with estimates of annual associated costs reaching up to $150 billion in the United States and $4.9 billion in Australia [6,7].
The causes of schizophrenia are not fully understood but are thought to be multifactorial, involving a complex interplay between multiple genetic variants and environmental factors. Genome-wide association studies have identified multiple common variants of small effect spanning over 250 genetic loci, suggesting that schizophrenia is a polygenic disorder in most patients [8,9,10]. Genes associated with schizophrenia risk are involved in various functions including the regulation of the postsynaptic membrane, synaptic transmission, and neurodevelopmental pathways including glutamate pathways [10,11,12]. Non-genetic factors that increase life-time risk for schizophrenia include obstetric complications, advanced paternal age, living in an urban setting, childhood trauma or adversity, cannabis use and first generation migration [2,13,14,15,16,17,18,19]. Although there have been advances in our understanding of risk factors associated with schizophrenia, the etiological complexity has made it a challenge to identify the underlying disease mechanisms and find effective cures and preventative strategies.
Currently available pharmacological treatments, chiefly conventional and atypical anti-psychotics, and psychotherapy, have proven clinical utility and can help manage positive symptoms in some patients. However, antipsychotic medications have limited efficacy and are poorly tolerated with substantial side effects in approximately 30% of people [20,21,22]. Moreover, they usually offer little benefit in improving negative and cognitive symptoms, and these symptom types therefore remain a pressing, unmet medical need [23]. The prevalence, burden and current lack of effective treatments for schizophrenia highlight a need to improve our understanding of the underlying mechanisms and neurobiology of the disorder in order to identify and develop better treatments.
2. NMDA receptor hypothesis of schizophrenia
Although the predominant literature has focused on dopaminergic dysfunction in schizophrenia [24,25,26], and many of the currently available anti-psychotics target dopaminergic receptors [27,28,29,30,31], another prevailing theory for the aetiology of schizophrenia gaining increasing support is the N-methyl-D-aspartate receptor (NMDAR) hypothesis [32,33,34]. This hypothesis attributes hypofunction of the NMDAR, one of the major glutamate receptor subtypes, to many of the symptoms of schizophrenia. From the late 1950s, phencyclidine (PCP) and ketamine have been reported to induce positive, negative, and cognitive phenotypes such as psychosis-like dissociative states and neurocognitive disturbances in healthy individuals like those observed in schizophrenia patients [35,36,37,38,39,40]. Furthermore, these drugs exacerbate symptoms including psychosis in individuals with schizophrenia [38,41,42]. The NMDAR hypothesis was proposed soon after, in the late 1970s – 1980s, when it was found that PCP and ketamine carry out their actions via NMDAR blockade [36,43,44,45]. The extensive literature drawing parallels between NMDAR antagonism and many of the unique features of schizophrenia have led some to postulate that these antagonists might act via the same mechanisms that become dysfunctional in this disorder [33,46,47].
Importantly, NMDAR antagonists can reproduce symptoms from all three schizophrenia symptom domains (positive, negative and cognitive) whereas alterations to other neurotransmitter systems such as dopamine have been predominantly linked only to positive symptoms [35,48]. Furthermore, NMDAR are ubiquitously expressed throughout the central nervous system, including several regions implicated in schizophrenia, like the frontal cortex where dopaminergic receptors are expressed at relatively lower levels [33]. It has been suggested that the disruptions to the dopaminergic and GABAergic systems seen in schizophrenia might be secondary to deficits in the glutamatergic system as NMDARs are known to regulate dopamine release and GABAergic interneuron activity (Figure 1) [46,49]. To further elaborate this proposition, presynaptic dopamine release in the prefrontal cortex (PFC) and striatum is thought to be regulated by GABAergic receptors found on presynaptic dopaminergic terminals. These GABAergic receptors are, in turn, activated by GABA released from inhibitory interneurons. The NMDARs localised on the interneurons are responsible for regulating this GABA release, such that stimulation of the receptors leads to enhanced GABA release [50]. In this way, any disruption to normal NMDAR function would lead to downstream GABAergic and dopaminergic dysregulation. In line with this theory, studies conducted in healthy humans and in rodent models show that administration of NMDAR antagonists can reproduce schizophrenia-like dopaminergic dysfunction including cortical disinhibition, and an increase in striatal D2 receptors and dopamine release [51,52,53,54]. Moreover, as NMDARs mediate excitatory drive onto inhibitory GABAergic interneurons, which in turn provide negative feedback to cortical pyramidal neurons, inhibition of these NMDARs leads counterintuitively to an increase in cortical excitation by disinhibition [55,56,57,58]. This could result in neurotoxicity and an alteration in the excitatory/inhibitory (E/I) balance in cortical circuits that is crucial for normal brain function. In this way, glutamatergic dysfunction might be driving the changes in the dopaminergic system and resulting in an E/I imbalance which in turn results in the positive, negative and cognitive symptoms of schizophrenia.
In recent years, several schizophrenia candidate genes that affect NMDAR signaling have been identified, including GRIN1, GRIN2A, GRIN2B and GRIN2D which are genes encoding the GluN1, GluN2A, GluN2B and GluN2D subunits of the NMDAR respectively, as well as the gene encoding D-amino acid oxidase (DAAO), which regulates the availability of D-serine, a co-agonist at NMDARs [10,59,60,61]. Findings from other genetic and post-mortem patient studies have further suggested that NMDAR signalling is disrupted in schizophrenia [62,63,64,65]. Additionally, studies in rodents have shown that knockdown of the obligatory GluN1 subunit of the NMDAR can result in phenotypes relevant to schizophrenia including hyperlocomotion (proposed to have translational relevance for psychosis in rodents), social deficits, impaired learning and abnormal neuronal oscillations [66,67,68,69,70].
Together these findings promote the hypothesis that underlying the key behavioral and cognitive symptoms of schizophrenia is the dysfunction and/or dysregulation of NMDAR-mediated neurotransmission [46]. The NMDAR hypofunction model has since become one of the most commonly adopted and supported models of schizophrenia and is often employed to study the etiology and pathology of the disorder as well as for the development of novel treatment strategies.
3. Glutamatergic neurotransmission in the CNS
Glutamate plays a key role in mediating the homeostatic balance between excitation and inhibition in the brain, cortico-cortical neurotransmission, neuronal development, neurodegeneration, nervous system plasticity and learning and memory [71]. All pyramidal neurons are glutamatergic, as are 60% of total brain neurons. Glutamate carries out its actions through its receptors which are divided into two groups. The ionotropic receptors, namely the NMDA, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) and kainate receptors, are integral membrane proteins composed of 4 large subunits (>900 residues) and act as ligand-gated cation channels, whilst the metabotropic receptors are G-protein coupled and activate intracellular biochemical cascades [72,73]. As this review is focused on NMDA receptors, specifically the GluN2D subunit and its relevance to schizophrenia, we direct readers to the following review articles [74,75,76] for further discussion of the role of AMPA, kainate and metabotropic receptors in schizophrenia.
4. NMDA receptor structure and function
NMDARs are a subtype of glutamate receptor that are widely distributed and can be found on both neuronal and non-neuronal cells. NMDARs are heteromeric, composed of multiple different subunits arranged to form a central ion channel pore that is highly permeable to calcium, but also conducts sodium and potassium [77]. These receptors play a key role in many physiological processes including neurodevelopment, synaptogenesis, locomotion, and due to their role as critical mediators of activity-dependent synaptic plasticity, they are especially important for learning, memory formation, and other forms of cognition [62,78,79]. NMDARs are unique in that they require the concomitant binding of glycine and glutamate along with membrane depolarisation for activation and are thus referred to as ‘molecular coincidence detectors’ [80,81,82]. At resting membrane potential, extracellular magnesium (Mg2+) can be found in the channel pore, blocking NMDARs in a voltage-dependent manner. Partial depolarisation of the neuron relieves the blockade, opening the channel. The subsequent Ca2+influx into the neuron triggers a cascade of events that can influence both local, acute functional synaptic plasticity and, via changes in gene expression, sustained neural plasticity [83]. The calcium permeability of NMDARs allows them to initiate long term potentiation (LTP) and long term depression (LTD) in the hippocampus and cortex [84,85].
The heteromeric composition of NMDARs enables different pharmacological, biophysical and functional properties for the receptor, and these vary in distribution and expression both regionally and temporally throughout development. The different subunits are termed GluN1, GluN2 and GluN3. The GluN1 subunit is encoded by a single gene but has eight splice variants [86]. There are four different GluN2 subunits (A-D), encoded by four different genes and two different Glun3 subunits (A and B) [78,87]. A functional NMDAR is typically composed of two GluN1 subunits and two subunits from among the GluN2A-D and GluN3A-B subunits [79,88]. The obligatory GluN1 subunit is ubiquitously expressed throughout the brain and over the lifespan [78,89]. Of the GluN2 subunits, which have a more varied and complex temporal and spatial expression, GluN2A and GluN2B are the predominant subtypes found in the adult human brain whilst the GluN2C and GluN2D subunits are more highly expressed in the developing brain [79,90]. The subunit composition of NMDARs influences its functional properties including agonist affinity, Mg2+ block, decay kinetics, and modulation by polyamines [91,92]. Each of the NMDAR subunits consists of four domains: the intracellular C-terminal domain is responsible for receptor trafficking and intracellular signalling, the transmembrane domain contains the ion channel including the pore, the ligand binding domain binds glycine or glutamate and the extracellular N-terminal domain contains the binding site for modulators like zinc ions [80,87,93]. Given the importance of NMDAR subunits in mediating normal brain function, it is not surprising that dysfunction of these subunits has been linked to various neurological diseases. This review will focus on the relatively understudied GluN2D subunit and its potential role in schizophrenia.
4.1. GluN2D receptor subunit expression and distribution
The GluN2D (common names: NMDAR2D, NR2D, GluRε4) subunit is encoded by the GRIN2D gene which consists of 13 exons, spanning 49.3kB and is located on chromosome 19q13.1-qter in the human genome [78]. The GRIN2D gene has two potential splice isoforms (NR2D-1 and NR2D-2), the longest of which contains 1356 amino acids [78,94]. In rodents, GluN2D has been extensively characterized; expression levels are first detected between embryonic day (E)15-18 during late-embryogenesis, with levels peaking by post-natal day (P)7-10 [95,96]. The GluN2D mRNA and protein expression levels decrease gradually after the early neonatal phase until late adolescence (P40-50) when they reach their relatively low steady-state expression level [91,95,96,97]. During the embryonic and early neonatal phases, the expression of GluN2D is widespread and detected in several regions including the spinal cord, midbrain nuclei, diencephalon (thalamus, hypothalamus), certain basal ganglia nuclei (substantia nigra and subthalamic nucleus), retina, olfactory bulb, cerebellum, the cerebral cortex, and the hippocampus [90,91,94,95,98]. The ubiquitous distribution of the GluN2D subunit during the early phases of life makes this subunit particularly interesting amongst the NMDAR subunits as it suggests that GluN2D plays a critical role in modulating circuit connectivity and function during neurodevelopment. This could be of significance when considering a disorder such as schizophrenia that is thought to have its origins in early development [99,100].
As the rodent ages and GluN2D expression levels reduce, it becomes more localized to distinct cell subtypes [91,95,96]. This is especially apparent in the hippocampus and cortex, where multiple studies have shown that GluN2D is enriched in parvalbumin(PV)-containing GABAergic interneurons in mature rodents whereas its expression and activity on glutamatergic pyramidal cells decreases [98,101,102,103]. The sustained expression of GluN2D-containing receptors on interneurons over development could imply that this subunit is required to maintain proper inhibitory drive and control overall circuit function [104].
In humans, the spatial and temporal expression of GluN2D is thought to be consistent with that reported in rodents, however, it is yet to be well characterised. In human fetal brains, GluN2D mRNA is abundantly expressed and is one of the predominant NMDAR subunits expressed between gestational weeks 8-20 [105]. In contrast, a study of neurologically normal, adult human post mortem brains reported only moderate expression of GluN2D mRNA in the prefrontal, parietal, and motor cortices, where instead the major subtypes expressed were the GluN2A and GluN2B subunits [106]. This study also reported that whilst GluN2D expression was low within most neurons in the hippocampus, expression was moderately intense within a small subset of hippocampal neurons particularly in the hilus, a region containing many interneurons [106]. A recent study which used laser microdissection to isolate pyramidal neurons and PV+ interneurons from the human postmortem dorsolateral PFC (dlPFC) combined with RNA sequencing and microarray provided evidence that the GluN2D subunit is indeed particularly enriched in PV+ interneurons, but not pyramidal neurons [102]. It should be noted however, that whilst the interneurons that were isolated for this study expressed markers specific to PV+ interneurons such as KCNS3, they also expressed markers common to other interneuron types such as somatostatin and thus might not be delimited to PV+ interneurons alone [102]. Another post-mortem in situ hybridization study revealed that in control human brains, the GluN2D subunit is preferentially expressed in layers II, IIIa-c, IV, Va and VIa of the cerebral cortex and at much higher levels in the prefrontal cortex compared with the parieto-temporal or cerebellar cortices [107].
Multiple studies examining the subunit composition of NMDARs in the central nervous system suggest that while the majority of GluN2D subunits are associated with the GluN1 subunit, the GluN2D subunit also forms heteromeric assemblies with GluN2A and/or GluN2B subunits in different brain regions and neuronal subpopulations [108,109,110]. For example, the GluN2D subunit is shown to primarily form a binary complex with GluN1 in the thalamus of young (P7) and adult rats, whereas this binary complex is not detected at all in the rat midbrain, where instead the GluN2D subunit forms triheteromeric assemblies with GluN1 and GluN2A or GluN2B [108]. The varying spatial expression of the different subtypes of GluN2D-containing NMDARs suggest distinct functional roles.
4.2. GluN2D receptor subunit function
Glutamate displays 5-6 times greater potency at GluN2D-containing NMDARs than GluN2A- or GluN2B-containing NMDARs [78,111]. Similarly, GluN1 agonists such as glycine are most potent when the GluN2 subunit in the NMDAR is GluN2D [112]. GluN2D-contaning NMDARs also have a weak Mg2+ block that is 10-fold lower than that of GluN2A- or GluN2B-containing receptors and are also reported to have a slightly lower Ca2+ permeability [113,114]. The resistance to Mg2+ block suggests that neurons expressing the GluN2D subunit may be more responsive to synaptic glutamate release. The non-competitive NMDAR antagonist ketamine is more potent at, and shows approximately 5-fold selectivity for GluN2D-containing receptors, compared with GluN2A or GluN2B subunits [115]. This, combined with the fact that GluN2D-containing receptors are predominantly expressed on interneurons, may suggest that the GluN2D-containing receptors might be involved in the cortical disinhibition induced by certain NMDAR antagonists such as ketamine [102,116]. Another unique feature of GluN2D-containing receptors is that they have the slowest deactivation rate of the GluN2 receptor subtypes; exhibiting 10-fold slower receptor deactivation than GluN2B and GluN2C receptors and 100-fold slower than GluN2A receptors [78,92,117]. Application of glutamate to NMDARs containing the GluN2A subunit was found to generate a macroscopic current with a deactivation time of tens of milliseconds compared with several seconds for GluN2D-containing NMDARs [117]. Importantly, this prolonged decay time is matched with an almost complete lack of desensitization giving this subunit the capacity to shuttle large quantities of ions across the plasma membrane in order to sustain depolarization for a long enough period to initiate burst-firing [91,117,118]. This ability to generate burst-firing would be especially important for interneurons on which they are expressed, possibly at least partially contributing to their rhythmic, oscillatory nature [116]. Thus, the unique electrophysiological properties of the GluN2D subunit including resistance to Mg2+ blockade and a remarkably long channel decay latency might prove important for the integrative functions of the interneurons where this subunit is predominantly expressed. However, it should be noted that this receptor has a very low open channel probability of 1-4% and as mentioned previously lower Ca2+ permeability than the other GluN subtypes and is considered a low-conductance NMDAR which might imply that it is less effective at depolarizing the postsynaptic membrane and triggering downstream signalling events [78,119].
GluN2D-containing receptors play a role in both presynaptic and postsynaptic neurotransmission. Studies have reported GluN2D-containing NMDARs on interneurons in the hippocampus and neocortex where they play a key role in postsynaptic signalling [103,120]. Aside from enabling interneurons to synchronise and coordinate the firing of large groups of cortical pyramidal neurons, a recent study showed that tonic activation of these GluN2D-containing NMDARs on developing cortical interneurons is required for proper intrinsic excitability, dendritic arborization, GABAergic synaptogenesis, and inhibitory tone onto excitatory pyramidal cells [120]. GluN2D-containing NMDARs have also been reported on the dendrites of neurons in the subthalamic nucleus (STN), on dopaminergic projection neurons in the substantia nigra pars compacta and in the dorsal horn of the spinal cord where they contribute to the modulation of the indirect pathway, mediate dopamine release to the striatum and play a role in pain perception, respectively [121,122]. GluN2D-containing NMDARs expressed at presynaptic sites are thought to play a modulatory role in the hippocampus and cerebellum. In the hippocampus, GluN2D, along with GluN2B and postsynaptic metabotropic glutamate receptors, have been shown to be critical for the induction of spike time dependent LTD [123]. This is a form of synaptic plasticity that occurs when the timing and order of spikes from the pre- and post-synaptic neurons leads to a long-lasting weakening of the synaptic connection between them, and is important for refining synapses and neural circuits both during development and learning and memory in adults [123,124]. In the cerebellum, presynaptic GluN2D-containing NMDARs help fine tune the release of GABA onto Purkinje cells [125].
Interestingly, it has been reported that the human GRIN2D gene contains 4 estrogen responsive elements which are highly preserved in the rat, suggesting that the GluN2D subunit might be under neuroendocrine control [126]. In line with this, a study using ovariectomised rats found an upregulation of GluN2D mRNA in the hypothalamus following 17β-estradiol treatment [126]. Given the well-established links between fluctuating levels of 17β-estradiol and schizophrenia onset and symptom severity [127], it is intriguing to consider that one of the actions by which 17β-estradiol may exert it’s effects is via regulation of GluN2D and other NMDAR subunits [128].
Overall, considering its expression peaks during development, and that it is predominantly located on interneurons where its unique properties likely contribute to the integrative function of the interneurons, it is possible that the GluN2D receptor subunit plays an important role in the NMDAR hypothesis of schizophrenia.
5. Alterations to GluN2D in schizophrenia
There is evidence to suggest that the GluN2D subunit is altered in schizophrenia. A study of approximately 200 Japanese patients found single nucleotide polymorphisms (SNPs) in the gene for the GluN2D receptor that might contribute to schizophrenia susceptibility [129]. They report that specific combinations of four SNPs within the GRIN2D gene were significantly associated with schizophrenia. These specific combinations were found in three pairs of SNPs; INT10SNP–EX13SNP2, EX13SNP2–EX13SNP3 and EX6SNP–EX13SNP2 [129]. A recent mutation-screening study also identified an ultra-rare, loss of function splice-site mutation (c.1412G>A) in the exonic region of the GRIN2D gene which may lead to a truncated, nonfunctional GluN2D receptor being created, thereby contributing to schizophrenia risk [130]. This study additionally found 4 missense mutations in schizophrenia patients in the GRIN2D gene and although the actual functional impact of these amino acid substitutions weren’t examined, in silico analysis classified each of the 4 variants as disease-causing based on their predicted effect on protein function [130]. An in-situ hybridization study on post-mortem human tissue reported a 53% increase in the proportion of GluN2D mRNA expression in the PFC of people with schizophrenia [107]. This increase in the proportion of GluN2D expression was not seen in non-schizophrenia diagnosed, but anti-psychotic-treated control brains proving that the change was specific to schizophrenia and could not be attributed to treatment with anti-psychotics [107]. However, in another in-situ hybridization study, no changes in the mRNA levels of the GluN2D subunit were observed in the dlPFC of people with schizophrenia [131]. In one human post-mortem study, laser microdissection was used to isolate a population of glutamatergic relay neurons and another mixed glial and GABAergic interneuron population (including PV+ interneurons) from the medial dorsal thalamus of people with schizophrenia and controls [132]. This study revealed decreased expression of the GluN2D transcript in glutamatergic relay neurons in the medial dorsal thalamus, whilst no changes were observed in the mixed glia and interneuron population in schizophrenia [132]. These relay neurons project to the PFC, suggesting that alterations to GluN2D in schizophrenia might contribute to the disruption of the thalamocortical circuit thought to be involved in the attention and sensory processing deficits and negative symptoms seen in schizophrenia [133,134,135]. Another study of small sample size (n=19) found that GluN2D gene expression was increased in the right cerebellum of post-mortem human schizophrenia brains compared to controls [136]. However, this study included people with schizophrenia who had been taking anti-psychotic medications chronically which could have contributed to any differences observed compared to controls. These discrepant results highlight the need for further studies examining the region specific and cell specific expression and function of the GluN2D subunit in the brains of people with schizophrenia. In the following section, the expected consequence of mutations in GRIN2D and changes in GluN2D expression are discussed.
6. How might alterations to the GluN2D subunit contribute to schizophrenia?
Several characteristics of the GluN2D subunit, including peak expression early in development, it’s localization to PV+ interneurons in the cortex and hippocampus and additionally, reports of alterations to this subunit in post-mortem tissue from people with schizophrenia, and evidence of GRIN2D being a schizophrenia candidate gene, suggest that it may be involved in schizophrenia pathology. Below, we propose a mechanism by which dysfunction of the GluN2D receptor could contribute to schizophrenia.
6.1. NMDAR hypofunction on GABAergic interneurons
Although NMDAR hypofunction has been linked to schizophrenia symptoms, the precise underlying mechanisms are still unclear. One hypothesis that has been gaining interest over the past few years is that it is primarily the dysfunction of NMDARs on GABAergic interneurons, rather than more widespread NMDAR dysfunction, which contributes to the molecular, physiological and behavioural characteristics of schizophrenia [49,137,138]. GABAergic interneurons are responsible for the precisely timed inhibitory control of cortical and subcortical circuits [139]. These interneurons are stimulated by NMDAR-mediated inputs and in turn control the inhibition of the excitatory glutamatergic pyramidal network throughout the brain, including the hippocampus, which drives downstream dopaminergic neurons. As such, it has been suggested that dysfunction of these interneurons could disrupt glutamatergic and dopaminergic signalling, ultimately resulting in the clinical manifestation of schizophrenia symptoms [49,137]. This hypothesis identifies the GABAergic interneuron as the key point of convergence of the dopaminergic, glutamatergic and GABAergic neurochemical hypotheses of schizophrenia.
Despite there being more than 20 different classes of GABAergic interneurons, it is the interneurons containing the calcium binding protein, PV, that have been proposed to be especially important in schizophrenia [137]. PV-containing interneurons are unique in that they each innervate hundreds of pyramidal neurons and have a fast-spiking firing pattern which enables them to maintain tight temporal control over the output and synchrony of pyramidal neurons [140]. Not only are PV-containing interneurons crucial for regulating cortical inhibition via the pyramidal neurons they innervate, but also for the generation of synchronous gamma-frequency oscillations [49,141]. For example, relatively recently, optogenetics studies in mice were able to show that the targeted activation of fast-spiking interneurons with light, selectively amplifies gamma oscillations [139,142]. Gamma oscillations are synchronous electrophysiological brain rhythms in the gamma frequency range (30-80 Hz) that are crucial for information processing, appropriate cortical function and underpin a wide range of cognitive processes including those disrupted in schizophrenia like working memory [141,143,144]. There is mounting evidence which suggests that abnormal gamma-frequency synchrony is a major pathological characteristic of schizophrenia [141]. Specifically, people with schizophrenia exhibit a reduction in gamma oscillations (30-80 Hz) evoked by sensory stimuli and an increase in the power of ongoing, spontaneous gamma oscillations in the frontal cortex [141]. For example, a recent study found lower amplitude gamma oscillations in people with schizophrenia while they were performing a working memory task [145]. Similarly, in another cohort of people with schizophrenia, gamma band activity was reduced during the sensory processing state during an auditory task, but baseline gamma power during the resting state was increased when compared with healthy controls [146,147]. Interestingly, NMDA antagonists like ketamine and PCP also induce these same gamma oscillatory disturbances in healthy humans and rodent models of schizophrenia [148,149,150,151].
Research suggests that the fast-spiking PV interneurons, which are responsible for this control and synchronisation of cortical circuits, are especially susceptible to genetic and early environmental insults, making them a vulnerable target and a likely contributor to schizophrenia etiology [49]. Although, there is some discrepancy in the literature as to whether it is the number of PV+ cells that are reduced in schizophrenia, or if there is instead an intrinsic deficit or down-regulation of genes or proteins that are necessary for these interneurons to maintain their proper function, it is widely reported that in schizophrenia, PV interneuron-mediated inhibition of postsynaptic pyramidal neurons is reduced [152]. This could increase excitation of pyramidal neurons by disinhibition, thereby altering the excitatory/inhibitory balance in cortical and subcortical circuits and affecting their ability to synchronise firing across neural networks, ultimately resulting in the clinical manifestations of schizophrenia including cognitive dysfunction such as impaired working memory [153].
There are several lines of evidence to link GABAergic interneuron dysfunction and NMDAR hypofunction [49,57]. Firstly, several studies have found that acute administration of NMDAR antagonists results in hyperactivity of cortical pyramidal neurons and enhanced cortical glutamate release [154,155,156,157]. This is corroborated by brain imaging data in both humans and rodents showing that administration of NMDAR antagonist results in net excitation and decreased coordinated burst firing in the prefrontal cortex [155,158,159,160,161]. This cortical disinhibition suggests that NMDAR antagonists may be preferentially targeting the NMDARs on GABAergic interneurons [55]. This has been shown in a study using awake rats where administration of the NMDAR antagonist, MK-801, resulted initially in a decrease in the activity of fast-spiking interneurons, and after a delay period of several minutes, was followed by an increase in the firing rate of the majority of surrounding pyramidal neurons [55]. Other studies have also shown that hippocampal GABAergic interneurons are disproportionately more sensitive to NMDAR antagonists compared with pyramidal neurons [162]. Animal studies and in vitro studies have shown that PV-containing interneurons are in particular more sensitive to NMDAR antagonists with a selective reduction seen in PV expression in the cortex, but no changes in calretinin- or calbindin-positive interneurons following administration of ketamine [163,164]. Repeated administration of NMDAR antagonists, specifically ketamine or PCP, leads to reductions in the expression of GAD67 and PV in cortical GABAergic interneurons, providing another link between NMDAR hypofunction and GABAergic dysfunction [165,166]. In mice where the essential NR1 subunit of the NMDAR was ablated from 40 – 50% of cortical and hippocampal interneurons in early postnatal development, deficits resembling the positive, negative and cognitive phenotypes of schizophrenia including hyperlocomotion, anhedonia (reduced preference for sucrose) and deficits in social and spatial working memory began to appear in adolescence [167]. This further lends support to the theory of interneuron-based NMDAR hypofunction which causes cortical excitation by disinhibiting pyramidal neurons which output to the PFC. This may be a critical target for the treatment of not only the positive and negative symptoms, but the currently untreated cognitive deficits associated with schizophrenia.
6.2. GluN2D subunit and Parvalbumin-positive GABAergic interneurons
As previously discussed, inhibitory interneurons have been identified as the key locus or point of convergence of the glutamatergic, GABAergic and dopaminergic hypotheses of schizophrenia and are also implicated in the cognitive deficits seen in schizophrenia. Interestingly, in-situ hybridization, electrophysiology and immunohistochemistry studies have revealed that GluN2D-containing NMDARs are expressed mainly in these interneurons in the hippocampus and PFC, two regions that underlie learning and memory function [101,103,120]. A recent study of post-mortem human tissue from control (no known neuropsychiatric or neurodegenerative diseases) subjects found, using both microarray and RNAseq, that the GluN2D subunit is selectively expressed in PV+ interneurons compared to pyramidal neurons in the dlPFC [102]. In this same study, electrophysiological analyses from the mouse medial prefrontal cortex (mPFC) showed that a GluN2C/D positive allosteric modulator, CIQ(+), increased the intrinsic excitability of interneurons and enhanced EPSCS from interneurons, whilst not having any effect on the surrounding pyramidal cells [102]. Another study, found that while CIQ(+) enhanced EPSCs in WT mice, this effect was missing in GluN2D KO mice [103]. Together this provides support for the theory that PV+ interneurons are the predominant cell type expressing the GluN2D subunit in the PFC and that NMDARs containing the GluN2D subunit are critical for excitatory synaptic transmission onto hippocampal interneurons. This study also reported that the GluN2D subunit is enriched in PV-containing interneurons relative to pyramidal neurons in the dlPFC using single-cell PCR and in situ hybridization [103]. Interestingly, as mentioned previously, several studies have shown that hypofunction of NMDARs at fast-spiking PV-containing interneurons is sufficient to produce schizophrenia-like symptoms [49]. A recent study showed that tonic activation of GluN2C/GluN2D-containing receptors during development is needed for proper cortical PV interneuron maturation including dendritic arborisation, maintaining GABAergic synaptic density and inhibitory tone onto excitatory pyramidal cells [120]. The selective expression of the GluN2D subunit in PV+ interneurons suggests it may be a potential therapeutic target that could reverse interneuronal hypofunction and the cognitive impairments that result from these deficits. Positive modulation of the GluN2D subunit-containing NMDA receptor, indeed increases the firing rates and restores the GABAergic network stability, which might lessen cognitive impairment in schizophrenia. Furthermore, selective modulation of PV+ interneurons, through optogenetic stimulation, was also able to rescue cognitive impairments in Dlx5/6(+/-) mice, a mouse model of impaired PV interneuron development and function, suggesting that pharmacological potentiation of GluN2D could be a viable and useful strategy for the treatment of cognitive deficits in schizophrenia [168]. However, as the GluN2D subunit is so critical to development, it may prove a challenge to reverse any deficits.
Multiple studies have found that the GluN2D subunit forms functional NMDAR channels in the substantia nigra pars compacta dopaminergic neurons [110]. Thus, it is possible that dysfunction of the GluN2D subunit could affect burst firing in these dopaminergic neurons, disrupting their function as seen in schizophrenia. Positive modulation of the GluN2D subunit containing NMDA receptor, which is specifically expressed in GABA-ergic interneurons, increases the firing rates and restores the GABA-ergic network stability, which is thought to lessen cognitive impairment in schizophrenia.
7. Consequences of loss of GluN2D function
Strategies to explore the influence of GluN2D function include using animal models and pharmacological tools to study behaviour and physiological measures which have relevance to schizophrenia.
7.1. Genetic models
GluN2D-knockout (KO) mice are viable, reproduce and grow normally, and have no overt changes in neuronal histology [169]. Moreover, mRNA levels of the other NMDAR subunits are unaffected in these mice [169]. These mice, however, exhibit unique behavioral phenotypes including diminished spontaneous motor movements in open-field tests [169,170]. GluN2D-KO mice also display deficits in spatial learning and memory, as well as impaired contextual fear memory, but show no deficits in the novel object recognition task [116,171,172]. Most studies report no abnormalities in motor function as measured by the rotarod test, nor any differences in anxiety when compared with WT mice during the light-dark compartment test and elevated maze test [169,173]. However, there is at least one report of reduced sensitivity to stress induced by the elevated-plus maze, the light-dark compartment test and forced swim tests in these KO mice [170]. This study also reported a downregulation of dopamine, serotonin, norepinephrine and their metabolites in the hippocampus and striatum of GluN2D-KO mice [170]. GluN2D-KO mice are reported to have a robust prepulse inhibition response, which is known to be deficient in schizophrenia [116,174]. This suggests that they might not model all aspects of schizophrenia. Interestingly, the hyperlocomotion- or psychosis-inducing effects of PCP and ketamine are reduced in GluN2D-KO mice indicating that the GluN2D subunit plays an important role in mediating the effects of these drugs [116,173,175]. Additionally, the motor impairments, locomotor sensitization and increase in extracellular dopamine seen following PCP or ketamine administration in WT mice is not seen in GluN2D-KO mice [173,175,176]. Autoradiography studies revealed that the increase in neuronal activity induced by sub-anesthetic ketamine in the medial prefrontal cortex, entorhinal cortex, presubiculum and caudate putamen in WT mice is not seen in GluN2D-KO mice [116,160,177]. These data suggest that the GluN2D subunit mediates the psychosis-inducing effects of the NMDAR antagonists, PCP and ketamine.
While administration of ketamine increased gamma frequency (30-140 Hz) oscillatory power in WT mice, it elicited a much less pronounced increase in GluN2D-KO mice, especially at the high frequency (>60Hz) range, suggesting a role for the GluN2D subunit in modulating high frequency neural network oscillations induced by ketamine [116]. Sapkota et al. also found reduced PV cell density in the substantia nigra and basolateral/lateral amygdala and a trend towards a reduction in the mPFC and hippocampus in GluN2D-KO mice compared with WT mice [116].
These studies suggest the GluN2D subunit expressing NMDA receptors are critical in mediating many of the effects of NMDAR antagonists like ketamine on behavior and electrophysiology that is relevant to schizophrenia, and thus may in turn suggest a role for the GluN2D subunit in the development of schizophrenia. There is evidence to suggest that GluN2D-containing NMDARs can contribute to neuronal networks that underlie cognition and which are found to be disrupted in schizophrenia. Targeting GluN2D-mediated signaling may prove to have therapeutic benefit in specific behavioural domains within schizophrenia.
7.2. Pharmacological manipulations
Although currently, to the best of our knowledge, there are no GluN2D-selective drugs available, competitive antagonists with 3-10 fold higher selectivity for GluN2C/GluN2D vs GluN2A/GluN2B-containing NMDARs have been developed. One such compound, (2R*,3S*)-1-(phenanthrenyl-2-carbonyl)piperazine-2,3-dicarboxylic acid (PPDA), resulted in more potent inhibition of LTD than LTP in rat hippocampal slices suggesting a role for the GluN2D (and GluN2C) NMDAR subunits in hippocampal LTD [178]. Another study using PPDA showed that GluN2D-containing NMDARs also contribute to extrasynaptic currents in rat CA1 neurons [179]. A recent study used an analogue of PPDA, (2R*,3S*)-1-(9-bromophenanthrene-3-carbonyl)piperazine-2,3-dicarboxylic acid (UBP145), in combination with GluN2D-KO mice to show that GluN2D-containing NMDARs play a role in short term potentiation (STP) as well as LTP in the mouse hippocampus [180]. This study reported that UBP145 partially inhibited LTP and the slow component of STP in WT but not GluN2D-KO mice [180]. Whilst the function of STP is lesser known than LTP, it has been hypothesized that it might play a role in forms of short term memory, such as working memory which is known to be disrupted in schizophrenia [181]. These data together suggest that GluN2D-containing NMDARs contribute to synaptic plasticity and thus cognitive processes in complex ways. Therefore, it’s possible that dysregulation of these receptors is involved in the cognitive dysfunction seen in neurological disorders including schizophrenia.
8. Conclusions
Precipitating factors, including any combination of genetic predisposition and environmental factors like maternal infection or obstetric complications, lead to NMDAR hypofunction disproportionately at fast-spiking PV-containing interneurons during development. This results in pathological phenotypes including impaired oscillatory activity and neuronal synchrony, cortical disinhibition and dopaminergic dysfunction ultimately giving rise to the various symptoms of schizophrenia. Disruption of the GluN2D subunit and alteration to GluN2D neurotransmission could be a molecular pathway contributing to the symptomatology of schizophrenia. This is of importance as it may provide new insights into the aetiology of this disorder and might even lead to the development of novel drugs for the treatment of specific schizophrenia symptoms, including cognitive dysfunction.
Author Contributions
Conceptualization, C.V., R.H., N.J. and S.S.; methodology, C.V.; software, C.V..; investigation, C.V.; resources, R.H., S.S.; writing—original draft preparation, C.V.; writing—review and editing, C.V., R.H., N.J., M.H., S.S..; supervision, R.H., N.J., S.S.; All authors have read and agreed to the published version of the manuscript.
Funding
C.V. is funded by a Monash University allocated research training program (RTP) scholarship provided by the Australian Government. R.H. is supported by a National Health and Medical Research Council ideas grant GNT2000893.
Data Availability Statement
No new data was created for this review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Saha, S.; Chant, D.; Welham, J.; McGrath, J. , A systematic review of the prevalence of schizophrenia. PLoS Med 2005, 2, e141. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, R.A.; Reis Marques, T.; Howes, O.D. , Schizophrenia-An Overview. JAMA Psychiatry 2020, 77, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Kahn, R.; Sommer, I.; Murray, R.; Meyer-Lindenberg, A.; Weinberger, D.; Cannon, T.; O’Donovan, M.; Correl, C.; Kane, J.; van Os, J. , Schizophrenia. Nature Reviews Disease Primers. Nov 2015, 12, 15067. [Google Scholar]
- Keefe, R.S.; Eesley, C.E.; Poe, M.P. , Defining a cognitive function decrement in schizophrenia. Biological psychiatry 2005, 57, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.G.; Voruganti, L.N. , The burden of schizophrenia on caregivers: a review. Pharmacoeconomics 2008, 26, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, M.; Aigbogun, M.S.; Guerin, A.; Nitulescu, R.; Ramanakumar, A.V.; Kamat, S.A.; DeLucia, M.; Duffy, R.; Legacy, S.N.; Henderson, C.; Francois, C.; Wu, E. , The Economic Burden of Schizophrenia in the United States in 2013. J Clin Psychiatry 2016, 77, 764–71. [Google Scholar] [CrossRef] [PubMed]
- Neil, A.L.; Carr, V.J.; Mihalopoulos, C.; Mackinnon, A.; Morgan, V.A. , Costs of psychosis in 2010: Findings from the second Australian National Survey of Psychosis. Australian & New Zealand Journal of Psychiatry 2014, 48, 169–182. [Google Scholar]
- Ripke, S.; Neale, B.M.; Corvin, A.; Walters, J.T.; Farh, K.-H.; Holmans, P.A.; Lee, P.; Bulik-Sullivan, B.; Collier, D.A.; Huang, H. , Biological insights from 108 schizophrenia-associated genetic loci. Nature 2014, 511, 421–427. [Google Scholar]
- Gottesman, I.I.; Shields, J. , A polygenic theory of schizophrenia. International Journal of Mental Health 1972, 1, 107–115. [Google Scholar] [CrossRef]
- Trubetskoy, V.; Pardiñas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.-Y.; Dennison, C.A.; Hall, L.S. , Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef]
- Walsh, T.; McClellan, J.M.; McCarthy, S.E.; Addington, A.M.; Pierce, S.B.; Cooper, G.M.; Nord, A.S.; Kusenda, M.; Malhotra, D.; Bhandari, A.; Stray, S.M.; Rippey, C.F.; Roccanova, P.; Makarov, V.; Lakshmi, B.; Findling, R.L.; Sikich, L.; Stromberg, T.; Merriman, B.; Gogtay, N.; Butler, P.; Eckstrand, K.; Noory, L.; Gochman, P.; Long, R.; Chen, Z.; Davis, S.; Baker, C.; Eichler, E.E.; Meltzer, P.S.; Nelson, S.F.; Singleton, A.B.; Lee, M.K.; Rapoport, J.L.; King, M.C.; Sebat, J. , Rare structural variants disrupt multiple genes in neurodevelopmental pathways in schizophrenia. Science 2008, 320, 539–43. [Google Scholar] [CrossRef] [PubMed]
- Fromer, M.; Roussos, P.; Sieberts, S.K.; Johnson, J.S.; Kavanagh, D.H.; Perumal, T.M.; Ruderfer, D.M.; Oh, E.C.; Topol, A.; Shah, H.R.; Klei, L.L.; Kramer, R.; Pinto, D.; Gumus, Z.H.; Cicek, A.E.; Dang, K.K.; Browne, A.; Lu, C.; Xie, L.; Readhead, B.; Stahl, E.A.; Xiao, J.; Parvizi, M.; Hamamsy, T.; Fullard, J.F.; Wang, Y.C.; Mahajan, M.C.; Derry, J.M.; Dudley, J.T.; Hemby, S.E.; Logsdon, B.A.; Talbot, K.; Raj, T.; Bennett, D.A.; De Jager, P.L.; Zhu, J.; Zhang, B.; Sullivan, P.F.; Chess, A.; Purcell, S.M.; Shinobu, L.A.; Mangravite, L.M.; Toyoshiba, H.; Gur, R.E.; Hahn, C.G.; Lewis, D.A.; Haroutunian, V.; Peters, M.A.; Lipska, B.K.; Buxbaum, J.D.; Schadt, E.E.; Hirai, K.; Roeder, K.; Brennand, K.J.; Katsanis, N.; Domenici, E.; Devlin, B.; Sklar, P. , Gene expression elucidates functional impact of polygenic risk for schizophrenia. Nat Neurosci 2016, 19, 1442–1453. [Google Scholar] [CrossRef] [PubMed]
- Ursini, G.; Punzi, G.; Chen, Q.; Marenco, S.; Robinson, J.F.; Porcelli, A.; Hamilton, E.G.; Mitjans, M.; Maddalena, G.; Begemann, M.; Seidel, J.; Yanamori, H.; Jaffe, A.E.; Berman, K.F.; Egan, M.F.; Straub, R.E.; Colantuoni, C.; Blasi, G.; Hashimoto, R.; Rujescu, D.; Ehrenreich, H.; Bertolino, A.; Weinberger, D.R. , Convergence of placenta biology and genetic risk for schizophrenia. Nat Med 2018, 24, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Malaspina, D.; Harlap, S.; Fennig, S.; Heiman, D.; Nahon, D.; Feldman, D.; Susser, E.S. , Advancing paternal age and the risk of schizophrenia. Arch Gen Psychiatry 2001, 58, 361–7. [Google Scholar] [CrossRef] [PubMed]
- Fountoulakis, K.N.; Gonda, X.; Siamouli, M.; Panagiotidis, P.; Moutou, K.; Nimatoudis, I.; Kasper, S. , Paternal and maternal age as risk factors for schizophrenia: a case–control study. International journal of psychiatry in clinical practice 2018, 22, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Radua, J.; Ramella-Cravaro, V.; Ioannidis, J.P.; Reichenberg, A.; Phiphopthatsanee, N.; Amir, T.; Yenn Thoo, H.; Oliver, D.; Davies, C.; Morgan, C. , What causes psychosis? An umbrella review of risk and protective factors. World Psychiatry 2018, 17, 49–66. [Google Scholar] [CrossRef] [PubMed]
- Stilo, S.A.; Murray, R.M. , Non-Genetic Factors in Schizophrenia. Curr Psychiatry Rep 2019, 21, 100. [Google Scholar] [CrossRef] [PubMed]
- Cantor-Graae, E.; Selten, J.-P. , Schizophrenia and migration: a meta-analysis and review. American Journal of Psychiatry 2005, 162, 12–24. [Google Scholar] [CrossRef]
- Varese, F.; Smeets, F.; Drukker, M.; Lieverse, R.; Lataster, T.; Viechtbauer, W.; Read, J.; Van Os, J.; Bentall, R.P. , Childhood adversities increase the risk of psychosis: a meta-analysis of patient-control, prospective-and cross-sectional cohort studies. Schizophrenia bulletin 2012, 38, 661–671. [Google Scholar] [CrossRef]
- Kane, J.M.; Correll, C.U. , Pharmacologic treatment of schizophrenia. Dialogues in clinical neuroscience 2022. [Google Scholar]
- Leucht, S.; Corves, C.; Arbter, D.; Engel, R.R.; Li, C.; Davis, J.M. , Second-generation versus first-generation antipsychotic drugs for schizophrenia: a meta-analysis. Lancet 2009, 373, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Stępnicki, P.; Kondej, M.; Kaczor, A.A. , Current concepts and treatments of schizophrenia. Molecules 2018, 23, 2087. [Google Scholar] [CrossRef]
- Keefe, R.S.; Bilder, R.M.; Davis, S.M.; Harvey, P.D.; Palmer, B.W.; Gold, J.M.; Meltzer, H.Y.; Green, M.F.; Capuano, G.; Stroup, T.S. , Neurocognitive effects of antipsychotic medications in patients with chronic schizophrenia in the CATIE Trial. Archives of general psychiatry 2007, 64, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Hietala, J.; Syvälahti, E. , Dopamine in schizophrenia. Annals of medicine 1996, 28, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Gründer, G.; Cumming, P. , The dopamine hypothesis of schizophrenia: Current status. In The neurobiology of schizophrenia, Elsevier: 2016; pp 109-124.
- Davis, K.L.; Kahn, R.S.; Ko, G.; Davidson, M. , Dopamine in schizophrenia: a review and reconceptualization. The American journal of psychiatry 1991. [Google Scholar]
- Seeman, P.; Lee, T.; Chau-Wong, M.; Wong, K. , Antipsychotic drug doses and neuroleptic/dopamine receptors. Nature 1976, 261, 717–719. [Google Scholar] [CrossRef] [PubMed]
- Van Rossum, J. , The significance of dopamine-receptor blockade for the mechanism of action of neuroleptic drugs. Arch. Int Pharmacodyn Ther 1996, 160, 492–494. [Google Scholar]
- Seeman, M.V. , History of the dopamine hypothesis of antipsychotic action. World Journal of Psychiatry 2021, 11, 355. [Google Scholar] [CrossRef]
- Seeman, P. , Dopamine receptors and the dopamine hypothesis of schizophrenia. Synapse 1987, 1, 133–152. [Google Scholar] [CrossRef]
- Snyder, S.H. , Dopamine receptors, neuroleptics, and schizophrenia. The American journal of psychiatry 1981. [Google Scholar]
- Lisman, J.E.; Coyle, J.T.; Green, R.W.; Javitt, D.C.; Benes, F.M.; Heckers, S.; Grace, A.A. , Circuit-based framework for understanding neurotransmitter and risk gene interactions in schizophrenia. Trends in neurosciences 2008, 31, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Kantrowitz, J.T.; Javitt, D.C. , N-methyl-d-aspartate (NMDA) receptor dysfunction or dysregulation: the final common pathway on the road to schizophrenia? Brain Res Bull 2010, 83, 108–21. [Google Scholar] [CrossRef] [PubMed]
- Coyle, J.T.; Tsai, G.; Goff, D. , Converging evidence of NMDA receptor hypofunction in the pathophysiology of schizophrenia. Annals of the New York Academy of Sciences 2003, 1003, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Krystal, J.H.; Karper, L.P.; Seibyl, J.P.; Freeman, G.K.; Delaney, R.; Bremner, J.D.; Heninger, G.R.; Bowers, M.B., Jr.; Charney, D.S. , Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Psychotomimetic, perceptual, cognitive, and neuroendocrine responses. Arch Gen Psychiatry 1994, 51, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Javitt, D. , Negative schizophrenic symptomatology and the PCP (phencyclidine) model of schizophrenia. The Hillside journal of clinical psychiatry 1987, 9, 12–35. [Google Scholar] [PubMed]
- Luby, E.D.; Cohen, B.D.; Rosenbaum, G.; Gottlieb, J.S.; Kelley, R. , Study of a new schizophrenomimetic drug—Sernyl. AMA Archives of Neurology & Psychiatry 1959, 81, 363–369. [Google Scholar]
- Luby, E.D.; Gottlieb, J.S.; Cohen, B.D.; Rosenbaum, G.; Domino, E.F. , Model psychoses and schizophrenia. American Journal of Psychiatry 1962, 119, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, G.; Cohen, B.D.; Luby, E.D.; Gottlieb, J.S.; Yelen, D. , Comparison of sernyl with other drugs: simulation of schizophrenic performance with sernyl, LSD-25, and amobarbital (amytal) sodium; I. Attention, motor function, and proprioception. AMA archives of general psychiatry 1959, 1, 651–656. [Google Scholar] [CrossRef]
- Cohen, B.D.; Luby, E.D.; Rosenbaum, G.; Gottlieb, J.S. , Combined sernyl and sensory deprivation. Comprehensive Psychiatry 1960, 1, 345–348. [Google Scholar] [CrossRef]
- Itil, T.; Keskiner, A.; Kiremitci, N.; Holden, J. , Effect of phencyclidine in chronic schizophrenics. Canadian Psychiatric Association Journal 1967, 12, 209–212. [Google Scholar] [CrossRef]
- Lahti, A.C.; Koffel, B.; LaPorte, D.; Tamminga, C.A. , Subanesthetic doses of ketamine stimulate psychosis in schizophrenia. Neuropsychopharmacology 1995, 13, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Anis, N.A.; Berry, S.C.; Burton, N.R.; Lodge, D. , The dissociative anaesthetics, ketamine and phencyclidine, selectively reduce excitation of central mammalian neurones by N-methyl-aspartate. Br J Pharmacol 1983, 79, 565–75. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kornhuber, H.; Schmid-Burgk, W.; Holzmüller, B. , Low cerebrospinal fluid glutamate in schizophrenic patients and a new hypothesis on schizophrenia. Neuroscience letters 1980, 20, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Javitt, D.C.; Zukin, S.R. , Recent advances in the phencyclidine model of schizophrenia. The American journal of psychiatry 1991. [Google Scholar]
- Javitt, D.C. , Glutamatergic theories of schizophrenia. Isr J Psychiatry Relat Sci 2010, 47, 4–16. [Google Scholar] [PubMed]
- Kantrowitz, J.; Javitt, D.C. , Glutamatergic transmission in schizophrenia: from basic research to clinical practice. Curr Opin Psychiatry 2012, 25, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S.H. , The dopamine hypothesis of schizophrenia: focus on the dopamine receptor. The American journal of psychiatry 1976. [Google Scholar]
- Nakazawa, K.; Zsiros, V.; Jiang, Z.; Nakao, K.; Kolata, S.; Zhang, S.; Belforte, J.E. , GABAergic interneuron origin of schizophrenia pathophysiology. Neuropharmacology 2012, 62, 1574–83. [Google Scholar] [CrossRef]
- Javitt, D.C.; Hashim, A.; Sershen, H. , Modulation of striatal dopamine release by glycine transport inhibitors. Neuropsychopharmacology 2005, 30, 649–656. [Google Scholar] [CrossRef]
- Nair, V.D.; Savelli, J.E.; Mishra, R.K. , Modulation of dopamine D 2 receptor expression by an NMDA receptor antagonist in rat brain. Journal of Molecular Neuroscience 1998, 11, 121–126. [Google Scholar] [CrossRef]
- Micheletti, G.; Lannes, B.; Haby, C.; Borrelli, E.; Kempf, E.; Warter, J.-M.; Zwiller, J. , Chronic administration of NMDA antagonists induces D2 receptor synthesis in rat striatum. Molecular brain research 1992, 14, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Kegeles, L.S.; Abi-Dargham, A.; Zea-Ponce, Y.; Rodenhiser-Hill, J.; Mann, J.J.; Van Heertum, R.L.; Cooper, T.B.; Carlsson, A.; Laruelle, M. , Modulation of amphetamine-induced striatal dopamine release by ketamine in humans: implications for schizophrenia. Biological psychiatry 2000, 48, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.S.; Schloesser, R.; Brodie, J.D.; Dewey, S.L.; Logan, J.; Vitkun, S.A.; Simkowitz, P.; Hurley, A.; Cooper, T.; Volkow, N.D. , Glutamate modulation of dopamine measured in vivo with positron emission tomography (PET) and 11C-raclopride in normal human subjects. Neuropsychopharmacology 1998, 18, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Homayoun, H.; Moghaddam, B. , NMDA receptor hypofunction produces opposite effects on prefrontal cortex interneurons and pyramidal neurons. Journal of Neuroscience 2007, 27, 11496–11500. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Clark, S.; Lewis, D.V.; Wilson, W.A. , NMDA receptor antagonists disinhibit rat posterior cingulate and retrosplenial cortices: a potential mechanism of neurotoxicity. Journal of Neuroscience 2002, 22, 3070–3080. [Google Scholar] [CrossRef] [PubMed]
- Olney, J.W.; Farber, N.B. , Glutamate receptor dysfunction and schizophrenia. Archives of general psychiatry 1995, 52, 998–1007. [Google Scholar] [CrossRef] [PubMed]
- Olney, J.; Labruyere, J.; Wang, G.; Wozniak, D.; Price, M.; Sesma, M. , NMDA antagonist neurotoxicity: mechanism and prevention. Science 1991, 254, 1515–1518. [Google Scholar] [CrossRef]
- Sun, J.; Jia, P.; Fanous, A.H.; Van Den Oord, E.; Chen, X.; Riley, B.P.; Amdur, R.L.; Kendler, K.S.; Zhao, Z. , Schizophrenia gene networks and pathways and their applications for novel candidate gene selection. PloS one 2010, 5, e11351. [Google Scholar] [CrossRef]
- Greenwood, T.A.; Light, G.A.; Swerdlow, N.R.; Radant, A.D.; Braff, D.L. , Association analysis of 94 candidate genes and schizophrenia-related endophenotypes. PLoS One 2012, 7, e29630. [Google Scholar] [CrossRef]
- Balu, D.T.; Coyle, J.T. , Neuroplasticity signaling pathways linked to the pathophysiology of schizophrenia. Neurosci Biobehav Rev 2011, 35, 848–70. [Google Scholar] [CrossRef]
- Balu, D.T. , The NMDA receptor and schizophrenia: from pathophysiology to treatment. In Advances in Pharmacology, Elsevier: 2016; Vol. 76, pp 351-382.
- Hashimoto, K.; Fukushima, T.; Shimizu, E.; Komatsu, N.; Watanabe, H.; Shinoda, N.; Nakazato, M.; Kumakiri, C.; Okada, S.; Hasegawa, H.; Imai, K.; Iyo, M. , Decreased serum levels of D-serine in patients with schizophrenia: evidence in support of the N-methyl-D-aspartate receptor hypofunction hypothesis of schizophrenia. Arch Gen Psychiatry 2003, 60, 572–6. [Google Scholar] [CrossRef] [PubMed]
- Weickert, C.S.; Fung, S.J.; Catts, V.S.; Schofield, P.R.; Allen, K.M.; Moore, L.T.; Newell, K.A.; Pellen, D.; Huang, X.F.; Catts, S.V.; Weickert, T.W. , Molecular evidence of N-methyl-D-aspartate receptor hypofunction in schizophrenia. Mol Psychiatry 2013, 18, 1185–92. [Google Scholar] [CrossRef] [PubMed]
- Labrie, V.; Lipina, T.; Roder, J.C. , Mice with reduced NMDA receptor glycine affinity model some of the negative and cognitive symptoms of schizophrenia. Psychopharmacology 2008, 200, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Mohn, A.R.; Gainetdinov, R.R.; Caron, M.G.; Koller, B.H. , Mice with reduced NMDA receptor expression display behaviors related to schizophrenia. Cell 1999, 98, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Halene, T.B.; Ehrlichman, R.S.; Liang, Y.; Christian, E.P.; Jonak, G.J.; Gur, T.L.; Blendy, J.A.; Dow, H.C.; Brodkin, E.S.; Schneider, F. , Assessment of NMDA receptor NR1 subunit hypofunction in mice as a model for schizophrenia. Genes, Brain and Behavior 2009, 8, 661–675. [Google Scholar] [CrossRef] [PubMed]
- Hakami, T.; Jones, N.C.; Tolmacheva, E.A.; Gaudias, J.; Chaumont, J.; Salzberg, M.; O'Brien, T.J.; Pinault, D. , NMDA receptor hypofunction leads to generalized and persistent aberrant γ oscillations independent of hyperlocomotion and the state of consciousness. PloS one 2009, 4, e6755. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.M.; Pinault, D.; O'Brien, T.J.; Jones, N.C. , Chronic administration of antipsychotics attenuates ongoing and ketamine-induced increases in cortical γ oscillations. International Journal of Neuropsychopharmacology 2014, 17, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Hudson, M.; Rind, G.; O'brien, T.; Jones, N. , Reversal of evoked gamma oscillation deficits is predictive of antipsychotic activity with a unique profile for clozapine. Translational psychiatry 2016, 6, e784–e784. [Google Scholar] [CrossRef]
- Zhou, Y.; Danbolt, N.C. , Glutamate as a neurotransmitter in the healthy brain. Journal of neural transmission 2014, 121, 799–817. [Google Scholar] [CrossRef]
- Hollmann, M.; Heinemann, S. , Cloned glutamate receptors. Annual review of neuroscience 1994, 17, 31–108. [Google Scholar] [CrossRef]
- Nakanishi, S.; Masu, M. , Molecular diversity and functions of glutamate receptors. Annual review of biophysics and biomolecular structure 1994, 23, 319–348. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, K.; Tani, H.; Nakajima, S.; Nagai, N.; Koizumi, T.; Miyazaki, T.; Mimura, M.; Takahashi, T.; Uchida, H. , AMPA receptors in schizophrenia: A systematic review of postmortem studies on receptor subunit expression and binding. Schizophrenia Research 2022, 243, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Rubio, M.D.; Drummond, J.B.; Meador-Woodruff, J.H. , Glutamate receptor abnormalities in schizophrenia: implications for innovative treatments. Biomolecules & therapeutics 2012, 20, 1. [Google Scholar]
- Maksymetz, J.; Moran, S.P.; Conn, P.J. , Targeting metabotropic glutamate receptors for novel treatments of schizophrenia. Molecular brain 2017, 10, 1–19. [Google Scholar]
- Scatton, B. , The NMDA receptor complex. Fundamental & clinical pharmacology 1993, 7, 389–400. [Google Scholar]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. , Glutamate receptor ion channels: structure, regulation, and function. Pharmacol Rev 2010, 62, 405–96. [Google Scholar] [CrossRef]
- Monyer, H.; Sprengel, R.; Schoepfer, R.; Herb, A.; Higuchi, M.; Lomeli, H.; Burnashev, N.; Sakmann, B.; Seeburg, P.H. , Heteromeric NMDA receptors: molecular and functional distinction of subtypes. Science 1992, 256, 1217–1221. [Google Scholar] [CrossRef]
- Karakas, E.; Furukawa, H. , Crystal structure of a heterotetrameric NMDA receptor ion channel. Science 2014, 344, 992–997. [Google Scholar] [CrossRef]
- Furukawa, H.; Singh, S.K.; Mancusso, R.; Gouaux, E. , Subunit arrangement and function in NMDA receptors. Nature 2005, 438, 185–192. [Google Scholar] [CrossRef]
- Lerma, J.; Zukin, R.S.; Bennett, M. , Glycine decreases desensitization of N-methyl-D-aspartate (NMDA) receptors expressed in Xenopus oocytes and is required for NMDA responses. Proceedings of the National Academy of Sciences 1990, 87, 2354–2358. [Google Scholar] [CrossRef]
- Greer, P.L.; Greenberg, M.E. , From synapse to nucleus: calcium-dependent gene transcription in the control of synapse development and function. Neuron 2008, 59, 846–860. [Google Scholar] [CrossRef] [PubMed]
- Artola, A.; Bröcher, S.; Singer, W. , Different voltage-dependent thresholds for inducing long-term depression and long-term potentiation in slices of rat visual cortex. Nature 1990, 347, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, E. , Molecular mechanism of neuronal plasticity: induction and maintenance of long-term potentiation in the hippocampus. Journal of pharmacological sciences 2006, 100, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, H.; Moriyoshi, K.; Ishii, T.; Masu, M.; Nakanishi, S. , Structures and properties of seven isoforms of the NMDA receptor generated by alternative splicing. Biochemical and biophysical research communications 1992, 185, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, P.; Bellone, C.; Zhou, Q. , NMDA receptor subunit diversity: impact on receptor properties, synaptic plasticity and disease. Nature Reviews Neuroscience 2013, 14, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Schorge, S.; Colquhoun, D. , Studies of NMDA receptor function and stoichiometry with truncated and tandem subunits. Journal of Neuroscience 2003, 23, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Henson, M.A.; Roberts, A.C.; Salimi, K.; Vadlamudi, S.; Hamer, R.M.; Gilmore, J.H.; Jarskog, L.F.; Philpot, B.D. , Developmental regulation of the NMDA receptor subunits, NR3A and NR1, in human prefrontal cortex. Cerebral Cortex 2008, 18, 2560–2573. [Google Scholar] [CrossRef]
- Tolle, T.; Berthele, A.; Zieglgansberger, W.; Seeburg, P.H.; Wisden, W. , The differential expression of 16 NMDA and non-NMDA receptor subunits in the rat spinal cord and in periaqueductal gray. Journal of Neuroscience 1993, 13, 5009–5028. [Google Scholar] [CrossRef]
- Monyer, H.; Burnashev, N.; Laurie, D.J.; Sakmann, B.; Seeburg, P.H. , Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron 1994, 12, 529–40. [Google Scholar] [CrossRef]
- Vicini, S.; Wang, J.F.; Li, J.H.; Zhu, W.J.; Wang, Y.H.; Luo, J.H.; Wolfe, B.B.; Grayson, D.R. , Functional and pharmacological differences between recombinant N-methyl-D-aspartate receptors. Journal of neurophysiology 1998, 79, 555–566. [Google Scholar] [CrossRef]
- Huggins, D.J.; Grant, G.H. , The function of the amino terminal domain in NMDA receptor modulation. Journal of Molecular Graphics and Modelling 2005, 23, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Moriyoshi, K.; Sugihara, H.; Sakurada, K.; Kadotani, H.; Yokoi, M.; Akazawa, C.; Shigemoto, R.; Mizuno, N.; Masu, M. , Molecular characterization of the family of the N-methyl-D-aspartate receptor subunits. Journal of Biological Chemistry 1993, 268, 2836–2843. [Google Scholar] [CrossRef] [PubMed]
- Akazawa, C.; Shigemoto, R.; Bessho, Y.; Nakanishi, S.; Mizuno, N. , Differential expression of five N-methyl-D-aspartate receptor subunit mRNAs in the cerebellum of developing and adult rats. J Comp Neurol 1994, 347, 150–60. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, A.; Villa, M.; Mohler, H.; Benke, D. , Developmental and regional expression of NMDA receptor subtypes containing the NR2D subunit in rat brain. Journal of neurochemistry 1996, 66, 1240–1248. [Google Scholar] [CrossRef]
- Dunah, A.W.; Yasuda, R.P.; Wang, Y. h.; Luo, J.; Dávila-García, M.I.; Gbadegesin, M.; Vicini, S.; Wolfe, B.B. , Regional and ontogenic expression of the NMDA receptor subunit NR2D protein in rat brain using a subunit-specific antibody. Journal of neurochemistry 1996, 67, 2335–2345. [Google Scholar] [CrossRef]
- Standaert, D.G.; Landwehrmeyer, G.B.; Kerner, J.A.; Penney Jr, J.B.; Young, A.B. , Expression of NMDAR2D glutamate receptor subunit mRNA in neurochemically identified interneurons in the rat neostriatum, neocortex and hippocampus. Molecular brain research 1996, 42, 89–102. [Google Scholar] [CrossRef]
- Cannon, M.; Murray, R.M. , Neonatal origins of schizophrenia. Archives of disease in childhood 1998, 78, 1–3. [Google Scholar] [CrossRef]
- Birnbaum, R.; Weinberger, D.R. , Genetic insights into the neurodevelopmental origins of schizophrenia. Nature Reviews Neuroscience 2017, 18, 727–740. [Google Scholar] [CrossRef]
- Engelhardt, J. v.; Bocklisch, C.; Tönges, L.; Herb, A.; Mishina, M.; Monyer, H. , GluN2D-containing NMDA receptors-mediate synaptic currents in hippocampal interneurons and pyramidal cells in juvenile mice. Frontiers in cellular neuroscience 2015, 9, 95. [Google Scholar] [CrossRef]
- Garst-Orozco, J.; Malik, R.; Lanz, T.A.; Weber, M.L.; Xi, H.; Arion, D.; Enwright III, J.F.; Lewis, D.A.; O’Donnell, P.; Sohal, V.S. , GluN2D-mediated excitatory drive onto medial prefrontal cortical PV+ fast-spiking inhibitory interneurons. Plos one 2020, 15, e0233895. [Google Scholar] [CrossRef]
- Perszyk, R.E.; DiRaddo, J.O.; Strong, K.L.; Low, C.M.; Ogden, K.K.; Khatri, A.; Vargish, G.A.; Pelkey, K.A.; Tricoire, L.; Liotta, D.C.; Smith, Y.; McBain, C.J.; Traynelis, S.F. , GluN2D-Containing N-methyl-d-Aspartate Receptors Mediate Synaptic Transmission in Hippocampal Interneurons and Regulate Interneuron Activity. Mol Pharmacol 2016, 90, 689–702. [Google Scholar] [CrossRef]
- Camp, C.R.; Yuan, H. , GRIN2D/GluN2D NMDA receptor: Unique features and its contribution to pediatric developmental and epileptic encephalopathy. Eur J Paediatr Neurol 2020, 24, 89–99. [Google Scholar] [CrossRef]
- Ritter, L.M.; Unis, A.S.; Meador-Woodruff, J.H. , Ontogeny of ionotropic glutamate receptor expression in human fetal brain. Developmental Brain Research 2001, 127, 123–133. [Google Scholar] [CrossRef]
- Scherzer, C.R.; Landwehrmeyer, G.B.; Kerner, J.A.; Counihan, T.J.; Kosinski, C.M.; Standaert, D.G.; Daggett, L.P.; Velicelebi, G.; Penney, J.B.; Young, A.B. , Expression of N-methyl-D-aspartate receptor subunit mRNAs in the human brain: hippocampus and cortex. J Comp Neurol 1998, 390, 75–90. [Google Scholar] [CrossRef]
- Akbarian, S.; Sucher, N.J.; Bradley, D.; Tafazzoli, A.; Trinh, D.; Hetrick, W.P.; Potkin, S.G.; Sandman, C.A.; Bunney, W.E., Jr.; Jones, E.G. , Selective alterations in gene expression for NMDA receptor subunits in prefrontal cortex of schizophrenics. J Neurosci 1996, 16, 19–30. [Google Scholar] [CrossRef]
- Dunah, A.W.; Luo, J.; Wang, Y.-H.; Yasuda, R.P.; Wolfe, B.B. , Subunit Composition ofN-Methyl-D-aspartate Receptors in the Central Nervous System that Contain the NR2D Subunit. Molecular Pharmacology 1998, 53, 429–437. [Google Scholar] [CrossRef]
- Brickley, S.G.; Misra, C.; Mok, M.S.; Mishina, M.; Cull-Candy, S.G. , NR2B and NR2D subunits coassemble in cerebellar Golgi cells to form a distinct NMDA receptor subtype restricted to extrasynaptic sites. Journal of Neuroscience 2003, 23, 4958–4966. [Google Scholar] [CrossRef]
- Jones, S.; Gibb, A.J. , Functional NR2B-and NR2D-containing NMDA receptor channels in rat substantia nigra dopaminergic neurones. The Journal of physiology 2005, 569, 209–221. [Google Scholar] [CrossRef]
- Erreger, K.; Geballe, M.T.; Kristensen, A.; Chen, P.E.; Hansen, K.B.; Lee, C.J.; Yuan, H.; Le, P.; Lyuboslavsky, P.N.; Micale, N. , Subunit-specific agonist activity at NR2A-, NR2B-, NR2C-, and NR2D-containing N-methyl-D-aspartate glutamate receptors. Molecular pharmacology 2007, 72, 907–920. [Google Scholar] [CrossRef]
- Chen, P.E.; Geballe, M.T.; Katz, E.; Erreger, K.; Livesey, M.R.; O'toole, K.K.; Le, P.; Lee, C.J.; Snyder, J.P.; Traynelis, S.F. , Modulation of glycine potency in rat recombinant NMDA receptors containing chimeric NR2A/2D subunits expressed in Xenopus laevis oocytes. The Journal of physiology 2008, 586, 227–245. [Google Scholar] [CrossRef]
- Kuner, T.; Schoepfer, R. , Multiple structural elements determine subunit specificity of Mg2+ block in NMDA receptor channels. Journal of Neuroscience 1996, 16, 3549–3558. [Google Scholar] [CrossRef]
- Clarke, R.J.; Johnson, J.W. , NMDA receptor NR2 subunit dependence of the slow component of magnesium unblock. J Neurosci 2006, 26, 5825–34. [Google Scholar] [CrossRef]
- Kotermanski, S.E.; Johnson, J.W. , Mg2+ imparts NMDA receptor subtype selectivity to the Alzheimer's drug memantine. Journal of Neuroscience 2009, 29, 2774–2779. [Google Scholar] [CrossRef]
- Sapkota, K.; Mao, Z.; Synowicki, P.; Lieber, D.; Liu, M.; Ikezu, T.; Gautam, V.; Monaghan, D.T. , GluN2D N-Methyl-d-Aspartate Receptor Subunit Contribution to the Stimulation of Brain Activity and Gamma Oscillations by Ketamine: Implications for Schizophrenia. J Pharmacol Exp Ther 2016, 356, 702–11. [Google Scholar] [CrossRef]
- Wyllie, D.J.; Behe, P.; Colquhoun, D. , Single-channel activations and concentration jumps: comparison of recombinant NR1a/NR2A and NR1a/NR2D NMDA receptors. J Physiol 1998, 510 Pt 1, 1–18. [Google Scholar] [CrossRef]
- Krupp, J.J.; Vissel, B.; Heinemann, S.F.; Westbrook, G.L. , Calcium-dependent inactivation of recombinant N-methyl-D-aspartate receptors is NR2 subunit specific. Molecular pharmacology 1996, 50, 1680–1688. [Google Scholar]
- Cull-Candy, S.; Brickley, S.; Farrant, M. , NMDA receptor subunits: diversity, development and disease. Current opinion in neurobiology 2001, 11, 327–335. [Google Scholar] [CrossRef]
- Hanson, E.; Armbruster, M.; Lau, L.A.; Sommer, M.E.; Klaft, Z.-J.; Swanger, S.A.; Traynelis, S.F.; Moss, S.J.; Noubary, F.; Chadchankar, J. , Tonic activation of GluN2C/GluN2D-containing NMDA receptors by ambient glutamate facilitates cortical interneuron maturation. Journal of Neuroscience 2019, 39, 3611–3626. [Google Scholar] [CrossRef]
- Swanger, S.A.; Vance, K.M.; Pare, J.-F.; Sotty, F.; Fog, K.; Smith, Y.; Traynelis, S.F. , NMDA receptors containing the GluN2D subunit control neuronal function in the subthalamic nucleus. Journal of Neuroscience 2015, 35, 15971–15983. [Google Scholar] [CrossRef]
- Pearlstein, E.; Gouty-Colomer, L.-A.; Michel, F.J.; Cloarec, R.; Hammond, C. , Glutamatergic synaptic currents of nigral dopaminergic neurons follow a postnatal developmental sequence. Frontiers in cellular neuroscience 2015, 9, 210. [Google Scholar] [CrossRef]
- Andrade-Talavera, Y.; Duque-Feria, P.; Paulsen, O.; Rodríguez-Moreno, A. , Presynaptic spike timing-dependent long-term depression in the mouse hippocampus. Cerebral Cortex 2016, 26, 3637–3654. [Google Scholar] [CrossRef]
- Feldman, D.E. , The spike-timing dependence of plasticity. Neuron 2012, 75, 556–571. [Google Scholar] [CrossRef]
- Dubois, C.J.; Lachamp, P.M.; Sun, L.; Mishina, M.; Liu, S.J. , Presynaptic GluN2D receptors detect glutamate spillover and regulate cerebellar GABA release. Journal of Neurophysiology 2016, 115, 271–285. [Google Scholar] [CrossRef]
- Watanabe, T.; Inoue, S.; Hiroi, H.; Orimo, A.; Muramatsu, M. , NMDA receptor type 2D gene as target for estrogen receptor in the brain. Molecular brain research 1999, 63, 375–379. [Google Scholar] [CrossRef]
- Riecher-Rössler, A.; Häfner, H.; Stumbaum, M.; Maurer, K.; Schmidt, R. , Can estradiol modulate schizophrenic symptomatology? Schizophrenia bulletin 1994, 20, 203–214. [Google Scholar] [CrossRef]
- McCarthny, C.R.; Du, X.; Wu, Y.C.; Hill, R.A. , Investigating the interactive effects of sex steroid hormones and brain-derived neurotrophic factor during adolescence on hippocampal NMDA receptor expression. International Journal of Endocrinology 2018, 2018. [Google Scholar] [CrossRef]
- Makino, C.; Shibata, H.; Ninomiya, H.; Tashiro, N.; Fukumaki, Y. , Identification of single-nucleotide polymorphisms in the human N-methyl-D-aspartate receptor subunit NR2D gene, GRIN2D, and association study with schizophrenia. Psychiatr Genet 2005, 15, 215–21. [Google Scholar] [CrossRef]
- Yu, Y.; Lin, Y.; Takasaki, Y.; Wang, C.; Kimura, H.; Xing, J.; Ishizuka, K.; Toyama, M.; Kushima, I.; Mori, D. , Rare loss of function mutations in N-methyl-D-aspartate glutamate receptors and their contributions to schizophrenia susceptibility. Translational psychiatry 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Beneyto, M.; Meador-Woodruff, J.H. , Lamina-specific abnormalities of NMDA receptor-associated postsynaptic protein transcripts in the prefrontal cortex in schizophrenia and bipolar disorder. Neuropsychopharmacology 2008, 33, 2175–2186. [Google Scholar] [CrossRef]
- Sodhi, M.S.; Simmons, M.; McCullumsmith, R.; Haroutunian, V.; Meador-Woodruff, J.H. , Glutamatergic gene expression is specifically reduced in thalamocortical projecting relay neurons in schizophrenia. Biological psychiatry 2011, 70, 646–654. [Google Scholar] [CrossRef]
- Andreasen, N.C. , A unitary model of schizophrenia: Bleuler's fragmented phrene as schizencephaly. Archives of general psychiatry 1999, 56, 781–787. [Google Scholar] [CrossRef]
- Anticevic, A.; Cole, M.W.; Repovs, G.; Murray, J.D.; Brumbaugh, M.S.; Winkler, A.M.; Savic, A.; Krystal, J.H.; Pearlson, G.D.; Glahn, D.C. , Characterizing thalamo-cortical disturbances in schizophrenia and bipolar illness. Cerebral cortex 2014, 24, 3116–3130. [Google Scholar] [CrossRef]
- Chen, P.; Ye, E.; Jin, X.; Zhu, Y.; Wang, L. , Association between thalamocortical functional connectivity abnormalities and cognitive deficits in schizophrenia. Scientific reports 2019, 9, 2952. [Google Scholar] [CrossRef]
- Schmitt, A.; Koschel, J.; Zink, M.; Bauer, M.; Sommer, C.; Frank, J.; Treutlein, J.; Schulze, T.; Schneider-Axmann, T.; Parlapani, E. , Gene expression of NMDA receptor subunits in the cerebellum of elderly patients with schizophrenia. European archives of psychiatry and clinical neuroscience 2010, 260, 101–111. [Google Scholar] [CrossRef]
- Lewis, D.A.; Hashimoto, T.; Volk, D.W. , Cortical inhibitory neurons and schizophrenia. Nat Rev Neurosci 2005, 6, 312–24. [Google Scholar] [CrossRef]
- Gonzalez-Burgos, G.; Lewis, D.A. , NMDA receptor hypofunction, parvalbumin-positive neurons, and cortical gamma oscillations in schizophrenia. Schizophrenia bulletin 2012, 38, 950–957. [Google Scholar] [CrossRef]
- Sohal, V.S.; Zhang, F.; Yizhar, O.; Deisseroth, K. , Parvalbumin neurons and gamma rhythms enhance cortical circuit performance. Nature 2009, 459, 698–702. [Google Scholar] [CrossRef]
- Cobb, S.R.; Buhl, E.H.; Halasy, K.; Paulsen, O.; Somogyi, P. , Synchronization of neuronal activity in hippocampus by individual GABAergic interneurons. Nature 1995, 378, 75–8. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Singer, W. , Abnormal neural oscillations and synchrony in schizophrenia. Nat Rev Neurosci 2010, 11, 100–13. [Google Scholar] [CrossRef]
- Cardin, J.A.; Carlén, M.; Meletis, K.; Knoblich, U.; Zhang, F.; Deisseroth, K.; Tsai, L.-H.; Moore, C.I. , Driving fast-spiking cells induces gamma rhythm and controls sensory responses. Nature 2009, 459, 663–667. [Google Scholar] [CrossRef]
- Buzsaki, G.; Wang, X.J. , Mechanisms of gamma oscillations. Annu Rev Neurosci 2012, 35, 203–25. [Google Scholar] [CrossRef] [PubMed]
- Alekseichuk, I.; Turi, Z.; de Lara, G.A.; Antal, A.; Paulus, W. , Spatial working memory in humans depends on theta and high gamma synchronization in the prefrontal cortex. Current Biology 2016, 26, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-M. A.; Stanford, A.D.; Mao, X.; Abi-Dargham, A.; Shungu, D.C.; Lisanby, S.H.; Schroeder, C.E.; Kegeles, L.S. , GABA level, gamma oscillation, and working memory performance in schizophrenia. NeuroImage: Clinical 2014, 4, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Spencer, K.M.; Niznikiewicz, M.A.; Nestor, P.G.; Shenton, M.E.; McCarley, R.W. , Left auditory cortex gamma synchronization and auditory hallucination symptoms in schizophrenia. BMC neuroscience 2009, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Spencer, K.M. , Baseline gamma power during auditory steady-state stimulation in schizophrenia. Frontiers in human neuroscience 2012, 5, 190. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.J.; Kasicki, S. , A systematic review of the effects of NMDA receptor antagonists on oscillatory activity recorded in vivo. Journal of Psychopharmacology 2013, 27, 972–986. [Google Scholar] [CrossRef]
- Kocsis, B.; Brown, R.E.; McCarley, R.W.; Hajos, M. , Impact of ketamine on neuronal network dynamics: Translational modeling of schizophrenia-relevant deficits. CNS neuroscience & therapeutics 2013, 19, 437–447. [Google Scholar]
- Hamm, J.P.; Gilmore, C.S.; Clementz, B.A. , Augmented gamma band auditory steady-state responses: support for NMDA hypofunction in schizophrenia. Schizophrenia research 2012, 138, 1–7. [Google Scholar] [CrossRef]
- Plourde, G.; Baribeau, J.; Bonhomme, V. , Ketamine increases the amplitude of the 40-Hz auditory steady-state response in humans. British journal of anaesthesia 1997, 78, 524–529. [Google Scholar] [CrossRef]
- Curley, A.A.; Lewis, D.A. , Cortical basket cell dysfunction in schizophrenia. The Journal of physiology 2012, 590, 715–724. [Google Scholar] [CrossRef]
- Lewis, D.A.; Curley, A.A.; Glausier, J.R.; Volk, D.W. , Cortical parvalbumin interneurons and cognitive dysfunction in schizophrenia. Trends Neurosci 2012, 35, 57–67. [Google Scholar] [CrossRef]
- Suzuki, Y.; Jodo, E.; Takeuchi, S.; Niwa, S.; Kayama, Y. , Acute administration of phencyclidine induces tonic activation of medial prefrontal cortex neurons in freely moving rats. Neuroscience 2002, 114, 769–779. [Google Scholar] [CrossRef]
- Jackson, M.E.; Homayoun, H.; Moghaddam, B. , NMDA receptor hypofunction produces concomitant firing rate potentiation and burst activity reduction in the prefrontal cortex. Proceedings of the National Academy of Sciences 2004, 101, 8467–8472. [Google Scholar] [CrossRef]
- Moghaddam, B.; Adams, B.; Verma, A.; Daly, D. , Activation of glutamatergic neurotransmission by ketamine: a novel step in the pathway from NMDA receptor blockade to dopaminergic and cognitive disruptions associated with the prefrontal cortex. Journal of Neuroscience 1997, 17, 2921–2927. [Google Scholar] [CrossRef]
- Lorrain, D.; Baccei, C.; Bristow, L.; Anderson, J.; Varney, M. , Effects of ketamine and N-methyl-D-aspartate on glutamate and dopamine release in the rat prefrontal cortex: modulation by a group II selective metabotropic glutamate receptor agonist LY379268. Neuroscience 2003, 117, 697–706. [Google Scholar] [CrossRef]
- Vollenweider, F.; Leenders, K.; Scharfetter, C.; Antonini, A.; Maguire, P.; Missimer, J.; Angst, J. , Metabolic hyperfrontality and psychopathology in the ketamine model of psychosis using positron emission tomography (PET) and [18F] fluorodeoxyglucose (FDG). European neuropsychopharmacology 1997, 7, 9–24. [Google Scholar] [CrossRef]
- Breier, A.; Malhotra, A.K.; Pinals, D.A.; Weisenfeld, N.I.; Pickar, D. , Association of ketamine-induced psychosis with focal activation of the prefrontal cortex in healthy volunteers. The American journal of psychiatry 1997. [Google Scholar]
- Miyamoto, S.; Leipzig, J.N.; Lieberman, J.A.; Duncan, G.E. , Effects of ketamine, MK-801, and amphetamine on regional brain 2-deoxyglucose uptake in freely moving mice. Neuropsychopharmacology 2000, 22, 400–412. [Google Scholar] [CrossRef]
- Duncan, G.E.; Moy, S.S.; Knapp, D.J.; Mueller, R.A.; Breese, G.R. , Metabolic mapping of the rat brain after subanesthetic doses of ketamine: potential relevance to schizophrenia. Brain research 1998, 787, 181–190. [Google Scholar] [CrossRef]
- Grunze, H.C.; Rainnie, D.G.; Hasselmo, M.E.; Barkai, E.; Hearn, E.F.; McCarley, R.W.; Greene, R.W. , NMDA-dependent modulation of CA1 local circuit inhibition. Journal of Neuroscience 1996, 16, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Kinney, J.W.; Davis, C.N.; Tabarean, I.; Conti, B.; Bartfai, T.; Behrens, M.M. , A specific role for NR2A-containing NMDA receptors in the maintenance of parvalbumin and GAD67 immunoreactivity in cultured interneurons. Journal of Neuroscience 2006, 26, 1604–1615. [Google Scholar] [CrossRef]
- Behrens, M.M.; Ali, S.S.; Dao, D.N.; Lucero, J.; Shekhtman, G.; Quick, K.L.; Dugan, L.L. , Ketamine-induced loss of phenotype of fast-spiking interneurons is mediated by NADPH-oxidase. Science 2007, 318, 1645–1647. [Google Scholar] [CrossRef]
- Keilhoff, G.; Becker, A.; Grecksch, G.; Wolf, G.; Bernstein, H.-G. , Repeated application of ketamine to rats induces changes in the hippocampal expression of parvalbumin, neuronal nitric oxide synthase and cFOS similar to those found in human schizophrenia. Neuroscience 2004, 126, 591–598. [Google Scholar] [CrossRef]
- Morrow, B.A.; Elsworth, J.D.; Roth, R.H. , Repeated phencyclidine in monkeys results in loss of parvalbumin-containing axo-axonic projections in the prefrontal cortex. Psychopharmacology 2007, 192, 283–290. [Google Scholar] [CrossRef]
- Belforte, J.E.; Zsiros, V.; Sklar, E.R.; Jiang, Z.; Yu, G.; Li, Y.; Quinlan, E.M.; Nakazawa, K. , Postnatal NMDA receptor ablation in corticolimbic interneurons confers schizophrenia-like phenotypes. Nature neuroscience 2010, 13, 76–83. [Google Scholar] [CrossRef]
- Cho, K.K.; Hoch, R.; Lee, A.T.; Patel, T.; Rubenstein, J.L.; Sohal, V.S. , Gamma rhythms link prefrontal interneuron dysfunction with cognitive inflexibility in Dlx5/6+/− mice. Neuron 2015, 85, 1332–1343. [Google Scholar] [CrossRef]
- Ikeda, K.; Araki, K.; Takayama, C.; Inoue, Y.; Yagi, T.; Aizawa, S.; Mishina, M. , Reduced spontaneous activity of mice defective in the ε4 subunit of the NMDA receptor channel. Molecular brain research 1995, 33, 61–71. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Yamada, K.; Noda, Y.; Mori, H.; Mishina, M.; Nabeshima, T. , Lower sensitivity to stress and altered monoaminergic neuronal function in mice lacking the NMDA receptor ε4 subunit. Journal of Neuroscience 2002, 22, 2335–2342. [Google Scholar] [CrossRef]
- Obiang, P.; Macrez, R.; Jullienne, A.; Bertrand, T.; Lesept, F.; Ali, C.; Maubert, E.; Vivien, D.; Agin, V. , GluN2D subunit-containing NMDA receptors control tissue plasminogen activator-mediated spatial memory. Journal of Neuroscience 2012, 32, 12726–12734. [Google Scholar] [CrossRef] [PubMed]
- Ide, S.; Ikekubo, Y.; Mishina, M.; Hashimoto, K.; Ikeda, K. , Cognitive impairment that is induced by (R)-ketamine is abolished in NMDA GluN2D receptor subunit knockout mice. International Journal of Neuropsychopharmacology 2019, 22, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Hagino, Y.; Kasai, S.; Han, W.; Yamamoto, H.; Nabeshima, T.; Mishina, M.; Ikeda, K. , Essential role of NMDA receptor channel ε4 subunit (GluN2D) in the effects of phencyclidine, but not methamphetamine. PloS one 2010, 5, e13722. [Google Scholar] [CrossRef]
- Takeuchi, T.; Kiyama, Y.; Nakamura, K.; Tsujita, M.; Matsuda, I.; Mori, H.; Munemoto, Y.; Kuriyama, H.; Natsume, R.; Sakimura, K. , Roles of the glutamate receptor ε2 and δ2 subunits in the potentiation and prepulse inhibition of the acoustic startle reflex. European Journal of Neuroscience 2001, 14, 153–160. [Google Scholar] [CrossRef]
- Yamamoto, H.; Kamegaya, E.; Sawada, W.; Hasegawa, R.; Yamamoto, T.; Hagino, Y.; Takamatsu, Y.; Imai, K.; Koga, H.; Mishina, M. , Involvement of the N-methyl-D-aspartate receptor GluN2D subunit in phencyclidine-induced motor impairment, gene expression, and increased Fos immunoreactivity. Molecular brain 2013, 6, 1–16. [Google Scholar] [CrossRef]
- Yamamoto, T.; Nakayama, T.; Yamaguchi, J.; Matsuzawa, M.; Mishina, M.; Ikeda, K.; Yamamoto, H. , Role of the NMDA receptor GluN2D subunit in the expression of ketamine-induced behavioral sensitization and region-specific activation of neuronal nitric oxide synthase. Neurosci Lett 2016, 610, 48–53. [Google Scholar] [CrossRef]
- Duncan, G.E.; Miyamoto, S.; Leipzig, J.N.; Lieberman, J.A. , Comparison of brain metabolic activity patterns induced by ketamine, MK-801 and amphetamine in rats: support for NMDA receptor involvement in responses to subanesthetic dose of ketamine. Brain research 1999, 843, 171–183. [Google Scholar] [CrossRef]
- Hrabetova, S.; Serrano, P.; Blace, N.; Heong, W.T.; Skifter, D.A.; Jane, D.E.; Monaghan, D.T.; Sacktor, T.C. , Distinct NMDA receptor subpopulations contribute to long-term potentiation and long-term depression induction. The Journal of Neuroscience 2000, 20, RC81. [Google Scholar] [CrossRef]
- Lozovaya, N.A.; Grebenyuk, S.E.; Tsintsadze, T.S.; Feng, B.; Monaghan, D.T.; Krishtal, O.A. , Extrasynaptic NR2B and NR2D subunits of NMDA receptors shape ‘superslow’afterburst EPSC in rat hippocampus. The Journal of physiology 2004, 558, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Eapen, A.V.; Fernández-Fernández, D.; Georgiou, J.; Bortolotto, Z.A.; Lightman, S.; Jane, D.E.; Volianskis, A.; Collingridge, G.L. , Multiple roles of GluN2D-containing NMDA receptors in short-term potentiation and long-term potentiation in mouse hippocampal slices. Neuropharmacology 2021, 201, 108833. [Google Scholar] [CrossRef] [PubMed]
- Ingram, R.; Kang, H.; Lightman, S.; Jane, D.E.; Bortolotto, Z.A.; Collingridge, G.L.; Lodge, D.; Volianskis, A. , Some distorted thoughts about ketamine as a psychedelic and a novel hypothesis based on NMDA receptor-mediated synaptic plasticity. Neuropharmacology 2018, 142, 30–40. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanisms by which dysfunction of GluN2D-containing NMDARs could potentially result in symptoms of schizophrenia. This simplified circuit shows that, in healthy individuals, GluN2D-containing NMDARs stimulate GABAergic interneurons on which they are expressed, which in turn mediate inhibition and coordinate the synchronized firing of networks of excitatory pyramidal neurons. These pyramidal neurons in turn stimulate dopaminergic (DA) neurons in the midbrain (via the ventral striatum and ventral pallidum, not shown) which project to the associative striatum. In healthy individuals, this circuit is under homeostatic balance, leading to gamma oscillations and healthy cognition. In schizophrenia, hypofunction of the GluN2D-containing NMDARs could lead to GABAergic interneurons increasing excitation of pyramidal neurons by reducing inhibition (disinhibition). The resulting aberrant increase in cortical excitation would lead to abnormal neuronal oscillations and impaired cognition. Moreover, this hyperactivity might lead to an overdrive in midbrain DA neurons and enhanced striatal DA which has been linked to the positive symptoms of schizophrenia.
Figure 1.
Mechanisms by which dysfunction of GluN2D-containing NMDARs could potentially result in symptoms of schizophrenia. This simplified circuit shows that, in healthy individuals, GluN2D-containing NMDARs stimulate GABAergic interneurons on which they are expressed, which in turn mediate inhibition and coordinate the synchronized firing of networks of excitatory pyramidal neurons. These pyramidal neurons in turn stimulate dopaminergic (DA) neurons in the midbrain (via the ventral striatum and ventral pallidum, not shown) which project to the associative striatum. In healthy individuals, this circuit is under homeostatic balance, leading to gamma oscillations and healthy cognition. In schizophrenia, hypofunction of the GluN2D-containing NMDARs could lead to GABAergic interneurons increasing excitation of pyramidal neurons by reducing inhibition (disinhibition). The resulting aberrant increase in cortical excitation would lead to abnormal neuronal oscillations and impaired cognition. Moreover, this hyperactivity might lead to an overdrive in midbrain DA neurons and enhanced striatal DA which has been linked to the positive symptoms of schizophrenia.
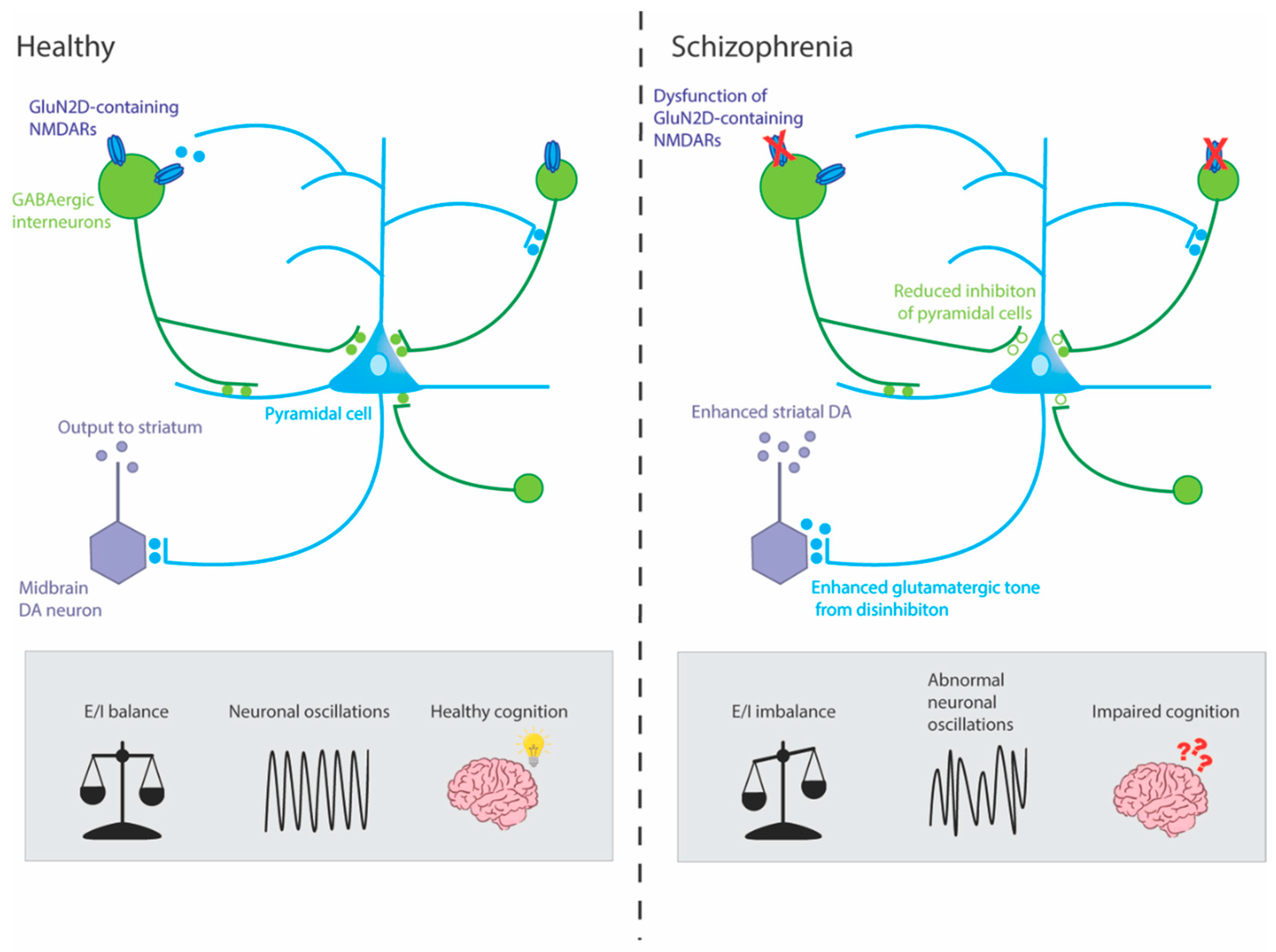
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



