
Submitted:
16 May 2023
Posted:
16 May 2023
You are already at the latest version
Abstract
Porcine parvovirus (PPV) 1 is a major causative pathogen of reproductive failure, and porcine circovirus (PCV) 2 is an etiological pathogen causing PCV-associated and PCV diseases worldwide. PPV2–6 and PCV3 were recently identified as newly emerging viruses with unknown pathogeneses. However, little is known of PPV1–6 and PCV3 effects or their co-infections with PCV2. In this study, we examined 926 samples (272 aborted pig fetuses and 654 pig lung tissues) from domestic pig farms for the presence of PPV1–6 or PCV3 and for PCV2 co-infection with these seven viruses. The prevalence of PPV and PCV was significantly greater in lung tissue samples than aborted pig fetus samples. PPV3–6 were detected for the first time in Korean domestic pigs. Co-infection rates with PCV2 and other viruses were significantly higher in abattoir samples than that in aborted pig fetus samples. Concurrent infections are more common than single infection in pig population and nu-merous infectious pathogens such as PPV and PCV2 can impact the respiratory diseases. Fur-thermore, Korean PPV1 strain (PPV1-82) was identical to the virulent PPV-27a strain in this respect. Our results describe, for the first time the prevalence of PPV1–6 and PCV3 infections and their co-infection rates with PCV2 in Korean domestic pigs.
Keywords:
Porcine parvovirus 1-6
; Porcine circovirus 2-3
; Prevalence
; Genetic diversity
1. Introduction
Porcine parvoviruses (PPVs) are small, non-enveloped, single-stranded linear genome DNA viruses of 4–6.3 kb [1]. PPVs are influential pathogens that induce reproductive failures in pigs, resulting in massive losses in the pig industry worldwide [2]. During the last two decades, several novel PPVs have been reported [2,3,4,5,6,7,8,9]. According to the International Committee on the Taxonomy of Viruses classification, to date, seven species of porcine parvoviruses (PPV) have been discovered in pigs, belonging to four genera: Protoparvovirus (PPV1), Tetraparvovirus (PPV2–3), Copiparvovirus (PPV4–6), and Chapparvovirus (PPV7) based on the similarity of the nonstructural protein 1 (NS1) [9,10]. PPV2 shares a high genetic similarity with parvoviruses detected in Chinese pigs with clinical symptoms of the post-weaning multisystemic wasting syndrome (PMWS) and high fever [5,7] and with parvoviruses first found in swine sera in Myanmar [4]. PPV3 was identified in Hong Kong in 2008 and was initially called porcine hokovirus (PHoV). However, phylogenetic analyses and comparative sequencing indicated that PHoV was more similar to the newly described bovine hokovirus and human parvoviruses four and five, which form a distinct cluster within parvoviruses [11]. Phylogenetic analysis showed PPV4, 5, and 6 were closely related and formed a distinct branch [8,12].
PPV co-infections with porcine circovirus (PCV) 2 in various combinations have been detected in wild boars and pigs [13,14,15,16]. Porcine circoviruses are classified into three types: PCV1 was detected in 1974 [17] and PCV2 was detected in 1997 [18] while PCV3 was first detected in pigs with multi-organ inflammation in the USA in 2015 [19]; PCV4 was recently detected in China in 2019 [20]. PCV2 is a small DNA virus with a circular genome and is the dominant causative pathogen of PMWS [21]. PCV2 has been associated with other porcine circovirus-associated diseases (PCVADs) in sows, resulting in litters of stillborn, mummified, or seropositive viable newborns, indicating the lack of antibodies in dams [22]. The most representative symptoms are porcine respiratory disease complex (PRDC), which typically occurs in 14–20-week-old pigs, PMWS in nursery or growing pigs, or porcine dermatitis and nephropathy syndrome (PDNS), which manifests during the final stages of growth [23,24,25,26,27]. PCV3 has been discovered in pigs with congenital tremor [28], PDNS [29,30], or reproductive failure [31].
The prevalence of PPV2, 3, 4, 5, and 6 and PCV3, and of their co-infections with PCV2, have not been determined in Korea. Therefore, this study was undertaken to investigate the prevalence of PPV1–6 and emerging PCV3, and their co-infection rates with PCV2. Furthermore, we characterized the genomes of PPV1–6, PCV2, and PCV3 and analyzed their phylogenetic trees, comparing the results for these newly detected viruses with those of other strains reported in Korea and elsewhere.
2. Materials and Methods
2.1. Sample preparation
The prevalence of PPV1, 2, 3, 4, 5, and 6 and PCV2 and 3 was assessed from 926 samples from 215 farms, including 272 aborted pig fetus-mixed tissue samples from 136 farms located in six different provinces and 654 lung tissue samples from 79 domestic pig farms obtained from abattoirs in five different provinces over 2.5 years from 2017 to 2019 (Supplementary Table S1). Among these, aborted pig fetuses were sought for pathogen differential diagnosis and domestic pigs aged five to six months were randomly selected from abattoirs. These samples were submitted to the Viral Disease Division of the Animal and Plant Quarantine Agency for the diagnosis of porcine reproductive failure disease, PMWS, PCVAD, and PRDC (OPTI-IAC-2001).
2.2. DNA extraction
Samples were homogenized using Precellys® CK28-R Lysing kit (bead tube for hard tissue homogenization; Bertin Technologies, Bretonneus, France) and Precellys® evolution (homogenizer; Bertin Technologies, Montigny-le-Bretonneux, France). Total DNA was extracted using a commercial DNeasy mini kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions.
2.3. Molecular Detection of viral nucleic acid
Individual PCR amplification for each pathogen was conducted using an AccuPower HotStart PCR Premix Kit (Bioneer, Daejeon, Korea) with detecting primers (PPV1-6 and PCV2-3) for PPV1–6 and PCV2 and 3 [3,13,32,33,34,35,36,37], respectively. Positive samples were PCR amplified for near-complete major capsid genes sequencing of PPV1 and PCV2 and near-complete sequencing of PPV2–6. When no signal was observed after up to 30 amplification cycles, the sample was ruled negative. All primers and amplification conditions used in this study are described in Supplementary Table S2.
2.4. DNA sequencing and phylogenetic analysis
Further sequencing of near-full-length major capsid genes was conducted to evaluate the possibility of genetic differences between strains. Target genes were amplified using the amplification sequencing primers listed in Supplementary Table S2. Positive amplicons were purified using an agarose gel extraction kit (Qiagen, Santa Clarita, CA, USA) and ligated into the pDrive vector (Qiagen, Santa Clarita, CA, USA) following the manufacturer’s instructions. Ligation products were transformed into Escherichia coli DH5 α-competent cells and incubated at 37℃ overnight. Ligated vectors were extracted using a plasmid miniprep kit (Inclone, Daejeon, Korea) and sequenced using Macrogen vector-sequencing primers (Macrogen Inc., Seoul, Korea). The obtained sequences were identified using the National Center for Biotechnology Information’s Basic Local Alignment Search Tool (BLAST) [38]. Multiple sequence alignments and homologies of nucleotide sequences of PPV1–6, PCV2, and PCV3 isolates were conducted using CLC Main Workbench (v. 7.0.3; CLC Bio, Qiagen, Aarhus, Denmark). Sequence alignment results were modified using BioEdit v. 7.2.5 (https://bioedit.software.informer.com/7.2/, accessed on March 1, 2021), and analyzed with a similarity matrix. Phylogenetic analysis was performed using MEGA v. 6.0 (https://megasoftware.net/, accessed on March 1, 2021) and reference sequences of major capsid genes [39] in the GenBank database (Supplementary Table S3), applying the maximum-likelihood approach and 1,000 bootstraps replicates values.
2.5. Statistical analysis
Pearson’s chi-squared test was used to determine the significance of intergroup differences. The analytical software package GraphPad Prism (v. 5.04; GraphPad Software Inc., La Jolla, CA, USA) was used for all statistical calculations. Statistical significance was set at P values < 0.05, and 95% confidence intervals (CI) were calculated.
The nucleotide sequences of the PPV1–6 and PCV2–3 obtained in this study were allocated the following GenBank accession numbers: PPV1 (MH447542-MH447550 and MZ856459), PPV2 (MH921914), PPV3 (MZ856460), PPV4 (MH921902, MH921910, MH921911, MH921915), PPV5 (MH21904-MH921905, MH921908, MH921912, MH921913), PPV6 (MH447535-MH447541), PCV2 (MK006032-MK006039), and PCV3 (OK362286) (Supplementary Table S4).
3. Results
3.1. Prevalence of PPV1 to 6 and PCV2 and 3 in pigs
Of the 926 samples tested, PPV4 was the least prevalent of both PPV and PCV viruses while PPV2 was the most prevalent. Virus prevalence in aborted fetuses was consistently much lower for all viruses than that in lung tissues (Table 1). The unique prevalence of these viruses was low, ranging from 0.3% for PPV4 (3/926, 95% CI: 0–0.7) to 3.1% for PPV1 (29/926, 95% CI: 2.0–4.3) (Supplementary Table S5).

Table 1.
Percentages of PPV1–6 and PCV2–3 positive samples among the 926 samples tested (aborted pig fetus samples (n = 272) and lung tissue abattoir samples (n = 654)).
Table 1.
Percentages of PPV1–6 and PCV2–3 positive samples among the 926 samples tested (aborted pig fetus samples (n = 272) and lung tissue abattoir samples (n = 654)).
| PPV1 | PPV2 | PPV3 | PPV4 | PPV5 | PPV6 | PCV2 | PCV3 | |
|---|---|---|---|---|---|---|---|---|
| Aborted pig fetus (n = 272) |
23.5% (64/272) |
16.9% (46/272) |
15.8% (43/272) |
7.0% (19/272) |
18.8% (51/272) |
18.0% (49/272) |
13.2% (36/272) |
9.9% (27/272) |
| Lung tissues from abattoirs (n = 654) |
59.6% (390/654) |
75.4% (493/654) |
71.6% (468/654) |
48.6% (318/654) |
62.5% (409/654) |
60.9% (398/654) |
67.4% (441/654) |
52.1% (341/654) |
| Total (n = 926) |
49.0% (454/926) |
58.2% (539/926) |
55.2% (511/926) |
36.4% (337/926) |
49.7% (460/926) |
48.3% (447/926) |
51.5% (477/926) |
39.7% (368/926) |
3.2. Prevalence of PPV1 co-infections with PPV2–6 or PCV3
PPV1 co-infection with PPV2–6 or PCV3 was present to a similar degree with all viruses, ranging from 22.9% with PPV4 (212/926, 95% CI: 20.2–25.6) to 34.8% with PPV2 (322/926, 95% CI: 31.7-37.8) (Table 2). Co-infection with PPV1 in aborted pig fetuses was low and was present from 1.1% to 5.9% of infections while this was much greater in the lung samples, ranging from 32.0% to 47.2%.
Table 2.
Percentage of PPV1 and PPV2 to PPV6 or PCV3 co-infection among all samples (n = 926), aborted pig fetus samples (n = 272), and lung tissues abattoir samples (n = 654).
Table 2.
Percentage of PPV1 and PPV2 to PPV6 or PCV3 co-infection among all samples (n = 926), aborted pig fetus samples (n = 272), and lung tissues abattoir samples (n = 654).
| PPV2 | PPV3 | PPV4 | PPV5 | PPV6 | PCV3 | |
|---|---|---|---|---|---|---|
| Aborted pig fetus (n = 272) |
4.8% (13/272) |
5.1% (14/272) |
1.1% (3/272) |
4.8% (13/272) |
5.9% (16/272) |
1.5% (4/272) |
| Lung tissues from abattoirs (n = 654) |
47.2% (309/654) |
45.1% (295/654) |
32.0% (209/654) |
43.3% (283/654) |
41.0% (268/654) |
33.3% (218/654) |
| Total (n = 926) |
34.8% (322/926) |
33.4% (309/926) |
22.9% (212/926) |
32.0% (296/926) |
30.7% (284/926) |
24.0% (222/926) |
3.3. Prevalence of PCV2 co-infections with PPV1–6 or PCV3
The prevalence of PCV2 co-infection with PPV1–6 or PCV3 was similar to that found for PCV1 co-infection, ranging from 25.8% with PPV4 (239/926, 95% CI: 23.0–28.6) to 38.9% with PPV2 (360/926, 95% CI: 35.7–42.0) (Table 3).
Table 3.
Percentages of PCV2 co-infection with PPV1 to PPV6 or PCV3 among all samples (n = 926), aborted pig fetus samples (n = 272), and lung tissue abattoir samples (n = 654).
Table 3.
Percentages of PCV2 co-infection with PPV1 to PPV6 or PCV3 among all samples (n = 926), aborted pig fetus samples (n = 272), and lung tissue abattoir samples (n = 654).
| PPV1 | PPV2 | PPV3 | PPV4 | PPV5 | PPV6 | PCV3 | |
|---|---|---|---|---|---|---|---|
| Aborted pig fetus (n = 272) |
1.8% (5/272) |
5.1% (14/272) |
5.1% (14/272) |
2.9% (8/272) |
5.1% (14/272) |
5.9% (16/272) |
4.0% (11/272) |
| Lung tissues from abattoirs (n = 654) |
40.4% (264/654) |
52.9% (346/654) |
51.2% (335/654) |
35.3% (231/654) |
43.0% (281/654) |
44.5% (291/654) |
39.9% (261/654) |
| Total (n = 926) |
29.0% (269/926) |
38.9% (360/926) |
37.7% (349/926) |
25.8% (239/926) |
31.9% (295/926) |
33.2% (307/926) |
29.4% (272/926) |
3.4. Genetic analyses of sequenced genomes
Sequencing of near full-length major capsid genes (PPV1, 1740–1749 bp) and near full-length genomes (PPV2–6, 5350–5979 bp) was conducted to evaluate the possibility of genetic variations between strains. Among the positive samples, representative samples were selected from the positive samples for phylogenetic analysis. Thus, we selected ten PPV1, one PPV2, one PPV3, four PPV4, four PPV5, and seven PPV6 positive samples. These strains were classified as VP2 (PPV1) or VP1 (PPV2 to 6) by amino acid-based phylogenetic tree analysis (Figure 1).
We detected ten PPV1 strains and sequenced each VP2 gene (designated 13, 20, 21, 22, 23, 25, 49, 82, 1220, and 190313). We detected nine sequences of VP2 genes of PPV1 in abattoirs samples and one sequence in aborted pig fetuses (190313). The amino acid similarities between the sequenced VP2 genes and the NADL-2 strain (GenBank acc. no. NC001718) ranged from 98.1% to 100%. Phylogenetic analysis showed that the Korean PPV1-82 strain in the abattoir samples clustered with the highly virulent strain PPV-27a strain (GenBank acc. no. AY684871) (Figure 1a).
Next, we detected and sequenced one PPV2 strain from the near full-length genome of PPV2 in abattoir samples. The Korean PPV2 sequence showed a high amino acid identity (98.5% in NS1 and 96.2% in VP1) with a Chinese strain (GenBank acc. no. KU745627). Parvoviruses genetically similar to the Korean PPV2 strain have been detected in China (Figure 1b).
Figure 1.
Phylogenetic trees based on near full-length major capsid genes (PPV1) and near full-length genomes (PPV2–6): PPV1 (a), PPV2 (b), PPV3 (c), PPV4 (d), PPV5 (e), and PPV6 (f). These strains were classified as VP2 (PPV1) or VP1 (PPV2 to 6) by amino acid-based phylogenetic tree analysis. The outgroup is K13-8 (a) and NADL-2 (b-f). The maximum-likelihood method with 1,000 bootstraps replicates values. The viral assembly sequences characterized in this study are denoted with red letters. GenBank accession numbers for the viral reference strains are shown in parentheses.
Figure 1.
Phylogenetic trees based on near full-length major capsid genes (PPV1) and near full-length genomes (PPV2–6): PPV1 (a), PPV2 (b), PPV3 (c), PPV4 (d), PPV5 (e), and PPV6 (f). These strains were classified as VP2 (PPV1) or VP1 (PPV2 to 6) by amino acid-based phylogenetic tree analysis. The outgroup is K13-8 (a) and NADL-2 (b-f). The maximum-likelihood method with 1,000 bootstraps replicates values. The viral assembly sequences characterized in this study are denoted with red letters. GenBank accession numbers for the viral reference strains are shown in parentheses.



One sequence for the near full-length genome of PPV3 was detected in abattoir samples. Regarding the PPV3 phylogenetic tree, complete major capsid genes were compared with those of initial isolates from Hong Kong, and the results showed that the Korean PPV3 capsid protein was highly conserved (Figure 1c). Korean PPV3 sequences indicated a high amino acid identity (98.1% in NS1 and 98.8% in VP1) with the Hong Kong strain (GenBank acc. no. EU200673).
Four sequences for near full-length PPV4 genomes were detected in two abattoir samples and two aborted pig fetuses (Figure 1d). Two PPV4 sequences (S180119-2, S180119-23) in aborted pigs exhibited 98.3% and 96.8% amino acid identity with a Chinese strain (GenBank acc. no. GU978965) and a US strain (GenBank acc. no. GQ387500), respectively. Additionally, two PPV4 sequences (S171206, S180605) in abattoir samples indicated high nucleotide identity (96.5% and 97.6%) with a Chinese strain (GenBank acc. no. GU978965) and a Brazilian strain (GenBank acc. no. KY586146), respectively.
The near full-length genomes of four novel Korean PPV5 strains were detected in three abattoir samples and one aborted pig fetus. Phylogenetic analysis based on VP1 amino acid sequences demonstrated that the four Korean PPV5 strains were closely related to a previously reported US strain (GenBank acc. no. JX896319). Additionally, at the nucleotide level (99.2%–99.4%), these four Korean PPV5 strains were closely related to each other and the US strain (Figure 1e).
Next in PPV6, the near full-length genomes of seven novel strains were detected in six abattoir samples and one aborted pig fetus sample. The near full-length genomes of four strains in abattoirs samples shared 98.5%–99.5% amino acid sequence identity with a Chinese PPV6 strain (GenBank acc. no. KF999685). Near full-length genomes of one PPV6 strain, discovered in abattoirs, and one PPV6 strain, discovered in aborted pig fetuses, shared 96.0% and 99.3% amino acid sequence identity with a US PPV6 strain, respectively (KR709266). The other PPV6 strains in abattoirs shared 98.2% amino acid sequence identity with a Brazilian PPV6 strain (GenBank acc. no. KY094494). Phylogenetic analysis of the VP1 genes indicated that PPV6 strains clustered in three distinct groups (Figure 1f).
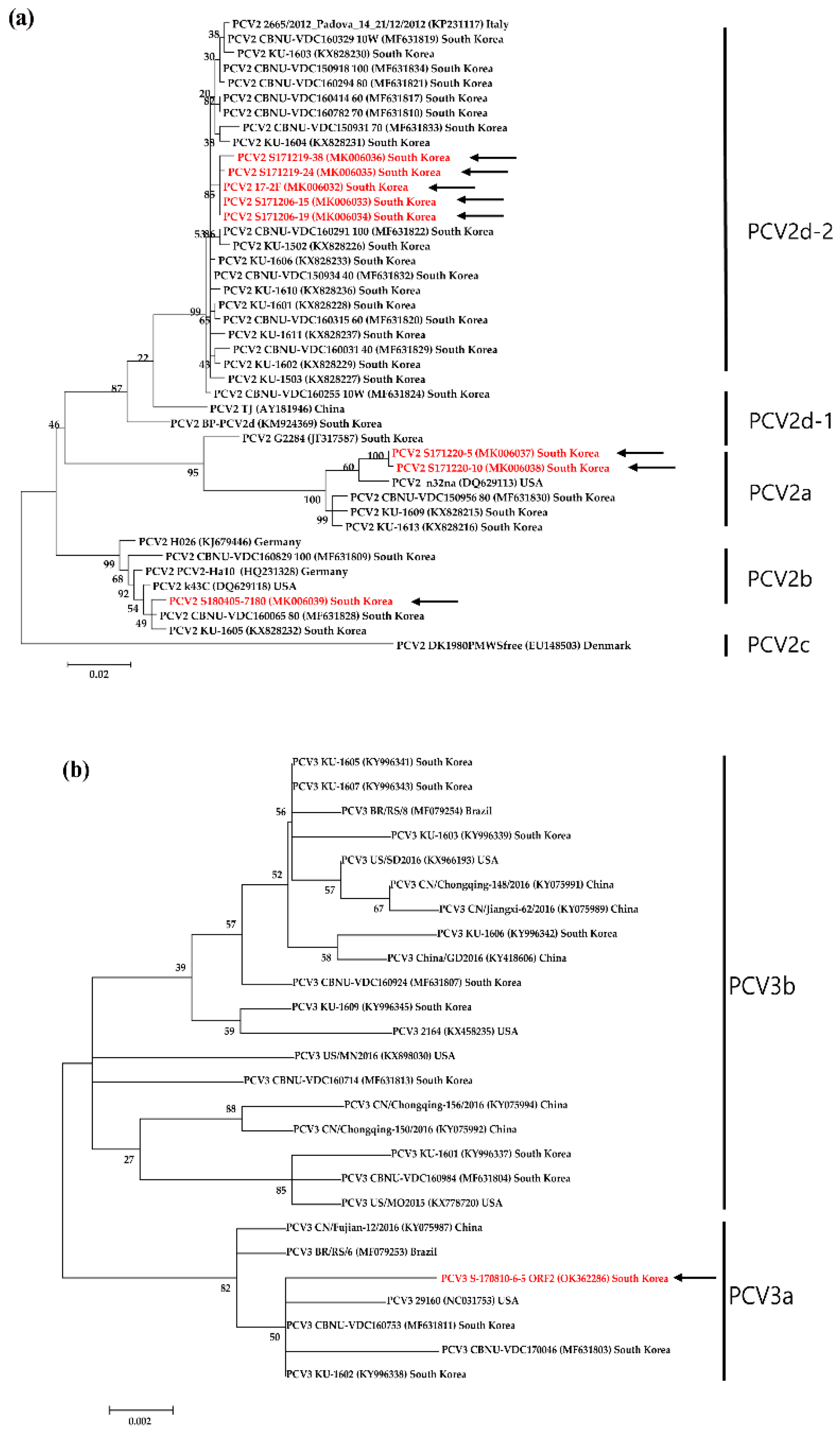
The PCV2 ORF2 strains were detected in seven abattoir samples (PCV2a: S171220-5, S171220-10, PCV2b: S180405-7180, PV2d: S171219-38, S171219-24, S171206-15, S171206-19) and in one aborted pig fetus sample (17-2F). The PCV2 ORF2 strains in abattoir samples shared 100% amino acid sequence identity with a Chinese PPV6 strain (GenBank acc. no. KX641127, MK305879, KX641127) and South Korea (GenBank acc. No. KF695388, MT376379). Phylogenetic analysis of the VP2 genes indicated that PCV2 strains clustered in five distinct groups. The PCV2 strains clustered in three groups: PCV2a (S171220-5 and S171220-10), PCV2b (S180405), and PV2d (S171219-38, S171219-24, 17-2F, S171206-15, and S171206-19) (Figure 2a).
PCV3 ORF2 strains were detected in one aborted pig fetus sample (PCV3 S-170810-6-5). The PCV3 strains shared 100% amino acid sequence identity with a Korean PCV3 strain (GenBank acc. no. MN698807). Phylogenetic analysis of the ORF2 genes indicated that PCV3 strains clustered in two distinct groups with the PCV3 strains clustered in the PCV3a group (Figure 2b).
Figure 2.
Phylogenetic trees of PCV2 (a) and PCV3 (b) based on the VP (ORF2) gene sequence by amino acid-based phylogenetic analysis. The maximum-likelihood method with 1,000 bootstraps replicates values. The viral assembly sequences characterized in this study are denoted with red letters. GenBank accession numbers for the viral reference strains are shown in parentheses.
Figure 2.
Phylogenetic trees of PCV2 (a) and PCV3 (b) based on the VP (ORF2) gene sequence by amino acid-based phylogenetic analysis. The maximum-likelihood method with 1,000 bootstraps replicates values. The viral assembly sequences characterized in this study are denoted with red letters. GenBank accession numbers for the viral reference strains are shown in parentheses.
4. Discussion
The prevalence of PPV1–7 in Europe has been shown to vary widely [32,40,41,42,43,44]. Here, we examined the molecular characteristics of the PPV1–6 and PCV2–3 strains circulating in domestic pigs in Korea and reported their detection rates and genetic characteristics. Notably, PPV3, PPV4, PPV5, and PPV6 were detected for the first time in Korean domestic pigs.
We found the PPV and PCV prevalence in lung tissue samples sourced from abattoirs was significantly greater than that found in aborted pig fetus samples. PPV1 has been previously detected in Korea, but only in one of 701 samples collected from 2013 to 2016 [45]. PPV2 has been detected in many other countries, including Hungary, the USA, Japan, Germany, and Thailand at rates of 6%, 21%, 58%, 78%, and 83%, respectively [46,47,48,49,50], and PPV2 was detected in two lung tissue samples in Korea in 2016 [51]. However, PPV3, 4, 5, nor 6 have not been previously detected in Korea. In this study, the PPV6 prevalence among aborted pig fetal samples (18.0%) was lower than that observed in China (50.0%), and the prevalence of PPV6 in abattoir samples (60.9%) was higher than that reported in China (15.6%) [8]. Previously, we detected PPV7 in aborted pig fetuses (24%) and domestic pigs (74.9%) in 2017 in Korea [52].
In the present study, we only detected unique infections of PPV and PCV viruses in a minority of aborted pig fetus samples. The genome sequences and phylogeny analyses indicate that the Korean strains are closely related to strains circulating in the US, Brazilian, and China Hong Kong. Novel PPVs, such as PPV4, PPV6, and PPV7, are suspected of causing reproductive failure, as they were detected in samples from aborted pig fetuses [8,46,52]. Furthermore, PPV4 and PPV6 were detected in adult female pigs with reproductive disorders [8,53]. Here, these pathogens (PPV1–6, PCV2, and PCV3) were all uniquely detected in aborted pig fetuses. The role of these viruses in reproductive failure disease pathogenesis and epidemiology needs to be studied in detail because of their current prevalence in major swine population countries worldwide.
Previous studies have detected PPV4/PPV5 co-infection in 15.6% of lung samples in infected pigs [3]. In this study, co-infections with PPV1 and PPV2, 3, 4, 5, or 6 or PCV3 were detected in Korean domestic pigs for the first time, and abattoir samples had considerably higher co-infection rates than in aborted pig fetus samples. Co-infections are more common than single infection in swine population and several infectious pathogens such as PPV and PCV2 can impact the respiratory diseases [54]. In addition, co-infection with PPV1 and PCV2 enhance PCVAD severity and pathological lesions in lymphoid tissues [55]. Remarkably, viremia of PPV2 was detected 2-3 weeks before the presence of respiratory signs, and the development of clinical PCVAD symptoms [5]. Since lung samples were collected at random from abattoir, we did not have any more detailed information in relation to the disease status of the animals from which the lungs studied here were originated. Therefore, the results show the prevalence of the studied viruses in the population but do not show the role of any of these viruses in the development of respiratory disease. Previous study proposes that PPV1 and PPV7 may rise the severity of PCV2 subclinical infections in fatteners through exciting PCV2 replication, and finally may lead to PCVAD in individuals. The mechanism of PPV1 impact on PCV2 infections is well documented, whereas PPV7 pathogenesis remains to be explained [40]. To address this, further studies are needed under controlled conditions to determine the pathogenic co-infection rates of PPV1 with PPV2–6 and PCV3.
PCV2 and PPV1–4 co-infection rates in domestic pigs detected in this study were higher than those previously reported in China (4% PPV1, 22.44% PPV2, 24% PPV3, and 12% PPV4) or Poland (2.9% for PPV1 to 26.6% for PPV2) [13,40]. Co-infection with PPV1 has previously been reported to enhance PCV2 infection by providing target cells for PCV2 replication or by altering expression profiles and cytokine formation, stimulating immune cells, and thereby suppressing PCV2 clearance [18]. Our study revealed that co-infection rates in lung tissue abattoir samples were significantly higher than in aborted pig fetus samples. PCV2/PPV4 co-infection has been detected in China, Hungary, Germany, and Thailand, with a prevalence of 2%, 6%, 7%, and 44%, respectively [44,46,48,50]. In previous studies, co-infections between PCV2 and at least one PPV species were common and were detected in 21.6% (112/519) of all tested serum samples, and as many as 70.4% of positive-PCV2 serum samples (112/159) [56]. Interestingly, PPV1, PPV3, PPV5, and PPV6 detection rates were significantly higher in positive-PCV2 than in negative-PCV2 serum samples, whereas no such correlation was found by Opriessnig et al. [56]. In this study, we discovered that the level of co-infection was significantly higher in PPV1–6, PCV2, and PCV3, although there was a lack of correlation between the co-infection prevalence and the pathogenesis of PPV1–6, PCV2, and PCV3 reproductive failure. However, we did identify co-infection in aborted pig fetuses.
Six PCV2/PPV1 co-infections were reported for the first time in Korean domestic pigs together with a higher prevalence of PCV2/PPV6 (5.9%) infection than PCV2/PPV1 to PPV5 or PCV2/PCV3 infections. Additionally, PCV3 positive clinical samples were consistently have been discovered to be co-infected with PCV2 at rates of 19.1% and 27.6%–39.4% in abattoirs and pig farms, respectively [57,58,59,60,61]. In Korea, co-infection rates for PCV2/PCV3 in serum and tissue were reported to be 10.8% and 35.0%, respectively, but no rate has been published for aborted fetuses [62]. Conversely, we detected PCV2/PCV3 co-infection in 4.0% of aborted pig fetuses. PCV3 and PCV2 preferentially target lymphoid tissues [29,63], and PCV2 infection causes immunosuppression in pigs and predispose animals to infections by other pathogens [64]. In previous studies, pathogen prevalence varied with pig age [65]. For instance, the PPV4 detection rates and viral loads were higher in serum samples from one- to four-month-old pigs with PMWS compared with healthy ones of the same age or pigs older than six months [65]. Furthermore, the fatteners had the highest PCV2 detection rates in serum [66]. The domestic pig samples from the abattoir were restricted to an age of five to six months. An additional study needs to be conducted to investigate the pathogenicity of each pathogen by collecting samples based on breeding stage and clinical symptoms.
Genetically, the PPV1-82 strain from domestic pigs clustered with those of the virulent PPV-27a strain. Cross-neutralization studies conducted against vaccine strains (IDT and NADL-2) that exhibit low neutralization activity against PPV-27a strain indicated that the existing PPV1 vaccine was ineffective in terms of preventing PPV1 spread [67,68]. Currently, inactivated vaccines are based on NADL-2 and related strains, which were isolated approximately 30 years ago. These vaccines are effective against homologous infections, although they do not prevent viral shedding and infection after challenge with the antigenically heterologous PPV-27a strain [68]. Infection of PPV-27a in pigs or injection of the virus into rabbits resulted in homologous neutralizing antibody titers that were 100- to 1,000-fold lower than heterologous titers against NADL-2, 143a, or MSV strains [67]. The presently available vaccines still seem matched for protecting individual pigs against PPV disease. In this study, except PPV1-82 strain, other detected PPV1 strains do not cluster with PPV1-27a, indicating that many strains are distributed in the Korean pig population. Therefore, more research should be conducted to develop effective PPV1 vaccine candidate strains.
Although the significance of amino acid substitutions has not been determined, amino acid changes in the PPV1 VP1 have been reported to be responsible for the pathogenicity of the Kresse strain of PPV1 [69]. It is unknown if the amino acid variability in PPV6 VP1 affects the pathogenicity or tissue tropism of the virus [12]. Nevertheless, this is the first study to determine the molecular characteristics of PPV3–6 strains circulating in tissue samples from abattoirs or aborted pig fetuses.
In conclusion, we report the prevalence of PPV1–6 and PCV3 co-infection and their co-infection rates with PCV2 in lung tissue samples from abattoirs and aborted pig fetuses. This study improves understanding of these viral infections in Korean pigs. Besides, results of this study are important that the investigation of the prevalence of the diseases for pork production or for the improvement of the biosecurity levels at the farm level in Korea. Multiple co-infections with these viruses were commonly detected in addition to co-infection with PCV2. Furthermore, the availability of near-full-length sequences of major capsid genes enabled us to compare the amino acid sequence between strains and comprehensive phylogenetic analysis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Summary of sample information collection, Table S2: PCR used in this study, Table S3: Sequence information for PPV and PCV strains and other reference isolates, Table S4: Prevalence of porcine parvovirus one to six (PPV1–6) and porcine circovirus two and three (PCV2 and PCV3) as classified by the number of pathogens detected, Table S5: Summary of PPV1–6 and PCV2–3 reference sequences.
Author Contributions
Conceptualization, I.O.O.; Formal analysis, H.C., and J.Y.L.; Funding acquisition, Y.H.L.; Methodology, S.Y.M.; Supervision, D.K.; Validation, J.Y.S., and B.H.H.; Writing–original draft, I.O.O.; Writing–editing and review, I.O.O., Y.H.L., and C.K.P.
Funding
This research was funded by the Ministry of Agriculture, Food and Rural Affairs, Republic of Korea (Grant no. N-1543083-2020-22-0202; B-1543083-2021-24-01).
Data Availability Statement
Data supporting the conclusions of this article are included within the article. The newly generated sequences were submitted to the GenBank database under the accession numbers MH447535-MH447550, MZ856459-MZ856460, MH921902, MH21904-MH921905, MH921908, MH921910-MH921915, and MK006032-MK006039. The datasets used and/or analyzed during this study are available from the corresponding author upon reasonable request.
Acknowledgments
None
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fernandes, S.; Boisvert, M.; Tijssen, P. Genetic Elements in the VP Region of Porcine Parvovirus Are Critical to Replication Efficiency in Cell Culture. J. Virol. 2011, 85 (6), 3025–3029; DOI: 10.1128/JVI.02215-10. [CrossRef]
- Hueffer, K.; Parrish, C. R. Parvovirus Host Range, Cell Tropism and Evolution. Curr. Opin. Microbiol. 2003, 6 (4), 392–398; DOI: 10.1016/s1369-5274(03)00083-3. [CrossRef]
- Xiao, C. T.; Giménez-Lirola, L. G.; Jiang, Y. H.; Halbur, P. G.; Opriessnig, T. Characterization of a Novel Porcine Parvovirus Tentatively Designated PPV5. PLOS ONE 2013, 8 (6), e65312; DOI: 10.1371/journal.pone.0065312. [CrossRef]
- Hijikata, M.; Abe, K.; Win, K. M.; Shimizu, Y. K.; Keicho, N.; Yoshikura, H. Identification of New Parvovirus DNA Sequence in Swine Sera from Myanmar. Jpn. J. Infect. Dis. 2001, 54 (6), 244–245.
- Wang, F.; Wei, Y.; Zhu, C.; Huang, X.; Xu, Y.; Yu, L.; Yu, X. Novel Parvovirus Sublineage in the Family of Parvoviridae. Virus Genes 2010, 41 (2), 305–308; DOI: 10.1007/s11262-010-0506-3. [CrossRef]
- Lau, S. K. P.; Woo, P. C. Y.; Tse, H.; Fu, C. T. Y.; Au, W. K.; Chen, X. C.; Tsoi, H. W.; Tsang, T. H. F.; Chan, J. S. Y.; Tsang, D. N. C.; Li, K. S. M.; Tse, C. W. S.; Ng, T. K.; Tsang, O. T. Y.; Zheng, B. J.; Tam, S.; Chan, K. H.; Zhou, B.; Yuen, K. Y. Identification of Novel Porcine and Bovine Parvoviruses Closely Related to Human Parvovirus 4. J. Gen. Virol. 2008, 89 (8), 1840–1848; DOI: 10.1099/vir.0.2008/000380-0. [CrossRef]
- Cheung, A. K.; Wu, G.; Wang, D.; Bayles, D. O.; Lager, K. M.; Vincent, A. L. Identification and Molecular Cloning of a Novel Porcine Parvovirus. Arch. Virol. 2010, 155 (5), 801–806; DOI: 10.1007/s00705-010-0646-8. [CrossRef]
- Ni, J.; Qiao, C.; Han, X.; Han, T.; Kang, W.; Zi, Z.; Cao, Z.; Zhai, X.; Cai, X. Identification and Genomic Characterization of a Novel Porcine Parvovirus (PPV6) in China. Virol. J. 2014, 11, 203; DOI: 10.1186/s12985-014-0203-2. [CrossRef]
- Palinski, R. M.; Mitra, N.; Hause, B. M. Discovery of a Novel Parvovirinae Virus, Porcine Parvovirus 7, by Metagenomic Sequencing of Porcine Rectal Swabs. Virus Genes 2016, 52 (4), 564–567; DOI: 10.1007/s11262-016-1322-1. [CrossRef]
- Streck, A. F.; Canal, C. W.; Truyen, U. Molecular Epidemiology and Evolution of Porcine Parvoviruses. Infect. Genet. Evol. 2015, 36, 300–306; DOI: 10.1016/j.meegid.2015.10.007. [CrossRef]
- Tuke, P. W.; Parry, R. P.; Appleton, H. Parvovirus PARV4 Visualization and Detection. J. Gen. Virol. 2010, 91 (2), 541–544; DOI: 10.1099/vir.0.014852-0. [CrossRef]
- Schirtzinger, E. E.; Suddith, A. W.; Hause, B. M.; Hesse, R. A. First Identification of Porcine Parvovirus 6 in North America by Viral Metagenomic Sequencing of Serum from Pigs Infected with Porcine Reproductive and Respiratory Syndrome Virus. Virol. J. 2015, 12, 170; DOI: 10.1186/s12985-015-0401-6. [CrossRef]
- Sun, J.; Huang, L.; Wei, Y.; Wang, Y.; Chen, D.; Du, W.; Wu, H.; Liu, C. Prevalence of Emerging Porcine Parvoviruses and Their Co-Infections with Porcine Circovirus type 2 in China. Arch. Virol. 2015, 160 (5), 1339–1344; DOI: 10.1007/s00705-015-2373-7. [CrossRef]
- Saekhow, P.; Kishizuka, S.; Sano, N.; Mitsui, H.; Akasaki, H.; Mawatari, T.; Ikeda, H. Coincidental Detection of Genomes of Porcine Parvoviruses and Porcine Circovirus type 2 Infecting Pigs in Japan. J. Vet. Med. Sci. 2016, 77 (12), 1581–1586; DOI: 10.1292/jvms.15-0167. [CrossRef]
- Sliz, I.; Vlasakova, M.; Jackova, A.; Vilcek, S. Characterization of Porcine Parvovirus type 3 and Porcine Circovirus type 2 in Wild Boars (sus Scrofa) in Slovakia. J. Wildl. Dis. 2015, 51 (3), 703–711; DOI: 10.7589/2015-01-005. [CrossRef]
- Sharma, R.; Saikumar, G. Porcine Parvovirus- and Porcine Circovirus 2-Associated Reproductive Failure and Neonatal Mortality in Crossbred Indian Pigs. Trop. Anim. Health Prod. 2010, 42 (3), 515–522; DOI: 10.1007/s11250-009-9454-0. [CrossRef]
- Tischer, I.; Rasch, R.; Tochtermann, G. Characterization of Papovavirus- and Picornavirus-Like Particles in Permanent Pig Kidney Cell Lines. Zentralbl. Bakteriol. Orig. A 1974, 226 (2), 153–167.
- Allan, G. M.; Kennedy, S.; McNeilly, F.; Foster, J. C.; Ellis, J. A.; Krakowka, S. J.; Meehan, B. M.; Adair, B. M. Experimental Reproduction of Severe Wasting Disease by Co-Infection of Pigs with Porcine Circovirus and Porcine Parvovirus. J. Comp. Pathol. 1999, 121 (1), 1–11; DOI: 10.1053/jcpa.1998.0295. [CrossRef]
- Phan, T. G.; Giannitti, F.; Rossow, S.; Marthaler, D.; Knutson, T. P.; Li, L.; Deng, X.; Resende, T.; Vannucci, F.; Delwart, E. Detection of a Novel Circovirus PCV3 in Pigs with Cardiac and Multi-Systemic Inflammation. Virol. J. 2016, 13 (1), 184; DOI: 10.1186/s12985-016-0642-z. [CrossRef]
- Zhang, H.H.; Hu, W.Q.; Li, J.Y.; Liu, T.N.; Zhou, J.Y.; Opriessnig, T.; Xiao, C.T. Novel Circovirus Species Identified in Farmed Pigs Designated as Porcine Circovirus 4, Hunan Province, China. Transbound. Emerg. Dis. 2020, 67 (3), 1057-1061; DOI: 10.1111/tbed.13446. [CrossRef]
- Meehan, B. M.; McNeilly, F.; Todd, D.; Kennedy, S.; Jewhurst, V. A.; Ellis, J. A.; Hassard, L. E.; Clark, E. G.; Haines, D. M.; Allan, G. M. Characterization of Novel Circovirus DNAs Associated with Wasting Syndromes in Pigs. J. Gen. Virol. 1998, 79 (9), 2171–2179; DOI: 10.1099/0022-1317-79-9-2171. [CrossRef]
- Dias, A. S.; Gerber, P. F.; Araújo, A. S.; Auler, P. A.; Gallinari, G. C.; Lobato, Z. I. Lack of Antibody Protection Against Porcine Circovirus 2 and Porcine Parvovirus in Naturally Infected Dams and Their Offspring. Res. Vet. Sci. 2013, 94 (2), 341–345; DOI: 10.1016/j.rvsc.2012.09.009. [CrossRef]
- Finsterbusch, T.; Mankertz, A. Porcine Circoviruses-Small but Powerful. Virus Res. 2009, 143 (2), 177–183; DOI: 10.1016/j.virusres.2009.02.009. [CrossRef]
- Ren, L.; Chen, X.; Ouyang, H. Interactions of Porcine Circovirus 2 with Its Hosts. Virus Genes 2016, 52 (4), 437–444; DOI: 10.1007/s11262-016-1326-x. [CrossRef]
- Segalés, J. Porcine Circovirus type 2 (pcv2) Infections: Clinical Signs, Pathology and Laboratory Diagnosis. Virus Res. 2012, 164 (1–2), 10–19; DOI: 10.1016/j.virusres.2011.10.007. [CrossRef]
- Liu, X.; Ouyang, T.; Ma, T.; Ouyang, H.; Pang, D.; Ren, L. Immunogenicity Evaluation of Inactivated Virus and Purified Proteins of Porcine Circovirus type 2 in Mice. B.M.C. Vet. Res. 2018, 14 (1), 137; DOI: 10.1186/s12917-018-1461-9. [CrossRef]
- Chae, C. Chae, C.A. Review of Porcine Circovirus 2-Associated Syndromes and Diseases. Vet. J. 2005, 169 (3), 326–336; DOI: 10.1016/j.tvjl.2004.01.012. [CrossRef]
- Chen, G. H.; Mai, K. J.; Zhou, L.; Wu, R. T.; Tang, X. Y.; Wu, J. L.; He, L. L.; Lan, T.; Xie, Q. M.; Sun, Y.; Ma, J. Y. Detection and Genome Sequencing of Porcine Circovirus 3 in Neonatal Pigs with Congenital Tremors in South China. Transbound. Emerg. Dis. 2017, 64 (6), 1650–1654; DOI: 10.1111/tbed.12702. [CrossRef]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B. M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2017, 91 (1), e01879-16; DOI: 10.1128/JVI.01879-16. [CrossRef]
- Stadejek, T.; Woźniak, A.; Miłek, D.; Biernacka, K. First Detection of Porcine Circovirus type 3 on Commercial Pig Farms in Poland. Transbound. Emerg. Dis. 2017, 64 (5), 1350–1353; DOI: 10.1111/tbed.12672. [CrossRef]
- Faccini, S.; Barbieri, I.; Gilioli, A.; Sala, G.; Gibelli, L. R.; Moreno, A.; Sacchi, C.; Rosignoli, C.; Franzini, G.; Nigrelli, A. Detection and Genetic Characterization of Porcine Circovirus type 3 in Italy. Transbound. Emerg. Dis. 2017, 64 (6), 1661–1664; DOI: 10.1111/tbed.12714. [CrossRef]
- Cadar, D.; Cságola, A.; Lorincz, M.; Tombácz, K.; Spînu, M.; Tuboly, T. Distribution and Genetic Diversity of Porcine Hokovirus in Wild Boars. Arch. Virol. 2011, 156 (12), 2233–2239; DOI: 10.1007/s00705-011-1125-6. [CrossRef]
- Gava, D.; Souza, C. K.; Schaefer, R.; Vincent, A. L.; Cantão, M. E.; Coldebella, A.; Ciacci-Zanella, J. R. A Taqman-Based Real-Time PCR for Detection and Quantification of Porcine Parvovirus 4. J. Virol. Methods 2015, 219, 14–17; DOI: 10.1016/j.jviromet.2015.03.011. [CrossRef]
- Schirtzinger, E. E.; Suddith, A. W.; Hause, B. M.; Hesse, R. A. First Identification of Porcine Parvovirus 6 in North America by Viral Metagenomic Sequencing of Serum from Pigs Infected with Porcine Reproductive and Respiratory Syndrome Virus. Virol. J. 2015, 12, 170; DOI: 10.1186/s12985-015-0401-6. [CrossRef]
- Ouardani, M.; Wilson, L.; Jetté, R.; Montpetit, C.; Dea, S. Multiplex PCR for Detection and Typing of Porcine Circoviruses. J. Clin. Microbiol. 1999, 37 (12), 3917–3924; DOI: 10.1128/JCM.37.12.3917-3924.1999. [CrossRef]
- Li, L.; Yuan, W.; Guo, H.; Ma, Z.; Song, Q.; Wang, X.; Li, H. Prevalence and Genetic Variation of Porcine Circovirus type 2 in Hebei, China from 2004 to 2014. Gene 2016, 586 (2), 222–227; DOI: 10.1016/j.gene.2016.04.014. [CrossRef]
- Ku, X.; Chen, F.; Li, P.; Wang, Y.; Yu, X.; Fan, S.; Qian, P.; Wu, M.; He, Q. Identification and Genetic Characterization of Porcine Circovirus type 3 in China. Transbound. Emerg. Dis. 2017, 64 (3), 703–708; DOI: 10.1111/tbed.12638. [CrossRef]
- Altschul, S. F.; Gish, W.; Miller, W.; Myers, E. W.; Lipman, D. J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215 (3), 403–410; DOI: 10.1016/S0022-2836(05)80360-2. [CrossRef]
- Cadar, D.; Dán, Á.; Tombácz, K.; Lőrincz, M.; Kiss, T.; Becskei, Z.; Spînu, M.; Tuboly, T.; Cságola, A. Phylogeny and Evolutionary Genetics of Porcine Parvovirus in Wild Boars. Infect. Genet. Evol. 2012, 12 (6), 1163–1171; DOI: 10.1016/j.meegid.2012.04.020. [CrossRef]
- Miłek, D.; Woźniak, A.; Podgórska, K.; Stadejek, T. Do Porcine Parvoviruses 1 Through 7 (PPV1-PPV7) have an Impact on Porcine Circovirus type 2 (PCV2) Viremia in Pigs?. Vet. Microbiol. 2020, 242, 108613; DOI: 10.1016/j.vetmic.2020.108613. [CrossRef]
- Lagan Tregaskis, P.; Staines, A.; Gordon, A.; Sheridan, P.; McMenamy, M.; Duffy, C.; Collins, P. J.; Mooney, M. H.; Lemon, K. Co-Infection Status of Novel Parvovirus’s (PPV2 to 4) with Porcine Circovirus 2 in Porcine Respiratory Disease Complex and Porcine Circovirus-Associated Disease from 1997 to 2012. Transbound. Emerg. Dis. 2021, 68 (4), 1979–1994; DOI: 10.1111/tbed.13846. [CrossRef]
- Miłek, D.; Woźniak, A.; Guzowska, M.; Stadejek, T. Detection Patterns of Porcine Parvovirus (PPV) and Novel Porcine Parvoviruses 2 Through 6 (PPV2-PPV6) in Polish Swine Farms. Viruses 2019, 11 (5), 474; DOI: 10.3390/v11050474. [CrossRef]
- Xiao, C. T.; Halbur, P. G.; Opriessnig, T. Complete Genome Sequence of a Novel Porcine Parvovirus (PPV) Provisionally Designated PPV5. Gnome Announc. Genome Announc. 2013, 1 (1), e00021-12; DOI: 10.1128/genomeA.00021-12. [CrossRef]
- Huang, L.; Zhai, S. L.; Cheung, A. K.; Zhang, H. B.; Long, J. X.; Yuan, S. S. Detection of a Novel Porcine Parvovirus, PPV4, in Chinese Swine Herds. Virol. J. 2010, 7, 333; DOI: 10.1186/1743-422X-7-333. [CrossRef]
- Oh, W. T.; Kim, R. Y.; Nguyen, V. G.; Chung, H. C.; Park, B. K. Perspectives on the Evolution of Porcine Parvovirus. Viruses 2017, 9 (8), 196; DOI: 10.3390/v9080196. [CrossRef]
- Cságola, A.; Lőrincz, M.; Cadar, D.; Tombácz, K.; Biksi, I.; Tuboly, T. Detection, Prevalence and Analysis of Emerging Porcine Parvovirus Infections. Arch. Virol. 2012, 157 (6), 1003–1010; DOI: 10.1007/s00705-012-1257-3. [CrossRef]
- Xiao, C. T.; Gerber, P. F.; Giménez-Lirola, L. G.; Halbur, P. G.; Opriessnig, T. Characterization of Porcine Parvovirus type 2 (PPV2) Which Is Highly Prevalent in the USA. Vet. Microbiol. 2013, 161 (3–4), 325–330; DOI: 10.1016/j.vetmic.2012.07.038. [CrossRef]
- Streck, A. F.; Homeier, T.; Foerster, T.; Fischer, S.; Truyen, U. Analysis of Porcine Parvoviruses in Tonsils and Hearts from Healthy Pigs Reveals High Prevalence and Genetic Diversity in Germany. Arch. Virol. 2013, 158 (6), 1173–1180; DOI: 10.1007/s00705-013-1603-0. [CrossRef]
- Saekhow, P.; Mawatari, T.; Ikeda, H. Coexistence of Multiple Strains of Porcine Parvovirus 2 in Pig Farms. Microbiol. Immunol. 2014, 58 (7), 382–387; DOI: 10.1111/1348-0421.12159. [CrossRef]
- Saekhow, P.; Ikeda, H. Prevalence and Genomic Characterization of Porcine Parvoviruses Detected in Chiangmai Area of Thailand in 2011. Microbiol. Immunol. 2015, 59 (2), 82–88; DOI: 10.1111/1348-0421.12218. [CrossRef]
- Lee, J. Y.; Kim, E. J.; Cho, I. S.; Lee, K. K.; Shin, Y. K. Complete Genome Sequences of Porcine Parvovirus 2 Isolated from Swine in the Republic of Korea. Genome Announc. 2017, 5 (15), e01738-16; DOI: 10.1128/genomeA.01738-16. [CrossRef]
- Ouh, I. O.; Park, S.; Lee, J. Y.; Song, J. Y.; Cho, I. S.; Kim, H. R.; Park, C. K. First Detection and Genetic Characterization of Porcine Parvovirus 7 from Korean Domestic Pig Farms. J. Vet. Sci. 2018, 19 (6), 855–857; DOI: 10.4142/jvs.2018.19.6.855. [CrossRef]
- Huang, L.; Zhai, S. L.; Cheung, A. K.; Zhang, H. B.; Long, J. X.; Yuan, S. S. Detection of a Novel Porcine Parvovirus, PPV4, in Chinese Swine Herds. Virol. J. 2010, 7, 333; DOI: 10.1186/1743-422X-7-333. [CrossRef]
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Créhan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; Zhu, J.; Gottschalk, M.; Summerfield, A.; Simon, G.; Bertho, N.; Meurens, F. Coinfections and Their Molecular Consequences in the Porcine Respiratory Tract. Vet. Res. 2020, 51 (1), 80; DOI: 10.1186/s13567-020-00807-8. [CrossRef]
- Opriessnig, T.; Fenaux, M.; Yu, S.; Evans, R.B.; Cavanaugh, D.; Gallup, J.M. Pallares, F.J.; Thacker, E.L.; Lager, K.M.; Meng, X.J.; Halbur, P.G. Effect of Porcine Parvovirus Vaccination on the Development of PMWS in Segregated Early Weaned Pigs Coinfected with Type 2 Porcine Circovirus and Porcine Parvovirus. Vet. Microbiol. 2004, 98 (3-4), 209–220; DOI: 10.1016/j.vetmic.2005.04.010. [CrossRef]
- Opriessnig, T.; Xiao, C. T.; Gerber, P. F.; Halbur, P. G. Identification of Recently Described Porcine Parvoviruses in Archived North American Samples from 1996 and Association with Porcine Circovirus Associated Disease. Vet. Microbiol. 2014, 173 (1–2), 9–16; DOI: 10.1016/j.vetmic.2014.06.024. [CrossRef]
- Li, X.; Qiao, M.; Sun, M.; Tian, K. A Duplex Real-Time PCR Assay for the Simultaneous Detection of Porcine Circovirus 2 and Circovirus 3. Virol. Sin. 2018, 33 (2), 181–186; DOI: 10.1007/s12250-018-0025-2. [CrossRef]
- Kim, H. R.; Park, Y. R.; Lim, D. R.; Park, M. J.; Park, J. Y.; Kim, S. H.; Lee, K. K.; Lyoo, Y. S.; Park, C. K. Multiple Real-Time Polymerase Chain Reaction for the Differential Detection of Porcine Circovirus 2 and 3. J. Virol. Methods 2017, 250, 11–16; DOI: 10.1016/j.jviromet.2017.09.021. [CrossRef]
- Wen, S.; Sun, W.; Li, Z.; Zhuang, X.; Zhao, G.; Xie, C.; Zheng, M.; Jing, J.; Xiao, P.; Wang, M.; Han, J.; Ren, J.; Liu, H.; Lu, H.; Jin, N. The Detection of Porcine Circovirus 3 in Guangxi, China. Transbound. Emerg. Dis. 2018, 65 (1), 27–31; DOI: 10.1111/tbed.12754. [CrossRef]
- Zheng, S.; Wu, X.; Zhang, L.; Xin, C.; Liu, Y.; Shi, J.; Peng, Z.; Xu, S.; Fu, F.; Yu, J.; Sun, W.; Xu, S.; Li, J.; Wang, J. The Occurrence of Porcine Circovirus 3 Without Clinical Infection Signs in Shandong Province. Transbound. Emerg. Dis. 2017, 64 (5), 1337–1341; DOI: 10.1111/tbed.12667. [CrossRef]
- Zhao, D.; Wang, X.; Gao, Q.; Huan, C.; Wang, W.; Gao, S.; Liu, X. Retrospective Survey and Phylogenetic Analysis of Porcine Circovirus type 3 in Jiangsu Province, China, 2008 to 2017. Arch. Virol. 2018, 163 (9), 2531–2538; DOI: 10.1007/s00705-018-3870-2. [CrossRef]
- Kim, S. C.; Nazki, S.; Kwon, S.; Juhng, J. H.; Mun, K. H.; Jeon, D. Y.; Jeong, C. G.; Khatun, A.; Kang, S. J.; Kim, W. I. The Prevalence and Genetic Characteristics of Porcine Circovirus type 2 and 3 in Korea. B.M.C. Vet. Res. 2018, 14 (1), 294; DOI: 10.1186/s12917-018-1614-x. [CrossRef]
- Jiang, H.; Wang, D.; Wang, J.; Zhu, S.; She, R.; Ren, X.; Tian, J.; Quan, R.; Hou, L.; Li, Z.; Chu, J.; Guo, Y.; Xi, Y.; Song, H.; Yuan, F.; Wei, L.; Liu, J. Induction of Porcine Dermatitis and Nephropathy Syndrome in Piglets by Infection with Porcine Circovirus type 3. J. Virol. 2019, 93 (4), e02045-18; DOI: 10.1128/JVI.02045-18. [CrossRef]
- Fu, X.; Fang, B.; Ma, J.; Liu, Y.; Bu, D.; Zhou, P.; Wang, H.; Jia, K.; Zhang, G. Insights into the Epidemic Characteristics and Evolutionary History of the Novel Porcine Circovirus type 3 in Southern China. Transbound. Emerg. Dis. 2018, 65 (2), e296–e303; DOI: 10.1111/tbed.12752. [CrossRef]
- Cibulski, S. P.; Teixeira, T. F.; Varela, A. P. M.; Scheffer, C. M.; Santos, H. F.; Lima, F. E. S.; Roehe, P. M. Ungulate Copiparvovirus 2 in Healthy and Postweaning Multisystemic Wasting Syndrome-Affected Pigs. Trop. Anim. Health Prod. 2017, 49 (5), 945–949; DOI: 10.1007/s11250-017-1279-7. [CrossRef]
- Woźniak, A.; Miłek, D.; Matyba, P.; Stadejek, T. Real-Time PCR Detection Patterns of Porcine Circovirus Type 2 (PCV2) in Polish Farms with Different Statuses of Vaccination against PCV2. Viruses 2019, 11 (12), 1135; DOI: 10.3390/v11121135. [CrossRef]
- Zeeuw, E. J. L.; Leinecker, N.; Herwig, V.; Selbitz, H. J.; Truyen, U. Study of the Virulence and Cross-Neutralization Capability of Recent Porcine Parvovirus Field Isolates and Vaccine Viruses in Experimentally Infected Pregnant Gilts. J. Gen. Virol. 2007, 88 (2), 420–427; DOI: 10.1099/vir.0.82302-0. [CrossRef]
- Jóźwik, A.; Manteufel, J.; Selbitz, H. J.; Truyen, U. Vaccination against Porcine Parvovirus Protects against Disease, but Does Not Prevent Infection and Virus Shedding After Challenge Infection with a Heterologous Virus Strain. J. Gen. Virol. 2009, 90 (10), 2437–2441; DOI: 10.1099/vir.0.012054-0. [CrossRef]
- Bergeron, J.; Hébert, B.; Tijssen, P. Genome Organization of the Kresse Strain of Porcine Parvovirus: Identification of the Allotropic Determinant and Comparison with Those of NADL-2 and Field Isolates. J. Virol. 1996, 70 (4), 2508–2515; DOI: 10.1128/JVI.70.4.2508-2515.1996. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



