
Submitted:
15 May 2023
Posted:
16 May 2023
You are already at the latest version
Abstract
The aim of this study was to examine whether H2O2, generated in such beverages as tea and coffee, is formed also in infusions of medicinal herbs and propose the possible mechanism of its formation. Infusions of sixteen herbs were studied and the presence of H2O2 was found in all cases. The highest concentrations of H2O2, exceeding 50 µM, were found in the infusions of leaves of Betula and Polygonum, and of the inflorescence of Tilia. These low amounts of H2O2 should not have a harmful effect and may even exert beneficial action, perhaps contributing to the health effects of the herbs. Generation of semiquinone and superoxide radicals was detected in herbal infusions, indicating that oxidation of phenolic compounds is the main source of H2O2 in these infusions. Herbal infusions were cytotoxic to human ovary cancer SKOV-3 and PEO1 cells; this cytotoxicity was compromised by catalase added to the growth medium, demonstrating the contribution of hydrogen peroxide to the cytotoxic action of herbal extracts.
Keywords:
hydrogen peroxide
; FOX assay
; medicinal herbs
; ovary cancer cells
; polyphenols
1. Introduction
Apart from the most popular beverages of plant origin such as tea and coffee, infusions of medicinal herbs are also often consumed as recommended by traditional or complementary medicine or as tea substituents. The commonly consumed beverages such as tea [1-4] and coffee [5-7] generate H2O2 upon brewing and subsequent standing. Hydrogen peroxide concentration in freshly brewed coffee was found to be 20-80 µM, depending on the type of coffee [6] while in the green and black tea it exceeded 200 µM and 700 µM after standing for 1 and 12 hours, respectively [2]. Lately, we documented the generation of H2O2 in cooked vegetables [8,9]. Wines, especially red wines, generate H2O2 [10,11] although wine components may also react with this compound [11]. Generation of hydrogen peroxide in these beverages is due to the autoxidation of antioxidants, mainly phenolic compounds contained in the plant material.
Hydrogen peroxide is a physiologically relevant compound, important in the defense against pathogenic microorganisms at micromolar up to millimolar concentrations and participating in intra- and intercellular signaling at nanomolar up to low micromolar concentrations [12,13]. Thus, not only endogenous generation but also exogenous sources of H2O2 in the human body are of interest.
Infusions of medicinal herbs are also rich in antioxidants including phenolic compounds and are an important source of dietary antioxidants. Part of the beneficial effects of medicinal herbs is attributed to the presence of their antioxidants, especially phenolics [14-17]. Phenolic compounds may be expected to generate hydrogen peroxide also in these infusions.
The aim of this study was to check whether H2O2 is generated in infusions of chosen medicinal herbs and inquire about the mechanism of H2O2 formation in the infusions. Sixteen various herbs were examined.
It has been reported that the cytotoxic effects of tea and wine in vitro were contributed by the hydrogen peroxide generated by them in the cell culture media [10]. Another aim of this study was thus to examine whether H2O2 also contributes to the cytotoxicity of several herbal infusions to human cancer cells in vitro.
2. Results and Discussion
2.1. Methodology of Hydrogen Peroxide Assay
The generation of hydrogen peroxide in the herbal infusions was assayed using the Ferrous Oxidation-Xylenol Orange method [18]. Although results obtained by this method are generally assumed to reflect the actual level of peroxides, our experience points to the possibility of artifacts when analyzing complex mixtures. Firstly, colored solutions can have basic absorbance at the analytical wavelength used, which may contribute to absorbance readings. Secondly, as the method is based on the oxidation of Fe2+ to Fe3+ by hydrogen peroxide, it can be affected by the presence of other compounds able to oxidize Fe2+. The presence of significant amounts of peroxides other than H2O2 is rather improbable in herbal infusions but polyphenols such as epigallocatechin gallate, gallic acid, and epicatechin, and their oxidation products, which can be present in plant infusions, can oxidize ferrous ions [19]. Therefore, we used two procedures, which should at least partly eliminate the artifacts but are also not free from drawbacks.
Procedure (i) using a Reagent Blank should eliminate the effect of an endogenous color of the solutions as well as the basic absorbance of the Xylenol Orange Reagent but it can be still subject to interference by compounds other than hydrogen peroxide able to oxidize Fe2+. Procedure (ii) based on the use absorbance difference between a sample not treated with catalase and a sample treated with catalase to decompose H2O2 should be specific for H2O2, irrespective of reactions of sample components with Fe2+. However, this procedure may lead to the underestimation of H2O2 concentration as some polyphenols inhibit catalase. The compounds most potent in catalase inhibition are epicatechin gallate and epigallocatechin gallate (IC50 values < 1 µM); they are even better inhibitors of the enzyme than the standard inhibitor azide [20]. As the strong inhibitors of catalase may be present in the infusions at various concentrations, the extent of underestimation of the H2O2 concentration may be different for various infusions, depending on their composition.
2.2. Hydrogen Peroxide is Generated in Herbal Infusions
Both applied procedures demonstrated the presence of hydrogen peroxide in herbal infusions (Figure 1). With Procedure (i), all the infusions showed the presence of hydrogen peroxide albeit at different concentrations; with Procedure (ii), the vast majority of the infusions contained significant amounts of H2O2, except for infusions of Artemisia, Taraxacum, Tussilago, and Urtica. The highest generation occurred during brewing and within the first hour but further incubation for up to 3 hours augmented the level of hydrogen peroxide in most cases, except for Cistus, Urtica (both Procedures), Tussilago (Procedure i), Gingko and Lavandula (Procedure ii). In Procedure (i), the highest concentrations of H2O2, reaching or even exceeding 50 µM, were detected in the infusions of the leaves of Betula (up to 70 µM) and Polygonum, and of the inflorescence of Tilia (up to over 50 µM). These concentrations exceeded that found in the green tea used (up to 35 µM). Generally, Procedure (ii) yielded lower H2O2 concentrations than Procedure (i).
It should be taken into account that the cultivar, location, growth conditions and time of collection, and even the extent of fragmentation may considerably affect the composition of phenolics and other compounds generating H2O2 so the values obtained in this study may be not representative of other batches of medicinal herbs. In any case, this study demonstrates that herbal infusions contain some amounts of H2O2.
The presence of micromolar concentrations of H2O2 in herbal infusions should not have deleterious consequences for health; on the contrary, it may even exert some beneficial effects. In the digestive tract, H2O2 may be partly decomposed by catalase and peroxidase activities present in the saliva and in gastric juice [21,22]. The residual H2O2 can exert bactericidal and virucidal action and thus may contribute to mouth hygiene and health [23,24] and act against Helicobacter pylori in the stomach [25]. Hydrogen peroxide promotes gastric motility [26]. High concentrations of H2O2 may damage colon cells, but low concentrations were suggested to stimulate cell divisions in the damaged intestine, thus contributing to epithelial repair [27]. Reaction with available iron in the intestine forms the hydroxyl radical and other free radicals, which facilitate digestion, since proteins subjected to free radical action may show enhanced susceptibility to proteolytic enzymes [28]. One can speculate whether the generation of H2O2 in the infusions of medicinal herbs can contribute to the prophylactic and therapeutic effects of at least some of these herbs.
2.3. Mechanism of Hydrogen Peroxide Generation
The generation of H2O2 in tea and coffee has been ascribed to the autoxidation of phenolic compounds present in these beverages such as epigallocatechin gallate, catechin, epicatechin, quercetin, delphinidin, hydroxytyrosol, rosmarinic acid and a range of other plant polyphenols [1-4, 6]. As polyphenols are secondary metabolites ubiquitous in plants, they are also present in other products of plant origin and may be expected to generate H2O2, especially upon cooking/heating, which inactivates enzymes decomposing this compound in vivo (mainly catalase and peroxidases). Formation of H2O2 due to phenolic oxidation is known to proceed in two steps: a polyphenol H2Q is oxidized to a semiquinone free radical HQ● in a reaction coupled to the reduction of molecular oxygen to the superoxide anion radical O2●- (1) and then semiquinone is oxidized to quinone Q producing second superoxide radical (2):
H2Q + O2 → HQ● + O2●- + H+
HQ● + O2 → Q + O2●- + H+,
Dismutation of superoxide radicals (3) or oxidation of another molecule by the superoxide radical (4) produces hydrogen peroxide
O2●- + O2●- + 2H+ → H2O2
H2S + O2●- + H+ → HS● + H2O2
where H2S is an oxidizable substrate and HS● represents a radical product of its one-electron oxidation.
EPR spectra of pulverized dry herbs and freshly prepared infusions demonstrated the presence of a singlet corresponding to the semiquinone radical. In spectra of dry herbs, This signal was superimposed on complex broad manganese and perhaps Fe signal, especially in dry samples; examples of signals of infusions are reported in Figure 2. The intensity of the signal varied for different herbs. EPR signals of dry samples of teas [29-31] and Salvia officinalis [32]; no signals of semiquinone radicals in herb extracts were published, to our best knowledge.
The plant material used in this study contained also other compounds producing hydrogen peroxide upon autoxidation, e.g. ascorbic acid [3] though the content of ascorbate is expected to be rather low in herbal infusions, and no signal of ascorbyl radical was detected in the ESR spectra.
Formation of superoxide radicals in herbal infusions was demonstrated by super-oxide-dismutase (SOD) inhibitable reduction of Nitro Blue Tetrazolium (NBT) and SOD-inhibitable oxidation of dihydroethidium (DHE). Both probes are not specific in their reactions. Reduction of NBT is often used to quantify superoxide production in animal and plant tissues [33,34] but other compounds may also reduce NBT so SOD-inhibitable reduction is a more reliable measure of superoxide production. Similarly, superoxide was suggested to be the main but not the only reactive oxygen species responsible for DHE oxidation [35,36]. The conditions of fluorescence measurements employed [37] were claimed to increase the selectivity of the method of DHE oxidation for superoxide, nevertheless, even this version of the method is far from being specific so SOD-inhibitable DHE oxidation is more reliable.
The presence of SOD-inhibitable components of NBT reduction and DHE oxidation in herbal infusions is demonstrated in Figure 3.
The concentration of NBT formazan formed in a SOD-inhibitable manner during 15-min brewing at 60 °C was 23.3±4.6 μM and 20.8.1±3.7 μM for 10% infusions of Betula leaves and Tilia inflorescence, respectively. These values probably underestimate the real superoxide production as a fraction of superoxide could still react with the probe or extract components avoiding dismutation by SOD. Nevertheless, these results document superoxide production, an intermediate in hydrogen formation, in herbal extracts.
3.3. Herbal Infusions Scavenge Hydrogen Peroxide
In most cases, the H2O2 concentration increased upon incubation of herbal infusions but in some cases, it decreased in time. It suggests that herbal infusions may not only generate but also scavenge H2O2. To test this possibility, some herbal extracts were added with H2O2, and the time course of the H2O2 concentration was monitored in infusions supplemented and not supplemented with hydrogen peroxide. Figure 4 shows that in all cases the H2O2 concentration measured in herbal infusions was significantly lower than the sum of concentration measured without the addition of exogenous H2O2 and the concentration of added H2O2 (finally 25 µM). This difference between the measured and expected hydrogen peroxide concentration was observed from the first measurement (“zero” time) and was higher in the infusion of herbs showing a decrease in the H2O2 concentration in the absence of exogenous H2O2 (Urtica and Taraxacum) than in the infusion of herbs in which the H2O2 concentration increased with time (exemplified by Betula and Tilia). The difference between the H2O2 concentrations measured in supplemented samples supplemented with H2O2 and not supplemented samples had a tendency to decrease in time (with the exception of Tilia inflorescence infusions). These results demonstrate that herbal infusions react with H2O2. The reaction was apparently the most rapid upon the addition of exogenous H2O2 (the “zero” time measurements were in fact performed 4±1 minutes after the addition of H2O2).
There are components of herbal infusions, especially polyphenols, that can react with H2O2 [38,39] and transition metal ions so a decrease in the H2O2 concentration can occur in these infusions. Thus, the H2O2 measured in herbal infusion represents a net outcome of the rate of production and scavenging of this compound.
2.4. Hydrogen Peroxide Contributes to the Cytotoxic Action of Herbal Infusions
An important consequence of H2O2 formation by herbal infusions concerns in vitro studies of their cytotoxic effect against malignant cells often performed as a measure of their potential anticancer activity. The H2O2 formed in these infusions may contribute to the cytotoxicity of these infusions, green tea, and wines [10], like isolated flavonoids such as epigallocatechin gallate and quercetin [3]. Cytotoxicity of various herbal extracts in vitro has been reported. E. g. G. biloba leaf extracts were found to be cytotoxic to carcinoma HSC-2 cells [40]. Plantago extracts were cytotoxic for human leukemia cells [41], MCF-7 breast cancer cells, A431 epidermal cells, and U87-MG glioma cell line [42].
We compared the cytotoxicity of five herbal infusions to human cancer ovary cells of two lines (SKOV-3 and PEO1) and human MRC-5 fibroblasts used as reference cells. PEO1 is a high-grade serous, and SKOV-3 is a non-serous, ovary cancer cell line. All the infusions studied except for the infusion of Taraxacum leaves, added at volumes of up to 20 µL (corresponding to infusions of 200 ng of dry herb) per 100 µL medium significantly decreased the survival of PEO1 cells; all the infusions except for those of Ginkgo leaves decreased the survival of SKOV-3 cells. The survival of MRC-5 fibroblasts was compromised only by the highest concentration of the infusions of Cistus and slightly enhanced by infusions of Taraxacum (Figure 5).
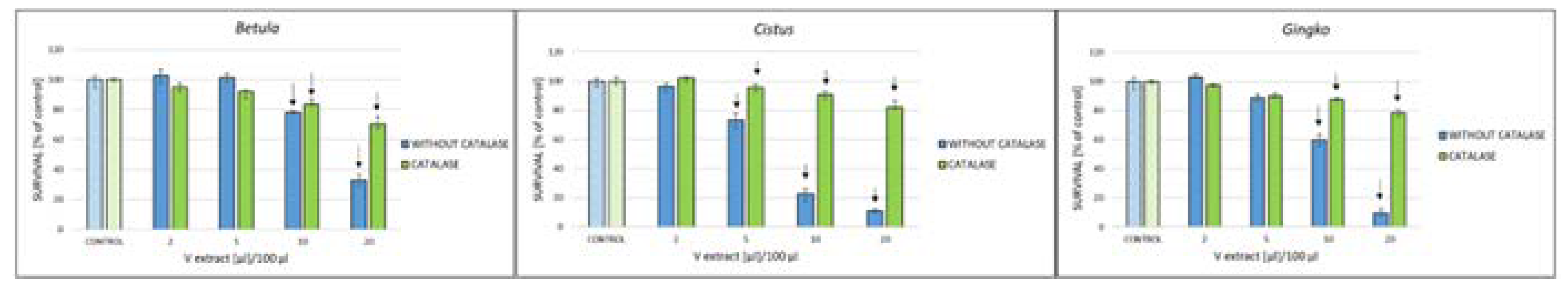
We checked whether the toxicity of three infusions, most effective with respect to the more sensitive PEO1 cells, is dependent on the production of H2O2. The presence of catalase, decomposing H2O2, significantly attenuated the cytotoxicity of all infusions (Figure 6). The toxic effects of the extracts included an H2O2-independent component, which was prevailing, but the effect of H2O2 also contributed to their toxicity as demonstrated by the protective action of catalase on cell survival. This effect is an artifact of in vitro experiments since in vivo conditions do not allow for an easy autoxidation of polyphenols due to lower oxygen concentration, and hydrogen peroxide, if formed, is efficiently disposed of, mainly by catalase and glutathione peroxidase. This difference between the in vitro and in vivo conditions should be kept in mind when evaluating the effect of herbal infusions on malignant cells. These infusions may be more effective in vitro, and one reason for it may be the formation of H2O2 under in vitro conditions (availability of oxygen). They may be less cytotoxic under in vivo conditions when the oxygen concentration is much smaller, the formation of hydrogen H2O2 due to autoxidation of herb components is much lower (if any) and the contribution of H2O2 to the herb cytotoxicity is much lower or absent.
3. Materials and Methods
3.1. Materials
Cell culture medium (McCoy’s 5A (cat. no 22330-021), RPMI + GlutaMAX (cat. no 72400-021), DMEM+GlutaMax (cat. no. 21885-025) and Dulbecco’s Phosphate Buffered Saline (DPBS) (cat. no. 14040-117) were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Fetal Bovine Serum (cat. no. S1813), Penicillin-Streptomycin solution (cat. no. L0022), Trypsin-EDTA solution (10×) (cat. no. X0930), and Phosphate-Buffered Saline without Ca2+ and Mg2+ (cat. no. P0750) were obtained from Biowest (Nuaillé, France). Ethanol (96%, cat. no. 396420113), glacial acetic acid (cat. no. 568760114), and Xylenol Orange were obtained from Avantor Performance Materials (Gliwice, Poland). Other reagents including catalase (cat. no. C40; ≥ 10,000 units/mg protein), Nitrotetrazolium Blue (NBT; cat. no. N6876), and dihydroethidium (cat. no. 309800) were purchased from Merck (Poznań, Poland).
75 cm2 flasks (cat. no. 156499) were provided by Thermo Fisher Scientific (Waltham, MA, USA). Transparent 96-well culture plates (cat. no 655180) were obtained from Greiner (Kremsmünster, Austria). Other sterile cell culture materials were provided by Nerbe (Winsen, Germany).
Human ovarian cancer cell lines (SKOV-3) and human lung normal fibroblast cell line (MRC-5) were obtained from American Type Culture Collection (ATCC). We also used a second ovarian cancer cell line derived from human ovarian cancer (PEO1) purchased in the European Collection of Authenticated Cell Cultures (ECACC).
Spectrophotometric and fluorimetric measurements were made in a Spark multimode microplate reader (Tecan Group Ltd., Männedorf, Switzerland).
3.2. Plant Material
The herbs of Achillea millefolium, Artemisia absinthium, Cistus incanus, Hypericum perforatum and Polygonum aviculare, leaves of Betula pendula, Taraxacum officinale, Melissa officinalis, Mentha piperica, Plantago lanceolata, Tussilago farfara, and Urtica dioica, inflorescence of Tilia cordata, and flowers of Calendula officinalis and Lavandula angustifolia were purchased in a local herbal store in Rzeszów, Poland. Leaves of Ginkgo biloba were collected from a tree grown in the garden of the corresponding author.
3.3. Preparation of Infusions
The material was crushed in a mortar and 250 mg of the dry material was treated with 25 ml of hot tap water. Immediately and after standing for 1 and 3 hours aliquots of the infusions were withdrawn for the assay of hydrogen peroxide. For the detection of semiquinone radicals and production of superoxide, 10% infusions were prepared (100 mg herb + 900 μL of water).
3.4. Determination of Hydrogen Peroxide
Hydrogen peroxide was estimated with Xylenol Orange [35] using two procedures:
Procedure (i): The infusion (180 µL) was added to two wells of a 96-well plate. One was added with 20 µL of the Xylenol Orange Reagent (2.5 mM Xylenol Orange/2.5 mM Mohr salt in 1.1 M perchloric acid) and another with a Blank Reagent containing the Mohr salt and perchloric acid, but no Xylenol Orange. In parallel, a blank for Xylenol Orange was prepared: two wells with 180 µL of water were added with 20 µL of the Xylenol Orange Reagent and Blank Reagent, respectively. After 30-min incubation, the absorbance of the samples was measured at 560 nm. Hydrogen peroxide concentration in the infusions was calculated on the basis of corrected absorbance Acorr:
Acorr = AS,XO – AS,B – (AW,XO – AW,B),
where AS,XO – absorbance of a sample added with the Xylenol Orange Reagent,
AS,B – absorbance of a sample added with the Blank Reagent,
AW,XO – absorbance of water added with the Xylenol Orange Reagent,
AS,B – absorbance of water added with the Blank Reagent.
Procedure (ii): The infusion (180 µL) was added to two wells of a 96-well plate. One well was added with 2 µL of water and another with 2 µL of a 1 mg/mL catalase solution. After 15-min incubation, 20.2 µL of the Xylenol Orange Reagent was added to both wells, and after 30-min incubation, absorbance was read at 560 nm. The difference in absorbance between the sample not treated with catalase and the catalase-treated sample was used as a measure of the hydrogen peroxide concentration. Preliminary experiments showed that the amount of catalase used was sufficient to fully decompose of 1 mM hydrogen peroxide present in a 200-µL sample during the incubation time employed.
The concentration of hydrogen peroxide was calculated using a calibration curve.
3.5. Detection of Semiquinone Radicals
Electron paramagnetic resonance (ESR) measurements were performed on a Bruker multifrequency and multiresonance FT-EPR ELEXSYS E580 spectrometer (Bruker Analytische Messtechnik, Rheinstetten, Germany) operating at the X-band (9.378989 GHz). The following settings were used: modulation amplitude, 0.4 G; modulation frequency, 100 kHz; microwave power, 94.64 mW; power attenuation, 2 dB; conversion time, 25 ms; sweep time, 102.4 s; powder sample: central field, 3501 G; scan range, 7000 G; liquid sample: central field, 3353.15 G; scan range, 100 G accumulation, 10.
The test samples (10% herbal infusions prepared as below) were deposited in quartz glass capillary tubes (Bruker, ER 221TUB/4 CFQ).
3.6. Detection of Superoxide Generation
Pulverized herbs were (10%) herbal extracts were added with NBT solution to a final concentration of 250 μM or dihydroethidium to a final concentration of 10 μM, SOD to a final concentration of 10 mg/ml (if present), and water to a final volume of 1 ml), heated in a thermoblock at 60 °C for 60 min, cooled and centrifuged. NBT reduction was measured at 530 nm; the concentration of the formazan was calculated in the supernatants using a millimolar absorption coefficient of 25.4 mM-1 cm-1 [43]. The fluorescence of ethidium was measured at the excitation wavelength of 405 nm and the emission wavelength of 570 nm to increase its specificity or the detection of the superoxide reaction product [37].
3.7. Cell Culture
SKOV-3 cells were cultured in McCoy’s 5A medium, PEO1 cells were cultured in RPMI+GlutaMAX and MRC-5 cells were cultured in DMEM+GlutaMax. Media used in the experiment were supplemented with 1% v/v penicillin and streptomycin solution and 10% heat-inactivated fetal bovine serum (FBS). Cells were incubated at 37 ℃ under 5% carbon dioxide and 95% humidity and were passaged at about 85% confluence. Cell morphology was examined under an inverted microscope with phase contrast Zeiss Primo Vert (Oberkochen, Germany). Ovarian adenocarcinoma cells and fibroblast viability was estimated by the Trypan Blue exclusion test; cells were counted in a Thoma hemocytometer (Superior Marienfeld, Lauda-Königshofen, Germany).
3.8. Estimation of Cytotoxicity of Herbal Infusions
SKOV-3 cells were seeded in wells of a clear 96-well plate at a density of 1 × 104 cells/well in 100 µL of culture medium. PEO1 cells were seeded at a density of 1.5 × 104 cells/well, and MRC-5 cells were seeded at a density of 7.5 × 103 cells/well and allowed to attach for 24 h at 37 ℃. Six 1% (w/v) herbal infusions (of leaves of Betula, Cistus, Ginkgo, Melissa, and Taraxacum) were used for determination of their toxicity towards human cancer ovary cells and fibroblasts. The infusions were prepared by treating 250 mg of the herbs with boiling phosphate-buffered saline (to provide isotonicity with the cell culture medium). After a 30-minute incubation, the infusions were centrifuged and sterilized using a syringe filter (0.2 μM). Cells were treated with different volumes of the infusions (0 μL, 2 μL, 5 μL, 10 μL and 20 μL added to 100 μL of the medium) for 24 h. After 24-h exposure, the medium was removed, replaced with 100 µL of 2% Neutral Red solution, and incubated for 1 h at 37 ℃. Then the cells were washed with PBS, fixed with 100 µL/well 50% ethanol, 49% H2O, and 1% glacial acetic acid, and shaken for 20 min (700 rpm) at room temperature. Absorbance was measured at 540 nm against 620 nm in a Spark multimode microplate reader (Tecan Group Ltd, Männedorf, Switzerland). Measurements were performed in sextuplicate.
3.9. Effect of Catalase on the Cytotoxicity of Herbal Infusions
PEO1 cells were seeded in a 96-well clear plate at density 1.5 × 104 cells/well and allowed to attach for 24 h at 37 ℃. After incubation cells were treated with the three most cytotoxic infusions as above but in the presence of 10 μg catalase/mL). After 24 h exposure, the medium was removed and cytotoxicity was estimated with Neutral Red as described above.
3.10. Statistical Analysis
To estimate differences between cells treated by chosen herbs and vegetable infusions and non-treated control Kruskal-Wallis test was performed. p ≤ 0.05 was considered as statistically significant. Differences between catalase-treated and non-treated trials were assessed by using the U Mann Whitney test with the same significance level (p ≤ 0.05). Statistical analysis of the data was performed using STATISTICA software package (version 13.1, StatSoft Inc. 2016, Tulsa, OK, USA).
4. Conclusions
Hydrogen peroxide is generated in herbal infusions and may be ingested with the infusions. These small amounts of H2O2 are unlikely to have adverse health effects; on the contrary, they may contribute to the beneficial effects of the infusions. Hydrogen peroxide contributes to the cytotoxic effects of herbal infusions found by in vitro studies.
Author Contributions
Conceptualization, I.S.-B.; methodology, I.S.-B., G.B. and I.S.; validation, I.S.-B.; investigation, A.T., N.P., I.S. and I.S.-B.; resources, writing—original draft preparation, G.B. and I.S.-B.; writing—review and editing, G.B. and I.S.-B.; supervision, I.S.-B.; project administration, I.S.-B. All authors have read and approved the final manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available from the corresponding author upon reasonable request.
Acknowledgments
We are indebted to Mrs. Edyta Bieszczad-Bedrejczuk, M. Sc., for the excellent technical help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Akagawa, M.; Shigemitsu, T.; Suyama, K. Production of hydrogen peroxide by polyphenols and polyphenol-rich beverages under quasi-physiological conditions. Biosci. Biotechnol. Biochem. 2003, 67(12), 2632–2640. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, H.; Maeda, M.; Okubo, S.; Shimamura, T. Role of hydrogen peroxide in bactericidal action of catechin. Biol. Pharm. Bull. 2004, 27(3), 277–281. [Google Scholar] [CrossRef]
- Grzesik, M.; Bartosz, G.; Stefaniuk, I.; Pichla, M.; Namieśnik, J.; Sadowska-Bartosz, I. Dietary antioxidants as a source of hydrogen peroxide. Food Chem. 2019, 278, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Q.; Gao, Y.; Long, D.; Yin, J.F.; Zeng, L.; Xu, Y.Q.; Xu, Y.Q. Effects Effects of Hydrogen Peroxide Produced by Catechins on the Aroma of Tea Beverages. Foods, 2022, 11(9), 1273.
- Hegele, J.; Münch, G.; Pischetsrieder, M. Identification of hydrogen peroxide as a major cytotoxic component in Maillard reaction mixtures and coffee. Mol. Nutr. Food Res. 2009, 53(6), 760–769. [Google Scholar] [CrossRef]
- Fujita, Y.; Wakabayashi, K.; Nagao, M.; Sugimura, T. Implication of hydrogen peroxide in the mutagenicity of coffee. Mut. Res. Lett. 1985, 144(4), 227–230. [Google Scholar] [CrossRef] [PubMed]
- Nagao, M.; Fujita, Y.; Wakabayashi, K.; Nukaya, H.; Kosuge, T.; Sugimura, T. Mutagens in coffee and other beverages. Environ. Health Perspect. 1986, 67, 89–91. [Google Scholar] [CrossRef]
- Bartosz, G.; Baran, S.; Grzesik-Pietrasiewicz, M.; Sadowska-Bartosz, I. The antioxidant capacity and hydrogen peroxide formation by black and orange carrots. Agric. Food Sci. 2022, 31(2), 71–77. [Google Scholar] [CrossRef]
- Bartosz, G.; Rajzer, K.; Grzesik-Pietrasiewicz, M.; Sadowska-Bartosz, I. Hydrogen peroxide is formed upon cooking of vegetables. Acta Biochim. Pol. 2022, 69(2), 471–474. [Google Scholar] [CrossRef]
- Chai, P.C.; Long, L.H.; Halliwell, B. Contribution of hydrogen peroxide to the cytotoxicity of green tea and red wines. Biochem. Biophys. Res. Commun. 2003, 304(4), 650–654. [Google Scholar] [CrossRef]
- Tama, A.; Bartosz, G.; Sadowska-Bartosz, I. Is hydrogen peroxide generated in wine? Food Biosci. 2022, 45, 101487. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21(7), 363–383. [Google Scholar] [CrossRef]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. (2022). Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23(7), 499-515.
- Ali, S.S.; Kasoju, N.; Luthra, A.; Singh, A.; Sharanabasava, H.; Sahu, A.; Bora, U. Indian medicinal herbs as sources of antioxidants. Food Res. Int. 2008, 41(1), 1–15. [Google Scholar] [CrossRef]
- Dragland, S.; Senoo, H.; Wake, K.; Holte, K.; Blomhoff, R. Several culinary and medicinal herbs are important sources of dietary antioxidants. J. Nutr. 2003, 133(5), 1286–1290. [Google Scholar] [CrossRef]
- Matkowski, A.; Jamiołkowska-Kozlowska, W.; Nawrot, I. Chinese medicinal herbs as source of antioxidant compounds-where tradition meets the future. Curr. Med. Chem. 2013, 20(8), 984–1004. [Google Scholar]
- Wojcikowski, K.; Stevenson, L.; Leach, D.; Wohlmuth, H.; Gobe, G. Antioxidant capacity of 55 medicinal herbs traditionally used to treat the urinary system: a comparison using a sequential three-solvent extraction process. J. Altern. Complement. Med. 2007, 13(1), 103–109. [Google Scholar] [CrossRef]
- Gay, C.A.; Gebicki, J.M. Measurement of protein and lipid hydroperoxides in biological systems by the ferric-xylenol orange method. Anal. Biochem. 2003, 315(1), 29–35. [Google Scholar] [CrossRef]
- Perron, N.R.; Wang, H.C.; Deguire, S.N.; Jenkins, M.; Lawson, M.; Brumaghim, J.L. Kinetics of iron oxidation upon polyphenol binding. Dalton Trans. 2010, 39(41), 9982–9987. [Google Scholar] [CrossRef]
- Krych, J.; Gebicka, L. Catalase is inhibited by flavonoids. Int. J. Biol. Macromol. 2013, 58, 148–153. [Google Scholar] [CrossRef]
- Durak, I.; Ormeci, N.; Akyol, O.; Canbolat, O.; Kavutçu, M.; Bülbül, M. Adenosine deaminase 5'-nucleotidase xanthine oxidase superoxide dismutase and catalase activities in gastric juices from patients with gastric cancer ulcer and atrophic gastritis. Dig. Dis. Sci. 1994, 39(4), 721–728. [Google Scholar] [CrossRef]
- Toczewska, J.; Konopka, T. Activity of enzymatic antioxidants in periodontitis: A systematic overview of the literature. Dent. Med. Probl. 2019, 56(4), 419–426. [Google Scholar] [CrossRef]
- Caruso, A.A.; Del Prete, A.; Lazzarino, A.I. Hydrogen peroxide and viral infections: A literature review with research hypothesis definition in relation to the current covid-19 pandemic. Med. Hypoth. 2020, 144, 109910. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, P.; Sager, B.; Fa, A.; Liang, T.; Lozano, C.; Khazzam, M. Bactericidal efficacy of hydrogen peroxide on Cutibacterium acnes. Bone Joint Res. 2019, 8(1), 3–10. [Google Scholar] [CrossRef] [PubMed]
- Di, J.; Zhang, J.; Cao, L.; Huang, T.T.; Zhang, J.X.; Mi, Y.N.; Xiao, X.; Yan, P.P.; Wu, M.L.; Yao, T.; Liu, D.Z.; Liu, J.; Cao, Y.X. Hydrogen Peroxide-Mediated Oxygen Enrichment Eradicates Helicobacter pylori In Vitro and In Vivo. Antimicrob. Agents Chemother. 2020, 64(5), e02192–e02219. [Google Scholar] [CrossRef]
- Fajardo, A.F.; Sobchak, C.; Shifrin, Y.; Pan, J.; Gonska, T.; Belik, J. Hydrogen peroxide promotes gastric motility in the newborn rat. Ped. Res. 2018, 84(5), 751–756. [Google Scholar] [CrossRef] [PubMed]
- Craven, P.A.; Pfanstiel, J.; DeRubertis, F.R. Role of reactive oxygen in bile salt stimulation of colonic epithelial proliferation. J. Clin. Invest. 1986, 77, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.P.; Dean, R.T. Fragmentation of proteins by free radicals and its effect on their susceptibility to enzymic hydrolysis. Biochem. J. 1986, 234(2), 399–403. [Google Scholar] [CrossRef]
- Morsy, M.A.; Khaled, M.M. Novel EPR characterization of the antioxidant activity of tea leaves. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2002, 58(6), 1271–1277. [Google Scholar] [CrossRef]
- Polovka, M.; Brezová, V.; Stasko, A. Antioxidant properties of tea investigated by EPR spectroscopy. Biophys. Chem. 2003, 106(1), 39–56. [Google Scholar] [CrossRef]
- Polat, M.; Korkmaz, M. Detection of irradiated black tea (Camellia sinensis) and rooibos tea (Aspalathus linearis) by ESR spectroscopy. Food Chem. 2008, 107(2), 956–961. [Google Scholar] [CrossRef]
- Çam, S.T.; Engin, B. Identification of irradiated sage tea (Salvia officinalis L.) by ESR spectroscopy. Radiat. Phys. Chem. 2010, 79(4), 540-544.
- Bournonville, C.F.; Díaz-Ricci, J.C. Quantitative determination of superoxide in plant leaves using a modified NBT staining method. Phytochem. Anal. 2011, 22(3), 268–271. [Google Scholar] [CrossRef]
- Wohlgemuth, H.; Mittelstrass, K.; Kschieschan, S.; Bender, J.; Weigel, H.J.; Overmyer, K.; Kangasjärvi, J.; Sandermann, H.; Langebartels, C. Activation of an oxidative burst is a general feature of sensitive plants exposed to the air pollutant ozone. Plant Cell Env. 2002, 25(6), 717–726. [Google Scholar] [CrossRef]
- Fink, B.; Laude, K.; McCann, L.; Doughan, A.; Harrison, D.G.; Dikalov, S. Detection of intracellular superoxide formation in endothelial cells and intact tissues using dihydroethidium and an HPLC-based assay. Am. J. Physiol. Cell Physiol. 2004, 287(4), C895–C902. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Kalyanaraman, B. Hydroethidine- and MitoSOX-derived red fluorescence is not a reliable indicator of intracellular superoxide formation: another inconvenient truth. Free Radic. Biol. Med. 2010, 48(8), 983–1001. [Google Scholar] [CrossRef] [PubMed]
- Nazarewicz, R.R.; Bikineyeva, A.; Dikalov, S.I. Rapid and specific measurements of superoxide using fluorescence spectroscopy. J. Biomol. Screen. 2013, 18(4), 498–503. [Google Scholar] [CrossRef] [PubMed]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41(6), 753–758. [Google Scholar] [CrossRef]
- Ozyürek, M.; Bektaşoğlu, B.; Güçlü, K.; Apak, R. Hydroxyl radical scavenging assay of phenolics and flavonoids with a modified cupric reducing antioxidant capacity (CUPRAC) method using catalase for hydrogen peroxide degradation. Anal. Chim. Acta 2008, 616(2), 196–206. [Google Scholar] [CrossRef]
- Babich, H.; Ackerman, N.J.; Burekhovich, F.; Zuckerbraun, H.L.; Schuck, A.G. Gingko biloba leaf extract induces oxidative stress in carcinoma HSC-2 cells. Toxicol. In Vitro. 2009, 23(6), 992–999. [Google Scholar] [CrossRef]
- Mohamed, I.K.; Osama, M.A.; Samiha, M.; Zahrat, E.M. Biochemical studies on Plantago major L. and Cyamopsis tetragonoloba L. Int. J. Biodivers. Conserv. 2011, 3, 83–91. [Google Scholar]
- Rezadoost, M.H.; Kumleh, H.H.; Ghasempour, A. Cytotoxicity and apoptosis induction in breast cancer, skin cancer and glioblastoma cells by plant extracts. Mol. Biol. Rep. 2019, 46(5), 5131–5142. [Google Scholar] [CrossRef]
- Oritani, T.; Fukuhara, N.; Okajima, T.; Kitamura, F.; Ohsaka, T. Electrochemical and spectroscopic studies on electron-transfer reaction between novel water-soluble tetrazolium salts and a superoxide ion. Inorgan. Chim. Acta 2004, 357(2), 436–442. [Google Scholar] [CrossRef]
Figure 1.
The concentration of hydrogen peroxide in the infusions of medicinal herbs and of green tea immediately after brewing and after 1 hour and 3 hours standing at ambient temperature. H2O2 concentration was estimated by two procedures, based on the use of a reagent blank (i) and on the decomposition of H2O2 by 10 µg/mL catalase in one of two parallel samples (ii). aP <0.05, bP < 0.01, cP < 0.001 with respect to zero generation.
Figure 1.
The concentration of hydrogen peroxide in the infusions of medicinal herbs and of green tea immediately after brewing and after 1 hour and 3 hours standing at ambient temperature. H2O2 concentration was estimated by two procedures, based on the use of a reagent blank (i) and on the decomposition of H2O2 by 10 µg/mL catalase in one of two parallel samples (ii). aP <0.05, bP < 0.01, cP < 0.001 with respect to zero generation.
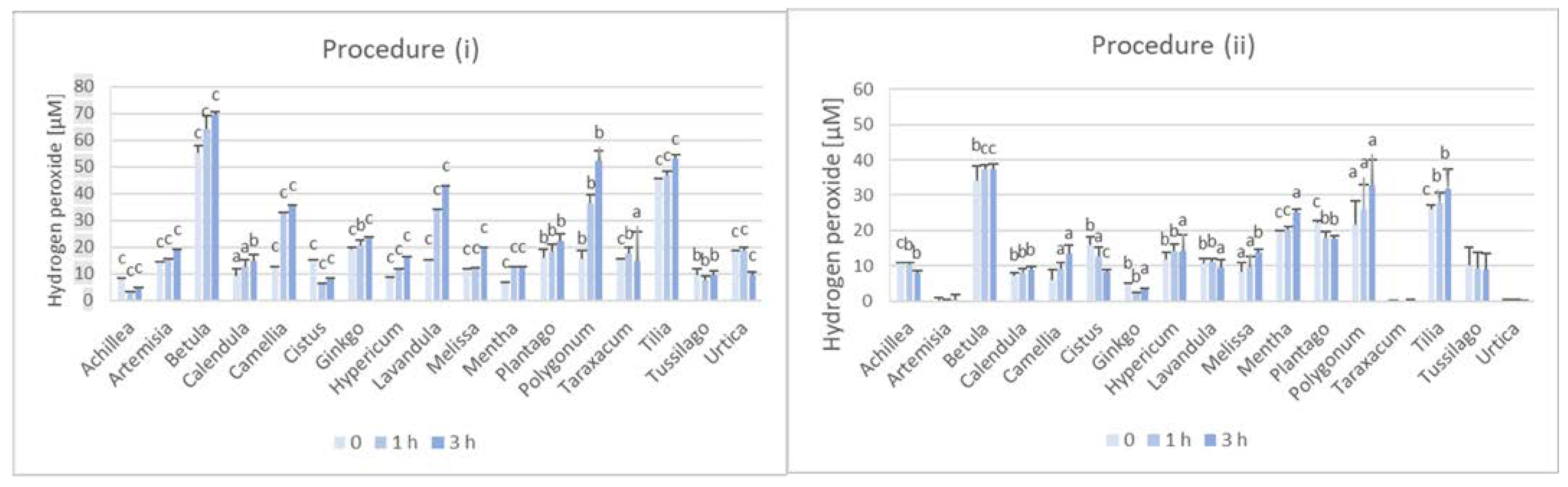
Figure 2.
EPR signals of the semiquinone radical measured in infusions of selected herbs.
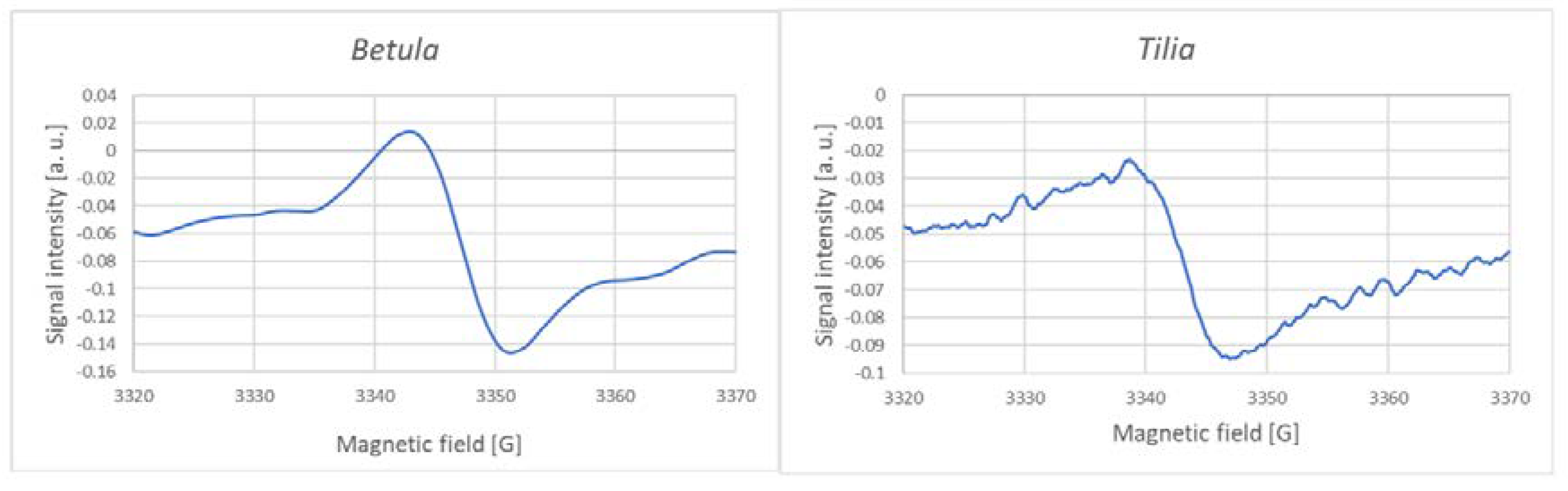
Figure 3.
Superoxide dismutase inhibition of NBT reduction (A) and DHE oxidation (B) in plant infusions. ap < 0.05, bp < 0.01 (with SOD vs no SOD).
Figure 3.
Superoxide dismutase inhibition of NBT reduction (A) and DHE oxidation (B) in plant infusions. ap < 0.05, bp < 0.01 (with SOD vs no SOD).
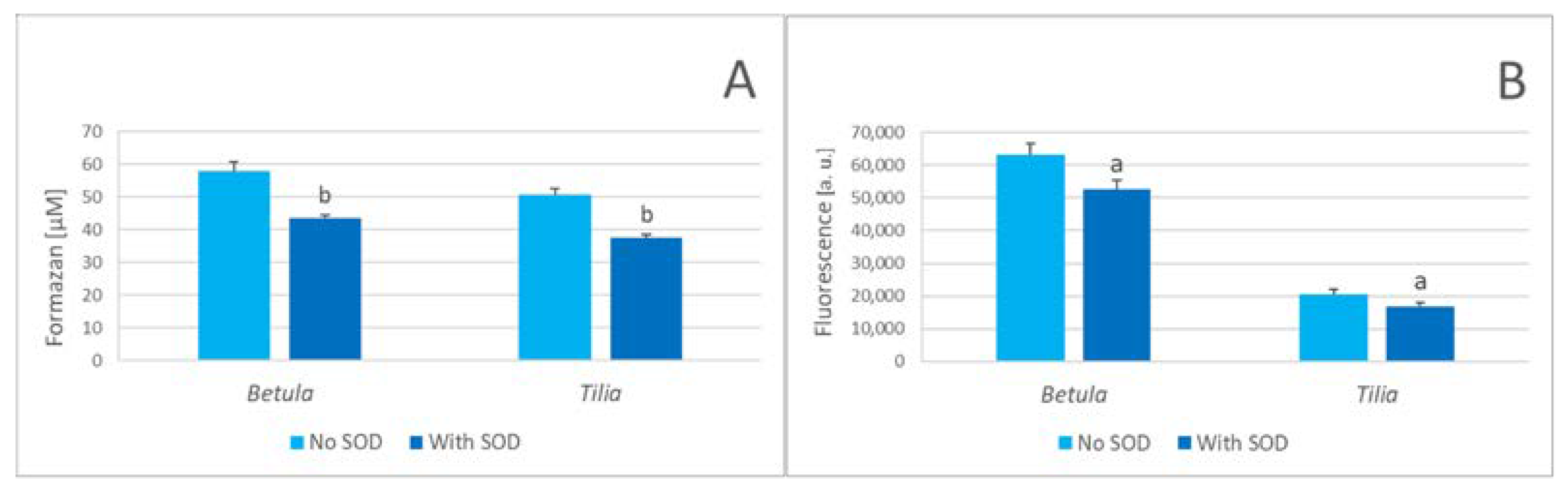
Figure 4.
Time course of hydrogen peroxide concentration in herbal infusions not supplemented with H2O2 and added with 25 μM (final) H2O2. Expected, H2O2 concentration calculated on the basis of concentration in non-supplemented infusions plus 25 μM. AP < 0.05, CP < 0.001 (supplemented vs not supplemented infusions), ♣P < 0.05, ♦P < 0.01, ♥P < 0.001 (supplemented with H2O2, measured vs expected).
Figure 4.
Time course of hydrogen peroxide concentration in herbal infusions not supplemented with H2O2 and added with 25 μM (final) H2O2. Expected, H2O2 concentration calculated on the basis of concentration in non-supplemented infusions plus 25 μM. AP < 0.05, CP < 0.001 (supplemented vs not supplemented infusions), ♣P < 0.05, ♦P < 0.01, ♥P < 0.001 (supplemented with H2O2, measured vs expected).
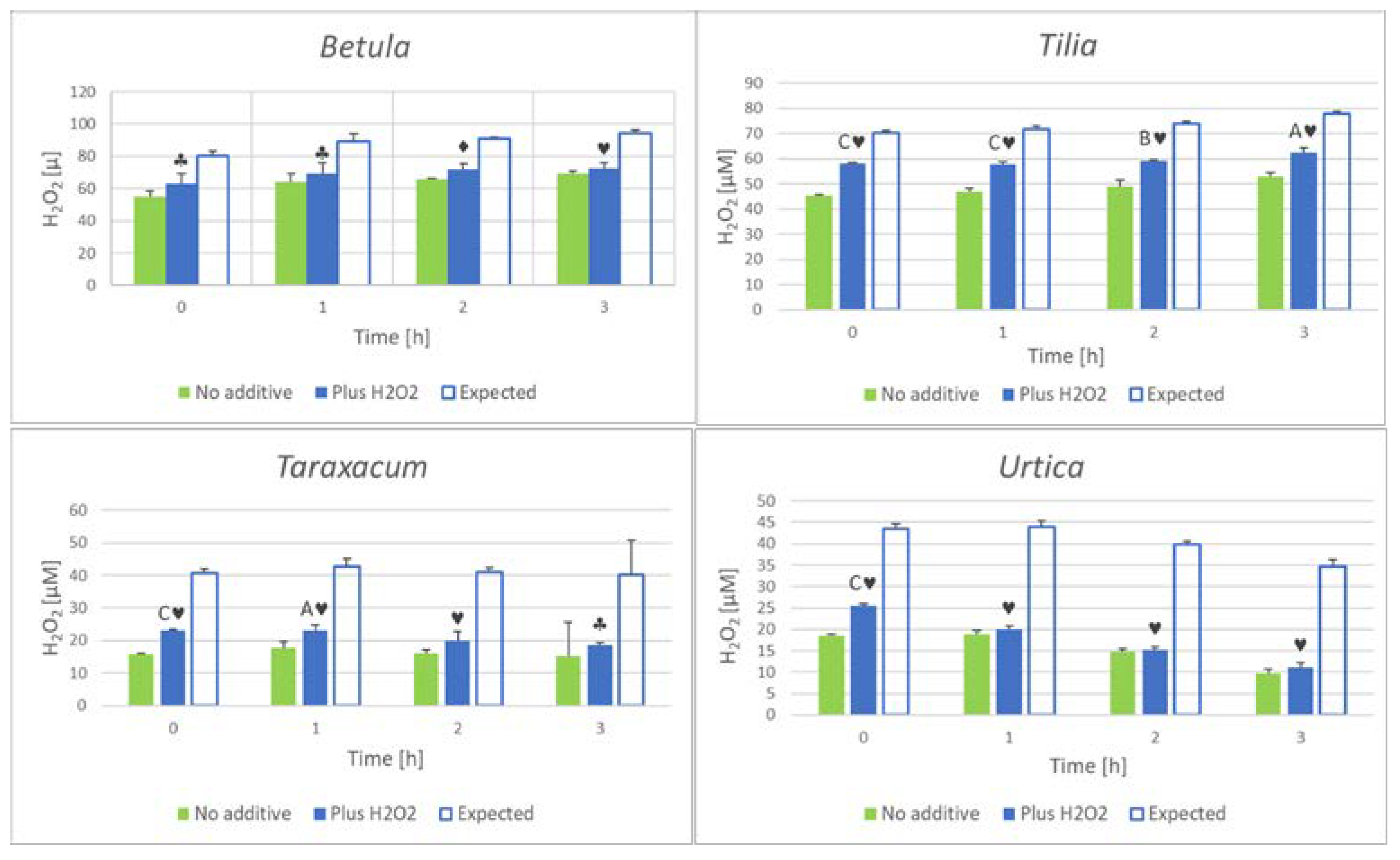
Figure 5.
Survival of SKOV-3 and PEO1 ovary cancer cells, and MRC-5 fibroblasts treated with various volumes of herbal infusions after 24-h incubation; aP <0.05.
Figure 5.
Survival of SKOV-3 and PEO1 ovary cancer cells, and MRC-5 fibroblasts treated with various volumes of herbal infusions after 24-h incubation; aP <0.05.
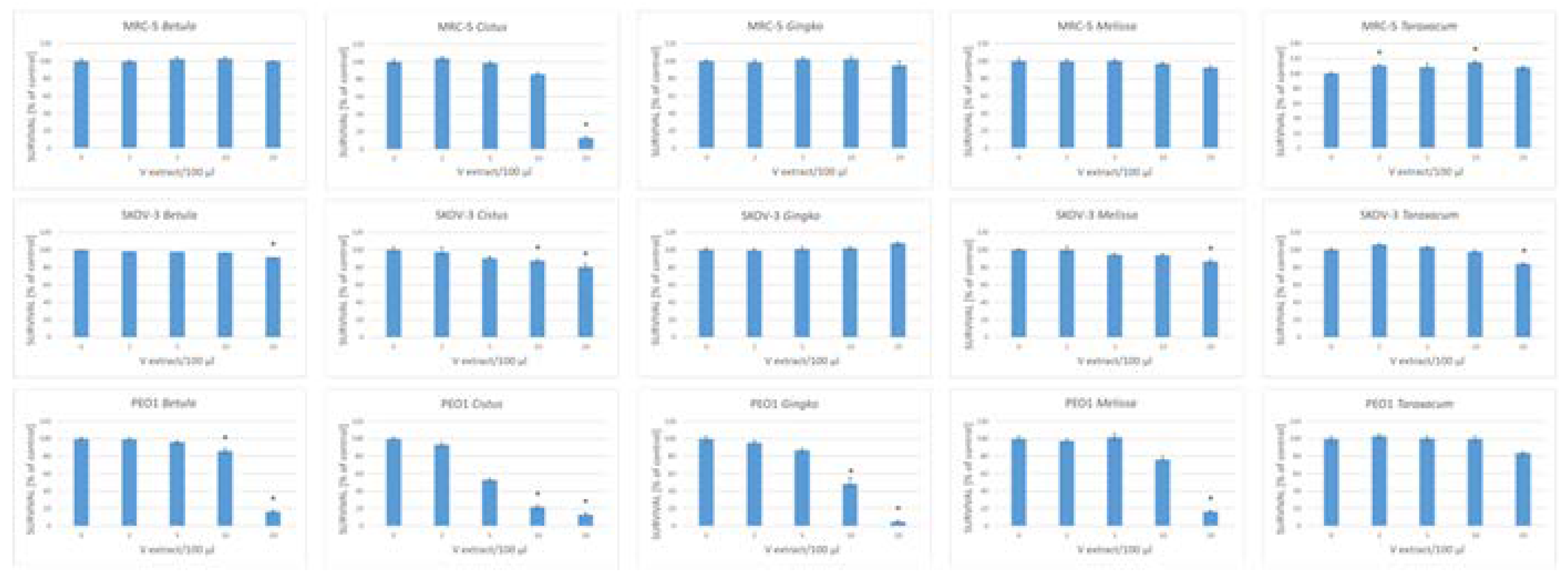
Figure 6.
Effect of catalase (10 µg/mL) on the survival of PEO1 cells treated with various volumes of infusions of Betula, Cistus, and Gingko infusions. aP < 0.05, bP < 0.01, cP < 0.001 (catalase vs no catalase).
Figure 6.
Effect of catalase (10 µg/mL) on the survival of PEO1 cells treated with various volumes of infusions of Betula, Cistus, and Gingko infusions. aP < 0.05, bP < 0.01, cP < 0.001 (catalase vs no catalase).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



