
Submitted:
11 May 2023
Posted:
16 May 2023
You are already at the latest version
Abstract
Breast cancer is a complex and heterogeneous disease that displays diverse molecular subtypes and clinical outcomes. Although it is known that the location of tumors can affect their biological behavior, the underlying mechanisms are not fully understood. In our previous study, we found a differential methylation profile and membrane potential between left (L) and right (R)-sided breast tumors. In this current study, we aimed to identify the ion channels responsible for this phenomenon and determine any associated phenotypic features. To achieve this, experiments were conducted in mammary tumors in mice, human patient samples, and with data from public datasets. The results revealed that L-sided tumors have a more depolarized state than R-ones. We identified a 6-ion channel-gene signature (CACNA1C, CACNA2D2, CACNB2, KCNJ11, SCN3A, and SCN3B) associated with side: L-tumors exhibit lower expression levels than R-ones. Additionally, in silico analyses show that the signature correlates inversely with DNA methylation writers and with key biological processes involved in cancer progression, such as proliferation and stemness. The signature also correlates inversely with patients’ survival rates. In an in vivo mouse model, we confirmed that KI67 and CD44 markers were increased in L-sided tumors and similar tendency for KI67 was found in patient L-tumors. Overall, this study provides new insights into the potential impact of anatomical location on breast cancer biology and highlights the need for further investigation into possible differential treatment options.
Keywords:
Laterality
; Ion Channel
; Breast Cancer
; Proliferation
; Stemness
INTRODUCTION
Ion channels (ICH) are transmembrane proteins that regulate the flow of ions across cell membranes, thereby modulating membrane potential, intracellular signaling, and gene expression. Rather than being confined to the nervous systems alone, these abilities are shared by all cells and are considered adaptations of an ancient bioelectric communication system that employed similar strategies to tackle challenges in the past [1]. The ability of cell groups to approach a specific morphology is largely reliant on bioelectric communication across tissues. This information processing occurs through the utilization of identical, well-conserved molecular components as those used by neurons [2]. ICH play crucial roles in many physiological processes, including cell proliferation, migration, invasion, and apoptosis[3]. Dysregulation of ICH can lead to altered membrane potential, intracellular signaling, and gene expression, promoting tumorigenesis and therapy resistance. Any bioelectric disruption can trigger cancerous characteristics [4,5], even in the absence of DNA damage or mutation. Essentially, when cells lose their tight informational connections with neighboring cells, they revert back to a primitive, unicellular state[6], breaking away from the multicellular group and regarding it as a mere external environment. This results in the reemergence of behaviors appropriate to the unicellular state, such as proliferation. So cancer can be seen as primarily a disorder of establishing informational boundaries between the cellular self and the outside world [7], which has already led to the development of effective treatments that can reverse and prevent cancer in amphibians[8] and are currently undergoing testing in human tissues [9]. Emerging evidence suggests that ICH also participate in the crosstalk between the TME and cancer cells[10,11], contributing to tumor initiation, progression, and therapy resistance (reviewed in [12,13,14,15]). For example, ICH expressed by cancer cells can sense and respond to changes in the TME, such as alterations in pH, oxygen tension, and ion concentration, leading to changes in intracellular signaling and gene expression that promote tumorigenesis[3]. Moreover, ICH expressed by TME components, such as immune cells and endothelial cells, can modulate their function and secretion of cytokines and growth factors, affecting tumor cell behavior[16,17]. Targeting ICH that are involved in the TME-tumor interaction may provide new opportunities for developing effective and specific anticancer therapies.
Breast cancer (BC) is a complex disease that exhibits heterogeneity with diverse subtypes that vary in their molecular and clinical characteristics. One aspect of this heterogeneity is the laterality of breast tumors, i.e., whether the tumor arises in the left(L) or right(R) breast[18]. Although breast tumors are generally thought to be similar regardless of laterality, several studies have suggested that the surrounding tissue of L-R breast tumors may differ [19]. The mammary glands are not identical [16,17,20]; each breast has its unique set of ducts, lobules, and supporting stroma. These structures can differ in size, shape, and branching patterns. In fact, all paired organs present physiological and molecular differences, as shown by microRNA profiling in paired eyes, lung and testes normal tissues[21]. Therefore, the normal differences between both breasts can lead to potential differences in the microenvironment surrounding tumors impacting tumor biology differently in each breast, making it worthwhile to consider when studying BC.
Interestingly, L-sided BC incidence is slightly increased[22], and even though the underlying causes are unclear[23], the tendency is consistent across populations. Similar differences are observed in other paired organs who laterally differ in cancer incidence and progression[24]. Although these differences seemed not to have a relevant clinical impact [25], growing evidence from research findings is indicating they are more important than it was considered[26]. For example, in animal studies using mouse models of BC, it has been shown that L-R-sided tumors had distinct gene expression profiles [27]. In humans, recent studies have discovered that L-R-sided tumors may respond differently to specific therapies depending on the local TME, with tumors in different regions of a single organ showing different expression profiles and properties [28,29]. Furthermore, L-sided BC has been found associated with a more aggressive biology and worse outcome as compared to R-BC [30]. Remarkably, recent literature has raised new questions about the relationship between BC laterality and prognosis[31]. Understanding these differences may have significant implications for diagnosis and treatment.
In our previous publications [32,33], we discovered a distinct methylation profile between L-R-sided breast tumors, which we associated with differences in membrane potential and ion concentration. Integrating the facts that L-R normal breast tissues have differences in gene expression and that the bioelectricity is known to play a crucial role in tumorigenesis, we hypothesized that bioelectric differences of both sides of the breast could contribute to the development of BC in a site-specific manner. In our current work, we sought to determine the primary ICH involved in these differences and explore the potential impact on the functional characteristics of the tumor cells. By investigating the role of specific ICH in L-R tumor development, we hope to uncover novel therapeutic targets for BC treatment.
METHODS
Human Primary Breast Tumors
RNA and tumor tissue from our patient’s tumor tissue bank was used for this study. Informed consent had been originally obtained from all subjects’ tissue included in the bank [34]. The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of the School of Medical Sciences, National University of Cuyo, Mendoza, Argentina. (Protocol code: 15856/2016, date of approval: September 16th 2016).
Mouse Model MMTV-PYMT
All animal procedures were reviewed and approved by the Tufts University Institutional Animal Care and Use Committee. Five MMTV-PyMT (002374, Jackson Laboratory, Bar Harbor, ME, USA) female mice were housed together in the same cage in an on-site housing facility, with access to standard food, water, and a 12/12 light cycle. At 13 weeks of age, the MMTV-PyMT mice were euthanized using CO2, and paired L-R breast tumors from 5 mice, all from the 4th gland, were excised. Tumors were fractionated into 5 pieces for measuring membrane potential by confocal microscopy and flow cytometry, and for DNA and RNA extraction.
Membrane Potential Measured by Flow Cytometry
Tumors were desegregated in a Petri dish with 1-2ml of disaggregation solution containing 1ml Collagenase 2mg/ml in PBS, 150µl of Hyaluronidase 10mg/ml in PBS (Sigma H4272), 100µl Antibiotic (Penicillin-Streptomycin-Glutamine 100X, #10378016) and 8,75ml of DMEM/F12 media (Gibco, #21041025) with FBS 2% (SH30071.03, Cytiva, Marlborough, MA). Tumors were cut in small pieces using vesture and all the remaining fat/normal mammary gland/skin was removed. The mix of tumor tissue and disaggregation solution was transposed to a tube and digested at 37°C in agitation for 2-4hs. Cells were filtered with 40µm cell strainers, then centrifuged for 5 min at 10000 RPM and washed 4 times with cold PBS. Afterwards, cells were resuspended in 200µl DMEM FluoroBrite (Gibco, #A1896701) with 2% FBS. DiBAC4(3) 2µM (positively charged, DB) (Invitrogen, #B438) and Mytotracker Deep Red FM 500nM (negatively charged, MT) (Cell Signaling, #8778) was added and incubated for 30min at 37°C. We used the MT probe instead of Rhodamine6G (which is a potentiometric dye [35]), because they have similar functionality and to preclude Rhodamine6G’s spectral overlap with DIBAC4(3). Fluorescence was measured by flow cytometry (Attune NxT flow cytometer®) using a 530/30 emission filter for DB measurement and a 670/14 emission filter for MT measurement. The autofluorescence of each tumor cell was measured and subtracted. Results were analyzed using FlowJo v X.0.7® software (RRID: SCR_008520).
Membrane Potential Measured by Confocal Microscopy
Thin slices of the paired L-R 4th gland tumors were cut and placed in Petri dishes with DMEM FluoroBrite (Gibco, #A1896701) plus 2% FBS. For measuring membrane potential, 2-3 slices per tumor were used, incubating 15 minutes at 37°C with media containing DB 2 μM and MT 500 nm in Fluorobrite DMEM. One slice was used to measure autofluorescence of the cells. Images were captured using a Leica Stellaris Sp8 confocal microscope at 10X magnification, at 37°C. 3-5 images were taken per tumor, with 3 z-slices per image. As control for normalized measurements, cells were treated with a high concentrated KCl (65 mM) as depolarizing agent, for 15 minutes at 37°C. Afterwards, slices were incubated with Fluorobrite DMEM containing DB (2 μM) and MT (500 nM). The images were processed using Image J software (National Institutes of Health, USA, RRID: SCR_003070). The sum of the z-slices was used for image projection. A ratio of MT/DB Integrated Density was calculated for each image, and this ratio was normalized to the same ratio obtained from the depolarized control images.
RNA Extraction
RNA was extracted from mice tumors using the Quick-RNA Miniprep Plus Kit (Zymo Research, #R1057), following the manufacturer's protocol. RNA from human breast tumors was extracted with a Trizol (Life Technologies, USA) based protocol.
Gene Expression Analysis in Mice Tumors by Droplet Digital PCR (ddPCR)
Purified RNA from mice tumors was converted to cDNA using M-MLV Reverse Transcriptase (Inbio #K1600, Argentina), and cDNA was stored at −20 °C until use. For each reaction, 1ng was used plus ddPCR Supermix for Probes (No dUTP) (BioRad, #1863023 ) and PrimeTime® Mini qPCR Assay probes for KI67, CD44 and HPRT (housekeeping) genes with FAM fluorescence for detection. Sequences for KI67 probe: 5'-/56-FAM/TGGCCTACC/ZEN/TGGTCTTAGTTCCGT/31ABkFQ/-3', primer 1: 5'-TTCCTTCAGCAAGCCTGAG-3', primer 2: 5'-CTTCATAGGCATTCCCTCACTC-3'. For CD44 probe: 5'-/56-FAM/ACCCATACC/ZEN/TGCATGTTTCAAAACCC/31ABkFQ/-3', primer 1: 5'-GCTTTCAACAGTACCTTACCCA-3', primer 2: 5'-GGATGAATCCTCGGAATTACCA-3'. For HPRT probe: 5’-/56-FAM/CTTGCTGGT/ZEN/GAAAAGGACCTCTCGAA/31ABkFQ/-3, primer 1: 5’-CCCCAAAATGGTTAAGGTTGC-3', primer 2: 5’-AACAAAGTCTGGCCTGTATCC-3’. We used the following probe concentrations to improve the multiplexed reaction: probe CD44 1.5X with HPRT 1X; and probe KI67 1X with HPRT 2X. For each assay, droplets were generated using Droplet Generation Oil for probes (Bio-Rad, #1863005) on the QX200 Droplet Generator (Bio-Rad) according to the manufacturer’s protocol adding the specific primers. Droplets were cycled on the C1000 Touch Thermal Cycler (Bio-Rad) for 40 cycles, with a 58°C annealing temperature. Droplets were read using the QX200 Droplet Reader (BioRad). Data was analyzed with QuantaSoft software (BioRad).
Gene Expression Analysis in Human Tumors by qPCR
1µg of total RNA was used for the synthesis of cDNA using M-MLV retro-transcriptase (Inbio #K1600, Argentina) and Random Hexamers (Qiagen #79236, USA) primers. 15ng were used to perform Real-Time PCR using specific primers for CD44, KI67 and β-actin (housekeeping) in an AriaMx Real-time PCR System (Agilent, CA, USA). The sequences of the primers for CD44 were: forward 5-TGCCGCTTTGCAGGTGTATT-3 and reverse 5-CCGATGCTCAGAGCTTTCTCC-3; for KI67: forward 5-TGACCCTGATGAGAAAGCTCAA-3 and reverse 5-CCCTGAGCAACACTGTCTTTT-3; for β-actin: forward 5-TGACGTGGACATCCGCAAAG-3 and reverse 5-CTGGAAGGTGGACAGCGAGG-3. For the reaction, a Master Mix qPCR 2X with SYBR was used (Inbio, Argentina). The amplification program consisted of 40 cycles of 15s at 95°C, 30s at 60°C and 30s at 72°C, followed by a final melting curve step. The analysis was performed using AriaMx Software. Relative expression normalization of genes of interest was carried out using β-actin gene expression as endogenous reference control by the ΔCq method.
Public Datasets and Platforms Used
Human ICH genes were obtained from the HUGO Gene Nomenclature Committee (HGNC) database (https://www.genenames.org/data/genegroup/#!/group/177).
Gene expression and laterality data of primary breast tumors was downloaded from The Cancer Genome Atlas (TCGA)- Xena Functional Genomics Explorer-UCSC (https://xenabrowser.net/) (TCGA-Breast Cancer dataset) and exploration tools for Differential Gene Expression (DGE), Enrichment Analyses and Kaplan Meier Curves were used.
Dataset Enrichment Score analyses to compare L-R tumors were performed using the computational method Gene Set Enrichment Analysis (GSEA). GSEA allows to determine whether pre-defined gene sets show significant differences between two phenotypes (in this case, L and R-sided tumors). Based on gene expression data from these two biological states, GSEA calculates and ranks the DGE, searches for the gene sets in the Human Molecular Signature Database (MSigDB) containing 33,591 gene sets and identifies the top ones that significantly contribute to the differences between the phenotypes. Leading-edge analysis using the GSEA tool, were used to find associated gene signatures.
Statistics
Statistical comparisons were performed using GraphPad Prism version 5.00 for Windows, GraphPad Software, San Diego California USA, www.graphpad.com. Un-paired or paired Student T test were used according to the experiment type, to compare means of fluorescence ratios. Data from different experiments was normalized to a maximum depolarized state by using high concentrated KCl. All results are means of 3 independent experiments with 2-3 technical replications each. Correlation analyses between two variables were performed using Spearman rank coefficient calculations. Sensitivity/specificity of the 6-ICH signature to distinguish between high vs low proliferation and stemness markers were calculated using the receiver operating characteristic curve (ROC). Overall survival, disease free survival, progression free survival and disease specific survival rates were analyzed by Kaplan Meier plots. P values <0.05 were considered statistically significant, using FDR correction (<0.1) when needed for repetitive comparisons.
RESULTS
Left-Sided Mice Tumors Exhibit Greater Depolarization
We chose to use a mouse model of breast cancer (MMTV-PyMT) which spontaneously develops mammary tumors on both the L and R side, to investigate the role of laterality in regulating the bioelectric properties of tumors. Pairwise comparisons of L-R tumors within the same mouse were possible using this model. After extracting paired tumors, we used two fluorescent probes with opposite charges to measure the bioelectric state establishing a ratio between both probes’ fluorescence (MT/DB). The paired comparison of 4 tumors, all located on the L-R 4th gland, revealed by confocal microscopy (Figure 1A, Supplementary Figure 1A and 1B) a difference in the MT/DB ratio: L-sided tumors showed a decreased MT/DB ratio (Figure 1B, Supplementary Files 1A-1D).
In parallel, samples from 3/5 mice were analyzed by flow cytometry, confirming the same trend as seen by microscopy: L-sided tumors had a decreased MT/DB ratio compared to R-sided tumors (Figure 2, Supplementary File 2).
This suggests that the L and R glands generate a difference in the bioelectric state of growing tumor cells. The consistent shift into the same direction suggests that L-sided tumors have a more depolarized state than R.
Bioinformatic Approach: ICH Gene Signature Involved in L-R Bioelectric Differences
Ion channels play a major role in regulating the bioelectrical properties of tumors. To further investigate which ICH may play a relevant role, we switched to a bioinformatic approach. We obtained a list of human ICH genes from the HGNC database. The list contained 330 genes, of which the expression levels of 319 were downloaded from 781 invasive breast tumors in The Cancer Genome Atlas (TCGA) dataset, comprising 393 L-sided and 388 R-sided tumors.
To determine whether a set of ICH genes contributed in a relevant way to the L-R differences, we used the computational method Gene Set Enrichment Analysis (GSEA). We selected the four gene sets that showed significant difference, particularly a consistent decrease of their expression in L-sided tumors (p < 0.05; FDR < 0.1): WP-ENERGY-METABOLISM (collection: C2), REACTOME-PHASE-0-RAPID-DEPOLARISATION (collection: C2), REACTOME_CARDIAC-CONDUCTION (collection: C2), and CANCER-MODULES (collection: C4). By combining these gene sets, we identified the leading genes that were associated with the tumor laterality: CACNA1C, CACNA2D2, CACNB2, KCNJ11, SCN3A, and SCN3B, signature that we called: 6-ICH signature.
Next, we analyzed the 6-ICH signature in 1,097 TCGA breast tumors using the Xena Functional Genomics Explorer from UCSC (571 left-sided, 526 right-sided). We obtained the expression levels of the signature from an equation where we assumed an equal contribution of each gene, i.e.,
=CACNA1C+CACNA2D2+CACNB2+KCNJ11+SCN3A+SCN3B.
We found a decreased expression of the signature in L-sided tumors compared to R-sided ones (Mann Whitney test for non-normal data, p=0.005, Supplementary File 3). Adding more genes to the signature did not significantly increase the association with laterality. However, removing genes one by one resulted in a loss of association. Interestingly, when we compared the expression levels of the individual genes of the 6-ICH signature in a one-by-one manner, we found a reduced or complete loss of association with side for some genes (data not shown). Therefore, we considered the 6-ICH signature as robust for further studies.
In silico: the 6-ICH signature inversely correlates with pro-mitotic and stemness markers, and survival rates.
ICH play a role in a wide range of biological processes and can regulate opposing cellular phenotypes. Therefore, the 6-ICH signature did not suggest a clearly defined phenotypic impact. To gain insight, we used a bioinformatic approach. We dichotomized the 6-ICH signature expression data into High and Low, using the mean expression value as a cut-off, and performed a Differential Gene Expression (DGE) analysis visualized in a Volcano Plot. In the Low-expressing group (as compared to the High-expressing one), a significant number of genes were up or down regulated (Figure 3A).
Next, we conducted an Enrichment Analysis to determine the principal biological processes the DGE were participating in. Interestingly, we found that in the Low-expressing subgroup, mitotic genes were overexpressed, and membrane potential regulating processes were less expressed (Figure 3B). These findings suggest that L-sided tumors with a low-expressing 6-ICH signature may exhibit increased mitotic activity compared to R-sided tumors.
From an in-silico cohort of 1,082 primary breast tumors from the Xena Functional Genomics Explorer-UCSC, we randomly generated a Discovery Cohort (533 tumors) and a Validation Cohort (549 tumors). We choose the markers “KI67” and “stemness scores” to analyze, as indicative of pro-mitotic behavior, with KI67 being a protein expressed throughout the cell cycle, and stemness scores measuring the extent to which a tissue is capable of self-renewal. So, we studied the correlation between the expression of the 6-ICH signature and the expression of KI67 and an RNA-seq based stemness score based [17], both available in the TCGA Pan-cancer Atlas. In the Discovery Cohort, we observed a strong inverse correlation between the 6-ICH signature and both markers, (r=-0.47 and r=-0.62 respectively, Spearman correlation test, p<0.0001) (Figure 4, panel A). Receiver operating characteristic curves (ROC analyses) demonstrated an acceptable discrimination value of the 6-ICH signature to distinguish between high and low expression of KI67 (AUC=0.74, SE=0.02, 95%CI=0.698 to 0.782, p<0.0001) and high or low stemness scores (AUC=0.79, SE=0.01, 95%CI=0.761 to 0.837, p<0.0001). We confirmed these results in the Validation Cohort of 549 primary breast tumors (r=-0.51 and r=-0.62 respectively, Spearman correlation test, p<0.0001; ROC curves for KI67: AUC=0.74, SE=0.02, 95%CI=0.707 to 0.788, and for CD44: AUC=0.81, SE=0.01, 95%CI=0.776 to 0.848, p<0.0001) (Figure 4, panel B). We also confirmed these correlations in a pan-cancer analysis on 9,546 tumors (r=-0.51 and r=-0.62 respectively, Spearman correlation test, p<0.0001) (Figure 4, panel C).
We then analyzed the 5-year (1,800 days) rates of overall survival, disease-specific survival, disease-free survival, and progression-free survival by comparing high vs low 6-ICH signature expression in 1,094 BC and in 9,546 pan-cancer patients. Remarkably, all survival rates were increased in high-expressing tumors (Figure 4, panel D).
We also examined the correlation between the 6-ICH signature and epigenetic regulators, particularly histone modifiers, methylation erasers, and methylation writers, by performing an expression signature of each group of epigenetic enzymes. We found a strong inverse correlation only between the 6-ICH signature and the signature of DNA methylation writers composed of DNMT1, DNMT3A, DNMT3B, DNMT3L, TRDMT1, and DMAP1 (r=-0.42, Spearman correlation test, p<0.0001) (Figure 5, Supplementary File 4).
Taken together, these results suggest that the bioelectric signature is associated with the side of the tumor and with cancer hallmarks such as proliferation and stemness markers and is also associated with regulators of DNA methylation (which are known to respond on microenvironmental factors). Moreover, it reveals an impact on the survival rate of cancer patients. The results are robust across different cohorts.
In vivo: the 6-ICH signature inversely correlates with proliferation marker and stemness scores.
After establishing in silico that the 6-ICH signature was associated with side and with proliferation and stemness divergences, we progressed to in vivo models in mice to confirm these findings. We utilized paired L-R tumors of 4 female mice and analyzed the expression of the proliferation marker KI67 and the stemness marker CD44.
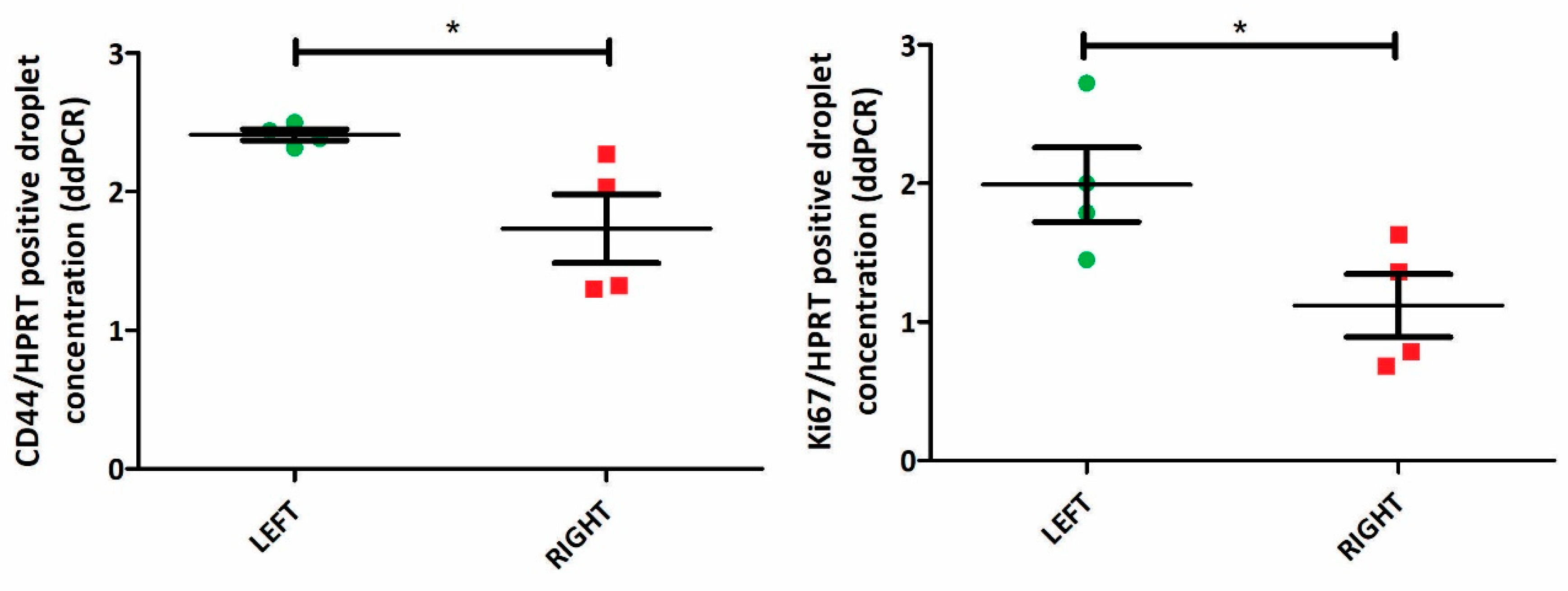
Our analyses revealed a significant increase in the expression of both markers in L-sided tumors compared to R-sided tumors (paired T-test, p<0.05) (Figure 6, Supplementary File 5).
We also investigated whether these differences were present in human breast tumor RNA samples. We analyzed the expression of the markers KI67 (in 6 L and 5 R) and CD44 (in 5 L and 4 R) by qPCR. Although not statistically significant, it is notable that we found a similar trend: an increased mean expression of both markers in L-sided tumors (unpaired t-test, p=0.2) (Supplementary File 6).
Taken together, our observations in mice and humans strongly suggest that L-R breast tumors are distinct in two critical cancer hallmarks, namely proliferation and stemness, which are shaped by the differential expression of ICH.
DISCUSSION
Breast cancer is a heterogeneous disease that encompasses a broad spectrum of molecular subtypes and clinical outcomes[36]. The association between the anatomical location of tumors and their biological behavior is still an open question. Diverse studies across different species have addressed the L-R issue of bilateral organs, like RNA differential profiles in mice paired eyes, lungs and testes[21], L-R differences acquired during embryonic development [16], L-R metabolic differences in Xenopus[37], in avian [38] and mice[39] models, and in humans[40]. It has been known for many decades that breast tumors occur more frequently in L sides[41], however the causes are still unclear[22,25,42]. In our previous research, we addressed the question of whether epigenetic differences existed between L-R human and mice-xenografted breast tumors. We found differential DNA methylation profiles in genomic regions[32], among which ICH genes were included. Furthermore, we found how L and R extracts from human mammary glands were able to generate membrane potential differences in cultured cells and how L-extract treated cells developed a more depolarized state [43]. So, we proposed that the different L-R environments generated methylation differences which ended up provoking different bioelectric states in the L-R tumors. In this study, we aimed to address the L-R asymmetry question at a whole organism level, in mice and in humans. Our results show that L-sided tumors consistently exhibit a more depolarized state, increased expression of proliferation and stemness markers, and that the lower level of ICH signature expression in L-tumors is associated with reduced survival rates, as compared to the R-sided ones. This is suggesting that the microenvironments of the L and R sides generate a consistent difference in the bioelectric state of growing tumor cells, with a direct impact on tumor behavior. To our knowledge, prior to our research, there were no published studies informing proliferation differences between L and R breast tumors. However, a recent publication supports our findings, demonstrating (in line with our observations) that L-sided BC is associated with more aggressive biology and worse outcome as compared to R BC [30]. As far as we know, these are the first conclusive findings differentiating breast tumors in bioelectric, proliferative and survival rate terms, based on their side.
We identified a 6-ICH gene expression signature that was consistently associated with the tumor side. The signature is composed of 3 calcium, 2 sodium and 1 potassium channel. CACNA1C, CACNA2D2, and CACNB2, are all subunits of voltage-gated calcium channels, which are involved in a wide range of physiological processes, including muscle contraction, neurotransmitter release, and gene expression [44]. KCNJ11, on the other hand, codes for a potassium channel that is involved in regulating insulin secretion in pancreatic beta cells [45]. Bioelectric signaling driven by potassium channels have been proposed as metastasis regulators in triple-negative breast cancer [46]. Finally, SCN3A and SCN3B are subunits of voltage-gated sodium channels, which are important for the initiation and propagation of action potentials in neurons and other excitable cells [47]. Recent findings have revealed the clinical importance of sodium channels and the ionic microenvironment of breast tumors [48]. Predicting the outcome of a gene expression shift in these 6 ICH entities, however, is challenging. For example, reduced expression of CACNA1C, CACNA2D2, and CACNB2 in cardiac myocytes could result in decreased calcium influx and reduced contractility, which could lead to a hyperpolarized state. And reduced expression of SCN3A and SCN3B in neurons could result in decreased sodium influx and reduced excitability, which could also lead to a hyperpolarized state. However, reduced expression of KCNJ11 in pancreatic beta cells could lead to decreased potassium efflux and increased insulin secretion, which could result in a depolarized state. Overall, the impact of changed expression of ICH on membrane potential and cellular excitability is complex and context-dependent and requires consideration of the specific ion channel and cell type involved. Therefore, the 6-ICH signature did not suggest a clearly defined phenotypic impact.
By bioinformatic analyses in breast tumors, the lower 6-ICH expression revealed to be associated with increased proliferation and stemness, and with worst survival in 5 years. Proliferation and stemness are key biological processes in tumorigenesis. Importantly, higher values for stemness indices are associated with increased number of cancer stem cells and with a greater dedifferentiated phenotype [49]. Stemness is defined as the potential of a cell for self-renewal and differentiation. Cancer progression involves gradual loss of differentiated phenotype and acquisition of stem cell-like features. Less differentiated tumors are more likely to spread to distant organs and make prognosis worse, particularly because metastases are usually resistant to available therapies[50]. Proliferation is also a critical aspect to consider when evaluating BC, and KI67 expression stands out as one of the most important and cost-effective surrogate markers for assessing tumor cell proliferation[51] [52]. Our results suggest that the 6-ICH signature may play a role in modulating these processes in breast cancer, probably by changing the bioelectric state in a side-dependent manner.
The tumor microenvironment (TME) is a complex and dynamic ecosystem that plays a critical role in tumor initiation, progression, and response to therapy. It comprises various cell types, extracellular matrix components, and signaling molecules that interact with tumor cells in a bidirectional manner[53]. Growing evidence suggests that the TME can alter the epigenetic landscape of cancer cells, leading to changes in gene expression patterns that promote tumorigenesis and therapy resistance. For example, TME-derived factors, such as growth factors, cytokines, and extracellular matrix proteins, can activate oncogenic signaling pathways that promote DNA methylation, histone modifications, and chromatin remodeling in cancer cells[54]. Moreover, TME components, such as cancer-associated fibroblasts, immune cells, and endothelial cells, can secrete exosomes and micro-vesicles that transfer epigenetic modifiers, such as microRNAs, DNA methyltransferases, and histone deacetylases, to cancer cells, further altering their epigenetic landscape[10,11]. The impact of TME on the tumoral epigenome and its effects on tumor behavior are of great interest because epigenetic changes are reversible and potentially targetable by therapeutic agents.
The finding that differences between L-R breast tumors are related to ICHs suggests possible therapeutic opportunities by applying already approved drugs used in other diseases to modulate channels. Dysregulation of ICH has been implicated in various diseases. In recent years, ICH modulating drugs have gained attention as potential cancer therapeutics. Drug repositioning is a strategy that involves the repurposing of existing drugs for new therapeutic indications. In the context of cancer treatment, repositioning ICH modulating drugs has emerged as a promising approach to identify new treatments for cancer [55] and the use of ICH drugs in cancer is increasingly being studied, as reviewed in [56]. An oncology-focused drug repurposing database has been developed, comprising drugs that show potential for repurposing in cancer treatment (https://www.anticancerfund.org/en/redo-db). Among the 369 drugs listed, to date, there are 20 that target some of the 6 channels included in our 6-ICH signature, which sounds very encouraging. By repositioning these drugs for cancer treatment, the development timeline and costs associated with drug development can be reduced, making it an attractive option for drug discovery.
Finally, another evidence of our study is that the ICH signature was found inversely associated with DNMTs expression, and not with other epigenetic modulators (e.g., histone deacetylases, acetylases, histone methyl transferases, DNA methyl-erasers). It is now well established that epigenetic dysregulations play pivotal roles in cancer onset and progression, including DNA methylation. To occur, these epigenetic events need to be triggered by genetic alterations or transcription shifts of the epigenetic regulators. In this, the TME plays a fundamental role. Therefore, it is valid to reason that the ICH signature is regulated by DNA methylation, and further studies should be performed to confirm this, by for example inhibiting in-vitro DNMTs function with 5-azacytidine or deoxycytidine.
In conclusion, our study suggests that the anatomical location of breast tumors may influence their bioelectric state, potentially mediated by ICH. The 6-ICH signature identified in our study may play a role in modulating key biological processes involved in cancer progression, and further studies are warranted to validate these findings. The potential impact of anatomical location on BC biology and clinical outcomes should also be further investigated, suggesting differential treatment options with repurposed ICH drugs.
Author Contributions
Conceptualization, SM, MR, MO and ML; Methodology, SM, SR, PM; Formal Analysis, MR; Investigation, SM, MR; Writing – Original Draft Preparation, SM, MR; Writing – Review & Editing, SM, MR, SR, PM, ML, MO; Supervision, MR, MO, ML; Project Administration, SM, MR; Funding Acquisition, MR, MO.
Funding
This work has been possible thanks to the following fundings: National Agency of Scientific and Technological Promotion (FONCTyT-ANPCyT)- PICT-A-2020, Argentina; National Cancer Institute-2016, Argentina; Travel Fellowship from the The Company of Biologist-2021<, UK, DP2CA271387 from NIH to MJO, USA.
Acknowledgements
We thank Dr. Cristian Acosta for his advice on the bioelectrical interpretation of the 6-ICH-signature, Sergio Laurito for his assistance with GSEA software, and Justinne Guarin for her collaboration in animal management at Dr. Oudin's laboratory animal facility.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fields, C.; Levin, M. Competency in Navigating Arbitrary Spaces as an Invariant for Analyzing Cognition in Diverse Embodiments. Entropy (Basel). 2022, 24. [Google Scholar] [CrossRef]
- Mathews, J.; Levin, M. Gap Junctional Signaling in Pattern Regulation: Physiological Network Connectivity Instructs Growth and Form. Dev. Neurobiol. 2017, 77, 643–673. [Google Scholar] [CrossRef] [PubMed]
- Hirata, E.; Sahai, E. Tumor Microenvironment and Differential Responses to Therapy. Cold Spring Harb. Perspect. Med. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Blackiston, D.; Adams, D.S.; Lemire, J.M.; Lobikin, M.; Levin, M. Transmembrane Potential of GlyCl-Expressing Instructor Cells Induces a Neoplastic-like Conversion of Melanocytes via a Serotonergic Pathway. Dis. Model. Mech. 2011, 4, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Morokuma, J.; Blackiston, D.; Adams, D.S.; Seebohm, G.; Trimmer, B.; Levin, M. Modulation of Potassium Channel Function Confers a Hyperproliferative Invasive Phenotype on Embryonic Stem Cells. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 16608–16613. [Google Scholar] [CrossRef] [PubMed]
- Trigos, A.S.; Pearson, R.B.; Papenfuss, A.T.; Goode, D.L. Altered Interactions between Unicellular and Multicellular Genes Drive Hallmarks of Transformation in a Diverse Range of Solid Tumors. Proc. Natl. Acad. Sci. U. S. A. 2017. [Google Scholar] [CrossRef]
- Levin, M. Bioelectrical Approaches to Cancer as a Problem of the Scaling of the Cellular Self. Prog. Biophys. Mol. Biol. 2021, 165, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Chernet, B.T.; Levin, M. Transmembrane Voltage Potential Is an Essential Cellular Parameter for the Detection and Control of Tumor Development in a Xenopus Model. Dis. Model. Mech. 2013, 6, 595–607. [Google Scholar] [CrossRef]
- Mathews, J.; Kuchling, F.; Baez-Nieto, D.; Diberardinis, M.; Pan, J.Q.; Levin, M. Ion Channel Drugs Suppress Cancer Phenotype in NG108-15 and U87 Cells: Toward Novel Electroceuticals for Glioblastoma. Cancers (Basel). 2022, 14. [Google Scholar] [CrossRef]
- Arcangeli, A.; Crociani, O.; Bencini, L. Interaction of Tumour Cells with Their Microenvironment: Ion Channels and Cell Adhesion Molecules. A Focus on Pancreatic Cancer. A Focus on Pancreatic Cancer. Philos. Trans. R. Soc. B Biol. Sci. 2014. [CrossRef]
- Arcangeli, A. Ion Channels and Transporters in Cancer. 3. Ion Channels in the Tumor Cell-Microenvironment Cross Talk. Am. J. Physiol. - Cell Physiol. 2011. [CrossRef]
- Lanni, J.S.; Peal, D.; Ekstrom, L.; Chen, H.; Stanclift, C.; Bowen, M.E.; Mercado, A.; Gamba, G.; Kahle, K.T.; Harris, M.P. Integrated K+ Channel and K+Cl- Cotransporter Functions Are Required for the Coordination of Size and Proportion during Development. Dev. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chernet, B.T.; Levin, M. Transmembrane Voltage Potential of Somatic Cells Controls Oncogene-Mediated Tumorigenesis at Long-Range. Oncotarget 2014. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.; Kane, A.; Harrison, C.; Levin, M. A Meta-Analysis of Bioelectric Data in Cancer, Embryogenesis, and Regeneration. Bioelectricity 2020. [Google Scholar] [CrossRef] [PubMed]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion Channels and the Hallmarks of Cancer. Trends Mol. Med. 2010. [CrossRef] [PubMed]
- Ma, K. Embryonic Left-Right Separation Mechanism Allows Confinement of Mutation-Induced Phenotypes to One Lateral Body Half of Bilaterians. Am. J. Med. Genet. A 2013, 161A, 3095–3114. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.; Klar, A.J.S.; Ramsdell, A.F. Introduction to Provocative Questions in Left–Right Asymmetry. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150399. [Google Scholar] [CrossRef] [PubMed]
- Perkins, C.I.; Hotes, J.; Kohler, B.A.; Howe, H.L. Association between Breast Cancer Laterality and Tumor Location, United States, 1994-1998. Cancer Causes Control 2004, 15, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Petitprez, F.; Sun, C.M.; Lacroix, L.; Sautès-Fridman, C.; de Reyniès, A.; Fridman, W.H. Quantitative Analyses of the Tumor Microenvironment Composition and Orientation in the Era of Precision Medicine. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef]
- Levin, M.; Thorlin, T.; Robinson, K.R.; Nogi, T.; Mercola, M. Asymmetries in H+/K+-ATPase and Cell Membrane Potentials Comprise a Very Early Step in Left-Right Patterning. Cell 2002. [Google Scholar] [CrossRef]
- Shi, J.; Hu, C.; Zhou, Y.; Cui, C.; Yang, J.; Cui, Q. MicroRNA Profiling in Paired Left and Right Eyes, Lungs, and Testes of Normal Mice. Mol. Ther. Nucleic Acids 2020, 21, 687–695. [Google Scholar] [CrossRef]
- Sughrue, T.; Brody, J.P. Breast Tumor Laterality in the United States Depends upon the Country of Birth, but Not Race. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Titus-Ernstoff, L.; Newcomb, P.A.; Egan, K.M.; Baron, J.A.; Greenberg, E.R.; Trichopoulos, D.; Willett, W.C.; Stampfer, M.J. Left-Handedness in Relation to Breast Cancer Risk in Postmenopausal Women. Epidemiology 2000, 11, 181–184. [Google Scholar] [CrossRef]
- Roychoudhuri, R.; Putcha, V.; Møller, H. Cancer and Laterality: A Study of the Five Major Paired Organs (UK). Cancer Causes Control 2006. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Yu, K. Da; Jiang, Y.Z.; Shao, Z.M.; Di, G.H. The Effect of Laterality and Primary Tumor Site on Cancer-Specific Mortality in Breast Cancer: A SEER Population-Based Study. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Mittwoch, U. Different Gene Expressions on the Left and the Right: A Genotype/Phenotype Mismatch in Need of Attention. Ann. Hum. Genet. 2008, 72, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Robichaux, J.P.; Hallett, R.M.; Fuseler, J.W.; Hassell, J.A.; Ramsdell, A.F. Mammary Glands Exhibit Molecular Laterality and Undergo Left-Right Asymmetric Ductal Epithelial Growth in MMTV-CNeu Mice. Oncogene 2014. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Duanmu, J.; Liu, Y.; Yan, M.; Li, T.; Jiang, Q. Analysis of Multi-Omics Differences in Left-Side and Right-Side Colon Cancer. PeerJ 2021, 9. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, Y.; Dai, Y.; Cheng, J.N.; Gong, Z.; Feng, Y.; Sun, C.; Jia, Q.; Zhu, B. Immune Landscape of Colorectal Cancer Tumor Microenvironment from Different Primary Tumor Location. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Abdou, Y.; Gupta, M.; Asaoka, M.; Attwood, K.; Mateusz, O.; Gandhi, S.; Takabe, K. Left Sided Breast Cancer Is Associated with Aggressive Biology and Worse Outcomes than Right Sided Breast Cancer. Sci. Rep. 2022, 12. [Google Scholar] [CrossRef]
- Al Saad, S.; Al Shenawi, H.; Almarabheh, A.; Al Shenawi, N.; Mohamed, A.I.; Yaghan, R. Is Laterality in Breast Cancer Still Worth Studying? Local Experience in Bahrain. BMC Cancer 2022, 22. [Google Scholar] [CrossRef]
- Campoy, E.M.; Laurito, S.R.; Branham, M.T.; Urrutia, G.; Mathison, A.; Gago, F.; Orozco, J.; Urrutia, R.; Mayorga, L.S.; Roqué, M. Asymmetric Cancer Hallmarks in Breast Tumors on Different Sides of the Body. PLoS One 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, S.; Real, S.; Campoy, E.; Branham, M.T.; Marzese, D.M.; Salomon, M.; De Blas, G.; Arias, R.; Levin, M.; Roque, M. Correction to: When Left Does Not Seem Right: Epigenetic and Bioelectric Differences between Left and Right-sided Breast Cancer (Molecular Medicine, (2022), 28, 1, (15), 10.1186/S10020-022-00440-5). Mol. Med. 2022, 28, 10020. [Google Scholar] [CrossRef]
- Marzese, D.M.; Hoon, D.S.B.; Chong, K.K.; Gago, F.E.; Orozco, J.I.; Tello, O.M.; Vargas-Roig, L.M.; Roqué, M. DNA Methylation Index and Methylation Profile of Invasive Ductal Breast Tumors. J. Mol. Diagnostics 2012, 14. [Google Scholar] [CrossRef]
- Mandalà, M.; Serck-Hanssen, G.; Martino, G.; Helle, K.B. The Fluorescent Cationic Dye Rhodamine 6G as a Probe for Membrane Potential in Bovine Aortic Endothelial Cells. Anal. Biochem. 1999, 274, 1–6. [Google Scholar] [CrossRef]
- Nolan, E.; Lindeman, G.J.; Visvader, J.E. Deciphering Breast Cancer: From Biology to the Clinic. Cell 2023, 186. [Google Scholar] [CrossRef] [PubMed]
- Onjiko, R.M.; Morris, S.E.; Moody, S.A.; Nemes, P. Single-Cell Mass Spectrometry with Multi-Solvent Extraction Identifies Metabolic Differences between Left and Right Blastomeres in the 8-Cell Frog (Xenopus) Embryo. Analyst 2016, 141, 3648–3656. [Google Scholar] [CrossRef]
- Monsoro-Burq, A.H.; Levin, M. Avian Models and the Study of Invariant Asymmetry: How the Chicken and the Egg Taught Us to Tell Right from Left. Int. J. Dev. Biol. 2018. [Google Scholar] [CrossRef]
- Robichaux, J.P.; Fuseler, J.W.; Patel, S.S.; Kubalak, S.W.; Hartstone-Rose, A.; Ramsdell, A.F. Left-Right Analysis of Mammary Gland Development in Retinoid X Receptor A+/- Mice. Philos. Trans. R. Soc. B Biol. Sci. 2016. [Google Scholar] [CrossRef]
- Mittwoch, U. Different Gene Expressions on the Left and the Right: A Genotype/Phenotype Mismatch in Need of Attention. Ann. Hum. Genet. 2008, 72, 2–9. [Google Scholar] [CrossRef]
- Busk, T.; Clemmesen, J. The Frequencies of Left-and Right-Sided Breast Cancer. Br. J. Cancer 1947. [Google Scholar] [CrossRef]
- Levin, M.; Klar, A.J.S.; Ramsdell, A.F. Introduction to Provocative Questions in Left-Right Asymmetry. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2016, 371. [Google Scholar] [CrossRef]
- Sofía, M.; Sebastián, R.; Emanuel, C.; Branham, M.T.; Marzese, D.M.; Matthew, S.; De Blas, G.; Rodolfo, A.; Michael, L.; María, R. When Left Does Not Seem Right: Epigenetic and Bioelectric Differences between Left- and Right-Sided Breast Cancer. Mol. Med. 2022, 28. [Google Scholar] [CrossRef]
- Karunasekara, Y.; Dulhunty, A.F.; Casarotto, M.G. The Voltage-Gated Calcium-Channel Beta Subunit: More than Just an Accessory. Eur. Biophys. J. 2009, 39, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, S.; Raza, S.T.; Rahman, Q.; Mahdi, F. Role of GNB3, NET, KCNJ11, TCF7L2 and GRL Genes Single Nucleotide Polymorphism in the Risk Prediction of Type 2 Diabetes Mellitus. 3 Biotech 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Payne, S.L.; Ram, P.; Srinivasan, D.H.; Le, T.T.; Levin, M.; Oudin, M.J. Potassium Channel-Driven Bioelectric Signalling Regulates Metastasis in Triple-Negative Breast Cancer. EBioMedicine 2022, 75. [Google Scholar] [CrossRef] [PubMed]
- Luiz, A.P.; Wood, J.N. Sodium Channels in Pain and Cancer: New Therapeutic Opportunities. Adv. Pharmacol. 2016, 75, 153–178. [Google Scholar] [CrossRef]
- Leslie, T.K.; Brackenbury, W.J. Sodium Channels and the Ionic Microenvironment of Breast Tumours. J. Physiol. 2023, 601. [Google Scholar] [CrossRef]
- Malta, T.M.; Sokolov, A.; Gentles, A.J.; Burzykowski, T.; Poisson, L.; Weinstein, J.N.; Kamińska, B.; Huelsken, J.; Omberg, L.; Gevaert, O.; et al. Machine Learning Identifies Stemness Features Associated with Oncogenic Dedifferentiation. Cell 2018, 173, 338–354. [Google Scholar] [CrossRef] [PubMed]
- Visvader, J.E.; Lindeman, G.J. Cancer Stem Cells: Current Status and Evolving Complexities. Cell Stem Cell 2012, 10, 717–728. [Google Scholar] [CrossRef]
- Deaton, A.M.; Bird, A. CpG Islands and the Regulation of Transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef]
- Lashen, A.G.; Toss, M.S.; Ghannam, S.F.; Makhlouf, S.; Green, A.; Mongan, N.P.; Rakha, E. Expression, Assessment and Significance of Ki67 Expression in Breast Cancer: An Update. J. Clin. Pathol. 2022; jcp-2022-208731. [Google Scholar] [CrossRef]
- Place, A.E.; Jin Huh, S.; Polyak, K. The Microenvironment in Breast Cancer Progression: Biology and Implications for Treatment. Breast Cancer Res. 2011, 13. [Google Scholar] [CrossRef] [PubMed]
- Flavahan, W.A.; Gaskell, E.; Bernstein, B.E. Epigenetic Plasticity and the Hallmarks of Cancer. Science 2017, 80, 357. [Google Scholar] [CrossRef] [PubMed]
- Pantziarka, P.; Verbaanderd, C.; Sukhatme, V.; Rica Capistrano, I.; Crispino, S.; Gyawali, B.; Rooman, I.; Van Nuffel, A.M.T.; Meheus, L.; Sukhatme, V.P.; et al. ReDO_DB: The Repurposing Drugs in Oncology Database. Ecancermedicalscience 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Awasthi, R.; Malviya, R. Bioelectronic Medicines: Therapeutic Potential and Advancements in next-Generation Cancer Therapy. Biochim. Biophys. acta. Rev. cancer 2022, 1877. [Google Scholar] [CrossRef]
Figure 1.
Differential MT/DB fluorescence ratio in L-R mice breast tumors. Panel A: representative confocal microscopy image of L (upper panel) and R (lower panel) of paired mouse breast tumor tissue, stained with MT (red) and DB (green) fluorescent probes. Fluorescence gradients of each fluorophore are shown. Panel B: Quantification of MT/DB ratio in 4 pairs of L-R mice breast tumors, as measured in different images by fluorescence microscopy. The L-mean of MT/DB fluorescence is lower than the R-one (unpaired T-test, p=0,01). Circles = Mouse 2, Triangles = Mouse 3, Squares= Mouse 4, Inverted Triangles = Mouse 5.
Figure 1.
Differential MT/DB fluorescence ratio in L-R mice breast tumors. Panel A: representative confocal microscopy image of L (upper panel) and R (lower panel) of paired mouse breast tumor tissue, stained with MT (red) and DB (green) fluorescent probes. Fluorescence gradients of each fluorophore are shown. Panel B: Quantification of MT/DB ratio in 4 pairs of L-R mice breast tumors, as measured in different images by fluorescence microscopy. The L-mean of MT/DB fluorescence is lower than the R-one (unpaired T-test, p=0,01). Circles = Mouse 2, Triangles = Mouse 3, Squares= Mouse 4, Inverted Triangles = Mouse 5.
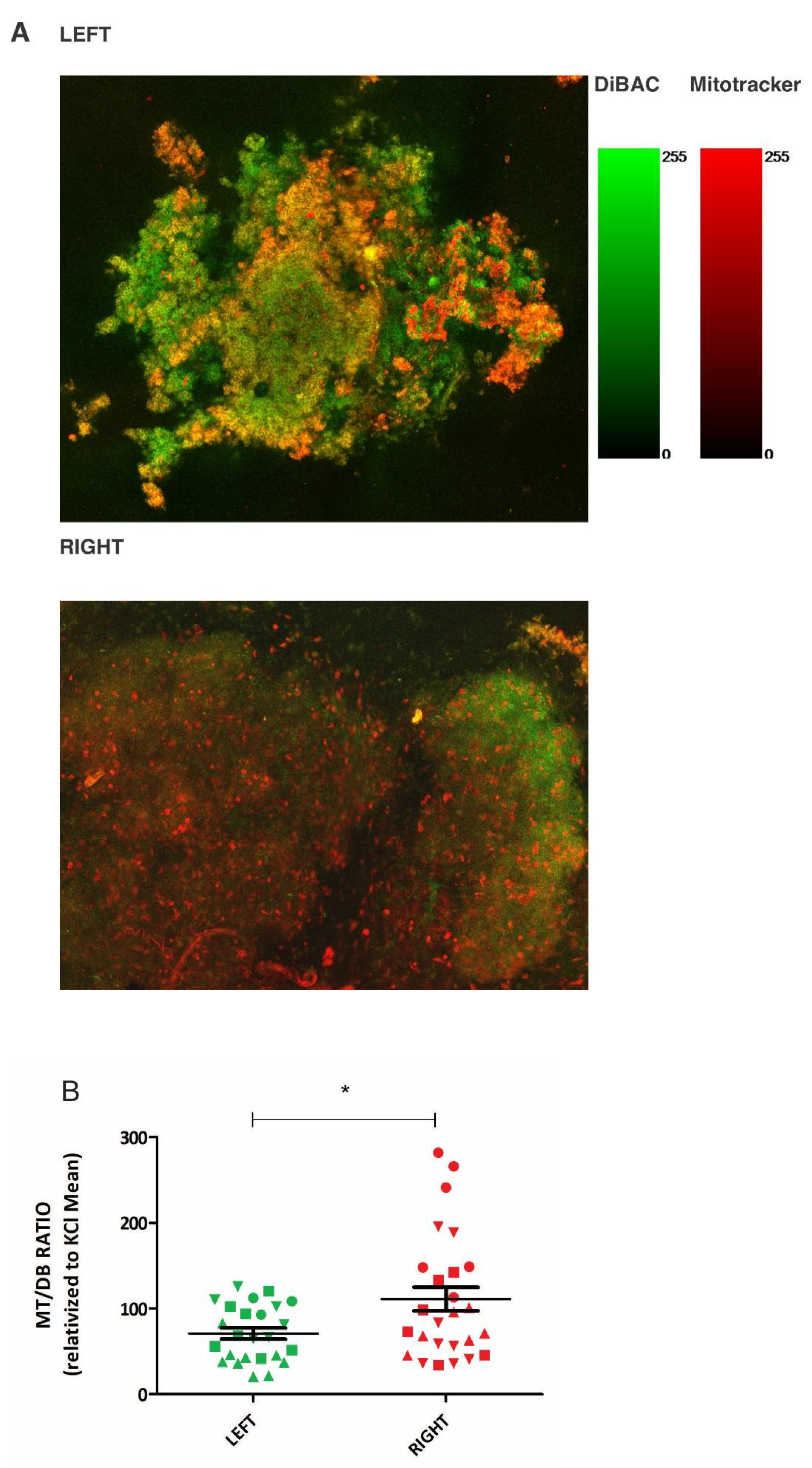
Figure 2.
Differential MT/DB fluorescence in L-R mice breast tumors. Panel A: representative flow cytometry image of breast tumor tissue, stained with MT and DB fluorescent probes. Left panel= L tumor cells, Right panel= R tumor cells. As can be seen, L-tumor cells show a decreased Q3 (% of MT-stained cells)/Q2 (% of DB-stained cells) ratio, as compared to R-cells. Panel B: quantification of MT /DB ratio in R-L tumor cells in 3 mice. In all, L-tumor cells show decreased MT/DB ratio (Two-Way Anova, p=0.05).
Figure 2.
Differential MT/DB fluorescence in L-R mice breast tumors. Panel A: representative flow cytometry image of breast tumor tissue, stained with MT and DB fluorescent probes. Left panel= L tumor cells, Right panel= R tumor cells. As can be seen, L-tumor cells show a decreased Q3 (% of MT-stained cells)/Q2 (% of DB-stained cells) ratio, as compared to R-cells. Panel B: quantification of MT /DB ratio in R-L tumor cells in 3 mice. In all, L-tumor cells show decreased MT/DB ratio (Two-Way Anova, p=0.05).
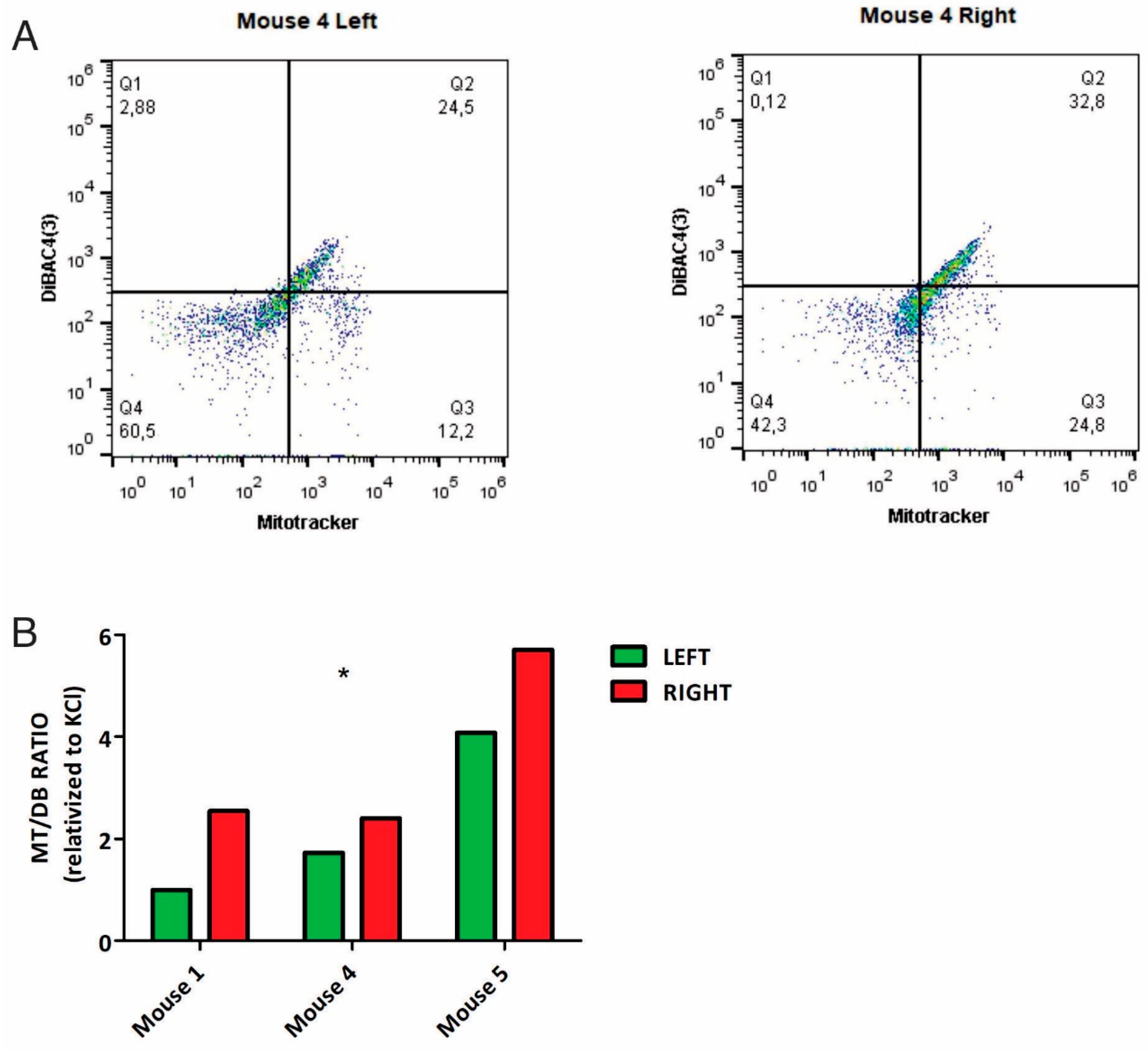
Figure 3.
Low expression of the 6-ICH signature associates with pro-mitotic pathways. Panel A: Differential gene expression between Low vs High expression of the 6-ICH signature. Volcano Plot showing the significantly down-regulated (blue dots) and up-regulated (red dots) genes in the Low-expressing subgroup, as compared to the High-expressing one. Performed in Xena Functional Genomics Explorer-USCS (https://xenabrowser.net/) (p values<0.05). Panel B: Enrichment Analyses of the DGE between Low-expressing vs High-expressing samples. Pro-mitotic biological processes and molecular functions (2018 version) are over-expressed in the Low-expressing subgroup.
Figure 3.
Low expression of the 6-ICH signature associates with pro-mitotic pathways. Panel A: Differential gene expression between Low vs High expression of the 6-ICH signature. Volcano Plot showing the significantly down-regulated (blue dots) and up-regulated (red dots) genes in the Low-expressing subgroup, as compared to the High-expressing one. Performed in Xena Functional Genomics Explorer-USCS (https://xenabrowser.net/) (p values<0.05). Panel B: Enrichment Analyses of the DGE between Low-expressing vs High-expressing samples. Pro-mitotic biological processes and molecular functions (2018 version) are over-expressed in the Low-expressing subgroup.
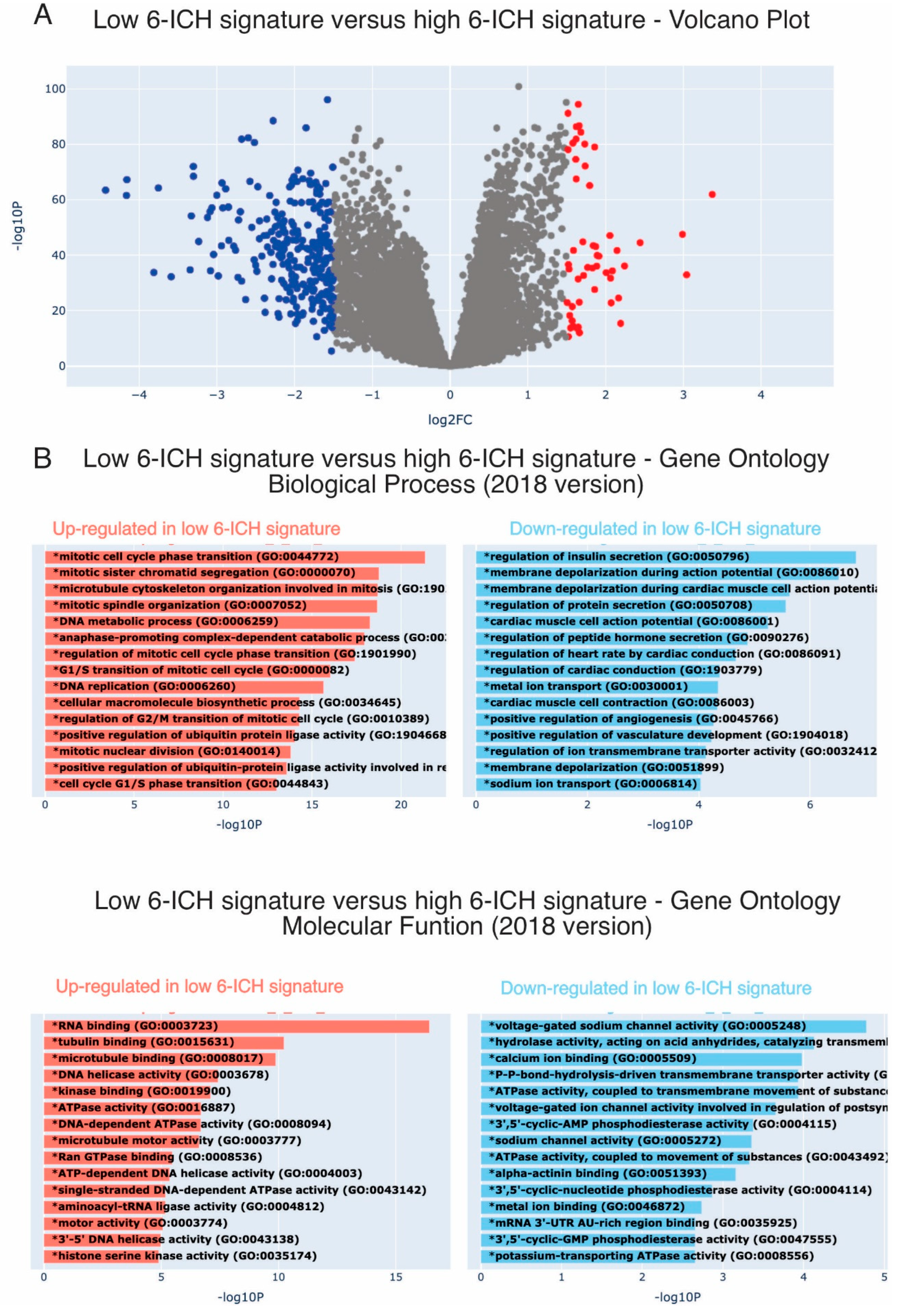
Figure 4.
Inverse correlation of the 6-ICH signature with proliferation, stemness markers, and survival rates. Panels A and B. Expression data in a Discovery Cohort of 533 primary breast tumors and a Validation Cohort of 549 primary breast tumors. Blue indicates less expression, red increased expression. Inverse correlation is shown between 6-ICH signature expression and markers KI67 and stemness score; ROC curves are presented showing the sensitivity and specificity of the 6-ICH signature to distinguish high from low KI67 and stemness score. Panel C. expression data of the 6-ICH signature in a Pan-cancer cohort of 9,546 tumors of different types. Panel D. Kapan Meier curves showing decreased survival (blue curves) in patients with tumors expressing lower 6-ICH signature, in a breast cancer and a pan-cancer cohort.
Figure 4.
Inverse correlation of the 6-ICH signature with proliferation, stemness markers, and survival rates. Panels A and B. Expression data in a Discovery Cohort of 533 primary breast tumors and a Validation Cohort of 549 primary breast tumors. Blue indicates less expression, red increased expression. Inverse correlation is shown between 6-ICH signature expression and markers KI67 and stemness score; ROC curves are presented showing the sensitivity and specificity of the 6-ICH signature to distinguish high from low KI67 and stemness score. Panel C. expression data of the 6-ICH signature in a Pan-cancer cohort of 9,546 tumors of different types. Panel D. Kapan Meier curves showing decreased survival (blue curves) in patients with tumors expressing lower 6-ICH signature, in a breast cancer and a pan-cancer cohort.
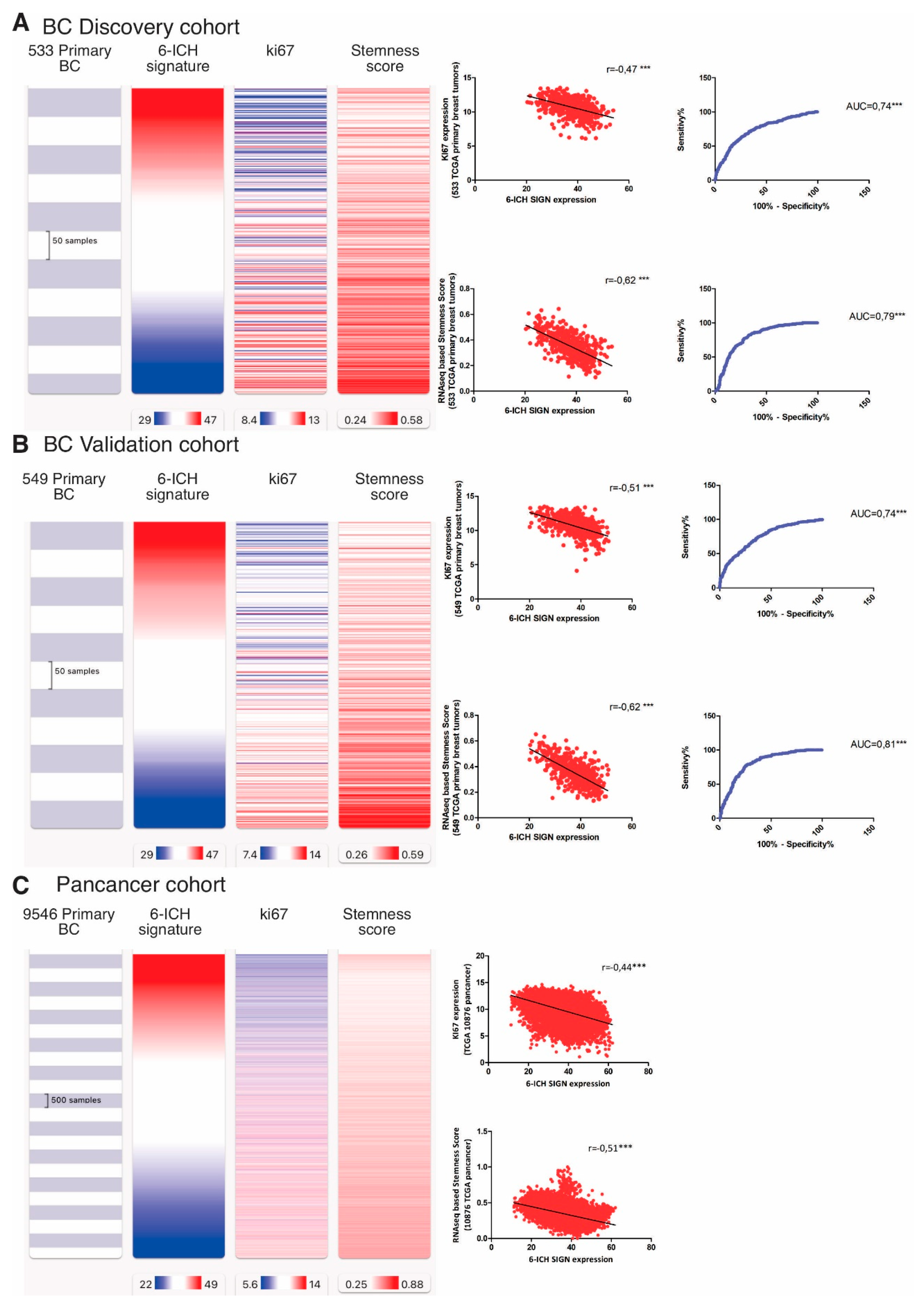
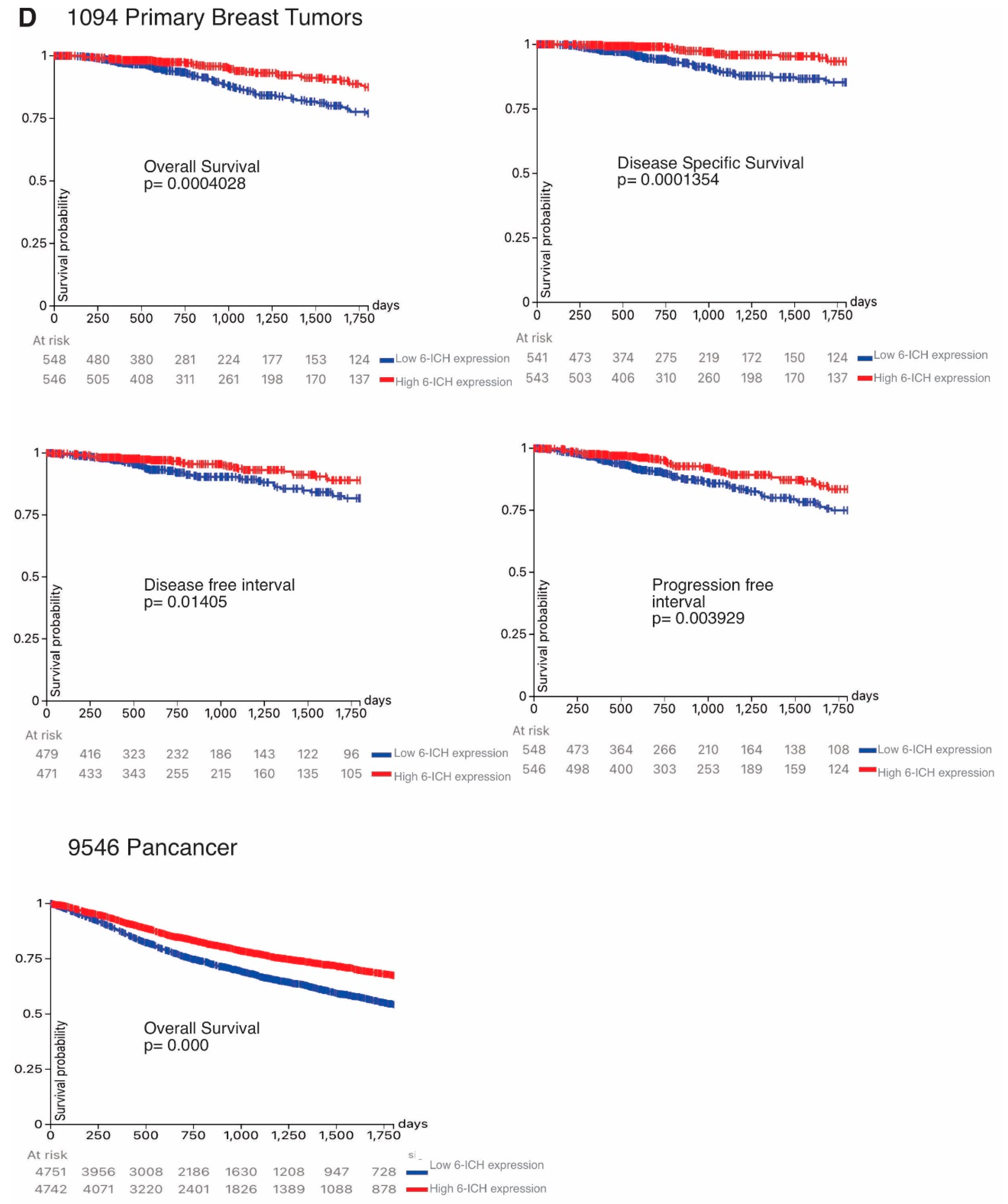
Figure 5.
Inverse correlation of the 6-ICH signature with DNA methylation writers’ expression. In 1,094 primary BC from TCGA, inverse correlation between the 6-ICH signature and the signature of DNA methylation writers composed of DNMT1, DNMT3A, DNMT3B, DNMT3L, TRDMT1, and DMAP1 (r=-0.42, Spearman correlation test, p<0.0001). Blue indicates less expression, red increased expression.
Figure 5.
Inverse correlation of the 6-ICH signature with DNA methylation writers’ expression. In 1,094 primary BC from TCGA, inverse correlation between the 6-ICH signature and the signature of DNA methylation writers composed of DNMT1, DNMT3A, DNMT3B, DNMT3L, TRDMT1, and DMAP1 (r=-0.42, Spearman correlation test, p<0.0001). Blue indicates less expression, red increased expression.
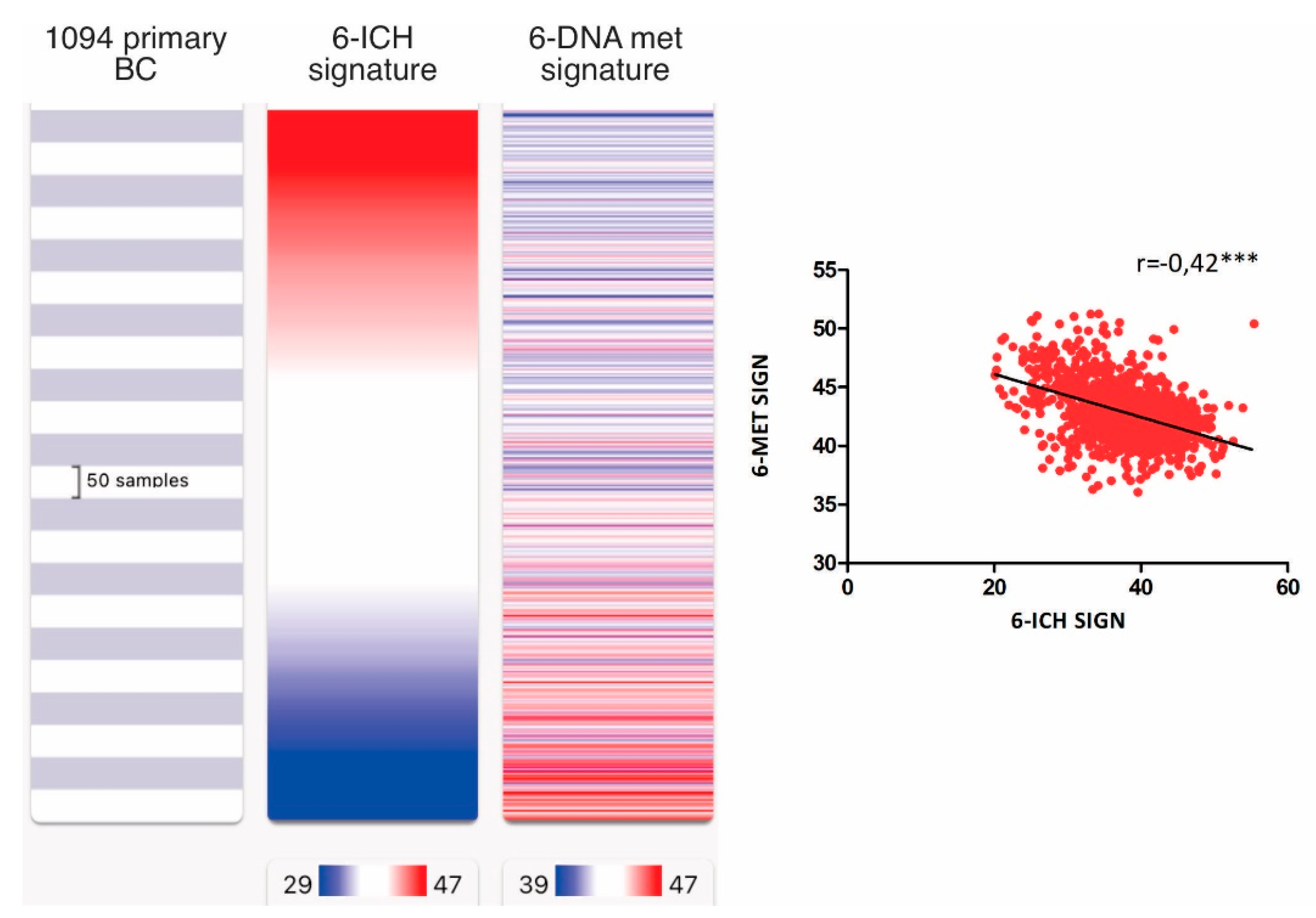
Figure 6.
Increased expression of proliferation and stemness markers in L-sided mice tumors. KI67 and CD44 expression in 4 paired L-R mice tumors (by ddPCR). Both markers are increased in L-tumors (paired T-test, p<0.05).
Figure 6.
Increased expression of proliferation and stemness markers in L-sided mice tumors. KI67 and CD44 expression in 4 paired L-R mice tumors (by ddPCR). Both markers are increased in L-tumors (paired T-test, p<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



