
Submitted:
16 May 2023
Posted:
17 May 2023
You are already at the latest version
Abstract
The inositol triphosphate associated proteins IRAG1 and IRAG2 are cGMP kinase substrate proteins, which regulate intracellular Ca2+. Previously, IRAG1 was discovered as a 125 kDa membrane protein at the endoplasmic reticulum, that is associated with the intracellular Ca2+ channel IP3R-I and the PKGIβ and inhibits IP3R-I upon PKGIβ-mediated phosphorylation. IRAG2 is a 75 kDa membrane protein homologue of IRAG1 and was recently also determined as a PKGI substrate protein. Several (patho-)physiological functions of IRAG1 and IRAG2 were meanwhile and recently elucidated in a variety of human and murine tissues, e.g. of IRAG1 in various smooth muscles, heart, platelets and other blood cells, of IRAG2 in pancreas, heart, platelets and taste cells. Hence, lack of IRAG1 or IRAG2 leads to diverse phenotypes in these organs, e.g. smooth muscle and platelet disorders or secretory deficiency, respectively. This review aims to highlight the recent research regarding these both regulatory proteins to envision its molecular and (patho-)physiological tasks and to unravel its functional interplay as possible (patho-)physiological counterparts.
Keywords:
cGMP
; cGKI
; IP3R-I
; IP3R-II
; IP3R-III
; IRAG
; IRAG1
; IRAG2
; Jaw1
; LRMP
; MRVI1
; PKGI
1. Introduction
The inositol 1,4,5-triphosphate receptor-associated cGMP kinase substrate 1 (IRAG1) and inositol 1,4,5-triphosphate receptor-associated 2 (IRAG2) are regulatory proteins, which interact with the intracellular Ca2+ channels inositol trisphosphate receptors (IP3-receptors/IP3R) and thereby modulate their physiological functions. The IRAG1 protein represents a 125 kDa membrane protein located at the endoplasmic reticulum (ER) or sarcoplasmic reticulum (SR), respectively. IRAG1 was previously identified as substrate protein, which interacts with cyclic guanosine monophosphate (cGMP) dependent protein kinase (PKG) Iβ isoform and regulates the IP3R-I function [1,2]. Then, it was elucidated that IRAG1 is mediating the Ca2+-dependent functions of cGMP in a variety of tissues, e.g. in the vasculature, in the gastrointestinal system and in platelets. These studies, performed with transgenic mice, showed that Irag1 deficiency leads to gastrointestinal and vascular dysfunction [3,4] and arterial thrombosis [5]. Interestingly, IRAG1 is homologous to the putative tumor suppressor protein murine retrovirus integration site 1 (MRVI1), which is expressed in myeloid immune cells [6]. Hence, it might be possible that IRAG1 is also involved in immunological functions.
IRAG2, which is also called lymphoid-restricted membrane protein (LRMP) or Jaw1, is a 75 kDa ER membrane protein and is particularly homologous to IRAG1 in its coiled-coil region (Figure 1) [7,8,9]. This region is essential for IP3R-I interaction of IRAG1 [1] and of IRAG2 [8]. Jaw1/LRMP was previously identified in lymphoid cells [7]. There were indications that it might be also involved in the development of lymphoma [10] or type 1 diabetes [11,12]. However, its molecular function was not established in these previous publications. Further investigations located Jaw1/LRMP in taste cells and thereby it was concluded that this protein might be associated with the perception taste [8].
Recently, several new studies revealed new molecular details and several novel functions of IRAG1 and IRAG2. Therefore, this review aims to summarize these important findings and shall give new aspects for studies of these regulatory proteins in future.
2. Functional features of IRAG1
2.1. Structure, interaction partners and cellular functions of IRAG1
The inositol 1,4,5-triphosphate receptor-associated cGMP kinase substrate 1 (IRAG1) or also known as its human homologue murine retrovirus integration site 1 (MRVI1) is, like IRAG2, a type II membrane protein linked to the ER via a C-terminal hydrophobic region. This hydrophobic region acts as a membrane anchor (Figure 1) [2,6]. Two isoforms of IRAG1 are existing, IRAG1a and IRAG1b, which differ in their amino acid sequence at the N-terminus, caused by alternative splicing. IRAG1a is the larger isoform due to 911 amino acids compared to IRAG1b with 859 amino acids [2,6]. For this reason, both molecules also differ in their molecular mass, that was calculated to be 90 kDa for IRAG1a and 98 kDa for IRAG1b [6]. However, the molecular weight for IRAG1a is 130 kDa and for IRAG1b 125 kDa, so that an average molecular mass of 125 kDa can be assumed [2]. A high content of prolines and glycosylations is discussed as a possible cause for the actually larger molecular masses compared to the predicted molecular mass [6]. Studies on the human IRAG1 gene revealed that splice variants of IRAG1 are truncated at the N- and C-terminal end. The C-terminal truncated variants act in a dominant-negative manner to counteract cGMP/PKGI signaling [13].
IRAG1 forms a macrocomplex with PKGIβ and IP3R-I, which is localized to the membrane of the ER to regulate the release of Ca2+ from this cellular structure via IP3R-I [1,2]. To this end, IRAG1 exhibits several structural features in addition to the membrane anchor already mentioned. For protein-protein interactions, it has a coiled-coil domain with a phosphorylation site for PKGIβ at each of its C- and N-terminal ends. [1,2,14]. Via amino acids 152 to 184, bovine IRAG1 interacts with the leucine zipper of PKGIβ located at its N-terminus but not with their linker region (Figure 1). By comparing the amino acids involved in the interaction between the two proteins, it can be assumed that the interaction occurs via electrostatic interactions [1]. Based on following studies, this assumption could be confirmed by showing that acidic residues in the leucine zipper of PKGIβ and small clusters of basic residues of IRAG1 are involved [14]. The interaction results in the phosphorylation of bovine IRAG1 at serine (Ser) 696 (Figure 1) respectively at Ser677 of human IRAG1 and inhibits the release of Ca2+ mediated via IP3R-I [1,5]. Furthermore, the interaction between IRAG1 and PKGIβ ensures that the PKGIβ reaches the ER, limiting its nuclear translocation and inhibiting its ability to mediate transcription [15]. This translocation could not be observed in vascular smooth muscle cells (VSMCs) of global Irag1-deficient mice [4]. However, the loss of the interaction between IRAG1 and PKGIβ may be the reason why protein expression of PKGIβ is reduced in Irag1 mouse mutants [3,4,16,17]. In contrast, the coiled-coil domain of IRAG1 is not involved in any interaction between IRAG1 and PKGIβ, but is necessary for the interaction of IRAG1 with the IP3R-I [1]. Furthermore, no protein interactions of IRAG1 with PKGIα and PKGII were observed [1,2,14]. Deletion of exon 12, which encodes the N-terminus of the coiled-coil domain, resulted in a 5 kDa smaller IRAG1 protein thus causing the interruption of the interaction with IP3R-I in COS-7 cells and murine smooth muscle tissue. In this context, it was further shown that the PKGIβ/IRAG1 interaction with IP3R-I is crucial for cGMP-dependent smooth muscle cell relaxation controlled by receptors (s. 2.4.) [3]. A graphical illustration of the PKGIβ/IRAG1 signaling pathway is shown in the following figure (Figure 2).
New results showed that IRAG1 is an interaction partner of hyperpolarization-activated cyclic nucleotide-sensitive isoform 4 (HCN4) channels. Expression of HCN4 and IRAG1 in HEK293 cells, followed by immunoprecipitation, resulted in IRAG1 as an interaction partner of HCN4. As an isoform-specific modulator of this channel, IRAG1 increases its activity to more depolarized potentials in the absence of cyclic adenosine monophosphate (cAMP) [18].
2.2. Expression pattern and localization of IRAG1
IRAG1 is a widely expressed protein in mammalian tissues. In the first description of MRVI1, RNA expression of the gene was detected in various human and murine tissues, e.g. heart, brain and skeletal muscle as well as myeloid cells [6]. Northern blot data of IRAG1 showed the same result in human tissues as when first described [2]. Further analysis on the expression and localization revealed that IRAG1 was detectable in a variety of murine tissues. High amounts were found in smooth muscle containing tissues like aorta, stomach or colon and in platelets. Only small amounts were found in the heart or the spleen [19]. In this context, expression and localization of PKGIβ, which is one of the interaction partners of IRAG1, were also investigated [2]. The expression and tissue distribution of PKGIβ correlated with that of IRAG1. In the brain this correlation was not associated with the localization of both proteins [19]. Cellular localization studies exposed that heterologous expressed IRAG1 was detectable perinuclearly, which supports the idea that IRAG1 is located in the membrane of the ER, where it forms a macrocomplex with the PKGIβ and IP3R-I [2,6,19]. In freshly isolated smooth muscle cells (SMCs) of cerebral arteries the macrocomplex, consisting of IRAG1, PKGIβ and IP3R-I, was localized to the SR, which is the equivalent of the ER in muscle cells [20].
2.3. Impact of IRAG1 on Ca2+ signaling
The formation of the trimeric complex of PKGIβ, IP3R-I and IRAG1 raised the question of the importance of IRAG1 for the release of Ca2+. Thereby, it could be shown in COS-7 cells that IP3R-I mediated Ca2+ release is suppressed in dependence on cGMP, after stimulation with bradykinin, as soon as PKGIβ and IRAG1 are coexpressed in these cells. Similarly, IRAG1 inhibits IP3-induced release of Ca2+ in a cGMP-dependent manner [2]. This regulation of Ca2+ release is achieved by phosphorylation of IRAG1 due to PKGIβ at Ser696 of bovine IRAG1 [1]. To investigate the influence of IRAG1 on Ca2+ release under physiological conditions, experiments similar to those performed in the COS-7 expression system were conducted in cells of the human colon. Again, IRAG1 was shown to inhibit the cGMP-dependent release of Ca2+ from the ER mediated by IP3R-I and seems to be involved in the control of nitric oxide (NO)-dependent relaxation of gastrointestinal muscles [21]. Measurements with aortic smooth muscle cells gave similar results. Therefore, the cGMP-dependent release of Ca2+ was not reduced in the VSMCs of Irag1Δ12/Δ12 mouse mutants compared to those of wild type mice. This result supports the fact that the interaction between IRAG1 and the IP3R-I is essential for cGMP-dependent regulation of Ca2+ release (Figure 2) [3]. Studies on IP3-receptors confirmed the inhibitory function of IRAG1 on Ca2+ release due to IP3R-I, whereas the absence of IRAG1 resulted in enhanced Ca2+ release. Interestingly, IRAG1 was also observed to inhibit direct modulation of IP3R-I by protein kinase A (PKA) and the PKA-dependent phosphorylation of IP3R-I [22].
In summary, IRAG1 is of critical importance in the regulation of Ca2+ release from the ER via IP3R-I.
2.4. (Patho-)physiological functions of IRAG1
2.4.1. IRAG1 and the gastrointestinal system
PKGI is of important relevance for the function of gastrointestinal SMCs [23]. The fact that IRAG1 forms a complex together with the PKGIβ and is phosphorylated by this enzyme combined with the strong expression of IRAG1 in the gastrointestinal tract suggests that IRAG1 is also of vital importance there [1,2,19,21]. Therefore, the (patho-)physiological function of IRAG1 was investigated in more detail by mouse models: the Irag1Δ12/Δ12 mouse and the global Irag1-deficient (Irag1-/-) mouse [3,4]. Irag1Δ12/Δ12 mice reveal an interruption of the interaction between IRAG1 and IP3R-I, caused by deletion of exon 12 of Irag1, which encodes the N-terminal part of the coiled-coil domain. The loss of the interaction between IRAG1 and IP3R-I did not affect the carbachol induced contraction but impaired the cGMP-mediated relaxation of SMCs of the colon as well as the tonic phase of depolarization induced contraction [3,24]. Similar results were found in colonic SMCs of global Irag1-deficient mice [4]. However, in jejunum of Irag1Δ12/Δ12 mice this was not observed [24]. This confirmed the important function of IRAG1 in the cGMP-mediated relaxation of receptor-triggered contraction of SMCs. Furthermore, these results approved the interaction between IRAG1 and the IP3R-I and that PKGIβ/IRAG1 signaling is of major importance for the cGMP-mediated relaxation of smooth muscles.
Studies on the physiological functions of IRAG1 in mouse models showed that IRAG1 is crucial for the functionality of the gastrointestinal tract in toto. Irag1Δ12/Δ12 mice developed an enlarged gastrointestinal tract, impaired gastrointestinal motility and pyloric stenosis [3]. These pathological changes could also be observed in global Irag1-/- mice [4]. Further studies with these transgenic mice revealed that they had an iron deficiency anemia as a result of gastrointestinal bleedings and subsequently developed a splenomegaly [16]. The results verified indications about splenomegaly in earlier studies of Irag1 mouse mutants [3,4]. Interestingly, these findings occurred mainly in female Irag1-deficient mice [16]. In this context, it was shown, that IRAG1 has a function in the development of achalasia. Achalasia is a disease of the gastrointestinal tract, which leads to impaired esophageal motility and incomplete relaxation of the lower esophageal sphincter. In two achalasia patients, a homozygous nonsense mutation of IRAG1 was detected. This mutation resulted in a loss of the known interaction of IRAG1 and PKGIβ. As a consequence, the central role of IRAG1 in regulation of Ca2+ levels and the associated regulation of cGMP-regulated smooth muscle relaxation was lost [25]. This case report is consistent with previously reported data showing that micro RNA (miRNA) regulate IRAG1 in esophageal SMCs in achalasia patients [26]. In Irag1 mouse mutants it was further observed that mutation or deficiency of IRAG1 caused a decrease in protein expression of PKGIβ [3,4,16]. The reduced protein expression of PKGIβ could not be explained by a decrease of the corresponding gene expression [16]. In this context, it is interesting to know that Prkg1 mouse mutants – which do not encode both isoforms of the PKGI – develop a similar phenotype to the Irag1 mouse mutants, only in a much stronger manifestation [27,28,29].
Combining the known data on IRAG1 and PKGIβ, this suggests that a disruption of PKGIβ/IRAG1 signaling pathway is significantly responsible for development of gastrointestinal disorders.
2.4.2. IRAG1 and (cardio-)vascular system
Besides the gastrointestinal SMCs IRAG1 is also highly expressed in VSMCs and the heart [19]. In vitro experiments with VSMCs of Irag1Δ12/Δ12 and Irag1-/- mice gave comparable results to those in SMCs of the colon. The loss of the interaction between IRAG1 and IP3R-I impaired cGMP-mediated relaxation of VSMCs of aorta [3,4]. Furthermore, in global Irag1-deficient mice the cGMP- and atrial natriuretic peptide (ANP)-mediated relaxation of VSMCs was reduced after acetylcholine induced contraction. Thus, IRAG1 has a central role in NO/cGMP- and ANP-mediated relaxation of VSMCs [4]. The detailed mechanism of cGMP-mediated relaxation of VSMCs has not yet been conclusively determined. A possible mechanism could be an interaction between IRAG1 and the transient receptor potential melastatin 4 channel (TRPM4). These sodium channels are activated by Ca2+ from the SR and take part in the vasoconstriction of blood vessels. In cerebral arteries IRAG1 and TRPM4 were colocalized in the SR of those VSMCs. When targeting Irag1 with morpholinos the NO-induced vasodilation was blunted and activity of TRPM4 was reduced. This indicates that NO/cGMP/PKG signaling decreases TRPM4 activity through an IRAG1-mediated inhibition of Ca2+ release from the SR [20]. Moreover, it was shown that not only cGMP promotes relaxation in VSMCs via IRAG1. The cyclic nucleotide cyclic cytidine 3’,5’-monophosphate (cCMP) also influences their relaxation. So, it was demonstrated that cCMP-mediated relaxation is also reduced in Irag1-deficient VSMCs in analogy to cGMP-mediated relaxation. It can be assumed that the effects of cCMP occur via stimulation of PGKI which then leads to phosphorylation of IRAG1 [30]. Physiological investigation of IRAG1 in Irag1-deficient mice exposed that a loss of IRAG1 caused a mild pulmonary arterial remodeling and an increasing percentage of muscularized arteries if these mice were kept under normoxic conditions [17]. A genetic analysis of a Caucasian family with moyamoya syndrome (MMS), which was associated with neurofibromatosis type 1 (NF1), revealed a polymorphism of IRAG1 in exon 5. This polymorphism might represent IRAG1 as a genetic susceptibility factor for MMS in NF1 [31]. Furthermore, IRAG1 was one of seven new identified genes in the context of lacunar stroke and was abundantly expressed on astrocytes [32].
Based on the physiological function of IRAG1 in VSMCs, the role of IRAG1 in the heart was of interest as it is also expressed there [6,17,18,19]. A good parameter to assess this is the heart function. Irag1 mouse mutants have a slight tendency for hypotension [3,4]. However, after induction of a sepsis, blood pressure remained constant in Irag1-deficient mice in contrast to wild type mice [4]. As already mentioned, Irag1-/- mice did not show any difference in the blood pressure. Though, under normoxic conditions an Irag1 deficiency caused a significant increase of the systolic pressure of the right ventricle (RV) and they develop a RV hypertrophy and dilatation. However, no evidence of heart fibrosis was recognized. Thus, an Irag1 deficiency causes the spontaneous development of pulmonary hypertonia (PH) under normoxic condition without any trigger, e.g. hypoxia [17]. This finding is consistent with the data of the pulmonary vascular remodeling of Irag1-/- mice, as pulmonary vascular remodeling is often associated with PH. The cardioprotective function of IRAG1 is also supported by other results. Burn-induced cardiomyopathy in rats causes a decrease of cGMP and various genes like Irag1 and Prkg. If these rats are treated with the phosphodiesterase (PDE) 5 inhibitor sildenafil, there was a normalization of cardiac function and an increase of cGMP and mRNA levels, among others of Irag1 and Prkg. This indicates that PDE5/cGMP/PKG are mediating the burn-induced heart dysfunction [33]. In the process of demonstrating HCN4 as a novel interaction partner of IRAG1, it was detected in the sinoatrial node of murine hearts. Based on a model of funny current (If) it was predicted that IRAG1 will increase the If in sinoatrial myocytes [18]. So, IRAG1, as a HCN4 modulator, is suggested to have a possible role in heart rate regulation [34].
Furthermore, IRAG1 is expressed in lungs [19]. Analysis of the expression of IRAG1 protein in lungs and isolated pulmonary artery of smooth muscle cells (PASMCs) of end stage idiopathic pulmonary arterial hypertension (IPAH) revealed an increased expression of IRAG1 and PKGIβ. This contrasted with hypoxic Irag1-deficient mice. They showed a decreased PKGIβ protein expression in lung and murine PASMCs [17].
IRAG1 is not only expressed in tissues of the cardiovascular system, but also in a huge amount in platelets [19]. The facts that PKGI has a crucial function in platelet aggregation and that IRAG1 is a substrate of the PKGIβ and also forms a macrocomplex with PKGIβ and IP3R-I, raised the question of the physiological function of IRAG1 in platelets [2,35]. This well-known macrocomplex, as well as the phosphorylation of IRAG1 by PKGIβ, which results in the inhibition of Ca2+ release from IP3R-I, was found in human and murine platelets. However, in human platelets two phosphorylated serine residues were identified – Ser664 and Ser677 – whereby the Ser677 in human IRAG1 corresponds to the Ser696 in bovine IRAG1 [2,5,36]. The examination of platelets from Irag1∆12/∆12 and global Irag1-deficient mouse mutants revealed that cGMP- and NO-induced platelet aggregation was inhibited in these mouse mutants [5,36]. cAMP- or prostacyclin-mediated aggregation was not affected and the cGMP- or NO-mediated and thrombin-induced Ca2+ release was not suppressed in the mutant platelets [5]. Therefore, IRAG1 has a crucial function in impeding NO/cGMP signaling in platelet aggregation by suppression of intracellular Ca2+ [5]. However, not only the cGMP-mediated inhibition of platelet aggregation is affected by IRAG1. The cCMP-mediated inhibition of aggregation was also inhibited in Irag1-deficient platelets. This suggests that, similar to VSMCs, cCMP-regulated effects are operated by the PGKI/IRAG1 pathway [30]. In addition, IRAG1 is involved in the NO- or cGMP-induced inhibition of adenosine triphosphate and serotonin secretion from dense granules and P-selectin secretion from alpha granules in platelets [36]. The circumstance that global Irag1-deficent mice showed reduced bleeding time confirmed the results on the physiological function of IRAG1 determined in in vitro experiments [36]. Furthermore, IRAG1 is necessary for cGMP-dependent inhibition of platelet activation and prevention of arterial thrombosis as well as for inhibition of thrombin-induced adhesion of platelets to fibrinogen [5,36]. These experimental data on the physiological function of IRAG1 are supported by several clinical data. Meta analyses discovered single nucleotide polymorphisms (SNPs) in the human IRAG1 gene, which cause an increased aggregability of platelets to agonists and reduce mean platelet volume [37,38,39].
Taken together, IRAG1 is an important protein for the physiological function of the (cardio-)vascular system and for platelet function. These findings and the participation in the development of disorders make IRAG1 an interesting target for further studies and possible pharmacological treatments.
2.4.3. IRAG1 and cancer disease
In addition to the (patho-)physiological functions of IRAG1 described so far, it is also important in tumor diseases. This was already evident in the first description of IRAG1 respectively MRVI1, where it was detected in BXH2 leukemias. It was concluded that IRAG1 can induce myeloid leukemia by altering the expression of a gene important for myeloid cell growth and/or differentiation, and it was suspected that IRAG1 may function as a tumor suppressor gene [6]. Involvement of IRAG1 in myeloid leukemias was also demonstrated in another study investigating the extent to which the recurrent chromosomal translocations of the tyrosine kinases BCR-ABL, TEL-PDGFRB, and TEL-JAK2 regulate distinct and overlapping gene transcription profiles. It was shown that IRAG1 was increasingly expressed in Ba/F3 cells after transfection with either BCR-ABL or TEL-PDGFRB. Again, it was concluded that IRAG1 plays an important role in the development of leukemia. However, this remains to be conclusively elucidated [40]. This statement is supported by the findings that CD300A was upregulated in patients with an acute myeloid leukemia of the intermediate or adverse risk category of the WHO criteria (2018) and predicts a poor survival. CD300A upregulation stimulates the cGMP/PKG signaling pathway and IRAG1 was positive correlated with CD300A [41].
Besides its involvement in the development of myeloid leukemia, IRAG1 is also important in solid tumors. In particular, association of IRAG1 with tumors of the female reproductive system was described so far. The response of patients suffering from serious ovarian carcinoma stage III to cytostatic therapies varies. It was reported that IRAG1 was upregulated in 61 % of these patients. Overexpression of IRAG1 has a direct impact on the survival because these patients have a significantly worse prognosis in terms of survival than those in whom IRAG1 was not upregulated. Furthermore, if the tumor was chemoresistant, the prognosis was even worse. These results implicate that IRAG1 is involved in chemoresistance of serious ovarian carcinoma [42]. While the involvement of IRAG1 in ovarian carcinoma is associated with a poor prognosis, this does not apply to endometrial carcinoma. MicroRNAs modulate cellular processes and there is growing evidence that they are linked to the progression of diverse cancers like the endometrial carcinoma. MicroRNA miR-940 acts as an oncogene during progression and high expression is associated with, among other things, reduced overall survival. In these tumors it was found that expression of IRAG1, which is a direct target of miR-940, was decreased. Moreover, IRAG1 expression was associated with, e.g. survival of the patients in the context that high expression levels of IRAG1 are linked to a good prognosis. Thus, miR-940 regulates progression of endometrial carcinoma by affecting the expression of IRAG1 [43]. Similar to endometrial carcinoma, low expression of IRAG1 in cervical carcinoma was associated with poor overall survival. This is caused by hypermethylation in the promotor regions of IRAG1 promoter, resulting in low gene expression of IRAG1 [44]. Furthermore, the gene expression of IRAG1 was upregulated in context of pancreatic ductal adenocarcinoma (PDAC), after silencing the transcription factor basic transcription factor 3 (BTF3) in pancreatic cancer cell lines [45]. Additionally, expression of IRAG1 was negatively associated with high expression of the cell division cycle associated protein 2 (CDCA2) in glioma [46].
Based on the current data, it can be concluded that it is not yet possible to make a clear statement on the function of IRAG1 in the development and/or progression of tumors. This is mainly due to the fact, that the data collected so far are established by bioinformatic analyses and investigations on corresponding cell lines. However, from these previous studies it can be assumed that IRAG1 has different functions depending on the tumor type. The exact role of IRAG1 in these tumors remains to be investigated in systematic experimental work.
2.4.4. Further (patho-)physiological functions of IRAG1
Apart from the previously mentioned (patho-)physiological functions, IRAG1 might also have more functions. For example, an increased expression of IRAG1 in addition to two other genes was described in keratoconus, but the role of IRAG1 in this eye disease is still unclear so far [47]. The attachment and motility of osteoclasts is regulated by NO and PGKI, which modulates the release of Ca2+ by the IP3R-I. Functional studies on osteoclasts revealed that IRAG1 is expressed in these cell types and is required for the Ca2+ release during motility. If the IRAG1 mediated Ca2+ release from the IP3R-I was disrupted, this might be a cause for dysfunction of osteoclasts [48]. IRAG1 reveals not only a significant role in the development of tumors of the female reproductive system, but it is also involved in other diseases in this body region. Proteomic analysis of tissue from intrauterine adhesions (IUA) – also known as Asherman’s syndrome – revealed IRAG1 as one of seven proteins which were upregulated in IUA. However, the exact mechanisms and signaling pathways remain to be investigated before IRAG1 can act as a potential target protein for clinical treatment of IUA [49]. Further investigations showed that IRAG1 participates in endothelial ANP/cGMP/PKGI signaling [50].
Taken together, IRAG1 is of importance for several physiological functions due to its large tissue distribution, but needs to be further investigated.
2.5. Polymorphisms of IRAG1 gene
In the last years several single nucleotide polymorphisms (SNPs) and variations (SNVs) were discovered in conjunction with IRAG1. IRAG1 was identified as one of four loci which are associated with arterial stiffness index. However, in the following secondary analysis of these data, IRAG1 was not genome-wide significant, but it represented an interesting candidate [51]. SNPs of IRAG1 were also identified in the context of migraine. There were associations of IRAG1 with migraine and combined with cervical artery dissection (CeAD) there were associations for CeAD mapped to the IRAG1 gene [52,53]. As described in section 2.4.2. polymorphisms of the IRAG1 gene were associated with NF1 associated MMS [31]. Platelet aggregation was also affected by variations of the IRAG1 gene. They caused an increased aggregability of the platelets to agonists [37,38,39]. Nevertheless, not all polymorphisms of IRAG1 were relevant for the development of dysfunctions. Patients with unstable angina, which had IRAG1 polymorphisms, did not show any association between the polymorphism and the disease [54]. Based on data of the UK Biobank study, there was an association between SNP of IRAG1 and childhood-onset asthma [55].
Thus, genetic variations of IRAG1 have an important role in cardiovascular disorders. However, their exact function and significance in the development and progression of the respective disease still needs to be investigated in further experimental studies with the aim of clarifying the potential role of IRAG1 in these processes and if it is suitable as a possible therapeutic target.
2.6. Significance of IRAG1 as a diagnostical/prognostic marker
Independent of the known (patho-)physiological functions of IRAG1, it was shown to be important in the context of diagnosis and prognosis of diseases. However, the extent to which IRAG1 is suitable as a marker is still unclear or under investigation. For example, IRAG1 was demonstrated to play an important role in the prognosis of idiopathic pulmonary fibrosis (IPF). This is a chronic respiratory disease characterized by peripheral distribution of bilateral pulmonary fibrosis, which is associated with a poor prognosis and short survival. Bronchoalveolar lavage cells from the Gene Expression Omnibus (GEO) database were evaluated and IRAG1 was identified as one of the relevant genes to the prognosis of IPF, among other genes [56,57].
As described in the previous section, expression of IRAG1 is mainly associated with the prognosis of survival depending on the respective types of tumors (s. 2.4.3.). It should be noted that these data are mainly evaluations of databases, which should be further investigated to verify the previous results. However, it must be mentioned that in cervical cancer expression and methylation levels of IRAG1 could be effectively differentiated between cancer and healthy tissue samples [44].
Based on the present knowledge about the (patho-)physiological function of IRAG1, the currently available data indicate that IRAG1 could also be used as a diagnostic and/or prognostic marker in some fibrotic or cancer diseases. Furthermore, it can be assumed that in the course of further research, IRAG1 may be used as a marker for diagnosis or progression in additional diseases.
3. Functional features of IRAG2
3.1. Structure of IRAG2
The inositol 1,4,5-triphosphate receptor associated 2 (IRAG2) is also known as Jaw1 or lymphoid restricted membrane protein (LRMP) and was first described in 1994 by Behrens et al. [7]. IRAG2 is a membrane protein consisting of 539 amino acids and is targeted to the cytoplasmic face of the ER [7,58]. Moreover, localization of IRAG2 was shown at the outer nuclear membrane [59,60]. The insertion into the ER membrane occurs posttranslationally [7,58]. After targeting to the ER, IRAG2 is cleaved at its C-terminus, leading to the existence of a shorter IRAG2 fragment besides the full-length IRAG2 [58,59,61]. This cleavage event occurs at the C-terminal, luminal domain between amino acid Ala509 and Ala510 and is achieved through the signal peptidase complex isoform SEC11A [61].
IRAG2 consists of a C-terminal anchor domain, which is important for its localization to the ER, a coiled-coiled domain, that is significant for protein-protein interactions and a N-terminal domain with a cytoplasmic orientation (Figure 1). Due to its C-terminal hydrophobic anchor domain and the cytosolic N-terminal domain, IRAG2 is classified as a type II membrane protein [7]. The short luminal domain of IRAG2 resides 39 carboxyl (C)-terminal amino acids and is important for the targeting of IRAG2 into the ER membrane. Replacement of the luminal domain through the luminal domain of other Klarsicht/ANC-1/Syne/homology (KASH) proteins leads to localization of IRAG2 at the outer nuclear membrane but not at the ER [61].
Especially in its coiled-coil domain IRAG2 reveals a homology of 44 % to the coiled-coil domain of IRAG1 [6,8,18]. Therefore, IRAG1 and IRAG2 share some similar interaction partners. Like IRAG1, IRAG2 interacts with the IP3-receptors in a variety of tissues and cell-lines. Besides the interaction with the IP3-receptors, IRAG2 – like IRAG1 – interacts with the HCN4 channel in CHO-cells and in HEK-cells [18,34]. The interaction with these ion channels and receptors suggests a function of IRAG2 in the gating-mechanisms of these channels and therefore an impact on physiological processes. Moreover, it was shown that IRAG2 interacts with Sad-1/UNC-84 (SUN) proteins and the microtubules [59].
3.2. Expression pattern and localization of IRAG2
First localization studies of IRAG2 showed mRNA expression mainly in lymphoid tissues and cell-lines, like B-cell-lines and T-cell-lines, where expression is regulated developmentally. The highest level of mRNA was found in pre-B-cells, pre-T-cells and mature B-cells. Only low expression was detected in mature T-cells and plasma B-cells, suggesting a role of IRAG2 in lymphoid development. Furthermore, protein expression of IRAG2 was seen in spleen and thymus. However, in nonhematopoietic cell-lines and tissues, no expression of IRAG2 was reported in these first studies [7]. Later, Tedoldi et al. performed immunohistological investigations, where further expression of IRAG2 was shown in peripheral lymphoid tissues like lymph nodes and the tonsil. Thereby, high expression was detected in germinal center B-cells and monocytoid B-cells but not in mantle zone B-cells and the interfollicular area. Furthermore, cortical thymocytes and splenic marginal zone cells were stained positive for IRAG2. In a bone marrow trephine, IRAG2 was detected in clustered normoblasts. This immunohistological analysis of IRAG2 also focused on hematolymphoid neoplasia like lymphomas, where IRAG2 expression was detectable in B-cell lymphomas but not in T-cell neoplasms. Additionally, IRAG2 is expressed in lymphomas arising from germinal centers, Burkitt´s lymphoma and lymphocyte-predominant Hodgkin’s disease. Hence, the expression in lymphoid neoplasms is reflecting the expression pattern that is observed in normal lymphoid tissues. Further, IRAG2 is detectable in almost all chronic lymphatic leukemias but not in classical Hodgkin’s disease. Besides the expression in lymphoid tissues, IRAG2 can be also found in non-lymphoid tissues and cells, like neuronal cells in the cerebral cortex, epithelial cells in the tonsil and in seminal vesicles, adrenal glands and zymogen-producing cells in the stomach [10]. Subsequent studies also showed presence of IRAG2 in various other tissues like sweet, bitter and umami taste-responsive cells, where it might play a role in taste signal transduction [8]. Furthermore, expression of IRAG2 is seen in sinoatrial nodes [18] and in intestinal tuft cells [62], where it might exert important physiological functions due to its interaction with different ion channels or receptors. We recently detected expression of IRAG2 in platelets and pancreatic acinar cells [9,63]. In these cells IRAG2 is involved in Ca2+ signaling, platelet aggregation and enzymatic secretion (s. 3.4., 3.6.4. and 3.6.5.).
3.3. Cellular functions of IRAG2
The C-terminus of IRAG2 shows a partial homology with the PPPX-motif of KASH proteins. This motif consists of four amino acids and is highly conserved between the five KASH proteins Nesprin1-4 and KASH5 [59,60]. Like IRAG2, KASH proteins also belong to the family of type II membrane proteins and are localized to the outer nuclear membrane. By its C-terminal luminal domain, KASH proteins interact with SUN proteins, that are localized to the inner nuclear membrane. This complex, called linker of nucleus and cytoskeletons (LINC), is formed in the perinuclear space by interaction of a trimeric KASH protein with trimeric SUN proteins, resulting in a hetero-hexamer complex, which is necessary for maintaining the nuclear shape. Furthermore, KASH proteins interact with microtubules through its cytosolic region [64,65,66,67]. This interaction allows a connection of the cytoplasm to the nucleoplasm and is important for maintaining shape and position of the nucleus [64,68]. For IRAG2, an interaction with SUN proteins and microtubules is also detected [59]. These findings suggest that IRAG2 acts as a KASH protein and is involved in maintaining the nuclear shape [59]. In a myeloma cell-line it is shown that depletion of IRAG2 leads to an aberrant nuclear shape, supporting this thesis. In conclusion, IRAG2 might function as part of the LINC-complex by interacting with microtubules on the cytosolic face and with SUN proteins in the perinuclear space via its KASH domain [59,60]. Furthermore, IRAG2 seems to act in an oligomeric state in this complex [59]. This oligomerization is probably achieved via the coiled-coil domain on the cytosolic face of the outer nuclear membrane [69].
Further studies showed that the N-terminal region of IRAG2 inhibits the formation of a smooth endoplasmic reticulum (OSER) [69]. It is known that overexpression and uncontrolled oligomerization of several ER resident proteins can cause OSER formation, which is characterized through highly dense structures like cisternae, nuclear karmellae and whorls [69,70,71,72]. Kozono et al. reported that deletion of the N-terminal region of IRAG2 leads to its aberrant oligomerization and in consequence to the formation of nuclear karmellae, where the ER membranes are stacked along the nuclear envelope. This phenomenon was observed for human and murine IRAG2, despite their relatively low percentage identity of the N-terminal region (43.6 %). However, a computational analysis indicates that the N-terminal regions of both murine and human IRAG2 are intrinsically disordered regions (IDR) [69]. This regions are characterized by higher polarity and lower hydrophobicity, compared to other, structured regions like coiled-coil domains or transmembrane domains, resulting in an unstable conformational state [73,74]. Moreover, IDRs regulate protein-protein interactions and are involved in the regulation of precise oligomerization [74,75,76]. Hence, it is suggested that the N-terminal region of IRAG2 regulates the oligomeric state of IRAG2 as an IDR by preventing the structural exposure of the coiled-coil as an oligomerization site. Loss of the N-terminal domain, however, leads to aberrant oligomerization of the protein. Thus, the N-terminal region of IRAG2 prevents the formation of an OSER, which is essential for maintaining the homeostatic localization of an IRAG2 oligomer and its interacting partners on the ER membrane [69].
Several components of the LINC-complex are involved in the morphology and position of the Golgi apparatus [77,78,79]. As IRAG2 functions as a KASH protein to maintain nuclear shape, it might play a role in the formation of the Golgi apparatus [59,80]. Investigations using B16F10-cells showed that IRAG2 is involved in maintaining the Golgi ribbon structure, associated with the microtubule network. Knockdown of IRAG2 in these cells causes fragmentation of the Golgi apparatus and a loss of the Golgi ribbon structure. Furthermore, depletion of IRAG2 disturbs the localization of the Golgi derived microtubule network, indicating that IRAG2 keeps the compact structure of the microtubule derived network and is needed to maintain the Golgi morphology. However, the mechanism how IRAG2 affects the maintenance of the Golgi apparatus still need to be clarified [80].
Moreover, these investigations of Okumura et al. indicate that IRAG2 might be associated with the position of the centrosome. As the correct position of the centrosome and Golgi apparatus is important for protein secretion, it is conceivable that IRAG2 has an impact on physiological functions derived from positional and morphological maintenance of the Golgi apparatus and the centrosome [80].
3.4. Impact of IRAG2 on Ca2+ signaling
IP3-receptors are important Ca2+ channels, which are activated upon binding of IP3, leading to the release of Ca2+ from intracellular stores [81]. An interaction of IRAG2 with IP3R-III was shown for the first time in COS-7-heterologous expression system, where IRAG2 was co-transfected with the IP3R-III [8]. Later, interaction was also detected with the IP3R-II in mouse embryo fibroblast (MEF) cells and with IP3R-I, IP3R-II and IP3R-III in HEK-cells, that were transfected with IRAG2 and one IP3-receptor subtype each [62,82]. Furthermore, we recently reported interaction of IRAG2 with all IP3-receptor subtypes in the murine pancreas as well as in murine platelets [9,63]. The interaction with these receptors occurs through the coiled-coil domain of IRAG2 and impacts the release of Ca2+ from intracellular stores [8,82].
In MEF-cells, which were co-transfected with IP3R-II and IRAG2, it is shown that IRAG2 enhances Ca2+ flux compared to MEF-cells not expressing IRAG2 [62]. Furthermore, Okumura et al. reported an increased release of Ca2+ in HEK-cells, which express IRAG2, compared to HEK-cells lacking IRAG2. Moreover, mutants of the coiled-coil domain of IRAG2 reveal no augmentative effect on Ca2+ release, suggesting that IRAG2 directly increases the Ca2+ release activity of the IP3-receptors. Interestingly, IRAG2 shows an augmentative effect on Ca2+ release on every IP3-receptor subtype, however, it modulates the activity of each IP3-receptor subtype in a subtle different manner [82].
In murine platelets we previously reported an augmentative effect of IRAG2 on Ca2+ release after stimulation with the agonists thrombin and collagen, which induce platelet aggregation [63]. Moreover, we detected an enhancement of basal Ca2+ release through IRAG2 in pancreatic acinar cells, which also impacts the basal exocrine amylase secretion. Furthermore, pancreatic acinar cells lacking IRAG2 reveal an increased frequency of Ca2+ oscillations, suggesting a modulation of the IP3-receptor activity through IRAG2 [9]. However, the mechanism how IRAG2 modulates Ca2+ oscillations through IP3-receptors and which IP3-receptor subtype is responsible for the IRAG2 derived oscillation pattern remains unclear and will be of interest in future investigations.
Recently, Kozono et al. stated that the cleavage of the C-terminal, luminal domain of IRAG2 enhances the augmentative effect of IRAG2 on the release of Ca2+ from intracellular stores. Mutation of the C-terminus – and therefore mutation of the cleavage site – results in a defect cleaving event and subsequently to a decreased release of Ca2+ via IP3-receptors. Therefore, this cleavage event might be a crucial step for the function of IRAG2 as an IP3-receptor regulator. However, the molecular mechanisms leading to this effect still remain unclear [61].
Taken together, these data suggest that IRAG2 enhances the release of Ca2+ from intracellular stores by regulating the activity of IP3-receptors (Figure 3). As IRAG1 inhibits Ca2+ release from the ER, IRAG2 might be a counterpart of IRAG1.
3.5. IRAG2 as a substrate of cGMP-dependent protein kinase I
As described in section 2.1., IRAG1 forms a ternary complex with PKGIβ and IP3R-I, in which IRAG1 is phosphorylated by PKGIβ. This phosphorylation results in an inhibition of Ca2+ release from the ER [1,2,3,5,36]. The interaction site of IRAG1 with PKGIβ is located between amino acids 152 and 184, however, this site is missing in IRAG2 [1,14]. Furthermore, we could not detect a stable interaction between IRAG2 and PKGIβ in murine platelets [63]. Despite the absence of a stable interaction site with PKGI, a quantitative phosphoproteomics study revealed phosphorylation of human LRMP at amino acids Ser363, threonine (Thr) 375 and Ser418 upon stimulation with NO-donors or riociguat in platelets [83]. Remarkably, the phosphorylation sites Ser363 and Thr375 in IRAG2 (consensus sequences: RSAS363 and RRVT375) (Figure 2) and the identified phosphorylation sites Ser664 and Ser677 in human IRAG1 (consensus sequences: RSMS664 and RRVS677) are very homologous. Moreover, we detected a cGMP-depen dent phosphorylation of IRAG2 in murine platelets [63]. As cGMP predominantly activates PKGI [35,84], we assume that phosphorylation of IRAG2 after cGMP-stimulation is achieved through PKGI (Figure 3). However, it still needs to be investigated which isoform of PKGI – PKGIα and/or PKGIβ – is responsible for phosphorylation of IRAG2. As murine platelets predominantly express PKGIβ and human platelets only express PKGIβ, IRAG2, like IRAG1, might be a substrate of PKGIβ [5].
3.6. (Patho-)physiological functions of IRAG2
3.6.1. Function of IRAG2 in intestinal type 2 immunity
The tumor suppressor p53 is crucial for the function of intestinal tuft cells to trigger type 2 immune response after parasitic infections. Previously, it was described that this regulation is mediated by IRAG2. Expression of IRAG2 is transcriptionally regulated by p53, where p53 ensures a high expression of IRAG2 [62]. Furthermore, IRAG2 is expressed in intestinal tuft cells and interacts with the IP3R-II in MEF-cells [62], which is the major IP3-receptor subtype in intestinal tuft cells [85]. Deficiency of p53 leads to lower levels of IRAG2 and results in an impaired Ca2+ flux [62]. However, Ca2+ flux in intestinal tuft cells is critical for the release of the cytokine IL-25, which triggers the type 2 immune response [85]. Taken together, it is suggested that p53 leads to a high expression of IRAG2 in tuft cells, which ensures Ca2+ flux in these cells and consequently the release of IL-25. In turn, the release of IL-25 is necessary for type 2 immune response upon parasitic infections. Further, it is stated that knockdown of IRAG2 results in a higher amount of parasites in the feces of Irag2-deficient mice, which might be a result of the impaired type 2 immune response [62].
3.6.2. Function of IRAG2 on HCN channels
Contrary effects of IRAG1 and IRAG2 on the function of ion channels were already described by Peters et al. [18]. As stated in section 2.1. and 2.4.2., HCN4 channels are regulated by IRAG1. These channels are activated in a voltage-dependent manner and are modulated by cAMP. Binding of cAMP results in a depolarizing shift, leading to an increased opening of the channels [86,87,88]. IRAG1 reveals a function on HCN4 channels by shifting the voltage dependent activation to more depolarized potentials in the absence of cAMP, resulting in an enhanced opening of the channels. Besides the interaction of HCN4 channels with IRAG1, interaction of IRAG2 with HCN4 channels was observed, too. However, IRAG2 does not lead to a gain of function, but causes a loss of function by reducing the cAMP-dependent shift of HCN4 channels to more depolarizing potentials. As IRAG1 and IRAG2 are both expressed in sinoatrial nodes, these data suggest important roles for both proteins in the regulation of cellular excitability. Thereby it is suggested that IRAG2 limits increases in If in response to stimulation of β-adrenoceptors [18,34]. Furthermore, these data contribute to our thesis that IRAG2 might be a counterpart to IRAG1.
3.6.3. Potential role of IRAG2 in taste-signal transduction
Expression of IRAG2 was shown in sweet, bitter and umami taste receptor expressing cells of murine circumvallate, foliate and fungiform papillae. In the circumvallate papillae coexpression with the IP3R-III was detected [8]. Moreover, direct interaction of IRAG2 and IP3R-III was shown in several cells-lines and tissues [8,9,63,82]. The IP3/Ca2+ signal cascade plays an important role in sweet, bitter and umami taste signal transduction. Activation of phospholipase Cβ2 (PLCβ2) in these cells results in production of IP3, which induces Ca2+ release through IP3R-III. This IP3-mediated Ca2+ release is necessary for transduction of taste signals. Mice lacking PLCβ2 or IP3R-III exert a deficiency in their ability to detect sweet, bitter and umami taste substances [89,90]. Consequently, the interaction of IRAG2 with IP3R-III suggests a role for IRAG2 in taste signal transduction [8]. However, to examine the exact function of IRAG2 in taste cells, further experiments are required.
3.6.4. Function of IRAG2 in exocrine pancreatic acinar cells
Recently we reported expression of IRAG2 in exocrine pancreatic acinar cells [9]. As IRAG2 interacts with all subtypes of IP3-receptors in the pancreas and enhances basal Ca2+ release in isolated pancreatic acinar cells (s. 3.4.), we investigated the impact of this effect on the physiological functions of the exocrine pancreas [9]. The main task of pancreatic acinar cells is the secretion of a pancreatic juice, consisting of different enzymes, that are needed for digestion of food components. The release of Ca2+ and the secretion of digestive enzymes, like amylase, are strongly linked. An increase of Ca2+ release triggers the secretion of amylase from granules [91,92,93]. In concordance with this, we found that the increased basal release of Ca2+ due to IRAG2 leads also to an increase of basal amylase secretion. Mice lacking IRAG2 reveal a decreased basal Ca2+ release and therefore also a reduced basal secretion of amylase. However, this in turn does not seem to affect the nutrient digestion dramatically, as Irag2-deficient animals show no significant differences in body weight compared to wild type animals [9]. Further, it remains unclear how IRAG2 affects other digestive enzymes, like lipase or trypsin, as well as bicarbonate production and secretion, which requires further investigation. Immunochemical studies also showed a higher amount of amylase in the pancreatic acinar cells of Irag2-deficient mice compared to wild type pancreata. As premature activation of digestive enzymes in the acinar cells contributes to the emergence of acute pancreatitis [93], IRAG2 might also reveal a protective effect against pancreatic diseases like acute pancreatitis. However, this topic must be investigated in future experiments using mouse models of acute pancreatitis.
3.6.5. Function of IRAG2 in platelets
Platelet activation and aggregation upon vascular injury is essential for primary hemostasis, but is also involved in the formation of occlusive thrombi [94]. IRAG1 is phosphorylated by PKGIβ, which causes an inhibition of IP3-receptor mediated Ca2+ release from the ER [1,2]. In turn, this results in a reduced aggregability of platelets and also prevents formation of occlusive thrombi [5,36]. We previously reported expression of IRAG2 in platelets, where it interacts with all subtypes of IP3-receptors. Interestingly, in contrast to IRAG1, IRAG2 enhances IP3-receptor mediated Ca2+ release from the ER in murine platelets. Moreover, we found that this enhanced Ca2+ release increases the aggregation rate of platelets. Platelets lacking IRAG2 reveal reduced Ca2+ release and platelet aggregation. This effect seems to be mediated by phosphorylation of IRAG2 upon stimulation with cGMP-analogues, as Irag2-deficient animals show an even reduced platelet aggregation after stimulation with cGMP or the NO-donor sodium nitroprusside [63]. Consequently, our results suggest that PKGI-dependent phosphorylation of IRAG2 in platelets causes an increase in Ca2+ release, which results in an enhanced platelet aggregation (Figure 3). This contrasts with the effect of IRAG1 in platelets. Therefore, we assume that IRAG1 and IRAG2 are counterparts in platelets. Furthermore, as IRAG2 shows an augmentative effect on Ca2+ release, it might also contribute to the formation of occlusive thrombi, which can cause severe cardiovascular diseases. Therefore, IRAG2 might be a potential target for prevention of those diseases.
3.7. Significance of IRAG2 as a prognostic marker of cancer
As described in 3.2., expression of IRAG2 was shown in a variety of lymphoid tissues and cell-lines, like germinal center (GC) B-cells [10]. Lymphochip array studies revealed that IRAG2 is one of the GC genes whose overexpression defines the good-prognosis GC subcategory of diffuse large B-cell lymphoma (DLBCL) [95,96]. Additionally, in a RT-PCR based study, high expression of IRAG2 was associated with good prognosis of DLBCL [97]. It is shown that IRAG2 is involved in the delivery of peptides to MHC class I molecules in a TAP-independent manner [98]. This could be of interest for the behavior of DLBCL upon IRAG2 overexpression, as there is evidence that loss of expression of MHC class II molecules is associated with a poor outcome. Also, MHC class II-negative cases show a decrease in tumor-infiltrating CD8-positive T-cells [99,100]. This might explain the association between IRAG2 overexpression and good prognosis of DLBCL [10].
In breast cancer, correlation is seen between disease-free survival and expression of genes which are involved in immune response regulation and normal development of lymphoid tissues. One of these genes is the IRAG2 gene. Therefore, high expression of IRAG2 might be a promising biomarker of life expectancy [101].
Preliminary studies also showed expression of IRAG2 in some cases of ovarian cancer, which suggests that IRAG2 may represent a potential marker in the field of ovarian cancer [10]. However, therefore further investigations are needed.
Recently, IRAG2 expression was examined in lung adenocarcinoma. Thereby it is shown that IRAG2 gene and protein expression is lower in patients with lung adenocarcinoma as well as in lung adenocarcinoma cell-lines. Also, high expression of IRAG2 correlates with a better prognosis of patients, indicating that IRAG2 is a positive prognostic predictor. In vitro experiments demonstrated that overexpression of the IRAG2 protein could decrease the proliferation, migration and invasion in A549-cells. Furthermore, multiple oncogenic signaling pathways, e.g. the p-STAT3, p-PI3K-p-AKT, p-MEK and EMT pathway, are downregulated when IRAG2 expression is enhanced. IRAG2 is also positively associated with various tumor-infiltrating immune cells and their markers. These data suggest that IRAG2 might act as a tumor suppressor gene. Additionally, high IRAG2 expression correlated with an upregulation of immune checkpoints. Hence, IRAG2 could also be involved in immunotherapy response of lung adenocarcinoma patients using checkpoint-inhibitors [102].
Lower IRAG2 expression was detected in many other solid cancers, e.g. breast invasive carcinoma, colon adenocarcinoma, glioblastoma multiforme, kidney chromophobe, kidney renal papillary cell carcinoma, prostate adenocarcinoma, rectum adenocarcinoma, and uterine corpus endometrial carcinoma [102]. This suggests a role for IRAG2 overexpression also in other solid cancers.
IRAG2 seems to be involved in the cellular response following boron neuron capture therapy (BNCR). In this therapy, 10B-boronophenylalanine can be preferentially taken up into tumor cells. In combination with radiation, this therapy results in an apoptosis of the tumor cells. Sato et al. described that a higher amount of fragmented IRAG2 is generated in human squamous carcinoma in response to BNCR. Hence, fragmentation of IRAG2 could be involved in the cellular response of BNCR and therefore in in BNCR-induced cell death. Additionally, IRAG2 fragmentation could be considered as a potential biomarker for cell damage in response to BNCR. However, further investigations are needed to evaluate this potential role [103].
3.8. Significance of IRAG2 polymorphisms
The pulmonary adenoma susceptibility 1 (Pas1) locus plays a major role in predisposition of lung tumors [104]. The IRAG2 gene is localized to this locus. Furthermore, expression of IRAG2 was detected in normal human lung tissue and human lung adenocarcinomas, as well as in mouse normal lung tissue and mouse lung tumors. Thereby, no obvious differences between normal lung and lung adenocarcinoma tissue in IRAG2 expression levels were observed [105,106,107]. This, however, is in contrast to the data of Jin et al. who detected a lower IRAG2 gene and protein expression in lung adenocarcinoma tissue [102]. It is also shown that an IRAG2 Pro537Leu nonconservative variation was linked with inflammatory response in the lung of several mouse strains [108]. Further, two SNPs were found for human IRAG2: a Val141Leu variation and a Ser197Cys variation. In animal studies, IRAG2 polymorphisms increased the risk of lung tumors. In humans, analysis revealed no significant association of these SNPs with enhanced lung cancer risk [102,105,106,107]. However, in patients with age at tumor onset ≤ 65 years, survival rates of patients carrying either the Leu/Leu or the Val/Leu genotype differed significantly from those with the Val/Val genotype. Kaplan–Meier analysis demonstrated that the median follow-up at death was 33 months for Leu allele carrier and 100 months for Val/Val cancer patients. These findings suggest that the IRAG2 Val141Leu polymorphism can predict survival in lung adenocarcinoma. A hypothesis for the impact of the Val141Leu SNP on the survival rate might be an altered modulation of patients’ immune system [107], as it was recently suggested that IRAG2 might act as tumor suppressor, which is involved in the progression of lung adenocarcinoma (s. 3.7.) [102]. The Ser197Cys SNP, however, showed no effect on survival of lung cancer patients [107].
The IRAG2 gene is localized at the insulin dependent diabetes (idd) susceptibility locus (idd6). A high number of SNPs were found on this locus, which are associated with type 1 diabetes. As idd6 seems to be involved in the control of T-cell survival and proliferation, candidate genes for idd6 are those who are implicated with the immune system [12]. Therefore, IRAG2 might be a strong candidate for this locus, as it is expressed in a variety of lymphoid tissues and cell-lines, like B-and T-cell-lines [7,12]. Also, a high number of IRAG2 SNPs is found on this locus in non-obese-diabetes (NOD) mice, which contributes to the thesis that IRAG2 might have an impact on development of diabetes [12].
Additionally, it is reported that the idd6 diabetes susceptibility region controls defective expression of the IRAG2 gene in NOD mice, where the NOD allele at this locus mediates lower mRNA expression levels of IRAG2. This leads to the hypothesis that decreased expression of IRAG2 in these NOD mice might constitute a type 1 diabetes susceptibility factor in this Idd6 region [11].
4. Conclusion and Outlook
As depicted in this review, IRAG1 and IRAG2 are membrane proteins, which regulate intracellular Ca2+ and are substrate proteins of PKGI. IRAG1 is located at the ER [2,6], whereas IRAG2 was not only found at the ER, but also at other cellular localization, like the outer nuclear membrane [59] or the Golgi apparatus [80]. Hence, it can be speculated whether IRAG1 might be localized at further intracellular positions and if it exerts further, up to now not established functions there. In this regard, it is interesting that IRAG1 and IRAG2 are modulators of HCN4 channels, that are located at the plasma membrane [34]. Hence, IRAG1 and IRAG2 might be found also at the plasma membrane or other locations where the ER plasma membrane is associated.
Until now, the regulation of the IP3R by IRAG1 and IRAG2 is not fully understood at the molecular level. Therefore, it would be tempting to reveal the molecular mechanism, by which the PKGI-mediated phosphorylation of IRAG1 inhibits and of IRAG2 activates the intracellular Ca2+ via the IP3R. These mechanisms might also point to new target sites for the development of novel pharmacological modulators of IP3R. Modulators of the IP3R could lead to valuable pharmacological drugs for various diseases, e.g. in the gastrointestinal tract, in the lung, in the heart, in immune cells or in cancer cells.
The functional interplay of IRAG1 and IRAG2 in tissues and cells is a topic, which needs further exploration. Several studies indicate that IRAG1 and IRAG2 act as functional counterparts in platelets or in pacemaker cells. However, this regulation might differ in diverse other cells and might depend on other factors, e.g. on PKG-mediated phosphorylation of IRAG1 and/or IRAG2, protein splice variants [21] on other protein modifications, e.g. glycosylation or truncation.
IRAG1 and IRAG2 could serve as biomarkers for several diseases in the future. Upregulated expression of IRAG1 was associated with bad prognosis of solid tumors [40,44], whereas the high expression level of IRAG2 could be a beneficial marker for prognosis of lung tumors or the development of type 1 diabetes. Several polymorphisms were elucidated for IRAG1 or IRAG2. For IRAG1, there were polymorphisms identified which are associated with migraine and CeAD, with achalasia and moyamoya syndrome, with asthma and with arterial thrombosis. For IRAG2, polymorphisms are coupled to the risk of cancer diseases, e.g. lung tumor, diabetes or immune diseases. Hence, these polymorphisms could serve as prognostic markers for these diseases. Regarding tumor diseases, it is noteworthy that both IRAG1 and IRAG2 were reported to possibly act as tumor suppressor genes [6,102]. However, the mechanism regarding this aspect was not elucidated up to now and would be tempting to determine in the future.
Taken together, these findings indicate that IRAG1 and IRAG2 might be applicable as potential targets for therapeutical and diagnostic biomarker/polymorphism approaches, e.g. in diverse cardiovascular, gastrointestinal and cancer diseases. Further molecular and functional research will be needed to exploit these different prospects.
Author Contributions
S.P., M.M. and J.S. planned and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Requests to access the figures should be directed to the corresponding author: Jens.Schlossmann@chemie.uni-regensburg.de
Acknowledgments
We thank Leonhard Jakob and Robert Peter for carefully proofreading the manuscript. The work was supported by the Bavarian state.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ammendola, A.; Geiselhöringer, A.; Hofmann, F.; Schlossmann, J. Molecular determinants of the interaction between the inositol 1,4,5-trisphosphate receptor-associated cGMP kinase substrate (IRAG) and cGMP kinase Iβ. Journal of Biological Chemistry 2001, 276, 24153–24159. [Google Scholar] [CrossRef] [PubMed]
- Schlossmann, J.; Ammendola, A.; Ashman, K.; Zong, X.; Huber, A.; Neubauer, G.; Wang, G.X.; Allescher, H.D.; Korth, M.; Wilm, M.; et al. Regulation of intracellular calcium by a signalling complex of IRAG, IP3 receptor and cGMP kinase Iβ. Nature 2000, 404, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Geiselhöringer, A.; Werner, M.; Sigl, K.; Smital, P.; Wörner, R.; Acheo, L.; Stieber, J.; Weinmeister, P.; Feil, R.; Feil, S.; et al. IRAG is essential for relaxation of receptor-triggered smooth muscle contraction by cGMP kinase. EMBO J. 2004, 23, 4222–4231. [Google Scholar] [CrossRef] [PubMed]
- Desch, M.; Sigl, K.; Hieke, B.; Salb, K.; Kees, F.; Bernhard, D.; Jochim, A.; Spiessberger, B.; Höcherl, K.; Feil, R.; et al. IRAG determines nitric oxide- and atrial natriuretic peptide-mediated smooth muscle relaxation. Cardiovasc. Res. 2010, 86, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Antl, M.; von Brühl, M.-L.; Eiglsperger, C.; Werner, M.; Konrad, I.; Kocher, T.; Wilm, M.; Hofmann, F.; Massberg, S.; Schlossmann, J. IRAG mediates NO/cGMP-dependent inhibition of platelet aggregation and thrombus formation. Blood 2007, 109, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Shaughnessy, J.D.; Largaespada, D.A.; Tian, E.; Fletcher, C.F.; Cho, B.C.; Vyas, P.; Jenkins, N.A.; Copeland, N.G. Mrvi1, a common MRV integration site in BXH2 myeloid leukemias, encodes a protein with homology to a lymphoid-restricted membrane protein Jaw1. Oncogene 1999, 18, 2069–2084. [Google Scholar] [CrossRef]
- Behrens, T.W.; Jagadeesh, J.; Scherle, P.; Kearns, G.; Yewdell, J.; Staudt, L.M. Jaw1, A lymphoid-restricted membrane protein localized to the endoplasmic reticulum. Journal of immunology (Baltimore, Md.: 1950) 1994, 153, 682–690. [Google Scholar] [CrossRef]
- Shindo, Y.; Kim, M.-R.; Miura, H.; Yuuki, T.; Kanda, T.; Hino, A.; Kusakabe, Y. Lrmp/Jaw1 is expressed in sweet, bitter, and umami receptor-expressing cells. Chem. Senses 2010, 35, 171–177. [Google Scholar] [CrossRef]
- Prüschenk, S.; Majer, M.; Schreiber, R.; Schlossmann, J. IRAG2 Interacts with IP3-Receptor Types 1, 2, and 3 and Regulates Intracellular Ca2+ in Murine Pancreatic Acinar Cells. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Tedoldi, S.; Paterson, J.C.; Cordell, J.; Tan, S.-Y.; Jones, M.; Manek, S.; Dei Tos, A.P.; Roberton, H.; Masir, N.; Natkunam, Y.; et al. Jaw1/LRMP, a germinal centre-associated marker for the immunohistological study of B-cell lymphomas. The Journal of pathology 2006, 209, 454–463. [Google Scholar] [CrossRef]
- Duarte, N.; Lundholm, M.; Holmberg, D. The Idd6.2 diabetes susceptibility region controls defective expression of the Lrmp gene in nonobese diabetic (NOD) mice. Immunogenetics 2007, 59, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Grimm, C.H.; Rogner, U.C.; Avner, P. Lrmp and Bcat1 are candidates for the type I diabetes susceptibility locus Idd6. Autoimmunity 2003, 36, 241–246. [Google Scholar] [CrossRef] [PubMed]
- von Werder, A.; Mayr, M.; Schneider, G.; Oesterle, D.; Fritsch, R.M.; Seidler, B.; Schlossmann, J.; Hofmann, F.; Schemann, M.; Allescher, H.D.; et al. Truncated IRAG variants modulate cGMP-mediated inhibition of human colonic smooth muscle cell contraction. Am. J. Physiol. Cell Physiol. 2011, 301, C1445–C1457. [Google Scholar] [CrossRef] [PubMed]
- Casteel, D.E.; Boss, G.R.; Pilz, R.B. Identification of the interface between cGMP-dependent protein kinase Iβ and its interaction partners TFII-I and IRAG reveals a common interaction motif. Journal of Biological Chemistry 2005, 280, 38211–38218. [Google Scholar] [CrossRef]
- Casteel, D.E.; Zhang, T.; Zhuang, S.; Pilz, R.B. cGMP-dependent protein kinase anchoring by IRAG regulates its nuclear translocation and transcriptional activity. Cell. Signal. 2008, 20, 1392–1399. [Google Scholar] [CrossRef]
- Majer, M.; Prueschenk, S.; Schlossmann, J. Loss of PKGIβ/IRAG1 Signaling Causes Anemia-Associated Splenomegaly. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Kojonazarov, B.; Hadzic, S.; Majer, M.; Bajraktari, G.; Novoyatleva, T.; Ghofrani, H.A.; Grimminger, F.; Seeger, W.; Weissmann, N.; et al. IRAG1 Deficient Mice Develop PKG1β Dependent Pulmonary Hypertension. Cells 2020, 9. [Google Scholar] [CrossRef]
- Peters, C.H.; Myers, M.E.; Juchno, J.; Haimbaugh, C.; Bichraoui, H.; Du, Y.; Bankston, J.R.; Walker, L.A.; Proenza, C. Isoform-specific regulation of HCN4 channels by a family of endoplasmic reticulum proteins. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 18079–18090. [Google Scholar] [CrossRef]
- Geiselhöringer, A.; Gaisa, M.; Hofmann, F.; Schlossmann, J. Distribution of IRAG and cGKI-isoforms in murine tissues. FEBS Lett. 2004, 575, 19–22. [Google Scholar] [CrossRef]
- Ali, S.; Solano, A.S.; Gonzales, A.L.; Thakore, P.; Krishnan, V.; Yamasaki, E.; Earley, S. Nitric Oxide Signals Through IRAG to Inhibit TRPM4 Channels and Dilate Cerebral Arteries. Function (Oxf) 2021, 2, zqab051. [Google Scholar] [CrossRef]
- Fritsch, R.M.; Saur, D.; Kurjak, M.; Oesterle, D.; Schlossmann, J.; Geiselhöringer, A.; Hofmann, F.; Allescher, H.-D. InsP3R-associated cGMP kinase substrate (IRAG) is essential for nitric oxide-induced inhibition of calcium signaling in human colonic smooth muscle. Journal of Biological Chemistry 2004, 279, 12551–12559. [Google Scholar] [CrossRef] [PubMed]
- Masuda, W.; Betzenhauser, M.J.; Yule, D.I. InsP3R-associated cGMP kinase substrate determines inositol 1,4,5-trisphosphate receptor susceptibility to phosphoregulation by cyclic nucleotide-dependent kinases. J. Biol. Chem. 2010, 285, 37927–37938. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, A.; Klatt, P.; Massberg, S.; Ny, L.; Sausbier, M.; Hirneiss, C.; Wang, G.X.; Korth, M.; Aszódi, A.; Andersson, K.E.; et al. Defective smooth muscle regulation in cGMP kinase I-deficient mice. EMBO J. 1998, 17, 3045–3051. [Google Scholar] [CrossRef] [PubMed]
- Frei, E.; Huster, M.; Smital, P.; Schlossmann, J.; Hofmann, F.; Wegener, J.W. Calcium-dependent and calcium-independent inhibition of contraction by cGMP/cGKI in intestinal smooth muscle. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G834–G839. [Google Scholar] [CrossRef] [PubMed]
- Koehler, K.; Hmida, D.; Schlossmann, J.; Landgraf, D.; Reisch, N.; Schuelke, M.; Huebner, A. Homozygous mutation in murine retrovirus integration site 1 gene associated with a non-syndromic form of isolated familial achalasia. Neurogastroenterol. Motil. 2020, 32, e13923. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, O.; Mazza, T.; Bassotti, G.; Merla, A.; Tolone, S.; Biagini, T.; Cuttitta, A.; Bossa, F.; Martino, G.; Latiano, T.; et al. microRNA-mRNA network model in patients with achalasia. Neurogastroenterol. Motil. 2020, 32, e13764. [Google Scholar] [CrossRef] [PubMed]
- Angermeier, E.; Domes, K.; Lukowski, R.; Schlossmann, J.; Rathkolb, B.; Angelis, M.H.; Hofmann, F. Iron deficiency anemia in cyclic GMP kinase knockout mice. Haematologica 2016, 101, e48–e51. [Google Scholar] [CrossRef]
- Föller, M.; Feil, S.; Ghoreschi, K.; Koka, S.; Gerling, A.; Thunemann, M.; Hofmann, F.; Schuler, B.; Vogel, J.; Pichler, B.; et al. Anemia and splenomegaly in cGKI-deficient mice. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 6771–6776. [Google Scholar] [CrossRef]
- Singh, A.K.; Spiessberger, B.; Zheng, W.; Xiao, F.; Lukowski, R.; Wegener, J.W.; Weinmeister, P.; Saur, D.; Klein, S.; Schemann, M.; et al. Neuronal cGMP kinase I is essential for stimulation of duodenal bicarbonate secretion by luminal acid. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 2012, 26, 1745–1754. [Google Scholar] [CrossRef]
- Desch, M.; Schinner, E.; Kees, F.; Hofmann, F.; Seifert, R.; Schlossmann, J. Cyclic cytidine 3',5'-monophosphate (cCMP) signals via cGMP kinase I. FEBS Lett. 2010, 584, 3979–3984. [Google Scholar] [CrossRef]
- Santoro, C.; Giugliano, T.; Kraemer, M.; Torella, A.; Schwitalla, J.C.; Cirillo, M.; Melis, D.; Berlit, P.; Nigro, V.; Perrotta, S.; et al. Whole exome sequencing identifies MRVI1 as a susceptibility gene for moyamoya syndrome in neurofibromatosis type 1. PLoS One 2018, 13, e0200446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Qin, F.; Li, X.; Du, X.; Li, T. Identification of novel proteins for lacunar stroke by integrating genome-wide association data and human brain proteomes. BMC medicine 2022, 20, 211. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.J.; Cummins, C.; Radhakrishnan, R.S. Sildenafil Recovers Burn-Induced Cardiomyopathy. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Peters, C.H.; Singh, R.K.; Bankston, J.R.; Proenza, C. Regulation of HCN Channels by Protein Interactions. Front. Physiol. 2022, 13, 928507. [Google Scholar] [CrossRef] [PubMed]
- Massberg, S.; Sausbier, M.; Klatt, P.; Bauer, M.; Pfeifer, A.; Siess, W.; Fässler, R.; Ruth, P.; Krombach, F.; Hofmann, F. Increased adhesion and aggregation of platelets lacking cyclic guanosine 3',5'-monophosphate kinase I. J. Exp. Med. 1999, 189, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Schinner, E.; Salb, K.; Schlossmann, J. Signaling via IRAG is essential for NO/cGMP-dependent inhibition of platelet activation. Platelets 2011, 22, 217–227. [Google Scholar] [CrossRef]
- Eicher, J.D.; Chami, N.; Kacprowski, T.; Nomura, A.; Chen, M.-H.; Yanek, L.R.; Tajuddin, S.M.; Schick, U.M.; Slater, A.J.; Pankratz, N.; et al. Platelet-Related Variants Identified by Exomechip Meta-analysis in 157,293 Individuals. Am. J. Hum. Genet. 2016, 99, 40–55. [Google Scholar] [CrossRef]
- Eicher, J.D.; Xue, L.; Ben-Shlomo, Y.; Beswick, A.D.; Johnson, A.D. Replication and hematological characterization of human platelet reactivity genetic associations in men from the Caerphilly Prospective Study (CaPS). J. Thromb. Thrombolysis 2016, 41, 343–350. [Google Scholar] [CrossRef]
- Johnson, A.D.; Yanek, L.R.; Chen, M.-H.; Faraday, N.; Larson, M.G.; Tofler, G.; Lin, S.J.; Kraja, A.T.; Province, M.A.; Yang, Q.; et al. Genome-wide meta-analyses identifies seven loci associated with platelet aggregation in response to agonists. Nat. Genet. 2010, 42, 608–613. [Google Scholar] [CrossRef]
- Kim, H.; Gillis, L.C.; Jarvis, J.D.; Yang, S.; Huang, K.; Der, S.; Barber, D.L. Tyrosine kinase chromosomal translocations mediate distinct and overlapping gene regulation events. BMC Cancer 2011, 11, 528. [Google Scholar] [CrossRef]
- Zhuang, H.; Li, F.; Si, T.; Renzhi, P.; Yu, M.; Chen, D.; Ye, P.; Lu, Y. High Expression of CD300A Predicts Poor Survival in Acute Myeloid Leukemia. Acta Haematol. 2023. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Hwan, J.D.; Bae, S.; Bae, D.-H.; Shick, W.A. Identification of differentially expressed genes using an annealing control primer system in stage III serous ovarian carcinoma. BMC Cancer 2010, 10, 576. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xu, Y.-P.; Wang, L.-J.; Kong, Y. miR-940 potentially promotes proliferation and metastasis of endometrial carcinoma through regulation of MRVI1. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Li, K.; Jiang, W.; Li, J.; Zhang, J.-A.; Zhu, X. MRVI1 and NTRK3 Are Potential Tumor Suppressor Genes Commonly Inactivated by DNA Methylation in Cervical Cancer. Front. Oncol. 2021, 11, 802068. [Google Scholar] [CrossRef] [PubMed]
- Kusumawidjaja, G.; Kayed, H.; Giese, N.; Bauer, A.; Erkan, M.; Giese, T.; Hoheise, J.D.; Friess, H.; Kleeff, J. Basic transcription factor 3 (BTF3) regulates transcription of tumor-associated genes in pancreatic cancer cells. Cancer Biol. Ther. 2007, 6, 367–376. [Google Scholar] [CrossRef]
- Jin, X.; Sun, Z.-Q.; Zhou, G.-L.; Li, G.-J.; Deng, S.-F. The Higher Expression of CDCA2 Associated with Poor Prognosis in Glioma. Dis. Markers 2022, 2022, 2184867. [Google Scholar] [CrossRef]
- Lee, J.-E.; Oum, B.S.; Choi, H.Y.; Lee, S.U.; Lee, J.S. Evaluation of differentially expressed genes identified in keratoconus. Mol. Vis. 2009, 15, 2480–2487. [Google Scholar]
- Yaroslavskiy, B.B.; Turkova, I.; Wang, Y.; Robinson, L.J.; Blair, H.C. Functional osteoclast attachment requires inositol-1,4,5-trisphosphate receptor-associated cGMP-dependent kinase substrate. Lab. Invest. 2010, 90, 1533–1542. [Google Scholar] [CrossRef]
- Ye, J.; Li, Y.; Kong, C.; Ren, Y.; Lu, H. Label-free proteomic analysis and functional analysis in patients with intrauterine adhesion. J. Proteomics 2023, 277, 104854. [Google Scholar] [CrossRef]
- Chen, W.; Oberwinkler, H.; Werner, F.; Gaßner, B.; Nakagawa, H.; Feil, R.; Hofmann, F.; Schlossmann, J.; Dietrich, A.; Gudermann, T.; et al. Atrial natriuretic peptide-mediated inhibition of microcirculatory endothelial Ca2+ and permeability response to histamine involves cGMP-dependent protein kinase I and TRPC6 channels. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2121–2129. [Google Scholar] [CrossRef]
- Fung, K.; Ramírez, J.; Warren, H.R.; Aung, N.; Lee, A.M.; Tzanis, E.; Petersen, S.E.; Munroe, P.B. Genome-wide association study identifies loci for arterial stiffness index in 127,121 UK Biobank participants. Scientific reports 2019, 9, 9143. [Google Scholar] [CrossRef] [PubMed]
- Gormley, P.; Anttila, V.; Winsvold, B.S.; Palta, P.; Esko, T.; Pers, T.H.; Farh, K.-H.; Cuenca-Leon, E.; Muona, M.; Furlotte, N.A.; et al. Meta-analysis of 375,000 individuals identifies 38 susceptibility loci for migraine. Nat. Genet. 2016, 48, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Daghals, I.; Sargurupremraj, M.; Danning, R.; Gormley, P.; Malik, R.; Amouyel, P.; Metso, T.; Pezzini, A.; Kurth, T.; Debette, S.; et al. Migraine, Stroke, and Cervical Arterial Dissection: Shared Genetics for a Triad of Brain Disorders With Vascular Involvement. Neurol. Genet. 2022, 8, e653. [Google Scholar] [CrossRef] [PubMed]
- Rudzik, R.; Dziedziejko, V.; Rać, M.E.; Sawczuk, M.; Maciejewska-Skrendo, A.; Safranow, K.; Pawlik, A. Polymorphisms in GP6, PEAR1A, MRVI1, PIK3CG, JMJD1C, and SHH Genes in Patients with Unstable Angina. Int. J. Environ. Res. Public Health 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.A.R.; Mathur, R.; Vonk, J.M.; Szwajda, A.; Brumpton, B.; Granell, R.; Brew, B.K.; Ullemar, V.; Lu, Y.; Jiang, Y.; et al. Genetic Architectures of Childhood- and Adult-Onset Asthma Are Partly Distinct. Am. J. Hum. Genet. 2019, 104, 665–684. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Wang, Y.; Zhou, S.; Xia, X.; Han, R.; Fei, G.; Zeng, D.; Wang, R. Identification of three hub genes related to the prognosis of idiopathic pulmonary fibrosis using bioinformatics analysis. Int. J. Med. Sci. 2022, 19, 1417–1429. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, C.; Fan, R.; Liu, X.; Zhang, W. A prognostic model based on clusters of molecules related to epithelial-mesenchymal transition for idiopathic pulmonary fibrosis. Frontiers in genetics 2022, 13, 1109903. [Google Scholar] [CrossRef]
- Behrens, T.W.; Kearns, G.M.; Rivard, J.J.; Bernstein, H.D.; Yewdell, J.W.; Staudt, L.M. Carboxyl-terminal targeting and novel post-translational processing of JAW1, a lymphoid protein of the endoplasmic reticulum. Journal of Biological Chemistry 1996, 271, 23528–23534. [Google Scholar] [CrossRef]
- Kozono, T.; Tadahira, K.; Okumura, W.; Itai, N.; Tamura-Nakano, M.; Dohi, T.; Tonozuka, T.; Nishikawa, A. Jaw1/LRMP has a role in maintaining nuclear shape via interaction with SUN proteins. J. Biochem. 2018, 164, 303–311. [Google Scholar] [CrossRef]
- Horn, H.F.; Kim, D.; Wright, G.D.; Wong, E.S.M.; Stewart, C.L.; Burke, B.; Roux, K.J. A mammalian KASH domain protein coupling meiotic chromosomes to the cytoskeleton. J. Cell Biol. 2013, 202, 1023–1039. [Google Scholar] [CrossRef]
- Kozono, T.; Jogano, C.; Okumura, W.; Sato, H.; Matsui, H.; Takagi, T.; Okumura, N.; Takao, T.; Tonozuka, T.; Nishikawa, A. Cleavage of the Jaw1 C-terminal region enhances its augmentative effect on the Ca2+ release via IP3 receptors. J. Cell Sci. 2023, 136. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-Y.; Wang, J.; Zhao, Y.; Liu, J.; Yang, X.; Yue, X.; Wang, H.; Zhou, F.; Inclan-Rico, J.M.; Ponessa, J.J.; et al. Tumor suppressor p53 regulates intestinal type 2 immunity. Nat. Commun. 2021, 12, 3371. [Google Scholar] [CrossRef] [PubMed]
- Prüschenk, S.; Schlossmann, J. Function of IRAG2 Is Modulated by NO/cGMP in Murine Platelets. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Starr, D.A. A nuclear-envelope bridge positions nuclei and moves chromosomes. J. Cell Sci. 2009, 122, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Ke, H.; Gao, F.; Ren, J.; Wang, M.; Huo, L.; Gong, W.; Feng, W. Coiled-Coil Domains of SUN Proteins as Intrinsic Dynamic Regulators. Structure 2016, 24, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Sosa, B.A.; Rothballer, A.; Kutay, U.; Schwartz, T.U. LINC complexes form by binding of three KASH peptides to domain interfaces of trimeric SUN proteins. Cell 2012, 149, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shi, Z.; Jiao, S.; Chen, C.; Wang, H.; Liu, G.; Wang, Q.; Zhao, Y.; Greene, M.I.; Zhou, Z. Structural insights into SUN-KASH complexes across the nuclear envelope. Cell Res. 2012, 22, 1440–1452. [Google Scholar] [CrossRef]
- Crisp, M.; Liu, Q.; Roux, K.; Rattner, J.B.; Shanahan, C.; Burke, B.; Stahl, P.D.; Hodzic, D. Coupling of the nucleus and cytoplasm: role of the LINC complex. J. Cell Biol. 2006, 172, 41–53. [Google Scholar] [CrossRef]
- Kozono, T.; Sato, H.; Okumura, W.; Jogano, C.; Tamura-Nakano, M.; Kawamura, Y.I.; Rohrer, J.; Tonozuka, T.; Nishikawa, A. The N-terminal region of Jaw1 has a role to inhibit the formation of organized smooth endoplasmic reticulum as an intrinsically disordered region. Scientific reports 2021, 11, 753. [Google Scholar] [CrossRef]
- Snapp, E.L.; Hegde, R.S.; Francolini, M.; Lombardo, F.; Colombo, S.; Pedrazzini, E.; Borgese, N.; Lippincott-Schwartz, J. Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol. 2003, 163, 257–269. [Google Scholar] [CrossRef]
- Parrish, M.L.; Sengstag, C.; Rine, J.D.; Wright, R.L. Identification of the sequences in HMG-CoA reductase required for karmellae assembly. Mol. Biol. Cell 1995, 6, 1535–1547. [Google Scholar] [CrossRef] [PubMed]
- Fasana, E.; Fossati, M.; Ruggiano, A.; Brambillasca, S.; Hoogenraad, C.C.; Navone, F.; Francolini, M.; Borgese, N. A VAPB mutant linked to amyotrophic lateral sclerosis generates a novel form of organized smooth endoplasmic reticulum. FASEB J. 2010, 24, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Mao, A.H.; Crick, S.L.; Vitalis, A.; Chicoine, C.L.; Pappu, R.V. Net charge per residue modulates conformational ensembles of intrinsically disordered proteins. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 8183–8188. [Google Scholar] [CrossRef] [PubMed]
- Babu, M.M.; Kriwacki, R.W.; Pappu, R.V. Structural biology. Versatility from protein disorder. Science (New York, N.Y.) 2012, 337, 1460–1461. [Google Scholar] [CrossRef]
- Faust, O.; Bigman, L.; Friedler, A. A role of disordered domains in regulating protein oligomerization and stability. Chem. Commun. (Camb) 2014, 50, 10797–10800. [Google Scholar] [CrossRef] [PubMed]
- Dyson, H.J.; Wright, P.E. Intrinsically unstructured proteins and their functions. Nat. Rev. Mol. Cell Biol. 2005, 6, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Hieda, M.; Matsumoto, T.; Isobe, M.; Kurono, S.; Yuka, K.; Kametaka, S.; Wang, J.-Y.; Chi, Y.-H.; Kameda, K.; Kimura, H.; et al. The SUN2-nesprin-2 LINC complex and KIF20A function in the Golgi dispersal. Scientific reports 2021, 11, 5358. [Google Scholar] [CrossRef] [PubMed]
- Gimpel, P.; Lee, Y.L.; Sobota, R.M.; Calvi, A.; Koullourou, V.; Patel, R.; Mamchaoui, K.; Nédélec, F.; Shackleton, S.; Schmoranzer, J.; et al. Nesprin-1α-Dependent Microtubule Nucleation from the Nuclear Envelope via Akap450 Is Necessary for Nuclear Positioning in Muscle Cells. Curr. Biol. 2017, 27, 2999–3009. [Google Scholar] [CrossRef]
- Roux, K.J.; Crisp, M.L.; Liu, Q.; Kim, D.; Kozlov, S.; Stewart, C.L.; Burke, B. Nesprin 4 is an outer nuclear membrane protein that can induce kinesin-mediated cell polarization. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 2194–2199. [Google Scholar] [CrossRef]
- Okumura, W.; Tadahira, K.; Kozono, T.; Tamura-Nakano, M.; Sato, H.; Matsui, H.; Dohi, T.; Rohrer, J.; Tonozuka, T.; Nishikawa, A. Jaw1/LRMP is associated with the maintenance of Golgi ribbon structure. J Biochem 2023. [Google Scholar] [CrossRef]
- Berridge, M.J. Inositol trisphosphate and calcium signalling. Nature 1993, 361, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Okumura, W.; Kozono, T.; Sato, H.; Matsui, H.; Takagi, T.; Tonozuka, T.; Nishikawa, A. Jaw1/LRMP increases Ca2+ influx upon GPCR stimulation with heterogeneous effect on the activity of each ITPR subtype. Scientific reports 2022, 12, 9476. [Google Scholar] [CrossRef] [PubMed]
- Makhoul, S.; Walter, E.; Pagel, O.; Walter, U.; Sickmann, A.; Gambaryan, S.; Smolenski, A.; Zahedi, R.P.; Jurk, K. Effects of the NO/soluble guanylate cyclase/cGMP system on the functions of human platelets. Nitric Oxide 2018, 76, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Butt, E.; Nolte, C.; Schulz, S.; Beltman, J.; Beavo, J.A.; Jastorff, B.; Walter, U. Analysis of the functional role of cGMP-dependent protein kinase in intact human platelets using a specific activator 8-para-chlorophenylthio-cGMP. Biochem. Pharmacol. 1992, 43, 2591–2600. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.-C.; Chen, Z.-H.; Xue, J.-B.; Zhao, D.-X.; Lu, C.; Li, Y.-H.; Li, S.-M.; Du, Y.-W.; Liu, Q.; Wang, P.; et al. Infection by the parasitic helminth Trichinella spiralis activates a Tas2r-mediated signaling pathway in intestinal tuft cells. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 5564–5569. [Google Scholar] [CrossRef] [PubMed]
- DiFrancesco, D.; Tortora, P. Direct activation of cardiac pacemaker channels by intracellular cyclic AMP. Nature 1991, 351, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Wainger, B.J.; DeGennaro, M.; Santoro, B.; Siegelbaum, S.A.; Tibbs, G.R. Molecular mechanism of cAMP modulation of HCN pacemaker channels. Nature 2001, 411, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Wicks, N.L.; Wong, T.; Sun, J.; Madden, Z.; Young, E.C. Cytoplasmic cAMP-sensing domain of hyperpolarization-activated cation (HCN) channels uses two structurally distinct mechanisms to regulate voltage gating. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 609–614. [Google Scholar] [CrossRef]
- Hisatsune, C.; Yasumatsu, K.; Takahashi-Iwanaga, H.; Ogawa, N.; Kuroda, Y.; Yoshida, R.; Ninomiya, Y.; Mikoshiba, K. Abnormal taste perception in mice lacking the type 3 inositol 1,4,5-trisphosphate receptor. Journal of Biological Chemistry 2007, 282, 37225–37231. [Google Scholar] [CrossRef]
- Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Mueller, K.L.; Cook, B.; Wu, D.; Zuker, C.S.; Ryba, N.J.P. Coding of sweet, bitter, and umami tastes: different receptor cells sharing similar signaling pathways. Cell 2003, 112, 293–301. [Google Scholar] [CrossRef]
- Matthews, E.K.; Petersen, O.H.; Williams, J.A. Pancreatic acinar cells: acetylcholine-induced membrane depolarization, calcium efflux and amylase release. J. Physiol. 1973, 234, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Petersen, O.H. Ca2+ signaling in pancreatic acinar cells: physiology and pathophysiology. Braz. J. Med. Biol. Res. 2009, 42, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Herbert, Y. Gaisano, Subhankar Dolai, and Toshimasa Takahashi. Physiologic Exocytosis in Pancreatic Acinar Cells and Pathologic Fusion Underlying Pancreatitis, 2020.
- Ruggeri, Z.M. Platelets in atherothrombosis. Nat. Med. 2002, 8, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.A.; Eisen, M.B.; Davis, R.E.; Ma, C.; Lossos, I.S.; Rosenwald, A.; Boldrick, J.C.; Sabet, H.; Tran, T.; Yu, X.; et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 2000, 403, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Rosenwald, A.; Wright, G.; Chan, W.C.; Connors, J.M.; Campo, E.; Fisher, R.I.; Gascoyne, R.D.; Muller-Hermelink, H.K.; Smeland, E.B.; Giltnane, J.M.; et al. The use of molecular profiling to predict survival after chemotherapy for diffuse large-B-cell lymphoma. N. Engl. J. Med. 2002, 346, 1937–1947. [Google Scholar] [CrossRef] [PubMed]
- Lossos, I.S.; Czerwinski, D.K.; Alizadeh, A.A.; Wechser, M.A.; Tibshirani, R.; Botstein, D.; Levy, R. Prediction of survival in diffuse large-B-cell lymphoma based on the expression of six genes. N. Engl. J. Med. 2004, 350, 1828–1837. [Google Scholar] [CrossRef] [PubMed]
- Snyder, H.L.; Bacík, I.; Bennink, J.R.; Kearns, G.; Behrens, T.W.; Bächi, T.; Orlowski, M.; Yewdell, J.W. Two novel routes of transporter associated with antigen processing (TAP)-independent major histocompatibility complex class I antigen processing. J. Exp. Med. 1997, 186, 1087–1098. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.P.; Lippman, S.M.; Spier, C.M.; Slymen, D.J.; Grogan, T.M. HLA-DR (Ia) immune phenotype predicts outcome for patients with diffuse large cell lymphoma. J. Clin. Invest. 1988, 82, 370–372. [Google Scholar] [CrossRef]
- Rimsza, L.M.; Roberts, R.A.; Miller, T.P.; Unger, J.M.; LeBlanc, M.; Braziel, R.M.; Weisenberger, D.D.; Chan, W.C.; Muller-Hermelink, H.K.; Jaffe, E.S.; et al. Loss of MHC class II gene and protein expression in diffuse large B-cell lymphoma is related to decreased tumor immunosurveillance and poor patient survival regardless of other prognostic factors: a follow-up study from the Leukemia and Lymphoma Molecular Profiling Project. Blood 2004, 103, 4251–4258. [Google Scholar] [CrossRef]
- Marchetti, P.; Antonov, A.; Anemona, L.; Vangapandou, C.; Montanaro, M.; Botticelli, A.; Mauriello, A.; Melino, G.; Catani, M.V. New immunological potential markers for triple negative breast cancer: IL18R1, CD53, TRIM, Jaw1, LTB, PTPRCAP. Discov. Oncol. 2021, 12, 6. [Google Scholar] [CrossRef]
- Jin, X.; Chen, L.; Zhou, N.; Ni, H.; Zu, L.; He, J.; Yang, L.; Zhu, Y.; Sun, X.; Li, X.; et al. LRMP Associates With Immune Infiltrates and Acts as a Prognostic Biomarker in Lung Adenocarcinoma. Front. Mol. Biosci. 2021, 8, 711928. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Itoh, T.; Imamichi, S.; Kikuhara, S.; Fujimori, H.; Hirai, T.; Saito, S.; Sakurai, Y.; Tanaka, H.; Nakamura, H.; et al. Proteomic analysis of cellular response induced by boron neutron capture reaction in human squamous cell carcinoma SAS cells. Appl. Radiat. Isot. 2015, 106, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Gariboldi, M.; Manenti, G.; Canzian, F.; Falvella, F.S.; Radice, M.T.; Pierotti, M.A.; Della Porta, G.; Binelli, G.; Dragani, T.A. A major susceptibility locus to murine lung carcinogenesis maps on chromosome 6. Nat. Genet. 1993, 3, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Manenti, G.; Dragani, T.A. Pas1 haplotype-dependent genetic predisposition to lung tumorigenesis in rodents: a meta-analysis. Carcinogenesis 2005, 26, 875–882. [Google Scholar] [CrossRef]
- Manenti, G.; Galbiati, F.; Giannì-Barrera, R.; Pettinicchio, A.; Acevedo, A.; Dragani, T.A. Haplotype sharing suggests that a genomic segment containing six genes accounts for the pulmonary adenoma susceptibility 1 (Pas1) locus activity in mice. Oncogene 2004, 23, 4495–4504. [Google Scholar] [CrossRef]
- Manenti, G.; Galbiati, F.; Pettinicchio, A.; Spinola, M.; Piconese, S.; Leoni, V.P.; Conti, B.; Ravagnani, F.; Incarbone, M.; Pastorino, U.; et al. A V141L polymorphism of the human LRMP gene is associated with survival of lung cancer patients. Carcinogenesis 2006, 27, 1386–1390. [Google Scholar] [CrossRef]
- Maria, D.A.; Manenti, G.; Galbiati, F.; Ribeiro, O.G.; Cabrera, W.H.K.; Barrera, R.G.; Pettinicchio, A.; de Franco, M.; Starobinas, N.; Siqueira, M.; et al. Pulmonary adenoma susceptibility 1 (Pas1) locus affects inflammatory response. Oncogene 2003, 22, 426–432. [Google Scholar] [CrossRef]
Figure 1.
Schematical illustration of the primary structure and protein motifs of IRAG1 and IRAG2. Numbers indicate position of amino acids. PKGI phosphorylation sites are depicted in red (S: serine, T: threonine).
Figure 1.
Schematical illustration of the primary structure and protein motifs of IRAG1 and IRAG2. Numbers indicate position of amino acids. PKGI phosphorylation sites are depicted in red (S: serine, T: threonine).
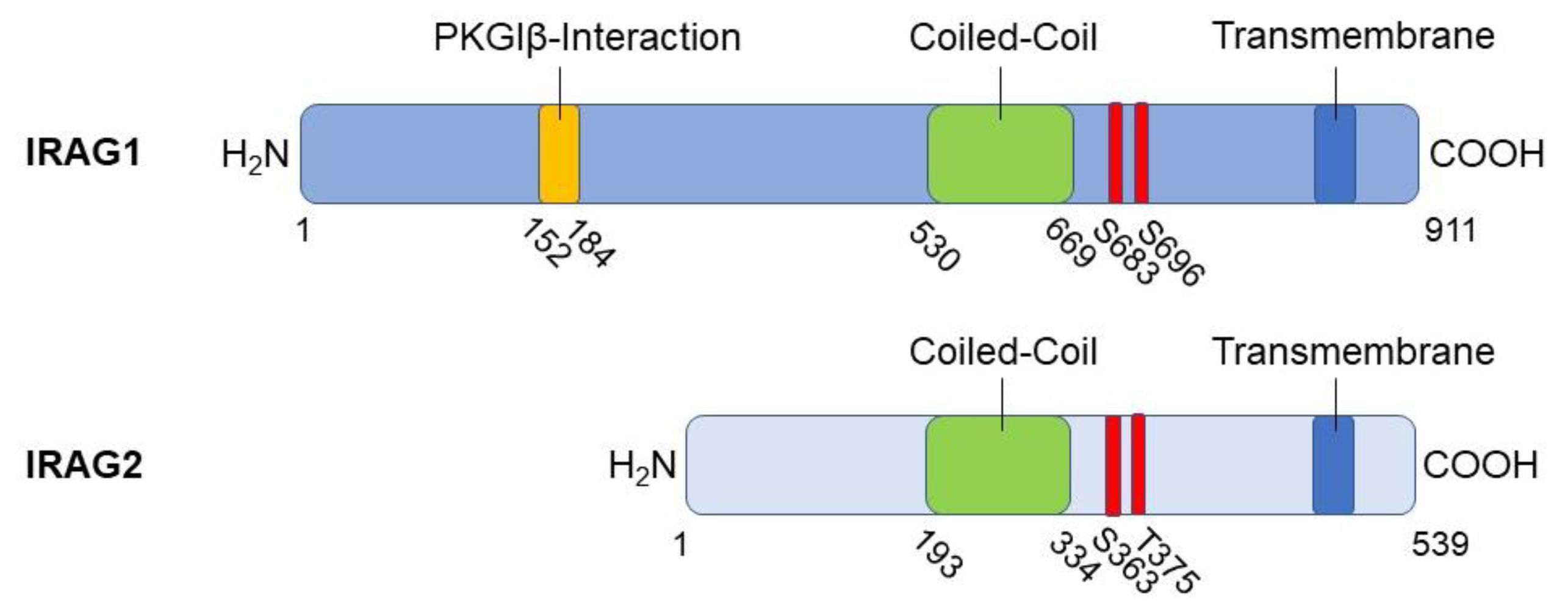
Figure 2.
Graphical illustration of the PKGIβ/IRAG1 signaling pathway. The PKGIβ (light green) interacts with IRAG1 (red) by its leucine zipper region (dark green). After cGMP-dependent phosphorylation, IRAG1 mediates Ca2+ release from the endoplasmic/sarcoplasmic reticulum via the IP3R-I (blue). For detailed description see text below.
Figure 2.
Graphical illustration of the PKGIβ/IRAG1 signaling pathway. The PKGIβ (light green) interacts with IRAG1 (red) by its leucine zipper region (dark green). After cGMP-dependent phosphorylation, IRAG1 mediates Ca2+ release from the endoplasmic/sarcoplasmic reticulum via the IP3R-I (blue). For detailed description see text below.
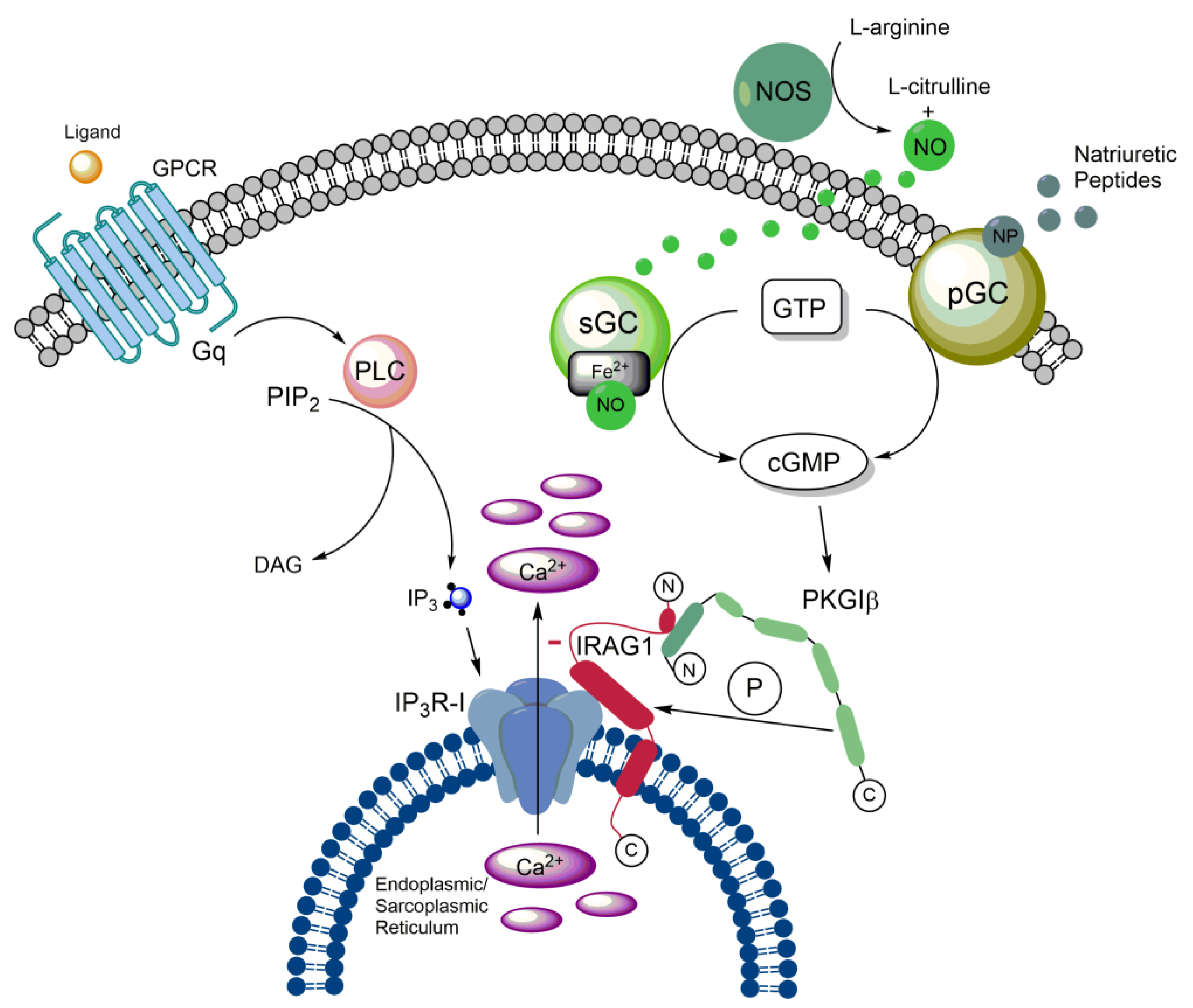
Figure 3.
Graphical illustration of the PKGI/IRAG2 signaling pathway. After cGMP-dependent phosphorylation of IRAG2 by PKGI, IRAG2 mediates Ca2+ release from the endoplasmic reticulum via IP3-receptors. A detailed description is in the text.
Figure 3.
Graphical illustration of the PKGI/IRAG2 signaling pathway. After cGMP-dependent phosphorylation of IRAG2 by PKGI, IRAG2 mediates Ca2+ release from the endoplasmic reticulum via IP3-receptors. A detailed description is in the text.
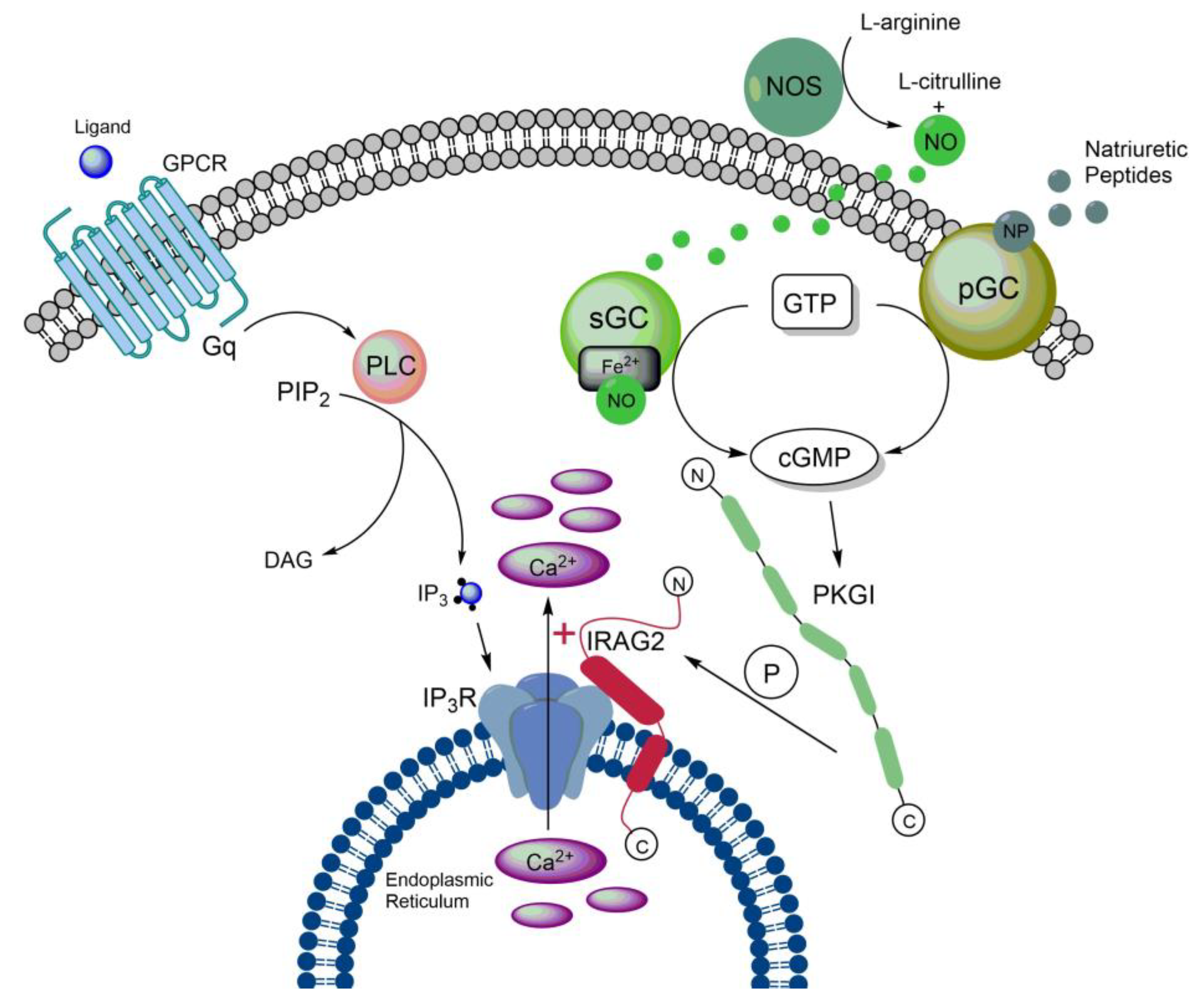
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



